Introduction
Esophageal cancer is the eighth most common
malignancy in the world and the fifth most commonly diagnosed
cancer in China (1,2). Esophageal squamous cell cancer (ESCC)
is the main histological subtype of esophageal cancer in China.
Spicy food consumption, alcohol drinking and smoking increase the
risk of esophageal cancer (3). Due
to insidious early symptoms, ESCC is often diagnosed at advanced
stages. Despite advances in multimodal therapy, the 5 year survival
rate of ESCC patients remains at ~10–15% (4). Therefore, it is urgent to better
understand the molecular mechanisms and identify biomarkers for the
diagnosis, prognosis and treatment of ESCC.
The human brain expressed X-linked gene family
consists of BEX1, BEX2, BEX3, BEX4 and BEX5, which are highly
conserved in phylogeny (5,6). The human BEX family proteins were
first identified in the central nervous system and play an
important role in neuronal differentiation and regeneration.
Further studies have revealed that BEX family members are involved
in the regulation of the cell cycle and apoptosis as well as muscle
differentiation. Emerging evidence has demonstrated that BEX family
proteins are distinctly expressed among different types of cancers,
but their roles in tumorigenesis are currently controversial. BEX1
is overexpressed in acute myeloid leukemia (AML) with mixed lineage
leukemia (MLL) mutation, indicating that BEX1 plays a role in
leukemogenesis (7). However, BEX1
expression was observed to be downregulated in a group of FMS-like
tyrosine kinase-3 (FLT3-ITD) driven AML patients (8). Ding et al reported that BEX1
was downregulated in imatinib-resistant chronic myeloid leukemia
K562 cell line and overexpression of BEX1 re-sensitized the cells
to imatinib-induced apoptosis (9).
Gene expression analyses have revealed that BEX1 promoter is
hypermethylated in intracranial ependymoma, malignant glioma and
oral squamous cell carcinoma (OSCC) (10–12).
The differential expression and promoter hypermethylation indicated
that BEX1 may be a potential tumor suppressor. However, the
expression status and clinical significance of BEX1 expression as
well as its role in ESCC remains to be determined.
In the present study, we determined BEX1 protein
expression in the tumor and adjacent normal tissues from 118 ESCC
patients and evaluated its relationship with clinicopathological
parameters and prognosis. We observed that BEX1 was downregulated
in late tumor stage ESCC tissues and low expression of BEX1
predicted poor prognosis of patients with ESCC. Furthermore, we
assessed BEX1 biological functions in ESCC by in vitro and
in vivo assays. We revealed that overexpression of BEX1
inhibited ESCC cell proliferation in vitro and tumor growth
in mouse xenografts. In addition, we observed that BEX1
overexpression inhibited NF-κB activation. Our results indicated
that BEX1 may function as a tumor suppressor in ESCC and may serve
as a potential target for therapies of ESCC.
Materials and methods
Patients and tissue samples
Tumor tissues and adjacent normal tissues from 118
ESCC patients were collected from Binzhou Medical University
Hospital from January 1, 2009 to December 31, 2010. None of the
patients had received neoadjuvant therapies prior to surgery.
Clinical data including sex, age, smoking history, drinking
history, tumor location, tumor size, tumor differentiation, T
stage, lymph node status and clinical stage, were obtained from
medical records. The pathological clinical stages of esophageal
cancer were classified according to the 7th edition of the
UICC/AJCC TNM system. Survival time was determined from the date of
surgery to the date of death. Patient follow-ups were conducted
every 3–5 months until December 2016 for all patients. This study
was approved by the Ethics Committee of Binzhou Medical University
Hospital and informed written consent was obtained from every
patient.
Immunohistochemistry
All specimens were embedded in paraffin and
sectioned at a thickness of 4 µm. Tissue sections were
deparaffinized in a series of alcohols and heated in citrate buffer
(pH 6.0) for antigen retrieval in a microwave oven. Following
blocking of nonspecific binding sites with normal goat blood serum,
the sections were incubated with BEX1 antibody (dilution 1:50; cat.
no. AP10669c; Abgent Biotech Co., Ltd., Suzhou, China) at 4°C
overnight. The sections were washed with phosphate-buffered saline
(PBS) and incubated with horseradish peroxidase (HRP) conjugated
secondary antibody (cat. no. PV-6000; OriGene Technologies, Inc.,
Beijing, China) for 30 min at room temperature followed by 3 washes
with PBS. The sections were then subjected to 3,3′-diaminobenzidine
(DAB; ZSGB-Bio, Beijing, China) staining. The sections were
counterstained with hematoxylin (ZSGB-Bio) and placed on coverslips
with neutrogum nutral balsam (Sinopharm Chemical Reagent Co., Ltd.,
Shanghai, China). Stained specimens were evaluated by two
independent pathologists. For scoring, five fields were randomly
selected from each tissue section and at least 100 cells were
counted. BEX1 immuno-intensity was classified into three
categories: 0, no staining or weak; 1, moderate; 2, strong. The
percentage of stained cells was scored as follows: 1, 0–25%; 2,
26–50%; 3, 51–75%; 4, 76–100%. The final score was calculated by
multiplying the staining intensity and the percentage of staining
cells. The expression level of BEX1 was graded as low (scores, 1–4)
or high (scores, 5–8).
Cell lines and cell culture
Human ESCC cell lines (Eca-109 and TE-1) were
obtained from the Chinese Academy of Medical Sciences (Shanghai,
China). Human EC9706 cells were purchased from Fenghuibio
(Changsha, China). Cell lines were cultured in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
containing 10% fetal bovine serum (FBS; Tianhang Biotechnology Co.,
Ltd., Huzhou, China). A normal human esophageal epithelial cell
line (SHEE) was purchased from Fenghuibio (Changsha, China) and
cultured in Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo
Fisher Scientific, Inc.) with 10% FBS. All cells were incubated in
5% CO2 atmosphere at 37°C and were routinely passaged
every 2 or 3 days.
RNA isolation, reverse transcription
and real-time PCR analysis (RT-PCR)
Total RNA was isolated from tissues or cells using
an RNeasy Mini kit (Qiagen GmbH, Hilden, Germany). cDNA was
synthesized using the PrimeScript RT reagent kit (Takara Bio, Inc.,
Otsu, Japan). BEX1 mRNA expression was quantified using SYBR Premix
Ex Taq II (Takara Bio, Inc.) and performed with an ABI 7500 System.
The primers were as follows: BEX1 forward,
5′-CCTCCCTTTGGATGCTGGTGAAT-3′ and reverse,
5′-CTCATCCTTGCCTGTGGTTCTCC-3′; GAPDH forward,
5′-TGACTTCAACAGCGACACCCA-3′ and reverse,
5′-CACCCTGTTGCTGTAGCCAAA-3′. GAPDH was used as an internal control.
RT-PCR reactions were performed at 95°C for 4 min followed by 40
cycles of 95°C for 30 sec, 58°C for 25 sec and 72°C for 30 sec.
Western blot analysis
Following washing twice with cold PBS, the cells
were lysed on ice in RIPA lysis buffer for 30 min followed by
centrifugation with 12,000 × g at 4°C for 15 min. The supernatant
total protein was quantified by BCA Protein Assay reagent (Pierce;
Thermo Fisher Scientific, Inc.). An equal amount of total protein
(30–50 µg) was separated on 8–15% SDS-polyacrylamide gel
electrophoresis and then transferred to 0.22-µm polyvinylidene
fluoride membranes (PVDF) (EMD Millipore, Billerica, MA, USA),
which were then blocked with blocking buffer (5% skim milk in
Tris-buffered saline with 0.5% Tween-20, TBST) at room temperature
for 1 h. Following blocking, the membranes were incubated overnight
with primary anti-BEX1 antibody (1:300; cat. no. AP10669c),
anti-AKT antibody (1:500; cat. no. AP7028B), anti-phosphorylated
AKT antibody (1:500; cat. no. AP3434a; Abgent Biotech Co.),
anti-p65 antibody (1:100; cat. no. sc-72675; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), anti-phosphorylated p65
(p-p65) (1:100; cat. no. sc-136548; Santa Cruz Biotechnology,
Inc.), or anti-GAPDH antibody (1:10,000; cat. no. KC-5G4; KangChen
Bio-tech, Shanghai, China) at 4°C. The HRP-conjugated secondary
antibodies (1:10,000; cat. nos. KC-MM-035 and KC-RB-035; KangChen
Bio-tech) were diluted at 1:5,000 and incubated with membranes at
room temperature for 1 h. The bands were visualized using EZ-ECL
kit (Biological Industries, Kibbutz Beit Haemek, Israel). GAPDH was
used as the loading control. The densitometry of Western blotting
band were analyzed by ImageJ software 1.8.0 (National Institutes of
Health, Bethesda, MD, USA).
Cell transfection and generation of
stable cells
BEX1 expression vector and lentivirus expressing
BEX1 were purchased from Vigene Biosciences (Jinan, China). Eca-109
and TE-1 cells were transfected with BEX1 expression construct or
an empty vector control via Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's
instructions. Eca-109 and TE-1 cells were transduced with medium
containing lentivirus. Stable cells were established by puromycin
selection and BEX1 expression was confirmed via RT-PCR and western
blotting.
Cell proliferation assay
The role of BEX1 on cell proliferation was evaluated
by MTT assay. Briefly, cells were seeded in 96-well plates at a
density of 5×103 cells/well and cultured for 24 h in a
5% CO2 atmosphere at 37°C. Then, MTT solution was added
to each well at a concentration of 5 mg/ml and incubated at 37°C
for 4 h. Subsequently, the culture solution was removed and DMSO
was added. The absorbance was determined at 490 nm with a
spectrophotometer.
Colony formation assay
A colony formation assay was performed in a 6-well
plate. Up to 500 cells were plated in one well and cultured in 4 ml
of RPMI-1640 medium (with 10% FBS) for two weeks. Then, the
colonies were fixed and stained with 0.5% crystal violet for 20
min. Cell clusters consisting of >50 cells were defined as a
colony and all the colonies were counted under a light
microscope.
Cell cycle and apoptosis analysis
Cells were harvested and fixed in 70% ethanol at 4°C
for 24 h. Following washing with cold PBS, the cells were incubated
with RNase at 37°C for 30 min, stained with propidium iodide (PI)
for 30 min, and subjected to cell cycle distribution analysis using
a flow cytometer (Beckman Coulter, Brea, CA, USA). For apoptosis
analysis, the cells were stained with both FITC-Annexin V and PI
(BioLegend, Inc., San Diego, CA, USA) and subjected to flow
cytometric analysis.
Xenograft tumor assay
Five-week-old, female nude mice were obtained from
the Shanghai Laboratory Animal Center (Shanghai, China) and used to
establish xenograft models. In brief, 1×106 Eca-109
cells stably expressing BEX1 or empty vector were suspended in 150
µl of PBS and subcutaneously injected into the flank of nude mice.
The length (L) and width (W) of the tumor were measured every 4
days and the tumor volume was calculated as follows: L × W × W/2.
The tumor weight was determined after the mice were sacrificed by
cervical dislocation. The Animal Committee of Binzhou Medical
University approved all the experimental protocols and animal
handling procedures. All experimental procedures and postoperative
animal care were conducted in accordance with the guidelines for
the Care and Use of Laboratory Animals from the National Institutes
of Health (Bethesda, MD, USA).
NF-κB-luciferase reporter assay
ESCC cells stably expressing BEX1 were transfected
with an NF-κB-luciferase expression vector (Genomeditech, Shanghai,
China). At 24 h post-transfection, the cells were stimulated with
or without 15 ng/ml TNFα (Sangon Biotech Co., Ltd., Shanghai,
China) for 24 h. The cells were collected and analyzed for
luciferase activities using the Dual-Luciferase Reporter assay
system (Promega Corp., Madison, WI, USA). Luciferase activity
values were normalized to the expression level of the control
vector.
Statistical analysis
All data were statistically analyzed by SPSS
software 16.0 (SPSS, Inc., Chicago, IL, USA). A Chi-square test was
used to assess the relationship between the expression of BEX1
protein and the clinicopathological characteristics. The
Kaplan-Meier method was used to draw the survival curve and the
log-rank test was performed to analyze the survival data. Cox's
proportional hazards model was performed to analyze variables
associated with survival. All data were represented as the mean ±
standard deviation (SD). Student's t-tests were used to compare the
data between the two groups. P<0.05 was considered to indicate a
statistically significant difference.
Results
The expression of BEX1 in ESCC tumor
tissues and cell lines
The protein expression of BEX1 was determined in 118
pairs of ESCC tumor and adjacent normal tissues via
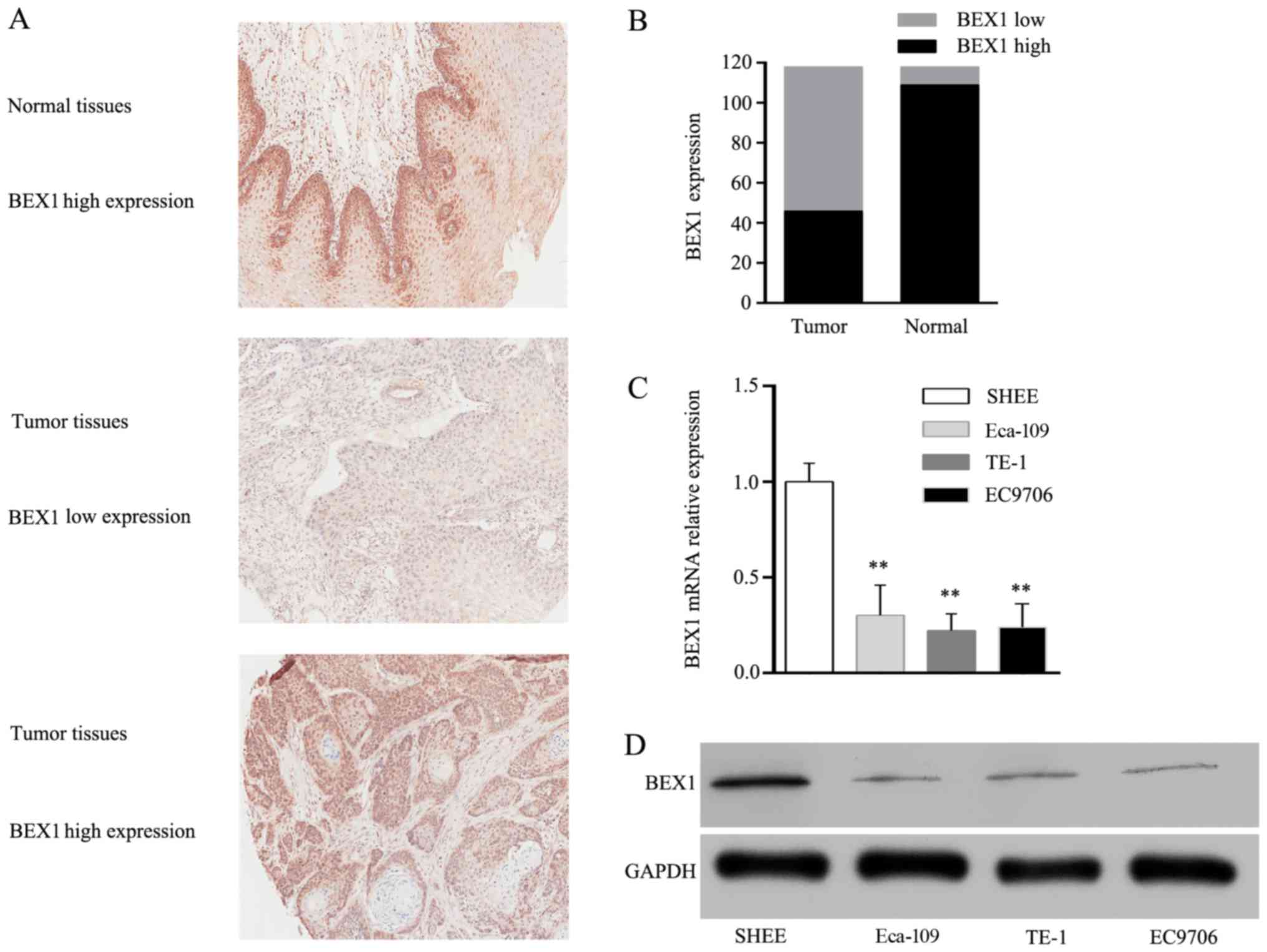
immunohistochemical analysis. As displayed in Fig. 1A, the immunoreactivity of BEX1 was
observed in the nucleus and the cytoplasm. Low BEX1 expression was
observed in 72 cases of tumor tissues (61.02%) and 9 cases of
normal tissues (7.63%, Fig. 1B). We
further assessed the expression of BEX1 in normal human esophageal
epithelial cells (SHEE) and three ESCC cell lines (TE-1, Eca-109
and EC9706). RT-PCR analysis revealed that BEX1 mRNA expression in
ESCC cell lines was significantly lower than that of SHEE cells
(Fig. 1C). Consistently, western
blot analysis revealed that BEX1 protein levels in ESCC cell lines
were significantly lower than that of SHEE cells (Fig. 1D). These results revealed BEX1 to be
downregulated in ESCC.
Association of BEX1 expression with
clinicopathological characteristics of ESCC
Subsequently, we classified the 118 ESCC patients
into the BEX1 low-expression group (n=72) and high-expression group
(n=46), and systematically analyzed the association of BEX1
expression with clinicopathological characteristics via chi-square
test. The clinicopathological variables and BEX1 expression are
listed in Table I. There was a
statistically significant negative association between the
expression level of BEX1 and tumor length (P<0.001), T stage
(P=0.011) and pTNM stage (P=0.039) (Table I). However, there was no significant
association of BEX1 expression with sex (P=0.688), age (P=0.312),
smoking history (P=0.266), alcohol drinking history (P=0.099),
tumor location (P=0.604), differentiation (P=0.077) and N stage
(P=0.178). These data indicated that low expression of BEX1
predicted poor prognosis of ESCC patients and that BEX1 may be an
independent negative prognostic factor for overall survival
rates.
 | Table I.Relationship between the number of
cases with BEX1 expression and the clinicopathological variables of
ESCC patients. |
Table I.
Relationship between the number of
cases with BEX1 expression and the clinicopathological variables of
ESCC patients.
|
|
| BEX1 expression
(%) |
|---|
|
|
|
|
|---|
| Variables | Case number
(n=118) | Low (n=72) | High (n=46) | P-value |
|---|
| Sex |
|
|
| 0.688 |
|
Male | 97 | 60 (61.9) | 37 (38.1) |
|
|
Female | 21 | 12 (57.1) | 9 (42.9) |
|
| Age (years) |
|
|
| 0.312 |
|
<60 | 60 | 39(65.0) | 21 (35.0) |
|
|
≥60 | 58 | 33 (56.9) | 25 (43.1) |
|
| Smoking
history |
|
|
| 0.266 |
| Never
or light | 44 | 24 (54.5) | 20 (45.5) |
|
|
Heavy | 74 | 48 (64.9) | 26 (35.1) |
|
| Alcohol drinking
history |
|
|
| 0.099 |
| Never
or light | 48 | 25 (52.1) | 23 (47.9) |
|
|
Heavy | 70 | 47 (67.1) | 23 (32.9) |
|
| Tumor location |
|
|
| 0.604 |
|
Cervical | 1 | 1 (100.0) | 0 (0.0) |
|
| Upper
thoracic | 12 | 6 (50.0) | 6 (50.0) |
|
| Middle
thoracic | 60 | 39 (65.0) | 21 (35.0) |
|
| Lower
thoracic | 45 | 26 (57.8) | 19 (42.2) |
|
| Tumor length
(cm) |
|
|
| <0.001 |
|
<4 | 57 | 22 (38.6) | 35 (61.4) |
|
| ≥4 | 61 | 50 (82.0) | 11 (18.0) |
|
|
Differentiation |
|
|
| 0.077 |
|
Poor | 19 | 15 (78.9) | 4 (21.1) |
|
|
Moderate | 53 | 34 (64.2) | 19 (35.8) |
|
|
Well | 46 | 23 (50.0) | 23 (50.0) |
|
| T stage |
|
|
| 0.011 |
| T1 | 10 | 3 (30.0) | 7 (70.0) |
|
| T2 | 17 | 7 (41.2) | 10 (58.8) |
|
| T3 | 69 | 44 (63.8) | 25 (36.2) |
|
| T4 | 22 | 18 (81.8) | 4 (18.2) |
|
| N stage |
|
|
| 0.178 |
| N0 | 58 | 32 (55.2) | 26 (44.8) |
|
| N1 | 26 | 15 (57.7) | 11 (42.3) |
|
| N2 | 29 | 20 (69.0) | 9 (31.0) |
|
| N3 | 5 | 5 (100.0) | 0 (0.0) |
|
| pTNM stage |
|
|
| 0.039 |
| I | 16 | 5 (31.2) | 11 (68.8) |
|
| II | 45 | 27 (60.0) | 18 (40.0) |
|
|
III | 57 | 40 (70.2) | 17 (29.8) |
|
Low expression of BEX1 is associated
with poor clinical outcome of ESCC patients
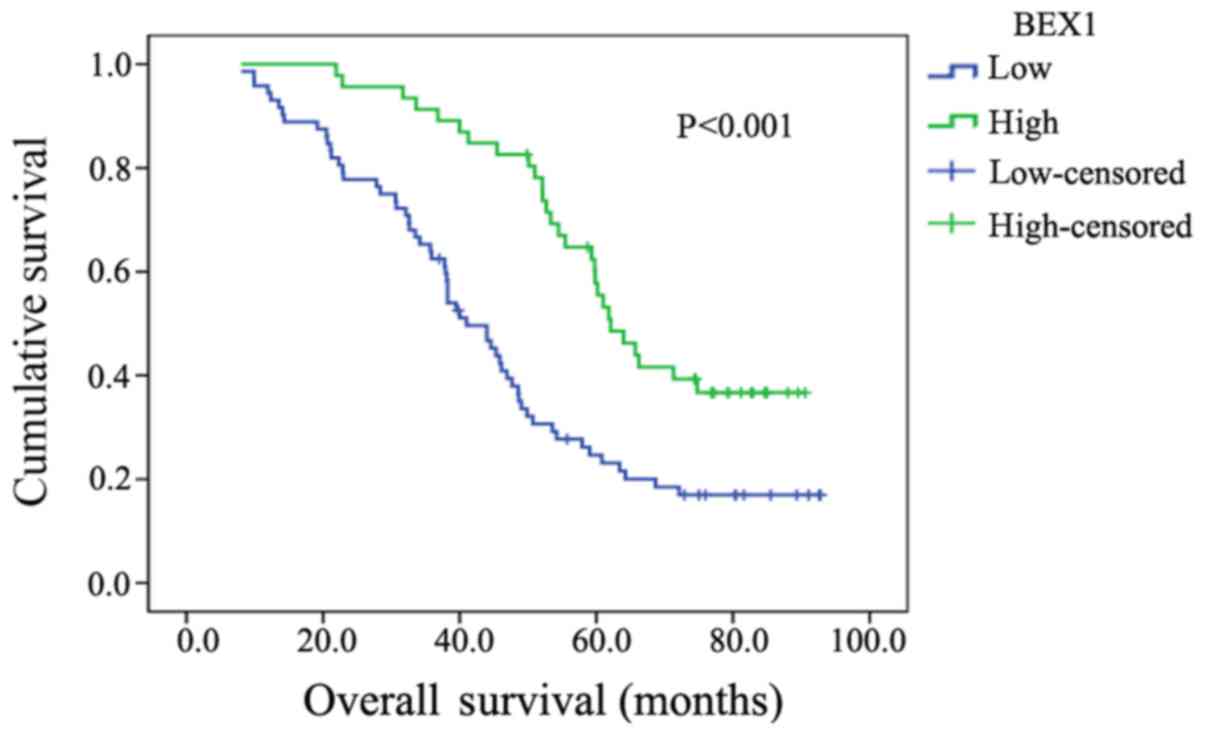
We further performed Kaplan-Meier analysis to
determine BEX1 expression level in association to patient survival
time. The 5-year survival rate of ESCC patients with high BEX1
expression was 63.04% (29/46), while that of patients with low BEX1
expression was 29.17% (21/72). The difference was statistically
significant (P<0.001; Fig. 2).
Univariate COX regression analysis demonstrated that BEX1
(P<0.001), tumor length (P=0.002), T stage (P<0.001), N stage
(P<0.001) and pTNM stage (P<0.001) were statistically
associated with overall survival (Table II). Multivariate COX regression
analysis revealed that the expression level of BEX1 (P=0.039) as
well as T stage (P=0.034), N stage (P=0.007) and pTNM stage
(P=0.025), were independent prognostic factors for overall survival
of ESCC patients (Table II).
| Table II.Univariate and multivariate analysis
of survival associated factors. |
Table II.
Univariate and multivariate analysis
of survival associated factors.
|
| OS (5 year) |
|---|
|
|
|
|---|
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Parameters | HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Sex (male vs.
female) | 0.732
(0.413–1.299) | 0.286 | 0.549
(0.272–1.110) | 0.095 |
| Age (<60 vs. ≥60
years) | 1.066
(0.698–1.627) | 0.767 | 1.289
(0.821–2.014) | 0.272 |
| Smoking (never vs.
heavy) | 1.392
(0.882–2.197) | 0.155 | 1.403
(0.741–2.660) | 0.299 |
| Drinking (never vs.
heavy) | 1.178
(0.761–1.824) | 0.463 | 0.594
(0.318–1.108) | 0.102 |
| Tumor length (<4
vs. ≥4cm) | 2.134
(1.327–3.431) | 0.002 | 1.070
(0.595–1.926) | 0.821 |
| Differentiation
(poor vs. moderate and well differentiated) | 1.145
(0.645–2.031) | 0.645 | 0.619
(0.329–1.163) | 0.136 |
| T stage (T1-2 vs.
T3-4) | 3.497
(1.886–6.484) | 0.000 | 2.298
(1.108–4.765) | 0.034 |
| N stage (N0 vs.
N1-3) | 2.361
(1.517–3.675) | 0.000 | 1.875
(1.191–2.952) | 0.007 |
| pTNM stage (I, II
vs. III) | 2.825
(1.807–4.419) | 0.000 | 2.160
(1.061–4.401) | 0.025 |
| BEX1 (low vs.
high) | 0.425
(0.269–0.670) | 0.000 | 0.565
(0.329–0.971) | 0.039 |
BEX1 overexpression inhibits the
proliferation and colony formation capacity of ESCC cells
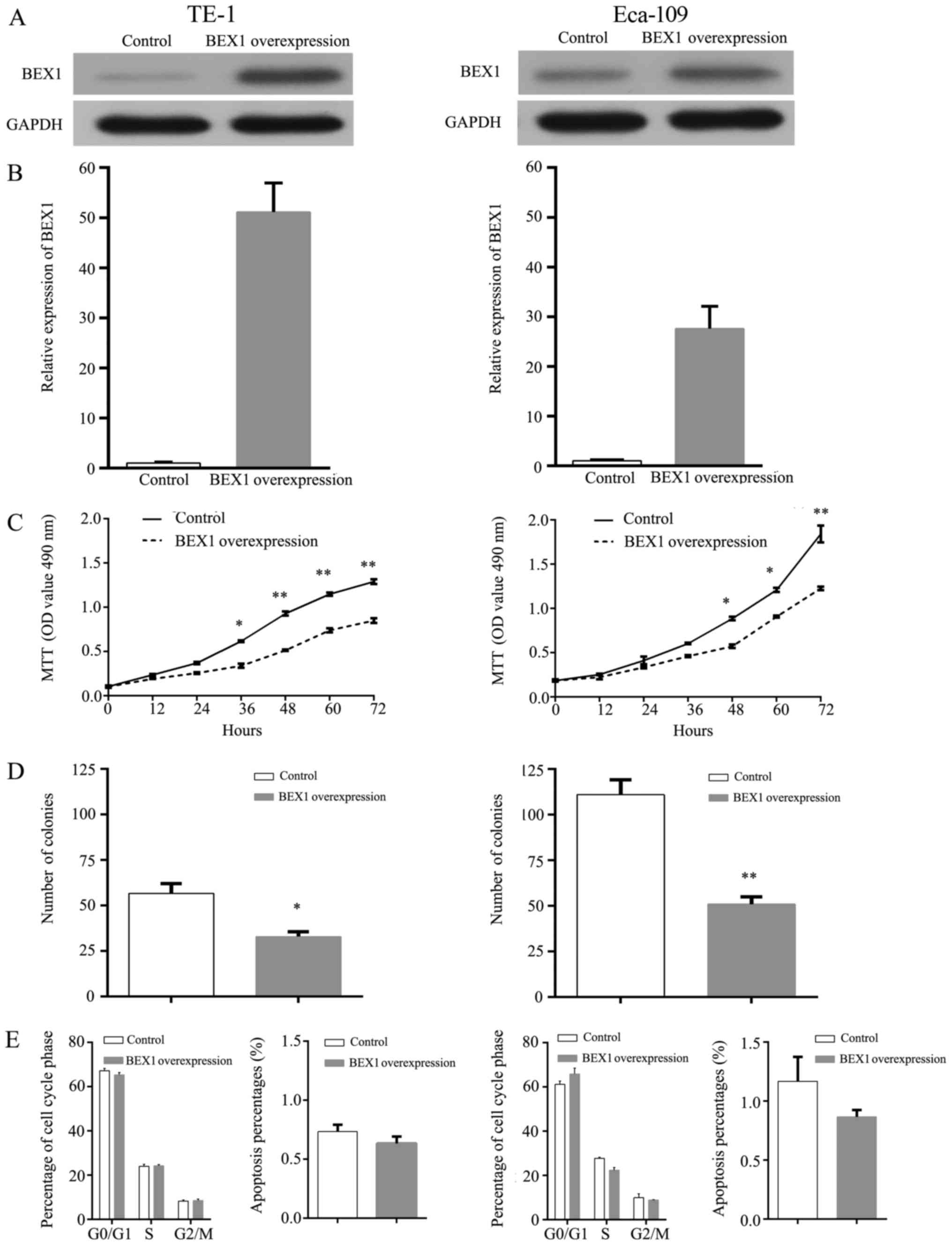
To explore the biological roles of BEX1 in ESCC
cancer cells, we transfected TE-1 and Eca-109 cells with BEX1
expression vector to increase BEX1 expression. Western blot
analysis and RT-PCR analyses revealed increased BEX1 expression in
the transfected TE-1 and Eca-109 cells (Fig. 3A and B). MTT assays revealed that
the proliferation of Eca-109 and TE-1 cells was significantly
inhibited following BEX1 overexpression in a time-dependent manner
(Fig. 3C). Consistent with the
decreased cellular proliferation rate, the colony formation assay
also revealed that BEX1 overexpression inhibited the colony
formation ability of TE-1 and Eca-109 cells (Fig. 3D). To examine whether BEX1 affected
the ability of cell proliferation by altering cell cycle
progression or apoptosis, we analyzed cell cycle distribution and
apoptosis via flow cytometry. We observed that ectopic BEX1
overexpression did not significantly change the cell cycle
distribution or apoptosis rates of ESCC cells (Fig. 3E). These results demonstrated that
overexpression of BEX1 inhibited the proliferation ability of ESCC
cells in vitro.
Overexpression of BEX1 inhibits ESCC
tumor growth in vivo
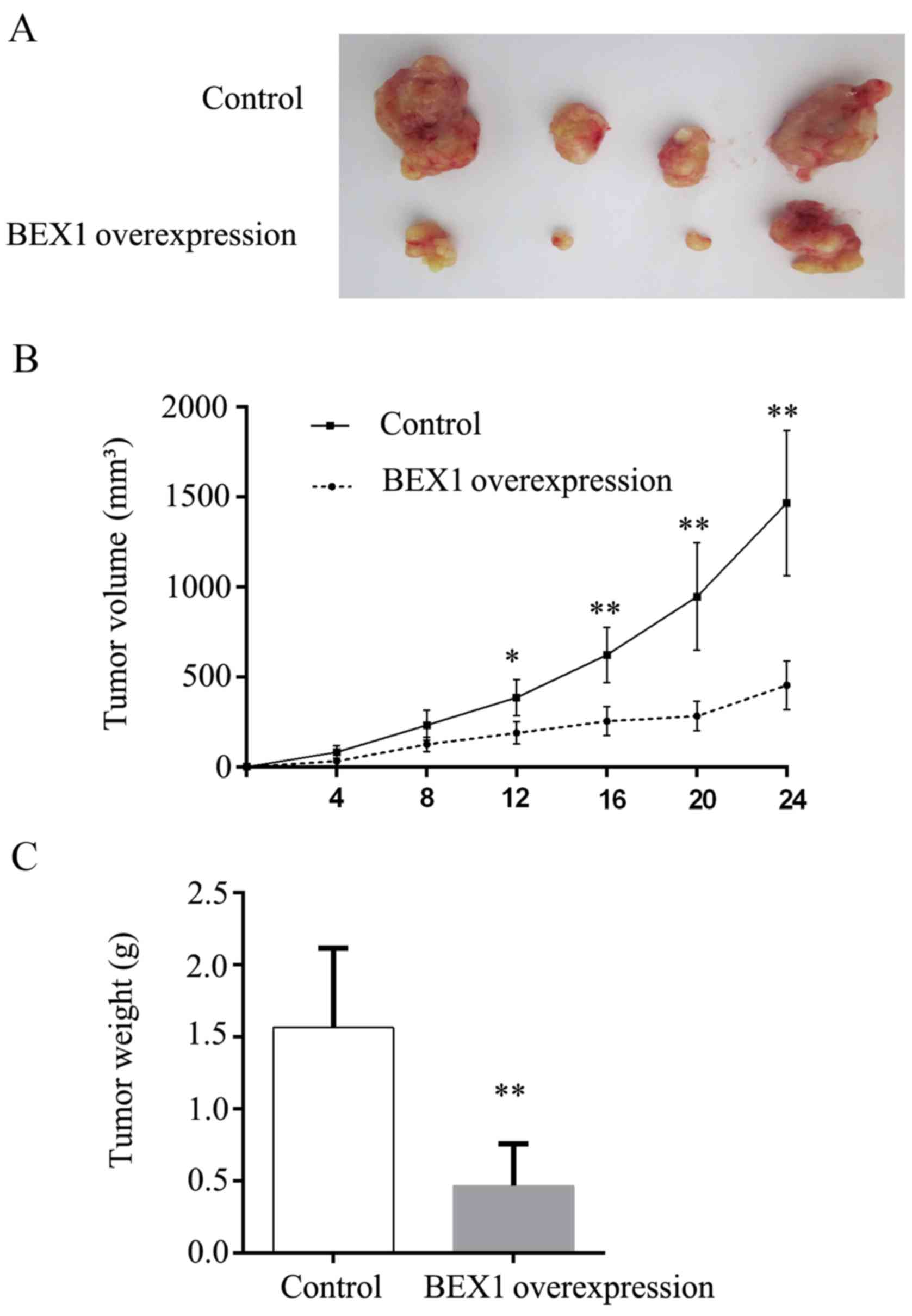
To extend our in vitro observations, we
assessed the effect of BEX1 overexpression on tumor growth in mouse
xenograft models. TE-1 cells stably expressing BEX1 or control
cells were subcutaneously injected into the flank of nude mice and
the tumor growth was monitored. The tumor volume was measured every
4 days, and the tumor weight was measured and analyzed 24 days
after injection. We observed that BEX1-overexpressing TE-1
×enografts grew significantly slower than the control xenografts
(Fig. 4A and B). Consistently, the
tumor weights from the BEX1-overexpressing group were significantly
lower than those from the control group (Fig. 4C). These data demonstrated that BEX1
inhibited ESCC tumor growth in vivo.
Overexpression of BEX1 inhibits the
activity of the NF-κB signaling pathway
To investigate the underlying mechanisms by which
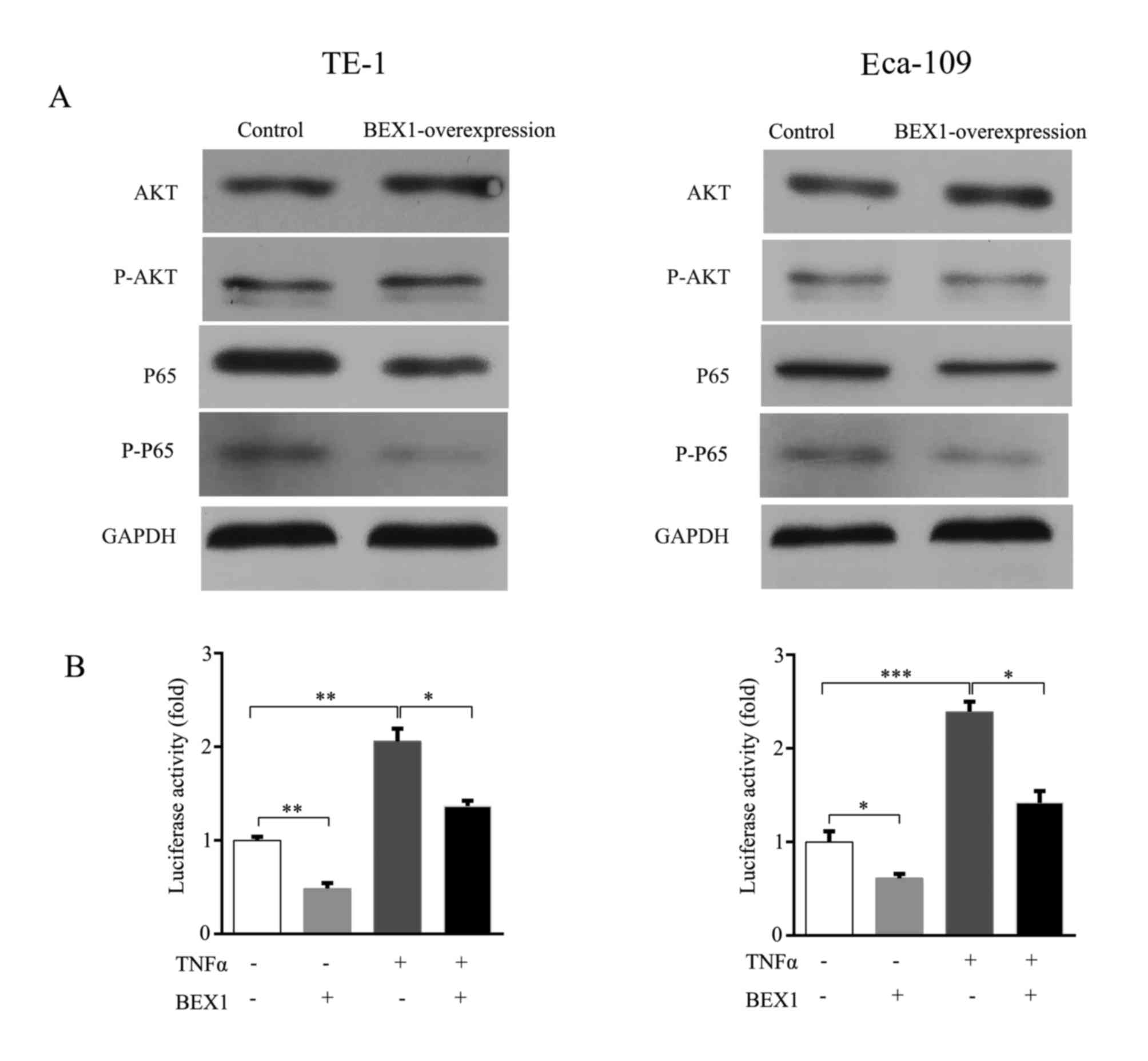
BEX1 inhibited the proliferation of ESCC cells, we firstly examined
the activity of the PI3K/AKT pathway, a canonical cell
growth-promoting signaling pathway, in TE-1 and Eca-109 cells with
or without stable BEX1 expression. We revealed that there was no
marked change in both total AKT and phosphorylated-AKT (pAKT)
levels between cells with and without BEX1 overexpression in TE-1
and Eca-109 cell lines (Fig. 5A).
The p65 subunit is an important member of canonical NF-κB and plays
a critical role in cell proliferation of ESCC cells. A previous
study demonstrated a close association of BEX1 with the NF-κB
signaling pathway in OSCC (11).
Therefore, we speculated that BEX1 may play tumor-suppressive roles
via the NF-κB signaling pathway in ESCC. To test our hypothesis, we
examined total p65 and phosphorylated-p65 (p-p65) expression levels
after overexpression of BEX1 in TE-1 and Eca-109 cells.
Immunoblotting revealed that overexpression of BEX1 decreased both
p65 and p-p65 levels (Fig. 5A). In
addition, NF-κB-luciferase reporter assays revealed that
BEX1-overexpresson significantly inhibited NF-κB activation and
prevented TNFα-stimulated NF-κB activation in both TE-1 and Eca-109
cells (Fig. 5B). These results
indicated that overexpression of BEX1 inhibited ESCC proliferation
via the downregulation of the NF-κB signaling pathway.
Discussion
Aberrant expression of BEX1 has been observed in
AML, CML, intracranial ependymoma, malignant glioma and OSCC
(8–12). In breast, gliomas and colorectal
cancers, BEX2 was found to be upregulated and BEX2 overexpression
enhanced cell proliferation (13–18),
indicating BEX2 to be a putative oncogene. However, BEX4
downregulation was also reported to promote the growth of OSCC,
suggesting a tumor suppressor role of BEX4 in OSCC (12). Therefore, the exact functions of
BEX1 in tumorigenesis remain elusive and warrant further extensive
investigation in different types of cancers with the advent of
precision cancer medicine.
Recent studies have revealed BEX1 to be
downregulated in OSCC, which has similar etiology and biological
phenotypes as ESCC (11,12). However, at present, little is known
about the expression level and function of BEX1 in ESCC. In the
present study, we investigated the expression of BEX1 in ESCC
tumors and cell lines and found that BEX1 expression was
downregulated ESCC tissues and cell lines compared to normal
tissues. Moreover, low expression of BEX1 was associated with
longer tumor length, as well as more advanced T stages and clinical
stages in ESCC patients. Additionally, low BEX1 expression
predicted poor prognosis of ESCC patients. Tumor size and T stage
are significantly correlated with cancer cell proliferation. In the
present study, we determined that BEX1 overexpression markedly
inhibited ESCC cell proliferation in vitro and tumor growth
in vivo. In agreement with our findings, BEX1 expression was
recently found to be downregulated in 27 out of 40 pediatric
intracranial ependymoma tissues, and overexpression of BEX1 to
significantly suppress cell proliferation and colony formation
(10). Similarly, Lee et al
found that the mRNA expression level of BEX1 was significantly
lower in OSCC patients with alcohol consumption, betel quid chewing
and cigarette smoking, and BEX1 demonstrated potent growth
inhibitory activities in OSCC cell culture and mouse xenografts
(11). Collectively, our findings
revealed that BEX1 may function as a tumor suppressor by inhibiting
the proliferation and survival of ESCC cells.
Emerging evidence suggests that BEX1 may be involved
in the regulation of cell apoptosis and cell cycle progression. In
AML, BEX1 induced cell apoptosis by selectively inhibiting
FLT3-ITD-induced AKT phosphorylation, without directly interacting
with FLT3-ITD (8). In malignant
glioma, BEX1 and BEX2 down-regulation resulting from promoter
hypermethylation led to increased sensitivity of glioma cells to
camptothecin-induced apoptosis (19). Furthermore, BEX1 was found to
re-sensitize K562 cells to imatinib-induced apoptosis by binding to
B-cell lymphoma 2 (BCL-2) (9,20).
Inconsistent with the previous studies, we observed no evident
alteration of cell cycle distribution and apoptosis following BEX1
overexpression, indicating that BEX1 regulates cancer cell
proliferation and proliferation via multiple mechanisms.
It has been well established that NF-κB plays an
important role in the regulation of inflammation, apoptosis,
angiogenesis as well as tumorigenesis (21). Activation of NF-κB, characterized by
phosphorylation of the p65 subunit, is closely associated with the
initiation and progression of many types of cancers including ESCC
(22–24). NF-κB p65 is overexpressed in human
ESCC tissues and NF-κB activation was revealed to be associated
with aggressive clinical biology and poor treatment outcome of ESCC
patients (25–27). In addition, blockade of the NF-κB
pathway by suppression of p65 not only inhibited cell
proliferation, but also suppressed the invasion and metastasis of
ESCC cells (28–31). Previous studies have disclosed a
close functional linkage between BEX family members and the NF-κB
signaling pathway (11,31). The potential role of BEX1 in the
inhibition of ESCC proliferation via regulation of the NF-κB
pathway remained unknown. Our results reevaled that p65 expression
was suppressed in BEX1-overexpressed ESCC cells. Notably, our
results demonstrated that both the constitutive and TNFα-stimulated
activation of NF-κB signaling were markedly inhibited by
BEX1-overexpression. Our findings indicated that the NF-κB pathway
may participate in BEX1-regulated ESCC initiation and progression
(data not shown). However, further investigations are required to
determine how BEX1 inhibits NF-κB activation in ESSC cells.
In conclusion, our study revealed that BEX1
expression was downregulated in ESCC and that low expression of
BEX1 was significantly associated with longer tumor length, more
advanced T stage and more advanced pTNM stage. Moreover, low
expression of BEX1 was associated with poor clinical outcome of
ESCC patients and was an independent prognostic factor for overall
survival. In addition, overexpression of BEX1 inhibited ESCC cell
proliferation and colony formation in vitro accompanied with
suppression of the NF-κB signaling pathway, and tumor growth in
vivo. Our findings collectively indicated that BEX1 may
function as a tumor suppressor by suppressing proliferation and
growth of ESCC tumors.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Beijing Medical
and Health Foundation (no. B17126), the Projects of Medical and
Health Technology Development Program of Shandong Province (no.
2015WSB30011) and the Science and Technology Planning Project of
Binzhou Medical University (no. BY2011KJ030).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author on reasonable request.
Authors' contributions
HTG, ZWC and YFC designed the study. HTG, ZWC, RJC,
ZBW, SZX and CG collected and analyzed the data. RJC, FW and CML
advised on histological staining and analysis. SZX, CG, FW and CML
contributed to the sample collection and intellectual input. HTG,
ZWC and RJC drafted and wrote the manuscript. SSC and YFC revised
the manuscript critically for intellectual content. All authors
gave intellectual input to the study and approved the final version
of the manuscript. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Binzhou Medical University Hospital. All procedures performed in
studies involving human participants were in accordance with the
ethical standards of the institutional and national research
committee and with the 1964 Helsinki declaration and its later
amendments or comparable ethical standards. Written informed
consent was obtained from all individual participants included in
the present study. The Animal Committee of Binzhou Medical
University approved all the experimental protocols and animal
handling procedures. All experimental procedures and postoperative
animal care were conducted in accordance with the guidelines for
the Care and Use of Laboratory Animals from the National Institutes
of Health (Bethesda, MD, USA).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wu M, Liu AM, Kampman E, Zhang ZF, Van't
Veer P, Wu DL, Wang PH, Yang J, Qin Y, Mu LN, et al: Green tea
drinking, high tea temperature and esophageal cancer in high- and
low-risk areas of Jiangsu Province, China: A population-based
case-control study. Int J Cancer. 124:1907–1913. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Paul S and Altorki N: Outcomes in the
management of esophageal cancer. J Surg Oncol. 110:599–610. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Alvarez E, Zhou W, Witta SE and Freed CR:
Characterization of the Bex gene family in humans, mice, and rats.
Gene. 357:18–28. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang L: Adaptive evolution and frequent
gene conversion in the brain expressed X-linked gene family in
mammals. Biochem Genet. 46:293–311. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Quentmeier H, Tonelli R, Geffers R,
Pession A, Uphoff CC and Drexler HG: Expression of BEX1 in
acute myeloid leukemia with MLL rearrangements. Leukemia.
19:1488–1489. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lindblad O, Li T, Su X, Sun J, Kabir NN,
Levander F, Zhao H, Lu G, Rönnstrand L and Kazi JU: BEX1 acts as a
tumor suppressor in acute myeloid leukemia. Oncotarget.
6:21395–21405. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ding K, Su Y, Pang L, Lu Q, Wang Z, Zhang
S, Zheng S, Mao J and Zhu Y: Inhibition of apoptosis by
downregulation of hBex1, a novel mechanism, contributes to the
chemoresistance of Bcr/Abl+ leukemic cells.
Carcinogenesis. 30:35–42. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Karakoula K, Jacques TS, Phipps KP,
Harkness W, Thompson D, Harding BN, Darling JL and Warr TJ:
Epigenetic genome-wide analysis identifies BEX1 as a
candidate tumour suppressor gene in paediatric intracranial
ependymoma. Cancer Lett. 346:34–44. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee CH, Wong TS, Chan JY, Lu SC, Lin P,
Cheng AJ, Chen YJ, Chang JS, Hsiao SH, Leu YW, et al: Epigenetic
regulation of the X-linked tumour suppressors BEX1 and
LDOC1 in oral squamous cell carcinoma. J Pathol.
230:298–309. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gao W, Li JZ, Chen SQ, Chu CY, Chan JY and
Wong TS: Decreased brain-expressed X-linked 4 (BEX4) expression
promotes growth of oral squamous cell carcinoma. J Exp Clin Cancer
Res. 35:922016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Naderi A, Liu J and Francis GD: A feedback
loop between BEX2 and ErbB2 mediated by c-Jun signaling in breast
cancer. Int J Cancer. 130:71–82. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Naderi A, Teschendorff AE, Beigel J,
Cariati M, Ellis IO, Brenton JD and Caldas C: BEX2 is overexpressed
in a subset of primary breast cancers and mediates nerve growth
factor/nuclear factor-kappaB inhibition of apoptosis in breast
cancer cell lines. Cancer Res. 67:6725–6736. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fischer C, Drexler HG, Reinhardt J,
Zaborski M and Quentmeier H: Epigenetic regulation of brain
expressed X-linked-2, a marker for acute myeloid leukemia with
mixed lineage leukemia rearrangements. Leukemia. 21:374–377. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou X, Xu X, Meng Q, Hu J, Zhi T, Shi Q
and Yu R: Bex2 is critical for migration and invasion in malignant
glioma cells. J Mol Neurosci. 50:78–87. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou X, Meng Q, Xu X, Zhi T, Shi Q, Wang Y
and Yu R: Bex2 regulates cell proliferation and apoptosis in
malignant glioma cells via the c-Jun NH2-terminal kinase pathway.
Biochem Biophys Res Commun. 427:574–580. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu Y, Xiao Q, Chen H, He J, Tan Y, Liu Y,
Wang Z, Yang Q, Shen X, Huang Y, et al: BEX2 promotes tumor
proliferation in colorectal cancer. Int J Biol Sci. 13:286–294.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Foltz G, Ryu GY, Yoon JG, Nelson T, Fahey
J, Frakes A, Lee H, Field L, Zander K, Sibenaller Z, et al:
Genome-wide analysis of epigenetic silencing identifies BEX1
and BEX2 as candidate tumor suppressor genes in malignant
glioma. Cancer Res. 66:6665–6674. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Xiao Q, Hu Y, Liu Y, Wang Z, Geng H, Hu L,
Xu D, Wang K, Zheng L, Zheng S, et al: BEX1 promotes
imatinib-induced apoptosis by binding to and antagonizing BCL-2.
PLoS One. 9:e917822014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sethi G, Ahn KS, Sung B and Aggarwal BB:
Pinitol targets nuclear factor-kappaB activation pathway leading to
inhibition of gene products associated with proliferation,
apoptosis, invasion, and angiogenesis. Mol Cancer Ther.
7:1604–1614. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pikarsky E, Porat RM, Stein I, Abramovitch
R, Amit S, Kasem S, Gutkovich-Pyest E, Urieli-Shoval S, Galun E and
Ben-Neriah Y: NF-kappaB functions as a tumour promoter in
inflammation-associated cancer. Nature. 431:461–466. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Karin M and Greten FR: NF-kappaB: Linking
inflammation and immunity to cancer development and progression.
Nat Rev Immunol. 5:749–759. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Karin M: Nuclear factor-kappaB in cancer
development and progression. Nature. 441:431–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang F, He W, Fanghui P, Wang L and Fan Q:
NF-κBP65 promotes invasion and metastasis of oesophageal squamous
cell cancer by regulating matrix metalloproteinase-9 and
epithelial-to-mesenchymal transition. Cell Biol Int. 37:780–788.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kang MR, Kim MS, Kim SS, Ahn CH, Yoo NJ
and Lee SH: NF-κB signalling proteins p50/p105, p52/p100, RelA, and
IKKepsilon are over-expressed in oesophageal squamous cell
carcinomas. Pathology. 41:622–625. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Izzo JG, Malhotra U, Wu TT, Ensor J,
Luthra R, Lee JH, Swisher SG, Liao Z, Chao KS, Hittelman WN, et al:
Association of activated transcription factor nuclear factor kappab
with chemoradiation resistance and poor outcome in esophageal
carcinoma. J Clin Oncol. 24:748–754. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tian F, Zhang C, Tian W, Jiang Y and Zhang
X: Comparison of the effect of p65 siRNA and curcumin in promoting
apoptosis in esophageal squamous cell carcinoma cells and in nude
mice. Oncol Rep. 28:232–240. 2012.PubMed/NCBI
|
|
29
|
Tian F, Fan T, Jiang Y, Zhang X and Wang
X: A small interfering RNA targeting NF-κB p65 alone or combined
with 5-FU inhibits growth of esophageal squamous cell carcinoma in
nude mice. Pathol Res Pract. 208:32–38. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Tian F, Zang WD, Hou WH, Liu HT and Xue
LX: Nuclear factor-κB signaling pathway constitutively activated in
esophageal squamous cell carcinoma cell lines and inhibition of
growth of cells by small interfering RNA. Acta Biochim Biophys Sin.
38:318–326. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Meng Q, Zhi T, Chao Y, Nie E, Xu X, Shi Q,
Hua L, Wang L, Zhan W, Wang Y, et al: Bex2 controls proliferation
of human glioblastoma cells through NF-κB signaling pathway. J Mol
Neurosci. 53:262–270. 2014. View Article : Google Scholar : PubMed/NCBI
|