Introduction
Renal cell carcinoma (RCC) is accountable for ~90%
of renal cancer patients and is the most prevalent malignancies of
kidney cancer (1). Among all RCC
histological subtypes, clear cell RCC (ccRCC) is the most malignant
form and leads to the most cancer-related deaths (2). Although marked development has been
achieved in the diagnosis and treatment of ccRCC, ~30% of patients
have metastatic disease (3).
Surgery is still the most effective treatment for localized primary
ccRCC, but ccRCC is resistant to conventional treatments such as
radiation, hormone treatment and chemotherapy. Despite the fact
that specific targeted therapies with favorable clinical outcomes
to a certain extent have been developed, individual differences in
response and the risk of adverse effects restrict the use of these
drugs (4). Therefore, appropriate
ccRCC biomarkers may contribute to improve early diagnosis and
patient therapy.
Dyskerin is a predominantly nucleolar protein
encoded by the DKC1 gene and is involved in dyskeratosis congenital
(5). It is a component of H/ACA
small nucleolar ribonucleoprotein with various characteristics,
such as bone marrow failure, mucocutaneous abnormalities and an
increased tumor susceptivity (6).
Several lines of evidence have demonstrated that DKC1 expression is
significantly upregulated and associated with poor prognosis in
some human cancers, such as prostate cancer (7), neuroblastomas (8) and hepatocellular carcinoma (9). Despite these survey results, the
significance of dyskerin expression in cancer has been disputed,
with some research revealing that dyskerin may have a function as a
tumor suppressor (10,11). In addition, the specific mechanism
and significance of DKC1 in ccRCC progression are not fully
known.
In this study, we sought to examine the expression
levels of DKC1 in ccRCC, and then explored its association with the
clinical characteristics, 5-year overall and disease-specific
survival of ccRCC patients. In addition, we explored the difference
in the expression levels of DKC1 in renal cancer and paracancerous
tissues. Moreover, we also investigated how DKC1 regulated ccRCC
cell proliferation, migration and invasion in vitro, and
tumor growth in vivo as well as the molecular
mechanisms.
Materials and methods
Patients and specimens
Two independent retrospective ccRCC cohorts with
tissue microarrays (TMAs) were examined in the present study. The
small ccRCC TMA was obtained from Shanghai Outdo Biotechnology
(Shanghai, China), which contained 75 pairs of ccRCC tissues and
matched paracancerous tissues. It contained 75 patients who
underwent radical nephrectomy between November 2006 and September
2008. Another large TMA was composed of 310 ccRCC tissues from
patients collected from the Affiliated Hospital of Xuzhou Medical
University who underwent radical nephrectomy without prior
treatment from February 2005 to December 2008, and their
clinicopathological characteristics containing age, sex, tumor
diameter, depth of invasion, lymph node and distant metastasis, as
well as tumor-node-metastasis (TNM) stage were obtained. Due to
specimen deficiency during antigen retrieval, finally 307 ccRCC
patients in the large TMA were utilized to explore the association
of DKC1 expression with clinicopathological features. Informed
consents from all patients were obtained and institutional approval
was obtained by the review board of the Affiliated Hospital of
Xuzhou Medical University prior to this study.
Construction of TMAs and
immunohistochemistry (IHC)
Construction of the large TMA was performed by
contract service at the National Engineering Center for Biochip
(Shanghai, China). Each tissue microarray dot was cut 1.5 mm in
diameter from the paraffin tumor block. The standard procedure for
IHC of TMA was performed as previously described (12). The polyclonal rabbit anti-DKC1
(1:50, cat. no. ab64667; Abcam, Cambridge, MA, USA) was used for
primary antibody incubation at 4°C overnight. The slides without
primary antibody incubation were used as negative controls.
Assessment of immunostaining
The evaluation of DKC1 staining was blindly examined
by two pathologists. Positive DKC1 immunostaining was defined
mainly in the cytoplasm but could also be observed in the nucleus
area. The tissues were scored according to both the intensity and
percentage of cells with positive staining. The staining intensity
of DKC1 was scored as 0–3 (0=negative; 1=weak; 2=moderate;
3=strong). The percentage of DKC1-positive stained cells was also
scored into 4 categories: 1 (0–25%); 2 (26–50%); 3 (51–75%); and 4
(76–100%). In the case of a discrepancy between copied cores, the
mean score from the two tissue cores was selected as the final
score. The level of DKC1 staining was evaluated by immunoreactive
score (IRS), which is calculated by multiplying the scores of the
staining intensity and the percentage of positive cells. For
statistical analysis, scores of 0–5 were considered as low
expression, while scores of 6–12 were considered as high
expression; the cut-off value was determined by receiver operating
characteristic curve analysis.
Animals and cell lines
A total of 20 female BALB/c nude mice weighing 14–18
g, 6–8 weeks old, were purchased from Beijing HFK Bioscience Co.,
Ltd. (Beijing, China) for studies approved by the Animal Care
Committee of Xuzhou Medical University and maintained under
specific pathogen-free conditions. Human embryo kidney epithelial
cell line HK-2 and human ccRCC cell lines ACHN, 786-O and OSRC-2
were purchased from the Shanghai Institute of Biochemistry and Cell
Biology, Chinese Academy of Sciences (Shanghai, China). HK-2 cells
were cultured in keratinocyte serum free medium (K-SFM)
supplemented with 10% fetal calf serum (FCS; both from Invitrogen;
Thermo Fisher Scientific, Inc., Shanghai, China). ACHN cells were
cultured in Minimum Essential Media medium (MEM) supplemented with
10% FCS (both from Invitrogen; Thermo Fisher Scientific, Inc.).
786-O and OSRC-2 cells were cultured in Roswell Park Memorial
Institute-1640 medium (RPMI-1640; Invitrogen; Thermo Fisher
Scientific, Inc., Shanghai, China) supplemented with 10% FCS. These
four cell lines were both incubated in a 37ºC humidified incubator
with 5% CO2.
Plasmid, siRNA and shRNA
transfections
The DKC1 siRNA and scrambled siRNA were purchased
from Shanghai GenePharma Co., Ltd., (Shanghai, China). The p65
overexpression plasmids were obtained from Dr Yu Wu (Nanjing
Medical University, Nanjing, China). DKC1 siRNA and scrambled siRNA
were transfected into the ACHN and 786-O cells by siLentFect™ Lipid
reagent (Bio-Rad Laboratories, Hercules, CA, USA) following the
manufacturer's protocol. Transfection of p65 overexpression plasmid
and control vector into the ACHN and 786-O cells was performed
using Lipofectamine 2000 transfection reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. The DKC1 knockdown ACHN cell lines
(DKC1KD-ACHN) and control ACHN cell lines (Ctrl-ACHN)
were established by transfecting with lentivirus packing DKC1 shRNA
expression and control vector, respectively (Shanghai GenePharma).
Target cells were transfected with the lentivirus for 48 h and then
selected with puromycin (Santa Cruz Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA) at a concentration of 5 µg/ml for 3 weeks.
Cell proliferation assay
Complete medium (2 ml) containing 1×105
stable DKC1 knockdown ACHN or 786-O cells and corresponding
controls were seeded in 6-well plates and cultured for 24, 48, 72
and 96 h. At exact time-points, the number of cells in the DKC1
knockdown ACHN or 786-O cells and corresponding controls was
counted respectively.
Cell migration and invasion
assays
Cell migration and invasion assays were carried out
using Transwell filter inserts (8.0-µm pore size with polycarbonate
membranes) precoated with or without Matrigel (BD Biosciences,
Franklin Lakes, NJ, USA). The detailed conditions were previously
described (13).
Antibodies and western blotting
(WB)
WB was performed as previously reported (14). The following antibodies against the
corresponding proteins were used for WB: DKC1 (1:1,000 for WB, cat.
no. ab64667; Abcam); NF-κB-p65 (1:1,000 for WB; 1:50 for IHC, cat.
no. 8242P; Cell Signaling Technology, Inc., Beverly, MA, USA);
MMP-2 (1:1,000 for WB, 1:50 for IHC, cat. no. 4022S; Cell Signaling
Technology, Inc.); p-AKT (1:1,000 for WB, cat. no. 4060T; Cell
Signaling Technology, Inc.); p-S6K (1:1,000 for WB, cat. no. 9234T;
Cell Signaling Technology, Inc.); MMP-9 (1:1,000 for WB, cat. no.
3852S; Cell Signaling Technology, Inc.) and GAPDH (1:5,000 for WB,
cat. no. sc-365062; Santa Cruz Biotechnology, Inc.). Then,
HRP-conjugated goat anti-mouse and goat anti-rabbit secondary
antibodies (1:10,000, anti-mouse, cat. no. SA00001-1, anti-rabbit
cat. no. SA00001-2; Proteintech Group, Inc., Rosemont, IL, USA)
were applied to the blot for 1 h at room temperature.
Subcutaneous tumor model in vivo
To produce a subcutaneous tumor model in
vivo, the BALB/c nude mice were randomly divided into two
groups consisting of 15 mice each. Stable DKC1 knockdown
(1×106) and control ACHN cells were suspended in 200 µl
phosphate-buffered saline (PBS) and subcutaneously injected through
axillary fossa, respectively. After 2 weeks, the two groups of mice
were sacrificed; their subcutaneous tumors were excised and fixed
in 10% buffered formalin for statistical analysis and further
histopathological analysis.
Statistical analysis
Paired Wilcoxon test was used to explore the
difference of DKC1 staining between tumors and their matched
non-tumor tissues. Fisher's exact test was carried out to assess
the correlation between DKC1 expression and clinicopathological
features. The Kaplan-Meier method with a log-rank test was used to
investigate the differences of the 5-year survival and
disease-specific survival between the positive DKC1 expression
group and the negative expression group. Univariate and
multivariate Cox proportional hazards regression analysis were used
to evaluate the hazard ratios (HRs) and 95% confidence interval
(CI) of HRs. Independent samples t-test was used in the
proliferation, migration and invasion assays. All the statistical
analyses were performed by SPSS 20.0 statistical software package
(IBM Corp., Armonk, NY, USA). A P-value of <0.05 was defined as
statistically significant, and all tests were two-sided.
Results
DKC1 expression is increased in ccRCC
tissues and RCC cell lines
To explore whether the expression level of DKC1 was
altered in RCC, a small TMA was used to investigate DKC1 protein
expression in 75 pairs of ccRCC and paracancerous tissues using
IHC. Our data revealed that DKC1 protein was localized in both the
cytoplasm and nucleolus (Fig. 1A).
As shown in Table I, high DKC1
expression was detected in 46 out of 75 (61.3%) ccRCC tissues and
in 26 out of 75 (34.7%) paracancerous tissues, and there was a
significant expression difference between ccRCC and paracancerous
tissues (Fig. 1B). Furthermore,
western blot analysis revealed that the expression of DKC1 was
significantly lower in HK-2, a type of human embryo kidney
epithelial cell line, as compared with that in all 3 analyzed RCC
cell lines, including ACHN, 786-O and OSRC-2 (Fig. 1C). Consequently, our results
demonstrated that DKC1 was increased in RCC.
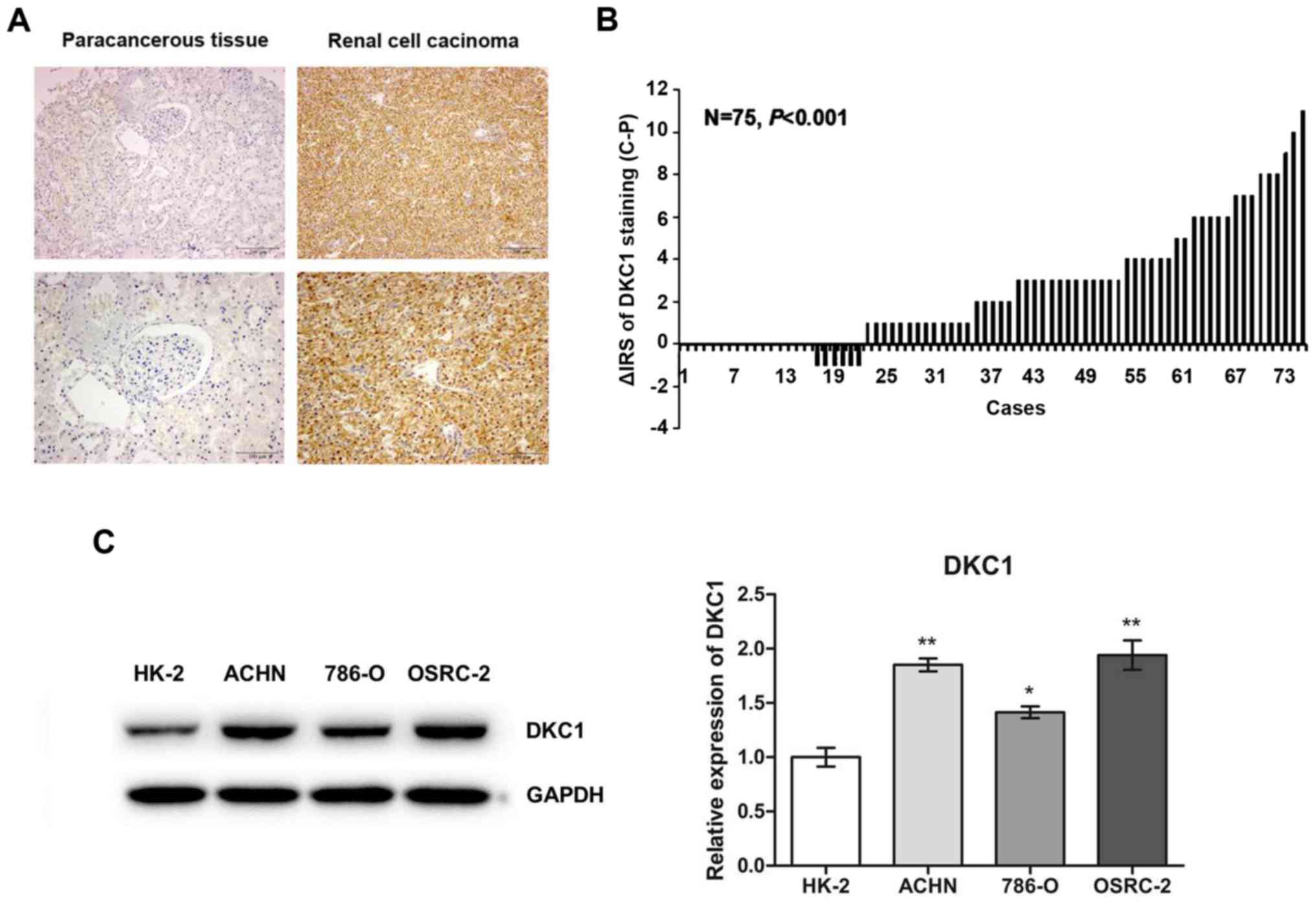 | Figure 1.DKC1 expression is increased in ccRCC
compared with normal renal tissues and cell lines. (A)
Representative images of DKC1 immunohistochemical staining in TMA
are shown. Top panel, magnification, ×100; Bottom panel,
magnification, ×200. (B) The distribution of the difference in
staining intensities of DKC1 in ccRCC tissues compared with
paracancerous tissues. C, ccRCC tissues; P, paracancerous tissues;
IRS, immunoreactivity score. (C) Western blot analysis of DKC1
expression in human embryo kidney epithelial cells HK-2 and renal
cell carcinoma cell lines, including ACHN, 786-O an OSRC-2. Data
are presented as the mean ± SD, *P<0.05, **P<0.01. ccRCC,
clear cell renal cell carcinoma. |
 | Table I.DKC1 expression in ccRCC and
paracancerous tissues. |
Table I.
DKC1 expression in ccRCC and
paracancerous tissues.
|
| DKC1 staining |
|---|
|
|
|
|---|
| Tissues | Low (%) | High (%) | Total |
P-valuea |
|---|
| Paracancerous
tissues | 49 (65.3) | 26 (34.7) | 75 | 0.001 |
| Renal cell
carcinoma | 29 (38.7) | 46 (61.3) | 75 |
|
DKC1 expression is associated with
clinicopathological characteristics in ccRCC patients
To further study the association between DKC1
expression and clinicopathological features, another TMA including
307 cases of ccRCC tissues was used to investigate DKC1 protein
expression. There were 204 male and 103 female patients. Their mean
age was 55.8 years. The distribution of the TNM stage was as
follows: 181 patients at stage I, 55 at stage II, 40 at stage III
and 31 at stage IV. Table II
contains the association between DKC1 and the clinicopathological
features of ccRCC patients. The data revealed that there were
significant correlations of DKC1 expression with tumor size
(P=0.002), pT status (P<0.001), pN status (P=0.041) and TNM
stage (P<0.001). However, no significance was found between DKC1
expression and other clinical characteristics, such as sex and age.
These results indicated that high DKC1 expression tends to be
associated with advanced clinicopathological parameters in ccRCC
patients.
| Table II.DKC1 staining and clinicopathological
characteristics of 307 renal cancer patients. |
Table II.
DKC1 staining and clinicopathological
characteristics of 307 renal cancer patients.
|
| DKC1 staining |
|---|
|
|
|
|---|
| Variables | Low (%) | High (%) | Total |
P-valuea |
|---|
| Age (years) |
|
|
| 0.674 |
|
≤56 | 119 (80.4) | 29 (19.6) | 148 |
|
|
>56 | 124 (78.0) | 35 (22.0) | 159 |
|
| Sex |
|
|
| 0.460 |
|
Male | 164 (80.4) | 40 (19.6) | 204 |
|
|
Female | 79 (76.7) | 24 (23.3) | 103 |
|
| Tumor size
(cm) |
|
|
| 0.002 |
| ≤7 | 115 (57.5) | 85 (42.5) | 200 |
|
|
>7 | 41 (38.3) | 66 (61.7) | 107 |
|
| pT status |
|
|
| <0.001 |
|
pT1-pT2 | 178 (74.8) | 60 (25.2) | 238 |
|
|
pT3-pT4 | 28 (40.6) | 41 (59.4) | 69 |
|
| pN status |
|
|
| 0.041 |
|
pN0 | 154 (54.0) | 131 (46.0) | 285 |
|
|
pN1-pN3 | 9 (40.9) | 13 (59.1) | 22 |
|
| pM status |
|
|
| 0.609 |
|
pM0 | 166 (57.6) | 122 (42.4) | 288 |
|
|
pM1 | 8 (42.1) | 11 (57.9) | 19 |
|
| TNM stage |
|
|
| <0.001 |
|
I–II | 221 (93.6) | 15 (6.4) | 236 |
|
|
III–IV | 22 (31.0) | 49 (69.0) | 71 |
|
DKC1 expression is correlated with the
prognosis of ccRCC patients
To evaluate the prognostic value of DKC1 expression
in ccRCC, the log-rank test combined with Kaplan-Meier survival
curves was established. Our data revealed that ccRCC patients with
high DKC1 expression were correlated with greater unfavorable
5-year overall and disease-specific survival than the rest of the
patients with low DKC1 expression (P<0.001 and P<0.001,
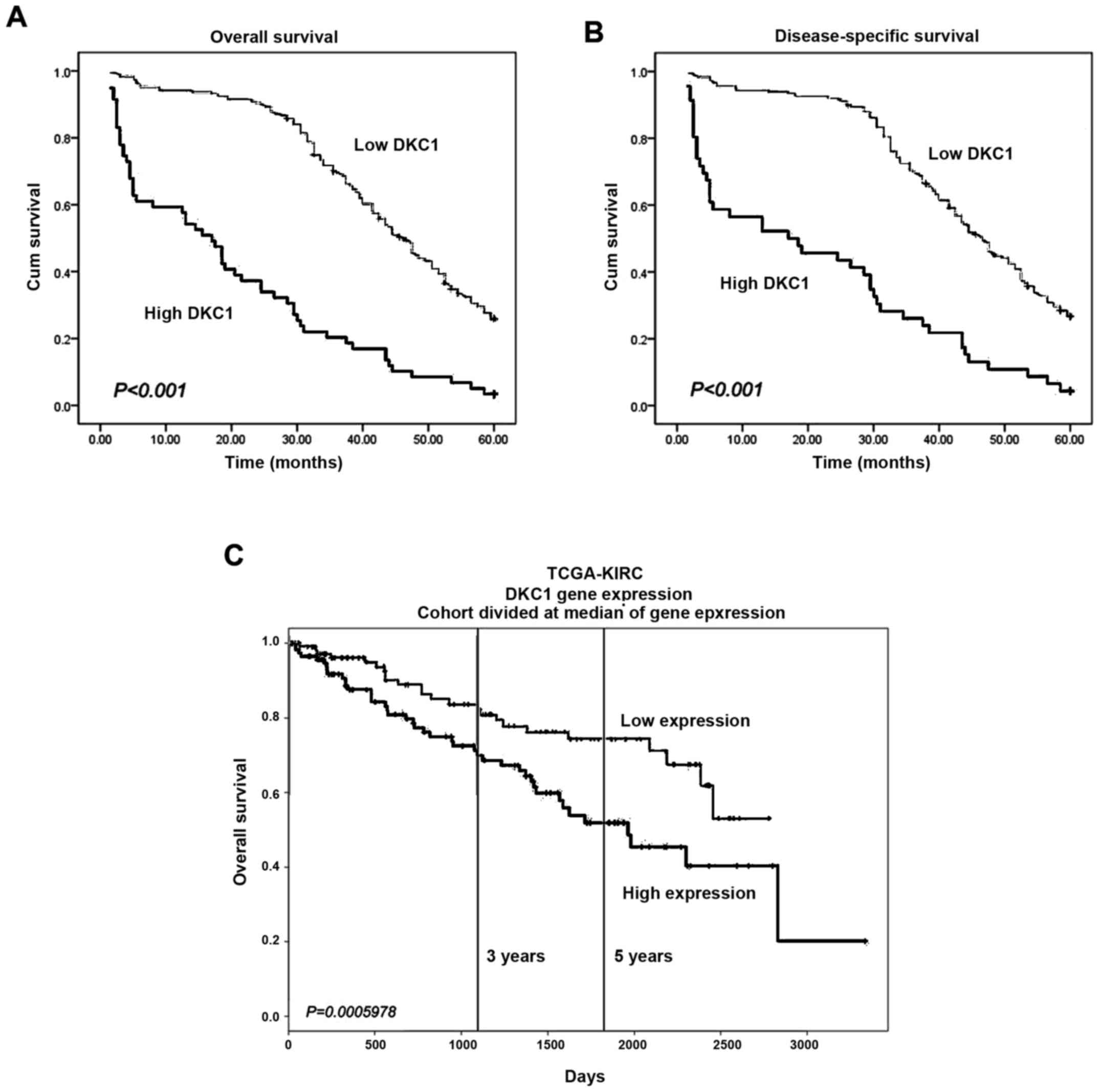
respectively) (Fig. 2A and B).
Furthermore, to investigate whether DKC1 was an independent
prognostic marker in ccRCC, univariate and multivariate COX
analysis was performed. In the univariate COX analysis, our data
suggested that increased DKC1 expression was significantly
correlated with a worse 5-year overall and disease-specific
survival, as well as with other prognostic markers, such as tumor
size, pT status, pN status, pM status and TNM stage (Table III). In addition, both increased
DKC1 expression (HR=1.932, 95% CI, 1.290–2.893, P=0.001 for 5-year
overall survival; HR=1.778, 95% CI, 1.150–2.748, P=0.010 for
disease-free survival) and TNM stage (HR=2.790, 95% CI,
1.861–4.184, P<0.001 for 5-year overall survival; HR=2.689, 95%
CI, 1.746–4.140, P<0.001 for disease-free survival) were
independent adverse prognostic markers for the 5-year overall and
disease-specific survival of ccRCC patients in multivariate COX
analysis (Table IV). By searching
the PROGgeneV2 platform, we found that ccRCC patients with high
DKC1 expression had significant poorer prognosis than those with
low DKC1 expression in The Cancer Genome Atlas (TCGA) which was
consistent with our findings (Fig.
2C).
| Table III.Univariate Cox proportional
regression analysis on the 5-year overall and disease-specific
survival of ccRCC patients. |
Table III.
Univariate Cox proportional
regression analysis on the 5-year overall and disease-specific
survival of ccRCC patients.
|
| Overall
survival | Disease-specific
survival |
|---|
|
|
|
|
|---|
| Variables | Hazard ratio | 95% CIb |
P-valuea | Hazard ratio | 95% CIb |
P-valuea |
|---|
| DKC1 |
|
|
|
|
| <0.001 |
|
Low | 1.000 |
| <0.001 | 1.000 |
|
|
|
High | 3.525 | 2.574–4.828 |
| 3.126 | 2.213–4.414 |
|
| Age (years) |
|
|
|
|
| 0.728 |
|
≤56 | 1.000 |
| 0.637 | 1.000 |
|
|
|
>56 | 1.067 | 0.810–1.410 |
| 1.053 | 0.786–1.411 |
|
| Tumor size
(cm) |
|
|
|
|
| 0.021 |
| ≤7 | 1.000 |
| 0.003 | 1.000 |
|
|
|
>7 | 1.676 | 1.194–2.352 |
| 1.549 | 1.069–2.244 |
|
| pT status |
|
|
|
|
| 0.014 |
|
pT1-pT2 | 1.000 |
| 0.002 | 1.000 |
|
|
|
pT3-pT4 | 1.574 | 1.178–2.102 |
| 1.473 | 1.081–2.007 |
|
| pN status |
|
|
|
|
| <0.001 |
|
pN0 | 1.000 |
| <0.001 | 1.000 |
|
|
|
pN1-pN3 | 3.421 | 1.842–6.352 |
| 3.046 | 1.686–5.502 |
|
| pM status |
|
|
|
|
| 0.021 |
|
pM0 | 1.000 |
| <0.001 | 1.000 |
|
|
|
pM1 | 4.162 | 2.282–7.594 |
| 2.870 | 1.172–7.027 |
|
| TNM stage |
|
|
|
|
| <0.001 |
|
I–II | 1.000 |
| <0.001 | 1.000 |
|
|
|
III–IV | 4.084 | 2.972–5.613 |
| 3.713 | 2.628–5.247 |
|
| Table IV.Multvariate Cox regression analysis
on the 5-year overall and disease-specific survival of ccRCC
patients. |
Table IV.
Multvariate Cox regression analysis
on the 5-year overall and disease-specific survival of ccRCC
patients.
|
| Overall
survival | Disease-specific
survival |
|---|
|
|
|
|
|---|
|
Variablesa | Hazard ratio | 95% CIb |
P-valuea | Hazard ratio | 95% CIb |
P-valuea |
|---|
| DKC1 | 1.932 | 1.290–2.893 | 0.001 | 1.778 | 1.150–2.748 | 0.010 |
| Age (years) | 1.189 | 0.897–1.576 | 0.228 | 1.126 | 0.838–1.513 | 0.432 |
| Tumor size | 1.319 | 0.930–1.871 | 0.120 | 1.293 | 0.885–1.888 | 0.184 |
| TNM stage | 2.790 | 1.861–4.184 | <0.001 | 2.689 | 1.746–4.140 | <0.001 |
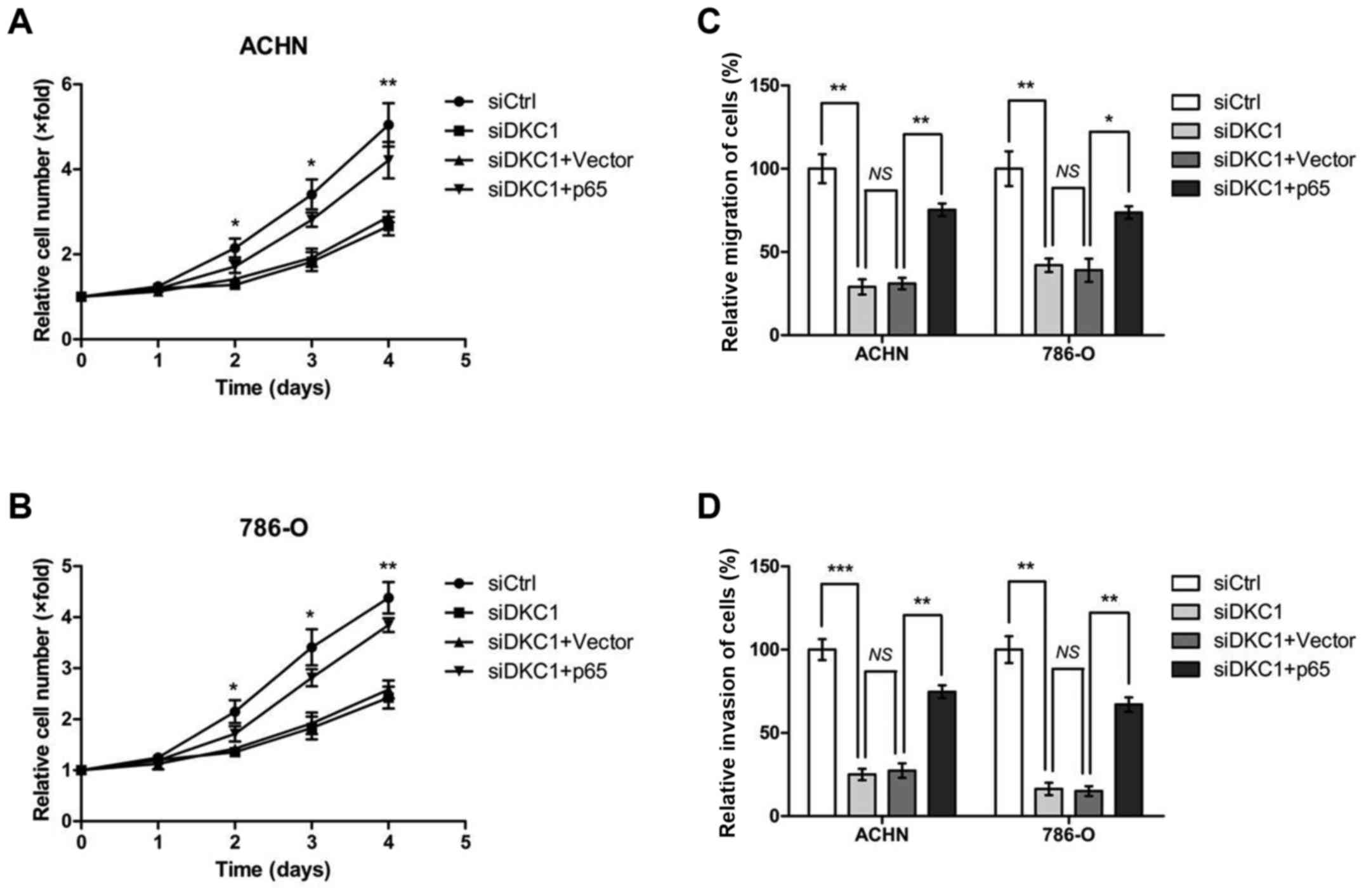
Knockdown of DKC1 suppresses ccRCC
cell proliferation, migration and invasion in vitro
To explore the role of DKC1 in ccRCC progression,
ACHN and 786-O cells were transiently transfected with control
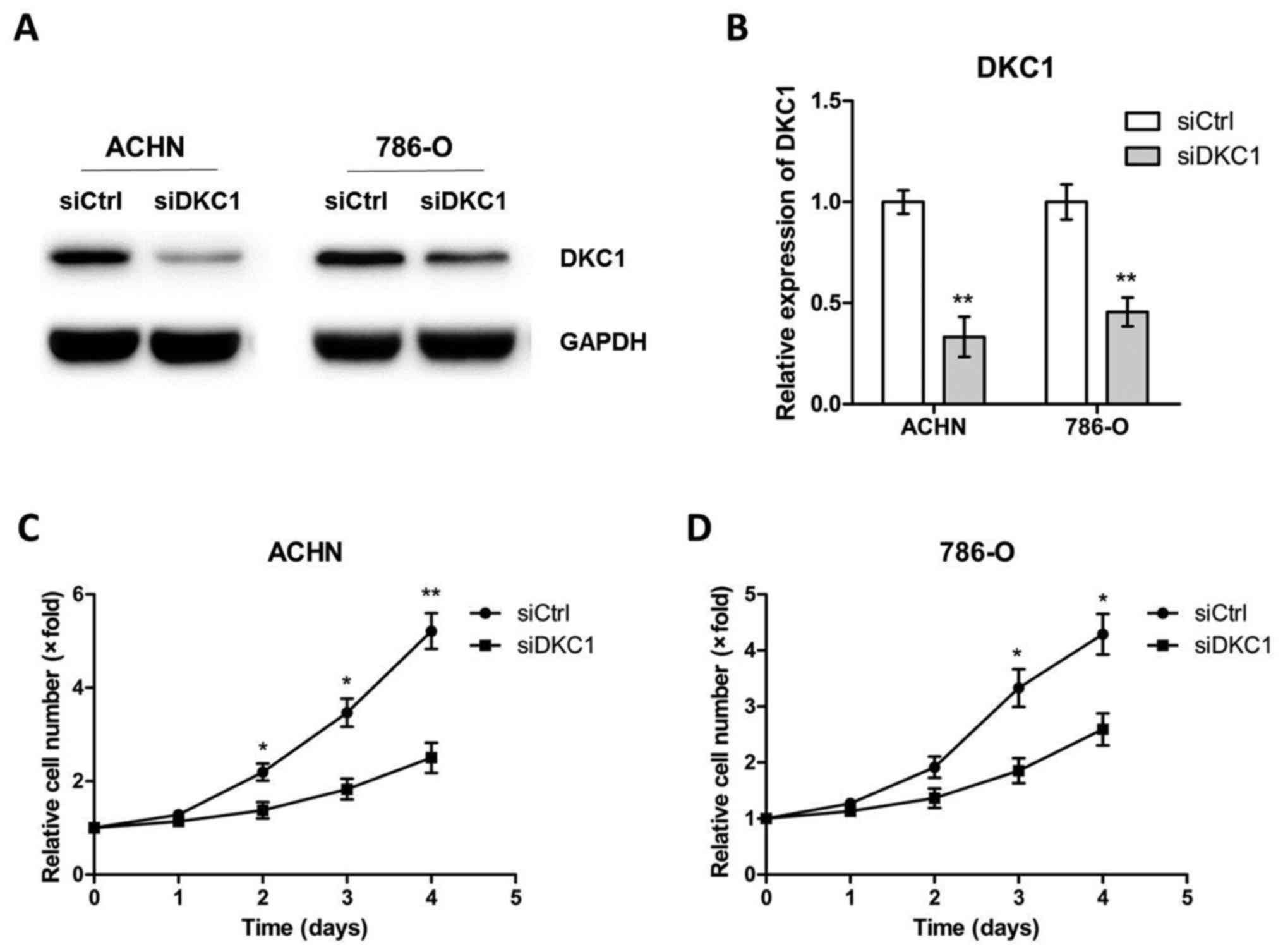
siRNA and DKC1 siRNA, respectively (Fig. 3A and B). Then the cell proliferation
assays were carried out and the data revealed that DKC1 depletion
led to a significant decrease in cell proliferation (Fig. 3C and D). Since high DKC1 expression
was relevant to poor prognosis in the ccRCC patient cohort, we
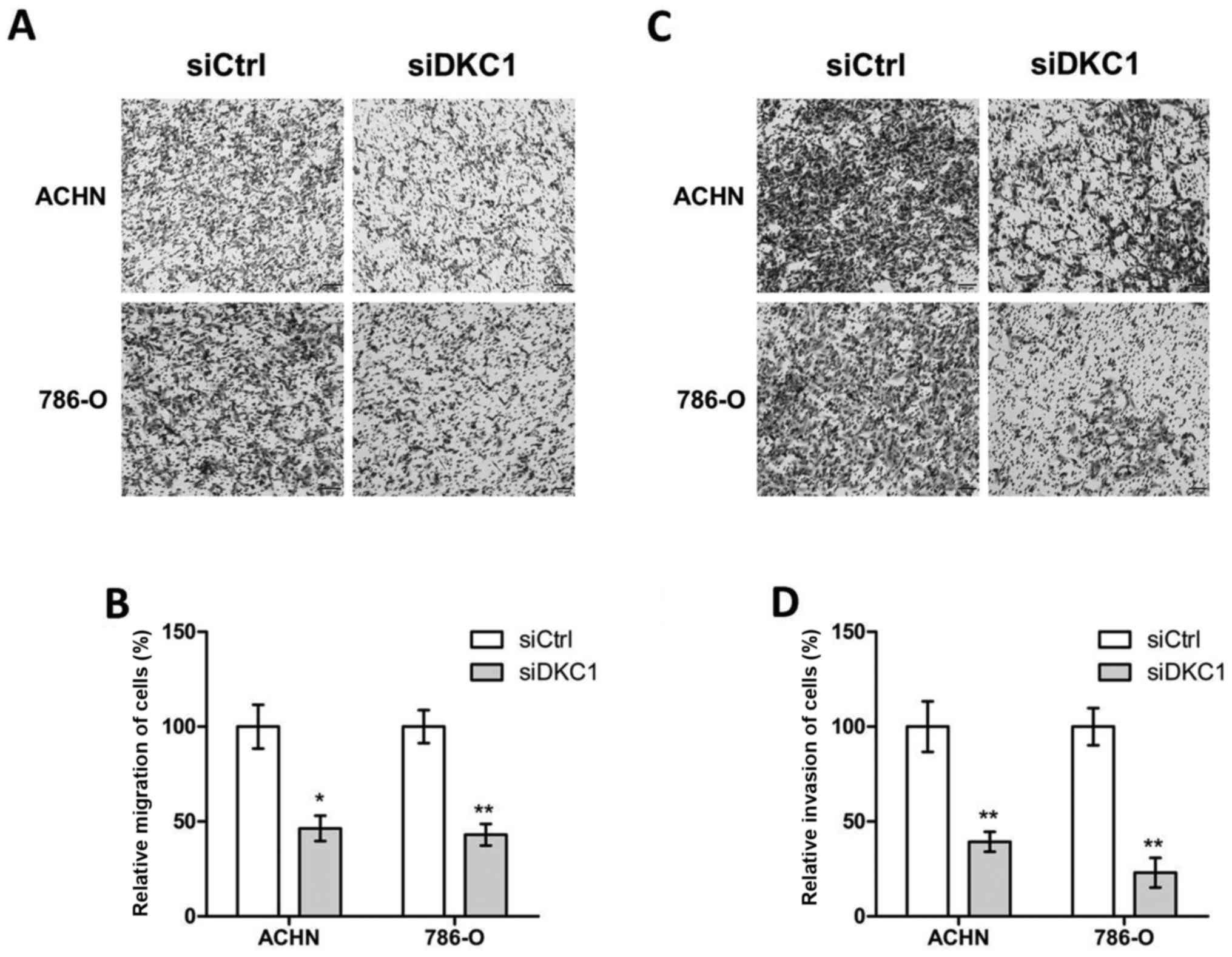
further explored the metastatic role of DKC1 in ccRCC cells. The
Transwell assays were performed and our data revealed that
knockdown of DKC1 decreased the abilities of cell migration in ACHN
and 786-O cells when compared with the corresponding controls
(Fig. 4A and B). In accordance with
these consequences, the abilities of cell invasion were
significantly decreased in ACHN and 786-O cells by DKC1 siRNAs when
compared with the respective controls (Fig. 4C and D).
Knockdown of DKC1 inhibits ccRCC cell
proliferation, migration and invasion via regulation of the
NF-κB/MMP-2 signaling pathway
NF-κB is a crucial transcriptional factor and plays
an important role in tumorigenesis. It regulates cell
proliferation, metastasis, angiogenesis and survival, thus it is
not surprising that NF-κB has been demonstrated to be activated in
many human cancers (15). Its κB
site was found in the promoters of genes encoding MMP-2 which plays
a critical role in cancer metastasis (16). In order to investigate the possible
mechanism of DKC1 regulation of proliferation and metastasis in
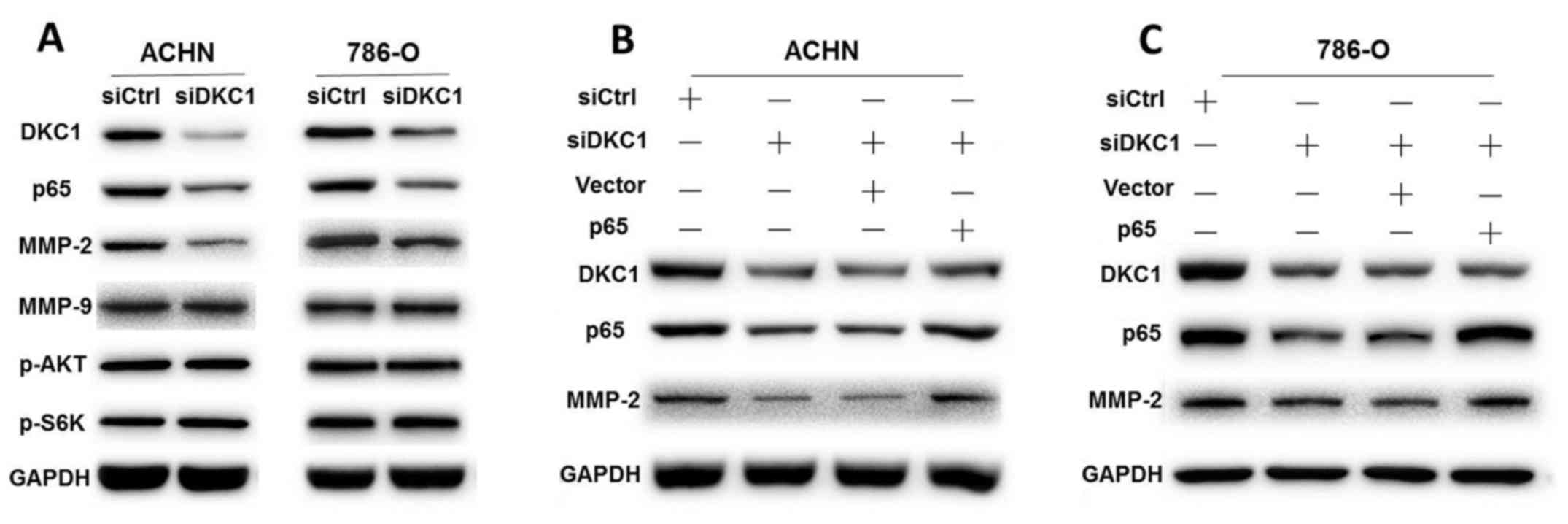
ccRCC cells, we carried out western blotting to explore the protein
levels of NF-κB and MMPs in ACHN and 786-O cells. Our data revealed
that DKC1 knockdown significantly inhibited p65 and MMP-2 protein
expression in ACHN and 786-O cells compared with the corresponding
controls, but not MMP-9 (Fig. 5A).
Since the PI3K/AKT pathway is also an important factor of cell
proliferation and survival (17),
we detected the expression levels of p-AKT and p-S6K. However, the
results revealed that the protein levels of p-AKT and p-S6K were
similar regardless of the expression levels of DKC1 (Fig. 5A).
To further investigate whether DKC1 regulated ccRCC
cell proliferation and metastasis though the NF-κB/MMP-2 signaling
pathway, p65 rescue assays were performed. We co-transfected p65
overexpression plasmids and DKC1 siRNAs in ACHN and 786-O cells,
respectively. Our data revealed that p65 overexpression
significantly rescued the expression of MMP-2 (Fig. 5B and C). The cell proliferation
assays revealed that the decreased abilities of cell proliferation
in the DKC1-knockdown ACHN and 786-O cells could be markedly
accelerated by p65 overexpression (Fig.
6A and B). Moreover, the results of cell migration and invasion
revealed that p65 overexpression markedly rescued the
DKC1-knockdown-decreased cell migration and invasion abilities
(Fig. 6C and D). These results
indicated that p65 and MMP-2 functioned as the downstream targets
of DKC1 in ccRCC cell proliferation, migration and invasion.
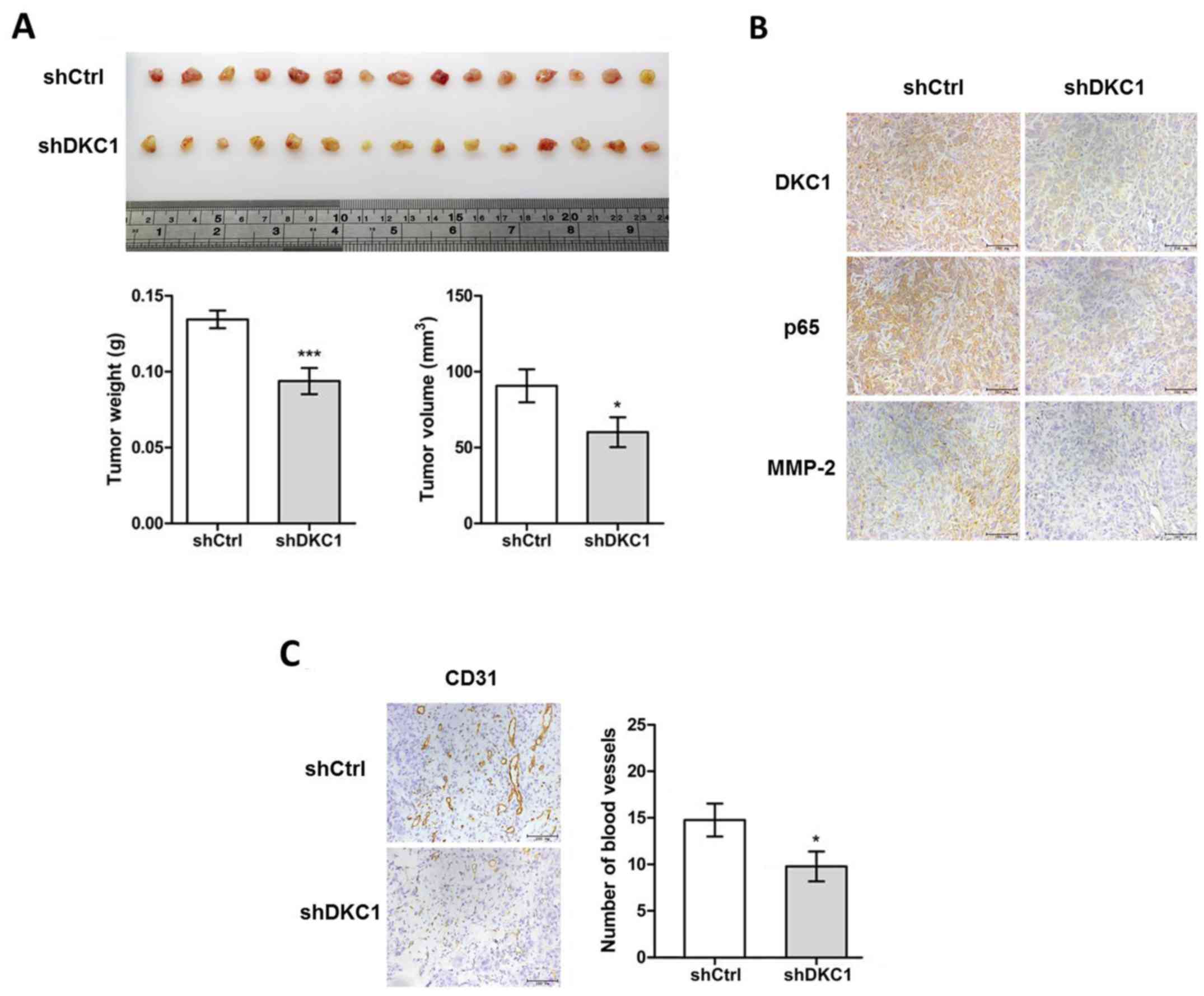
DKC1 accelerates the proliferation and
angiogenesis of ccRCC cells in vivo
To further validate the function of DKC1 in the
regulation of ccRCC proliferation in vivo,
DKC1KD-ACHN cell lines and Ctrl-ACHN cell lines were
established. The stable DKC1 knockdown and control ACHN cells were
subcutaneously injected into BALB/c nude mice. After 2 weeks, the
two groups of mice were sacrificed and their subcutaneous tumors
were excised (Fig. 7A) and
statistical analysis was performed on the differences in tumor
weight and volume between the two groups of mice. Our data revealed
that a significant decrease of tumor weight and volume was observed
in the DKC1KD group when compared with the control group
(Fig. 7A). Then, we carried out
immunohistochemical staining on paraffin-sectioned neoplastic
tissues. The data revealed that the expression of DKC1, p65 and
MMP-2 were markedly reduced in the knockdown group compared with
the control group (Fig. 7B). In
addition, the visual examination revealed that neovessels in the
subcutaneous tumors were reduced in the DKC1KD group
when compared with those in the control group (Fig. 7A). Previous research had
demonstrated that NF-κB promoted angiogenesis (18). To further validate the difference of
angiopoiesis between the two groups, we detected the expression of
CD31 in the two groups. The results revealed that the number of
vessels labeled by CD31 were significantly fewer in the
DKC1KD group than the control group (Fig. 7C). These results further confirmed
our conclusion which had been previously demonstrated in
vitro.
Discussion
Updated research has demonstrated that functional
genes which play a role in tumorigenesis can be regarded as
potential biomarkers for the diagnosis and prognosis of ccRCC
patients (19). DKC1, an X-linked
gene encoding dyskerin at Xq28, is a crucial ingredient of the
telomerase complex and is indispensable for normal telomere
function and the post-transcriptional modification of precursor
rRNA (20,21). It has been revealed to exert diverse
biological functions and have prognostic value in numerous types of
cancer (22–24). In the present study, we examined the
prognostic value and biological role of DKC1 in ccRCC. Using
retrospective cohorts of ccRCC patients with TMAs, we discovered
that DKC1 expression was significantly upregulated in ccRCC
compared with normal renal tissues, and positively associated with
TNM stage. Moreover, ccRCC patients with increased DKC1 expression
had an unfavorable survival, and multivariate COX regression
analysis revealed that positive DKC1 expression was an independent
hazardous indicator for the prognosis of ccRCC patients. Our
findings from survival analysis conformed with the data obtained in
The Cancer Genome Atlas (TCGA) collected from PROGgeneV2 platform.
These results revealed that DKC1 was a prognostic factor for ccRCC
patients and may act as an oncogene in ccRCC progression.
RCC is the predominant malignancy of kidney cancer
and is greatly resistant to chemotherapy and radiation. The status
of the p53 tumor-suppressor gene has been associated with the
efficacy of chemotherapy and radiation, where aberrant p53 function
can be attributed to defective responsiveness to treatment
(25,26). However, p53 is rarely mutated in
RCC, which suggests that other genes may be involved in the
regulation of tumorigenesis in RCC (27). To date, few studies have reported
the potential role of DKC1 in tumorigenesis and these results are
disputable. von Stedingk et al (28) revealed that a potential function of
DKC1 was to increase telomerase activity and contribute to advanced
tumors, suggesting an oncogenic role of DKC1. However, Montanaro
et al (29) reported that
low DKC1 expression was correlated with tumor progression, thus
revealing that DKC1 may serve as a tumor suppressor. The molecular
mechanism and significance of DKC1 in ccRCC progression are still
unclear. Our results demonstrated that DKC1 expression tends to be
associated with adverse clinicopathological characteristics in
ccRCC patients, which supports its role in tumor promotion.
Tumor development and progression require six
necessary changes to normal cell physiology: An independent growth
signaling pathway; sustained angiogenesis; resistance to growth
inhibition; escape of apoptosis; tissue invasion and metastasis;
and cell immortality (30). In the
present study, we investigated the role of DKC1 in several
important processes, such as proliferation, migration, invasion and
angiogenesis. Our data revealed that DKC1 expression regulated
ccRCC cell proliferation in vitro and in vivo, which
was consistent with our clinical data that DKC1 expression was
associated with tumor diameter. Moreover, we demonstrated that
knockdown of DKC1 markedly inhibited ccRCC cell migration and
invasion in vitro and angiogenesis in vivo. These
findings could explain our previous assertion that positive DKC1
expression was associated with enhanced lymph node metastases.
Nuclear factor of κB (NF-κB) is a transcriptional
factor and it has been most extensively studied for its function in
immunity and inflammation (31).
Until recent decades, NF-κB has been characterized by its critical
role in cancer development and progression and induces the
expression of MMPs (32,33). Matrix metalloproteinases (MMPs) can
break down the extracellular matrix (ECM) in numerous malignant
tumors and are crucial for metastasis-promoting genes (34). Except for their role in the
extracellular matrix and migration of cancer cells, MMPs also
regulate signaling pathways which control cell growth and
angiogenesis (35).
In the present study, we investigated whether DKC1
regulated ccRCC proliferation and migration via NF-κB and MMPs, and
we demonstrated that knockdown of DKC1 significantly inhibited the
protein expression of p65 and MMP-2, which may well account for our
proliferation, migration and invasion results. However, in our
study, MMP-9 expression did not appear to be activated by p65 in
ccRCC cells. The functions of p65 and MMP-2 in DKC1-mediated ccRCC
cell proliferation, migration and invasion were further explored by
co-transfection of both DKC1 siRNAs and p65 overexpression
plasmids, which rescued the decreased cell proliferation, migration
and invasion after DKC1 knockdown. Moreover, our subcutaneous tumor
model in vivo revealed that knockdown of DKC1 significantly
suppressed proliferation and angiogenesis of tumors and the
expression of p65, MMP-2 and CD31 in tumor tissues, which conformed
to the in vitro results and cohort of ccRCC patients.
Unfortunately, we did not explore the potential mechanism between
DKC1 and the NF-κB pathway, and therefore whether the regulation of
NF-κB by DKC1 is direct or indirect is not clear. However, we did
determine a possible relationship between them. Recently, it has
been established that DKC1 is a direct target of c-Myc in several
types of cancer (36). Thus, we
speculated that NF-κB could also directly activate DKC1 expression,
and p65 was regulated by DKC1 via negative feedback. In future
research, we will explore the potential molecular mechanisms
between DKC1 and the NF-κB signaling pathway in ccRCC cells
consistently.
In conclusion, DKC1 expression was markedly enhanced
in ccRCC compared with normal renal tissues. Positive DKC1
expression in ccRCC tissues was markedly associated with
unfavorable clinicopathological characteristics and a dismal
prognosis of patients, which can to some extent be explained by the
NF-κB/MMP-2 signaling pathway which regulated proliferation,
migration, invasion and angiogenesis of ccRCC cells. Therefore,
these results demonstrated that DKC1 may act as a significant
prognostic indicator and therapeutic target for ccRCC.
Acknowledgements
Not applicable.
Funding
The present study was funded by grants from the
National Natural Science Foundation of China (nos. 81472663,
81502280 and 81672845), the Education Department of Jiangsu
Province (no. 15KJA320006) and the Project of Invigorating Health
Care through Science, Technology and Education from Jiangsu
Province.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
MZ, JB and YP conceived and designed the
experiments; YP, RJ and FC conducted the experiments; MZ and PH
carried out the statistical analysis; HS and TJ supported the
experiments and helped to draft the manuscript. All authors have
read and approved the manuscript and agree to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
Informed consents from all patients were obtained
and institutional approval was obtained by the Review Board of the
Affiliated Hospital of Xuzhou Medical University prior to this
study. The animal studies were approved by the Animal Care
Committee of Xuzhou Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ccRCC
|
clear cell renal cell carcinoma
|
|
IHC
|
immunohistochemistry
|
|
NF-κB
|
nuclear factor κB
|
|
MMPs
|
matrix metalloproteinases
|
|
TMA
|
tissue microarray
|
|
IRS
|
immunoreactive score
|
|
TCGA
|
The Cancer Genome Atlas
|
|
GAPDH
|
glyceraldehyde-3-phosphate
dehydrogenase
|
|
ECM
|
extracellular matrix
|
|
HR
|
hazard ratio
|
|
CI
|
confidence interval
|
References
|
1
|
Volpe A and Patard JJ: Prognostic factors
in renal cell carcinoma. World J Urol. 28:319–327. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rini BI, Campbell SC and Escudier B: Renal
cell carcinoma. Lancet. 373:1119–1132. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lu X, Gu W, Zhang H, Zhu Y, Shi G and Ye
D: Oligometastatic state predicts a favorable outcome for renal
cell carcinoma patients with bone metastasis under the treatment of
sunitinib. Oncotarget. 7:26879–26887. 2016.PubMed/NCBI
|
|
4
|
Singer EA, Gupta GN and Srinivasan R:
Update on targeted therapies for clear cell renal cell carcinoma.
Curr Opin Oncol. 23:283–289. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Heiss NS, Knight SW, Vulliamy TJ, Klauck
SM, Wiemann S, Mason PJ, Poustka A and Dokal I: X-linked
dyskeratosis congenita is caused by mutations in a highly conserved
gene with putative nucleolar functions. Nat Genet. 19:32–38. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kirwan M and Dokal I: Dyskeratosis
congenita: A genetic disorder of many faces. Clin Genet.
73:103–112. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sieron P, Hader C, Hatina J, Engers R,
Wlazlinski A, Müller M and Schulz WA: DKC1 overexpression
associated with prostate cancer progression. Br J Cancer.
101:1410–1416. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
O'Brien R, Tran SL, Maritz MF, Liu B, Kong
CF, Purgato S, Yang C, Murray J, Russell AJ, Flemming CL, et al:
MYC-Driven neuroblastomas are addicted to a telomerase-independent
function of dyskerin. Cancer Res. 76:3604–3617. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu B, Zhang JL, Huang C and Liu H:
Dyskerin overexpression in human hepatocellular carcinoma is
associated with advanced clinical stage and poor patient prognosis.
PloS one. 7:e431472012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bellodi C, Krasnykh O, Haynes N,
Theodoropoulou M, Peng G, Montanaro L and Ruggero D: Loss of
function of the tumor suppressor DKC1 perturbs p27 translation
control and contributes to pituitary tumorigenesis. Cancer Res.
70:6026–6035. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Montanaro L, Calienni M, Bertoni S, Rocchi
L, Sansone P, Storci G, Santini D, Ceccarelli C, Taffurelli M,
Carnicelli D, et al: Novel dyskerin-mediated mechanism of p53
inactivation through defective mRNA translation. Cancer Res.
70:4767–4777. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mei P, Bai J, Shi M, Liu Q, Li Z, Fan Y
and Zheng J: BRMS1 suppresses glioma progression by regulating
invasion, migration and adhesion of glioma cells. PloS One.
9:e985442014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bai J, Yong HM, Chen FF, Song WB, Li C,
Liu H and Zheng JN: RUNX3 is a prognostic marker and potential
therapeutic target in human breast cancer. J Cancer Res Clin Oncol.
139:1813–1823. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bai J, Zhou Y, Chen G, Zeng J, Ding J, Tan
Y, Zhou J and Li G: Overexpression of Cullin1 is associated with
poor prognosis of patients with gastric cancer. Hum Pathol.
42:375–383. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Karin M: Nuclear factor-kappaB in cancer
development and progression. Nature. 441:431–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Takeshita H, Yoshizaki T, Miller WE, Sato
H, Furukawa M, Pagano JS and Raab-Traub N: Matrix metalloproteinase
9 expression is induced by Epstein-Barr virus latent membrane
protein 1 C-terminal activation regions 1 and 2. J Virol.
73:5548–5555. 1999.PubMed/NCBI
|
|
17
|
Cully M, You H, Levine AJ and Mak TW:
Beyond PTEN mutations: The PI3K pathway as an integrator of
multiple inputs during tumorigenesis. Nat Rev Cancer. 6:184–192.
2006. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang S, Robinson JB, Deguzman A, Bucana
CD and Fidler IJ: Blockade of nuclear factor-kappaB signaling
inhibits angiogenesis and tumorigenicity of human ovarian cancer
cells by suppressing expression of vascular endothelial growth
factor and interleukin 8. Cancer Res. 60:5334–5339. 2000.PubMed/NCBI
|
|
19
|
Audenet F, Yates DR, Cancel-Tassin G,
Cussenot O and Roupret M: Genetic pathways involved in
carcinogenesis of clear cell renal cell carcinoma: Genomics towards
personalized medicine. BJU Int. 109:1864–1870. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mochizuki Y, He J, Kulkarni S, Bessler M
and Mason PJ: Mouse dyskerin mutations affect accumulation of
telomerase RNA and small nucleolar RNA, telomerase activity, and
ribosomal RNA processing. Proc Natl Acad Sci USA. 101:10756–10761.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Filipowicz W and Pogacic V: Biogenesis of
small nucleolar ribonucleoproteins. Curr Opin Cell Boil.
14:319–327. 2002. View Article : Google Scholar
|
|
22
|
Westermann F, Henrich KO, Wei JS, Lutz W,
Fischer M, König R, Wiedemeyer R, Ehemann V, Brors B, Ernestus K,
et al: High Skp2 expression characterizes high-risk neuroblastomas
independent of MYCN status. Clin Cancer Res. 13:4695–4703. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schaner ME, Ross DT, Ciaravino G, Sorlie
T, Troyanskaya O, Diehn M, Wang YC, Duran GE, Sikic TL, Caldeira S,
et al: Gene expression patterns in ovarian carcinomas. Mol Boil
Cell. 14:4376–4386. 2003. View Article : Google Scholar
|
|
24
|
Poncet D, Belleville A, t'kint de
Roodenbeke C, Roborel de Climens A, Ben Simon E, Merle-Beral H,
Callet-Bauchu E, Salles G, Sabatier L, Delic J and Gilson E:
Changes in the expression of telomere maintenance genes suggest
global telomere dysfunction in B-chronic lymphocytic leukemia.
Blood. 111:2388–2391. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lowe SW: Cancer therapy and p53. Curr Opin
Oncol. 7:547–553. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lowe SW, Bodis S, McClatchey A, Remington
L, Ruley HE, Fisher DE, Housman DE and Jacks T: p53 status and the
efficacy of cancer therapy in vivo. Science. 266:807–810. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tomasino RM, Morello V, Tralongo V, Nagar
C, Nuara R, Daniele E, Curti M and Orestano F: p53 expression in
human renal cell carcinoma: An immunohistochemical study and a
literature outline of the cytogenetic characterization.
Pathologica. 86:227–233. 1994.PubMed/NCBI
|
|
28
|
von Stedingk K, Koster J, Piqueras M,
Noguera R, Navarro S, Påhlman S, Versteeg R, Ora I, Gisselsson D,
Lindgren D and Axelson H: snoRNPs regulate telomerase activity in
neuroblastoma and are associated with poor prognosis. Transl Oncol.
6:447–457. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Montanaro L, Brigotti M, Clohessy J,
Barbieri S, Ceccarelli C, Santini D, Taffurelli M, Calienni M,
Teruya-Feldstein J, Trerè D, et al: Dyskerin expression influences
the level of ribosomal RNA pseudo-uridylation and telomerase RNA
component in human breast cancer. J Pathol. 210:10–18. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Karin M, Cao Y, Greten FR and Li ZW:
NF-kappaB in cancer: From innocent bystander to major culprit. Nat
Rev Cancer. 2:301–310. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Uhlik M, Good L, Xiao G, Harhaj EW, Zandi
E, Karin M and Sun SC: NF-kappaB-inducing kinase and IkappaB kinase
participate in human T-cell leukemia virus I Tax-mediated NF-kappaB
activation. J Biol Chem. 273:21132–21136. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kumar A, Takada Y, Boriek AM and Aggarwal
BB: Nuclear factor-kappaB: Its role in health and disease. J Mol
Med (Berl). 82:434–448. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim A, Kim MJ, Yang Y, Kim JW, Yeom YI and
Lim JS: Suppression of NF-kappaB activity by NDRG2 expression
attenuates the invasive potential of highly malignant tumor cells.
Carcinogenesis. 30:927–936. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: Regulators of the tumor microenvironment. Cell.
141:52–67. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Alawi F and Lee MN: DKC1 is a direct and
conserved transcriptional target of c-MYC. Biochem Biophys Res
Commun. 362:893–898. 2007. View Article : Google Scholar : PubMed/NCBI
|