Introduction
Gastric cancer (GC) is one of the most common
malignant tumors with a high mortality rate. There are no specific
clinical symptoms or signs in the early stage of GC. Therefore, the
majority of cases of GC are diagnosed at moderate or advanced
stages (1). The five-year overall
survival rate of GC is <30% in the worldwide (2). There are several factors that
influence the prognosis of GC, and its invasion and metastasis.
Chemotherapy is an important method in the comprehensive treatment
for GC (3). However, the presence
of multi-drug resistance (MDR) of GC often leads to the failure of
chemotherapy, which is a main contributor to mortality rates in
patients with GC (4). Therefore,
reversing the MDR of GC offers potential for significant
enhancement of the effect of chemotherapy and improvement of
prognosis (5). In the present
study, the molecular mechanism underlying the MDR of GC, and the
correlation between differentiation and MDR in GC were examined.
The aim was to provide a therapeutic target for GC.
MicroRNAs are a group of small, non-coding RNAs,
which are 21–23 nucleotides in length. MicroRNAs can regulate
downstream gene expression as post-transcriptional regulators.
Mature microRNAs can bind to the 3′-untranslated region (UTR) of
target gene mRNAs and negatively regulate protein expression. It
has been reported that microRNAs are involved in several biological
processes, including cell proliferation, differentiation and
apoptosis, and aberrant microRNA expression can attribute to the
pathogenesis of GC. For example, the upregulation of microRNA
(miR)-185 can promote GC cell apoptosis via regulating B-cell
lymphoma 2 (Bcl-2), survivin and X-linked inhibitor of apoptosis
(6). miR-320 acts as a suppressor
of GC cell proliferation and metastasis via the Eph receptor
A2/Wnt/b-catenin/epithelial-mesenchymal transition signal pathway
(7). miR-340 and miR-124 act as
tumor-suppressive factors by regulating SLIT-ROBO Rho GTPase
activating protein 1 in GC cells (8). miR-155 regulates cell growth and
migration by negatively regulating transforming growth factor-β
receptor 2 in GC cells (9). miR-204
can negatively regulate Cyclin-dependent kinase regulatory subunit
1, Chemokine (C-X-C motif) ligand 1 and G protein-coupled receptor
class C group 5 member A, and suppress the proliferation of GC
cells (10). miR-195-5p belongs to
the miR-15 family. It has been reported that miR-195-5p is involved
in various types of cancer, including thyroid cancer,
hepatocellular cancer and breast cancer (11–16).
However, the effects of miR-195-5p on the MDR of GC cells remains
to be elucidated. In the present study, it was determined that
miR-195-5p regulated the MDR of GC cells by targeting Zing finger
139 (ZNF139).
Materials and methods
Gastric adenocarcinoma tissue
specimens
A total of 12 tissue samples of gastric
adenocarcinoma were collected at the Fourth Hospital of Hebei
Medical University (Hebei, China) between April 2014 and 2015. The
patients included six male patients and six female patients (age
38–78 years, average age 60 years). Histopathologic features of the
tissue samples were determined by hematoxylin and eosin staining,
and histological grades of tumor samples were assigned according to
the World Health Organization classification criteria (17). All gastric adenocarcinoma tissues
were divided into two groups, the well differentiated group and
poorly differentiated group. Each group contained six samples. All
tissue samples were stored in 4% paraform at −80°C.
Cell culture
The MKN28 GC cell line, a derivative of the MKN74
cell line (18), was used, which
was from the Central Laboratory of the Fourth Hospital of Hebei
Medical University. All cells were cultured in Dulbecco's modified
Eagle's medium supplemented with 10% fetal calf serum (Sigma; EMD
Millipore, Billerica, MD, USA), 80 U/ml penicillin and 0.1 mg/ml
streptomycin (Thermo Fisher Scientific, Inc., Waltham, MA, USA) in
a 5% CO2 humidified atmosphere at 37°C.
miR-195-5p mimic and inhibitor
transfection
The miR-195-5p mimic and inhibitor were synthesized
by GenePharm, Inc. (Sunnyvale, CA, USA). The sequences were as
follows: miR-195-5p mimic, sense 5′-UAGCAGCACAGAAAUAUUGGC-3′ and
miR-195-5p mimic, antisense 5′-CAAUAUUUCUGUGCUGCUAUU-3′; miR-195-5p
inhibitor: 5′-GCCAAUAUUUCUGUGCUGCUA-3′. The miR-195-5p mimic and
inhibitor were transfected into cells using Lipofectamine™ 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. At 2 day pre-transfection, the cells
were cultured in a 6-well-plate at 1.0×105 cells per
well. In brief, 20 ng miRNA oligo was diluted in 50 µl Opti-MEM
medium, and 2 µl Lipofectamine™ 2000 reagent was diluted in 50 µl
Opti-MEM reduced serum medium. Following gentle mixing and
incubation at room temperature for 5 min, the two mixtures were
combined and incubated for 20 min. The mixture was added into each
well and the cells were incubated at 37°C. The medium was replaced
6 h later and incubated for 44 h prior to harvesting cells.
Quantification of miR-19a levels by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) analysis
Total RNA from cells and tissues were isolated using
TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol. RNA (1 µg) was reverse-transcribed
into cDNA using the EasyScript First-strand cDNA Synthesis kit
(Beijing Transgen Biotech Co., Ltd., Beijing, China). The analysis
of miR-195-5p levels was performed by qPCR analysis according the
protocol of the SYBR-Green II kit (Takara Biotechnology Co., Ltd.,
Dalian, China). cDNA (1 µl), 10 µl SYBR Green mixture, 0.8 µl
primers (10 µM) and 8.2 µl ddH2O were mixed. The
procedures of PCR were described as follow: 95°C for 30 sec,
followed by 40 cycles at 95°C for 5 sec and 60°C for 20 sec. The
relative level of miR-195-5p was normalized by U6 small nucleolar
RNA, which was used as the housekeeping gene. The gene expression
level was calculated using the 2−ΔΔCq method (19). All analyses were performed in
triplicate. The primers were designed using Primer Premier 5
(Premier Biosoft International, Palo Alto, CA, USA. The sequences
of the RT primers were as follows: miR-195-5p,
5′-TTCCGATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTCAGTT-3′; U6,
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAAAAATATG-3′. The
nucleotide primers used for qPCR were as follows: miR-195-5p,
forward, 5′-GCGATAGCAGCACAGAAATA-3′; U6, forward
5′-GCGCGTCGTGAAGCGTTC-3′ and universal reverse,
5′-GTGCAGGGTCCGAGGT-3′.
Western blot analysis
The total protein from tissues and cells was
extracted with RIPA (Beijing Solarbio Science & Technology Co.,
Ltd., Beijing, China). The protein concentration was measured using
a BCA kit (Thermo Fisher Scientific, Inc.). The proteins (15 µg)
from each sample were separated by 10% SDS-PAGE and
electrotransferred onto a polyvinylidene fluoride (PVDF) membrane
(EMD Millipore). Then membranes were blocked with 5% non-fat milk
in TBST at room temperature for 2 h, and then incubated with the
following 1:1,000 diluted primary antibodies at 4°C for 12–18 h:
ZNF139 (cat. no. ab32124; Abcam, Cambridge, UK), P-glycoprotein
(P-gp; cat. no. 13978; Cell Signaling Technology, Inc., Danvers,
MA, USA), Multi-drug resistance-associated protein 1 (MRP1; cat.
no. 14182; Cell Signaling Technology, Inc.), Bcl-2-associated ×
protein (Bax; cat. no. 2772; Cell Signaling Technology, Inc.),
Bcl-2 (cat. no. 3498; Cell Signaling Technology, Inc.), or GAPDH
(cat. no. 5174; Cell Signaling Technology, Inc.). Following washing
with TBST five times, the membranes were incubated with 1:10,000
diluted horseradish peroxidase-conjugated secondary antibody (cat.
no. 7074; Cell Signaling Technology, Inc.) at room temperature for
2 h. Following washing with TBST five times, the specific bands
were detected with an enhanced chemiluminescence detection system
(EMD Millipore). GAPDH was used as the housekeeping gene to
normalized other genes. The quantification of protein was performed
by using ImageJ version 1.42 (National Institutes of Health,
Bethesda, MD, USA). The relative protein levels were calculated as
the ration of treatment group to control group. The experiments
were performed in triplicate.
Luciferase reporter assay
It was predicted that there was a binding site of
miR-195-5p on the 3′-UTR of ZNF139 using miRNA target prediction
databases, including miRanda (http://www.microrna.org/), TargetScan (http://www.targetscan.org/) and PicTar (http://www.pictar.org/). The fragment of ZNF139 3′-UTR
containing the binding site of miR-195-5p was amplified from human
gastric adenocarcinoma cells using the following primers: znf139,
forward 5′-ATGAGCTCAGAAGAATTTGCCATCAAGCC-3′
(SacI); znf139, reverse 5′-ATTCTAGAGTTCTGATCTCTGGGATGAGGAG-3′
(Xbal). The pmiRGLO vector was purchased from Promega
Corporation (Madison, WI, USA). The PCR product was digested with
SacI and XbaI endonuclease. The fragment was purified
and inserted into the pmiRGLO vector (pmiRGLO-ZNF139). On the day
prior to transfection, the MKN28 cells were seeded into a 96-well
plate at 5,000 cells/well in 100 µl of DMEM medium. The MKN28 cells
were co-transfected with the pmiRGLO vector, pmiRGLO-ZNF139 and a
miR-195-5p mimic (or negative control) using Lipofectamine™ 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) for 48 h.
Luciferase activity was assessed by using the Dual-Glo Luciferase
assay system (Promega Corp.). All the assays were performed with
six repeats.
MTT assay
At 24 h prior to transfection, the MNK28 cells were
seeded into a 96-well plate at 1×104 cells/well. At 24 h
post-transfection, 20 µl MTT (Sigma; EMD Millipore) assay solution
(5 mg/ml) was added into the wells for 4 h at 37°C. The supernatant
medium was then discarded, and 150 µl dimethyl sulfoxide was added.
The absorbance was measured at 490 nm on a microplate reader
(Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Chemosensitivity of cancer cells
The MNK28 cells (1×104 per well) were
cultured in a 96-well plate. The cells were treated with 50 µM
5-fluorouracil (5-FU; Sigma; EMD Millipore) or 10 µM oxaliplatin
(L-OHP; Sigma; EMD Millipore) for 24 h at 37°C. The cell viability
was then determined using the MTT assay. The MNK28 cells
(8×105 per well) were cultured in a 6-well plate. The
miR-195-5p mimic and inhibitor were transfected into MNK28 cells
for 48 h, followed by treatment with 50 µM 5-FU or 10 µM L-OHP for
24 h. The cells were harvested and total protein was extracted for
western blot analysis.
Statistical analysis
Statistical analysis was performed using the SPSS
13.0 statistical software package (SPSS, Inc., Chicago, IL, USA).
Data are presented as the mean ± standard error of the mean. The
non-parametric Mann-Whitney U test was used to compare two groups.
One-way analysis of variance followed by Tukey's post hoc test was
used to compare three or more groups. P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression of miR-195-5p is decreased
in poorly differentiated GC tissues with higher
chemosensitivity
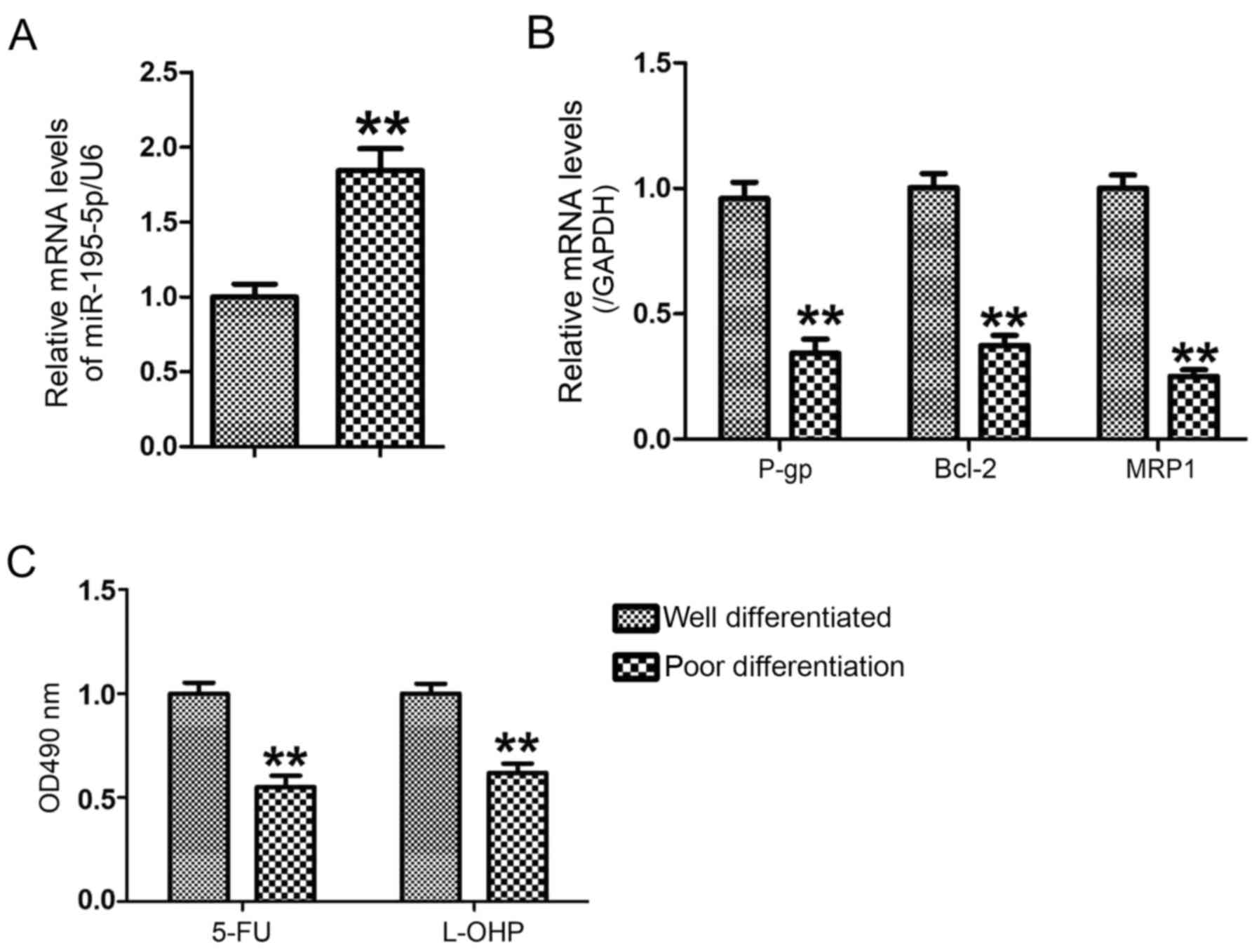
To analyze the changes of miR-195-5p between well
differentiated GC tissues and poorly differentiated GC tissues, the
levels of miR-195-5p in well differentiated GC tissues (n=6) and
poorly differentiated GC tissues (n=6) were measured using RT-qPCR
analysis. Compared with the well differentiated GC tissues, the
expression of miR-195-5p was significantly upregulated in the
poorly differentiated GC tissues (P<0.01; Fig. 1A), accompanied by decreased mRNA
levels of P-gp, MRP1 and Bcl-2 (Fig.
1B). The GC cancer cells were separated into well
differentiated GC tissues and poorly differentiated GC tissues,
respectively. The results of the MTT assay showed that the poorly
differentiated GC cells were more sensitive to 5-FU and L-OHP than
the well differentiated GC cells (Fig.
1C). This result suggested that decreased miR-195-5p may be
associated with the chemosensitivity of GC cells.
miR-195-5p regulates the MDR of GC
cells
In order to further investigate the impact of
miR-195-5p on the MDR of GC cells, miR-195-5p mimic and miR-195-5p
inhibitor were transfected into MNK28 cells for 48 h, respectively.
The expression of miR-195-5p was increased ~10-fold in the MNK28
cells transfected with miR-195-5p mimic, whereas the level of
miR-195-5p was reduced by 60% in the MNK28 cells transfected with
the miR-195-5p inhibitor (Fig. 2A and
B). In addition, the overexpression of miR-195-5p reduced the
protein levels of P-gp, MRP1 and Bcl-2 (Fig. 2C). By contrast, the downregulation
of miR-195-5p induced the expression of P-gp, MRP1 and Bcl-2
(Fig. 2D). To determine the effect
of miR-195-5p on the chemosensitivity of GC cells, the MKN28 cells
were transfected with miR-195-5p mimic and miR-195-5p inhibitor,
respectively for 48 h, followed by treatment with 5-FU and L-OHP,
respectively, for 24 h. The results of the MTT assay showed that
the survival rate of the miR-195-5p mimic-transfected cells was
lower than that of the negative control miRNA mimic-transfected
cells (Fig. 2E). The survival rate
of the miR-195-5p inhibitor cell group was higher than that of the
negative control miRNA inhibitor group (Fig. 2F). These results suggested that
miR-195-5p regulated the MDR of MNK28 cells.
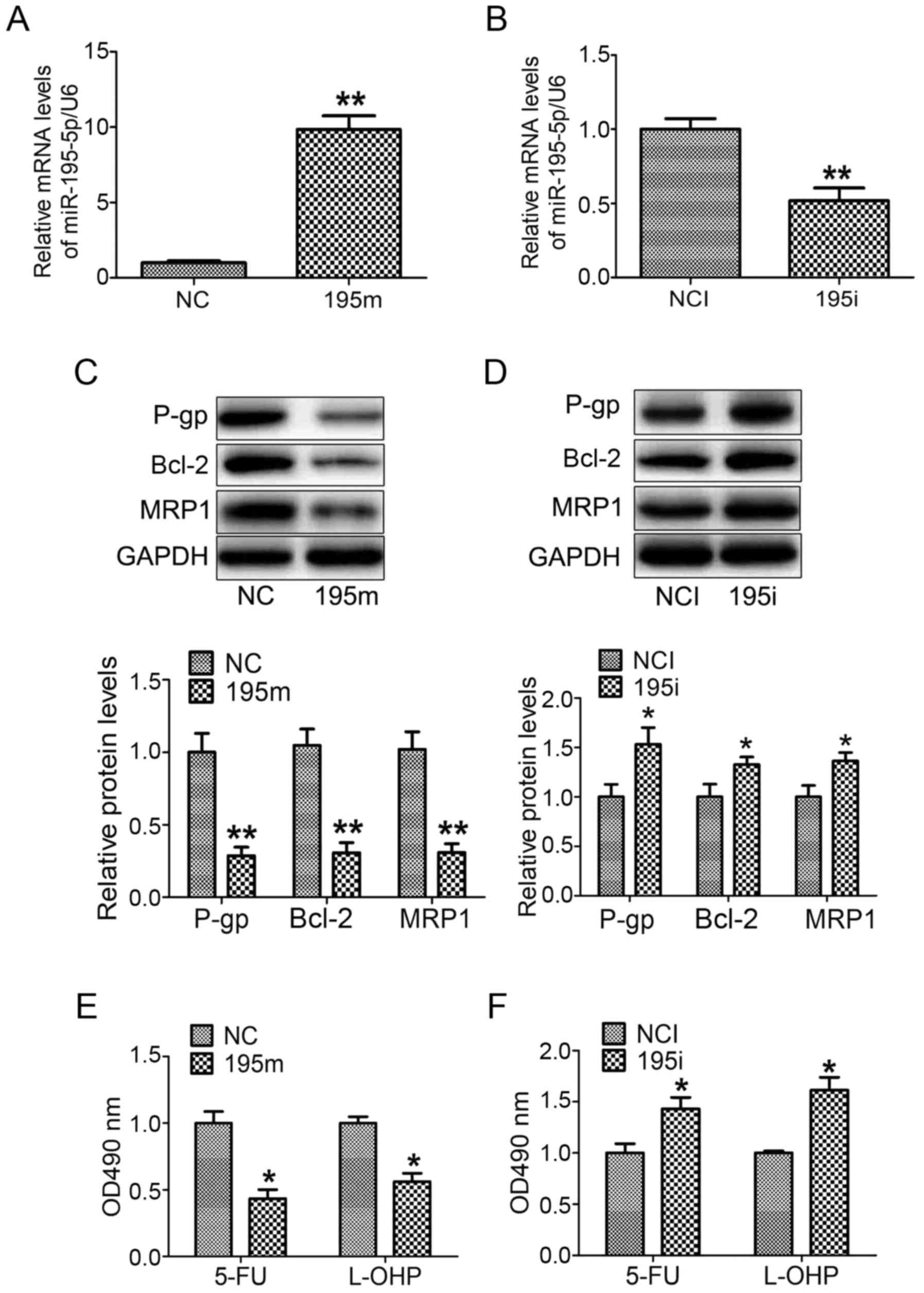 | Figure 2.miR-195-5p regulates the multi-drug
resistance of gastric cancer cells. The levels of miR-195-5p were
analyzed in MNK28 cells transfected with (A) miR-195-5p mimic and
(B) miR-195-5p inhibitor. The protein levels of P-gp, BCL-2 and
MRP1 were measured by western blot analysis in MNK28 cells
transfected with (C) miR-195-5p mimic and (D) miR-195-5p inhibitor.
The chemosensitivities of 5-FU and L-OHP were determined by MTT in
MNK28 cells transfected with (E) miR-195-5p mimic and (F)
miR-195-5p inhibitor. *P<0.05 and **P<0.01, vs. control
group. miR, microRNA; P-gp, P-glycoprotein; BCL-2, B-cell lymphoma
2; MRP1, Multi-drug resistance-associated protein 1; 5-FU,
5-fluorouracil; L-OHP, oxaliplatin; NC, negative control; 195m,
miR-195-5p mimic; NCI, negative control inhibitor; 195i, miR-195-5p
inhibitor. |
miR-195-5p negatively regulates the
expression of ZNF139 by binding to its 3′-UTR
It was predicted that ZNF139 was a target gene of
miR-195-5p by the MiRanda, TargetScan and PicTar miRNA databases.
There was a binding site of miR-195-5p at the ZNF139 3′-UTR
(Fig. 3A). The fragment of ZNF139
3′-UTR was then amplified and inserted into a pmiRLGO vector to
construct a recombinant vector. The nucleotides of the binding site
were mutated as shown in Fig. 3B.
As shown in Fig. 3C, the miR-195-5p
mimic significantly decreased the luciferase activity only in the
MNK28 cells transfected with pmiRLGO vector containing the
wild-type ZNF139 3′-UTR. The miR-195-5p mimic did not affect the
luciferase activity in the MNK28 cells transfected with pmiRLGO
vector containing the mutated ZNF139 3′-UTR. In addition,
transfection with the miR-195-5p mimic decreased the protein
expression of ZNF139 in MNK28 cells (Fig. 3D). By contrast, the miR-449a
inhibitor upregulated the protein expression of ZNF139 (Fig. 3E). These results suggested that
miR-195-5p negatively regulated the expression of ZNF139 by binding
to its 3′-UTR.
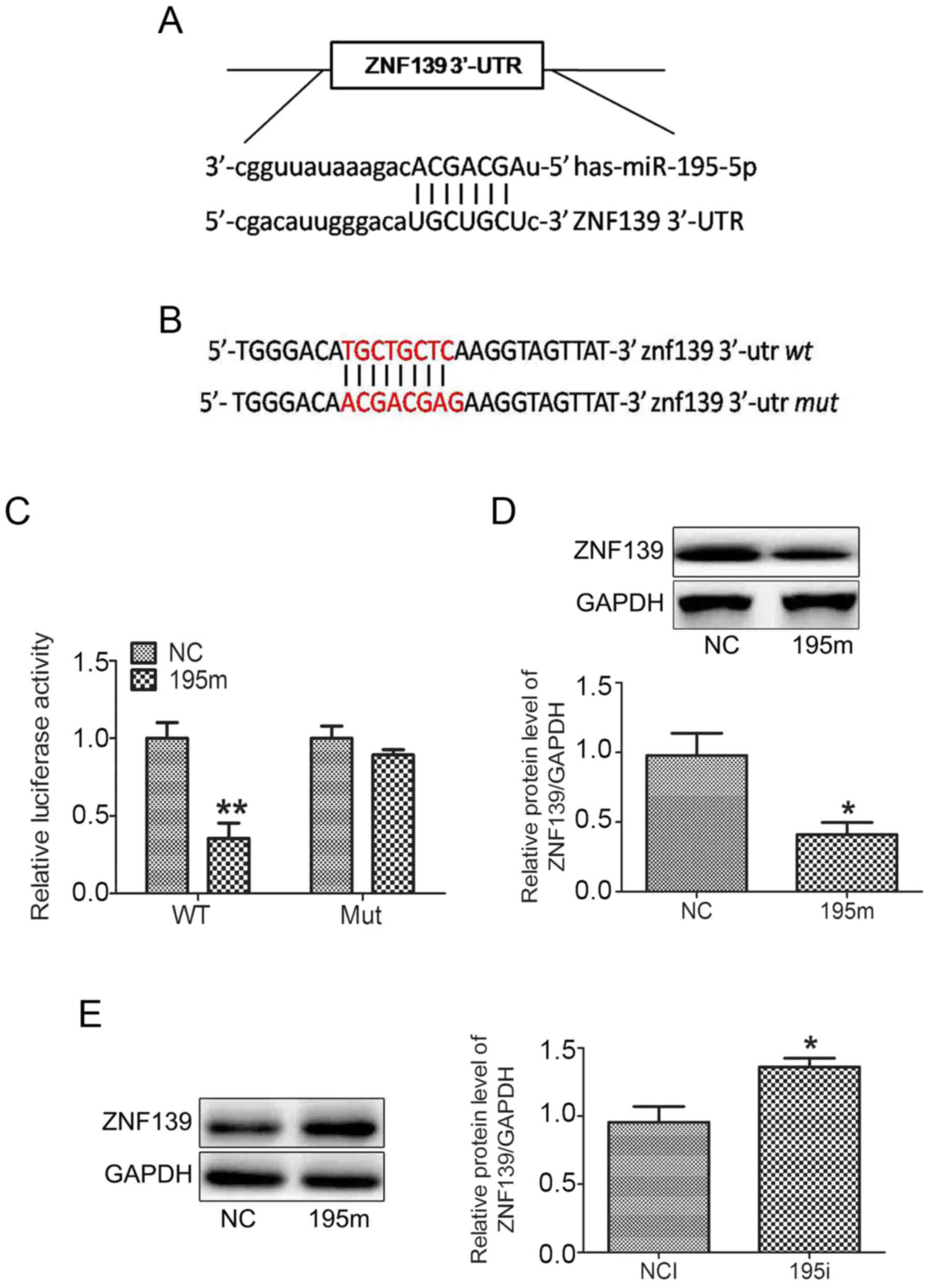 | Figure 3.miR-195-5p negatively regulates the
expression of ZNF139 by binding to its 3′-UTR. (A) Bioinformatics
database prediction of the binding site of miR-195-5p on the ZNF139
3′-UTR. (B) Nucleotides of the binding site were mutated. (C)
Luciferase assay confirmed that miR-195-5p was able to bind to the
3′-UTR of ZNF139. The protein levels of ZNF139 were determined by
western blot analysis in in MNK28 cells transfected with (D)
miR-195-5p mimic and (E) miR-195-5p inhibitor. *P<0.05, vs.
control group. miR, microRNA; ZNF139, Zing finger 139; 3′-UTR,
3′-untranslated region; WY, wild-type; Mut, mutated; NC, negative
control; 195m, miR-195-5p mimic; NCI, negative control inhibitor;
195i, miR-195-5p inhibitor. |
Silencing of ZNF139 promotes the
chemosensitivity of GC cells
In our previous studies, it was found that ZNF139
was involved in the progression of GC (20,21).
In order to determine the effect of ZNF139 on the chemosensitivity
of GC cells, three pairs of siRNAs specifically targeting ZNF139
were selected. The results of the western blot analysis showed that
only siRNA-3 effectively inhibited the protein expression of ZNF139
(Fig. 4A). Silencing of ZNF139
reduced the protein levels of P-gp, MRP1 and Bcl-2 (Fig. 4B), and the chemosensitivity of MNK28
cells was enhanced via the downregulation of ZNF139 (Fig. 4C).
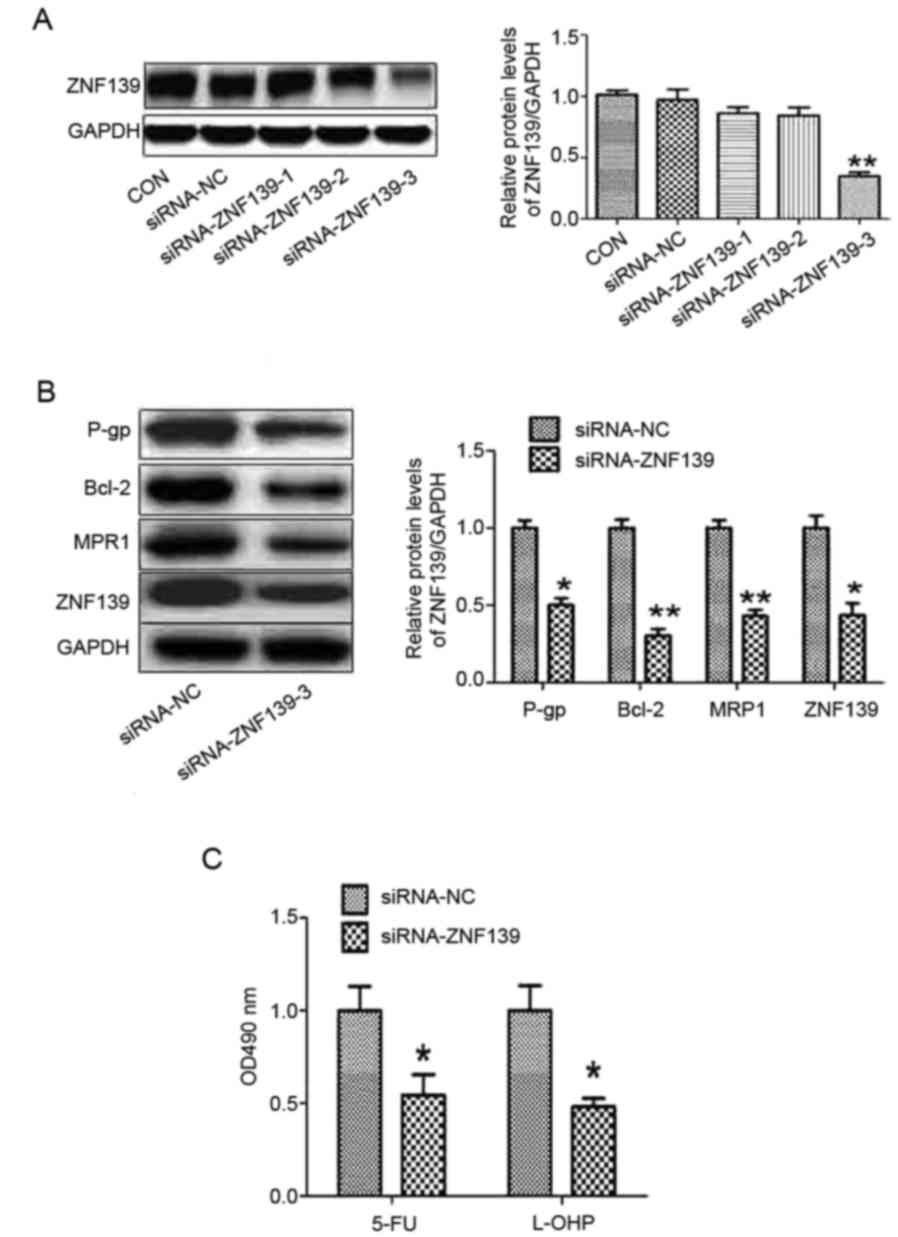 | Figure 4.Silencing of ZNF 139 promotes the
chemosensitivity of gastric cancer cells. (A) Three pairs of siRNAs
targeting ZNF139 were designed. (B) Protein levels of P-gp, BCL-2,
MRP1 and ZNF139 were measured using western blot analysis in MNK28
cells transfected with siRNA-ZNF139. (C) Chemosensitivities of 5-FU
and L-OHP were determined by MTT in MNK28 cells transfected with
siRNA-ZNF139. *P<0.05 and **P<0.01, vs. control group. miR,
microRNA; ZNF139, Zing finger 139; p-gp, p-glycoprotein; bcl-2,
B-cell lymphoma 2; mrp1, multi-drug resistance-associated protein
1; 5-FU, 5-fluorouracil; L-OHP, oxaliplatin; CON, control; siRNA,
small interfering RNA; NC, negative control. |
miR-195-5p regulates the MDR of GC
cells via targeting ZNF139
As it was suggested that ZNF139 may regulate the MDR
of GC cells, in order to further determine the role of ZNF139 in
the miR-195-5p inhibitor-induced MDR of MNK28 cells, siRNA
targeting ZNF139 and miR-449a inhibitor was co-transfected into
MNK28 cells. The silencing of ZNF139 reversed the effects of the
miR-195-5p inhibitor on the protein levels of P-gp, MRP1 and Bcl-2,
and the chemosensitivity of the MNK28 cells (Fig. 5A and B). These results demonstrated
that miR-195-5p regulated the MDR of MNK28 cells via targeting
ZNF139.
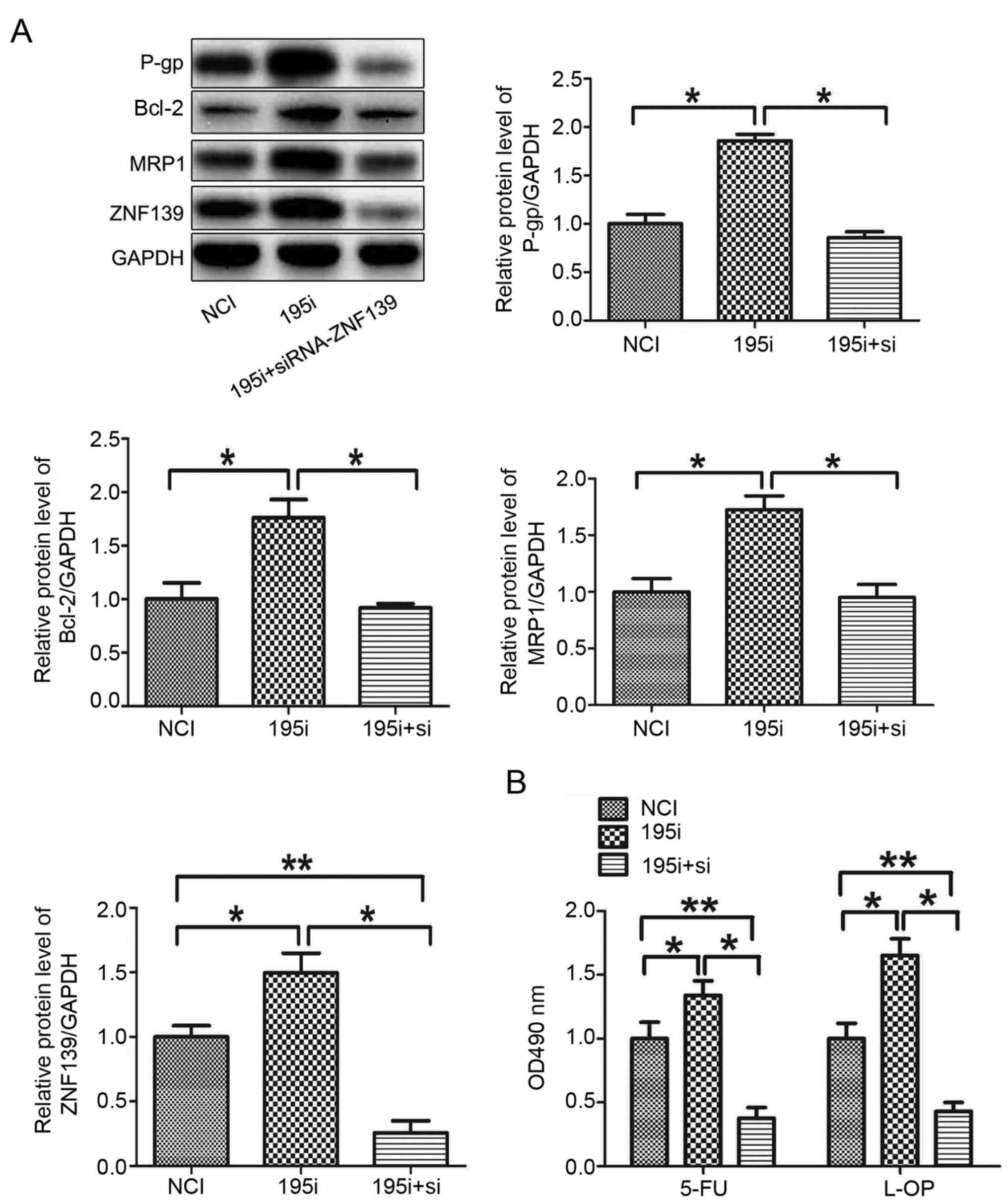 | Figure 5.miR-195-5p regulates the multi-drug
resistance of gastric cancer cells via targeting ZNF139. (A)
Protein levels of P-gp, BCL-2, MRP1 and ZNF139 were measured in
MNK28 cells using western blot analysis. (B) Chemosensitivities of
5-FU and L-OHP were determined by MTT in MNK28 cells transfected
with siRNA-ZNF139 *P<0.05 and **P<0.01. miR, microRNA;
ZNF139, Zing finger 139; P-gp, P-glycoprotein; BCL-2, B-cell
lymphoma 2; MRP1, multi-drug resistance-associated protein 1; NCI,
negative control inhibitor; 195i, miR-195-5p inhibitor; si, small
interfering RNA; 5-FU, 5-fluorouracil; L-OHP, oxaliplatin. |
Discussion
GC is one of the most common malignant tumors with a
high mortality rate. The majority of patients with GC are diagnosed
at an advanced stage. Therefore, chemotherapy is important in the
treatment of GC. The MDR of GC cells is a major factor leading to
the failure of chemotherapy (22).
Therefore, preventing the MDR of GC is beneficial for improving the
effect of chemotherapy against GC.
It had been suggested that the degree of tumor
differentiation is associated with various biological
characteristics, including invasion, adhesion, proliferation,
metastasis and drug resistance. Previous studies have shown that
the degree of GC tumor differentiation affects their MDR.
Consistent with this, the results of the present study showed that
poorly differentiated GC cells were more sensitive to 5-FU and
L-OHP than well differentiated GC cells. In addition, miR-195-5p
was upregulated in poorly differentiated GC cells. These data
suggested that miR-195-5p may regulate the MDR of GC cells.
miR-195-5p is a member of the miR-15 family, which
is located on chromosome 17. Aberrant expression of miR-195-5p is
associated with various types of cancer. For example, miR-195-5p
serves as a tumor suppressor in renal cell carcinoma via
REGγ-mediated regulation of the wnt/β-catenin pathway (23). The overexpression of miR-195-5p
promotes apoptosis, and reduces cell growth, migration and invasion
in oral squamous cell carcinoma via targeting tripartite
motif-containing 14 (11). The
results of the present study revealed that the overexpression of
miR-195-5p enhanced chemotherapy sensitivity to 5-FU and L-OHP. The
protein levels of P-gp, BCL-2 and MRP1 were decreased in GC cells
transfected with the miR-195-5p mimic. P-gp and MRP1 are produced
by the mdr1 and mrp1 genes, which regulate tumor MDR. P-gp and MRP1
are cell membrane proteins, which function as an efflux pump to
transport chemotherapy drugs out of cells and lead to the MDR of
cancer cells (24). BCL-2 induces
the MDR of cancer cell by promoting cell proliferation and
inhibiting cell apoptosis (25,26).
MicroRNAs are a group of small, non-coding RNAs,
which exert their biological function by negatively regulating
their target genes. In the present study, bioinformatics databases
predicted that ZNF139 was a target gene of miR-195-5p. The results
of the luciferase assay confirmed that miR-195-5p was able to
directly bind to the 3′-UTR of ZNF139. ZNF139 is a zinc finger
protein; the zinc finger protein family function as transcriptional
factors to regulate gene expression, which are associated with
tumorigenesis, metastasis and drug resistance (27). It has been determined that ZNF139
affects the MDR of GC cells by promoting the expression of P-gp,
MRP1 and BCL-2 (20,21). In accordance, the results of the
present study indicated that silencing of ZNF139 reversed the
effects of miR-195-5p inhibitor on the MDR of GC cells.
In conclusion, the present study demonstrated that
miR-195-5p was associated with the degree of differentiation and
MDR of GC cells. The upregulation of miR-195-5p negatively
regulated the protein level of ZNF139 and promoted the
chemosensitivity of GC cells via affecting the protein expression
of P-gp, BCL-2 and MRP1.
Acknowledgements
Not applicable.
Funding
The present study was supported by The National
Science Foundation of China (grant no. 81072033).
Availability of data and materials
The analysed datasets generated during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
HN and YL planned the experiments and wrote the
manuscript; JM and JW performed the experiments and analyzed the
data. All authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethical Review
committee of the Fourth Hospital of Hebei Medical University.
Written informed consent was obtained from each patient.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Barra WF, Moreira FC, Pereira Cruz AM,
Khayat AS, Calcagno DQ, Carneiro Dos Santos NP, Mascarenhas Junior
RW, Thomaz Araújo TM, Ishak G, Demachki S and Rodríguez Burbano RM:
GEJ cancers: Gastric or esophageal tumors? searching for the answer
according to molecular identity. Oncotarget. 8:104286–104294. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cunningham SC, Kamangar F, Kim MP, Hammoud
S, Haque R, Maitra A, Montgomery E, Heitmiller RE, Choti MA,
Lillemoe KD, et al: Survival after gastric adenocarcinoma
resection: Eighteen-year experience at a single institutuion. J
Gastrointest Surg. 9:718–725. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hwang JH: Understanding gastric cancer
risk factors: We need to close the gap. Gut Liver. 12:1–2. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Peng PL, Zhou XY, Yi GD, Chen PF, Wang F
and Dong WG: Identification of a novel gene pairs signature in the
prognosis of gastric cancer. Cancer Med. 7:344–350. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nienhüser H and Schmidt T: Angiogenesis
and anti-angiogenic therapy in gastric cancer. Int J Mol Sci.
19:E432017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fan L, Tan B, Li Y, Zhao Q, Yuan H, Liu Y,
Wang D and Zhang Z: Upregulation of miR185 promotes apoptosis of
the human gastric cancer cell line MGC803. Mol Med Rep.
17:3115–3122. 2018.PubMed/NCBI
|
|
7
|
Huang J, He Y, McLeod HL, Xie Y, Xiao D,
Hu H, Chen P, Shen L, Zeng S, Yin X, et al: miR-302b inhibits
tumorigenesis by targeting EphA2 via Wnt/β-catenin/EMT signaling
cascade in gastric cancer. BMC Cancer. 17:8862017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang T, Zhou Y, Zhang J, Wong CC, Li W,
Kwan JSH, Yang R, Chan AKY, Dong Y, Wu F, et al: SRGAP1, a crucial
target of miR-340 and miR-124, functions as a potential oncogene in
gastric tumorigenesis. Oncogene. 37:1159–117. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Qu Y, Zhang H, Sun W, Han Y, Li S, Qu Y,
Ying G and Ba Y: MiR-155 promotes gastric cancer growth and
invasion by negatively regulating transforming growth factor β
receptor 2. Cancer Sci. 109:618–62. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shrestha S, Yang CD, Hong HC, Chou CH, Tai
CS, Chiew MY, Chen WL, Weng SL, Chen CC, Chang YA, et al: :
Integrated MicroRNA-mRNA analysis reveals miR-204 inhibits cell
proliferation in gastric cancer by targeting CKS1BCXCL1GPRC5A. Int
J Mol Sci. 19:E872017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang T, Ren Y, Liu R, Ma J, Shi Y, Zhang L
and Bu R: miR-195-5p suppresses the proliferation, migration, and
invasion of oral squamous cell carcinoma by targeting TRIM14.
Biomed Res Int. 2017:73781482017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yu S, Jing L, Yin XR, Wang MC, Chen YM,
Guo Y, Nan KJ and Han LL: MiR-195 suppresses the metastasis and
epithelial-mesenchymal transition of hepatocellular carcinoma by
inhibiting YAP. Oncotarget. 8:99757–99771. 2017.PubMed/NCBI
|
|
13
|
He B, Yan F and Wu C: Overexpressed
miR-195 attenuated immune escape of diffuse large B-cell lymphoma
by targeting PD-L1. Biomed Pharmacother. 98:95–101. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hong Z, Zhang R and Qi H: Diagnostic and
prognostic relevance of serum miR-195 in pediatric acute myeloid
leukemia. Cancer Biomark. 21:269–275. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu Y, Liu J, Wang L, Yang X and Liu X:
MicroRNA195 inhibits cell proliferation, migration and invasion in
laryngeal squamous cell carcinoma by targeting ROCK1. Mol Med Rep.
16:7154–7162. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qattan A, Intabli H, Alkhayal W, Eltabache
C, Tweigieri T and Amer SB: Robust expression of tumor suppressor
miRNA's let-7 and miR-195 detected in plasma of Saudi female breast
cancer patients. BMC Cancer. 17:7992017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fléjou JF: WHO classification of digestive
tumors: The fourth edition. Ann Pathol. 31 Suppl 5:S27–S31.
2011.(In French). View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Capes-Davis A, Theodosopoulos G, Atkin I,
Drexler HG, Kohara A, Macleod RA, Master JR, Nakamura Y, Reid YA,
Reddel RR and Freshney RI: Check your cultures! A list of
cross-contaminated or misidentified cell lines. Int J Cancer.
127:1–8. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Y, Tan BB, Zhao Q, Fan LQ, Liu Y and
Wang D: Regulatory mechanism of ZNF139 in multi-drug resistance of
gastric cancer cells. Mol Biol Rep. 41:3603–3610. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nie HF, Li Y, Li ZX, Mu JX and Wang JS:
Effects of ZNF139 on gastric cancer cells and mice with gastric
tumors. Oncol Lett. 12:2550–2554. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu W, Yang Z and Lu N: Molecular targeted
therapy for the treatment of gastric cancer. J Exp Clin Cancer Res.
35:12016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen S, Wang L, Yao X, Chen H, Xu C, Tong
L, Shah A, Huang T, Chen G, Chen J, et al: miR-195-5p is critical
in REGγ-mediated regulation of wnt/β-catenin pathway in renal cell
carcinoma. Oncotarget. 8:63986–64000. 2017.PubMed/NCBI
|
|
24
|
Stefan K, Schmitt SM and Wiese M:
9-Deazapurines as broad-spectrum inhibitors of the ABC transport
proteins P-glycoprotein, multidrug resistance-associated protein 1,
and breast cancer resistance protein. J Med Chem. 60:8758–8780.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang Y, Wang X, Zhao H, Liang B and Du Q:
Clusterin confers resistance to TNF-alpha-induced apoptosis in
breast cancer cells through NF-kappaB activation and Bcl-2
overexpression. J Chemother. 24:348–357. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tomek M, Akiyama T and Dass CR: Role of
Bcl-2 in tumour cell survival and implications for pharmacotherapy.
J Pharm Pharmacol. 64:1695–1702. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jen J and Wang YC: Zinc finger proteins in
cancer progression. J Biomed Sci. 23:532016. View Article : Google Scholar : PubMed/NCBI
|