Introduction
Head and neck squamous cell carcinoma (HNSCC) is one
of the most common types of cancer worldwide (1). Approximately 550,000 new patients are
diagnosed, and 30,000 patients succumb to this disease each year
(2). HNSCC has varying levels of
differentiation, and a propensity for lymph node metastasis and
poor prognosis (3,4). Over the past few years, there has been
little improvement in the five-year survival rate using the current
therapies available. Therefore, in light of the high recurrence
rates and limited treatment options, it is necessary to investigate
the invasive molecular mechanism of HNSCC.
Tumor-associated macrophages (TAMs) are a key
component of the tumor microenvironment and are crucial for cancer
development (5). The majority of
macrophages originate from monocytes in the circulation and form an
extremely heterogeneous population. Activated macrophages are
classified into M1 and M2 macrophages. M1 is classically activated,
while M2 is activated via an alternative pathway (6,7). TAMs,
similar to the M2 phenotype, promote cancer progression by
accelerating new blood cell formation and mediating
immunoregulation (8,9). High levels of TAMs are often
associated with a poor prognosis (10). Therefore, studying the mechanism of
the interaction between TAMs and tumors may provide important
targets for cancer therapy.
Epithelial to mesenchymal transition (EMT) increases
the motility of cancer cells by altering epithelial cell morphology
and apicobasal polarity, changing cellular contact patterns and
reducing cell adhesion (11,12).
It has been reported that macrophages may directly induce the EMT
of breast cancer cells in a juxtacrine manner, thereby establishing
an environment suitable for the growth of tumor stem cells
(13). TAMs are often located in
the invasive front of cancer cells, where the majority of cancer
cells are undergoing EMT (14). Our
previous study indicated that HNSCC cells could induce monocytes to
differentiate into M2 macrophages via a C-C motif chemokine ligand
2 and C-C chemokine receptor type 2 pathway (15). In addition, it was identified that
macrophages were likely to induce the EMT of HNSCC cells, as the
majority of macrophages were located at the edge of the scratch in
the wound healing assay. Pirilä et al (16) reported that M2 macrophages
co-cultured with HSC-3 cells increased the expression of epidermal
growth factor (EGF), transforming growth factor-β (TGF-β) and
macrophage colony-stimulating factor (M-CSF). Activation of the EGF
and/or TGF-β signaling pathways and their downstream cascade may
trigger the EMT process in various types of cancer cells (17,18).
However, the mechanism by which TAMs in HNSCC induce the EMT of
tumor cells remains unknown.
In the present study, the expression of TAMs and
EMT-associated proteins in the HNSCC tissues were detected, and the
correlations between them were evaluated. Direct and indirect
co-culture systems of TAMs and HNSCC cells were established, and
the involved extracellular and intracellular signaling pathways
were examined. To the best of our knowledge, this is the first
study to suggest that TAMs induce the EMT of HNSCC cells primarily
by activating the EGF receptor (EGFR)/extracellular
signal-regulated kinase1/2 (ERK1/2) signaling pathway. This may
provide a potential therapeutic strategy for suppressing tumor
invasion and migration in HNSCC.
Materials and methods
Patient samples
A total of 56 paraffin-embedded human HNSCC
specimens and 10 normal adjacent mucous samples that were
histopathologically diagnosed at Second Hospital of Dalian Medical
University (Dalian, China) from January 2010 to December 2014 were
included in the present study. The detailed pathological and
clinical data for all of the samples are presented in Table I. The use of human tissues was
approved by the Medical Ethics Committee of Dalian Medical
University and written informed consent was provided by each
patient. Specimens that were obtained from patients treated with
radiotherapy and chemotherapy were excluded from the present study.
The procedure followed the USA National Institutes of Health
guidelines (19) regarding use of
human tissues.
 | Table I.Clinical characteristics of patients
and the 56 HNSCC and 10 normal tissues. |
Table I.
Clinical characteristics of patients
and the 56 HNSCC and 10 normal tissues.
|
|
| Macrophages
infiltration |
|---|
|
|
|
|
|---|
| Clinical
characteristic | Total cases
(n) | Negative | Low | High |
|---|
| Normal and adjacent
tissue | 10 | 7 | 3 | – |
| HNSCC | 56 | – | 25 | 31 |
| Age, years |
|
≤45 | 12 | – | 4 | 8 |
|
>45 | 44 | – | 21 | 23 |
| Sex |
|
Male | 36 | – | 12 | 24 |
|
Female | 20 | – | 13 | 7 |
| TNM grading |
| Stage
I | 21 | – | 14 | 7 |
| Stage
II | 24 | – | 8 | 16 |
| Stage
III | 8 | – | 3 | 5 |
| Stage
IV | 3 | – | 0 | 3 |
| Histological
differentiation |
|
Well | 33 | – | 18 | 15 |
|
Moderately | 18 | – | 6 | 12 |
|
Poorly | 5 | – | 1 | 4 |
Cell culture
THP1 [human acute monocytic leukemia cell line;
China Center for Type Culture Collection (CCTCC), Wuhan, China]
cells were maintained in RPMI-1640 (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and Cal27 (oral tongue squamous
carcinoma cell line; CCTCC) cells were maintained in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc.). SCC25 [oral tongue squamous carcinoma cell line; American
Type Culture Collection (ATCC), Manassas, VA, USA] cells were
cultured in a 1:1 mixture of DMEM and Ham's F12 medium (Thermo
Fisher Scientific, Inc.) and Fadu (hypopharyngeal squamous
carcinoma cell line; ATCC) cells were cultured in DMEM. All the
cells were cultured at 37°C in a 5% CO2 humidified
atmosphere with medium containing 10% fetal bovine serum (FBS), 100
IU/ml penicillin and 100 µg/ml streptomycin (Thermo Fisher
Scientific, Inc.).
Induction of macrophage
polarization
According to our previous study, M0, M1 and M2
macrophages were induced from THP1 cells (15,20).
During this induction process, cells were cultured at 37°C in a 5%
CO2 humidified atmosphere. First,
phorbol-12-myristate-13-acetate (PMA; 320 nM; Cell Signaling
Technology, Inc., Danvers, MA, USA) was added to
1×106/ml THP1 cells. Following 24 h, THP1 cells were
induced into the M0 phenotype. For M1 and M2 macrophages, THP1
cells were treated with PMA for 6 h, and then induced into M1
macrophages by interferon-γ (IFN-γ; 20 ng/ml) and
lipopolysaccharide (LPS; 100 ng/ml) for 18 h, or induced into M2
macrophages by interleukin-4 (IL-4; 20 ng/ml) and IL-13 (20 ng/ml)
for 18 h. Following a further 24 h culture of each macrophage
phenotype in FBS-free medium, the macrophage conditional medium was
collected for use in the next step.
Immunohistochemistry and
evaluation
Immunohistochemical staining was performed according
to the method described previously (21). Briefly, tissues were fixed with 4%
paraformaldehyde for 48 h at room temperature. Then 4-µm thick
tissue sections were cut, dried, deparaffinized with xylene and
rehydrated using a descending series of alcohol following standard
procedures. Then, the sections were subjected to heat-induced
antigen retrieval at 98°C for 10 min with citrate buffer (Abcam,
Cambridge, MA, USA) and washed thrice with PBS for 3 min each. The
sections were then blocked with 10% non-immune goat serum (Gibco;
Thermo Fisher Scientific, Inc.) for 1 h at room temperature.
Immunohistochemical staining was performed overnight a 4°C with the
following primary antibodies: Mouse monoclonal anti-cluster of
differentiation (CD)-68 antibodies (Santa Cruz Biotechnology, Inc.,
Dallas, TX, USA; dilution, 1:50; cat. no. sc-17832), mouse
monoclonal anti-CD163 antibodies (Santa Cruz Biotechnology, Inc.;
dilution, 1:50; cat. no. sc-33715) and antibodies against EMT
markers, including mouse anti-Vimentin (Abcam; dilution, 1:200;
cat. no. ab8978) and rabbit anti-E-cadherin (Abcam; dilution,
1:100; cat. no. ab40772). Tissues were then incubated with
horseradish peroxidase (HRP)-conjugated secondary antibodies
(Abcam; dilution, 1:500; cat. nos. ab6112 and ab97046) for 1 h at
room temperature. The macrophage markers of CD68 and CD163 were
semi-quantitatively evaluated in HNSCC on the basis of staining
intensity and distribution using the immunoreactive score (IRS)
(22): IRS = staining intensity ×
percentage of positive cells. The staining intensity was defined as
follows: 0, negative; 1, weak; 2, moderate; and 3, strong. The
percentage of positive cells was defined as follows: 0, negative;
1, 1–20% positive cells; 2, 21–50% positive cells; and 3, 51–100%
positive cells. The IRS ranged from 0 to 9, which was divided into
3 groups on the basis of the final score: i) IRS 0, negative; ii)
IRS 1–4, low immunoreactivity; and iii) IRS >4, high
immunoreactivity. The percentage of positive cells was quantified
by 3 pathologists who counted in ten different visual fields at
random using a light microscope (Leica Microsystems, Inc., Buffalo
Grove, IL, USA; magnification, ×200) and the average IRS was
calculated as the final value. HNSCC cells with high
immunoreactivity for CD68 and CD163 were classified as high
macrophage HNSCC (HM-SCC); cells with low immunoreactivity for one
or both of the macrophage markers (CD68 and CD163) were classified
as low macrophage HNSCC (LM-SCC). All of the HNSCC samples were
divided into HM-SCC and LM-SCC, as shown in Table I.
Immunofluorescence
Tissue section processing was similar to that
performed for immunohistochemistry. The treated tissue was
incubated overnight with primary antibodies against EMT markers
(E-cadherin and Vimentin) and M2 macrophage markers (CD68 and
CD163) at 4°C (all antibodies as aforementioned). Then, the tissue
was incubated with secondary anti-mouse or anti-rabbit antibodies
conjugated with Alexa Fluor 488 (Invitrogen; Thermo Fisher
Scientific, Inc.; dilution, 1:400; cat. no. A-11034) or Alexa Fluor
568 (Invitrogen; Thermo Fisher Scientific, Inc.; dilution, 1:400;
cat. no. A-21144) for 1 h at room temperature. Finally, the nuclei
were stained with mounting medium containing DAPI for 5 min at room
temperature. The tissues were observed and photographed with a
fluorescence microscope (Leica Microsystems, Inc.). For the
evaluation, as described previously (23), the images were analyzed using the
Image-Pro Plus 7.0 version software (Media Cybernetics, Inc.,
Rockville, MD, USA). Briefly, five representative high-power fields
were selected randomly. The reference value that identified the
positive staining was set. Then, the sum integrated optical density
(IOD) and the sum area (area) would be generated automatically. The
semi-quantitative value of each marker was expressed as a mean
density (IOD/area).
Flow cytometry
To analyze the macrophage marker expression in the
induced THP1 cells, flow cytometry was performed. Different
phenotype macrophages were harvested, washed in ice-cold PBS and
blocked with PBS containing 3% bovine serum albumin (Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany) on ice for 30 min. The cells were
centrifuged at 300 × g for 5 min at 4°C and resuspended with cold
PBS to a final concentration of 2×107 cells/ml. Then
1×106 cells/tube in 100 µl were prepared and incubated
with fluorescein isothiocyanate conjugated CD68 antibody (BD
Biosciences, Franklin Lakes, NJ, USA; cat. no. 562117; dilution,
1:20) and phycoerythrin-conjugated antibody CD163 (BD Biosciences;
cat. no. 563887; dilution, 1:20) for 20 min on ice avoiding light.
Cells were washed 3 times in PBS, and detected on
Fluorescence-Activated Cell Sorting (FACS) Calibur flow cytometer
(BD Biosciences). The data was analyzed using BD Cell Quest 5.1 (BD
Biosciences) and WinMDI 2.9 software (The Scripps Institute, Flow
Cytometry Core Facility, La Jolla, CA, USA; www.cyto.purdue.edu/flowcyt/software/Winmdi.htm).
Cell invasion and migration assay
A 24-well Transwell chamber (pore size, 8 mm) with
or without Matrigel coating (BD Biosciences) was used to detect
tumor cell invasion and migration, respectively. Tumor cells
(1×105) co-cultured with or without TAMs were plated in
the upper chamber, and different conditional media (CM) containing
10% FBS (Thermo Fisher Scientific, Inc.) with or without U0126
[1,4-diamino-2, 3-dicyano-1,4-bis (2-aminophenylmercapto)
butadiene; ERK1/2 inhibitor; 10 µg/ml; Cell Signaling Technology,
Inc.] were added to the lower chamber. Once the cells were
incubated for 24 h, the non-penetrated cells were scraped from the
upper surface of the filter, and the penetrated cells, representing
invaded or migrated cells, were stained with crystal violet
solution (Sigma-Aldrich; Merck KGaA) at room temperature for 10
min. Cells were counted under a phase microscope, photographed and
analyzed using ImageJ 1.42 software (National Institutes of Health,
Bethesda, MD, USA).
Wound healing assay
Cal27 cells (1×106) were seeded in 6-well
plates with or without TAM-CM and M2 macrophages. When tumor cells
reached 70–80% confluence, a straight scratch was made using a 200
µl pipette tip, and an artificial wound was formed. Following 18 h
at 37°C in a 5% CO2 humidified atmosphere, the migration
of tumor cells across this artificial wound was photographed using
a light microscope (Leica Microsystems, Inc.) and assessed using
ImageJ 1.42 software (National Institutes of Health).
Enzyme-linked immunosorbent assay
(ELISA)
Cytokine secretion of EGF and TGF-β in the THP1 M0,
M1 and M2 macrophage culture supernatant was measured using EGF
ELISA kits (R&D Systems, Inc., Minneapolis, MN, USA; cat. no.
#DY236) and TGF-β ELISA kits (R&D Systems, Inc.; cat. no.
#DY240) according to the manufacturer's protocol. Briefly, 100 µl
of prepared standard and samples were added to the 96-wells, then
the plate was covered and incubated at room temperature for 2 h.
The samples were removed and the plate was washed 3 times with 200
µl PBS. Then, 100 µl diluted antibody was added into each well and
tapped gently on the side of the plate to mix. Following incubation
for 1 h at room temperature, the liquid was discarded and the wells
were washed 3 times. Then, 100 µl of diluted HRP conjugate was
added to each well and incubated at room temperature for 30 min.
The liquid was removed and washed 3 times. Then, 100 µl chromogenic
substrate was added to each well at room temperature in the dark
for 30 min. Following the addition of 100 µl of stop solution to
each well, the absorbance of each sample was evaluated using a
microplate reader at 450 and 550 nm. By comparing the absorbance of
the standard points against the standard concentrations, a standard
curve was constructed, and the results for the test samples were
calculated.
Western blot analysis
Cal27 cells labeled with carboxyfluorescein
diacetate succinimidyl ester (CFDA-SE; Thermo Fisher Scientific,
Inc.; cat. no. V12883) were added to the co-culture system.
Following treatment with the EGFR inhibitors Cetuximab (10 µg/ml;
Merck KGaA) or U0126 (10 µg/ml; Cell Signaling Technology, Inc.)
for the indicated times 24 h, Cal27 cells were sorted from the
co-culture by FACS. Tumor cells (2×106) were then lysed
with radioimmunoprecipitation assay lysis and extraction buffer
(Thermo Fisher Scientific, Inc.; cat. no. #89900) with Halt
Protease and Phosphatase Inhibitor (Thermo Fisher Scientific, Inc.;
cat. no. #78447) at 4°C, and the concentration of protein in each
cell lysate was measured using the bicinchoninic acid method. Then,
20 µg of protein was separated by 10% SDS-PAGE and electroblotted
onto polyvinylidene fluoride membranes. The blots were blocked with
5% nonfat dry milk at room temperature for 1 h, and probed
overnight at 4°C with primary antibodies, including rabbit
anti-total ERK1/2 (1:1,000; cat. no. 4695), rabbit anti-total P65
(1:500; cat. no. 8242), rabbit anti-protein kinase B (Akt; 1:1,000;
cat. no. 9272), anti-phospho-Akt (p-Akt; 1:1,000; cat. no. 9611),
rabbit anti-Mothers against decapentaplegic homolog 3 (Smad3;
1:1,000; cat. no. 9513), anti-p-Smad3 antibodies (1:500; cat. no.
9520; all from Cell Signaling Technologies, Inc.), mouse
anti-α-smooth muscle actin (α-SMA; 1:1,000; cat. no. ab7817), mouse
anti-GADPH antibody (1:1000; cat. no. ab8245; both Abcam), rabbit
anti-p-ERK1/2 (1:500; cat. no. MAB18251) and anti-p-P65 antibodies
(1:500; cat. no. MAB72261; both R&D Systems, Inc.). Then, the
immunoblots were incubated with HRP-conjugated secondary antibodies
(1:5,000; cat. nos. 32230 and 32260; Pierce; Thermo Fisher
Scientific, Inc.) at room temperature for 2 h. The protein bands
were visualized using an enhanced chemiluminescence detection kit
(Pierce; Thermo Fisher Scientific, Inc.). ImageJ 1.42 software
(National Institutes of Health) was used for densitometric
analysis.
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), and reverse
transcribed using PrimeScript RT (Takara Bio, Inc., Otsu, Japan)
according to the manufacturer's protocol. Briefly, the template
RNA/Primer Mixture was incubated at 65°C for 5 min, then
immediately cooled on ice. PrimeScript Buffer and Reverse
Transcriptase were then added to the reaction mixture, creating a
20-µl reaction, and incubated at 42°C for 50 min and 70°C for 15
min, followed by cooling on ice. qPCR was performed using the 7500
Fast System (Applied Biosystems; Thermo Fisher Scientific, Inc.)
and the Fast SYBR-Green Master Mix (Thermo Fisher Scientific,
Inc.). RT-qPCR was performed with the following thermocycling
conditions: 40 cycles of 95°C for 5 sec, 60°C for 20 sec and 72°C
for 20 sec. The internal control of the experiment was GADPH, and
the primer sequences used for qPCR are presented in Table II. The relative expression levels
were analyzed using the 2−ΔΔCq method (24).
| Table II.Primer sequences used to determine
gene expression by reverse transcription-quantitative polymerase
chain reaction. |
Table II.
Primer sequences used to determine
gene expression by reverse transcription-quantitative polymerase
chain reaction.
| Gene | Type | Primers sequence
(5′-3′) |
|---|
| TNF-α | Sense |
TCTTCTCGAACCCCGAGTGA |
|
| Antisense |
CCTCTGATGGCACCACCAG |
| IL-1β | Sense |
TACGAATCTCCGACCACCACTACAG |
|
| Antisense |
TGGAGGTGGAGAGCTTTCAGTTCATATG |
| TGF-β | Sense |
GGGACTATCCACCTGCAAGA |
|
| Antisense |
CCTCCTTGGCGTAGTAGTCG |
| IL-10 | Sense |
AGAACAGCTGCACCCACTTC |
|
| Antisense |
GCATCACCTCCTCCAGGTAA |
| EGF | Sense |
GTGCAGCTTCAGGACCACAA |
|
| Antisense |
AAATGCATGTGTCGAATATCTTGAG |
| GAPDH | Sense |
AGTCCTTCCACGATACCAAAGT |
|
| Antisense |
CATGAGAAGTATGACAACAGCCT |
Statistical analysis
The results of immunohistochemistry and
immunofluorescence were assessed using Image Pro Plus 7.0 software
(Media Cybernetics, Inc.). Western blotting and wound healing assay
results were analyzed using ImageJ 1.42 software (National
Institutes of Health). The hierarchical analysis was conducted on
the basis of Pearson's correlation using Cluster version 3.0 (Eisen
Lab, University of California, Berkeley, CA, USA; bonsai.hgc.jp/~mdehoon/software/cluster/) and
visualized as a heatmap using the Java Tree View version 1.0.5
software (Eisen Lab, University of California; bitbucket.org/TreeView3Dev/treeview3/). All
experiments included at least 3 independent repeats. The data were
analyzed using GraphPad Prism 6 for Windows (GraphPad Software,
Inc., La Jolla, CA, USA). One-way analysis of variance followed by
the post hoc Tukey's multiple comparison test was performed for
analysis of 3 or more groups. Two-tailed Pearson's statistics was
performed for correlation analysis of EMT and macrophage markers.
P<0.05 was considered to indicate a statistically significant
difference.
Results
TAMs are associated with EMT markers
in HNSCC
The 57 HNSCC specimens were classified into two
groups of HM-SCC and LM-SCC on the basis of CD68 and CD163
immunohistochemical staining using the immunoreactive score
(Table I). To explore the
association between TAMs and EMT in HNSCC, the expression levels of
EMT-associated proteins were evaluated. Using immunofluorescence,
the results indicated that the epithelial marker, E-cadherin, was
significantly downregulated in HM-SCC compared with LM-SCC
(P<0.01) and normal mucosa (NM; P<0.005, Fig. 1A-D). Immunohistochemistry was also
performed to confirm the expression of EMT-associated proteins in
HNSCC and NM samples. The results indicated higher expression of
Vimentin and lower expression of E-cadherin in HM-SCC when compared
with LM-SCC and NM (Fig. 1E). These
results suggested that HNSCC cells with high infiltration of M2
macrophages may undergo EMT. Furthermore, Pearson correlation
analysis revealed correlations between EMT-associated proteins
(E-cadherin and Vimentin) and M2-macrophage markers (CD68 and
CD163). The results indicated that the mean density of E-cadherin
were negatively correlated with CD163+ cells (P=0.0004; Fig. 1F), while the expression of Vimentin
in HNSCC was positively correlated with CD163+ cells (P=0.0011;
Fig. 1F). These results indicated
that the EMT of HNSCC cells may be induced by TAMs, which may
contribute to the aggressive behaviors of HNSCC.
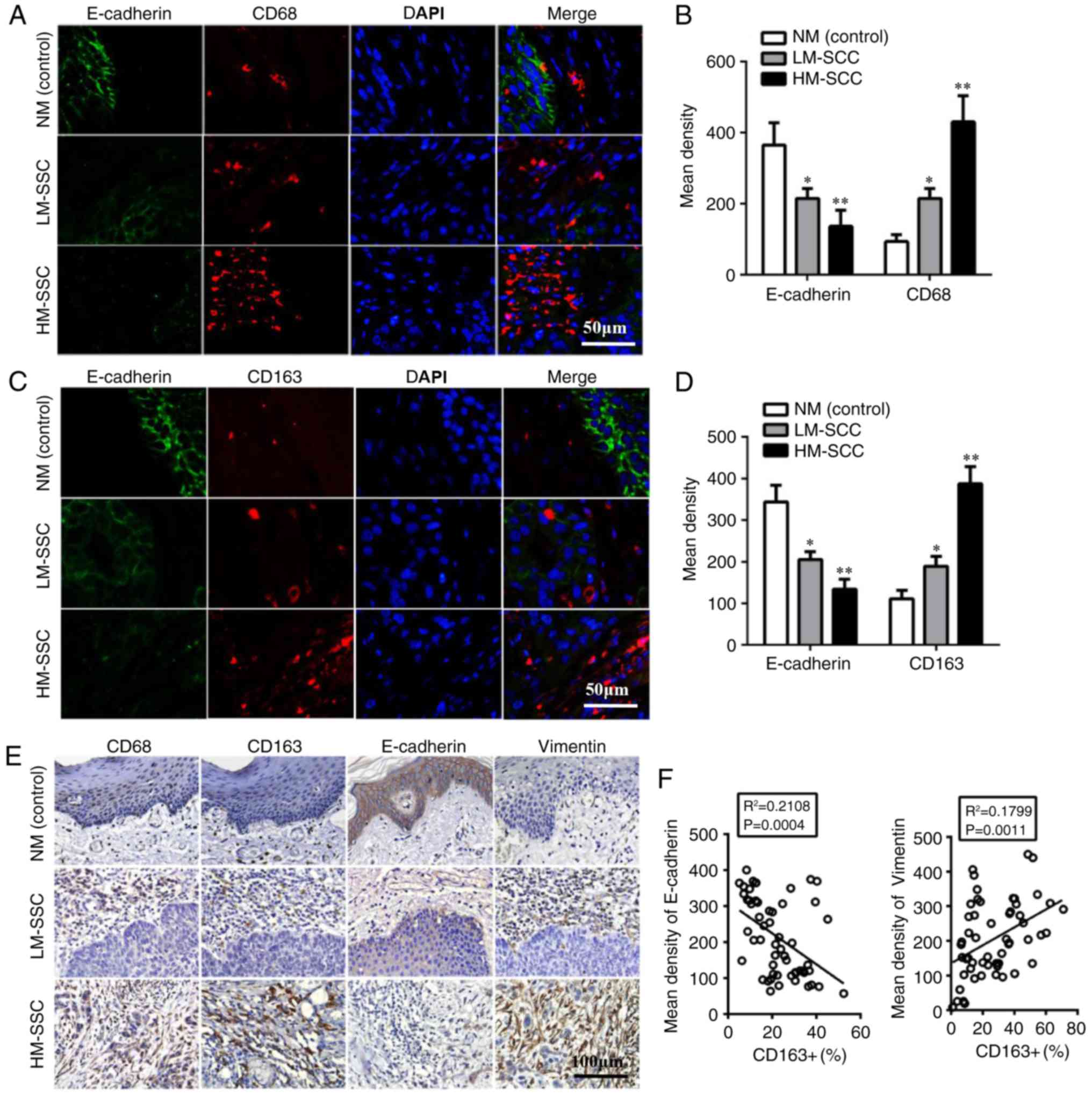 | Figure 1.Tumor associated macrophages
biomarkers are associated with EMT-associated proteins in HNSCC.
(A) Detection of CD68 and E-cadherin expression by double
immunofluorescent staining in HNSCC tissues. Magnification, ×200.
(B) Quantitative data of CD68 and E-cadherin expression from
immunofluorescent staining in HNSCC tissues. (C) Detection of CD163
and E-cadherin by double immunofluorescent staining in HNSCC
tissues. Magnification, ×200. (D) Quantitative data of CD163 and
E-cadherin from immunofluorescent staining in HNSCC tissues. Data
are expressed as the mean ± standard error of the mean. *P<0.05
and **P<0.01 vs. NM (control). (E) Detection of CD68, CD163 and
EMT-associated protein expression by immunohistochemical staining
in HNSCC tissues. Magnification, ×200. (F) Pearson correlation
analysis of EMT-associated protein and CD163+ expression. HNSCC,
head and neck squamous cell carcinoma; EMT, epithelial to
mesenchymal transition; CD, cluster of differentiation; HM-SCC,
HNSCC with high macrophages; LM-SCC, HNSCC with low macrophages;
NM, normal mucosa. |
TAMs induce the EMT of HNSCC
cells
To investigate whether TAMs induce EMT in HNSCC
cells, direct and indirect co-culture systems were established
between HNSCC cells and TAMs. Human THP-1 cells are widely used as
models for monocyte/macrophage differentiation; when treated with
PMA (320 nM) for 24 h, THP-1 cells quickly stop proliferating,
become attached and differentiate into macrophages (25). According to this method, THP1 cells
were induced into M0, M1 and M2 phenotype macrophages. Then, the
surface markers of macrophages were detected by flow cytometry. The
results indicated that M0 (PMA only) and M1 (PMA/IFN-γ/LPS) cells
were positive for CD68, and M2 (PMA/IL-4/IL-13) cells were positive
for CD68 and CD163 (Fig. 2A). To
further confirm the different polarization of macrophages, the
cytokine profiles of THP1, M0-phenotype, M1-polarized and
M2-polarized THP-1 cells were assessed using RT-qPCR. The results
indicated that M1-polarized THP-1 macrophages expressed high levels
of TNF-α and IL-1β mRNA and low levels of TGF-β and IL-10 mRNA,
while the opposite results were observed in M2-polarized THP-1
macrophages (Fig. 2B). In
PMA-treated THP1 cells (M0 macrophages), the expression of TNF-α,
IL-1β, TGF-β and IL-10 was slightly increased when compared with
the THP1 control. The cytokine profiles and surface markers
provided evidence that the PMA-treated, M1-polarized and
M2-polarized THP-1 groups had been established successfully.
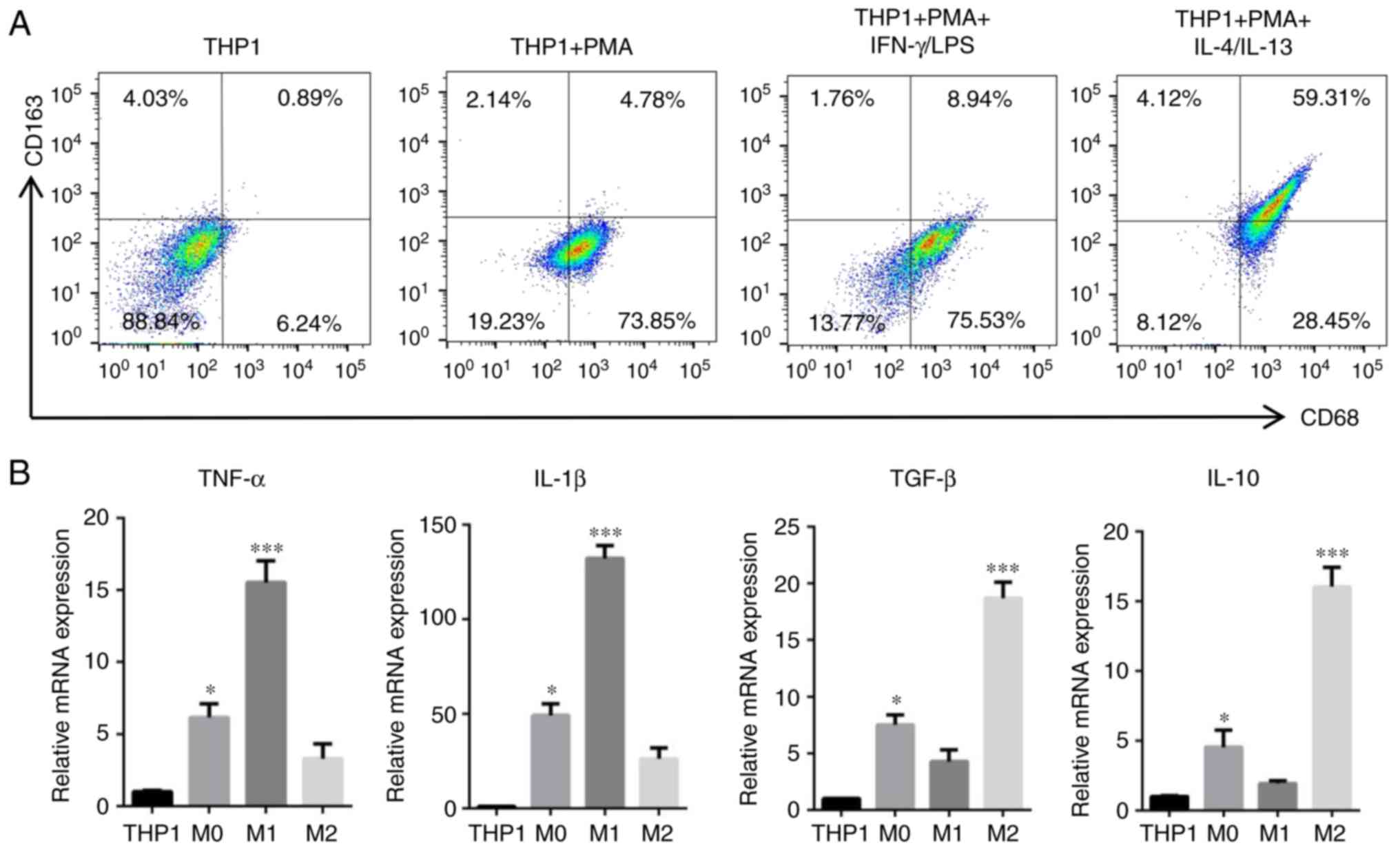 | Figure 2.Induction and acquirement of
macrophage polarization. Macrophages with M0, M1 and M2 functional
profiles were acquired from THP1 cells following treatment with
PMA, PMA/LPS/IFN-γ and PMA/IL-4/IL-13, respectively. (A) Expression
of classical M0, M1 and M2 polarized macrophage surface markers
(CD68 and CD163) was evaluated by flow cytometry. (B) mRNA
expression levels of TNF-α, IL-1β, TGF-β and IL-10 in
differentiated cells following induction were detected by reverse
transcription-quantitative polymerase chain reaction. Data are
expressed as the mean ± standard error of the mean. *P<0.05 and
***P<0.001 vs. THP1. PMA, phorbol-12-myristate-13-acetate; LPS,
lipopolysaccharide; CD, cluster of differentiation; IFN-γ,
interferon-γ; IL, interleukin; TNF-α, tumor necrosis factor-α;
TGF-β, transforming growth factor-β. |
In the direct co-culture system, THP1 cells were
first induced into M2 macrophages, then Cal27 cells labeled with
CFDA-SE were added to the dishes. Following co-culture for 24 h,
the Cal27 cells were separated by FACS. In the indirect co-culture
system, TAMs-CM was added to the Cal27 dishes for 24 h. Then, the
proteins of HNSCC cells were extracted and assessed (Fig. 3). The results from western blot
analysis indicated that E-cadherin in co-cultured Cal27 cells
(direct and indirect) was downregulated, while N-cadherin, Vimentin
and α-SMA were upregulated (Fig. 3A and
D). Similar results were obtained in SCC25 and Fadu cells
(Fig. 3B, C, E and F). Furthermore,
the protein expression of EMT-associated transcription factors,
including Slug, Snail and Twist, was increased in HNSCC cells that
were co-cultured with TAMs, particularly Slug (Fig. 3D-F). As the EMT of cancer cells
accelerates their migration, invasion and metastasis, the migration
and invasion ability of Cal27 cells was assessed using wound
healing and Transwell assays. In the wound healing assay, the
horizontal migration ability of Cal27 cells co-cultured with TAM-CM
or TAMs was markedly enhanced when compared with Cal27 only. In the
Transwell assay, for the Cal27 (TAM) group, 1×105 THP-1
cells were first seeded into upper inserts, treated with PMA for 6
h, then induced into M2-polarized macrophages with IL-4/IL-13 for
18 h. The inserts with M2-macrophages were washed to remove all PMA
and co-cultured with Cal27 cells (1×105 cells/well) in
24-well plates for a further 24 h. The results indicated that the
vertical migratory and invasive abilities of Cal27 were increased
by TAMs-CM or TAMs, particularly by TAMs (Fig. 3G). These results suggested that TAMs
induced EMT-like transformation of HNSCC cells in the direct and
indirect co-culture systems.
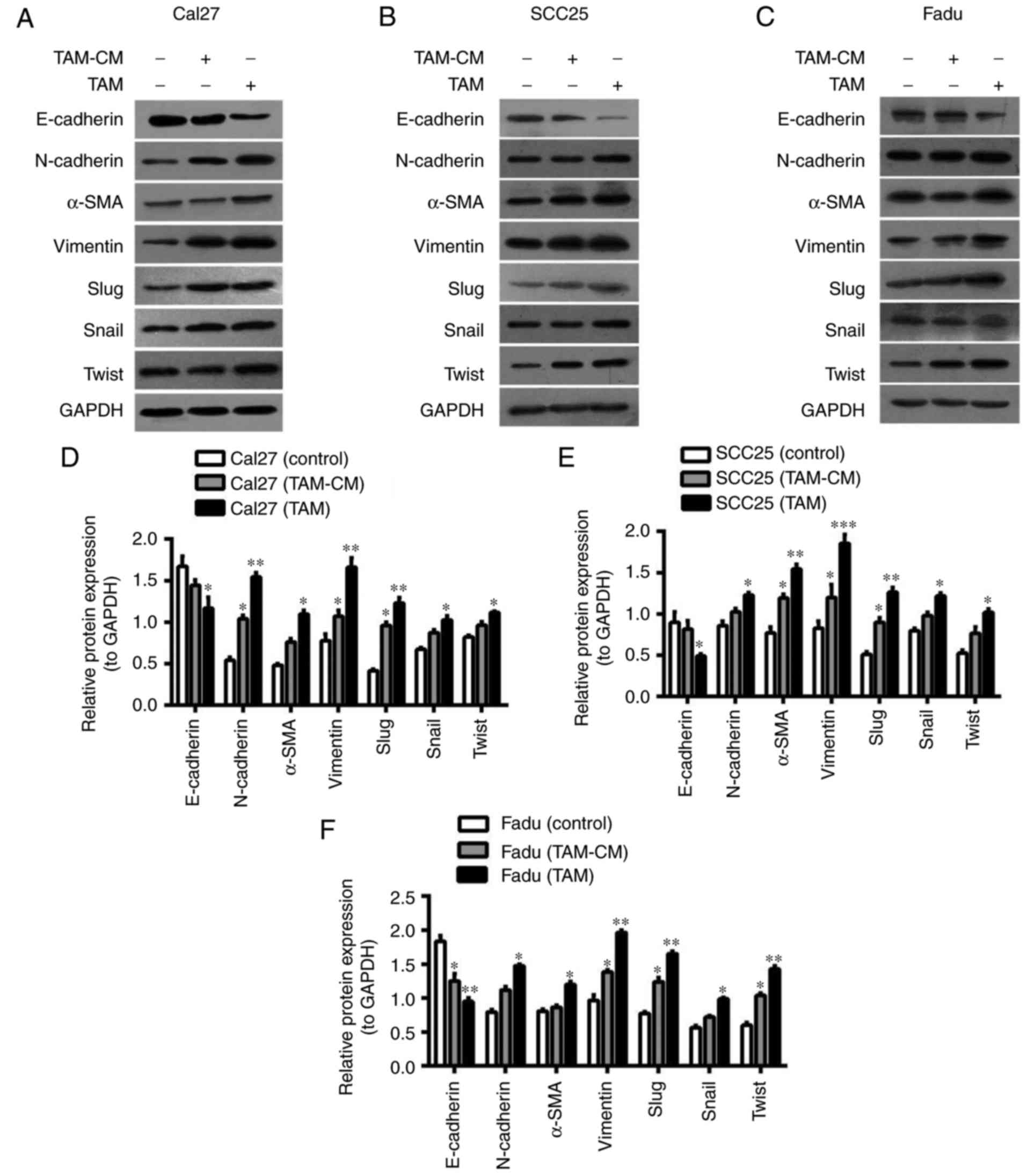 | Figure 3.TAMs induce the EMT of HNSCC cells.
Cal27 cells were labeled by carboxyfluorescein diacetate
succinimidyl ester, then added to the dishes that M2-macrophages
had been seeded in for 24 h. Following co-culture for 24 h, the
cells were sorted by fluorescence-activated cell sorting. (A-C)
Western blot analysis of the expression of EMT-associated proteins
(E-cadherin, N-cadherin, α-SMA, Vimentin, Slug, Snail and Twist) in
HNSCC cells: (A) Cal27, (B) SCC25 and (C) Fadu cells were
co-cultured with TAMs and TAM-CM. GADPH was used as an internal
control. (D-F) Statistical analysis of EMT-associated protein
expression of (D) Cal27, (E) SCC25 and (F) Fadu cells in the
co-culture system. (G) The migratory and invasive abilities of
Cal27 cells were measured by wound healing and Transwell assay
(magnification, ×200). Data are expressed as the mean ± standard
error of the mean. *P<0.05, **P<0.01 and ***P<0.001 vs.
the associated control. TAMs, tumor associated macrophages; EMT,
epithelial to mesenchymal transition; α-SMA, α-smooth muscle actin;
CM, conditional media. |
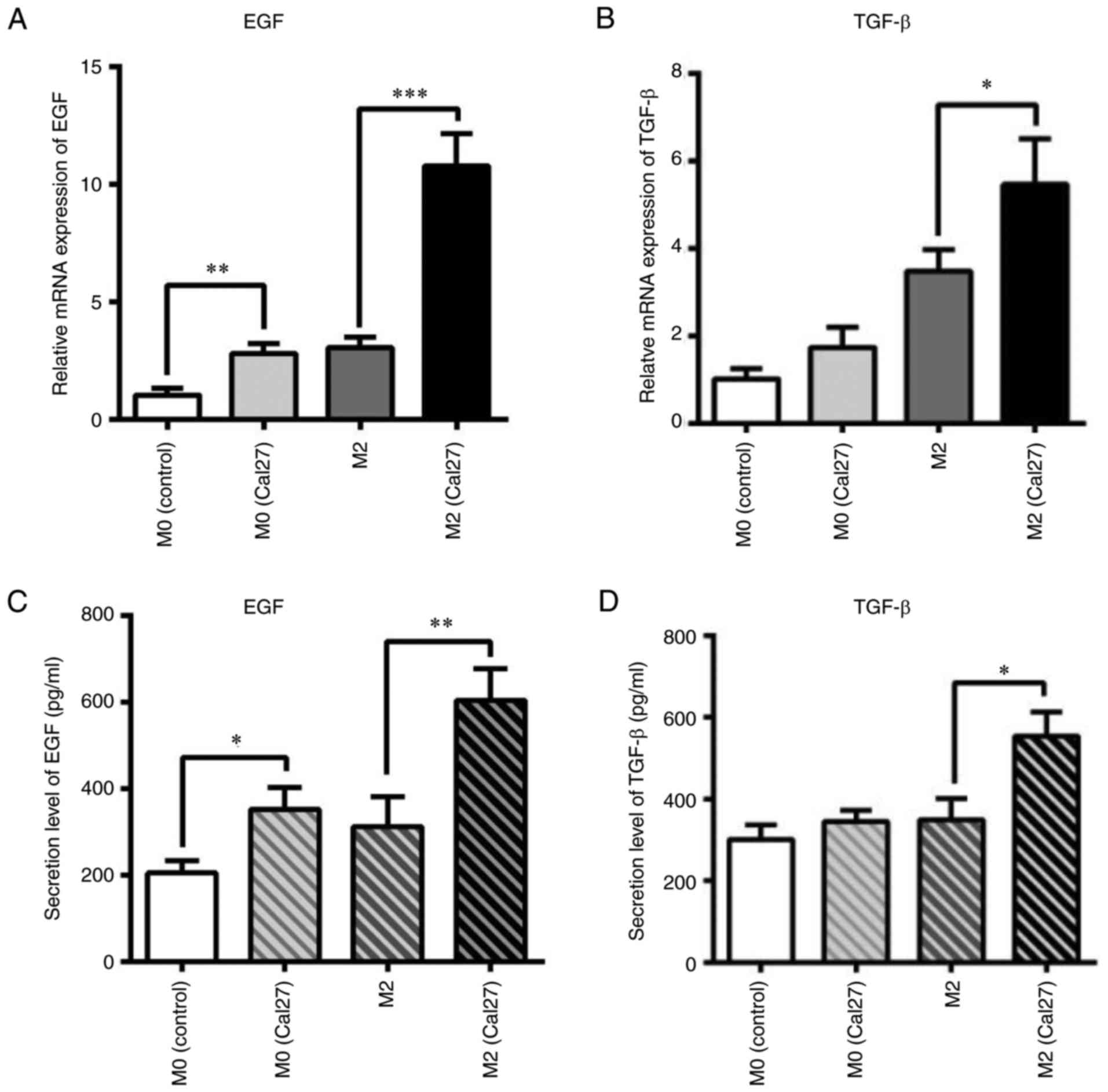
Macrophages secrete EGF and TGF-β
depending on the co-culture system in HNSCC cells
The aforementioned results suggested that TAMs may
promote an EMT-like process by secreting growth factors and
cytokines in the direct and indirect co-culture systems. Thus,
macrophages were labeled with CFDA-SE and sorted from the
co-cultured system. The mRNA expression of EMT-associated growth
factors and cytokines, including EGF and TGF-β, was measured by
qPCR. M0 macrophages induced from THP1 cells by treatment with PMA
were used as control. As shown in Fig.
4A and B, the mRNA expression of EGF and TGF-β was
significantly increased in the co-cultured macrophages when
compared with the control group. In particular, EGF was upregulated
by 5-fold (P<0.001). Furthermore, the secreted EGF and TGF-β
were analyzed by ELISA. The results were similar to those for mRNA
expression levels (Fig. 4C and D).
The EGF protein level in the direct co-culture system increased
from ~400 to ~700 pg/ml when compared with the control (P<0.01).
TGF-β level in the co-culture was also upregulated from ~400 to
~500 pg/ml when compared with the control (P<0.05).
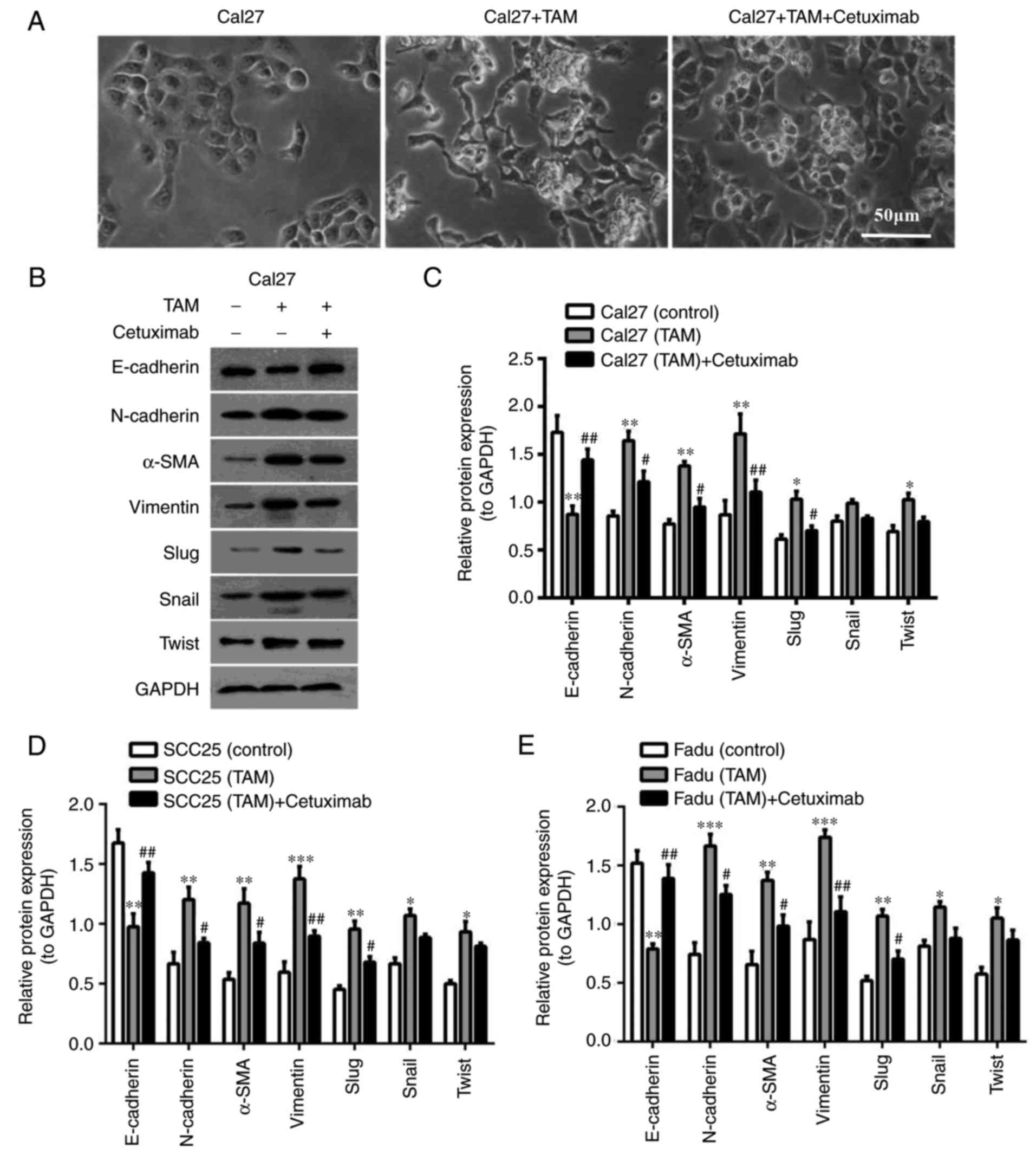
To investigate the function of EGF in this process,
the EGFR inhibitor Cetuximab (10 µg/ml) was added into the
co-culture system. As shown in Fig.
5A, the morphology of HNSCC cells was transformed from an
elongated, spindle-shaped appearance to cobblestones in the direct
co-culture with Cetuximab. Following co-culture for 24 h, the Cal27
cells were sorted. Western blot analysis of protein expression
indicated that E-cadherin was slightly increased, while N-cadherin,
Vimentin, α-SMA and transcription factors (Slug, Snail and Twist)
were significantly decreased in the co-cultured Cal27 cells treated
with Cetuximab (Fig. 5B and C). In
the co-culture with SCC25 or Fadu cells, the epithelial marker
E-cadherin was also upregulated and the mesenchymal markers were
downregulated with Cetuximab treatment (Fig. 5D and E). These results suggested
that EGF/EGFR may contribute to the TAM-induced EMT of HNSCC
cells.
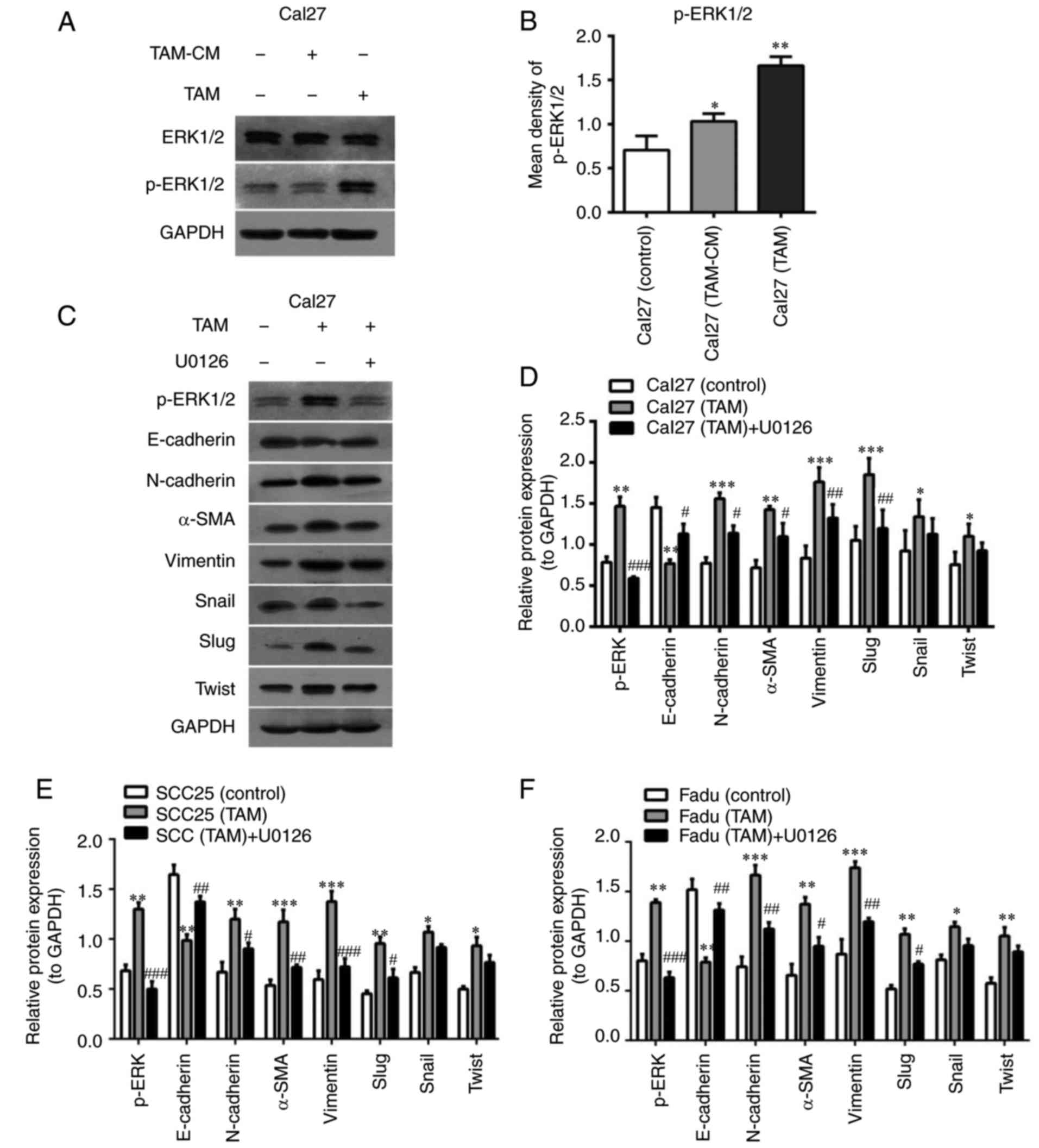
TAMs induce EMT via the ERK1/2
signaling pathway in HNSCC cells
Previous reports have suggested that certain
signaling pathways that are upregulated by EGF and TGF-β activation
lead to EMT and cellular migration (17,18,26).
Thus, signaling pathways including mitogen-activated protein kinase
(MAPK), Akt, Smad3 and nuclear factor-kB P65 (P65) were evaluated
to identify possible cascades involved in macrophage-mediated EMT.
Western blot analysis indicated that the p-ERK1/2 pathway was
significantly activated in the co-cultured HNSCC cells (Fig. 6A and B), while the signaling of Akt,
Smad3 and P65 was not markedly activated (data not shown).
U0126 is commonly used as an MAPK inhibitor. U0126
may selectively inhibit MAPK kinase 1 (MEK1) and MEK2, thereby
suppressing the phosphorylation and activation of ERK1/2. In the
present study, U0126 (10 mM) was added into the co-culture system,
which abolished the increase of N-cadherin, α-SMA and Vimentin
expression and restored E-cadherin levels in HNSCC cells induced by
TAMs (Fig. 6C and D). Furthermore,
the protein expression of transcription factors that repress
epithelial genes, including Slug, Snail and Twist, was also
assessed in the presence of U0126. The results indicated that the
expression of Slug was significantly downregulated (P<0.01), and
Snail and Twist were slightly decreased in the co-cultured Cal27
cells with U0126. The results from the co-cultured SCC25 and Fadu
cells were similar to those of Cal27 cells (Fig. 6E and F).
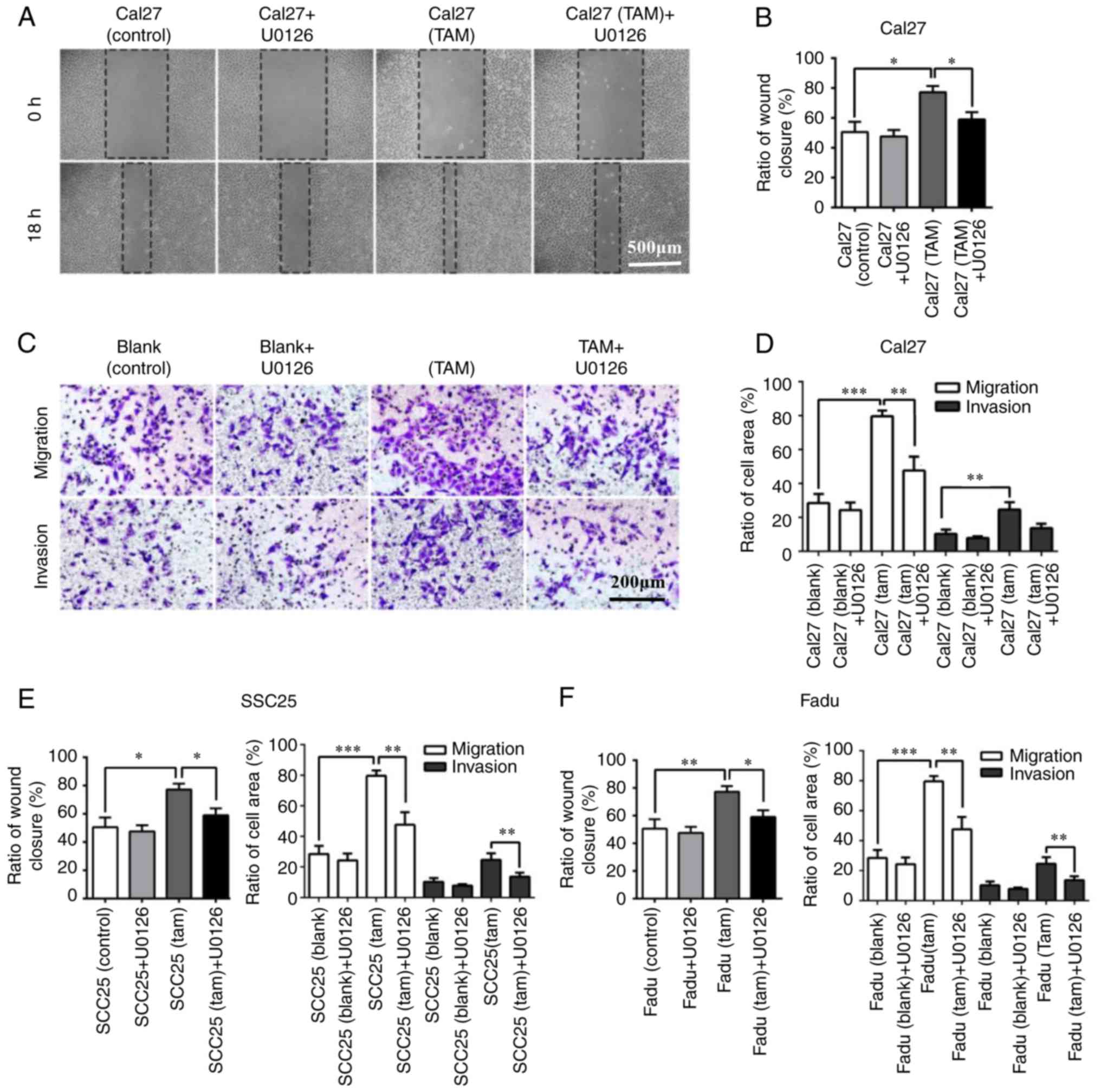
In the wound healing and Transwell assay, the
horizontal invasive and migratory abilities of HNSCC cells were
significantly impaired following 18 h of U0126 (10 µg/ml) treatment
in the co-culture system (Fig. 7A and
B). In addition, the vertical migratory and invasive abilities
of Cal27 cells were also inhibited following 24 h incubation with
U0126 (10 µg/ml; Fig. 7C and D).
The results of SCC25 and Fadu cells were consistent with the
results observed in Cal27 cells (Fig.
7E and F). In conclusion, these results indicated that TAMs
induced the EMT of HNSCC cells via the ERK1/2 signaling pathway,
and suppression of ERK1/2 markedly inhibited the EMT process of
HNSCC cells and diminished the potential motility of tumor cells in
the co-culture system.
Activation of the ERK1/2 signaling
pathway is positively correlated with EMT in HNSCC induced by
TAMs
The expression of p-ERK1/2, as well as the
biomarkers of EMT-associated genes, was evaluated in patient
tissues. These results were subjected to cluster analysis and then
visualized as a heatmap. The NM samples were strongly positive for
E-cadherin and negative or weakly positive for Vimentin, Slug and
p-ERK1/2. By contrast, the expression levels of E-cadherin were
significantly downregulated, while the expression levels of
Vimentin, α-SMA, Slug and p-ERK1/2 were markedly upregulated in the
majority of HNSCC samples, particularly the samples with high
infiltration of macrophages (data not shown). In Fig. 8, the correlation between tested
markers (left) and samples (top) was indicated by the length and
subdivision of the branches in the heatmap. The TAM marker CD163
and EMT-associated gene Vimentin were closely clustered, which was
consistent with the results from the Pearson correlation analysis.
In addition, the cluster analysis demonstrated the closest
association between EGFR and Slug. Furthermore, a close association
between Slug and p-ERK1/2 was also verified. All of the HNSCC
samples were clustered and could be divided into two groups, with
high and low infiltration of macrophages. Collectively, the cluster
analysis revealed that the EGFR/ERK1/2 signaling pathway was
closely associated with the EMT-associated markers and the
infiltration of TAMs.
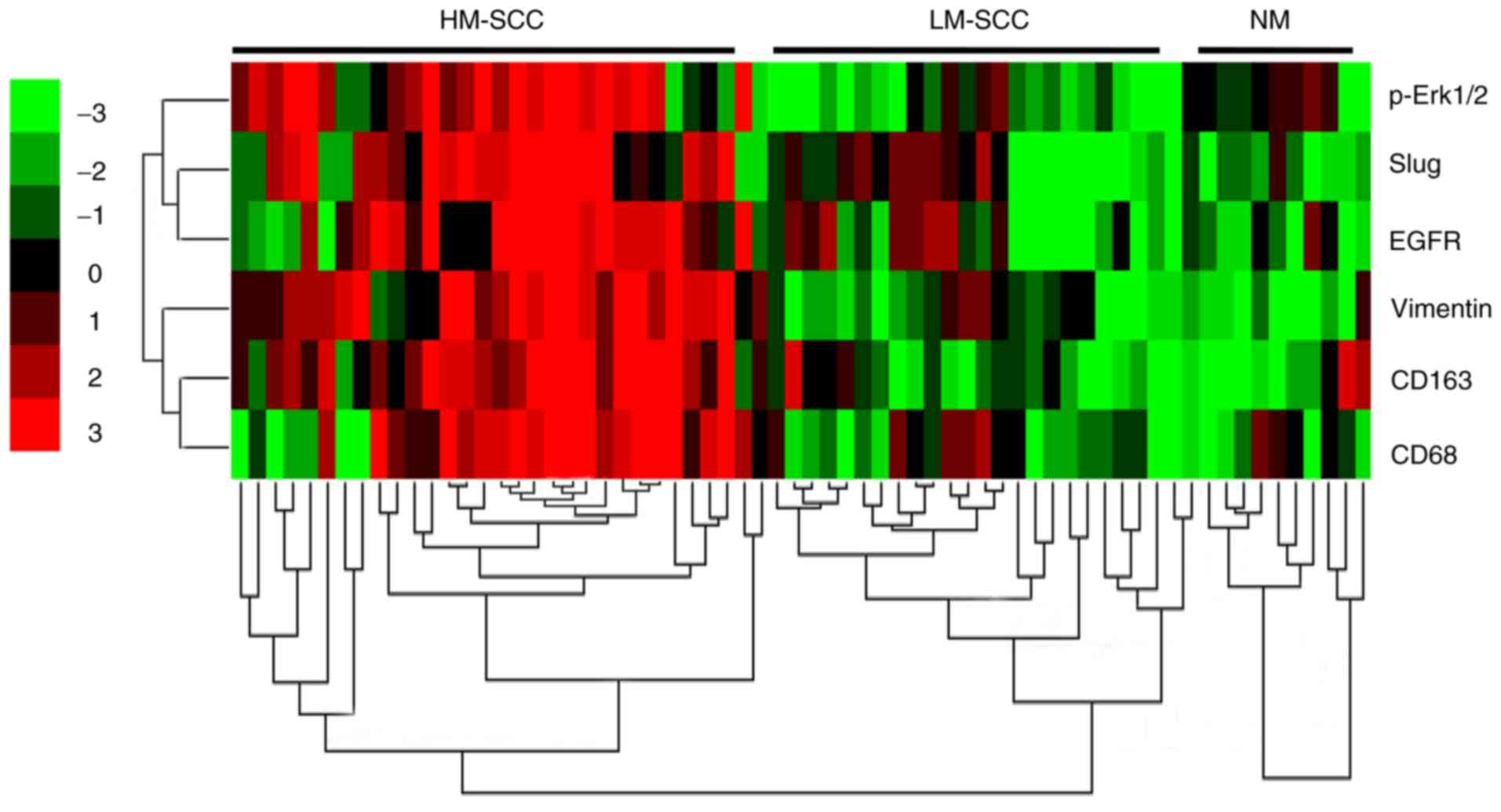 | Figure 8.Activation of the ERK1/2 signaling
pathway was positively correlated with the epithelial to
mesenchymal transition of tumor cells induced by tumor associated
macrophages in HNSCC tissues. Hierarchical clustering analysis of
CD68, CD163, Vimentin, EGFR, Slug and p-ERK1/2 in HNSCC and normal
tissues. ERK1/2, extracellular signal-regulated protein kinase 1/2;
CD, cluster of differentiation; EGFR, epidermal growth factor
receptor; p-, phosphorylated; HNSCC, head and neck squamous cell
carcinoma; HM-SCC, HNSCC with high macrophages; LM-SCC, HNSCC with
low macrophages; NM, normal mucosa. |
Discussion
HNSCC is thought to be caused by chronic
inflammation through smoking, repeated injuries or human
papillomavirus infection (27–29). A
number of previous studies have indicated that poor prognosis in
patients with HNSCC is associated with macrophage infiltration, and
the amount of macrophage infiltration may potentially be used as a
prognostic marker (21,30). Recently, we investigated the
interaction between monocytes and HNSCC cells (15). The results revealed that HNSCC cells
could recruit and differentiate monocytes into M2 macrophages when
HNSCC cells were co-cultured with monocytes. This process, in turn,
promoted the formation of aggressive invadopodia of cancer cells
(15). According to these results,
the aim of the present study was to investigate whether HNSCC cells
underwent EMT-like transformation induced by TAMs, so as to promote
invasion and metastasis. Furthermore, the underlying mechanism by
which TAMs induce the EMT of tumor cells was evaluated, which may
reveal novel targets for HNSCC therapy.
In the present study, M2 macrophage markers were
detected in HNSCC and normal adjacent mucosa. The results confirmed
our previous findings that the expression of CD68 and CD163 in
HNSCC was significantly higher when compared with NM, which
indicated that M2 macrophages are a primary cellular component in
the tumor stroma in HNSCC. M2-polarized macrophages may facilitate
tumor maintenance and growth by enhancing tumor-associated
angiogenesis, immunosuppression, invasion and metastasis (31,32).
EMT is a powerful metastasis process that describes a change from
polarized immotile epithelial cells to motile mesenchymal cells,
which leads to an increase in cell mobilization. This transition is
usually accompanied by a decrease in intercellular adhesive
molecules such as E-cadherin and β-catenin, and an increase in
mesenchymal cell markers such as Vimentin and N-cadherin, as well
as the upregulation of matrix metalloproteinase (12,33,34).
EMT is not only critical for appropriate embryonic development,
wound healing, tissue regeneration, organ fibrosis and cancer
progression, but also an important process in tumor invasion,
metastasis and tumor drug resistance (11,35,36). A
large body of evidence has indicated that TAMs also serve an
important role during the EMT of cancer cells, through which cancer
cells could disperse from primary sites and form metastatic lesions
(14,37–39).
In the present study, a close association between TAM infiltration
and the EMT of tumor cells was demonstrated in HNSCC, and the
mechanism of interaction between them was evaluated.
In the present study, EMT-associated markers were
detected in the human HNSCC samples, and then the correlation
between EMT and TAM infiltration was analyzed. The results
indicated that the expression of E-cadherin was downregulated,
while Vimentin and α-SMA were upregulated in the samples with high
infiltration of macrophages when compared with those with low
infiltration of macrophages and NM. Through correlation analysis,
the immunohistochemical staining of E-cadherin and Vimentin was
identified to be correlated with CD163+ cells. These results
indicated that EMT induced by TAMs may serve a role in the
aggressive behavior of HNSCC. To investigate the association
between TAMs and EMT in HNSCC cell lines, a direct and indirect
co-culture system was established between TAMs and HNSCC cells,
including Cal27, SCC25 and Fadu cells; the direct co-culture may
better reflect the real situation in tumor tissues. Tumor cells
were sorted from the co-culture system by FACS for further
analysis. All 3 HNSCC cell lines exhibited a significant decrease
in the epithelial marker E-cadherin, and an increase in the
mesenchymal markers N-cadherin, Vimentin and α-SMA. In conclusion,
these results indicated that TAMs could induce the EMT of HNSCC
cells in the indirect and direct co-culture system. Notably, the
EMT-like transformation appeared to be stronger in the direct
co-culture system when compared with the indirect co-culture
system, indicating that direct contact between cancer cells and
macrophages also contributes to the process.
Subsequently, the underlying mechanism was explored
further. A previous study has reported that TAMs promote the EMT of
tumor cells by secreting growth factors and cytokines (16). In the present study, it was observed
that the cytokines EGF and TGF-β in M2-macrophages directly
co-cultured with Cal27 were significantly increased when compared
with M2-macrophages alone. EGF is a growth factor highly associated
with the progression of HNSCC (18). TGF-β is associated with M2
macrophages and tumor cell proliferation, tissue fibrosis,
immunosuppressive activity and EMT. The natural ligands of EGFR
include EGF and TGF-β (40).
Therefore, Cetuximab, a monoclonal antibody that binds EGFR, was
added to the co-culture system. The data revealed that Cetuximab
could reverse the expression of EMT-associated proteins in Cal27
cells co-cultured with TAMs. These results indicated that the EGFR
signaling pathway may serve an important role in the EMT process in
HNSCC.
According to these results, the indirect and direct
co-culture systems could induce HNSCC cells to undergo the EMT
process. That is to say, TAMs induced the EMT of tumor cells, via
juxtacrine and paracrine signaling. Therefore, it was important to
evaluate intracellular signaling pathways in the present study. EMT
is governed by multiple molecular mechanisms, leading to the
phosphorylation and activation of intracellular signaling events,
including the MAPK, phosphoinositide 3-kinase/Akt, Smad3 and
protein kinase C signaling pathways. Together, these signaling
pathways result in the loss of apico-basal polarity, and the
acquisition of the motile and invasive phenotype (26,41).
In the present study, it was observed that the ERK1/2 signaling
pathway acted as the major regulator for EMT-like transformation in
HNSCC cells induced by TAMs. Signaling by receptor tyrosine kinases
to RAS-ERK-MAPK has been demonstrated to participate in the EMT of
various types of tumor cells (42,43).
In the present study, it was further revealed that pretreatment
with the ERK1/2-specific inhibitor U0126 reversed the EMT-like
process induced by TAMs due to the upregulation of E-cadherin and
downregulation of Vimentin and α-SMA. Simultaneously, the
EMT-associated transcription factors Slug, Snail and Twist were
also downregulated, particularly Slug in all 3 HNSCC cell lines. In
addition, it was demonstrated that U0126 in the co-culture system
could prevent the tumor cell migration and invasion of human HNSCC
cells. Collectively, these results indicated that activation of the
ERK1/2 signaling pathway served a major role in the induction of
EMT by TAMs in HNSCC cells. As the RAF-MEK-ERK cascade serves a
central role in the regulation of EMT, and cell migration and
invasion, together with the frequent deregulation of this pathway
in human cancer, it makes an attractive target for drug development
(44).
To improve the clinical relevance of these findings,
the expression status of p-ERK1/2 signaling and the EMT inducer
Slug was evaluated in HNSCC specimens, as well as EGFR, Vimentin
and CD163 expression. Hierarchical clustering analysis indicated a
close association between p-ERK1/2 and Slug, and also indicated
that the expression of TAM marker CD163 and EMT marker Vimentin
were closely associated with the expression of p-ERK1/2 and Slug.
This suggested that ERK1/2 may serve an important role in the EMT
process induced by TAMs in HNSCC, and could be a promising target
for the prevention of cancer metastasis.
In conclusion, the present study provides evidence
that TAMs could induce the EMT of HNSCC cells via the secretion of
EGF and TGF-β, which further enhanced the invasive ability of HNSCC
cells. In addition, it was indicated that the EMT process of HNSCC
cells induced by TAMs was partly via the ERK1/2 signaling pathway,
which may provide a novel target for cancer therapy in HNSCC.
Acknowledgements
Not applicable.
Funding
The present study was partially supported by the
National Natural Science Foundation of China (grant nos. 81570994,
81371159 and 81602780). It was also supported by the Young Elite
Scientist Sponsorship Program by China Association for Science and
Technology (CAST; grant no. 2015QNRC001) and the Open Research Fund
Program of Hubei-MOST KLOS & KLOBM (grant no. 201803).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YZ, WZ and LG conceived and designed the study. LG,
WqZ, ZL and HL performed the experiments. LG drafted the article.
ZY was also involved in the conception of the study and performed
the experiment. YZ and WZ reviewed and edited the manuscript. All
authors have read and approved the manuscript to be published.
Ethics approval and consent to
participate
All procedures were approved by the Medical Ethics
Committee of Wuhan University (Hubei, China). Experiments involving
human were approved by the Medical Ethics Committee of Dalian
Medical University (Dalian, China) and written informed consent was
provided by each patient.
Patient consent for publication
Patient consent for publication was obtained from
each patient.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
TAMs
|
tumor associated macrophages
|
|
EMT
|
epithelial to mesenchymal
transition
|
|
HNSCC
|
head and neck squamous cell
carcinoma
|
|
EGF
|
epithelial growth factor
|
|
ERK
|
extracellular regulated protein
kinases
|
References
|
1
|
Jemal A, Siegel R, Xu J and War E: Cancer
statistics, 2010. CA Cancer J Clin. 60:277–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jou A and Hess J: Epidemiology and
molecular biology of head and neck cancer. Oncol Res Treat.
40:328–332. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thompson L: World health organization
classification of tumours: Pathology and genetics of head and neck
tumours. Ear Nose Throat J. 85:742006.PubMed/NCBI
|
|
4
|
Rivera C: Essentials of oral cancer. Int J
Clin Exp Pathol. 8:11884–11894. 2015.PubMed/NCBI
|
|
5
|
Solinas G, Germano G, Mantovani A and
Allavena P: Tumor-associated macrophages (TAM) as major players of
the cancer-related inflammation. J Leukoc Biol. 86:1065–1073. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gordon S and Taylor PR: Monocyte and
macrophage heterogeneity. Nat Rev Immunol. 5:953–964. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gordon S: Alternative activation of
macrophages. Nat Rev Immunol. 3:23–35. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Mantovani A, Sozzani S, Locati M, Allavena
P and Sica A: Macrophage polarization: Tumor-associated macrophages
as a paradigm for polarized M2 mononuclear phagocytes. Trends
Immunol. 23:549–555. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pollard JW: Tumour-educated macrophages
promote tumour progression and metastasis. Nat Rev Cancer. 4:71–78.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pollard JW: Macrophages define the
invasive microenvironment in breast cancer. J Leukoc Biol.
84:623–630. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nieto MA, Huang RY, Jackson RA and Thiery
JP: EMT: 2016. Cell. 166:21–45. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lu H, Clauser KR, Tam WL, Fröse J, Ye X,
Eaton EN, Reinhardt F, Donnenberg VS, Bhargava R, Carr SA, et al: A
breast cancer stem cell niche supported by juxtacrine signalling
from monocytes and macrophages. Nat Cell Biol. 16:1105–1117. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Su S, Liu Q, Chen J, Chen J, Chen F, He C,
Huang D, Wu W, Lin L, Huang W, et al: A positive feedback loop
between mesenchymal-like cancer cells and macrophages is essential
to breast cancer metastasis. Cancer Cell. 25:605–620. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gao L, Wang FQ, Li HM, Yang JG, Ren JG, He
KF, Liu B, Zhang W and Zhao YF: CCL2/EGF positive feedback loop
between cancer cells and macrophages promotes cell migration and
invasion in head and neck squamous cell carcinoma. Oncotarget.
7:87037–87051. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pirilä E, Väyrynen O, Sundquist E, Päkkilä
K, Nyberg P, Nurmenniemi S, Pääkkönen V, Pesonen P, Dayan D, Vered
M, et al: Macrophages modulate migration and invasion of human
tongue squamous cell carcinoma. PLoS One. 10:e01208952015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Quan J, Elhousiny M, Johnson NW and Gao J:
Transforming growth factor-β1 treatment of oral cancer induces
epithelial-mesenchymal transition and promotes bone invasion via
enhanced activity of osteoclasts. Clin Exp Metastasis. 30:659–670.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu Q, Zhang Q, Ishida Y, Hajjar S, Tang X,
Shi H, Dang CV and Le AD: EGF induces epithelial-mesenchymal
transition and cancer stem-like cell properties in human oral
cancer cells via promoting Warburg effect. Oncotarget. 8:9557–9571.
2017.PubMed/NCBI
|
|
19
|
National Institutes of Health (NIH): HIPAA
Authorization for Research. NIH Publication No. 04-5529. Bethesda,
MD: April. 2004
|
|
20
|
Zhang W, Chen G, Wang FQ, Ren JG, Zhu JY,
Cai Y, Zhao JH, Jia J and Zhao YF: Macrophages contribute to the
progression of infantile hemangioma by regulating the proliferation
and differentiation of hemangioma stem cells. J Invest Dermatol.
135:3163–3172. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
He KF, Zhang L, Huang CF, Ma SR, Wang YF,
Wang WM, Zhao ZL, Liu B, Zhao YF, Zhang WF and Sun ZJ: CD163+
tumor-associated macrophages correlated with poor prognosis and
cancer stem cells in oral squamous cell carcinoma. Biomed Res Int.
2014:8386322014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chui X, Egami H, Yamashita J, Kurizaki T,
Ohmachi H, Yamamoto S and Ogawa M: Immunohistochemical expression
of the c-kit proto-oncogene product in human malignant and
non-malignant breast tissues. Br J Cancer. 73:1233–1236. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhong WQ, Chen G, Zhang W, Xiong XP, Ren
JG, Zhao Y, Liu B and Zhao YF: Down-regulation of connexin43 and
connexin32 in keratocystic odontogenic tumours: Potential
association with clinical features. Histopathology. 66:798–807.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tjiu JW, Chen JS, Shun CT, Lin SJ, Liao
YH, Chu CY, Tsai TF, Chiu HC, Dai YS, Inoue H, et al:
Tumor-associated macrophage-induced invasion and angiogenesis of
human basal cell carcinoma cells by cyclooxygenase-2 induction. J
Invest Dermatol. 129:1016–1025. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gan Y, Shi C, Inge L, Hibner M, Balducci J
and Huang Y: Differential roles of ERK and Akt pathways in
regulation of EGFR-mediated signaling and motility in prostate
cancer cells. Oncogene. 29:4947–4958. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lambert R, Sauvaget C, de Camargo Cancela
M and Sankaranarayanan R: Epidemiology of cancer from the oral
cavity and oropharynx. Eur J Gastroenterol Hepatol. 23:633–641.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li H, Wawrose JS, Gooding WE, Garraway LA,
Lui VW, Peyser ND and Grandis JR: Genomic analysis of head and neck
squamous cell carcinoma cell lines and human tumors: A rational
approach to preclinical model selection. Mol Cancer Res.
12:571–582. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Suh Y, Amelio I, Urbano Guerrero T and
Tavassoli M: Clinical update on cancer: Molecular oncology of head
and neck cancer. Cell Death Dis. 5:e10182014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ji WT, Chen HR, Lin CH, Lee JW and Lee CC:
Monocyte chemotactic protein 1 (MCP-1) modulates pro-survival
signaling to promote progression of head and neck squamous cell
carcinoma. PLoS One. 9:e889522014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Colegio OR, Chu NQ, Szabo AL, Chu T,
Rhebergen AM, Jairam V, Cyrus N, Brokowski CE, Eisenbarth SC,
Phillips GM, et al: Functional polarization of tumour-associated
macrophages by tumour-derived lactic acid. Nature. 513:559–563.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Valastyan S and Weinberg RA: Tumor
metastasis: Molecular insights and evolving paradigms. Cell.
147:275–292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Garg M: Epithelial-mesenchymal
transition-activating transcription factors-multifunctional
regulators in cancer. World J Stem Cells. 5:188–195. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mego M, Gao H, Lee BN, Cohen EN, Tin S,
Giordano A, Wu Q, Liu P, Nieto Y, Champlin RE, et al: Prognostic
value of EMT-circulating tumor cells in metastatic breast cancer
patients undergoing high-dose chemotherapy with autologous
hematopoietic stem cell transplantation. J Cancer. 3:369–380. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Heldin CH, Vanlandewijck M and Moustakas
A: Regulation of EMT by TGFβ in cancer. FEBS Lett. 586:1959–1970.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fu XT, Dai Z, Song K, Zhang ZJ, Zhou ZJ,
Zhou SL, Zhao YM, Xiao YS, Sun QM, Ding ZB and Fan J:
Macrophage-secreted IL-8 induces epithelial-mesenchymal transition
in hepatocellular carcinoma cells by activating the
JAK2/STAT3/Snail pathway. Int J Oncol. 46:587–596. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bonde AK, Tischler V, Kumar S, Soltermann
A and Schwendener RA: Intratumoral macrophages contribute to
epithelial-mesenchymal transition in solid tumors. BMC Cancer.
12:352012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Techasen A, Loilome W, Namwat N, Dokduang
H, Jongthawin J and Yongvanit P: Cytokines released from activated
human macrophages induce epithelial mesenchymal transition markers
of cholangiocarcinoma cells. Asian Pac J Cancer Prev. 13
Suppl:S115–S118. 2012.
|
|
40
|
Kalyankrishna S and Grandis JR: Epidermal
growth factor receptor biology in head and neck cancer. J Clin
Oncol. 24:2666–2672. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu ZC, Chen XH, Song HX, Wang HS, Zhang
G, Wang H, Chen DY, Fang R, Liu H, Cai SH, et al: Snail regulated
by PKC/GSK-3β pathway is crucial for EGF-induced
epithelial-mesenchymal transition (EMT) of cancer cells. Cell
Tissue Res. 358:491–502. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Han M, Liu M, Wang Y, Chen X, Xu J, Sun Y,
Zhao L, Qu H, Fan Y and Wu C: Antagonism of miR-21 reverses
epithelial-mesenchymal transition and cancer stem cell phenotype
through AKT/ERK1/2 inactivation by targeting PTEN. PLoS One.
7:e395202012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Xie YX, Liao R, Pan L and Du CY: ERK
pathway activation contributes to the tumor-promoting effects of
hepatic stellate cells in hepatocellular carcinoma. Immunol Lett.
188:116–123. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Montagut C and Settleman J: Targeting the
RAF-MEK-ERK pathway in cancer therapy. Cancer Lett. 283:125–134.
2009. View Article : Google Scholar : PubMed/NCBI
|