Introduction
Prostate cancer (PCa) is the most common
non-cutaneous malignancy in men (1). PCa tumorigenesis is involved in
multiple genomic alterations (2).
For example, the oncogene c-Myc is upregulated in PCa, which
is associated with tumor progression (3,4). A
previous study revealed that an HP1γ/miR-451a/c-Myc feedback loop
exists in PCa cells and this loop is essential for PCa development
(5). However, the underlying
molecular mechanisms of c-Myc in PCa remain incompletely
understood.
Accumulating evidence indicates that non-coding RNAs
(ncRNAs), such as microRNAs (miRNAs, <200 nt) and long ncRNAs
(lncRNAs, >200 nt) (6), are
involved in numerous processes of tumorigenesis (7,8).
Accumulating data suggest that lncRNAs, such as PCA3
(9), PCAT-1 (4), SChLAP1 (10), PCGEM1 and PRNCR1
(11) play important roles in PCa
tumorigenesis. lncRNAs can regulate coding gene mRNAs via
competitively binding to co-target miRNAs, termed as competing
endogenous RNA (ceRNA) network, thus functioning in cancer
(12,13). The molecular mechanism of most
lncRNAs in PCa remains unknown due to their enormousness.
A c-Myc-upregulated lncRNA MYU
(ENSG00000261373) has been reported to exert an oncogenic effect in
colorectal cancer (CRC) (14,15),
but functions as a tumor-suppressor gene in gastric cancer (GC)
(16). A recent study suggests that
MYU as a target for Wnt/c-Myc signaling can promote cell
cycle progression by stabilizing CDK6 expression via associating
with hnRNP-K (14). However, the
functional roles and underlying molecular mechanisms of MYU
in PCa have not been explored. Thus, we were motivated to study the
roles of MYU in PCa.
In the present study, we revealed the clinical
significance and function of MYU in PCa, and demonstrated
that MYU promotes the progression of PCa as a ceRNA to
modulate miR-184 binding, consequently regulating c-Myc expression.
Our results shed new light on the prospect for MYU as a
potential novel diagnostic biomarker as well as a therapeutic
target.
Materials and methods
Cell culture
Human PCa cell lines DU145, PC3, 22Rv1 and LNCaP
were obtained from the Shanghai Institute of Biochemistry and Cell
Biology (CAS). Lenti-X™293T cells were obtained from Clontech
Laboratories, Inc. (Mountainview, CA, USA). Cell lines were
cultured routinely in RPMI-1640 medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) or DMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% fetal bovine serum (FBS,
Gibco; Thermo Fisher Scientific, Inc.), penicillin and streptomycin
(×100). All cells were maintained in 5% CO2 at 37°C. PCR
detection method was used to check mycoplasma contamination. All
cell lines were verified using short tandem repeat assays by
Genetic Testing Biotechnology, Suzhou, China.
5′/3′ Rapid amplification of cDNA ends (5′/3′ RACE).
The complementary DNA (cDNA) was synthesized using RNA (1.0 µg)
extracted from PC3 cells. The 5′/3′ RACE was carried out using the
SMARTer™ RACE cDNA Amplification kit (Clontech Laboratories, Inc.)
according to the manufacturer's instructions. The gene-specific
primers used for the PCR of the RACE analysis are given in Table I.
 | Table I.Primers for the 5′/3′ RACE. |
Table I.
Primers for the 5′/3′ RACE.
| Primers | Sequence |
|---|
| P1 | 5′
CTCTGGGAAGTGGCCGTTTT 3′ |
| P2 | 5′
GCGCTCCAAGATTGAGGAGT 3′ |
| P3 | 5′
CTGTTCGTACTCCAGGATGGC3′ |
| P4 | 5′
CATGCCAAGCTACGGGAAGG 3′ |
| P5 | 5′
TCTGTCTTCTGGGACTTTGCTC 3′ |
| P6 | 5′
AGGATGGGAGCAGTAAACGG 3′ |
| 5′-F | 5′
AGGCGGCTCTGACCCCG 3′ |
| 3′-R | 5′
ATCTCCCAGGTGTTACTAGAAGAAGTGG 3′ |
| GSP1 | 5′
GATTACGCCAAGCTTTTTGCTCCTGCCCCATCTGCC 3′ |
| GSP2 | 5′
GATTACGCCAAGCTTGGCAGATGGGGCAGGAGCAAA 3′ |
Plasmid construction
The full-length of MYU (Fig. 1) and VPS9D1, which is fused
with a FLAG epitope tag with the C terminus, Fig. 2) were subcloned into pLVX-IRES-Neo
vector. The short hairpin RNAs (shRNAs) that target different sites
of MYU, VPS9D1 and c-Myc CDS region were inserted
into the pSIH1-H1 vector. The shRNA primers are listed in Table II. MYU full-length and 3′
untranslated regions (UTR) of c-Myc containing the miR-184 binding
sites were synthesized in the dual luciferase reporter vector
pmirGLO (Promega Corporation, Madison, WI, USA).
| Table II.Primer sequences of MYU,
VPS9D1 and c-Myc shRNAs. |
Table II.
Primer sequences of MYU,
VPS9D1 and c-Myc shRNAs.
| shRNA | Primers | Primer
sequence |
|---|
| MYU shRNA-1 | Forward | 5′
GATCCGACCATGGACCTGCTCACACAGCCCTTCTTTCAAGAGAAGAAGGGCTGTGTGAGCAGGTCCATGGTCTTTTTTG
3′ |
|
| Reverse | 5′
AATTCAAAAAAGACCATGGACCTGCTCACACAGCCCTTCTTCTCTTGAAAGAAGGGCTGTGTGAGCAGGTCCATGGTCG
3′ |
| MYU shRNA-2 | Forward | 5′
GATCCGCCACTGGAGTTCCTCTGTCTTCTGGGACTTCAAGAGAGTCCCAGAAGACAGAGGAACTCCAGTGGCTTTTTTG
3′ |
|
| Reverse | 5′
AATTCAAAAAAGCCACTGGAGTTCCTCTGTCTTCTGGGACTCTCTTGAAGTCCCAGAAGACAGAGGAACTCCAGTGGCG
3′ |
| VPS9D1 shRNA-1 | Forward | 5′
GATCCTCCACTATATCTCCCAGGTGTTACTAGAATTCAAGAGATTCTAGTAACACCTGGGAGATATAGTGGATTTTTTG
3′ |
|
| Reverse | 5′
AATTCAAAAAATCCACTATATCTCCCAGGTGTTACTAGAATCTCTTGAATTCTAGTAACACCTGGGAGATATAGTGGAG
3′ |
| VPS9D1 shRNA-2 | Forward | 5′
GATCCGCAGATGATGGAGAACCTAGTTTCAAGAGAACTAGGTTCTCCATCATCTGCTTTTTTG
3′ |
|
| Reverse | 5′
AATTCAAAAAAGCAGATGATGGAGAACCTAGTTCTCTTGAAACTAGGTTCTCCATCATCTGCG
3′ |
| c-Myc shRNA | Forward | 5′
GATCCCAGCGAGGATATCTGGAAGAAATTCGAGCTTCAAGAGAGCTCGAATTTCTTCCAGATATCCTCGCTGTTTTTTG
3′ |
|
| Reverse | 5′
AATTCAAAAAACAGCGAGGATATCTGGAAGAAATTCGAGCTCTCTTGAAGCTCGAATTTCTTCCAGATATCCTCGCTGG
3′ |
RNA extraction, quantitative real-time
PCR (qRT-PCR) and Western blotting
Total RNA was isolated from PCa cells using RNAiso
Plus reagent (Takara, Shiga, Japan) and RNA (1 µg) was reverse
transcribed using the PrimerScript RT-PCR kit (Takara) according to
the manufacturer's instructions. The expression levels of
MYU were determined by qRT-PCR using the SYBR®
Green dye (Takara) detection method. We used the comparative Ct
method to quantify transcripts. The relative gene expression was
defined using the equation: ∆Ct=Ct target - Ct
reference, and the fold-change for target genes
normalized by GAPDH was determined by the formula
2−∆∆Ct.
Total cell lysates were prepared using RIPA reagent
(Beyotime, Shanghai, China) and sodium dodecyl sulfate (SDS)
loading buffer (Beyotime). The concentration of total proteins was
examined using BCA protein kit (Beyotime). Equivalent quantities of
proteins were loaded to each lane. SDS-polyacrylamide gel
electrophoresis (SDS-PAGE) was used to separate proteins and then
the proteins were transferred from the gel onto NC or PVDF
membranes. Membranes were incubated with primary antibodies against
GAPDH (AG019, 1:5,000), H3 (AH433, 1:5,000) and β-actin (AA128,
1:5,000) all from Beyotime, Flag (F3165, 1:1,000) from
Sigma-Aldrich; Merck KGaA (Darmstadt, Germany), ZO-1 (8193S,
1:1,000), caludin-1 (13995S, 1:1,000), E-cadherin (3195S, 1:1,000),
and c-Myc (13987S, 1:1,000) from Cell Signaling Technology, Inc.
(Danvers, MA, USA), N-cadherin (ab76011, 1:1,000), vimentin
(ab92547, 1:1,000) from Abcam (Cambridge, MA, USA). After
incubation with suitable HRP-conjugated secondary antibodies,
immunoreactive proteins were finally visualized using ECL
chemiluminescence system (CLiNX, Shanghai, China). All qRT-PCR
primers are listed in Table
III.
| Table III.qPCR sequences of MYU, VPS9D1, c-Myc
and U1. |
Table III.
qPCR sequences of MYU, VPS9D1, c-Myc
and U1.
| Gene | Primers | Sequence |
|---|
| MYU | Forward | 5′
AGTGGCCGTTTTACAGAGACA 3′ |
|
| Reverse | 5′
CATGCCAAGCTACGGGAAGG 3′ |
| GAPDH | Forward | 5′
GGAGCGAGATCCCTCCAAAAT 3′ |
|
| Reverse | 5′
GGCTGTTGTCATACTTCTCATGG 3′ |
| VPS9D1 | Forward | 5′
AGATCCACAATGCCGTAGAC 3′ |
|
| Reverse | 5′
CTTGGAGGCAGCTGTGTTTAG 3′ |
| c-Myc | Forward | 5′
TAGTGGAAAACCAGCAGCCT 3′ |
|
| Reverse | 5′
AGAAATACGGCTGCACCGAG 3′ |
| U1 | Forward | 5′
GGGAGATACCATGATCACGAAGGT 3′ |
|
| Reverse | 5′
CCACAAATTATGCAGTCGAGTTTCCC 3′ |
Subcellular fractionation
LNCaP cells were captured with 1 ml PBS and 200 µl
were extracted as the positive control, the reminder were
snap-frozen with liquid nitrogen and then quickly dissolved in a
water bath, repeating this operation twice. Then, 200 µl buffer A1
(10 mM HEPES, pH 7.9, 1.5 mM MgCl2, 10 mM KCl, 0.1%
NP-40 and 1% protease inhibitor) was added to the cells, and ground
15–20 times with a grinding pestle. After incubation on ice for 5
min, the lysate was centrifuged at 2,000 rpm for 5 min at 4°C. The
supernatant was used to separate the cytoplasmic fraction. The
remaining pellet was resuspended in 200 µl of buffer A2 (10 mM
HEPES, pH 7.9, 1.5 mM MgCl2, 10 mM KCl, and 1% protease
inhibitor) and ground 5–10 times with a grinding pestle. Then, the
lysate was resuspended in another 300 µl of buffer A2. After
incubating on ice for 5 min, the lysate was centrifuged at 2000 rpm
for 5 min at 4°C. The remaining pellet was resuspended in 500 µl of
buffer S1 (0.25 mM sucrose, 10 mM MgCl2, and 1% protease
inhibitor), and then added 500 µl of buffer S2 (0.35 mM sucrose,
0.5 mM MgCl2, and 1% protease inhibitor) slowly from the
top of the liquid. The lysate was centrifuged at 3,500 rpm for 5
min at 4°C. The remaining pellet was resuspended in 200 µl RIPA
buffer (50 mM Tris, pH 7.5, 150 mM NaCl, 1% NP-40, and 0.5%
deoxycholate) and incubated on ice for 30 min. The supernatant
containing the nuclear fraction was collected by centsrifugation at
12,000 rpm for 15 min at 4°C. qRT-PCR and western blot analysis
were used to assess the total RNA and protein isolated from the
subcellular fractions. GAPDH served as the cytoplasmic control, and
U1 as nuclear control. GAPDH, U1 and MYU expression levels
were measured by qRT-PCR.
Cell transfection
The packaging plasmids, recombined plasmids and LNAs
for overexpression and knockdown were transfected into Lenti-X™
293T cells. Two LNAs (LNA-1 and LNA-2) and LNA-Ctrl (300610,
Negative control A Antisense LNA GapmeR) were purchased from Exiqon
A/S (Vedbaek, Denmark). The hsa-miR-184 (miR-184) mimics and the
negative control miR-NC were purchased from RiboBio (Guangzhou,
China). Cells were harvested 48 h after transfection. These
corresponding plasmids or miRNA mimics were transfected with PEI or
Lipofectamine 3000 (Invitrogen; Thermo Fisher Scientific, Inc.).
The final concentrations of miRNAs/LNAs employed in this study were
as follows: miR-184 mimic/negative control mimic 50 nM/ml and
LNA-1/LNA-2/LNA-Ctrl 50 nM/ml.
Cell viability, colony formation and
migration assay
Cells were seeded into 96-well plates at a density
of 1.0×103 cells/well. Cell viability was assessed using
CellTiter-Glo (CTG) reagent (Promega Corporation) every 48 h
following the manufacturer's instructions. In addition,
1.0×103 cells were seeded per well of 6-well plates.
After 2 weeks, the number of clones was counted with staining using
ImageJ (https://imagej.nih.gov/ij/). Wound
healing and Transwell chamber assays were performed to determine
cell migration ability. For wound healing assay, cells were seeded
into two well silicones (ibidi, GmbH, Munich, Germany) in 24-well
plates. When the cell density reached 90–100% confluence, the
inserts were removed and washed with PBS, and then cultured with
serum-free medium. Images were collected at 0 and 24 h under an
inverted microscope (Ziess, Jena, Germany). Wound healing images
were analyzed using ImageJ. For the Transwell chamber assay, the
cell suspension, 3.5×104 cells/well in 200 µl, was added
to a 8.0-µm pore-size insert of a 24-well plate (Corning Costar,
Corning, NY, USA). The lower well was filled with 500 µl medium
with 10% FBS. The Transwell plate was then incubated for 24 h at
37°C with 5% CO2. Hereafter, the cells on the top
surface of the insert were wiped off and the cells on the lower
surface of the insert were fixed with 95% ethanol for 10–15 min,
and then dyed with 0.5% crystal violet. Images were collected under
an inverted microscope. Finally, the cells were decolorized with
95% ethanol and the same amount of eluent was used to measure the
absorbance at 580 nm wavelength. These experiments were conducted
in triplicate.
Exosome isolation and treatment
PC3 cells were grown to a 70–80% confluent
monolayer, washed three times with PBS, and grown in serum-free
culture medium for 3 days. Exosomes were collected by gradient
centrifugation, cleaned by PBS, and stored at −80°C.
Exosomes were isolated from the
MYU-overexpressing or control PC3 supernatant. An
appropriate amount of serum-free 1640 medium was added and filtered
with a 0.22-µm filter. For cell viability assay, PC3 cells were
plated in a 96-well plate on the first day, and then treated with
exosomes from MYU-overexpressing or control PC3 cells, which
were assessed by CTG assays after incubation for 5 days. For
Transwell assays, the PC3 cell suspension, 3.5×104
cells/well in 200 µl, was added to an 8.0-µm pore-size insert of a
24-well plate. The lower well contained 500 µl exosomes and 10%
FBS. The Transwell plate was then incubated for 24 h at 37°C with
5% CO2.
Dual luciferase reporter assay
Lenti-X™ 293T cells grown in a 24-well plate were
co-transfected with 200 ng of dual luciferase reporter vector
comprising MYU and its mutant or 3′UTR of c-Myc, 50 nM miRNA
mimics using Lipofectamie 3000. Cells were harvested 48 h after
transfection and analyzed using the dual luciferase reporter assay
kit (Promega Corporation) with a microplate reader (BioTek
Instruments, Inc., Winooski, VT, USA) according to the
manufacturer's protocol.
lncRNA, miRNA, and mRNA expression
data
The expression level of lncRNA MYU in
prostate adenocarcinoma (PRAD) based on the clinical information
was obtained from The Cancer Genome Atlas (TCGA) (17) and the matched expression profiles of
MYU in Mitranscriptome (18). TCGA PRAD patient miRNA expression
(Level 3 data, illuminahiseq mirnaseq) and mRNA expression (level 3
data, RNA-seq Version 2) was download from the Broad GDAC Firehose
(19).
ChIP-Seq data
Sequencing data from GSE96019, GSE96399 and GSE96418
were downloaded from GEO. Reads from the PC3 H3K4me3, H3K27ac and
H3K36me3 ChIP-Seq samples were mapped to human genome version hg19
using HISAT2. Peak calling was performed using MACS (20) according to the published protocols
(21). Data were visualized using
the UCSC Genome Browser (22).
Statistical analysis
The data were collected and expressed as means ±
SEM. Statistical differences (P-values) between two groups were
obtained by using a two-tailed Student's t-test. Statistical
differences (P-values) in multiple groups were obtained by using a
two-way ANOVA analysis. All experiments were carried out in
triplicate. A two-tailed P<0.05 was considered to indicate
statistical significance. All statistical analyses were undertaken
using GraphPad Prism 5.0 (GraphPad Software, Inc., La Jolla, CA,
USA).
Results
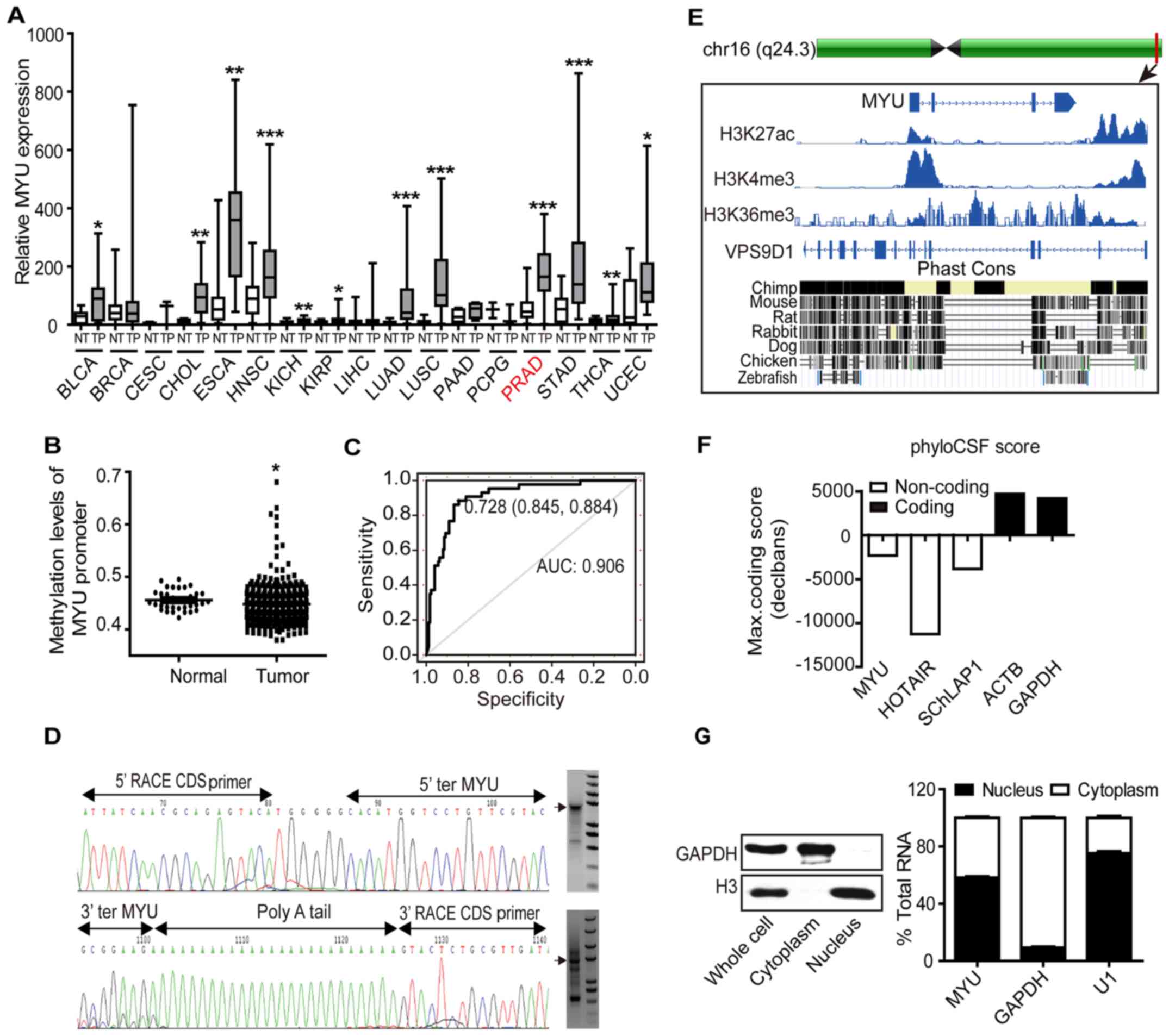
MYU is significantly upregulated in
PCa
In order to search for the differentially expressed
lncRNAs associated with PCa, we analyzed clinical patient data in
TCGA and matched expression profiles of lncRNAs from
Mitranscriptome. We noted that lncRNA MYU was significantly
upregulated in multiple cancers compared with adjacent normal
tissues, and focused on PCa (P<0.0001, Fig. 3A). To investigate the potential
cause of higher MYU expression in PCa, we examined the
methylation levels of the MYU promoter region, which is a
major cause of gene deregulation in tumorigenesis (23). Apparently, the methylation levels of
histone H3 of the MYU promoter region in PCa tissues was
significantly lower than those of adjacent normal tissues
(P=0.0163, Fig. 3B). Moreover,
receiver operating characteristic (ROC) curve was used to evaluate
the sensitivity and specificity of MYU expression in
predicting PCa tissues from normal tissues. MYU displayed
considerable predictive significance, with an area under the curve
(AUC) of 0.906 (Fig. 3C). In
summary, MYU is downregulated in PCa and may be a
potentially diagnostic indicator.
The molecular characterization of
Myu
To explore the function of MYU more
precisely, we performed 5′/3′-RACE to obtain the accurate
transcript of MYU, which is a 1,461 bp transcript with four
exons (Fig. 3D and Fig. 1). Published PCa chromatin
immunoprecipitation and sequencing (ChIP-seq) data confirmed that
the transcriptional start site (TSS) of MYU was marked by
H3-lysine-27-acetylation (H3K27ac) (24), H3-lysine-4-trimethylation (H3K4me3)
(25) and that its gene body
harbored H3-lysine-36-trimethylation (H3K36me3) (26), an epigenetic signature consistent
with lncRNAs. In addition, the UCSC Genome Browser revealed that
MYU sequences across the different species were extremely
unconserved (Fig. 3E). The PhyloCSF
analysis showed that MYU has no potential protein-coding
ability (Fig. 3F). Moreover,
subcellular localization of MYU showed about 58% MYU
in the nucleus in LNCaP cells (Fig.
3G). Taken together, these data revealed that MYU is an
active transcribed lncRNA, which exists both in the nucleus and
cytoplasm.
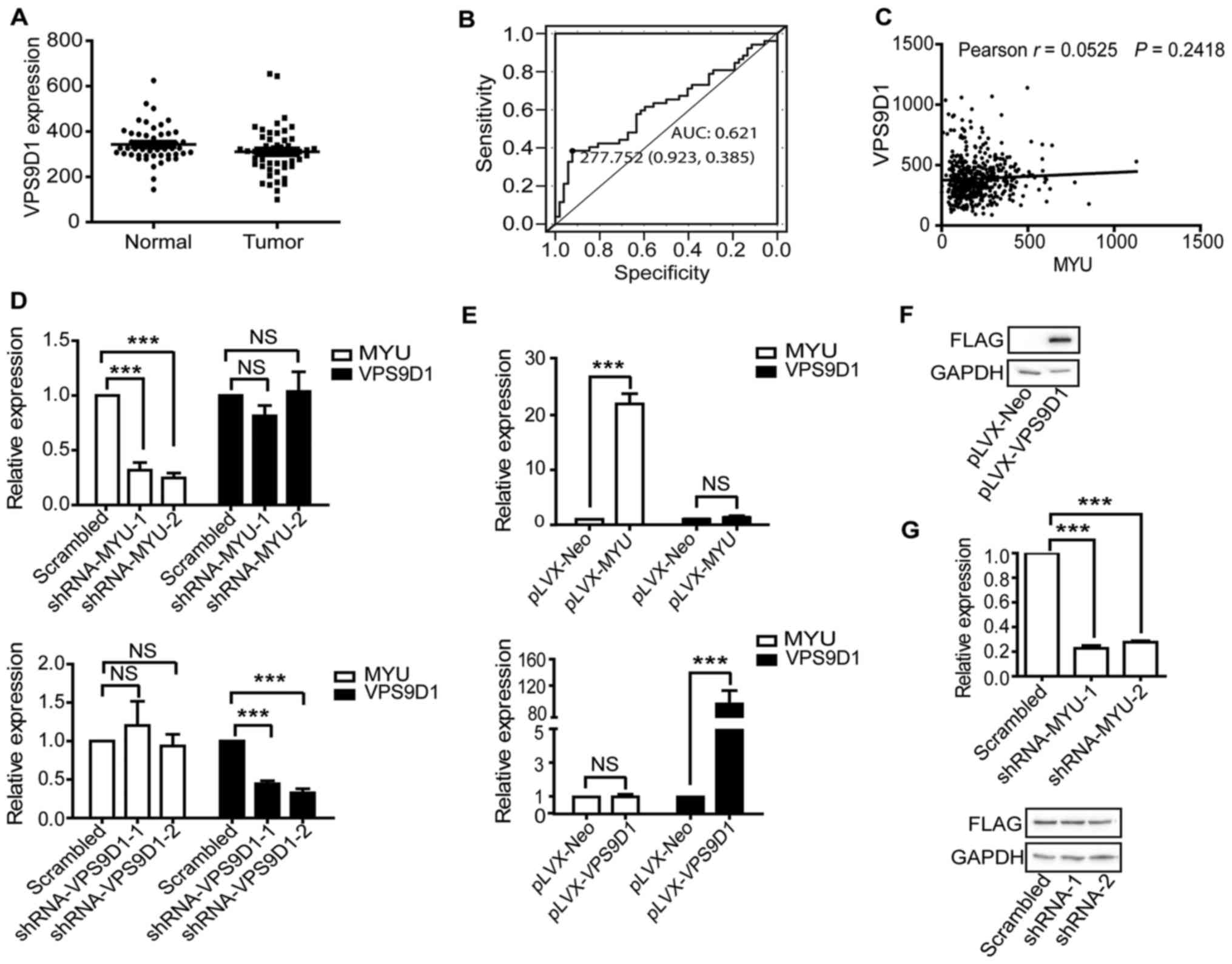
VPS9D1 is not regulated by MYU
We found that MYU has a corresponding
sense-cognate coding gene VPS9D1 from the UCSC Genome
Browser. Many studies have shown that antisense transcripts
regulate the expression of sense genes, particular for ncRNAs as
antisense transcripts (27,28). We investigated the potential
relationship between MYU and VPS9D1. There was no
difference in expression levels of VPS9D1 between PCa
tissues and adjacent normal tissues (Fig. 4A). The ROC analysis showed that the
AUC of VPS9D1 was 0.621 (Fig.
2B). Moreover, there was no statistically significant
correlation in RNA level between MYU and VPS9D1
(r=0.0525, P=0.2418) (Fig. 4C).
Furthermore, we examined the potential regulation between
MYU and VPS9D1 using qRT-PCR and western blotting
assays. The qRT-PCR results revealed that knockdown of either gene
did not affect the mRNA level of another gene (Fig. 4D), and the result of overexpression
was the same (Fig. 4E). In order to
determine whether MYU expression affected the protein level
of VPS9D1, we overexpressed VPS9D1 in DU145 cells (Fig. 4F). Then we knocked down MYU
in the VPS9D1-overexpressed cells, and observed the same protein
levels of VPS9D1 in either MYU-knockdown or control cells
(Fig. 4G). These results suggest
that MYU did not regulate VPS9D1 at either the mRNA
or protein level. Thus, we focused on studying lncRNA
MYU.
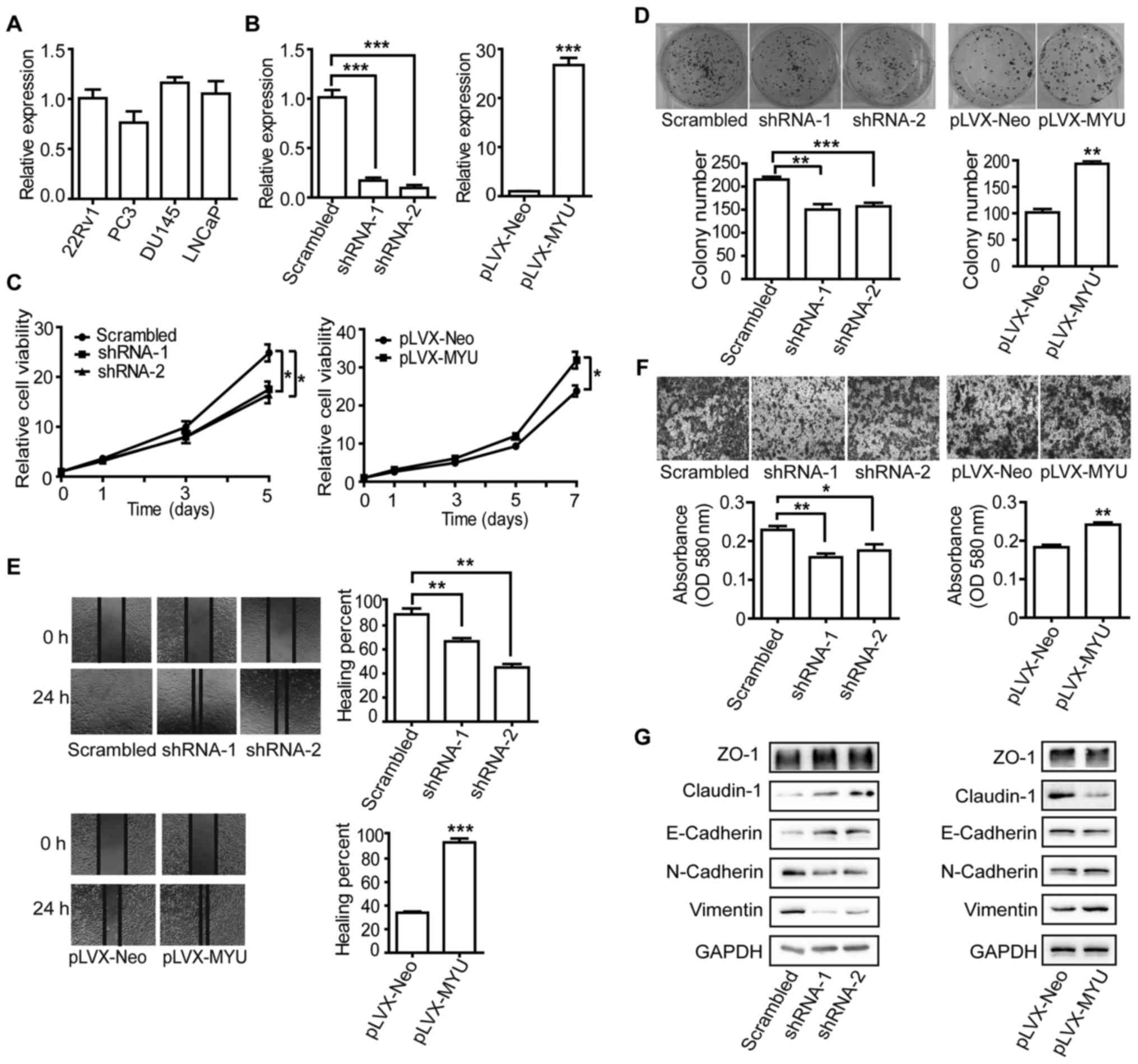
MYU promotes PCa cell proliferation
and migration in vitro
To evaluate the biological effects of MYU on
PCa, we firstly performed qRT-PCR to examine MYU expression
level in four PCa cell lines (Fig.
5A). To clarify the function of MYU, we established cell
lines with stably overexpressed MYU and MYU-specific
shRNAs (Fig. 5B). CTG assays
indicated that PCa cell proliferation was repressed by MYU
shRNAs compared to that of cells stably expressing the scrambled
shRNA, whereas enhancement of cell viability occurred after
overexpression of MYU (Fig.
5C). In colony formation assay, MYU-upregulated PC3
cells exhibited increased colony growth, while knockdown of
MYU in DU145 cells reduced colony growth compared to the
control cells (Fig. 5D).
Additionally, wound healing (Fig.
5E), Transwell chamber (Fig.
5F) and western blotting (Fig.
5G) assays indicated that knockdown of MYU reduced the
DU145 cell migration rate and the protein level of N-cadherin and
vimentin, but increased ZO-1, claudin-1 and E-cadherin. In
contrast, overexpression of MYU in the PC3 cells resulted in
a higher migration rate, accompanied by downregulated ZO-1,
claudin-1 and E-cadherin and upregulated N-cadherin and vimentin.
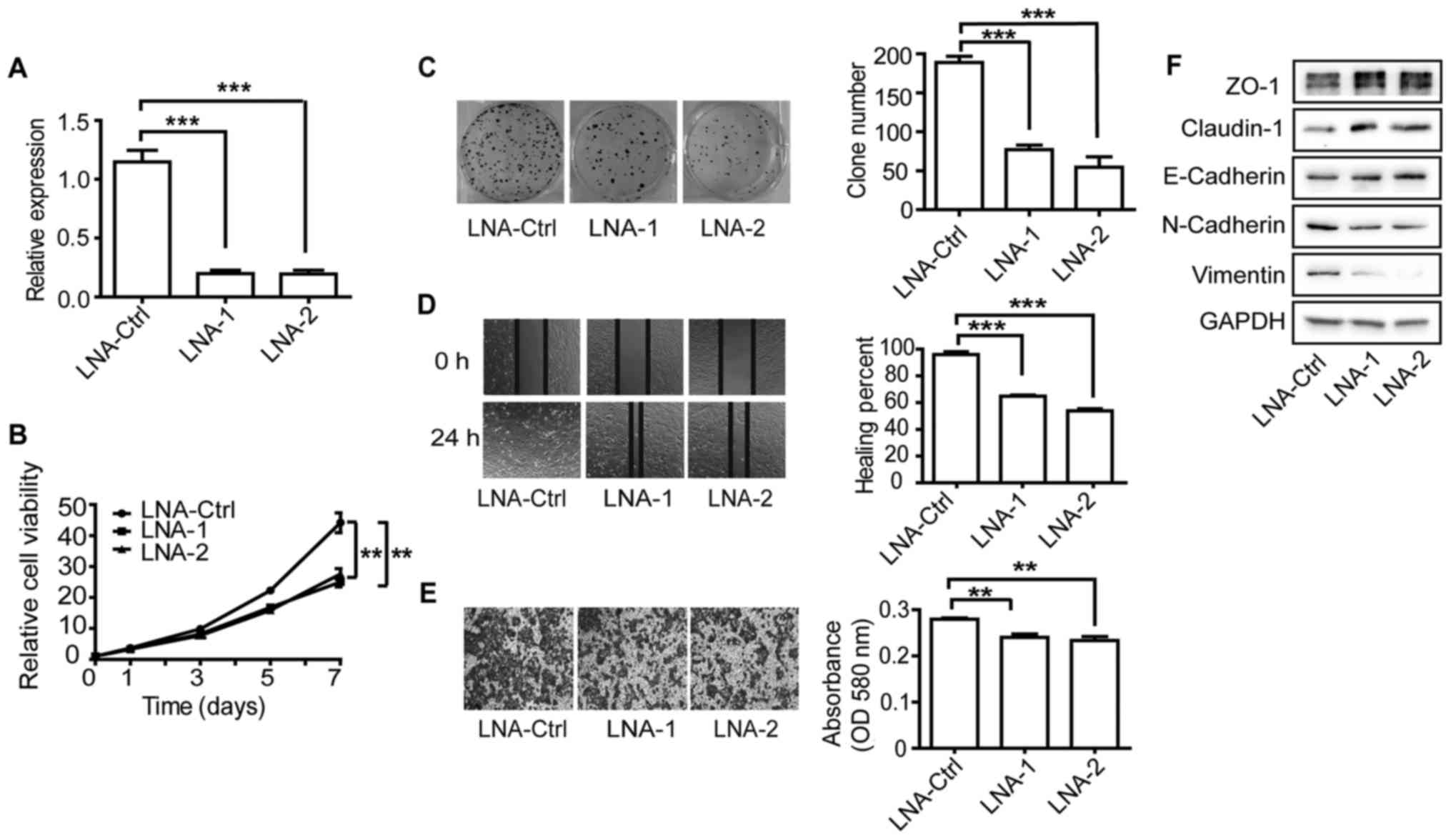
To further confirm these results, we also used locked nucleic acids
(LNAs) to knock down the expression of MYU. Similar results
for cell viability, clonogenicity and motility were obtained in the
DU145 cells (Fig. 6A-F). These
results suggest that MYU may act as a non-coding oncogene in
PCa cells.
MYU upregulates c-Myc expression by
competitively binding miR-184
We aimed to ascertain the potential underlying
mechanism of MYU, which may mediate PCa progression. lncRNAs
are known to work as sponges to recruit miRNAs, resulting in the
activation of miRNA target genes (13,29).
We analyzed the correlation of the expression level between
MYU and mature miRNAs and found that MYU showed a
significantly negative correlation with 15 miRNAs (r< −0.3,
P<0.0001, Table V). We then
tested if three miRNAs including miR-187-3p, miR-143-3p and miR-184
(r< −0.35, P<0.0001) can bind to MYU. The dual
luciferase reporter assays revealed that only miR-184 downregulated
the luciferase activity of lncRNA MYU. Then, we searched the
reported target coding genes of miR-184 in PubMed. Noteworthy,
miR-184 has been reported as a tumor-suppressor gene in multiple
cancers (30,31), and downregulated c-Myc expression
(32,33). It has been reported that MYU
is a direct target of c-Myc (14).
We hypothesized that MYU may promote the proliferation of
PCa as a ceRNA to sponge miR-184, thus upregulating c-Myc.
| Table V.Fifteen miRNAs show significantly
negative correlation with MYU in TCGA PCa tissues (Spearman
r<-0.3, P<0.0001). |
Table V.
Fifteen miRNAs show significantly
negative correlation with MYU in TCGA PCa tissues (Spearman
r<-0.3, P<0.0001).
| miRNA | Spearman r | P-value |
|---|
| hsa.miR.187.3p | −0.420633196 | 1.14E-10 |
| hsa.miR.221.3p | −0.40755072 | 4.74E-10 |
| hsa.miR.143.3p | −0.395310718 | 1.71E-09 |
| hsa.miR.873.5p | −0.369065673 | 2.25E-08 |
| hsa.miR.187.5p | −0.365681346 | 3.09E-08 |
| hsa.miR.184 | −0.3619539 | 4.36E-08 |
| hsa.miR.378c | −0.359757081 | 5.33E-08 |
| hsa.miR.222.3p | −0.350223717 | 1.25E-07 |
| hsa.miR.222.5p | −0.349057455 | 1.39E-07 |
| hsa.miR.23c | −0.343942199 | 2.16E-07 |
| hsa.miR.27b.3p | −0.334526294 | 4.81E-07 |
| hsa.miR.133b | −0.332952086 | 5.48E-07 |
| hsa.miR.767.5p | −0.327593572 | 8.51E-07 |
| hsa.miR.452.5p | −0.31499159 | 2.32E-06 |
| hsa.miR.152.3p | −0.306935385 | 4.30E-06 |
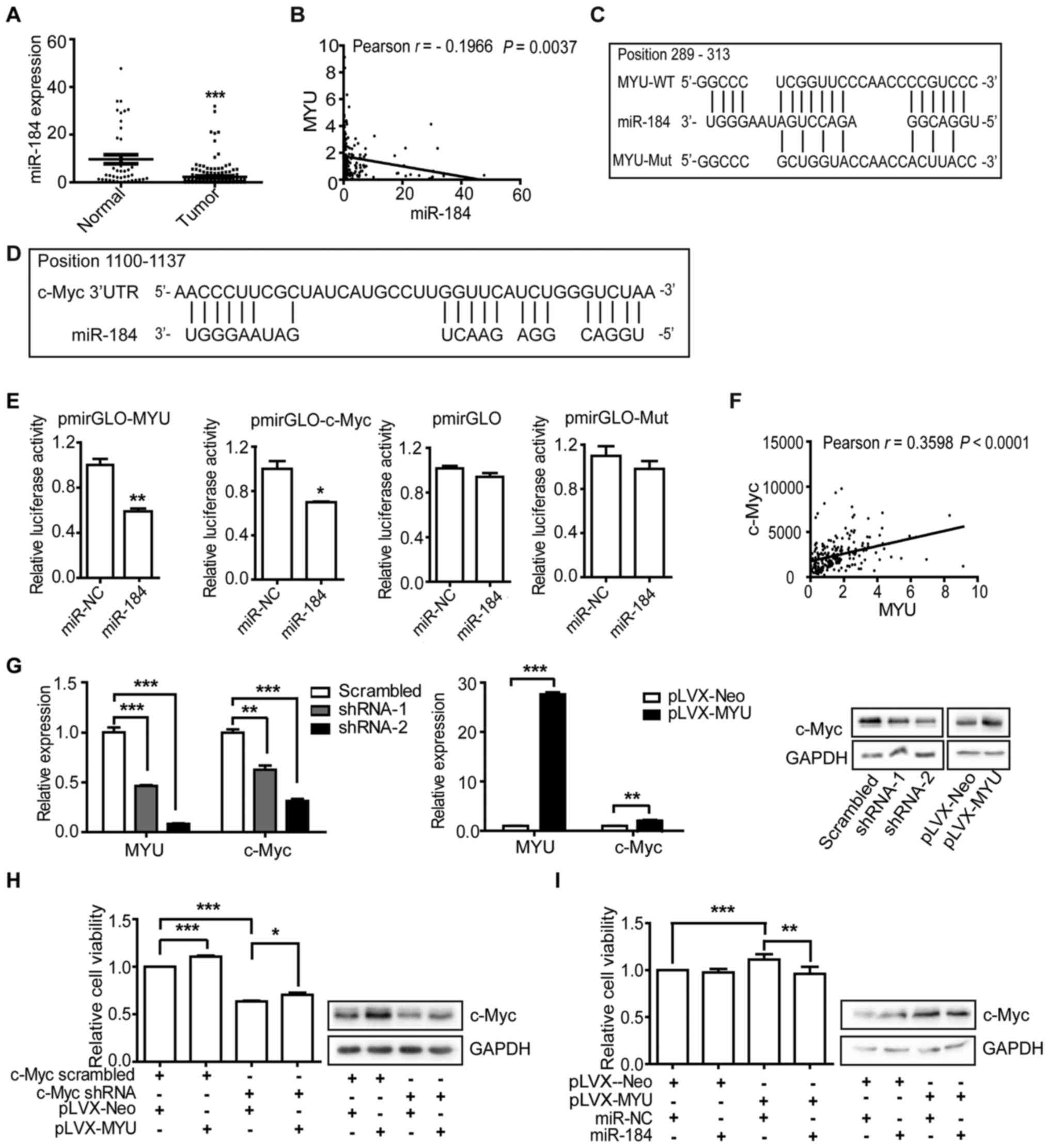
TCGA data revealed that miR-184 was significantly
downregulated in PCa tissues (Fig.
7A) and negatively correlated with MYU (r=−0.1966,
P=0.0037) (Fig. 7B). RNAhybrid
(34) was used to predict the
binding sites of miR-184 /MYU and miR-184/3′ UTR of c-Myc
(Fig. 7C and D). We constructed
luciferase reporters containing full-length MYU or mutated
miR-184 binding sites or 3′ UTR of c-Myc. We found that
overexpression of miR-184 mimic (Table
IV) reduced luciferase activity of MYU and c-Myc 3′UTR,
but not of the empty vector or the mutated reporter vector
(Fig. 7E). Next, we explored the
effect of MYU on the expression of c-Myc. The expression
level of c-Myc was positively correlated with MYU (r=0.3598,
P<0.0001) in PCa tissues (Fig.
7F). Furthermore, we detected that MYU knockdown in
DU145 cells decreased the mRNA and protein level of c-Myc.
Inversely, MYU overexpression in PC3 cells increased c-Myc
expression (Fig. 7G). In addition,
c-Myc knockdown suppressed the proliferation of PC3 cells and
inhibited the effect of MYU overexpression on promoting cell
proliferation. Additionally, the protein level of c-Myc was
consistent with the cell proliferation (Fig. 7H), indicating that c-Myc can partly
influence MYU. To further clarify the relationship of
MYU, miR-184 and c-Myc, we transfected miR-184 mimics into
MYU-overexpressing PC3 cells. We found that overexpression
of miR-184 significantly abrogated the effect of MYU
overexpression on promoting cell proliferation and suppressed the
protein level of c-Myc (Fig. 7I).
On the base of the above findings, MYU likely regulates
c-Myc by functioning as ceRNA to sponge miR-184.
| Table IV.Sequences of miR-184 mimics and
negative control. |
Table IV.
Sequences of miR-184 mimics and
negative control.
| Mimics | Sequence |
|---|
| miR-184 mimics | 5′
UGGACGGAGAACUGAUAAGGGU 3′ |
| Negative
control | 5′
UUCUCCGAACGUGUCACGUTT 3′ |
Intercellular transfer of MYU by
exosome functions in PCa cells
Recent studies suggest that exosomes, which contain
proteins, mRNAs, miRNAs and lncRNAs, may participate in the
biological effects in cancer cells (35,36).
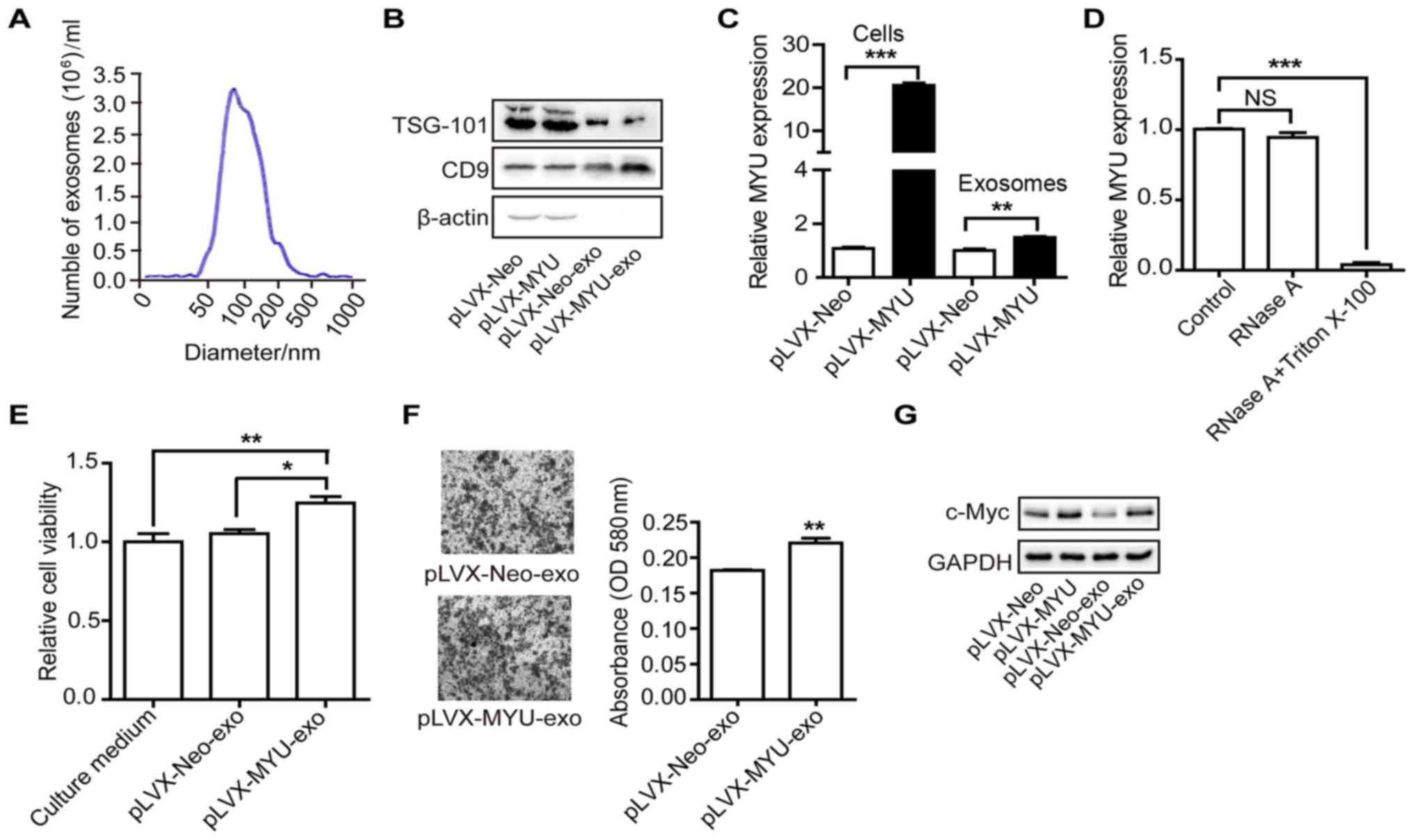
To examine whether MYU is present in exosomes, we isolated
exosomes from the culture medium of MYU-overexpressing or
control PC3 cells. Nanosight particle tracking analysis
demonstrated the size and number of exosomes (Fig. 8A). Western blot analysis further
confirmed their identity by the exosomal markers TSG-101 and CD9
(Fig. 8B). We detected the
expression level of MYU by qRT-PCR in the exosomes of both
PC3 cell lines. There was a significant upregulation of MYU
in the exosomes isolated from the MYU-overexpressing cells,
which was consistent with the cellular expression (Fig. 8C). We next investigated the existing
pattern of MYU from exosomes. The levels of MYU were
unchanged upon RNase A treatment but significantly decreased after
being treated with RNase A and Triton X-100 (Fig. 8D), indicating that MYU was
mainly wrapped by the membrane of exosomes. These findings suggest
that MYU can be delivered into the extracellular milieu by
exosomes. In order to explore whether exosome-contained MYU
can function in neighboring cells, we purified exosomes and then
fed PC3 cells. The CTG (Fig. 8E)
and Transwell chamber assays (Fig.
8F) demonstrated that exosomes from the
MYU-overexpressing PC3 cells significantly increased
proliferation and migration ability of recipient cells, compared to
those from control cells. Correspondingly, the c-Myc protein level
was significantly increased in the recipient PC3 cells fed with
exosomes from the MYU-overexpressing cells (Fig. 8G). These findings suggest that
MYU can be delivered into the extracellular milieu by
exosomes, and then function in other PCa cells.
Discussion
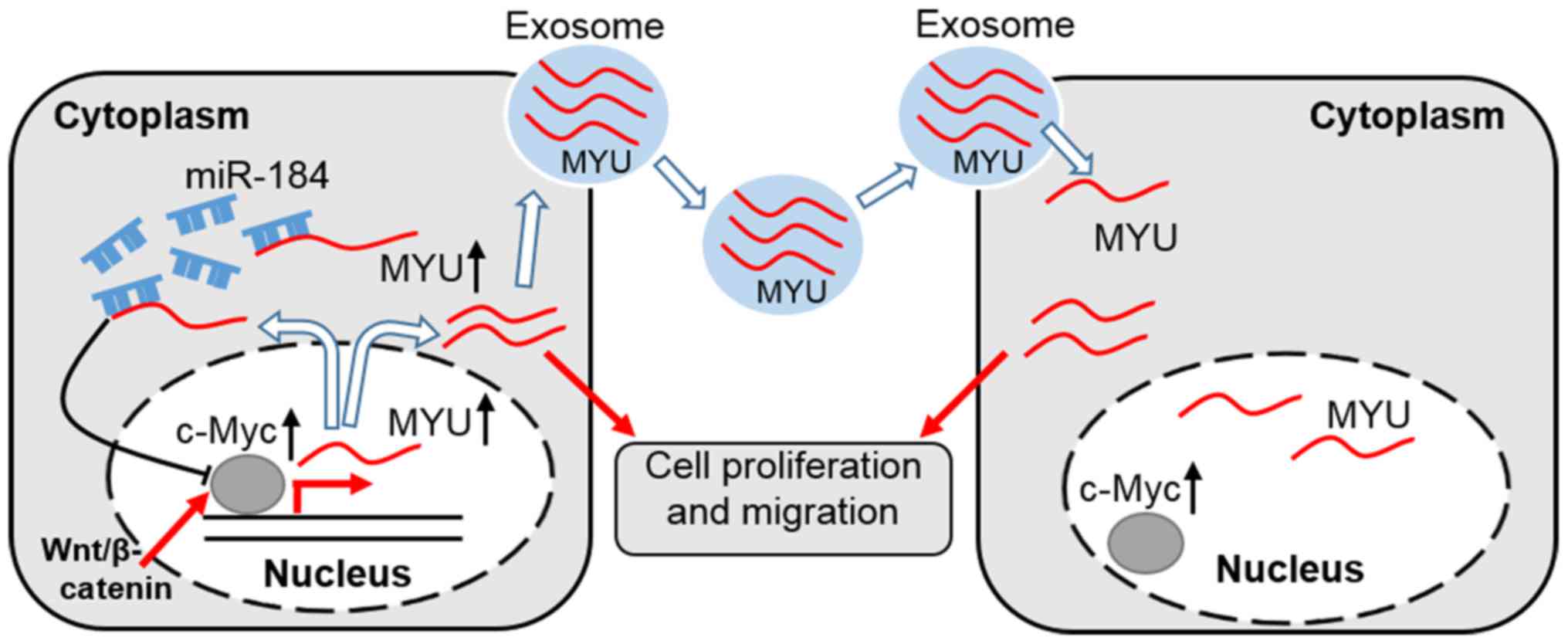
In the present study, we found that MYU
promotes PCa cell proliferation, migration and the exosome is the
carrier for extracellular MYU. Mechanistically, MYU
likely promotes proliferation of PCa cells by competitively binding
miR-184, increasing the expression of c-Myc (Fig. 9).
MYU is expressed in various tumors with a
broad spectrum. MYU has been reported to regulate CRC based
on its annotated sequence (14,15).
However, its future application may be limited due to the lack of
an accurate transcript. Generally, lncRNAs play oncogenic or
suppressive roles in the biological processes of cancer through
various mechanisms (37,38). A recent study indicates that the
expression of MYU in CRC is higher than that noted in normal
tissues and shortens overall survival time (15). However, Chen et al found that
the deregulation of MYU can be used as an indicator for
predicting poor overall and disease-free survival of GC patients
(16). These results showed that
MYU plays different roles in CRC and GC. In this study, we
found that MYU likely promotes PCa cell proliferation and
migration. Therefore, MYU may function as an oncogene in
PCa.
The exosome is a discoid vesicle that is 40–150 nm
in diameter and is released into the extracellular milieu upon
fusion with the cellular membrane (39). Exosomes participate in intercellular
communication by transmitting intracellular cargoes, such as
proteins and nucleic acids (40). A
recent study reported that lncRNAs are present in exosomes and
account for 20.19% of exosomal RNA extracted from the plasma of
castration-resistant PCa patients (41). In this study, we detected that
lncRNA MYU exists in the extracellular milieu of PC3 cells,
which can be wrapped by exosomes, and promotes recipient cell
proliferation and migration. Overall, MYU functions in the
extracellular milieu of PCa cells by exosome transportation.
LncRNAs act as ceRNAs to sponge miRNAs leading to
release of downstream target genes, which was a canonical model to
study the molecular mechanism of lncRNAs (12,13).
In this study, we found that MYU likely upregulates c-Myc
expression by sponging miR-184 (Fig.
9). A recent report demonstrated that MYU is a direct
target lncRNA of the Wnt/c-Myc pathway and participates in the
tumorigenicity of colon cancer cells (14). These results indicate that
MYU and c-Myc may form a feedback loop to regulate each
other. In conclusion, we demonstrated that MYU displays
oncogenic activity by regulating c-Myc expression in PCa,
suggesting that MYU maybe as a potential diagnostic
predictor and therapeutic target for PCa.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the National
Natural Science Foundation of China (81773023 and 81472827), the
National Key R&D Program of China (2016YFC1302100),
Hundred-Talent Program (Y521031102) and Frontier Research Program
(QYZDB-SSW-SMC038) of the Chinese Academy of Sciences to SG.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
SG and JW conceived and designed the study. JW
performed the experiments. JW and SG wrote the paper. All authors
read and approved the manuscript and agree to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cheng W, Zhang Z and Wang J: Long
noncoding RNAs: New players in prostate cancer. Cancer Lett.
339:8–14. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hawksworth D, Ravindranath L, Chen Y,
Furusato B, Sesterhenn IA, McLeod DG, Srivastava S and Petrovics G:
Overexpression of C-MYC oncogene in prostate cancer predicts
biochemical recurrence. Prostate Cancer Prostatic Dis. 13:311–315.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Prensner JR, Chen W, Han S, Iyer MK, Cao
Q, Kothari V, Evans JR, Knudsen KE, Paulsen MT, Ljungman M, et al:
The long non-coding RNA PCAT-1 promotes prostate cancer cell
proliferation through cMyc. Neoplasia. 16:900–908. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chang C, Liu J, He W, Qu M, Huang X, Deng
Y, Shen L, Zhao X, Guo H, Jiang J, et al: A regulatory circuit
HP1gamma/miR-451a/c-Myc promotes prostate cancer progression.
Oncogene. 37:415–426. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang C, Wang L, Wang X and Gao S: The
mystery of ‘junk’ DNA. Chin Sci Bull. 61:3079–3084. 2016.
|
|
7
|
Adams BD, Anastasiadou E, Esteller M, He L
and Slack FJ: The Inescapable Influence of Noncoding RNAs in
Cancer. Cancer Res. 75:5206–5210. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schwarzer A, Emmrich S, Schmidt F, Beck D,
Ng M, Reimer C, Adams FF, Grasedieck S, Witte D, Käbler S, et al:
The non-coding RNA landscape of human hematopoiesis and leukemia.
Nat Commun. 8:218–234. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Merola R, Tomao L, Antenucci A, Sperduti
I, Sentinelli S, Masi S, Mandoj C, Orlandi G, Papalia R, et al:
PCA3 in prostate cancer and tumor aggressiveness detection on 407
high-risk patients: a National Cancer Institute experience. J Exp
Clin Cancer Res. 34:15–20. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Prensner JR, Iyer MK, Sahu A, Asangani IA,
Cao Q, Patel L, Vergara IA, Davicioni E, Erho N, Ghadessi M, et al:
The long noncoding RNA SChLAP1 promotes aggressive prostate cancer
and antagonizes the SWI/SNF complex. Nat Genet. 45:1392–1398. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Parolia A, Crea F, Xue H, Wang Y, Mo F,
Ramnarine VR, Liu HH, Lin D, Saidy NR, Clermont PL, et al: The long
non-coding RNA PCGEM1 is regulated by androgen receptor activity in
vivo. Mol Cancer. 14:46–52. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu XH, Sun M, Nie FQ, Ge YB, Zhang EB,
Yin DD, Kong R, Xia R, Lu KH, Li JH, et al: Lnc RNA HOTAIR
functions as a competing endogenous RNA to regulate HER2 expression
by sponging miR-331-3p in gastric cancer. Mol Cancer. 13:92–105.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fang C, Qiu S, Sun F, Li W, Wang Z, Yue B,
Wu X and Yan D: Long non-coding RNA HNF1A-AS1 mediated repression
of miR-34a/SIRT1/p53 feedback loop promotes the metastatic
progression of colon cancer by functioning as a competing
endogenous RNA. Cancer Lett. 410:50–62. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kawasaki Y, Komiya M, Matsumura K, Negishi
L, Suda S, Okuno M, Yokota N, Osada T, Nagashima T, Hiyoshi M, et
al: MYU, a target lncRNA for Wnt/c-Myc signaling, mediates
induction of CDK6 to promote cell cycle progression. Cell Reports.
16:2554–2564. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang L, Xu L, Wang Q, Wang M and An G:
Dysregulation of long non-coding RNA profiles in human colorectal
cancer and its association with overall survival. Oncol Lett.
12:4068–4074. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen M, Wu X, Ma W, Zhou Q, Wang X, Zhang
R, Wang J and Yang X: Decreased expression of lncRNA VPS9D1-AS1 in
gastric cancer and its clinical significance. Cancer Biomark.
21:23–28. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Weinstein JN, Collisson EA, Mills GB, Shaw
KR, Ozenberger BA, Ellrott K, Shmulevich I, Sander C and Stuart JM:
Cancer Genome Atlas Research Network: The Cancer Genome Atlas
Pan-Cancer analysis project. Nat Genet. 45:1113–1120. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Iyer MK, Niknafs YS, Malik R, Singhal U,
Sahu A, Hosono Y, Barrette TR, Prensner JR, Evans JR, Zhao S, et
al: The landscape of long noncoding RNAs in the human
transcriptome. Nat Genet. 47:199–208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cheng PF, Dummer R and Levesque MP: Data
mining The Cancer Genome Atlas in the era of precision cancer
medicine. Swiss Med Wkly. 145:w141832015.PubMed/NCBI
|
|
20
|
Zhang Y, Liu T, Meyer CA, Eeckhoute J,
Johnson DS, Bernstein BE, Nusbaum C, Myers RM, Brown M, Li W, et
al: Model-based analysis of ChIP-Seq (MACS). Genome Biol.
9:R1372008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Feng J, Liu T and Zhang Y: Using MACS to
identify peaks from ChIP-Seq data. Curr Protoc Bioinformatics
Chapter. 2:Unit2.14. 2011.
|
|
22
|
Kent WJ, Sugnet CW, Furey TS, Roskin KM,
Pringle TH, Zahler AM and Haussler D: The human genome browser at
UCSC. Genome Res. 12:996–1006. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liang G and Weisenberger DJ: DNA
methylation aberrancies as a guide for surveillance and treatment
of human cancers. Epigenetics. 12:416–432. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Creyghton MP, Cheng AW, Welstead GG,
Kooistra T, Carey BW, Steine EJ, Hanna J, Lodato MA, Frampton GM,
Sharp PA, et al: Histone H3K27ac separates active from poised
enhancers and predicts developmental state. Proc Natl Acad Sci USA.
107:21931–21936. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu X, Wang C, Liu W, Li J, Li C, Kou X,
Chen J, Zhao Y, Gao H, Wang H, et al: Distinct features of H3K4me3
and H3K27me3 chromatin domains in pre-implantation embryos. Nature.
537:558–562. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Pokholok DK, Harbison CT, Levine S, Cole
M, Hannett NM, Lee TI, Bell GW, Walker K, Rolfe PA, Herbolsheimer
E, et al: Genome-wide map of nucleosome acetylation and methylation
in yeast. Cell. 122:517–527. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang B, Song JH, Cheng Y, Abraham JM,
Ibrahim S, Sun Z, Ke X and Meltzer SJ: Long non-coding antisense
RNA KRT7-AS is activated in gastric cancers and supports cancer
cell progression by increasing KRT7 expression. Oncogene.
35:4927–4936. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun J, Wang X, Fu C, Wang X, Zou J, Hua H
and Bi Z: Long noncoding RNA FGFR3-AS1 promotes osteosarcoma growth
through regulating its natural antisense transcript FGFR3. Mol Biol
Rep. 43:427–436. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yuan JH, Yang F, Wang F, Ma JZ, Guo YJ,
Tao QF, Liu F, Pan W, Wang TT, Zhou CC, et al: A long noncoding RNA
activated by TGF-β promotes the invasion-metastasis cascade in
hepatocellular carcinoma. Cancer Cell. 25:666–681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Emdad L, Janjic A, Alzubi MA, Hu B,
Santhekadur PK, Menezes ME, Shen XN, Das SK, Sarkar D and Fisher
PB: Suppression of miR-184 in malignant gliomas upregulates SND1
and promotes tumor aggressiveness. Neuro-oncol. 17:419–429. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cheng Z, Wang HZ, Li X, Wu Z, Han Y, Li Y,
Chen G, Xie X, Huang Y, Du Z, et al: MicroRNA-184 inhibits cell
proliferation and invasion, and specifically targets TNFAIP2 in
Glioma. J Exp Clin Cancer Res. 34:272015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang YB, Zhao XH, Li G, Zheng JH and Qiu
W: MicroRNA-184 inhibits proliferation and promotes apoptosis of
human colon cancer SW480 and HCT116 cells by downregulating C-MYC
and BCL-2. J Cell Biochem. 119:1702–1715. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhen Y, Liu Z, Yang H, Yu X, Wu Q, Hua S,
Long X, Jiang Q, Song Y, Cheng C, et al: Tumor suppressor PDCD4
modulates miR-184-mediated direct suppression of C-MYC and BCL2
blocking cell growth and survival in nasopharyngeal carcinoma. Cell
Death Dis. 4:e8722013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bhattacharjya S, Roy KS, Ganguly A, Sarkar
S, Panda CK, Bhattacharyya D, Bhattacharyya NP and Roychoudhury S:
Inhibition of nucleoporin member Nup214 expression by miR-133b
perturbs mitotic timing and leads to cell death. Mol Cancer.
14:42–56. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Qu L, Ding J, Chen C, Wu ZJ, Liu B, Gao Y,
Chen W, Liu F, Sun W, Li XF, et al: Exosome-transmitted lncARSR
promotes sunitinib resistance in renal cancer by acting as a
competing endogenous RNA. Cancer Cell. 29:653–668. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gao Y, Huang KH, Tang Y, Huang Y and Gao
S: Exosome: A rising star in the era of precision oncology. Chin
Sci Bull. 63:1–8. 2018.
|
|
37
|
He W, Zhong G, Jiang N, Wang B, Fan X,
Chen C, Chen X, Huang J and Lin T: Long noncoding RNA BLACAT2
promotes bladder cancer-associated lymphangiogenesis and lymphatic
metastasis. J Clin Invest. 128:861–875. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Marín-Béjar O, Mas AM, González J,
Martinez D, Athie A, Morales X, Galduroz M, Raimondi I, Grossi E,
Guo S, et al: The human lncRNA LINC-PINT inhibits tumor cell
invasion through a highly conserved sequence element. Genome Biol.
18:202–216. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Théry C: Exosomes: Secreted vesicles and
intercellular communications. F1000 Biol Rep. 3:15–22. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Melo SA, Sugimoto H, O'Connell JT, Kato N,
Villanueva A, Vidal A, Qiu L, Vitkin E, Perelman LT, Melo CA, et
al: Cancer exosomes perform cell-independent microRNA biogenesis
and promote tumorigenesis. Cancer Cell. 26:707–721. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Huang X, Yuan T, Liang M, Du M, Xia S,
Dittmar R, Wang D, See W, Costello BA, Quevedo F, et al: Exosomal
miR-1290 and miR-375 as prognostic markers in castration-resistant
prostate cancer. Eur Urol. 67:33–41. 2015. View Article : Google Scholar : PubMed/NCBI
|