Introduction
Breast cancer is the most common malignant tumor of
women that originates in breast epithelial tissue, and it is found
in women aged 40–60 years either before or after menopause. The
incidence of breast cancer has been increasing year by year in
recent years (1), and it has become
the second most common cancer worldwide. Breast cancer accounts for
~25% of all cancer-associated mortalities (2). Triple-negative breast cancer (TNBC)
accounts for ~12–20% of all incidences of breast cancer and is
associated with early recurrence and poor prognosis (2,3). TNBC
is a tumor that lacks the expression of the three common cell
surface receptors-estrogen receptor (ER), progesterone receptor
(PR) and human epidermal growth factor receptor-2 (HER2) (3,4).
Therefore, TNBC evades effective treatment as far as the majority
of the available targeted therapies are concerned (4). Owing to the high degree of malignancy,
invasive ability and the risk of distant metastasis, TNBC has
become a major focus of current research. As a representative cell
line of TNBC, MDA-MB-231 cells have been extensively studied,
achieving some meaningful results (5).
The family of bone morphogenetic proteins (BMPs)
belongs to the transforming growth factor-β (TGF-β) superfamily
(6). BMPs were originally viewed as
being osteoinductive cytokines that promote bone and cartilage
formation in vivo (7).
Recently, BMPs have been shown to be involved in the regulation of
tumorigenesis, development and bone metastases (8). BMP9 [also known as growth
differentiation factor 2 (GDF2)] is an integral member of the BMP
family that performs different functions in different tumor types
(9). For example, BMP9 may promote
the proliferation and migration of liver cancer cells (10), although it inhibits the growth of
osteosarcoma cells (11). Our
previous study revealed that BMP9 could inhibit the invasion,
migration and bone metastasis of MDA-MB-231 cells via the classical
BMP/SMAD signaling pathway (12).
Upon performing further studies, we discovered that BMP9 may also
have a role in other non-SMAD-dependent signaling pathways that are
involved in breast cancer, including the phosphoinositide 3-kinase
(PI3K)/Akt signaling pathway.
The PI3K/Akt pathway is one of the predominant
pathways that are involved in the malignant progression of various
types of tumors and mediates the proliferation, migration and
invasion of breast cancer tumors. The overexpression of BMP9
reduces phosphorylation of Akt, whereas interfering with BMP9
expression increases phosphorylation of Akt (13). Furthermore, the aberrant activation
of Akt is an important component in the recurrence and metastasis
of tumors (14–16). Certain BMPs are able to activate the
PI3K/Akt signaling pathway. Kang et al (17) identified that BMP2 may promote the
invasion and migration of gastric cancer cells by activating the
PI3K/Akt pathway and by upregulating matrix metalloproteinase 2
(MMP2). BMPs 4 and 7 were found to inhibit the apoptosis of ovarian
granulosa cells via the PI3K/phosphoinositide-dependent kinase 1
(PDK-1)/Akt and PI3K/PDK-1/protein kinase C (PKC) signaling
pathways, respectively (18). A
previous study published by our research group revealed that BMP9
was able to inhibit the growth, invasion and migration of
MDA-MB-231 cells (19). However,
whether the inhibitory mechanism is associated with the PI3K/Akt
signaling pathway has yet to be elucidated. Therefore, the present
study aimed to investigate the effect of BMP9 on the proliferation
of breast cancer cells and to explore the possible mechanism to
potentially provide a basis for targeted therapy of breast
cancer.
Materials and methods
Cell lines and adenoviral vectors
MDA-MB-231 breast cancer cells were purchased from
the Cell Resource Center, Shanghai Institute for Biological
Sciences (Chinese Academy of Sciences). The adenovirus that
overexpresses BMP9 and the adenovirus that expresses green
fluorescent protein (GFP), were obtained from the Molecular Medical
Laboratory of Yongchuan Hospital of Chongqing Medical University,
Chongqing, China.
Reagents
TRIzol® reagent was purchased from Takara
Biotechnology Co., Ltd. (Dalian, China); L15 medium was purchased
from Life Technologies (Thermo Fischer Scientific, Inc., Waltham,
MA, USA). Cell protein extraction and western blot detection
reagents RIPA lysis and extraction buffer were purchased from
Beyotime Biotechnology Co., Ltd. (Shanghai, China). Enhanced
chemiluminescence (ECL) detection kit Enlight™ was purchased from
Engreen Biological Co., Ltd. (Beijing, China). Phosphorylase and
protease inhibitor cocktail tablets PhosSTOP were purchased from
Roche Diagnostics (Basel, Switzerland). Mouse monoclonal GAPDH
antibody (cat. no. 97166), Akt total protein mouse monoclonal (cat.
no. 2920) and Akt phosphoprotein rabbit monoclonal (specific to
Ser473; cat. no. 4060) antibodies, cyclin E1 monoclonal antibody
(cat. no. 4129), cyclin D1 rabbit monoclonal antibody (cat. no.
2978) and cyclin B1 mouse monoclonal antibody (cat. no. 4135) were
purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA).
MM9 mouse monoclonal antibody (cat. no. ab58803) and BMP9 rabbit
polyclonal antibody (cat. no. ab71809) were obtained from Abcam
(Cambridge MA, USA). PV9001 horseradish enzyme anti-rabbit
immunoglobulin G (IgG) polymer, PV9002 horseradish enzyme
anti-mouse IgG polymer, and the DAB display reagent were purchased
from Zhongshan Golden Bridge Biological Co., Ltd. (Beijing,
China).
Patients and animals
A total of 23 pairs of breast cancer and adjacent
normal tissues (23 females, aged 25–77 years) were obtained from
the subjects (Table I), which were
confirmed by pathological examination using H&E staining. All
samples were collected from the Yongchuan Hospital of Chongqing
Medical University from May 2016 to August 2017. All the
experiments concerning the tumor specimens were approved by the
Ethics Association of Yongchuan Hospital of the Chongqing Medical
University, and all the patients signed the informed consent
form.
 | Table I.Clinicopathological features of the
23 breast cancer patients recruited to the study. |
Table I.
Clinicopathological features of the
23 breast cancer patients recruited to the study.
| Patient | Age (years) | Sex | Tumor grade | Tumor stage | Lymph node
metastasis | Tumor size
(cm) |
|---|
| 1 | 51 | Female | II | T2N3M0 | Yes | 2.5×2×1 |
| 2 | 53 | Female | I | T1N1M0 | Yes | 1.5×1.5×1 |
| 3 | 47 | Female | II | T1N0M0 | No | 2×2×1 |
| 4 | 54 | Female | II | T1N1M0 | Yes | 3×2×1 |
| 5 | 53 | Female | II | T1N1M0 | Yes | 1.8×1.5×1 |
| 6 | 49 | Female | II | T1N1M0 | Yes | 1.5×1×1 |
| 7 | 45 | Female | III | T1N1M0 | Yes | 2.5×1×1.5 |
| 8 | 64 | Female | II | T2N0M0 | No | 2×1.8×1.5 |
| 9 | 37 | Female | II | T1N0M0 | No | 1.5×1.2×1 |
| 10 | 53 | Female | II | T1N0M0 | No | 1×1×1 |
| 11 | 52 | Female | II | T1N0M0 | No | 0.8×0.8×0.7 |
| 12 | 50 | Female | II | T2N0M0 | No | 2.5×2×1.8 |
| 13 | 55 | Female | II | T1N0M0 | No | 1.5×1.3×1 |
| 14 | 74 | Female | II | T1N1M0 | Yes | 1.5×1.5×1.2 |
| 15 | 53 | Female | II | T1N0M0 | No | 2×1.5×1 |
| 16 | 58 | Female | II | T1N0M0 | No | 0.8×0.8×0.6 |
| 17 | 25 | Female | II | T2N1M0 | Yes | 2.5×1.5×1 |
| 18 | 43 | Female | III | T1N0M0 | Yes | 2×2×1.2 |
| 19 | 53 | Female | II | T1N0M0 | Yes | 1×0.8×0.8 |
| 20 | 43 | Female | II | T2N0M0 | Yes | 2.5×1.8×1.6 |
| 21 | 77 | Female | II | T1N0M0 | No | 2×1.6×1.5 |
| 22 | 50 | Female | II | T1N0M0 | Yes | 2×1.8×1.5 |
| 23 | 42 | Female | I | T2N3M0 | No | 3×2.5×1.2 |
Four-week-old male nude mice (weight 21 g) (BALB/c;
Beijing Hua Fukang Biological Polytron Technologies Inc., Beijing,
China) were used for generation of the xenograft model. All
experiments were approved by the Institutional Animal Care and Use
Committee of the Chongqing Medical University, as well as regional
authorities [reference number, SYXK(YU)2014-0001] in accordance
with the ‘Guidelines for the Welfare of Animals in Experimental
Neoplasia’ (The China Coordinating Committee on Cancer Research).
The animals were housed in individually ventilated cages under
sterile conditions and were granted unrestricted access to food and
water. At the end of the experimental period, the animals were
sacrificed by excessive ether inhalation.
Immunohistochemistry
The prepared tissue sections were dewaxed, hydrated
and antigen retrieval was performed. After washing three times with
PBS (5 min each wash), antibodies against BMP9, Akt and
phosphorylated Akt were added dropwise followed by incubation in
the humid chamber at 37°C for 2 h. Following three further washes
with PBS (5 min each wash), a secondary antibody (PV9001 or PV9002)
was added, and the sections were incubated at 37°C for a further 20
min. DAB and hematoxylin counterstain were added for 3 min followed
by routine dehydration to maintain transparency and sealing. Each
slice was photographed using the BX51P Olypmus polarizing
microscope (Olympus Corp., Tokyo, Japan) at a low magnification
×100 (with 10 randomly selected fields of view), and the cumulative
optical density of the dyed yellow areas was assessed using ImageJ
software (National Institutes of Health, Bethesda, MD, USA).
Cell culture and infection
MDA-MB-231 cells were cultured in L15 culture medium
containing 10% fetal bovine serum(FBS) (including 1% penicillin and
streptomycin) at 37°C, in the absence of CO2. When the
cells had reached a confluency of ~70–80%, they were digested and
passaged with 0.25% trypsin (containing 0.02% EDTA 2Na), and the
culture was continued. MDA-MB-231 cells in the exponential phase of
growth were seeded at a density of 2×106/culture flask.
After 24 h, the cells were subsequently infected with adenovirus
vehicles that express GFP and recombinant BMP-9. After a further
8–12 h cultivation, the medium was replaced by fresh medium, and
the fluorescence was subsequently observed 24 h later using a
fluorescence microscope (Olympus Imaging IX71; Olympus Corp.).
Real-time quantitative polymerase
chain reaction (RT-qPCR)
The total RNA of the MDA-MB-231 cells, and the total
RNA of the MDA-MB-231/BMP9 and MDA-MB-231/GFP cells were extracted
using the TRIzol® method after 24 h of adenovirus
infection (12). Aliquots (2 µg) of
total RNA were used to synthesize cDNA using a reverse
transcription kit, and subsequently 1 µl cDNA was used as a
template for PCR amplification. The following primer sequences were
used: GAPDH, sense, 5′-CAGCGACACCCACTCCTC-3′ and antisense,
5′-TGAGGTCCACCACCCTGT-3′; cyclin B1 sense, 5′-GCACTTTCCTCCTTCTCA-3′
and antisense, 5′-CGATGTGGCATACTTGTT-3′; cyclin D1 sense,
5′-GCGAGGAACAGAAGTGCG-3′andantisense,5′-GGAGTTGTCGGTGTAGATGC-3′;cyclinE1sense,5′-TGGATGTTGACTGCCTTGA-3′andantisense,5′-GTCGCACCACTGATACCCT-3′;c-Mycsense,5′-ACACCCTTCTCCCTTCG-3′
and antisense, 5′-CCGCTCCACATACAGTCC-3′; and MMP9 sense,
5′-GAACTTTGACAGCGACAAGA-3′ and antisense, 5′-GGCGAGGACCATAGAGG-3′.
The PCR reaction conditions were as follows: An initial step at
95°C for 30 sec; denaturation at 95°C for 5 sec; followed by
annealing for 30 sec at 55°C and extension for 10 sec at 72°C in a
total of 40 cycles. The experiment was repeated three times. The
2−ΔΔCq method was utilized to normalize the expression
of target mRNAs relative to the reference gene, GAPDH (12).
Western blotting
A density of 5×105 MDA-MB-231
cells/culture flask were passaged into 6-cm diameter Petri dishes.
When the cell confluency reached ~50–60%, the adenovirus that
overexpresses BMP9 was added, and the medium was subsequently
replaced with serum-free L15 medium after 24 h. The cells were
cultivated for a further 48 h, at which time the culture solution
was discarded, and the cells were subsequently washed twice with
pre-cooled PBS. Cell lysate (150 µl/ Petri dish) was added,
followed by lysis for 30 min on ice, and the cells were centrifuged
at 4°C for 15 min at 12,000 rpm (10,000 × g/min). The supernatant
was then transferred to a 1.5 ml EP tube, prior to utilizing the
bicinchoninic acid (BCA) method for the determination of protein
concentration. Aliquots (15 µl) of the protein solution were
extracted for protein electrophoresis (SDS-PAGE; concentration of
gel 5%, separation gel, 10%). The proteins were electrotransferred
onto a polyvinylidene difluoride (PVDF) membrane, and then blocked
with 5% bovine serum albumin (BSA) at 37°C for 1 h. The primary
antibodies against GAPDH (dilution 1:1,000), Akt (dilution
1:1,000), phosphorylated Akt (p-Akt; dilution 1:1,000), c-Myc
(dilution 1:1,000), cyclin D1 (dilution 1:1,000), cyclin B1
(dilution 1:1,000), cyclin E1 (dilution 1:1,000) and MMP9 (dilution
1:500) were added and incubated overnight at 4°C. The membranes
were then washed three times (15 min each wash) in 0.1%
Tris-buffered saline/Tween (TBST) solution, after which the
corresponding secondary antibody (goat anti-rabbit or goat
anti-mouse) (dilution, 1:5,000) was added. The cells were incubated
for 1 h at 37°C, and the PVDF membranes were washed three times (15
min each wash) in 0.1% TBST solution. Finally, the ECL chromogenic
solution was used for imaging. GEL EQ (Bio-Rad Laboratories,
Hercules, CA, USA) was used in the gel imager, and Quantity
One® software (Bio-Rad Laboratories, Inc.) was used to
analyze the relative optical densities. The relative expression
level of the target gene protein was calculated according to the
gray value of the target gene band/the gray value of the internal
reference band.
Animal models
Trypsin solution (0.25%) was used to digest the
MDA-MB-231 cells, and the MDA-MB-231/GFP and MDA-MB-231/BMP9 cells
in the exponential growth phase 24 h following adenovirus
infection. Tumor cells were injected subcutaneously into the backs
of the nude mice for each group of five nude mice (0.1 ml/nude
mouse; adjusted cell density, 5×106 cells/ml). Once the
tumors of any group were visible and palpable, the starting point
measurements were taken. The diameters of the tumor were
subsequently recorded every three days. The volume (V) was
calculated based on the following formula: V = 4/3π l/2 w/2
h/2, (where π=3.14, w is the width, l is the length and h is the
height), and the growth curves of the xenografts were plotted.
Following inoculation of the tumor cells, all nude mice were
sacrificed on the 25th day after the animal model had been
successfully constructed. The tumor tissue was dissected, and the
tumor size was measured. The transplanted tumors were subsequently
stripped, fixed in 4% polyformaldehyde solution, paraffin-embedded,
and then sectioned. The tumor tissue sections were stained with
H&E prior to immunohistochemistry and analysis using ImageJ
software (National Institutes of Health).
Statistical analysis
SPSS version 20.0 statistical software (IBM Corp.,
Armonk, NY, USA) was used to analyze the experimental data. The
results are expressed as the mean ± standard deviation (SD). The
expression of BMP9 or p-AKT using immunohistochemical staining in
breast cancer specimens was analyzed by paired Student's t-test.
The correlation between BMP9 and p-Akt was analyzed by the Spearman
correlation coefficient method. The other data were analyzed by
analysis of variance (ANOVA). Multiple comparison between the
groups was performed using the S-N-K method. The expression levels
of cyclin D1, cyclin B1, cyclin E1, c-Myc and MMP9 were analyzed
using RT-qPCR and the 2−ΔΔCq method. P<0.05 was
considered to indicate a statistically significant difference.
Results
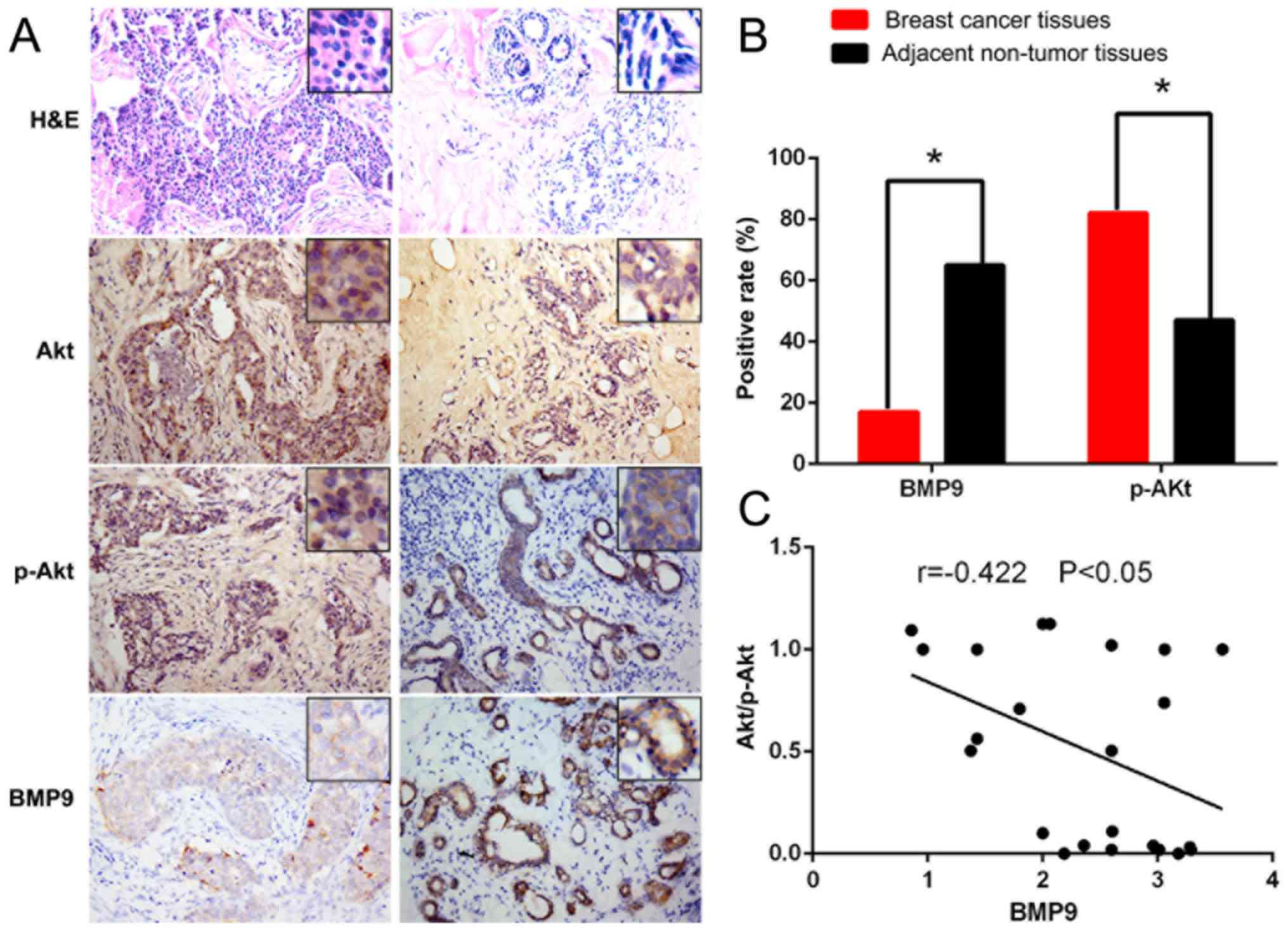
Expression level of BMP9 is
downregulated in breast cancer tissues, whereas the expression of
p-Akt is upregulated
In the 23 clinical breast cancer specimens,
immunohistochemical staining showed that the positive rate of BMP9
was 17.4% (4/23) in the tumor tissues, whereas the positive rate of
BMP9 was 65.2% (15/23) in the paracancerous tissues. The expression
level of BMP9 was significantly lower in the tumor tissues compared
with the level noted in the paracancerous tissues (P<0.05). The
positive rate of p-Akt was 82.6% (19/23) in the cancer tissues,
whereas the positive rate of p-Akt was 47.8% (11/23) in the
paracancerous tissues. The protein expression level of p-Akt was
significantly higher (P<0.05) in cancer tissues than the level
noted in the paracancerous tissues (Fig. 1A and B). Spearman correlation
coefficient analysis revealed that the expression level of BMP9 was
negatively correlated with the p-Akt protein expression level in
tumor tissue (r=−0.422; P<0.05) (Fig. 1C). These results supported that the
PI3K/Akt signaling pathway is involved in the development and
progression of breast cancer, since the expression level of BMP9
was negatively correlated with the expression level of p-Akt.
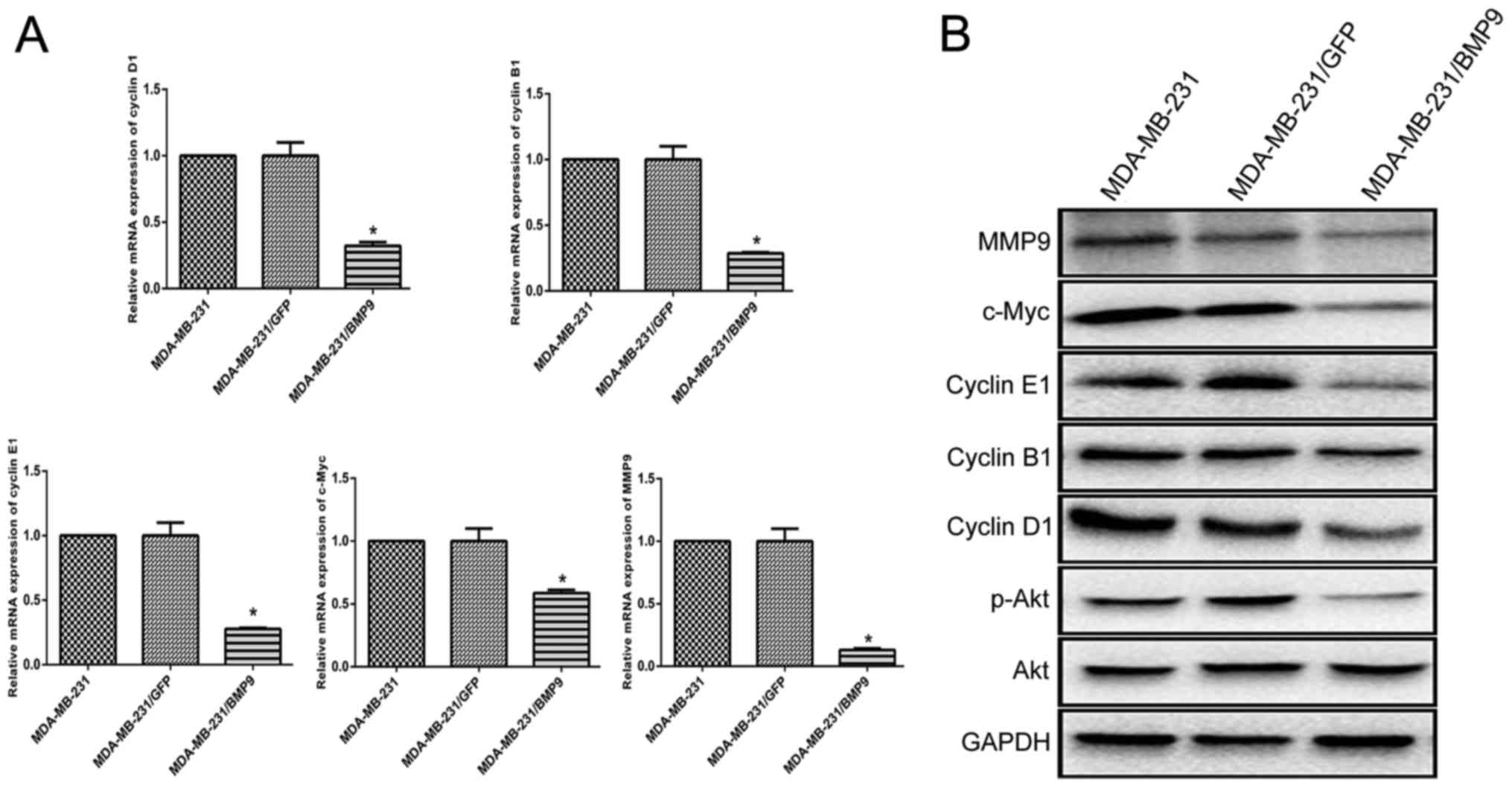
BMP9 inhibits the PI3K/Akt signaling
pathway and downregulates the expression levels of cyclin D1,
cyclin B1, cyclin E1, c-Myc and MMP9 in MDA-MB-231 cells
To investigate whether BMP9 exerts an inhibitory
effect on the PI3K/Akt signaling pathway, western blotting was
performed. Western blotting revealed that the protein expression
level of p-Akt in the BMP9 overexpression group (MDA-MB-231/BMP9)
was significantly lower compared with the GFP group
(MDA-MB-231/GFP) and the blank control group (MDA-MB-231) after the
MDA-MB-231 cells had been infected with the adenovirus
overexpressing BMP9 for 48 h. These results suggested that BMP9
inhibited the PI3K/Akt signaling pathway.
Cyclins D1, B1 and E1, c-Myc, and MMP9 have been
reported to be involved in the proliferation and invasion of tumor
cells (20–24). Therefore, RT-qPCR was used to
estimate their expression levels in recombinant MDA-MB-231/BMP9
cells. The expression levels of cyclin D1, cyclin B1, cyclin E1,
c-Myc and MMP9 were all shown to be downregulated in the
MDA-MB-231/BMP9 cells (cyclin D1: 2−ΔΔCq=0.321; cyclin
B1: 2−ΔΔCq=0.287; cyclin E1: 2−ΔΔCq=0.277;
c-Myc: 2−ΔΔCq=0.586; MMP9: 2−ΔΔCq=0.130)
(Fig. 2A). Western blotting was
performed to confirm the expression levels of cyclin D1, cyclin B1,
cyclin E1, c-Myc and MMP9 in the MDA-MB-231/BMP9 groups. The
results demonstrated that these proteins were significantly lower
in MDA-MB-231/BMP9 cells compared with these level in the
MDA-MB-231/GFP and MDA-MB-231 groups (Fig. 2B). These results suggested that BMP9
inhibited the proliferation of MDA-MB-231 cells via downregulation
of the expression levels of cyclin D1, cyclin B1, cyclin E1, c-Myc
and MMP9.
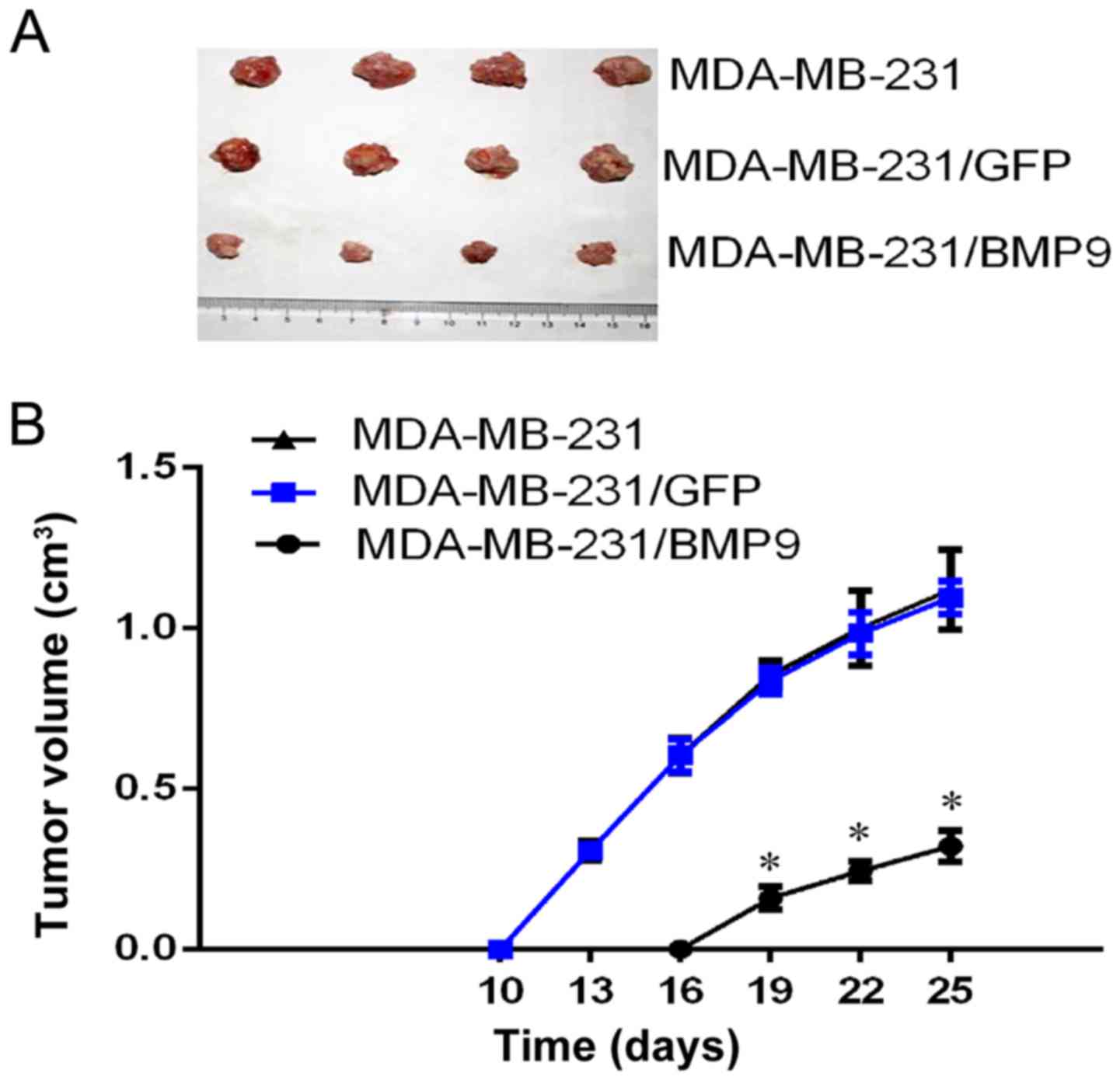
Overexpression of BMP9 inhibits the
growth of breast cancer tumors in a nude mouse xenograft tumor
model
After the successful generation of the nude mouse
xenograft tumor model, tumors were identified on the 10th day in
the MDA-MB-231/GFP and the MDA-MB-231 groups, although tumors were
found on the 16th day in the MDA-MB-231/BMP9 group. All the nude
mice were sacrificed on the 25th day after the animal model had
been successfully constructed, and the tumor tissue was dissected.
The size of the tumors was measured using a Vernier caliper (the
maximum tumour diameter: MDA-MB-231/BMP9 group was 1.08 cm;
MDA-MB-231/GFP group was 1.48 cm; MDA-MB-231 group was 1.49 cm on
the 25th day). The tumor volume in the MDA-MB-231/BMP9 group
(0.32±0.05 cm3) was markedly lower when compared with
that in the MDA-MB-231/GFP group (1.10±0.05 cm3) and
MDA-MB-231 group (1.12±0.12 cm3) (P<0.001; Fig. 3A and B). At the same time, the
tumors of the MDA-MB-231/GFP and MDA-MB-231 groups were found to be
necrotic and ulcerated when the tumors were peeled off. Owing to
the adhesion between the tumor and the skin of the nude mice, the
tumors were not easy to peel off. By contrast, the tumor masses of
the MDA-MB-231/BMP9 group were found to be hard, there was no
obvious necrosis, and the tumors were easy to peel off. These
results demonstrated that the overexpression of BMP9 may
considerably inhibit the growth of breast cancer cells.
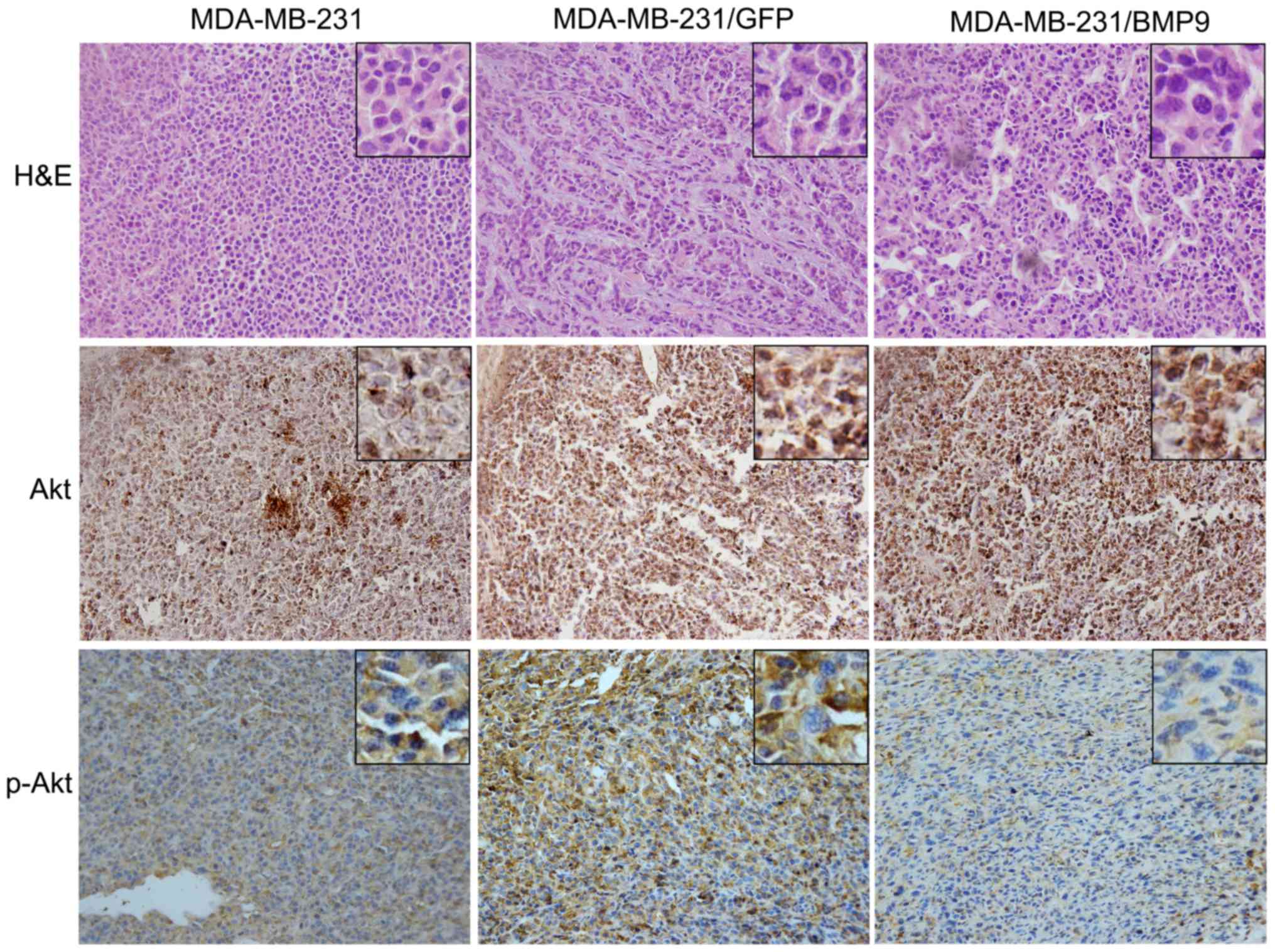
BMP9 inhibits PI3K/Akt expression in
the nude mouse xenograft tumor model
In the nude mouse xenograft tumor model,
immunohistochemical staining revealed that p-Akt protein expression
was obviously decreased in the MDA-MB-231/BMP9 group compared with
that noted in the MDA-MB-231/GFP and MDA-MB-231 groups. The t-Akt
protein expression exhibited no difference in the three groups.
This result further confirmed that BMP9 inhibits the growth of
breast cancer by inhibiting the PI3K/Akt signaling pathway in
vivo.
Discussion
Previous studies have revealed that BMPs are able to
promote bone formation, cell invasion and metastasis via the
classical BMP/SMAD signaling pathway (12). Recent studies have reported the
effect of BMPs on certain non-classical signal transduction
pathways, including those involving PI3K/Akt and mitogen-activated
protein kinases (MAPKs) (25–28).
These non-classical BMP/SMAD signaling pathways have an important
role in tumor growth, invasion and metastasis. Pancholi et
al (29) demonstrated that the
estrogen receptor (ER) could bind to both ERBB2 and PI3K, leading
to the phosphorylation of Akt. Another study by Lin et al
(30) confirmed that Akt is the
main downstream target of PI3K, demonstrating that PI3K and Akt may
work together to participate in tumorigenesis, development and
metastasis. It has been reported that most BMPs can promote tumor
growth, invasion and metastasis via the PI3K/Akt signaling pathway.
Ren et al (13) showed that
PI3K/Akt activation could promote the proliferation, migration and
invasion of HER2-positive SK-BR-3 breast cancer cells, whereas Duan
et al (31) showed that
inhibition of the PI3K/Akt pathway could inhibit cell proliferation
and migration in gastric cancer.
BMP9 was originally cloned from a fetal mouse liver
cDNA. It has been shown as a pleiotropic cytokine involved in a
number of physiologic events. These include hepatic
reticuloendothelial system, bone morphogenesis, neuronal
differentiation, hematopoiesis, angiogenesis, iron homeostasis,
glucose homeostasis and cancer growth or metastasis. In our
previous studies, HCT116 cells were infected with the BMP9
adenovirus to construct conditioned medium, and we found that BMP9
mRNA was readily detectable in the HCT116/BMP9 cells, and the
conditioned medium indeed contained detectable levels of BMP9
protein (9). Our research confirm a
previous study by Ye et al that BMP9 was released into the
medium following transfection of the gene into cells (32).
BMP9, as a multifunctional cytokine, not only
induces bone and cartilage formation but is also involved in the
proliferation, differentiation, invasion and metastasis of tumor
cells. A previous study by our research laboratory demonstrated
that BMP9 could inhibit the proliferation of lung adenocarcinoma
A549 cells via the SMAD-dependent BMP/SMAD signaling pathway
(33). Our previous research also
revealed that BMP9 can inhibit the proliferation, invasion, and
migration of breast cancer cells via the activin receptor-like
kinase-2 (ALK2)-dependent BMP/SMAD signaling pathway (19). This prompted us to consider whether
BMP9 exerts an effect on certain non-classical signal transduction
pathways, and therefore our research studies were aimed to
elucidate the mechanism of BMP9 in breast cancer cells in order to
provide a theoretical basis for the targeted therapy of breast
cancer.
In the present study, 23 cases of patients with
clinical breast cancer were first identified. Immunohistochemical
staining showed that the positive rate of BMP9 expression in breast
cancer was significantly lower when compared with that in adjacent
normal tissues (P<0.05). In addition, the protein expression
level of p-Akt was significantly reduced in the samples with BMP9
expression. Furthermore, the expression level of BMP9 in tumor
tissue was negatively correlated with the expression level of p-Akt
protein (r=−0.422; P<0.05) (Fig.
1C). These preliminary results suggest that BMP9 may
participate in the growth, invasion, and metastasis of breast
cancer cells via the PI3K/Akt signaling pathway. Subsequently, BMP9
was overexpressed in MDA-MB-231 breast cancer cells by adenovirus
infection in vitro, revealing that the overexpression of
BMP9 inhibited the protein expression level of p-Akt (Fig. 2B). Experiments performed in
vivo further confirmed that BMP9 serves an inhibitory role in
the growth, invasion, and migration of breast cancer cells, and
that a clear association exists with the PI3K/Akt signaling
pathway. In addition to acting via the classical BMP/SMAD signaling
pathway, BMP9 also functions through the non-SMAD-dependent
PI3K/Akt signaling pathway.
To further explore the carcinogenic mechanism of
BMP9 in breast cancer, a nude mouse xenograft tumor model was
constructed. The animal experiments demonstrated that BMP9 can
inhibit breast cancer growth (Fig. 3A
and B). After constructing the nude mouse model over a 28-day
period, the nude mice were sacrificed and the tumor tissues were
isolated and paraffin-embedded. Immunohistochemistry results showed
that the positive percentage of p-Akt protein in the
MDA-MB-231/BMP9 group was significantly lower compared with the
MDA-MB-231 and MDA-MB-231/GFP groups (Fig. 4). These results suggested that BMP9
can indeed inhibit the growth of MDA-MB-231 cells by inhibiting the
PI3K/Akt signaling pathway in vivo, thereby providing
experimental data in support of the clinical application of BMP9 in
the treatment of breast cancer.
As a multifunctional factor, the role of BMP9 in
osteogenesis and breast cancer has been progressively determined.
Accumulating evidence has shown that BMP9 may exert a role through
multiple signaling pathways (34–36).
The study has also, in part, explained why BMPs serve different
roles in different tumors, and even for the same BMP in different
tumors, its role may significantly differ. For example, BMP9 was
found to have an inhibitory role in prostate cancer and breast
cancer, whereas BMP9 had a role in promoting ovarian cancer
(32,37). We demonstrated that BMP9 can inhibit
the PI3K/Akt signaling pathway. Yet, we did not ascertain how BMP9
regulates the PI3K/Akt signaling pathway. We will knock down the
BMP9 type I receptor (ALK1 or ALK2) to inhibit the BMP9-independent
SMAD signaling pathways in order to determine the effect of BMP9
overexpression on breast tumor cell growth in future studies. The
published data indicate that ALK1 or ALK2 belongs to BMP9 type I
receptors and can bind with BMP9. Knocking down each of the BMP9
type I receptors can directly inhibit BMP9 signaling pathways. It
will confirm whether BMP9 can directly target the PI3K/AKT
signaling pathway or not (13).
The present study has shown that BMP9 can inhibit
the PI3K/Akt signaling pathway consequently suppressing breast
cancer growth. In addition, it has been shown by RT-qPCR and
western blotting that BMP9 may inhibit the expression levels of
cyclin D1, cyclin B1, cyclin E1, c-Myc and MMP9 (Fig. 2A and B), which subsequently leads to
inhibition of the proliferation, invasion, and metastasis of breast
cancer cells. These results may explain, in part, how the degree of
malignancy of MDA-MB-231 cells was increased in our previous
studies after the inhibitory effect of BMP9 on connective tissue
growth factor (CTGF) had been restored in MDA-MB-231 cells by using
an adenovirus co-infection technique in vitro (12), although certain differences existed
compared with the blank control group. Since BMP9 can exert a role
via the BMP/SMAD signaling pathway as well as through the
non-classical PI3K/Akt signal transduction pathway, this may also
explain why BMP9 considerably inhibited the growth of MDA-MB-231
cells.
In conclusion, the present study has shown that BMP9
expression was downregulated in breast cancer cells. This study has
shown, to the best of our knowledge for the first time, that BMP9
inhibits the growth of breast cancer by inhibiting the PI3K/Akt
signaling pathway. At the same time, BMP9 can also inhibit the
proliferation, invasion, and metastasis of breast cancer by
inhibiting the expression of cyclin D1, cyclin B1, cyclin E1,
c-Myc, and MMP9. Therefore, BMP9-mediated PI3K/Akt signaling may
serve as a novel target for potential breast cancer therapy to
prevent proliferation, invasion, and metastasis of breast cancer
cells.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Foundation Project of Yongchuan (grant nos. Ycstc.2016nc510
and Ycstc.2017nc5020), the Natural Science Foundation Project of
Chongqing Education Committee (grant no. KJ1500202) and the Natural
Science Foundation Project of Yongchuan Hospital (grant no.
YJZQN201517).
Availability of data and material
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
HD, SL, TZ, YW, CL and SP performed the research. KW
and SL designed the research study. SL, YH and HD analyzed the
data. SL wrote the paper. KW, as the corresponding author, modified
the article and proposed its written content. All authors have read
and approved the final version of the manuscript and agree to be
accounTable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All the experiments concerning the tumor specimens
were approved by the Ethics Association of Yongchuan Hospital of
the Chongqing Medical University, and all the patients signed the
informed consent form. All experiments were approved by the
Institutional Animal Care and Use Committee of the Chongqing
Medical University, as well as regional authorities [reference no.
SYXK(YU)2014-0001] in accordance with the ‘Guidelines for the
Welfare of Animals in Experimental Neoplasia’ (The China
Coordinating Committee on Cancer Research).
Patient consent for publication
All the patients signed the informed consent
form.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Zhang Z, Ni C, Chen W, Wu P, Wang Z, Yin
J, Huang J and Qiu F: Expression of CXCR4 and breast cancer
prognosis: A systematic review and meta-analysis. BMC Cancer.
14:492014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Richards CE, Vellanki SH, Smith YE and
Hopkins AM: Diterpenoid natural compound C4 (Crassin) exerts
cytostatic effects on triple-negative breast cancer cells via a
pathway involving reactive oxygen species. Cell Oncol. 41:35–46.
2018. View Article : Google Scholar
|
|
3
|
Chacón RD and Costanzo MV: Triple-negative
breast cancer. Breast Cancer Res. 12 Suppl 2:S32010. View Article : Google Scholar
|
|
4
|
Foulkes WD, Smith IE and Reis-Filho JS:
Triple-negative breast cancer. N Engl J Med. 363:1938–1948. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ma H, Xu X, Clague J, Lu Y, Togawa K, Wang
SS, Clarke CA, Lee E, Park HL, Sullivan-Halley J, et al:
Recreational physical activity and risk of triple negative breast
cancer in the California Teachers Study. Breast Cancer Res.
18:622016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wang K, Feng H, Ren W, Sun X, Luo J, Tang
M, Zhou L, Weng Y, He TC and Zhang Y: BMP9 inhibits the
proliferation and invasiveness of breast cancer cells MDA-MB-231. J
Cancer Res Clin Oncol. 137:1687–1696. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang RN, Green J, Wang Z, Deng Y, Qiao M,
Peabody M, Zhang Q, Ye J, Yan Z, Denduluri S, et al: Bone
morphogenetic protein (BMP) signaling in development and human
diseases. Genes Dis. 1:87–105. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ye L, Bokobza SM and Jiang WG: Bone
morphogenetic proteins in development and progression of breast
cancer and therapeutic potential (Review). Int J Mol Med.
24:591–597. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wan S, Liu Y, Weng Y, Wang W, Ren W, Fei
C, Chen Y, Zhang Z, Wang T, Wang J, et al: BMP9 regulates
cross-talk between breast cancer cells and bone marrow-derived
mesenchymal stem cells. Cell Oncol. 37:363–375. 2014. View Article : Google Scholar
|
|
10
|
García-Álvaro M, Addante A, Roncero C,
Fernández M, Fabregat I, Sánchez A and Herrera B: BMP9-induced
survival effect in liver tumor cells requires p38MAPK activation.
Int J Mol Sci. 16:20431–20448. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lv Z, Wang C, Yuan T, Liu Y, Song T, Liu
Y, Chen C, Yang M, Tang Z, Shi Q, et al: Bone morphogenetic protein
9 regulates tumor growth of osteosarcoma cells through the
Wnt/β-catenin pathway. Oncol Rep. 31:989–994. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ren W, Sun X, Wang K, Feng H, Liu Y, Fei
C, Wan S, Wang W, Luo J, Shi Q, et al: BMP9 inhibits the bone
metastasis of breast cancer cells by downregulating CCN2
(connective tissue growth factor, CTGF) expression. Mol Biol Rep.
41:1373–1383. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ren W, Liu Y, Wan S, Fei C, Wang W, Chen
Y, Zhang Z, Wang T, Wang J, Zhou L, et al: BMP9 inhibits
proliferation and metastasis of HER2-positive SK-BR-3 breast cancer
cells through ERK1/2 and PI3K/AKT pathways. PLoS One. 9:e968162014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zardavas D, Fumagalli D and Loi S:
Phosphatidylinositol 3-kinase/AKT/mammalian target of rapamycin
pathway inhibition: A breakthrough in the management of luminal
(ER+/HER2-) breast cancers? Curr Opin Oncol. 24:623–634. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ciruelos Gil EM: Targeting the
PI3K/AKT/mTOR pathway in estrogen receptor-positive breast cancer.
Cancer Treat Rev. 40:862–871. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Grunt TW and Mariani GL: Novel approaches
for molecular targeted therapy of breast cancer: Interfering with
PI3K/AKT/mTOR signaling. Curr Cancer Drug Targets. 13:188–204.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kang MH, Oh SC, Lee HJ, Kang HN, Kim JL,
Kim JS and Yoo YA: Metastatic function of BMP-2 in gastric cancer
cells: The role of PI3K/AKT, MAPK, the NF-κB pathway, and MMP-9
expression. Exp Cell Res. 317:1746–1762. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shimizu T, Kayamori T, Murayama C and
Miyamoto A: Bone morphogenetic protein (BMP)-4 and BMP-7 suppress
granulosa cell apoptosis via different pathways: BMP-4 via
PI3K/PDK-1/Akt and BMP-7 via PI3K/PDK-1/PKC. Biochem Biophys Res
Commun. 417:869–873. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang K, Liu D, Zhu TJ, Tang ZG and Dai HY:
BMP9 inhibit the proliferation, invasion and migration of
MDA-MB-231 breast cancer cells through binding ALK2 receptor to
activate BMPs/SMAD cell signaling pathway. Genomics and Applied
Biology. 35:1569–1576. 2016.
|
|
20
|
Bolat Kucukzeybek B, Vedat Bayoglu I,
Kucukzeybek Y, Alacacioglu A, Yigit S, Akder Sari A, Akyol M and
Oktay Tarhan M: The prognostic significance of cyclin D1 expression
in patients with triple-negative breast cancer. J BUON. 22:947–952.
2017.PubMed/NCBI
|
|
21
|
Khan S, Brougham CL, Ryan J, Sahrudin A,
O'Neill G, Wall D, Curran C, Newell J, Kerin MJ and Dwyer RM:
JmiR-379 regulates cyclin B1 expression and is decreased in breast
cancer. PLoS One. 8:e687532013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Guo X, Connick MC, Vanderhoof J, Ishak MA
and Hartley RS: MicroRNA-16 modulates HuR regulation of Cyclin E1
in breast cancer cells. Int J Mol Sci. 16:7112–7132. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang J, Li M, Chen D, Nie J, Xi Y, Yang X,
Chen Y and Yang Z: Expression of C-myc and β-catenin and their
correlation in triple negative breast cancer. Minerva Med.
108:513–517. 2017.PubMed/NCBI
|
|
24
|
Padala C, Tupurani MA, Puranam K, Gantala
S, Shyamala N, Kondapalli MS, Gundapaneni KK, Mudigonda S, Galimudi
RK, Kupsal K, et al: Synergistic effect of collagenase-1 (MMP1),
stromelysin-1 (MMP3) and gelatinase-B (MMP9) gene polymorphisms in
breast cancer. PLoS One. 12:e01844482017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Li B, Yang Y, Jiang S, Ni B, Chen K and
Jiang L: Adenovirus-mediated overexpression of BMP-9 inhibits human
osteosarcoma cell growth and migration through downregulation of
the PI3K/AKT pathway. Int J Oncol. 41:1809–1819. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pal I and Mandal M: PI3K and Akt as
molecular targets for cancer therapy: Current clinical outcomes.
Acta Pharmacol Sin. 33:1441–1458. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Z and Guo J: Mechanical induction of
BMP-7 in osteocyte blocks glucocorticoid-induced apoptosis through
PI3K/AKT/GSK3β pathway. Cell Biochem Biophys. 67:567–574. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zheng Y, Wang X, Wang H, Yan W, Zhang Q
and Chang X: Bone morphogenetic protein 2 inhibits hepatocellular
carcinoma growth and migration through downregulation of the
PI3K/AKT pathway. Tumour Biol. 35:5189–5198. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pancholi S, Lykkesfeldt A, Johnston SRD,
Dowsett M and Martin LA: The interaction of the ER with ERBB2 and
PI3K results in elevated levels of AKT and p90RSK in
tamoxifen-resistant MCF-7 cells. Breast Cancer Res. 7 Suppl
2:P2.082005. View
Article : Google Scholar :
|
|
30
|
Lin M, Bi H, Yan Y, Huang W, Zhang G,
Zhang G, Tang S, Liu Y, Zhang L, Ma J, et al: Parthenolide
suppresses non-small cell lung cancer GLC-82 cells growth via
B-Raf/MAPK/Erk pathway. Oncotarget. 8:23436–23447. 2017.PubMed/NCBI
|
|
31
|
Duan L, Ye L, Wu R, Wang H, Li X, Li H,
Yuan S, Zha H, Sun H, Zhang Y, et al: Inactivation of the
phosphatidylinositol 3-kinase/Akt pathway is involved in
BMP9-mediated tumor-suppressive effects in gastric cancer cells. J
Cell Biochem. 116:1080–1089. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ye L, Kynaston H and Jiang WG: Bone
morphogenetic protein-9 induces apoptosis in prostate cancer cells,
the role of prostate apoptosis response-4. Mol Cancer Res.
6:1594–1606. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dai HY, Xia WY, Zhu TJ, Tang ZG and Wang
K: BMP9 inhibits the proliferation of human lung adenocarcinoma
A549 cells through BMPs/SMAD signaling pathway. TUMOR. 35:997–1005.
2015.
|
|
34
|
Liu P, Man Y, Wang Y and Bao Y: Mechanism
of BMP9 promotes growth of osteosarcoma mediated by the Notch
signaling pathway. Oncol Lett. 11:1367–1370. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Muñoz-Félix JM, Cuesta C, Perretta-Tejedor
N, Subileau M, López-Hernández FJ, López-Novoa JM and
Martínez-Salgado C: Identification of bone morphogenetic protein 9
(BMP9) as a novel profibrotic factor in vitro. Cell Signal.
28:1252–1261. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yuan SX, Wang DX, Wu QX, Ren CM, Li Y,
Chen QZ, Zeng YH, Shao Y, Yang JQ, Bai Y, et al: BMP9/p38 MAPK is
essential for the antiproliferative effect of resveratrol on human
colon cancer. Oncol Rep. 35:939–947. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Herrera B, van Dinther M, TenDijke P and
Inman GJ: Autocrine bone morphogenetic protein-9 signals through
activin receptor-like kinase-2/Smad1/Smad4 to promote ovarian
cancer cell proliferation. Cancer Res. 69:9254–9262. 2009.
View Article : Google Scholar : PubMed/NCBI
|