Introduction
Multiple myeloma (MM) is a clonal B-cell malignancy
characterized by the proliferation of plasma cells (PCs) within the
bone marrow (BM). MM is the second most common hematological
malignancy, but it remains incurable, with a survival rate ranging
between a few months and over 10 years (1,2).
Although treatment strategies have evolved from traditional
chemotherapy and autologous hematopoietic stem cell transplantation
to novel targeted drug therapies, patient outcomes have not
improved significantly (3). The
5-year survival rate remains at only 30–40%, mainly due to the
development of drug resistance (4).
Therefore, identifying the mechanisms that regulate the key genes
responsible for the progression of MM and the underlying mechanisms
governing the malignant behavior of MM cells is important for
establishing treatments to improve patient prognosis. One topic of
interest is chromosomal abnormalities and other types of genetic or
epigenetic alterations that may contribute to microRNA (miRNA)
dysregulation in cancer (5–7).
miRNAs, a newly identified class of endogenous,
small, non-coding RNAs, regulate gene expression by inhibiting
protein translation or by inducing mRNA degradation (5). miRNAs are involved in various
biological processes, including cellular growth, development,
metabolism, apoptosis and development (8). Increasing evidence has demonstrated
that differential miRNA expression, including that of miRNA
(miR)-21, miR-155, miR-17-92 and miR-125a-5p, is involved in MM,
suggesting the importance of miRNAs in the progression of MM
(9–11). Deregulated miRNAs can act as
oncogenes or tumor suppressors, with the former being upregulated
and the latter being downregulated in cancer cells (12–14).
miR-125b is an important member of the miRNA family, which is
involved in various types of human, including prostate (15), colorectal cancer (16), leukemia (17), breast (18) and lung cancer (19). However, the biological role of
miR-125b in MM remains to be elucidated. Several studies have
suggested that the dysregulation of miRNAs initiates activation of
the Akt signaling pathway, which is involved in the progression of
breast, bladder and non-small cell lung cancer (20–23).
However, the biological role of miR-125b and its effects on the Akt
signaling pathway in the pathogenesis and progression of MM remain
to be fully elucidated. The aim of the present study was to
characterize the expression of miR-125b in MM and identify its
function in MM cell lines in vitro and in vivo, and
to determine the effect of miR-125b on the Akt signaling pathway.
Finally, screening for target genes of miR-125b was performed. It
was found that miR-125b promoted MM cell proliferation by targeting
tumor suppressor PH domain and leucine rich repeat protein
phosphatase 2 (PHLPP2) and that the downregulation of PHLPP2
rescued the effect of an miR-125b inhibitor in MM cells. These
findings provide novel insights into the function of miR-125b
during the development of MM and suggest that this miRNA is
involved in tumorigenesis.
Materials and methods
Study subjects and sample
collection
The specimens used in the present study were
obtained from 41 patients who were diagnosed according to the
National Comprehensive Cancer Network clinical practice guidelines
for MM at the First Affiliated Hospital of Nanchang University
(Nanchang, China) between October 2015 and July 2017 (24). Plasma samples from 41 patients
diagnosed with MM were assessed; six samples were used for
microarray tests, and 35 samples formed the validation set. Any
patients who received chemotherapy and/or biotherapy were excluded
from the study, and those with other types of malignant tumor were
also eliminated. To investigate the expression of miR-125b in
different genetic subtypes of MM, 35 samples were cytogenetically
classified using the FISH technique (Abbott Diagnostics, Berkshire,
UK). Cytogenetic abnormalities involving 13q deletions and
immunoglobulin heavy-chain gene rearrangements were investigated.
The patient details are shown in Tables
I and II. In addition, 20
plasma samples in the validation set from healthy individuals were
used as a control. Venous blood was collected in EDTA tubes (BD
Biosciences, Franklin Lakes, NJ, USA). The blood samples were
centrifuged at 800 × g for 10 min. The plasma was then transferred
into RNase/DNase-free tubes, followed by a 10-min high-speed
centrifugation at 16,000 × g to completely remove the cell debris.
The plasma was transferred to a fresh tube and stored at −80°C on
being frozen with liquid nitrogen. The present study was approved
by the Medical Research Ethics Committee of The First Affiliated
Hospital of Nanchang University, and written informed consent was
obtained from all the study subjects. The IRB number of human
sample study approved by the institutional committee is
2015(096).
 | Table I.Summary of the clinical details of 35
patients with multiple myeloma used for reverse
transcription-quantitative polymerase chain reaction analysis. |
Table I.
Summary of the clinical details of 35
patients with multiple myeloma used for reverse
transcription-quantitative polymerase chain reaction analysis.
| Clinical
details | n (%) |
|---|
| Sex |
|
|
Male | 23 (65.7) |
|
Female | 12 (34.3) |
| Age range
(years) | 35-75 |
| Mean age
(years) | 59 |
| Durie-Salmon
stage |
|
| I | 10 (28.57) |
| II | 6 (17.14) |
|
III | 19 (54.29) |
| Karyotype |
|
| t
(4:14) | 9 (25.71) |
| t
(11:14) | 5 (14.28) |
| t
(14:16) | 6 (17.14) |
| del
(13q) as a unique abnormality | 10 (28.57) |
| Normal
FISH | 5 (14.29) |
| Table II.Association between the expression of
miR-125b and the clinical pathological characteristics of patients
with multiple myeloma. |
Table II.
Association between the expression of
miR-125b and the clinical pathological characteristics of patients
with multiple myeloma.
| Characteristic | Cases (%) | miR-125b
expression | P-value |
|---|
| Age (years) |
|
≤50 | 16 | 1.66±0.03 | 0.241 |
|
>50 | 19 | 1.72±0.05 |
|
| Sex |
|
Male | 23 | 1.68±0.03 | 0.439 |
|
Female | 12 | 1.72±0.05 |
|
| Durie-Salmon
stage |
| I/II
phase | 16 | 1.62±0.04 | 0.009 |
| III
phase | 19 | 1.76±0.03 |
|
| Extramedullary
infiltration |
|
Yes | 8 | 1.82±0.05 | 0.012 |
| No | 27 | 1.66±0.03 |
|
| Karyotype |
| Normal
FISH | 5 | 1.65±0.12 | 0.455 |
|
Abnormal FISH | 30 | 1.72±0.03 |
|
miRNA array
Total RNA was extracted from six patients with MM
and six healthy controls using the phenol-chloroform method
(TRIzol; Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA
USA). The quality of the RNA was assessed by capillary
electrophoresis (Agilent Technologies Inc., Santa Clara, CA, USA).
Libraries for small RNA sequencing were prepared using the NEBNext
Multiplex Small RNA Library Prep Set for Illumina (New England
BioLabs, Inc., Ipswich, MA, USA) according to the manufacturer's
protocol. The libraries were quantified using the Agilent
Bioanalyzer 2100 system with DNA high-sensitivity chips. The raw
sequence files were subjected to quality control analysis with the
Fast QC quality control tool. To avoid low-quality data, adaptors
were removed by Cutadapt (version 1.2.1; DOI:10.14806/ej.17.1.200),
and lower-quality sequences were trimmed. The clean reads were
screened at a length of 21–22 nt as miRNA and were located to the
reference sequence with Bowtie software (version 2; CGE Risk
Management Solutions B.V., Leidschendam, The Netherlands). The
functions of novel miRNAs were analyzed with miRDeep2 software
(version 2.0.0.8; https:www.mdc-berlin.de/content/mirdeep2-documentation).
The differential expression sequence was used to calculate
differential expression levels and to evaluate the statistical
significance of detected alterations between the control and case
samples.
RNA isolation from human plasma and
reverse transcription- quantitative polymerase chain reaction
(RT-qPCR) analysis
Total RNA was extracted from human plasma using the
mirVana PARIS RNA Isolation kit (Ambion; Thermo Fisher Scientific,
Inc.) following the manufacture's protocol for liquid samples. The
concentration and purity of the extracted RNA were measured at 260
and 280 nm optical densities (ODs). cDNA was synthesized from the
RNA for RT-qPCR analysis using gene-specific primers (Applied
Biosystems; Thermo Fisher Scientific, Inc.) with the M-MLV Reverse
Transcriptase kit (GeneCopoeia, Inc., Rockville, MD, USA) according
to the manufacturer's protocol. To determine the expression levels
of miR-125b, qPCR analysis was performed with SYBR Green (Takara
Bio, Inc., Osaka, Japan). β-actin and U6 were used as internal
controls. The PCR conditions were as follows: 40 cycles of 95°C for
5 min, 95°C for 45 sec, 55°C for 15 sec, and 72°C for 50 sec, in
which each reaction (25 µl) contained cDNA (100 ng), primers (10
µM) and 10X Ex Taq E buffer (25 µl). The samples were analyzed in
triplicate, and gene expression was quantified by normalizing
target gene expression to that of the internal control using the
2−ΔΔCq formula (25).
The primer sequences used were as follows: miR-125b, forward
5′-TGCGCTAAAGTGCTTATAGTGC-3′ and reverse
5′-CCAGTGCAGGGTCCGAGGTATT-3′; PHLPP2, forward
5′-CCAATGAGCAAGGACAGGAT-3′ and reverse 5′-GGTCCTCTGGTTCCATCTGA-3′;
β-actin, forward 5′-AGCGAGCATCCCCCAAAGTT-3′ and reverse
5′-GGGCACGAAGGCTCATCATT-3′; and U6, forward 5′-CTCGCTTCGGCAGCACA-3′
and reverse 5′-AACGCTTCACGAATTTGCG-3′. The samples were analyzed in
triplicate, and gene expression was quantified by normalizing
target gene expression to that of the internal control using the
2−ΔΔCq formula.
Cell lines and transfection
Human MM cell lines (MM.1S, U266, and RPMI-8226) and
normal plasma cells (nPCs) were cultured in RPMI-1640 (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 10% fetal bovine
serum (FBS; HyClone Laboratories, Logan, UT, USA) and 1%
penicillin/streptomycin at 37°C in a 5% CO2 atmosphere.
When the cells reached 80% confluency, cells at the logarithmic
growth phase were collected. The cell lines were obtained from
American Type Culture Collection (Manassas, VA, USA). The cells
were assigned to the following groups: Blank group (no
transfection), negative control (NC) group (transfected with the
miR-125b NC sequence), miR-125b mimic (transfected with miR-125b
mimic), and miR-125b inhibitor (transfected with miR-125b
inhibitor). The cells were also transfected with small interfering
(si)RNA against PHLPP2 (siPHLPP2)
(5′-CCGGAATTCCAATCCCTAAATTTCCCTGGGA-3′). All sequences were
purchased from Shanghai GenePharma Co, Ltd. (Shanghai, China).
Transfection of plasmids was performed using Lipofectamine 2000
reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according to
the manufacturer's protocol.
Cell proliferation analysis
For the cell proliferation assays, the cells were
plated in individual wells of a 96-well plate (1,500 cells/well)
and were examined 48 h following transfection using a Cell Counting
Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc., Shanghai,
China). The OD values were determined at a wavelength of 450 nm
using a microplate reader. For the survival rate assay, the number
of viable cells was counted using Trypan blue dye (Beijing Solarbio
Science & Technology Co. Ltd., Beijing, China) and a Countstar
Cell Counter (Inno-Alliance Biotech, Inc. Wilmington, DE, USA)
according to the manufacturer's protocol. All assays were performed
in triplicate.
Migration and invasion assays
For the migration experiment at 72 h
post-transfection, the cells were collected and resuspended in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc.), following
which they were inoculated into the Transwell upper chamber and
placed in an incubator at 37°C with 5% CO2 for 48 h. The
cells that failed to penetrate the upper chamber were removed, and
the membrane was fixed in 95% ethanol for 15–20 min and then
immersed in water. The membrane was stained with crystal violet for
10 min and re-immersed in water, followed by observation and image
capture under a high-magnification laser confocal microscope (ZEISS
LSM888; Carl Zeiss, Jena, Germany). The number of cells on the back
of the membrane was then calculated. Five high-power fields were
randomly selected, and the number of cells penetrating through the
polycarbonate membrane was used to evaluate the cell migration
ability. For the invasion experiment, Matrigel matrix was dissolved
at 4°C overnight and diluted 1:3 with serum-free DMEM. A total
volume of 30 µl of Matrigel was added to the Transwell upper
chamber until it covered the bottom of the upper chamber. The cell
suspension was added to the upper chamber, and 0.5 ml of DMEM
(Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China) containing 10% FBS was added to the lower chamber of the
24-well plate. The number of cells penetrating through the Matrigel
was determined to assess the cell invasion ability.
Apoptosis analysis
The cells in each group were collected at 24, 48 and
72 h post-transfection, and cold phosphate-buffered saline (PBS)
was used to wash the cells three times. The cells were resuspended
with 500 µl of precooled binding buffer at a concentration of
5×106 cells/ml. The cell suspension (100 µl) was added
to flow cytometry tubes, and 5 µl of Annexin V-fluorescein
isothiocyanate (Beyotime Institute of Biotechnology, Haimen, China)
was added. Following mixing, the samples were incubated at room
temperature in the dark for 15 min, and 5 min prior to
measurements, 5 µl of 10 mg/l propidium iodide (PI) dye (Beyotime
Institute of Biotechnology) was added. The samples were immediately
analyzed with a FACSort system without washing or fixation. Each
assay was repeated three times.
Luciferase reporter analysis
The pGL3-PHLPP2-3′ untranslated region
(3′UTR)-WT/MUT vector (Guangzhou Ribobio, Co., Ltd., Guangzhou,
China) was cotransfected with the control plasmid or
miR-125b-expressing plasmid into 293T cells (Guangzhou Ribobio,
Co., Ltd.) using Lipofectamine 2000 (Thermo Fisher Scientific,
Inc.). Firefly and Renilla luciferase activities were
measured consecutively 24 h following transfection using the
Dual-Luciferase assay kit.
Western blot analysis
Whole-cell protein extracts were obtained from the
MM cell lines. The cell lysates were loaded and separated via
polyacrylamide gel electrophoresis. The proteins were transferred
onto membranes using a Trans-Blot Turbo Transfer Starter system
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) for 7 min.
Following protein transfer, the membranes were probed with primary
antibodies. Cells in the logarithmic growth phase at 80% confluency
were collected, and radioimmunoprecipitation assay lysis buffer
containing protease inhibitors (Beyotime Institute of
Biotechnology) was used for conventional extraction of total
cellular protein. A bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology) was used for protein quantification.
Total proteins (100 µg) were separated by 10% SDS gel
electrophoresis under denaturing and non-reducing conditions and
then were transferred onto a nitrocellulose membrane. Following the
addition of 5% skim milk, the membranes were agitated in a sealed
container for 1 h. The blotted membranes were incubated with the
following antibodies: Anti-Akt (1:500; cat. no. sc-5298),
anti-PHLPP2 (1:300; cat. no. sc-71973) and anti-phosphorylated
(p)-Akt (1:500; cat. no. sc-7985-R), all of which were obtained
from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA), and then
probed with a goat anti-rabbit secondary antibody (1:3,000; cat.
no. sc-2030; Santa Cruz Biotechnology, Santa Cruz, CA, USA),
β-actin (1:200; cat. no. sc-58673; Abcam, Cambridge, MA, USA) was
used as a loading control. The immunoblots were visualized using
the ImageQuant LAS 4000 digital imaging system (GE Healthcare,
Chicago, IL, USA).
Tumor xenograft model and
immunohistochemistry
Briefly, 5×106 U266 cells were
subcutaneously injected into the left upper flank region of male
nude mice BALB/c (3–4 weeks of age). Male BALA/C nude mice (n=20)
were purchased from the Beijing Hua Fukang Bioscience Company
(Beijing, China) and housed under specific pathogen-free
conditions. The housing condition were the following: Light-dark
cycle was setting as 10 h of light and 14 h of dark every day;
temperature was maintained at 26–28°C (78–82°F); relative humidity
was 40–60%. Tumor growth was monitored every 5 days. At 6 weeks
post-implantation, the mice were sacrificed by cervical
dislocation, and tumor size was calculated as follows: Tumor volume
(mm3) = (L × W2)/2, where L is the long axis,
and W is the short axis. All animal experiments were undertaken in
accordance with the National Institute of Health Guide for the care
and use of laboratory animals, with the approval of the Affiliated
Hospital of Nanchang University, the number of the animal
experiment approved by the institutional committee is 2017(095).
The tissues were cut into 4-µm sections by serial sectioning, and
the sections were dewaxed, alcohol rehydrated and subjected to
microwave antigen retrieval. The tissue samples were immersed in 3%
hydrogen peroxide to block endogenous peroxidase activity. The
expression levels of Akt, PHLPP2 and Ki-67 in the local tumor
issues were determined by immunostaining with antibodies against
Akt (1;200; cat. no. sc-5298), PHLPP2 (1:100; cat. no. sc-71973)
and Ki-67 (1:100) (rabbit monoclonal antibody, cat. no. ab16667;
Abcam). After incubation for 20 min at room temperature with the
addition of polymerase auxiliary agent, horseradish
peroxidase-labeled goat anti-mouse or goat anti-rabbit secondary
antibody (cat. nos. ab6789 and ab6721; Abcam, Inc., Cambridge, MA,
USA) was added, and the sections were incubated at room temperature
for 30 min and stained with diaminobenzidine (DAB) from the
Sigma-Aldrich; EMD Millipore, Billerica, MD, USA). The sections
were counterstained with hematoxylin and then mounted. PBS was used
instead of a primary antibody in the NC, and normal mucosa was used
as a positive control. Cytoplasm and cell membranes containing
brown-yellow granules were defined as having positive expression of
Akt, PHLPP2 and Ki-67 proteins, and nuclei containing brown-yellow
or brown staining indicated positive expression of Akt, PHLPP2 and
Ki-67 proteins. The integral optical density (IOD) level of the
protein of interest was quantified using Gel-Pro-Analyzer software
6.0 (Media Cybernetics, Inc., Rockville MD, USA).
Statistical analysis
Each experiment was performed at least three times,
and the values are reported as the mean ± standard deviation.
Differences between two groups were evaluated using the unpaired
t-test. For multiple-group comparisons, one-way analysis of
variance was followed by Tukey's multiple comparisons test.
P<0.05 was considered to indicate a statistically significant
difference. Event-free survival (EFS) curves were plotted
separately for patients with low, vs. high expression of miR-125b
using the Kaplan-Meier method. The curves were compared by log-rank
analysis (GraphPad Prism version 5.0; GraphPad Software, Inc., La
Jolla, CA, USA). The EFS times were calculated from the time of
diagnosis to the date of relapse, mortality, or last contact.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-125b is overexpressed in plasma
from MM patients and in MM cell lines
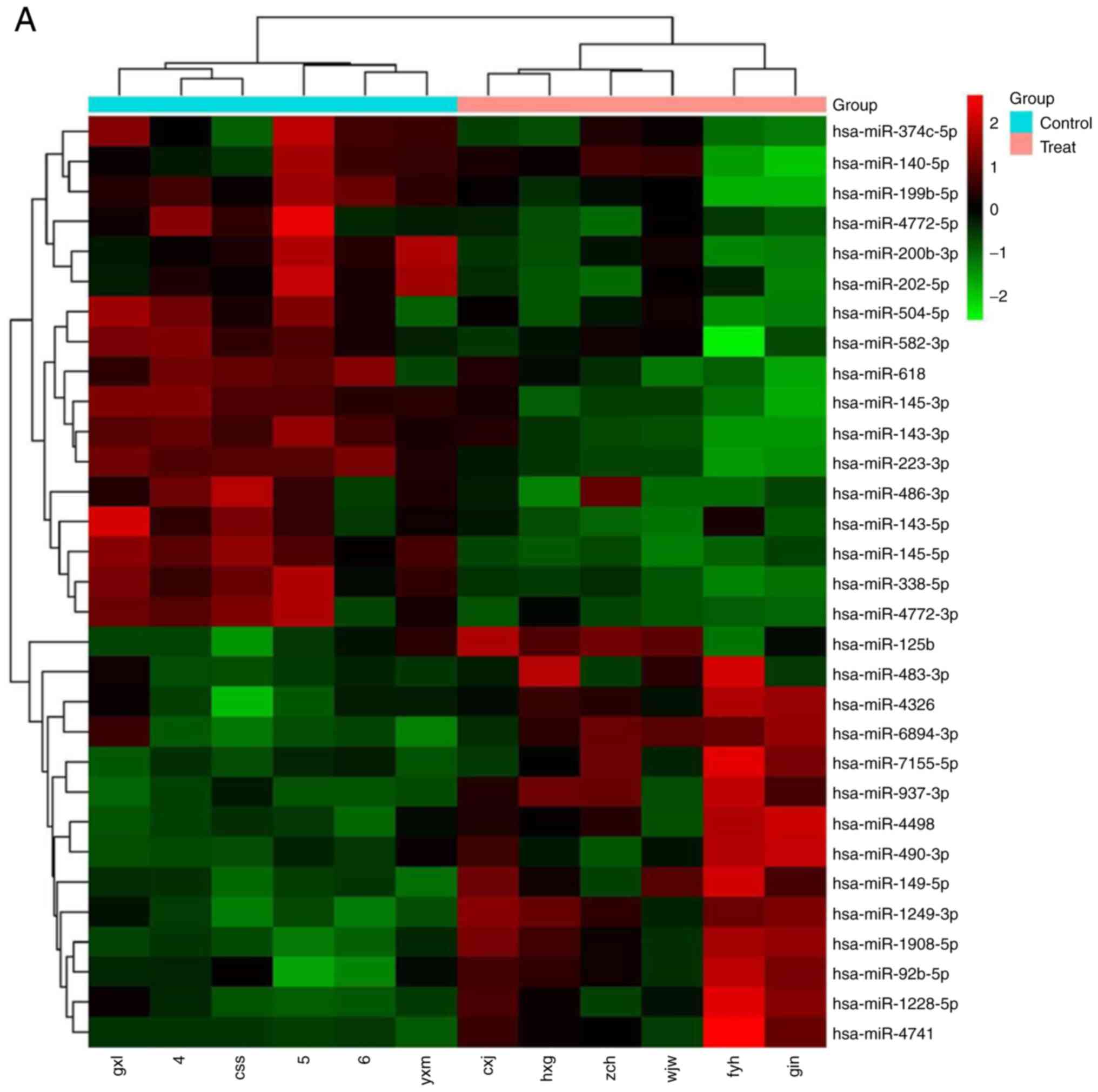
The miRNA microarray analysis revealed that miR-125b
was significantly upregulated in MM plasma (Fig. 1A). To confirm this result, RT-qPCR
analysis was performed in 35 plasma samples from patients with MM
and 20 samples from healthy control individuals. It was found that
the level of circulating miR-125b was upregulated in cells from the
MM group, compared with those from the control subjects. The
normalized expression levels of miR-125b in the patients with MM
and the control subjects were 1.69±0.03 and 1.30±0.08,
respectively, and the expression of miR-125b in the patients with
MM was significantly higher than that in the control individuals
(P<0.05) (Fig. 1B). Consistent
with these data, the expression of miR-125b in the U266, MM.1S, and
RPMI-8226 MM cell lines was significantly increased compared with
the expression in the normal, healthy bone marrow-derived PCs
(Fig. 1C). The expression levels of
circulating miR-125b were significantly higher in patients with MM
with extramedullary infiltration (1.82±0.05) than in patients
without extramedullary infiltration (1.66±0.03), and the expression
level in stage III patients (1.76±0.03) was significantly higher
than that in stage I/II patients (1.62±0.04) (both P<0.05).
However, there was no association between the expression of
miR-125b with and the age, sex or karyotype of the patients (all
P>0.05; Table II).
Association between the expression of
miR-125b and EFS
Kaplan-Meier analyses were performed to assess the
association between the expression levels of miR-125b and clinical
outcome, using the median as a cut-off value. The median follow-up
time was 12.5 months (range 1–26). High levels of circulating
miR-125b in patients with MM were associated with shorter EFS
(P=0.02, Fig. 1D).
Effects of the expression of miR-125b
on cellular growth, migration, invasion and apoptosis
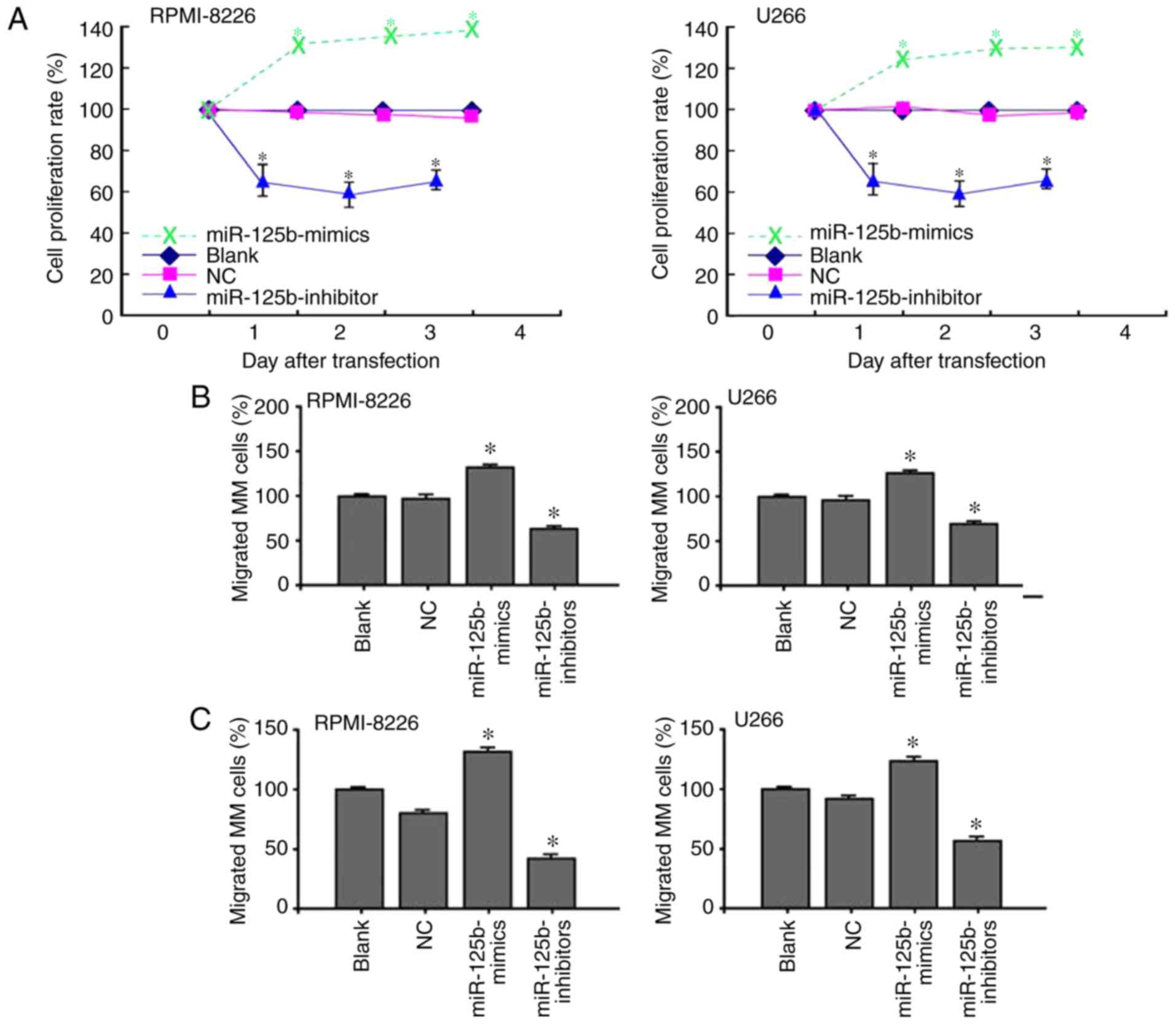
The CCK-8 assay results showed that the viability of
U266 and RPMI-8226 cells was inhibited by transfection of the
miR-125b inhibitor (Fig. 2A). Cell
growth in the miR-125b mimic group was significantly increased
compared with that in the blank and NC groups (P<0.05).
Furthermore, the present study examined whether treatment with the
miR-125b inhibitor inhibited MM cell migration using a Transwell
assay. Consistent with the CCK-8 assay, the Transwell assay
demonstrated that the migration rate of cells transfected with
miR-125b inhibitor was inhibited (Fig.
2B). The number of migrated cells in the miR-125b inhibitor
group was significantly lower than the numbers in the blank and NC
groups (P<0.05). The cell invasion results showed that the
number of invaded cells in the miR-125b inhibitor group was
significantly lower than the numbers in the blank and NC groups
(Fig. 2C) (P<0.05). In addition,
the apoptotic rates in the miR-125b inhibitor group were 23.8% in
the U266 cells and 19.6% in the RPMI-8226 cells (Fig. 2D). Compared with the two control
groups, the apoptotic rate in the miR-125b inhibitor group was
significantly higher (P<0.05) (Fig.
2E).
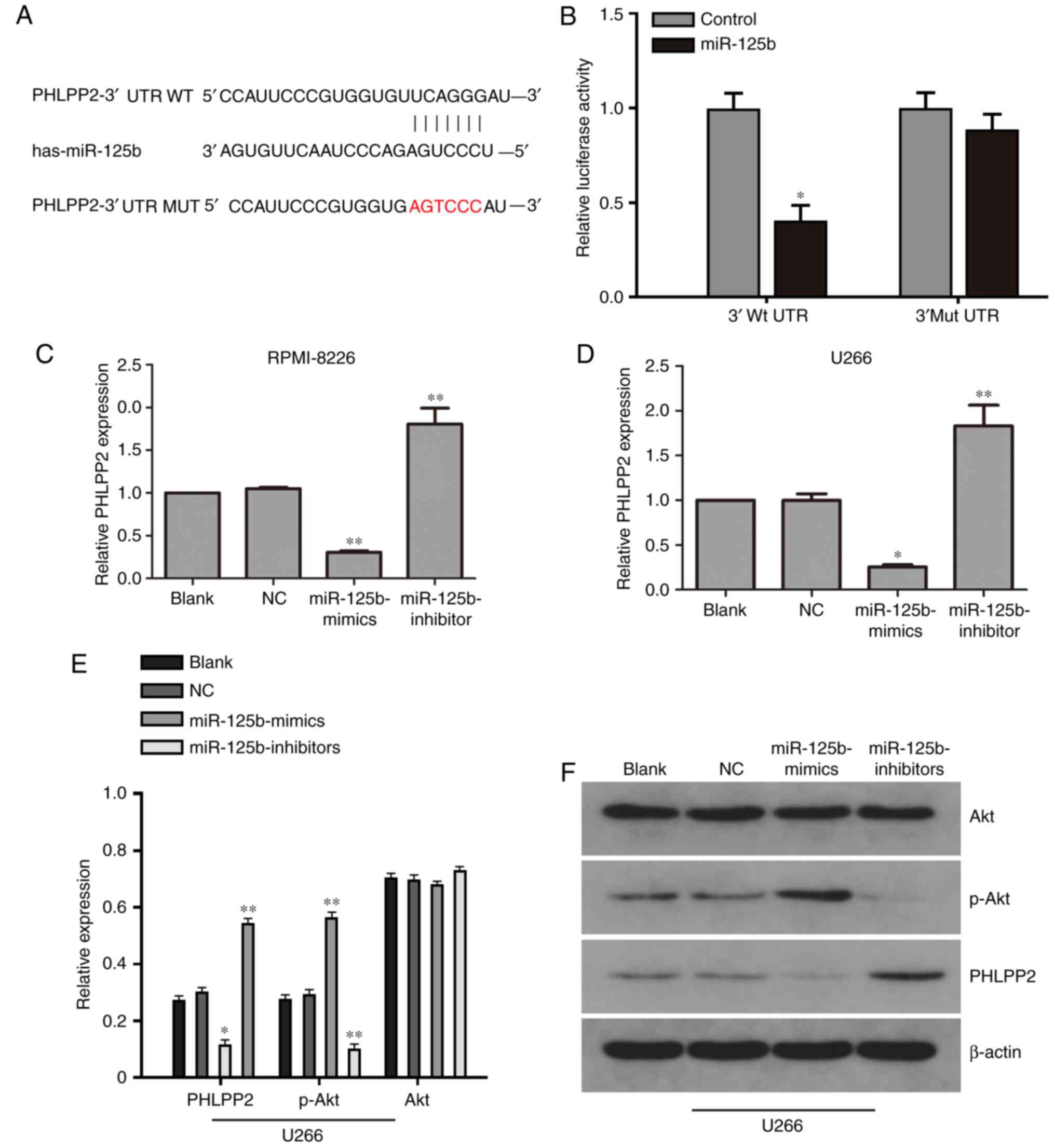
PHLPP2 as a target gene of
miR-125b
On further analyzing the regulatory mechanism
through which miR-125b promotes MM cell proliferation, the present
study found that the tumor suppressor PHLPP2 was the target of
miR-125b, as predicted by TargetScan (http://genes.mit.edu/targetscan.test/ucsc.html)
(Fig. 3A). To determine whether
miR-125b binds to the 3′-UTR of PHLPP2, a luciferase reporter assay
was performed. As expected, the overexpression of miR-125b
inhibited luciferase activity. By contrast, cells with a mutant
PHLPP2 3′-UTR exhibited higher luciferase activity (Fig. 3B). To determine whether miR-125b
downregulates PHLPP2 at the mRNA level, the expression of PHLPP2
was examined by RT-qPCR analysis. As shown in Fig. 3C and D, the mRNA expression of
PHLPP2 was upregulated in MM cells transfected with miR-125b
inhibitor. By contrast, the mRNA level of PHLPP2 was decreased in
MM cells transfected with miR-125b mimic.
Protein expression of PHLPP2, AKT and
p-AKT
In the U266 cells, the western blot analysis results
showed that the expression of p-Akt in the miR-125b mimic group was
downregulated compared with that in the blank and NC groups
(P<0.05). miR-125b knockdown promoted the expression of PHLPP2,
suggesting that miR-125b inhibited PHLPP2 (Fig. 3E and F). No statistically
significant differences in protein expression were found between
the blank and NC groups.
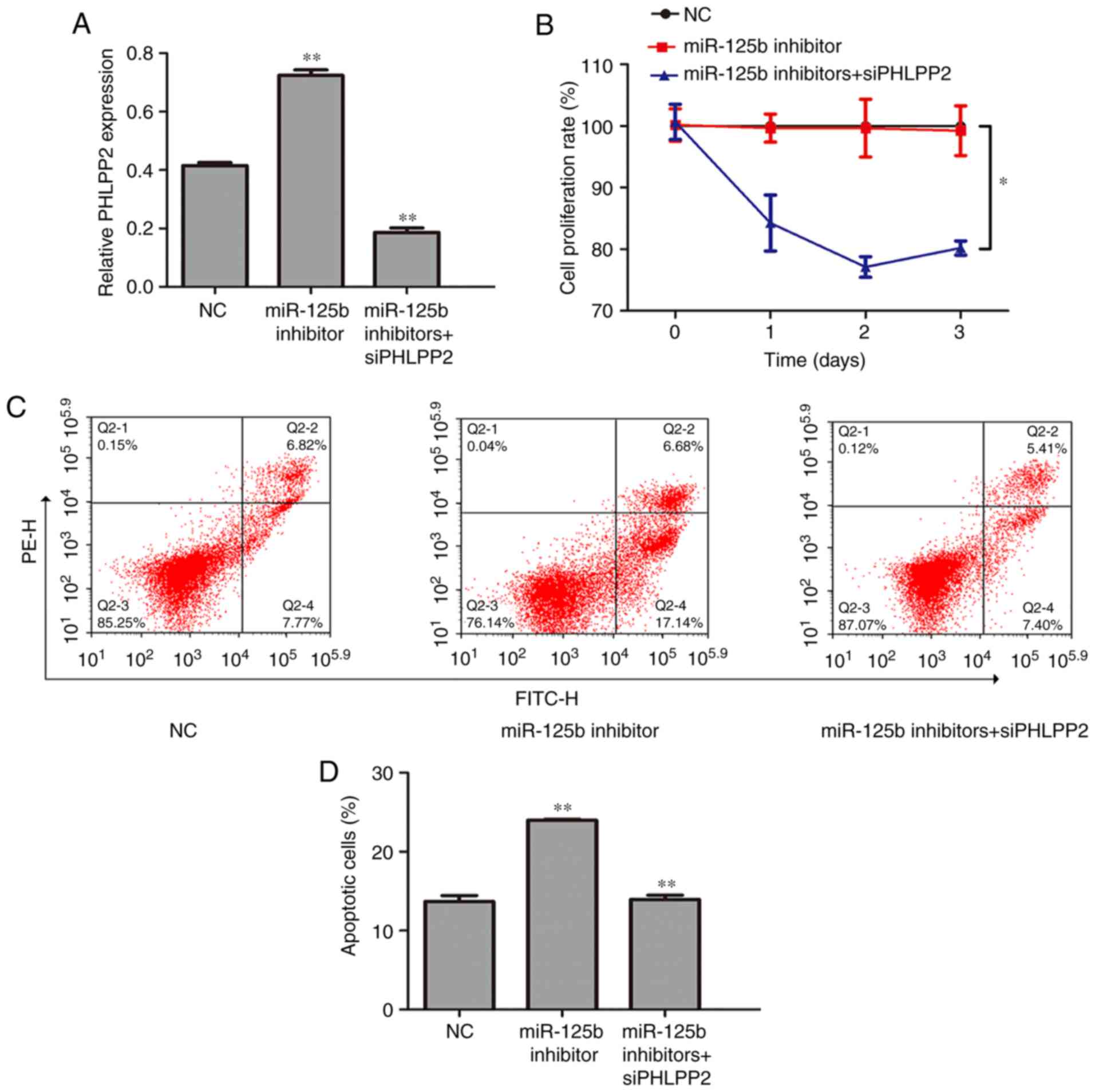
PHLPP2 reverses the effect of
miR-125b
To furthere whether PHLPP2 is a functional target of
miR-125b, rescue experiments were performed by inhibiting PHLPP2.
The U266 cells were cotransfected with miR-125b inhibitor and
siPHLPP2. Transfection with siPHLPP2 successfully decreased the
expression of PHLPP2 in the cotransfected U266 cells (Fig. 4A, P<0.01). The CCK-8 assay
indicated that the cell growth inhibition induced by miR-125b
knockdown was relieved by transfection with siPHLPP2 (Fig. 4B, P<0.05). The results of the
flow cytometry showed that cell apoptosis was increased in miR-125b
inhibitor cells, whereas siPHLPP2 partially reversed this promotion
in cell apoptosis (Fig. 4C and D,
P<0.05).
Effect of the expression of miR-125b
on tumor growth in a nude mouse model
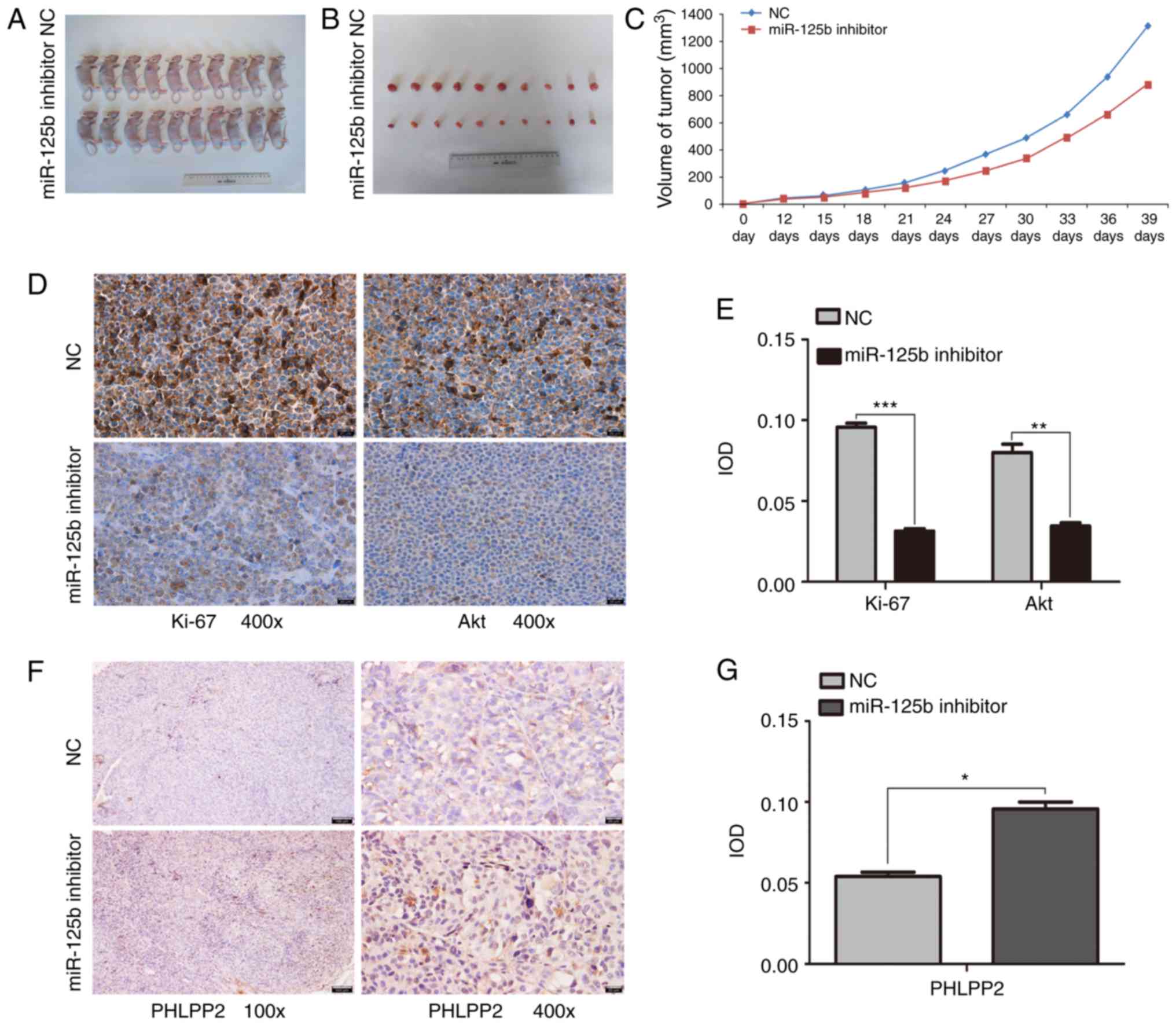
The present study investigated whether the miR-125b
inhibitor was able to inhibit tumor growth in the xenograft tumor
model. Tumor growth was monitored for 39 days. The results showed
that the tumor weights and volumes in the mice treated with
miR-125b inhibitor were significantly reduced compared with those
treated with NC (P<0.05; Fig.
5A-C). In addition, the immunohistochemistry results (Fig. 5D-G) showed that Ki-67 and Akt were
mainly expressed in the NC group. The tumors from the miR-125b
inhibitor group exhibited higher levels of PHLPP2 than the tumors
from the NC group. The immunohistochemistry showed that treatment
with miR-125b inhibitor reduced the expression of Ki-67 and Akt,
and increased the expression of PHLPP2.
Discussion
MM is a PC disorder that represents a relatively
high proportion of hematological malignancies. Despite significant
progress in elucidating the mechanism of MM and identifying optimal
treatments, the disease remains incurable, necessitating the
development of novel and effective therapies (26). The therapeutic modulation of miRNAs
that function as an oncogene or tumor suppressor may be a potential
strategy for cancer treatment. The aim of the present study was to
examine the roles of miR-125b in the proliferation, migration and
invasion of MM cells.
The dysregulation of miRNA expression is important
in cancer development, and circulating miRNAs serve as biomarkers
for several malignancies (27–29).
Aberrant miRNA expression profiles in plasma have also been
reported in MM (30,31). In the present study, miRNA
microarray was used to compare the miRNA expression patterns of
plasma samples from six patients with MM and six healthy controls.
It was found that miR-125b, miR-4326, miR-4498, miR-490-3p and
miR-7155-5p were overexpressed in the MM patient samples. Among
these aberrant miRNAs, miR-125b had the highest degree of
overexpression in samples from patients with MM compared with those
from healthy control patients. Additionally, the expression of
miR-125b was examined in MM cell lines and nPCs. miR-125b was
upregulated in the MM cell lines compared with the nPCs.
Notably, miR-125b belongs to the miR-17-92 cluster
and has been reported to act as an oncogene in several human
malignancies. Wang et al demonstrated that miR-125b was
overexpressed in nasopharyngeal carcinoma (32). Shen et al reported that the
expression of miR-125b was markedly increased in type II diabetes
mellitus (33). In addition, it has
been shown that high levels of miR-125b are associated with
shortened progression-free survival (PFS) times (34). In the present study, the expression
of miR-125b in plasma was significantly higher in patients with
stage III MM than in patients with stage I/II MM, suggesting the
essential role of miR-125b in survival diagnosis. MM cell lines
were utilized for subsequent loss-of-function experiments. The
results showed a marked reduction in MM cell viability following
transfection with an miR-125b inhibitor. The transfected cells also
exhibited reduced migratory ability and an upregulated apoptosis
level. Furthermore, the xenograft mouse model demonstrated the
suppressive effects of miR-125b knockdown on tumor growth in
vivo. The immunohistochemistry results showed that treatment
with miR-125b inhibitor reduced the expression of Ki-67. The
expression of Ki67 is associated with tumor cell proliferation and
growth, and is widely used in routine pathological investigation as
a proliferation marker. It has been reported that the inhibition of
Ki67 either by antibodies or antisense oligonucleotides leads to
the arrest of cell proliferation. Notably, antisense
oligonucleotides and antibodies against p-Ki67 have been shown to
inhibit progression of the cell cycle (35). Therefore, the suppression of Ki67 s
may be a promising strategy in MM therapy. These data suggest that
miR-125b is relevant to the development and progression of MM.
As an oncogenic pathway, the Akt pathway is
frequently activated during tumorigenesis and is crucial in the
growth, proliferation, migration and invasion of malignant cells
(36,37), making this pathway an attractive
candidate for drug development (38). Studies have shown that PI3K/Akt
pathway activation reduces levels and of p21Cip1 and increased the
expression of cyclin D1 (39,40),
which regulate cell cycle progression through the G1 phase. In
addition, activated phosphoinositide 3-kinase (PI3K) catalyzes
3,4-phosphatidylinositol trisphosphate phosphorylation and
subsequently activates protein kinase Akt to promote cell growth
and proliferation (36). Chen et
al reported that the activation of AKT was associated with
intrahepatic metastasis, tumor grade and a high proliferation
index. Furthermore, the Akt pathway is a diagnostic and prognostic
indicator of hepatocellular carcinoma invasion and metastasis, and
a therapeutic target for this type of cancer (41). PHLPP2, a characterized member of the
PHLPP family and suppressor of AKT signaling, exhibits an
inhibitory effect on cell cycle progression and apoptosis in
various tumor cell types (42).
PHLPP2 has been predicted to be a direct target of miR-125b;
therefore, the present study confirmed this prediction via RT-qPCR,
western blot and dual-luciferase reporter assays. The mRNA and
protein levels of PHLPP2 were increased upon the downregulation of
miR-125b in U266 cells. To confirm whether PHLPP2 is a target that
mediates the function of miR-125b, a loss-of-function approach was
used to functionally characterize the roles of PHLPP2 in the growth
and apoptosis of MM. It was found that PHLPP2 silencing partially
attenuated the effects of miR-125b inhibition on cell proliferation
and apoptosis. Evidence suggests that the activation of PI3K/Akt by
miR-181 is mainly mediated by the suppression of PHLPP2 (43). The results of the present study
revealed that p-AKT was downregulated in MM cells in which miR-125b
was suppressed. A previous study examined the mechanism of miR-125b
in the occurrence and progression of MM (44). The present study investigated the
effect of miR-125b on the migration and apoptosis of MM, and on the
Akt signal pathway. The prometastatic effect of miR-125b in MM was
also examined, and a novel target gene of miR-125b was reported.
The results demonstrated a novel role for miR-125b in AKT pathway
activation by the direct suppression of PHLPP2.
In conclusion, the present study demonstrated that
miR-125b regulated MM cell proliferation and migration by
modulating the Akt signaling pathway. Therefore, this pathway may
serve as a novel therapeutic target for MM. Notably, miR-125b
suppressed the expression of PHLPP2, leading to activation of the
AKT pathway. Elucidating the association between miR-125b and the
Akt signaling pathway may assist in identifying novel molecular
markers and potential therapeutic targets for MM treatment. The
effects of miR-125b on MM cell proliferation, migration and
invasion require confirmation using animal models in further
investigations.
Acknowledgements
Not applicable.
Funding
This study was supported by the International
Collaboration Fund of the National Science and Technology Committee
of China (grant no. 2011DFA32820), the National Natural Science
Fund Project (grant no. 81460037) and the Innovation Fund Project
in Jiangxi Province (grant no. YC2016-B018).
Availability of data and materials
The datasets used during in the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
YJ performed the molecular biology experiments and
drafted the manuscript. JD collected, analyzed and interpreted the
data. JL performed the molecular biology experiments and revised
the manuscript. GC conceived and designed the study. All authors
read and approved the manuscript and agree to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
The study was approved by the Medical Research
Ethics Committee of The First Affiliated Hospital of Nanchang
University and written informed consent was obtained from all study
subjects. The IRB number of human sample study approved by
institutional committee is 2015(096). The use of mice in animal
experiments was approved by the Affiliated Hospital of Nanchang
University. The number of animal experiment approved by
institutional committee is 2017(095).
Patient consent for publication
Consent to publish has been obtained from the
participants.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sonneveld P, Avet-Loiseau H, Lonial S,
Usmani S, Siegel D, Anderson KC, Chng WJ, Moreau P, Attal M, Kyle
RA, et al: Treatment of multiple myeloma with high-risk
cytogenetics: a consensus of the International Myeloma Working
Group. Blood. 127:2955–2962. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Abdi J, Chen G and Chang H: Erratum: Drug
resistance in multiple myeloma: Latest findings and new concepts on
molecular mechanisms. Oncotarget. 6:73642015.PubMed/NCBI
|
|
3
|
Mimura N, Hideshima T and Anderson KC:
Novel therapeutic strategies for multiple myeloma. Exp Hematol.
43:732–741. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Costa LJ, Brill IK, Omel J, Godby K, Kumar
SK and Brown EE: Recent trends in multiple myeloma incidence and
survival by age, race, and ethnicity in the United States. Blood
Adv. 1:282–287. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lionetti M, Biasiolo M, Agnelli L,
Todoerti K, Mosca L, Fabris S, Sales G, Deliliers GL, Bicciato S,
Lombardi L, et al: Identification of microRNA expression patterns
and definition of a microRNA/mRNA regulatory network in distinct
molecular groups of multiple myeloma. Blood. 114:e20–e26. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Calin GA and Croce CM: MicroRNAs and
chromosomal abnormalities in cancer cells. Oncogene. 25:6202–6210.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Calin GA and Croce CM: MicroRNA-cancer
connection: The beginning of a new tale. Cancer Res. 66:7390–7394.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Seckinger A, Meißner T, Moreaux J, Benes
V, Hillengass J, Castoldi M, Zimmermann J, Ho AD, Jauch A,
Goldschmidt H, et al: miRNAs in multiple myeloma-a survival
relevant complex regulator of gene expression. Oncotarget.
6:39165–39183. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Leotta M, Biamonte L, Raimondi L,
Ronchetti D, Di Martino MT, Botta C, Leone E, Pitari MR, Neri A,
Giordano A, et al: A p53-dependent tumor suppressor network is
induced by selective miR-125a-5p inhibition in multiple myeloma
cells. J Cell Physiol. 229:2106–2116. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chi J, Ballabio E, Chen XH, Kusec R,
Taylor S, Hay D, Tramonti D, Saunders NJ, Littlewood T, Pezzella F,
et al: MicroRNA expression in multiple myeloma is associated with
genetic subtype, isotype and survival. Biol Direct. 6:232011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Luo X, Gu J, Zhu R, Feng M, Zhu X, Li Y
and Fei J: Integrative analysis of differential miRNA and
functional study of miR-21 by seed-targeting inhibition in multiple
myeloma cells in response to berberine. BMC Syst Biol. 8:822014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Esquela-Kerscher A: The lin-4 microRNA:
The ultimate micromanager. Cell Cycle. 13:1060–1061. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Iorio MV and Croce CM: microRNA
involvement in human cancer. Carcinogenesis. 33:1126–1133. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Garzon R and Marcucci G: Potential of
microRNAs for cancer diagnostics, prognostication and therapy. Curr
Opin Oncol. 24:655–659. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ma Q, Chen Z, Jia G, Lu X, Xie X and Jin
W: The histone demethylase PHF8 promotes prostate cancer cell
growth by activating the oncomiR miR-125b. Onco Targets Ther.
8:1979–1988. 2015.PubMed/NCBI
|
|
16
|
Yu X, Shi W, Zhang Y, Wang X, Sun S, Song
Z, Liu M, Zeng Q, Cui S and Qu X: CXCL12/CXCR4 axis induced
miR-125b promotes invasion and confers 5-fluorouracil resistance
through enhancing autophagy in colorectal cancer. Sci Rep.
7:422262017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wong TN and Link DC: miR-125b
promotes leukemogenesis via VEGFA. Blood. 129:1409–1410.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang Q, Wang Y, Lu X, Zhao Z, Zhu L, Chen
S, Wu Q, Chen C and Wang Z: MiR-125b regulates
epithelial-mesenchymal transition via targeting Sema4C in
paclitaxel-resistant breast cancer cells. Oncotarget. 6:3268–3279.
2015.PubMed/NCBI
|
|
19
|
Zhang Y and Huang S: Up-regulation of
miR-125b reverses epithelial-mesenchymal transition in
paclitaxel-resistant lung cancer cells. Biol Chem. 2015. View Article : Google Scholar
|
|
20
|
Luo M, Tan X, Mu L, Luo Y, Li R, Deng X,
Chen N, Ren M, Li Y, Wang L, et al: MiRNA-21 mediates the
antiangiogenic activity of metformin through targeting PTEN and
SMAD7 expression and PI3K/AKT pathway. Sci Rep. 7:434272017.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu MH, Yang L, Liu XJ, Nie ZY and Luo JM:
Targeted suppression of miRNA-21 inhibit K562 cells growth through
PTEN-PI3K/AKT signaling pathway. Zhonghua Xue Ye Xue Za Zhi.
37:982–986. 2016.(In Chinese). PubMed/NCBI
|
|
22
|
Yang X, Cheng Y, Li P, Tao J, Deng X,
Zhang X, Gu M, Lu Q and Yin C: A lentiviral sponge for miRNA-21
diminishes aerobic glycolysis in bladder cancer T24 cells via the
PTEN/PI3K/AKT/mTOR axis. Tumour Biol. 36:383–391. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang F, Li L, Chen Z, Zhu M and Gu Y:
MicroRNA-214 acts as a potential oncogene in breast cancer by
targeting the PTEN-PI3K/Akt signaling pathway. Int J Mol Med.
37:1421–1428. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Anderson KC: Progress and paradigms in
multiple myeloma. Clin Cancer Res. 22:5419–5427. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bates SE: Multiple myeloma: Multiplying
therapies. Clin Cancer Res. 22:54182016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Gasparri ML, Casorelli A, Bardhi E,
Besharat AR, Savone D, Ruscito I, Farooqi AA, Papadia A, Mueller
MD, Ferretti E, et al: Beyond circulating microRNA biomarkers:
Urinary microRNAs in ovarian and breast cancer. Tumour Biol.
39:10104283176955252017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zeng Z, Chen X, Zhu D, Luo Z and Yang M:
Low expression of circulating MicroRNA-34c is associated with poor
prognosis in triple-negative breast cancer. Yonsei Med J.
58:697–702. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang X, Wang W, Yang Y, Du L, Yang X,
Wang L, Zheng G, Duan W, Wang R, Zhang X, et al: Identification of
circulating microRNA signatures as potential noninvasive biomarkers
for prediction and prognosis of lymph node metastasis in gastric
cancer. Oncotarget. 8:65132–65142. 2017.PubMed/NCBI
|
|
30
|
Qu X, Zhao M, Wu S, Yu W, Xu J, Xu J, Li J
and Chen L: Circulating microRNA 483-5p as a novel biomarker for
diagnosis survival prediction in multiple myeloma. Med Oncol.
31:2192014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Huang JJ, Yu J, LiJ Y, Liu YT and Zhong
RQ: Circulating microRNA expression is associated with genetic
subtype and survival of multiple myeloma. Med Oncol. 29:2402–2408.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li LN, Xiao T, Yi HM, Zheng Z, Qu JQ,
Huang W, Ye X, Yi H, Lu SS, Li XH and Xiao ZQ: MiR-125b increases
nasopharyngeal carcinoma radioresistance by targeting A20/NF-κB
signaling pathway. Mol Cancer Ther. 16:2094–2106. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shen Y, Xu H, Pan X, Wu W, Wang H, Yan L,
Zhang M, Liu X, Xia S and Shao Q: miR-34a and miR-125b are
upregulated in peripheral blood mononuclear cells from patients
with type 2 diabetes mellitus. Exp Ther Med. 14:5589–5596.
2017.PubMed/NCBI
|
|
34
|
Piatopoulou D, Avgeris M, Marmarinos A,
Xagorari M, Baka M, Doganis D, Kossiva L, Scorilas A and Gourgiotis
D: miR-125b predicts childhood acute lymphoblastic leukaemia poor
response to BFM chemotherapy treatment. Br J Cancer. 117:801–812.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li LT, Jiang G, Chen Q and Zheng JN: Ki67
is a promising molecular target in the diagnosis of cancer
(review). Mol Med Rep. 11:1566–1572. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang Z, Fang S, Di Y, Ying W, Tan Y and Gu
W: Modulation of NF-κB/miR-21/PTEN pathway sensitizes non-small
cell lung cancer to cisplatin. PLoS One. 10:e01215472015.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang Y, Zheng L, Ding Y, Li Q, Wang R,
Liu T, Sun Q, Yang H, Peng S, Wang W and Chen L: MiR-20a induces
cell radioresistance by activating the PTEN/PI3K/Akt signaling
pathway in hepatocellular carcinoma. Int J Radiat Oncol Biol Phys.
92:1132–1140. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Carnero A, Blanco-Aparicio C, Renner O,
Link W and Leal JF: The PTEN/PI3K/AKT signalling pathway in cancer,
therapeutic implications. Curr Cancer Drug Targets. 8:187–198.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Medema RH, Kops GJ, Bos JL and Burgering
BM: AFX-like forkhead transcription factors mediate cell-cycle
regulation by ras and PKB through p27kip1. Nature.
404:782–787. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Roy SK, Srivastava RK and Shankar S:
Inhibition of PI3K/AKT and MAPK/ERK pathways causes activation of
FOXO transcription factor, leading to cell cycle arrest and
apoptosis in pancreatic cancer. J Mol Signal. 5:102010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen JS, Wang Q, Fu XH, Huang XH, Chen XL,
Cao LQ, Chen LZ, Tan HX, Li W, Bi J and Zhang LJ: Involvement of
PI3K/PTEN/AKT/mTOR pathway in invasion and metastasis in
hepatocellular carcinoma: Association with MMP-9. Hepatol Res.
39:177–186. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wei XE, Zhang FY, Wang K, Zhang QX and
Rong LQ: Assembly of the FKBP51-PHLPP2-AKT signaling complex in
cerebral ischemia/reperfusion injury in rats. Brain Res.
1566:60–68. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Strotbek M, Schmid S, Sanchez-Gonzalez I,
Boerries M, Busch H and Olayioye MA: miR-181 elevates Akt signaling
by co-targeting PHLPP2 and INPP4B phosphatases in luminal breast
cancer. Int J Cancer. 140:2310–2320. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gao D, Xiao Z, Li HP, Han DH and Zhang YP:
The mechanism study of miR-125b in occurrence and progression of
multiple myeloma. Cancer Med. 7:134–145. 2018. View Article : Google Scholar : PubMed/NCBI
|