Introduction
Lung cancer is a leading cause of cancer-related
mortality with ~1.59 million deaths annually worldwide based on the
most recent WHO survey (available at: http://www.who.int/en/news-room/fact-sheets/detail/cancer).
In China, lung cancer was estimated to account for 21.6% of all
cancer-related deaths in 2015 (1).
Lung cancer consists of NSCLC (~85%) and small cell lung cancer
(SCLC; ~15%). This study focused on the major histological subtype
NSCLC, which can be further classified as lung adenocarcinoma
(LUAD) and lung squamous cell carcinoma (LUSC) (2).
Gasdermin D (GSDMD) belongs to the structurally and
evolutionarily conserved GSDM protein superfamily, the biochemical
functions of which are largely unknown (3). Humans possess GSDMA, GSDMB, GSDMC,
GSDMD, DFNA5 and DFNB59. Mice harbor three GSDMAs (GSDMA1-3) and
four GSDMCs (GSDMC1-4) (4).
In 2015, two independent studies identified GSDMD as
the key executioner of pyroptosis (5,6).
Pyroptosis is a lytic and inflammatory form of programmed cell
death characterized by cell swelling and lysis, concurrent with
cellular extrusion of various substances, such as the
pro-inflammatory mediators lL-1β and IL-18 (7). Inflammatory caspases cleave GSDMD into
a 32-kDa N-terminal domain (GSDMD-N) and a 22-kDa C-terminal domain
(GSDMD-C), thus relieving the autoinhibitory effect of GSDMD-C on
GSDMD-N. The GSDMD-N domain, acting as the direct executor of
pyroptosis, then translocates to the plasma membrane and forms
pores (8–11). Mechanistically, the activation of
inflammasomes, a group of multiprotein complexes categorized on the
basis of their major constituent, such as NLRP1, NLRP3 and AIM2,
triggers pyroptosis via caspase-1-mediated canonical inflammasome
signaling pathways. Internalized lipopolysaccharides, which
directly stimulate murine caspase-11 or its human homologs,
caspase-4 and −5, can also initiate pyroptosis through
non-canonical signaling pathways (4,12).
Inflammation exerts wide and sometimes contrasting
effects during carcinogenesis (13–15).
Pyroptotic cell lysis releases large amounts of damage-associated
molecular patterns (DAMPs) and proinflammatory cytokines, resulting
in inflammation. The involvement of targets or products of
pyroptotic pathways in tumorigenesis has been investigated,
focusing on diverse inflammasomes, inflammatory caspases, or
cytokines, such as IL-1β and IL-18. However, the conclusions are
controversial. For example, mice lacking NLRP3 are reported to be
hypersusceptible to colitis-associated colorectal cancer as shown
by several studies (16,17), whereas another study demonstrated
that a lack of NLRP3 attenuated DSS-induced colitis in mice
(18). Recently, accumulating
evidence tends to support a procarcinoma role of inflammation
(19,20).
Despite the critical role of GSDMD in pyroptosis and
inflammation, its precise relevance and function in cancer remains
unknown. In the present study, we investigated whether GSDMD has a
distinct biological role in NSCLC tumorigenesis.
Materials and methods
Tissue microarrays
Two commercial tissue microarrays (HLugA180Su02 and
HLugSqu150Sur01; Shanghai Outdo Biotech, Shanghai, China) were used
to evaluate GSDMD expression in LUAD and LUSC. Antigen retrieval
was performed by microwave heating in citrate buffer (pH 6.0) for 5
min. Microarrays were incubated with the primary antibody (GSDMD;
1:100; cat. no. sc-81868; Santa Cruz Biotechnology, Inc., Santa
Cruz, CA, USA) at 4°C overnight. Slides were analyzed separately by
two independent pathologists. Staining intensity was scored as
negative (0), weak (1), medium
(2) or strong as previously
described (3). An overall GSDMD
expression score was calculated by multiplying the intensity and
positive percentage scores.
Cell lines and cell culture
NSCLC cell lines (PC9, H1703, A549, SPC-A1, H1915,
H1975, H1299 and H1650) and a normal human bronchial epithelial
cell line (HBE) were purchased from the Institute of Biochemistry
and Cell Biology of the Chinese Academy of Sciences (Shanghai,
China). PC9/SPC-A1/HBE cells were cultured in Gibco™ DMEM medium
(Thermo Fisher Scientific, Inc., Waltham, MA, USA). H1703, A549,
H1915, H1975, H1299 and H1650 cells were cultured in RPMI-1640
medium (Gibco™). All culture media contained 10% fetal bovine serum
(FBS; HyClone Laboratories; GE Healthcare, Chicago, IL, USA). Cells
were cultured in a humidified atmosphere at 37°C with 5%
CO2.
Cell proliferation assays
Cell viability was assessed using a Cell
Proliferation Reagent Kit I (MTT; Roche, Basel, Switzerland).
Briefly, cells were cultured in the presence of 0.5 mg/ml MTT for 4
h. The medium was removed, 150 µl dimethyl sulfoxide (DMSO) was
added to each well and incubated at room temperature (RT) for 20
min before absorbance detection at 490 nm. For the clonogenic
assay, in order for cells to form colonies, 1,500 cells were
cultured on a 6-well plate. Following 2 weeks, the colonies were
fixed with methanol, stained with 0.1% crystal violet
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and captured with a
camera. The caspase inhibitor z-VAD-FMK was purchased from Selleck
Chemicals (Shanghai, China). Cells were pretreated with 30 µM
Z-VAD-FMK for 24 h as needed.
RNA extraction, cDNA synthesis and
reverse transcription quantitative PCR (RT-qPCR)
Total RNA was extracted using Invitrogen™ TRIzol
reagent (Thermo Fisher Scientific, Inc.), following the
manufacturers protocol. The cDNA was synthesized using a
PrimeScript RT-PCR kit (Takara Biotechnology Co., Ltd., Dalian,
China) and RT-qPCR was performed using a SYBR Premix Ex Taq II Kit
(Takara Biotechnology) on a QuantStudio 3 (Applied Biosystems;
Thermo Fisher Scientific, Inc.), according to the manufacturers
instructions. The specific primers used were: Actin forward,
TGACGTGGACATCCGCAAAG and reverse, CTGGAAGGTGGACAGCGAGG; GSDMD
forward, GAGTGTGGCCTAGAGCTGG and reverse, GGCTCAGTCCTGATAGCAGTG.
The qPCR conditions were 95°C for 30 sec, followed by 40 cycles
with denaturation at 95°C for 5 sec and annealing/elongation at
60°C for 30 sec. Relative expression fold changes were calculated
according to the 2−∆∆Cq method (21).
Cell transfection
Lipofectamine 2000 (Invitrogen™) was used to
transfect small interference RNA (siRNA) into cells following the
manufacturers instructions. Sequences of siRNAs were: GSDMD-1031:
GGAGACCAUCUCCAAGGAATT (sense), UUCCUUGGAGAUGGUCUCCTT (antisense),
GSDMD-1244: GGAACUCGCUAUCCCUGUUTT (sense), AACAGGGAUAGCGAGUUCCTT
(antisense).
Western blotting
Total protein was isolated using a lysis buffer
containing the mammalian protein extraction reagent RIPA (Beyotime
Institute of Biotechnology, Nantong, China), a protease inhibitor
cocktail (Roche), and PMSF (Roche). Samples were electrophoresed on
a 10% SDS-PAGE gel and electrotransferred onto nitrocellulose
membranes (EMD Millipore, Billerica, MA, USA). Membranes were
blocked with 5% skim milk, and then incubated with specific primary
antibodies overnight at 4°C and secondary antibodies for 1 h at
37°C. The ECL chromogenic substrate (EMD Millipore) was used to
detect specific bands. Specific primary antibodies included: GSDMD
(1:500; cat. no. sc-81868; Santa Cruz Biotechnology), PARP-1
(1:500; cat. no. sc-7150; Santa Cruz Biotechnology), caspase-3
(1:1,000; cat. no. cst-9662; Cell Signaling Technology, Inc.,
Billerica, MA, USA), caspase-1 (1:1,000; cat. no. ab108362; Abcam,
Cambridge, UK), Nlrp3 (1:1,000; cat. no. cst-5101; Cell Signaling
Technology, Inc.), Akt (1:1,000; cat. no. cst-4961; Cell Signaling
Technology, Inc.), p-Akt (1:1,000; cat. no. cst-4060; Cell
Signaling Technology, Inc.), mTOR (1:1,000; cat. no. cst-2983; Cell
Signaling Technology, Inc.), p-mTOR (1:1,000; cat. no. cst-5536;
Cell Signaling Technology, Inc.), p-EGFR (1:1,000; cat. no.
cst-3777; Cell Signaling Technology, Inc.), β-actin (1:5,000; cat.
no. cst-4970; Cell Signaling Technology, Inc.) and GAPDH (1:5,000;
cat. no. ab181602; Abcam). Secondary antibodies included: Goat
anti-mouse IgG-HRP (1:50,000; cat. no. abs20001; Absin, Shanghai,
China), goat anti-rabbit IgG-HRP (1:50,000; cat. no. abs20002;
Absin).
Flow cytometry for cell apoptosis
Cells were resuspended in binding buffer and stained
with FITC-Annexin V and propidium iodide (PI), using an Annexin
V-FITC Apoptosis Detection kit (BD Biosciences, San Jose, CA, USA),
following the manufacturers instructions. The cells were analyzed
by flow cytometry (BD Biosciences, Franklin Lakes, NJ, USA).
JC-1 test
Mitochondrial membrane potential (MMP) was
determined using a mitochondrial membrane potential detection kit
(JC-1) (KGA602; Nanjing KeyGen Biotech Co., Ltd., Nanjing, China)
according to the manufacturers instructions and analyzed by BD
Calibur flow cytometer (BD Biosciences).
Lactate dehydrogenase (LDH) assay
Cells transfected with siRNA were cultured in
96-well plates, and the release of LDH was determined using the
Pierce LDH cytotoxicity assay kit (cat. no. 88954; Thermo Fisher
Scientific, Inc.), in a time-dependent manner, according to the
manufacturer's instructions.
Enzyme-linked immunosorbent assay
(ELISA)
The concentrations of lL-1β in the culture
supernatants were determined using commercial Human lL-1β Valukine
ELISA kit (VAL101; R&D Systems, Minneapolis, MN, USA) following
the manufacturers instructions.
Inflammasome stimulation
For NLRP3 inflammasome stimulation, cells
transfected with siRNA were primed with 1 µg/ml lipopolysaccharide
(LPS) (Sigma-Aldrich; Merck KGaA) for 5 h and subsequently
incubated with 5 mM adenosine 5-triphosphate (ATP) (SunShine
Biotechnology, Nanjing, China) for 12 h prior to apoptosis
detection.
Lentiviral transduction
Lentiviral vectors encoding GSDMD shRNA (shGsd) and
negative control shRNA (shNeg) were purchased from GenePharma
Biotechnology Co., Ltd. (Shanghai, China). The lentiviral vectors
were added to tumor cells, and the culture medium was replaced with
fresh complete medium after 24 h. At 96 h post-transduction, cells
were harvested for further experiments. The sequence of the shGsd
was: GGAGACCATCTCCAAGGAACT. The sequence of the negative control
shRNA was: TTCTCCGAACGTGTCACGT.
Xenograft experiments
Five six-week-old male Balb/c nude mice (15–18 g)
were purchased from the Model Animal Research Center of Nanjing
University and were used for xenograft experiments. Animal care and
experimental protocols were approved by the Ethics Committee of the
Animal Research Center of Jinling Hospital, (project no. 81572273;
12 January 2017) and performed in strict accordance with the
Institutional Animal Care and Use guidelines. The mice were housed
under a 12-h light/dark cycle with constant temperature (22–25°C)
and relative humidity of 55%, and had free access to standard diet
and tap water. PC9 cells stably transduced with either shGsd or
shNeg were cultured for 96 h and then subcutaneously injected into
the right and left sides of the posterior flank of mice
(2×106 cells/side). Tumor volumes were determined every
three days until 18 days post-infection.
Immunohistochemistry (IHC) and
hematoxylin and eosin (H&E) staining
Tumor tissues were harvested from nude mice, fixed
in 4% paraformaldehyde and embedded in paraffin, with 5-µm sections
cut from the paraffin blocks. Staining was performed using Ki-67
(1:400; cat. no. cst-12202; Cell Signaling Technology, Inc.) and
H&E.
TUNEL staining
Cell death (apoptosis) in the tumor tissue was
detected in situ using a commercial terminal
deoxynucleotidyl transferase dUTP nick end-labeling TUNEL assay kit
(Roche). The TUNEL staining was performed following the
manufacturers protocol.
Bioinformatics analysis
A normalized Gene Expression Ombibus (GEO) array was
downloaded at MERAV database (available at http://merav.wi.mit.edu/) and co-expressing genes were
identified using Morpheus tools (available at: http://software.broadinstitute.org/morpheus/). KEGG
enrichment analysis was performed using the OmicShare tools
(available at: www.omicshare.com/tools).
Statistical analysis
Statistical analyses were performed using GraphPad
6.01 (GraphPad Software, Inc., La Jolla, CA, USA) and SPSS 22.0
(IBM Corp., Armonk, NY, USA) software programs. Comparisons between
two groups were performed by a two-tailed Students t-test.
Comparisons among multiple groups were performed by ANOVA test.
Bonferroni's method for equal variances and Games-Howell method for
unequal variances were used for further post-hoc testing. P<0.05
was considered to indicate a statistically significant
difference.
Results
Expression profile of GSDMD in human
NSCLC tissue
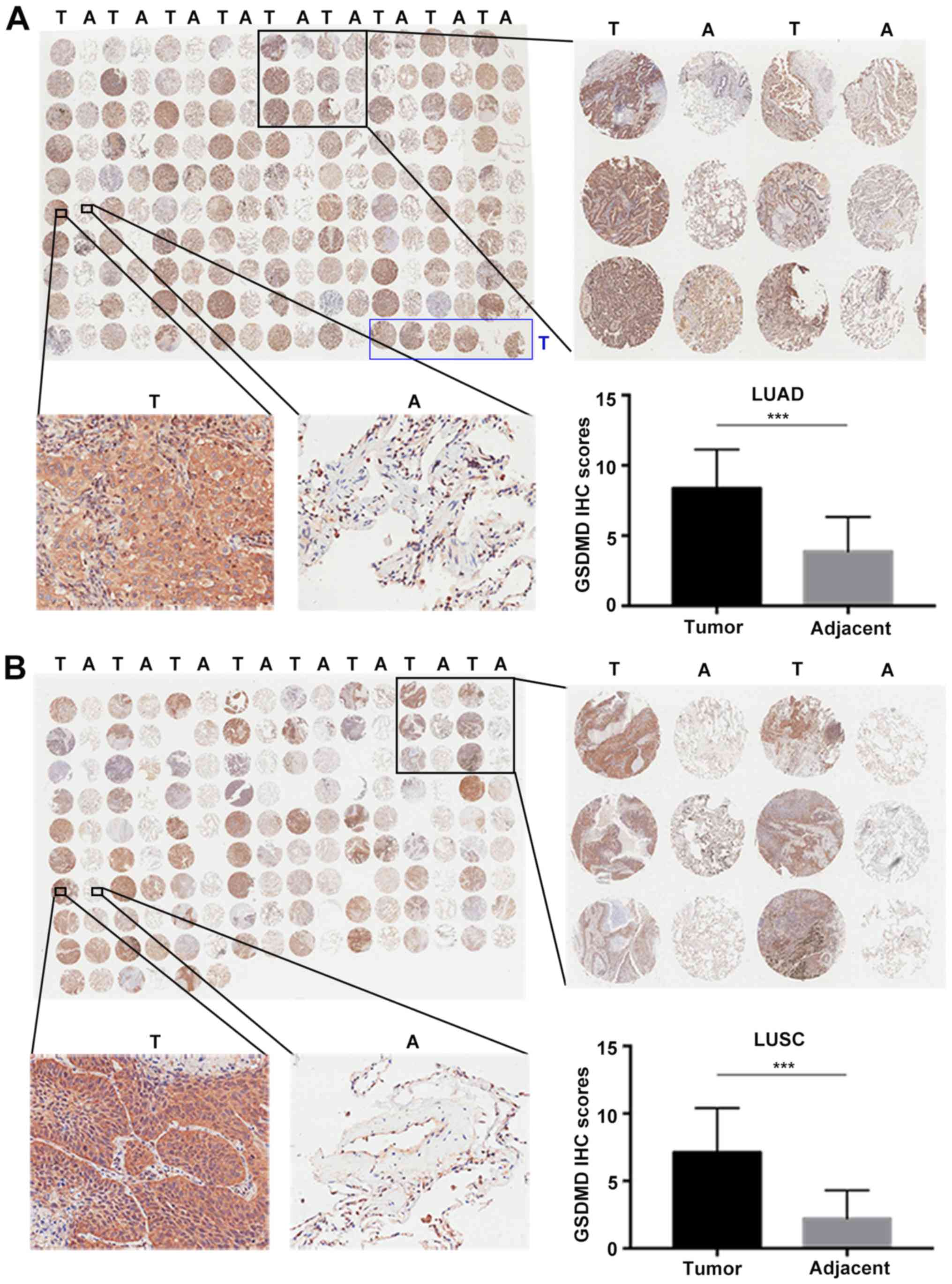
Two commercial tissue microarrays, including 93 LUAD
plus 87 matched adjacent tumor specimens and 75 paired LUSC, were
used to analyze the protein expression profile of GSDMD by IHC
(Fig. 1A and B). IHC scores were
defined as the product of intensity and positivity scores as
mentioned in ‘Materials and methods’ and as previously described
(3). GSDMD was predominantly
expressed in the cytoplasm of tumor cells, demonstrating
significant upregulation in both LUAD (P<0.001) (Fig. 1A) and LUSC (P<0.001) compared to
the adjacent tumor tissues (Fig.
1B).
Correlation between GSDMD expression,
clinicopathological characteristics and prognosis in NSCLC
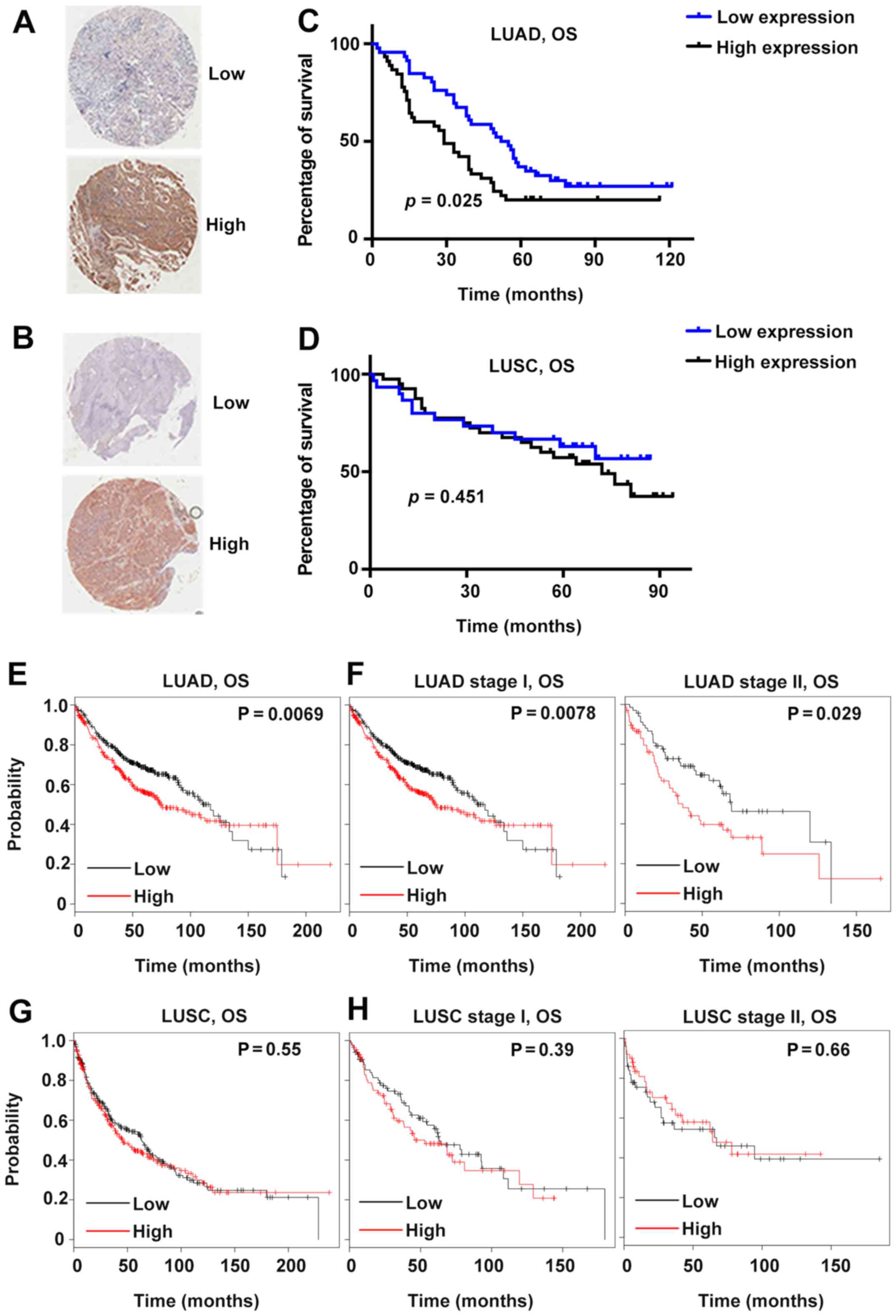
Patients were further divided into two groups based
on the average IHC scores. Specifically, the average score of LUAD
was 8.4; therefore, the patients with GSDMD IHC scores <8.4 were
allocated to the low-expression group, and the rest were assigned
to the high-expression group (Fig. 2A
and B). Patients with LUSC were grouped according to the same
principle, with a cut-off value of 7.1. Several clinicopathological
characteristics were analyzed, including age, sex, tumor size,
lymph node metastasis and tumor-node-metastasis (TNM) stages. GSDMD
protein expression was significantly associated with the tumor size
(P=0.045) in LUAD and with the TNM stages (P=0.048 for LUAD and
P=0.037 for LUSC) in both LUAD and LUSC (Table I).
 | Table I.Association between GSDMD protein
expression and clinicopathological characteristics of the NSCLC
cases. |
Table I.
Association between GSDMD protein
expression and clinicopathological characteristics of the NSCLC
cases.
|
| LUAD | LUSC |
|---|
|
|
|
|
|---|
|
Characteristics | Total | Low | High |
P-valuea | Total | Low | High |
P-valuea |
|---|
| All patients | 92 | 47 | 45 |
| 70 | 30 | 40 |
|
| Age, years |
|
<65 | 52 | 25 | 27 | 0.510 | 35 | 17 | 18 | 0.387 |
|
≥65 | 40 | 22 | 18 |
| 34 | 13 | 21 |
|
| Sex |
|
Male | 43 | 23 | 20 | 0.666 | 66 | 29 | 37 | 0.457 |
|
Female | 49 | 24 | 25 |
| 4 | 1 | 3 |
|
| Tumor size, cm |
| ≤3 | 34 | 22 | 12 | 0.045b | 18 | 9 | 9 | 0.558 |
|
>3 | 58 | 25 | 33 |
| 50 | 21 | 29 |
|
| Lymph node
metastasis |
| N0 | 38 | 21 | 17 | 0.298 | 37 | 20 | 17 | 0.066 |
|
N1-3 | 37 | 16 | 21 |
| 18 | 5 | 13 |
|
| TNM stage |
|
I–II | 45 | 27 | 18 | 0.048b | 39 | 20 | 19 | 0.037b |
|
III–IV | 30 | 11 | 19 |
| 15 | 3 | 12 |
|
Subsequently, the prognostic predictive value of
GSDMD was investigated. High GSDMD protein expression was
significantly associated with poor prognosis in LUAD (P=0.025,
Fig. 2C). However, the GSDMD
protein level revealed no association with the survival rate of
LUSC patients (Fig. 2D).
Furthermore, according to the Kaplan-Meier Plotter public database,
lower GSDMD mRNA expression indicated better survival in
patients suffering from either stage I or stage II LUAD (Fig. 2E and F). On the contrary, no obvious
association was observed between the survival of patients suffering
from LUSC and GSDMD mRNA expression profiles (Fig. 2G and H). Further multivariate
analysis indicated that GSDMD protein level was an independent
prognostic factor of LUAD (Table
II).
| Table II.Univariate and multivariate Cox
proportional hazards analysis for overall survival of LUAD
patients. |
Table II.
Univariate and multivariate Cox
proportional hazards analysis for overall survival of LUAD
patients.
|
|
| Univariate | Multivariate |
|---|
|
|
|
|
|
|---|
|
Characteristics |
| HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Age, years | <65/≥65 | 1.073
(0.669–1.720) | 0.771 |
|
|
| Sex | Male/female | 1.344
(0.837–2.157) | 0.221 |
|
|
| Tumor size, cm | ≤3/>3 | 1.520
(0.919–2.513) | 0.103 | 1.258
(0.705–2.244) | 0.437 |
| Lymph node
metastasis | N0/N1-3 | 2.637
(1.520–4.573) | 0.001b | 2.174
(1.005–4.701) | 0.049a |
| TNM stage | I–II/III–IV | 3.085
(1.777–5.356) |
<0.001c | 1.662
(0.776–3.563) | 0.191 |
| GSDMD level | Low/high | 1.706
(1.059–2.750) | 0.028a | 1.934
(1.096–3.412) | 0.023a |
Depletion of GSDMD attenuates the
proliferation of NSCLC cell lines
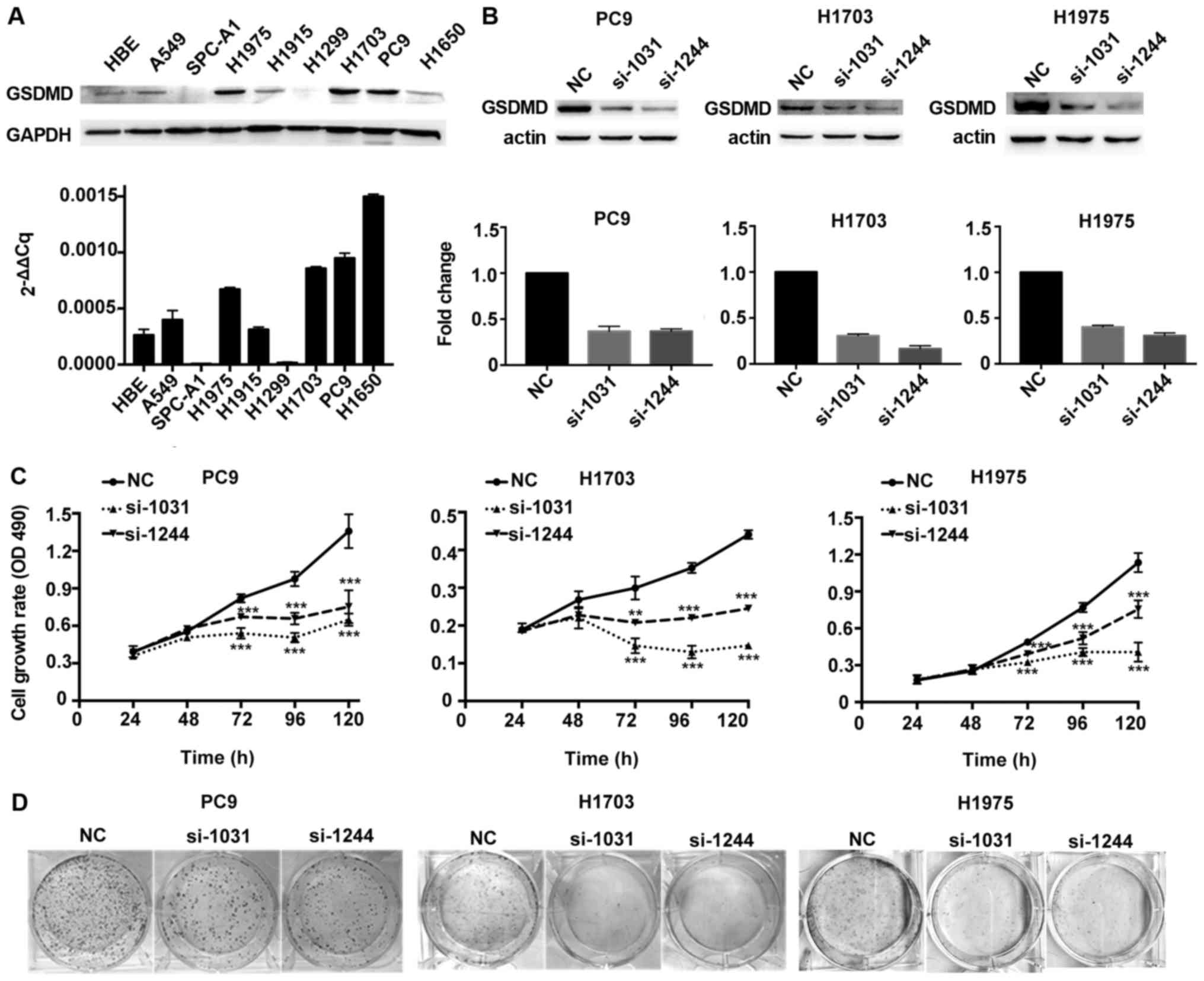
The biological function of GSDMD was assessed in
vitro by analysis of GSDMD protein and mRNA expression levels
in eight different human NSCLC cell lines, as well as in a normal
human bronchial epithelial (HBE) cell line. GSDMD was expressed in
HBE and most NSCLC cell lines, except for SPC-A1 and H1299. Protein
and mRNA expression profiles of GSDMD were consistent in all cell
lines except in H1650 (Fig. 3A).
PC9, H1703 and H1975 cell lines (human LUAD, LUSC and LUAD cell
lines, respectively) with high levels of GSDMD were used for
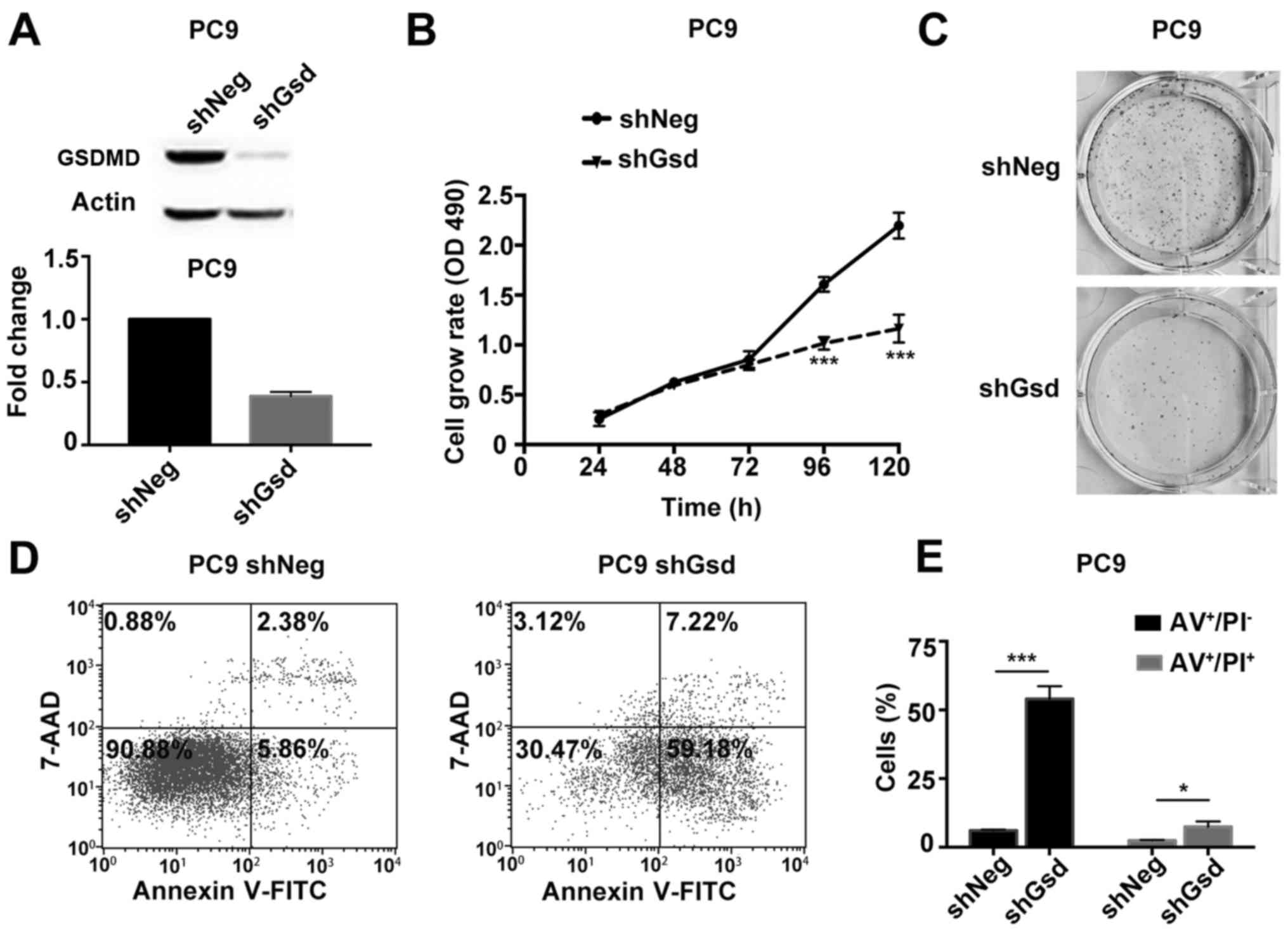
further investigation. Knockdown efficiency of GSDMD by two
artificial siRNAs is displayed in Fig.
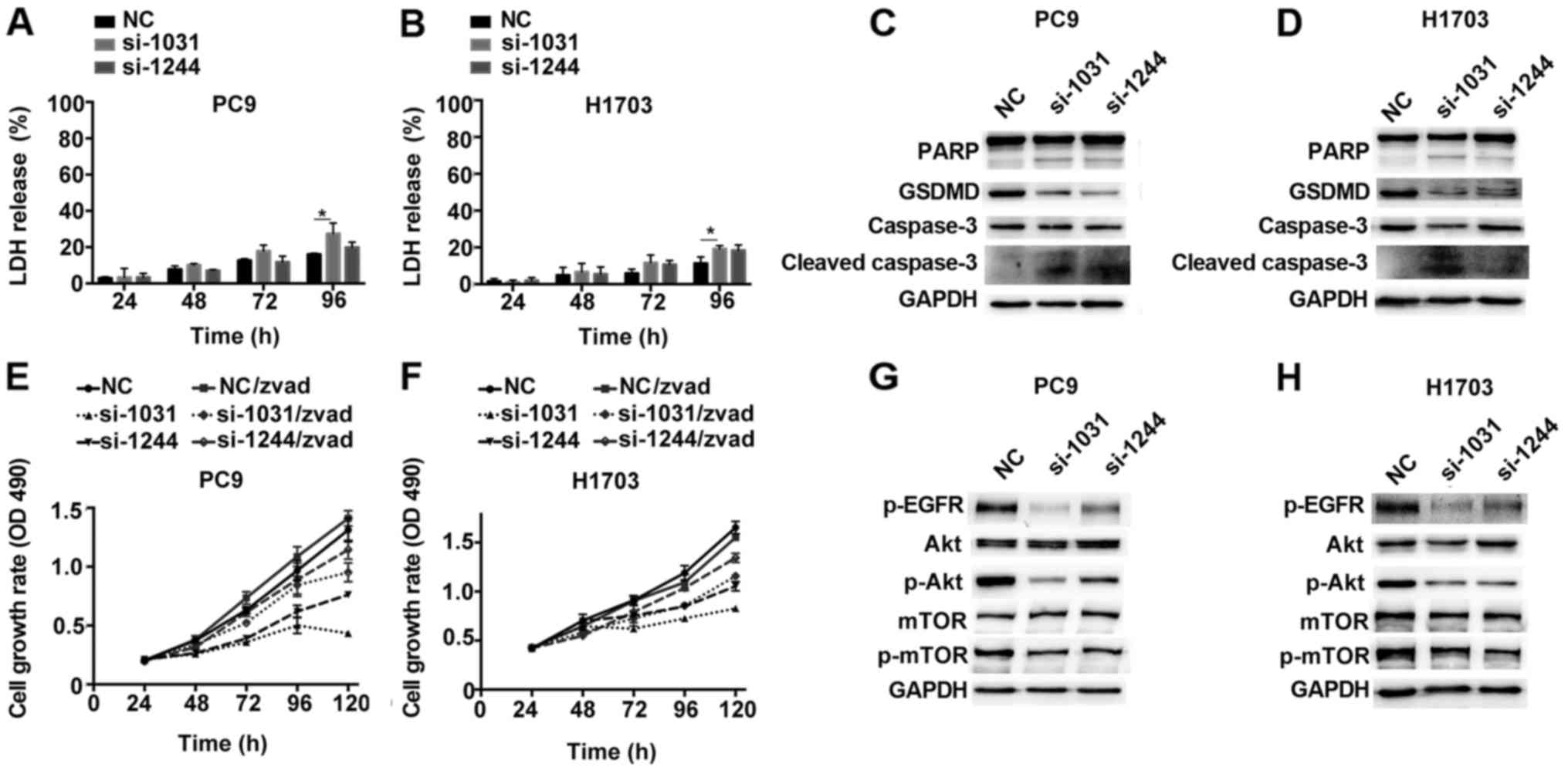
3B. The MTT assay demonstrated that depletion of GSDMD
significantly attenuated cell viability in all three cell lines
(Fig. 3C). Similarly, a colony
formation assay revealed fewer cell colonies in these three cell
lines after cells were treated with GSDMD siRNAs (Fig. 3D). Thus, these findings indicated
that knockdown of GSDMD inhibited the proliferation of NSCLC cells
in vitro.
Depletion of GSDMD facilitates the
apoptosis of NSCLC cell lines
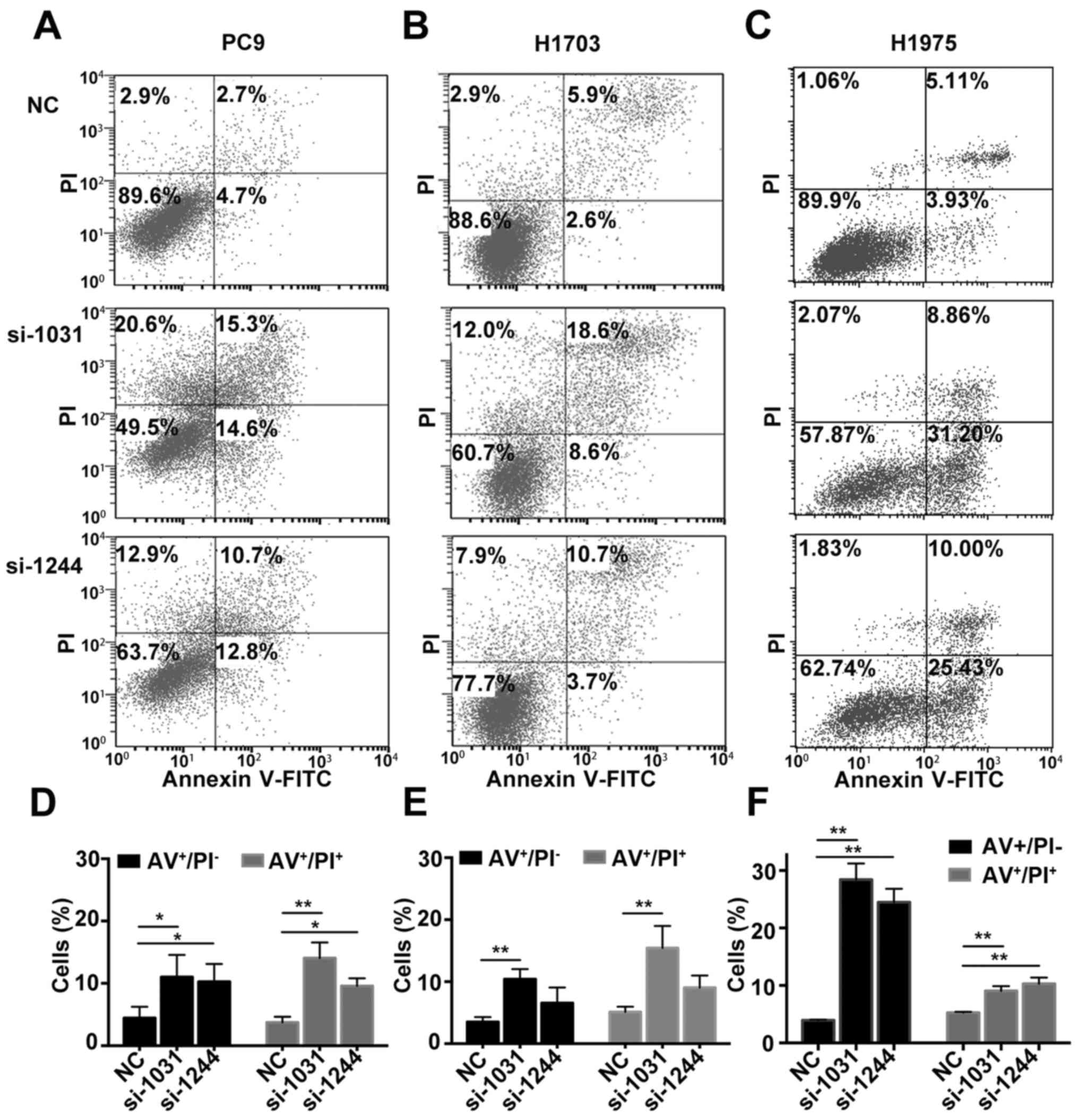
Subsequently, we investigated whether apoptosis was
activated in tumor cells in the absence of GSDMD. Combined staining
with Annexin V and PI indicated there were more Annexin
V+/PI− and Annexin
V+/PI+ cells in cell lines lacking GSDMD
(Fig. 4). Generally, Annexin
V+/PI− cells represent early apoptotic cells
and Annexin V+/PI+ cells represent late
apoptotic cells. As the pyroptotic cells also tend to be
simultaneously Annexin V+/PI+ (pyroptosis is
faster than apoptosis), an obvious increase in Annexin
V+/PI− cells indicated the increase in
apoptotic cells (22). The JC-1
test revealed that the mitochondrial membrane potential (MMP) was
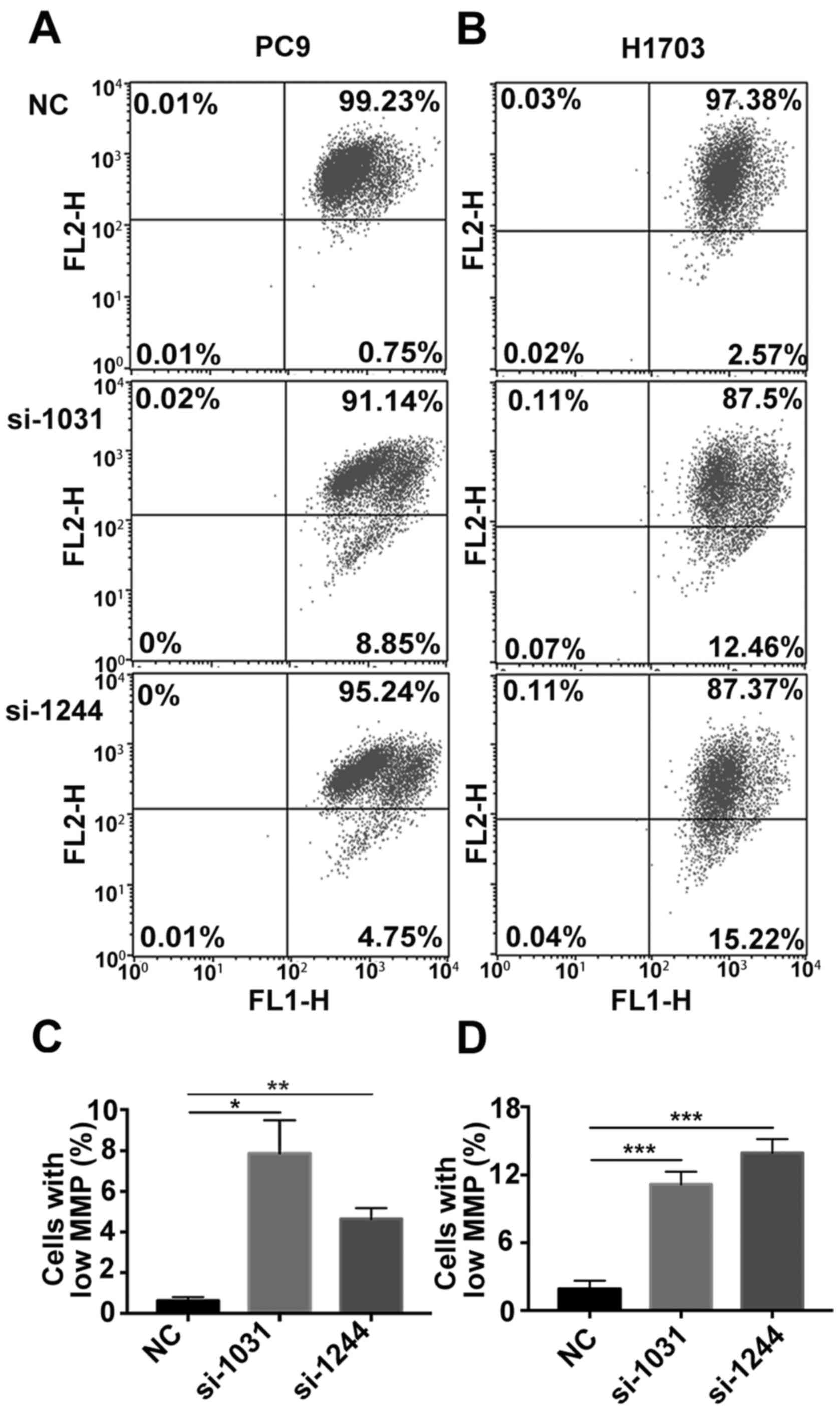
decreased following the knockdown of GSDMD (Fig. 5). These results indicated that
siRNA-induced depletion of GSDMD facilitated the intrinsic
mitochondrial apoptotic pathway.
NLRP3 inflammasome stimulation in
GSDMD-deficient cells induces apoptosis, instead of pyroptosis
Taabazuing et al recently described a
bidirectional crosstalk between pyroptosis and apoptosis in
monocytes and macrophages (23). In
response to pyroptotic stimuli, the upstream inflammasome/caspase-1
pathway may lead to the activation of apoptotic caspases when GSDMD
is absent in monocytes and macrophages (23,24).
We therefore, investigated whether the tumor cells shared the
similar mechanism for cell death.
Considering that GSDMD-deficient cancer cells
underwent apoptosis without any pyroptotic stimuli, we firstly
investigated whether there was an endogenous activation of
NLRP3/caspase-1 signaling pathway in cancer cells. Both PC9 and
H1703 cells, with either absent or scrambled GSDMD, revealed
detectable levels of NLRP3 by western blot analysis. Intact
caspase-1 and cleaved p20 caspase-1 were also expressed (Fig. 6A). We also detected the release of
IL-1β in the culture supernatant in PC9 cells by ELISA. PC9 cells
can release IL-1β spontaneously (control group), but in the absence
of GSDMD expression, PC9 cells demonstrated reduced secretion of
IL-1β [GSDMD contributes to the release of IL-1β (24)] (Fig.
6B). Collectively, these data indicated that there was an
endogenous activation of the NLRP3/caspase-1 signaling pathway in
tumor cells.
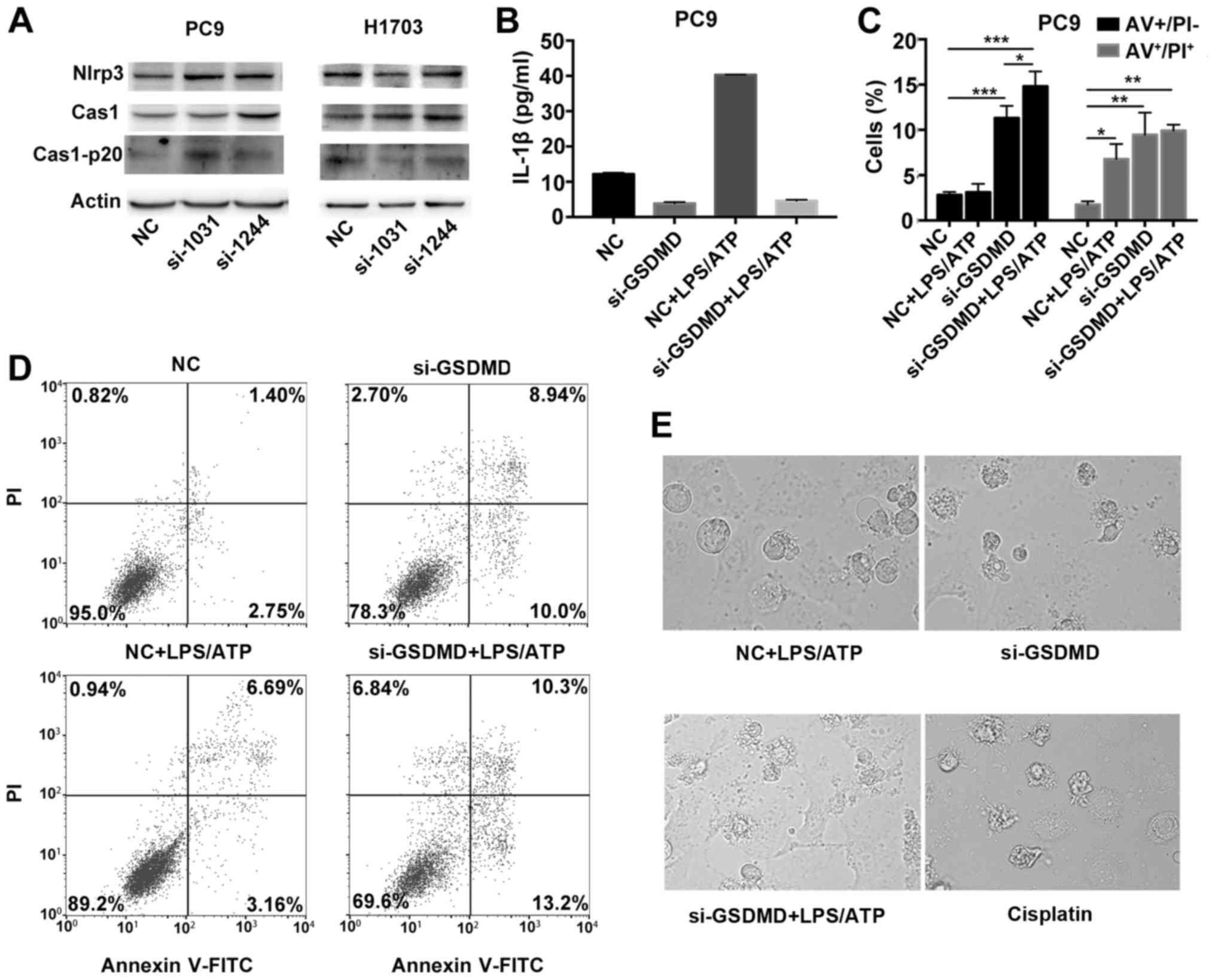 | Figure 6.NLRP3 inflammasome stimulation in
GSDMD-deficient cells induce apoptosis, instead of pyroptosis. (A)
Western blotting was used to examine the expression of proteins
upstream of GSDMD in classical pyroptotic pathways: NLRP3,
caspase-1 and its cleaved form in transfected PC9 and H1703 cells.
(B) PC9 cells transfected with indicated siRNAs (NC or si-GSDMD)
were primed with 1 µg/ml LPS for 5 h and subsequently incubated
with 5 mM ATP for 12 h. IL-1β levels in the supernatants of
cultured cell lines were detected by ELISA. Data are represented as
the mean ± SD (n=3). (C) Statistical analysis of Annexin V-FITC
(AV)+/PI− and AV+/PI+
PC9 cells treated as described in B. (D) Cell death was analyzed by
flow cytometry with Annexin V-FITC/PI double staining. Generally,
AV+/PI−, AV+/PI+,
AV−/PI− and AV−/PI+
indicate early apoptotic, late apoptotic, live and necrotic cells,
respectively. Notably, pyroptotic cells tend to be simultaneously
Annexin V/PI positive. (E) Representative light microscopy images
of PC9 cells treated as described in B. PC9 cells were also treated
with cisplatin at a concentration of 80 µg/ml for 24 h. *P<0.05,
**P<0.01, ***P<0.001 (ANOVA test). GSDMD, gasdermin D; LPS,
lipopolysaccharide; ATP, adenosine 5′-triphosphate. |
To further confirm that the activated pyroptotic
signaling, upstream of GSDMD, may contribute to the proapoptotic
phenotype in GSDMD-deficient lung cancer cells, we treated PC9
cells with an exogenous pyroptotic stimulus (LPS plus ATP) to
further activate NLRP3 inflammasomes. After NLRP3 inflammasome
stimulation, PC9 cells, transfected with the scrambled siRNA,
showed higher secretion of IL-1β (Fig.
6B). In addition, when treated wirh LPS/ATP, the cell line
demonstrated an increase in the Annexin
V+/PI+ cells, but no significant increase in
Annexin V+/PI− cells was observed compared to
the untreated group, indicating that without GSDMD knockdown, the
cells had undergone pyroptosis (pyroptotic cells tend to be
simultaneously Annexin V+/PI+, but are seldom
Annexin V+/PI−) following NLRP3 inflammasome
stimulation (Fig. 6C and D).
Furthermore, there was a high number of Annexin
V+/PI− and Annexin
V+/PI+ cells in the GSDMD-deficient PC9 cell
line whether or not treated with LPS/ATP, indicating that
GSDMD-deficient cells had undergone apoptosis either with or
without LPS/ATP stimulation (Fig. 6C
and D). In addition, cells transfected with GSDMD siRNA and
scrambled siRNA exhibited morphological differences following
LPS/ATP treatment. LPS/ATP-treated control cells exhibited a
swelling cellular morphology (characteristic of pyroptosis). While
LPS/ATP-treated GSDMD-deficient cells were more likely to have
apoptotic morphology similar to cisplatin-treated PC9 cells
(Fig. 6E). These data further
confirmed that NLRP3 inflammasome stimulation (intrinsic or
extrinsic activation) in GSDMD-deficient cells induced apoptosis
instead of pyroptosis.
Depletion of GSDMD activates
caspase-3-mediated apoptosis, which is partially reversed by
caspase inhibition
To further confirm that cancer cells lacking GSDMD
underwent apoptotic pathway, distinct from the lytic cell death
pathway (such as pyroptosis), we observed the release of LDH over
time. The expected release of LDH caused by toxicity of
Lipofectamine used for siRNA transfection was detected 24 h
post-transfection, but no obvious increase in the LDH release was
observed after GSDMD knockdown at early time-points compared to
those observed in cells transfected with the scrambled siRNA
(Fig. 7A and B). Notably, although
LDH release is a hallmark of lytic cell death, the slight increase
in LDH release at 96 h post-transfection was likely due to
secondary necrosis of apoptotic cells (25).
Initiator caspases lead to the processing of
executioner caspase-3, subsequently mediating the apoptotic
cascade, including PARP cleavage (26). Western blotting assay indicated that
GSDMD knockdown was associated with the activation of caspase-3 and
inactivation of PARP, as indicated by the presence of their cleaved
forms (Fig. 7C and D).
We used z-VAD-FMK, a general caspase inhibitor, to
pretreat the cells before siRNA transfection. Caspase inhibition by
z-VAD-FMK partially alleviated the decreased cell proliferation
induced by GSDMD depletion (Fig. 7E and
F). At the last time-point (120 h), PC9 cells treated with
GSDMD siRNAs demonstrated 33% (si-1031) and 58% (si-1244) cell
viability, while PC9 cells which were treated with GSDMD siRNAs and
also pretreated with z-VAD-FMK maintained 67% (si-1031) and 81%
(si-1244) cell viability, although significant inhibition of
proliferation still remained (Fig.
7E). H1703 cell lines exhibited similar outcomes (Fig. 7F). Collectively, these results
revealed that the knockdown of GSDMD led to the induction of
apoptosis by activation of caspase-3, which was partially reversed
by the caspase inhibition.
GSDMD modulates the EGFR/Akt signaling
pathway
As the z-VAD-FMK inhibitor could not completely
reverse the apoptosis of GSDMD-deficient cells, there may be other
key pathways, which are involved in cell proliferation, that are
altered. We downloaded a normalized Gene Expression Ombibus (GEO)
array (available at http://merav.wi.mit.edu/) that includes 79 NSCLC
samples and analyzed genes that were co-expressed with GSDMD
using Morpheus tools (available at: http://software.broadinstitute.org/morpheus/). A total
of 584 co-expressed genes with a Pearson's correlation coefficient
equal to or >0.3 were identified and subsequent KEGG enrichment
analysis, using the OmicShare tools (available at: www.omicshare.com/tools), showed that the
co-expressed genes were enriched in several classical signal
transduction pathways (Table
III). The top five hits were Rap1, ErbB, PI3K-Akt,
phospholipase D, and MAPK signaling pathways, of which we chose the
PI3K-Akt signaling pathways (P=0.0077) for further investigation
because we focused on the regulation of cell proliferation and
apoptosis by GSDMD, and the PI3K-Akt signaling pathway is mainly
involved in regulation of cell survival and apoptosis (27).
| Table III.Pathway enrichment analysis based on
coexpressing genes of GSDMD from expression Ombibus (GEO)
array. |
Table III.
Pathway enrichment analysis based on
coexpressing genes of GSDMD from expression Ombibus (GEO)
array.
| Pathway | Out (119) | All (8,248) | P-value | Q-value |
|---|
| Rap1 signaling
pathway | 8 | 157 | 0.001895346 | 0.05907162 |
| ErbB signaling
pathway | 5 | 67 | 0.002720708 | 0.06275739 |
| Hippo signaling
pathway | 4 | 52 | 0.006542435 | 0.10558077 |
| PI3K-Akt signaling
pathway | 9 | 240 | 0.007739042 | 0.10558077 |
| Phospholipase D
signaling pathway | 5 | 100 | 0.01458475 | 0.15151935 |
| MAPK signaling
pathway | 7 | 190 | 0.01981771 | 0.15933242 |
| Hippo signaling
pathway | 5 | 109 | 0.02044908 | 0.15933242 |
| Ras signaling
pathway | 6 | 167 | 0.03352433 | 0.20583959 |
| TGF-β signaling
pathway | 3 | 67 | 0.07217501 | 0.2871644 |
| NF-κB signaling
pathway | 3 | 90 | 0.1404493 | 0.39659881 |
| Calcium signaling
pathway | 4 | 141 | 0.1462627 | 0.40222242 |
| Wnt signaling
pathway | 3 | 103 | 0.1858804 | 0.4634618 |
| MAPK signaling
pathway | 1 | 17 | 0.2190935 | 0.51861373 |
| Jak-STAT signaling
pathway | 3 | 130 | 0.2891426 | 0.60752434 |
| HIF-1 signaling
pathway | 2 | 76 | 0.3000821 | 0.62350392 |
| AMPK signaling
pathway | 2 | 84 | 0.3425983 | 0.66347952 |
| cAMP signaling
pathway | 3 | 147 | 0.3564909 | 0.66347952 |
| FoxO signaling
pathway | 2 | 100 | 0.4248353 | 0.72222001 |
| VEGF signaling
pathway | 1 | 39 | 0.4333989 | 0.73014049 |
| Sphingolipid
signaling pathway | 1 | 78 | 0.6798369 | 0.80974204 |
| TNF signaling
pathway | 1 | 90 | 0.7315566 | 0.81262773 |
| cGMP-PKG signaling
pathway | 1 | 118 | 0.8222264 | 0.86379964 |
As predicted, western blotting revealed
downregulation of phosphorylated Akt (p-Akt) in PC9 and H1703 cells
lacking GSDMD. Phosphorylated EGFR (p-EGFR), an upstream mediator,
was also distinctly downregulated, while a downstream target,
phosphorylated mTOR (p-mTOR) was slightly downregulated (Fig. 7G and H). This data indicated that
GSDMD could modulate the EGFR/Akt signaling pathway in NSCLC cell
lines.
Depletion of GSDMD suppresses tumor
growth in xenograft mouse models
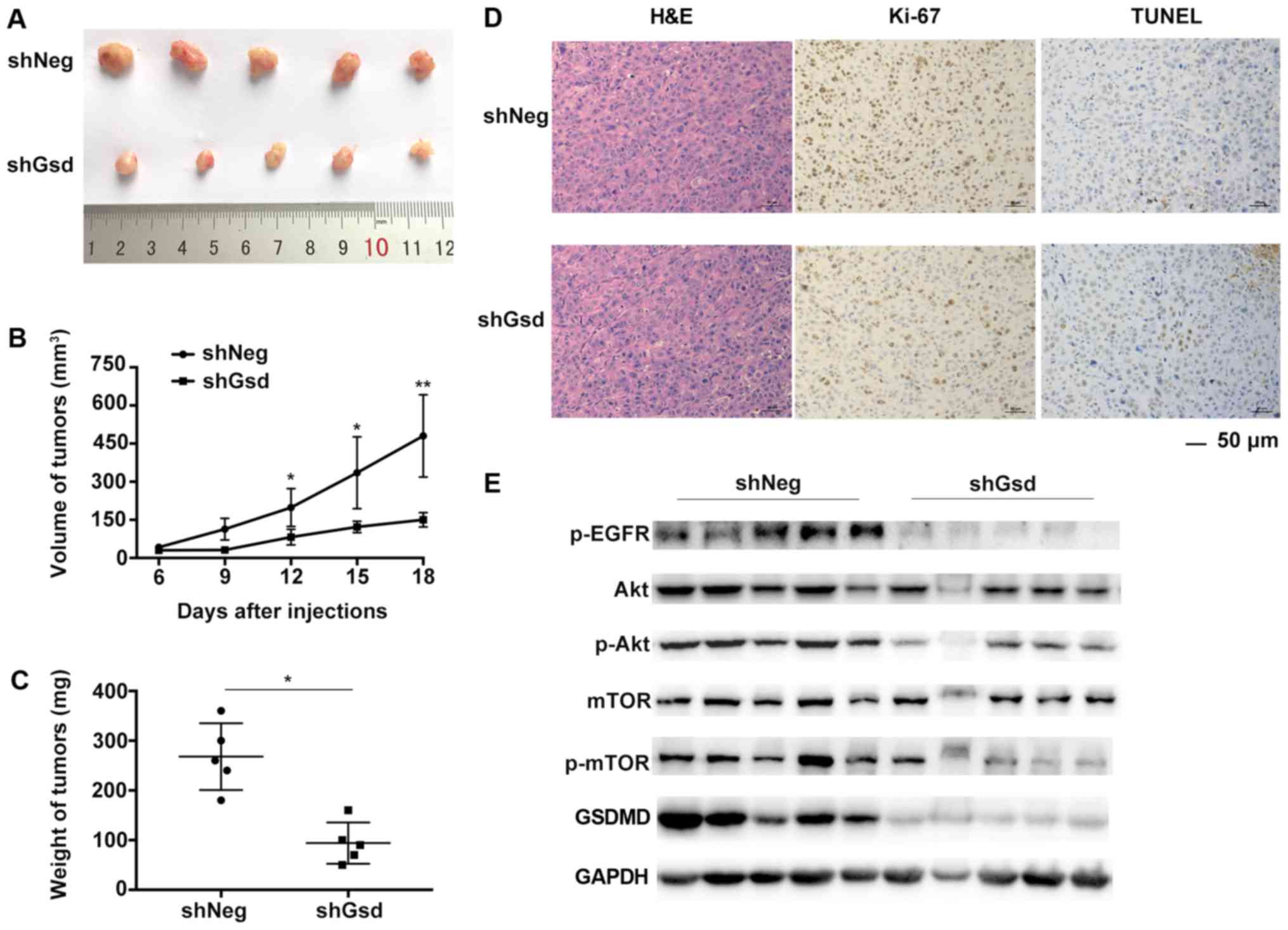
Subsequently, we validated the tumor growth
suppression by GSDMD depletion in a PC9 mouse xenograft model. PC9
cells were stably transduced with shNeg or shGsd lentiviral
vectors. PC9 cells transduced with shGsd demonstrated decreased
cell proliferation and increased cell apoptosis in vitro as
displayed in Fig. 8. Six-week-old
male Balb/c nude mice were injected subcutaneously with PC9 cells
transduced with shGsd or shNeg. Tumor volumes were determined every
3 days until 18 days post infection. Eighteen days following
injection, growth of the PC9 tumors was significantly inhibited in
the shGsd group (Fig. 9A). Both
tumor volumes and weights were significantly lower in the shGsd
group (Fig. 9B and C). In addition,
decreased Ki-67 staining and increased TUNEL staining were observed
during IHC analysis of shGsd derived tumors (Fig. 9D). The evidence provided in the
present study indicated that depletion of GSDMD can suppress tumor
growth in vivo. Furthermore, as predicted, western blotting
revealed downregulation of p-EGFR/p-Akt/p-mTOR levels in shGsd
tumors (Fig. 9E).
Discussion
Pyroptosis is a pro-inflammatory form of regulated
cell death. GSDMD is a newly characterized pyroptotic executioner
(28) and its homologous GSDM
family members (GSDMA-C, DFNA5) have been reported to play a
potential role in several types of tumors (4). Our study elucidated the role of GSDMD
in tumorigenesis for the first time and indicated its
procarcinogenic function in NSCLC. We demonstrated by MTT and
colony assays that depletion of GSDMD inhibited NSCLC growth in
PC9, H1703 and H1975 cell lines. Despite the fact that GSDMD could
execute pyroptosis in monocytes and macrophages, GSDMD-deficient
cancer cells seemed to induce another classical programed cell
death pathway (apoptosis) that is generally considered
immunologically silent.
Apoptotic cells show early positive Annexin V
staining due to active phosphatidylserine (PS) exposure on the
outer plasma membrane leaflet, whereas pyroptosis involved rapid
membrane rupture and pore formation, allowing early Annexin
V+/PI+ staining (22). Thus, Annexin V staining alone does
not discriminated between apoptosis and pyroptosis. However,
combined Annexin V and PI staining may be used to distinguish
between them. We classified the form of cell death using combined
staining. Both Annexin V+/PI− and Αnnexin
V+/PI+ cells, indicative of early and late
apoptosis, respectively, were increased in GSDMD-deficient cells.
Mitochondrial outer membrane permeability is a hallmark of
intrinsic apoptotic pathway (29).
Similarly, our JC-1 data confirmed that the MMP was significantly
decreased in GSDMD-depleted cells. Subsequent LDH release assays
revealed non-lytic apoptotic cell death, although secondary
necrosis was detected at later stages.
Crosstalk between pyroptosis and apoptosis has been
observed in several recent studies, mainly concentrating on
monocytes and macrophages (23,24,30).
Potentially, molecules upstream of GSDMD in the pyroptotic pathway
may be responsible for the induction of apoptosis in the absence of
GSDMD. AIM2 and NLRP3 inflammasomes have been reported to activate
pro-caspase-8 and induce apoptosis in caspase-1 null macrophages
(30). Additionally, contribution
of caspase-1 was confirmed by a higher activity of apoptotic
caspases in GSDMD null macrophages compared to the macropahges
lacking caspase-1 (24). Another
study further elucidated the role of caspase-1 in activating
caspase-3 and −7 using DPP8/9 inhibitors in monocytes and
macrophages (23).
Although inflammation is a fundamental pathological
process mainly involving immune cells, activated inflammasomes
concurrent with inflammatory responses have been described in tumor
cells (31). Subsequently we
investigated whether apoptotic GSDMD-deficient cancer cells shared
the same apoptotic mechanism as GSDMD-deficient immune cells.
Notably, GSDMD-deficient NSCLC cells underwent apoptosis without
any exogenous stimuli, while GSDMD-deficient immune cells were
found to be apoptotic when cocultured with a pyroptotic stimulus.
Thus, we hypothesized that there may be a mechanism of endogenous
activation of pyroptotic signaling in cancer cells. Western blot
results showed that NLRP3 and cleaved caspase-1 were intrinsically
expressed in PC9 and H1703 cells without any stimuli, regardless of
GSDMD status. Release of IL-1β in culture supernatant was also
detected in PC9 cells.
Perhaps, the most critical question to be asked is
why tumor cells had endogenous activation of pyroptotic signaling
without undergoing pyroptotic cell death. In fact, this phenomenon
has previously been reported in melanoma cells. Constitutive
cleaved caspase-1, IL-1β production and secretion were observed in
melanoma cells without exogenous stimuli (20).
We also investigated the crosstalk between apoptosis
and pyroptosis in PC9 cells with an exogenous stimulus (ATP/LPS).
Early Annexin+/PI+ staining and swollen
morphology of the cells indicated pyroptotic characteristics in PC9
cells treated with scrambled siRNA and ATP/LPS. Both Annexin
V+/PI− and Annexin
V+/PI+ cells, and cells with a shriveled
morphology indicated apoptotic characteristics in PC9 cells treated
with GSDMD siRNA and whether or not treated with ATP/LPS. Thus, we
suggested that intrinsic or extrinsic activation of NLRP3/caspase-1
signaling in GSDMD-deficient cells induced apoptosis, instead of
pyroptosis. Furthermore, increased expression of cleaved caspase-3
and PARP was induced in GSDMD-knockdown cells, confirming the
activation of apoptotic signaling. Cells pretreated with z-VAD-FMK,
the broad-spectrum caspase inhibitor, demonstrated decreased growth
inhibition compared with untreated GSDMD-deficient cells; however,
significant growth inhibition was still observed in
z-VAD-FMK-treated GSDM.D-knockdown cells. Incomplete inhibition by
z-VAD-FMK is a possible reason, although additional pathways
modified by GSDMD cannot be excluded.
The PI3K/Akt pathway has long been recognized as an
important regulatory pathway for a wide array of cellular processes
including metabolism, growth or survival and apoptosis (32). Coexpression analysis based on a
published GEO array identified the co-expression of genes involved
in the PI3K/Akt signaling pathway with GSDMD. EGFR mutation
and increased copy numbers are common in NSCLC, activating
downstream PI3K/Akt/mTOR signaling and resulting in uncontrolled
growth and cell proliferation (33). Evidence that GSDMD could modify this
vital pathway in cancer cells was provided by the apparent
downregulation of p-EGFR and p-Akt, as well as a slight decrease in
p-mTOR. However, to the best of our best knowledge, there are no
studies reporting the correlation between GSDMD and the PI3K/Akt
signaling pathway in cancer or other diseases. It may be that
knockdown of GSDMD impairs the inflammatory responses, thus
interrupting the negative feedback regulation for activating the
PI3K/Akt pathway (32), but it
could also be that the full length GSDMD has a role in regulating
this pathway, particularly in cancer cells.
Clinical data indicated that GSDMD was upregulated
in NSCLC. GSDMD is an independent prognostic biomarker for LUAD
based on our tissue array and Kaplan-Meier plot database analysis.
However, although correlation analysis demonstrated that GSDMD
overexpression was associated with a more aggressive phenotype in
LUSC, no significant prognostic difference existed in LUSC.
Notably, disparity between preclinical and clinical data may be
attributed to the expression of GSDMD in the tumor
microenvironment, particularly in immune cells; for example,
macrophages may have a more important role in regulating
inflammation. However, the inherent apoptotic cell death in LUSC
initiated by blocking expression of GSDMD in tumor cells should not
be ignored and it may contribute to the targeted therapy and
apoptosis-associated resistance in cancer treatment.
In summary, we demonstrated that expression of GSDMD
by tumor cells is involved in regulating the proliferation in
NSCLC. Clinical analysis indicated the association of GSDMD
expression with the tumor size and TNM stages. GSDMD is an
independent prognostic biomarker in LUAD, but not LUSC. Knockdown
of GSDMD induced apoptosis by activating caspase-3, which can be
partially rescued by caspase inhibition. Crosstalk between
pyroptosis and apoptosis may be attributed to intrinsically or
extrinsically active upstream pyroptotic signaling. Coexpression
analysis indicates the modification of the PI3K.Akt signaling
pathway by GSDMD.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (nos. 81401903,
81572273 and 81602015), the Natural Science Foundation of Jiangsu
province (BK20161386) and the Jiangsu Provincial Special Program of
Medical Science (BL2013026). The funders had no role in the study
design, data collection and analysis, the decision to publish, or
preparation of the manuscript.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
TL and YS conceived and designed the experiments;
JG, XQ and GX performed the experiments. JG and XQ analyzed the
data. HL and FZ acquired the reagents, the materials and the
analysis tools. JG wrote the paper. All authors read and approved
the manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Animal care and experimental protocols were approved
by the Animal Research Center of Jinling Hospital (project no.
81572273, 12 January 2017; Nanjing, P.R. China) and were performed
in strict accordance with the Institutional Animal Care and Use
guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
GSDMD
|
gasdermin D
|
|
GSDM
|
gasdermin
|
|
NSCLC
|
non-small cell lung cancer
|
|
LUAD
|
lung adenocarcinoma
|
|
LUSC
|
lung squamous cell carcinoma
|
|
DAMPs
|
damage-associated molecular
patterns
|
|
MMP
|
mitochondrial membrane potential
|
|
shNeg
|
negative control shRNA
|
|
shGsd
|
GSDMD shRNA
|
|
IHC
|
immunohistochemistry
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
LDH
|
lactate dehydrogenase
|
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reck M and Rabe KF: Precision diagnosis
and treatment for advanced non-small-cell lung cancer. N Engl J
Med. 377:849–861. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Saeki N, Usui T, Aoyagi K, Kim DH, Sato M,
Mabuchi T, Yanagihara K, Ogawa K, Sakamoto H, Yoshida T and Sasaki
H: Distinctive expression and function of four GSDM family
genes (GSDMA-D) in normal and malignant upper
gastrointestinal epithelium. Genes Chromosomes Cancer. 48:261–271.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shi J, Gao W and Shao F: Pyroptosis:
Gasdermin-mediated programmed necrotic cell death. Trends Biochem
Sci. 42:245–254. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kayagaki N, Stowe IB, Lee BL, O'Rourke K,
Anderson K, Warming S, Cuellar T, Haley B, Roose-Girma M, Phung QT,
et al: Caspase-11 cleaves gasdermin D for non-canonical
inflammasome signalling. Nature. 526:666–671. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shi J, Zhao Y, Wang K, Shi X, Wang Y,
Huang H, Zhuang Y, Cai T, Wang F and Shao F: Cleavage of GSDMD by
inflammatory caspases determines pyroptotic cell death. Nature.
526:660–665. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fink SL and Cookson BT: Apoptosis,
pyroptosis, and necrosis: Mechanistic description of dead and dying
eukaryotic cells. Infect Immun. 73:1907–1916. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Aglietti RA, Estevez A, Gupta A, Ramirez
MG, Liu PS, Kayagaki N, Ciferri C, Dixit VM and Dueber EC: GsdmD
p30 elicited by caspase-11 during pyroptosis forms pores in
membranes. Proc Natl Acad Sci USA. 113:7858–7863. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen X, He WT, Hu L, Li J, Fang Y, Wang X,
Xu X, Wang Z, Huang K and Han J: Pyroptosis is driven by
non-selective gasdermin-D pore and its morphology is different from
MLKL channel-mediated necroptosis. Cell Res. 26:1007–1020. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ding J, Wang K, Liu W, She Y, Sun Q, Shi
J, Sun H, Wang DC and Shao F: Pore-forming activity and structural
autoinhibition of the gasdermin family. Nature. 535:111–116. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sborgi L, Ruhl S and Mulvihill E: GSDMD
membrane pore formation constitutes the mechanism of pyroptotic
cell death. EMBO J. 35:1766–1778. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wallach D, Kang TB, Dillon CP and Green
DR: Programmed necrosis in inflammation: Toward identification of
the effector molecules. Science. 352:aaf21542016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zitvogel L, Kepp O, Galluzzi L and Kroemer
G: Inflammasomes in carcinogenesis and anticancer immune responses.
Nat Immunol. 13:343–351. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Karki R, Man SM and Kanneganti TD:
Inflammasomes and cancer. Cancer Immunol Res. 5:94–99. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Munn LL: Cancer and inflammation. Wiley
Interdiscip Rev Syst Biol Med. 9:2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Allen IC, TeKippe EM, Woodford RM, Uronis
JM, Holl EK, Rogers AB, Herfarth HH, Jobin C and Ting JP: The NLRP3
inflammasome functions as a negative regulator of tumorigenesis
during colitis-associated cancer. J Exp Med. 207:1045–1056. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zaki MH, Vogel P, Body-Malapel M, Lamkanfi
M and Kanneganti TD: IL-18 production downstream of the Nlrp3
inflammasome confers protection against colorectal tumor formation.
J Immunol. 185:4912–4920. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bauer C, Duewell P, Mayer C, Lehr HA,
Fitzgerald KA, Dauer M, Tschopp J, Endres S, Latz E and Schnurr M:
Colitis induced in mice with dextran sulfate sodium (DSS) is
mediated by the NLRP3 inflammasome. Gut. 59:1192–1199. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Diakos CI, Charles KA, McMillan DC and
Clarke SJ: Cancer-related inflammation and treatment effectiveness.
Lancet Oncol. 15:e493–e503. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhai Z, Liu W, Kaur M, Luo Y, Domenico J,
Samson JM, Shellman YG, Norris DA, Dinarello CA, Spritz RA, et al:
NLRP1 promotes tumor growth by enhancing inflammasome activation
and suppressing apoptosis in metastatic melanoma. Oncogene.
36:3820–3830. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vande Walle L and Lamkanfi M: Pyroptosis.
Curr Biol. 26:R568–R572. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Taabazuing CY, Okondo MC and Bachovchin
DA: Pyroptosis and apoptosis pathways engage in bidirectional
crosstalk in monocytes and macrophages. Cell Chem Biol.
24(507–514): e5042017.
|
|
24
|
He WT, Wan H, Hu L, Chen P, Wang X, Huang
Z, Yang ZH, Zhong CQ and Han J: Gasdermin D is an executor of
pyroptosis and required for interleukin-1beta secretion. Cell Res.
25:1285–1298. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Silva MT: Secondary necrosis: The natural
outcome of the complete apoptotic program. FEBS Lett.
584:4491–4499. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yuan J, Najafov A and Py BF: Roles of
caspases in necrotic cell death. Cell. 167:1693–1704. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yip PY: Phosphatidylinositol
3-kinase-AKT-mammalian target of rapamycin (PI3K-Akt-mTOR)
signaling pathway in non-small cell lung cancer. Transl Lung Cancer
Res. 4:165–176. 2015.PubMed/NCBI
|
|
28
|
Aglietti RA and Dueber EC: Recent insights
into the molecular mechanisms underlying pyroptosis and gasdermin
family functions. Trends Immunol. 38:261–271. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jorgensen I, Rayamajhi M and Miao EA:
Programmed cell death as a defence against infection. Nat Rev
Immunol. 17:151–164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sagulenko V, Thygesen SJ, Sester DP, Idris
A, Cridland JA, Vajjhala PR, Roberts TL, Schroder K, Vince JE, Hill
JM, et al: AIM2 and NLRP3 inflammasomes activate both apoptotic and
pyroptotic death pathways via ASC. Cell Death Differ. 20:1149–1160.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kong H, Wang Y, Zeng X, Wang Z, Wang H and
Xie W: Differential expression of inflammasomes in lung cancer cell
lines and tissues. Tumour Biol. 36:7501–7513. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Weichhart T and Saemann MD: The
PI3K/Akt/mTOR pathway in innate immune cells: Emerging therapeutic
applications. Ann Rheum Dis. 67 Suppl 3:iii70–iii74. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Papadimitrakopoulou V: Development of
PI3K/AKT/mTOR pathway inhibitors and their application in
personalized therapy for non-small-cell lung cancer. J Thorac
Oncol. 7:1315–1326. 2012. View Article : Google Scholar : PubMed/NCBI
|