Introduction
It has been recognized that less than 2% of the
total genome sequence are protein coding genes, while more than 80%
of the genome are non-protein coding genes (1). Apart from small amount of non-coding
RNAs, such as microRNAs, a large portion of transcribed RNA
constituents are long non-coding RNAs (lncRNAs), which have a
length of more than 200 nucleotides with no open reading frame, and
are controlled by both epigenetic and transcriptional factors
(2). In recent years, thousands of
lncRNAs have been found to be differentially expressed between
cancers and normal tissues (3,4).
Notably, the aberrant expression of lncRNAs was revealed to be an
important contributor to the development of cancers and several
other diseases (5,6). lncRNAs have been implicated in the
regulation of various biological processes, such as chromosomal
imprinting, growth, differentiation, pluripotency, apoptosis and
cell cycle arrest (7). Owing to the
secondary and tertiary structure, lncRNAs act as decoys, guides or
scaffolds to post-transcriptionally regulate gene expression
(8). Despite the increasing studies
on lncRNAs, the role of lncRNAs in cancer remains unclear. To date,
lncRNA-targeted approaches have been translated to clinics. It is
an important task to unravel the functions of lncRNAs in different
cancers, so as to achieve optimal diagnostic and therapeutic
efficacies in cancer.
Osteosarcoma is one of most common cancers in
children and adolescents, comprising 20% of all primary bone
cancers (9). The overall survival
of osteosarcoma remains dismal and approximately 35% of patients
succumb to the disease within five years, despite several
improvements in the treatment approaches. Understanding the
mechanisms associated with osteosarcoma cell proliferation,
differentiation, invasion, and metastasis is critical in advancing
the clinical management of the disease. Consequently, it is
imperative to investigate the molecular mechanism of osteosarcoma
and pinpoint key targets for efficient diagnosis and treatment of
this disease. Recent evidence has indicated that lncRNAs are of
great significance in the pathogenesis of osteosarcoma. lncRNA UCA1
(10), HULC (11) and TUG1 (12) have been reported to play an
important role in promoting osteosarcoma progression, leading to
poor prognosis of patients. Recently, long intergenic
non-protein-coding RNA 1296 (LINC01296) has been implicated in the
regulation of a plethora of cancers. LINC01296 has been revealed to
be a tumor-promoting molecule in prostate (13), colorectal (14) and gastric cancer. However, the role
of LINC01296 in osteosarcoma has never been reported.
The aim of the present study was to explore the
functional role of LINC01296 in osteosarcoma. The expression of
LINC01296 in patients with osteosarcoma tissues was analyzed. The
mechanism of action of LINC01296 in osteosarcoma invasion,
migration and the cell cycle was explored.
Materials and methods
Patient samples
The present study was approved by the Research
Ethics Committee of Jilin University (Changchun, China). Tissues
were collected from 30 patients (17 males and 13 females from 40 to
76 years old, averaged 62.26±6.87 years old), among which 18
patients had a tumor size <6 cm and 12 patients had a tumor size
>6 cm. Tumors of these patients were graded as well
differentiated (6 patients), moderately differentiated (17
patients) and poorly differentiated (7 patients). Lymph node
metastases were observed in 20 patients. Four patients were at
stage II, 19 at stage III and 7 at stage IV. Informed consent was
obtained from all patients.
Quantitative real-time PCR
Total RNA was extracted from human tissues and cells
using TRIzol Total RNA Isolation kit (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). Purified RNA (1 µg) was transcribed to
cDNA using the High-Capacity cDNA Reverse Transcription kit (Thermo
Fisher Scientific, Inc.). Real-time PCR was performed using the
SYBR Green Master Mix (Invitrogen; Thermo Fisher Scientific, Inc.)
and Mastercycler (Eppendorf, Hamburg Germany). The following
sequences were used: LINC01296 forward, AACTGGCACCAGCCTCACT and
reverse, CGGCCAACTTCTTTACCATC; GAPDH forward, ACTGGAACGTGAAGGTG and
reverse, AGAGAAGTGGGGTGGCTT; cyclin E1 forward,
TGGATTGGTTAATGGAGGTGTGTG and reverse, AGCTGTTGGATCTCTGTGTCCTG; CDK4
forward, TAACCCTGGTGTTTGAGCATGTAG and reverse,
GTCGGCTTCAGAGTTTCCACAGA; CDK2 forward, TCTGCCATTCTCATCGGGTCC and
reverse, GAAATCCGCTTGTTAGGGTCGTA; cyclin D1 forward,
GCATCTACACCGACAACTCCATC and reverse, CGTGTGAGGCGGTAGTAGGA. GAPDH
was used as an internal control. The thermocycling conditions were
based on the Tm of primer (initial denaturation was 95°C for 1 min,
in 40 cycles, 95°C for 30 sec, 55-60°C for 30 sec and 68°C for 1
min, final 68°C for 10 min). Quantification of RNA levels was
performed using the 2−ΔΔCq method (15). For the qRT-PCR analysis of LINC01296
expression in patient samples, the top 50% (15 cases) was divided
into the high-expressing group, and the bottom 50% (15 cases) was
divided into the low-expressing group.
Cell lines and cell culture
All cells used in this study, hFOB1.19, MG63 and
143B, were purchased from the American Type Culture Collection
(ATCC; Rockville, MD, USA). HOS-8603 was purchased from Second
Military Medical University Cell Bank (Shanghai, China). It is a
type of Chinese-established human osteosarcoma cell line with
phenotypic characteristics of osteoblasts (16), and has been used in some OS studies
from 1994 (17,18). The medium and atmosphere of those
cell lines cultured in were following: hFOB1.19 [DMEM (cat. no.
11039-02125030149; Gibco; Thermo Fisher Scientific, Inc.) at 34°C],
SW1353 [Leibovitz's L-15 Medium (cat. no. 21083-027; Gibco; Thermo
Fisher Scientific, Inc.) at 37°C in 100% air], MG63 [MEM (cat. no.
11095-080; Gibco; Thermo Fisher Scientific, Inc.) at 37°C in 95%
air+5% CO2], HOS-8603 [DMEM (cat. no. 10566016; Gibco;
Thermo Fisher Scientific, Inc.) at 37°C in 95% air+5%
CO2] and 143B [MEM (cat. no. 11095-080B23151; Gibco;
Thermo Fisher Scientific, Inc.) at 37°C in 100% air]. Due to
experimental conditions and the similar pre-test results among
these OS cell lines, we selected MG63 and HOS-8603 as research cell
lines in functional assays.
Gene knockdown and overexpression
The following RNA sequences were used for gene
knockdown: sh-LINC01296-1,
CCGGcatatgatacatttgtgttaaCTCGAGttaacacaaatgtatcatatgTTTTTG
(forward) and
AATTCAAAAAcatatgatacatttgtgttaaCTCGAGttaacacaaatgtatcatatg
(reverse); sh-LINC01296-2,
CCGGcaggaagcagacagtccccttCTCGAGaaggggactgtctgcttcctgTTTTTG
(forward) and
AATTCAAAAAcaggaagcagacagtccccttCTCGAGaaggggactgtctgcttcctg
(reverse); si-Cyclin D1,
ACCTCGGATGCTGGAGATGTGAAGTTTCAAGAGAACTTCACATCTCCAGCATCCTT.
Transfection of shRNA was performed using the pLKO.1 transfer
plasmid (cat. no. 10878; Addgene, Inc., Cambridge, MA, USA).
Transfection of siRNAs was carried out using the Lipofectamine 2000
system (Invitrogen; Thermo Fisher Scientific, Inc.). For gene
overexpression, cDNA was cloned into the pLV plasmid and
transfected using Lipofectamine 2000.
Cell Counting Kit-8 assay
Cell Counting Kit-8 assay (CCK-8; Dojindo Molecular
Laboratories, Tokyo, Japan) was used to monitor cell proliferation.
First, cells (5×103) were plated in 96-well plates.
Then, 10 µl of CCK-8 solution was added to the cells and incubation
followed at 37°C for 0.5–4 h. The absorbance was measured at 450 nm
using a plate reader (Tecan Trading AG, Mannedorf, Switzerland).
The viability of cells was quantified using the following equation:
Viability (%) = (ODcontrol -
ODexperiment)/ODcontrol × 100%.
Flow cytometry
Cell cycle progression analysis was performed using
flow cytometry. Cells were harvested and fixed in 70% ethanol.
Then, the cells were lysed with 0.2% Triton X-100 at 4°C for 30 min
and pelleted by centrifugation (1,200 × g). Subsequently, the cells
were resuspended in PBS containing RNAse (10 mg/ml), stained with
propidium iodide (PI), and finally analyzed on FACSCalibur (BD
Biosciences, San Jose, CA, USA). The generated histogram was used
to calculate the ratio of cells in the G0/G1, S or G2/M phases.
Western blot analysis
Western blot analysis was carried out to analyze
cell-cycle checkpoint protein expression. Protein extracted by RIPA
buffer (cat. no. 89900; Thermo Fisher Scientific, Inc.) and was
measured concentration by BCA (cat. no. 23225; Thermo Fisher
Scientific, Inc.) and UV-5800H spectrophotometer (Shanghai Metash
Instruments Co., Ltd., Shanghai, China). Cell lysates (20 µg) were
used for 3–8% SDS-PAGE (cat. no. EA0375BOX; Thermo Fisher
Scientific, Inc.) and then transferred to polyvinylidene fluoride
(PVDF) membranes. BSA (1%) was used to block the PVDF membranes.
Then, primary antibodies were applied to the membranes and
incubated at room temperature for 1 h. Primary antibodies such as
cyclin D1 (cat. no. ab16663), cyclin E1 (cat. no. ab133266), CDK4
(cat. no. ab108357) and CDK2 (cat. no. ab32147) were purchased from
Abcam (Cambridge, MA, USA). GAPDH antibody (cat. no. 2118) was
obtained from Cell Signaling Technology (Danvers, MA, USA). Primary
antibodies were diluted 1:1,000. After extensive washing, GAPDH
antibody (cat. no. 2118), the secondary rabbit antibody, was added
to the membrane (1:5,000). Following incubation for 1 h, the
membranes were washed and ECL chemiluminescent reagents (cat. no.
6883; Cell Signaling Technology) were added for visualization of
the protein bands.
Statistical analysis
All experiments were performed in triplicates unless
otherwise stated. Data were expressed in the form of the mean ±
standard deviation (SD). Comparisons between two groups was
performed using Student's t-test. Multiple comparisons among more
than three groups were performed using one-way (ANOVA), and
subsequently, SNK-q test was used additionally in the comparison
between two groups. For the comparison of rates a Chi-square test
was used. Correlation analysis was performed using Pearson's
correlation analysis. Kaplan-Meier analysis was used for overall
survival analysis. Intergroup differences were considered
statistically significant when the P-value was <0.05.
Results
Malignant osteosarcoma is
characterized by high LINC01296 expression
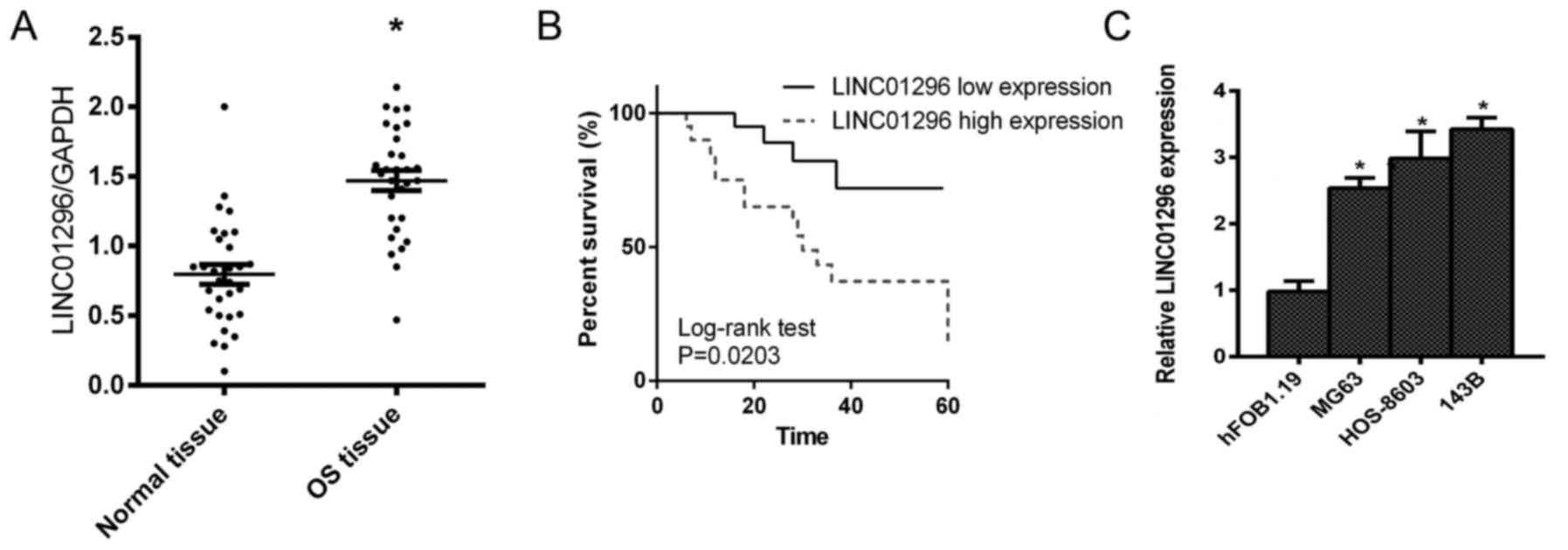
To validate the role of LINC01296 in osteosarcoma,
we performed qPCR analysis from tissues collected from patients
with osteosarcoma. LINC01296 expression was revealed to be
significantly higher in osteosarcoma tissues, compared to that in
normal tissues (Fig. 1A).
Consistently, the survival of patients with high LINC01296
expression was poorer (Fig. 1B).
These data ascertained the potential of LINC01296 as a biomarker
for malignant osteosarcoma. For further confirmation, LINC01296
expression in normal Hfob1.19 bone cells and several osteosarcoma
cells was analyzed, respectively. As anticipated, LINC01296 was
revealed to be greatly upregulated in osteosarcoma cells (Fig. 1C), which further validated the
potential role of LINC01296 as an oncogenic factor in osteosarcoma.
Based on this, we proceeded to analyze the role of LINC01296 in
cell proliferation, migration and invasion.
Indispensable role of LINC01296 in the
promotion of proliferation, migration and invasion of osteosarcoma
cells
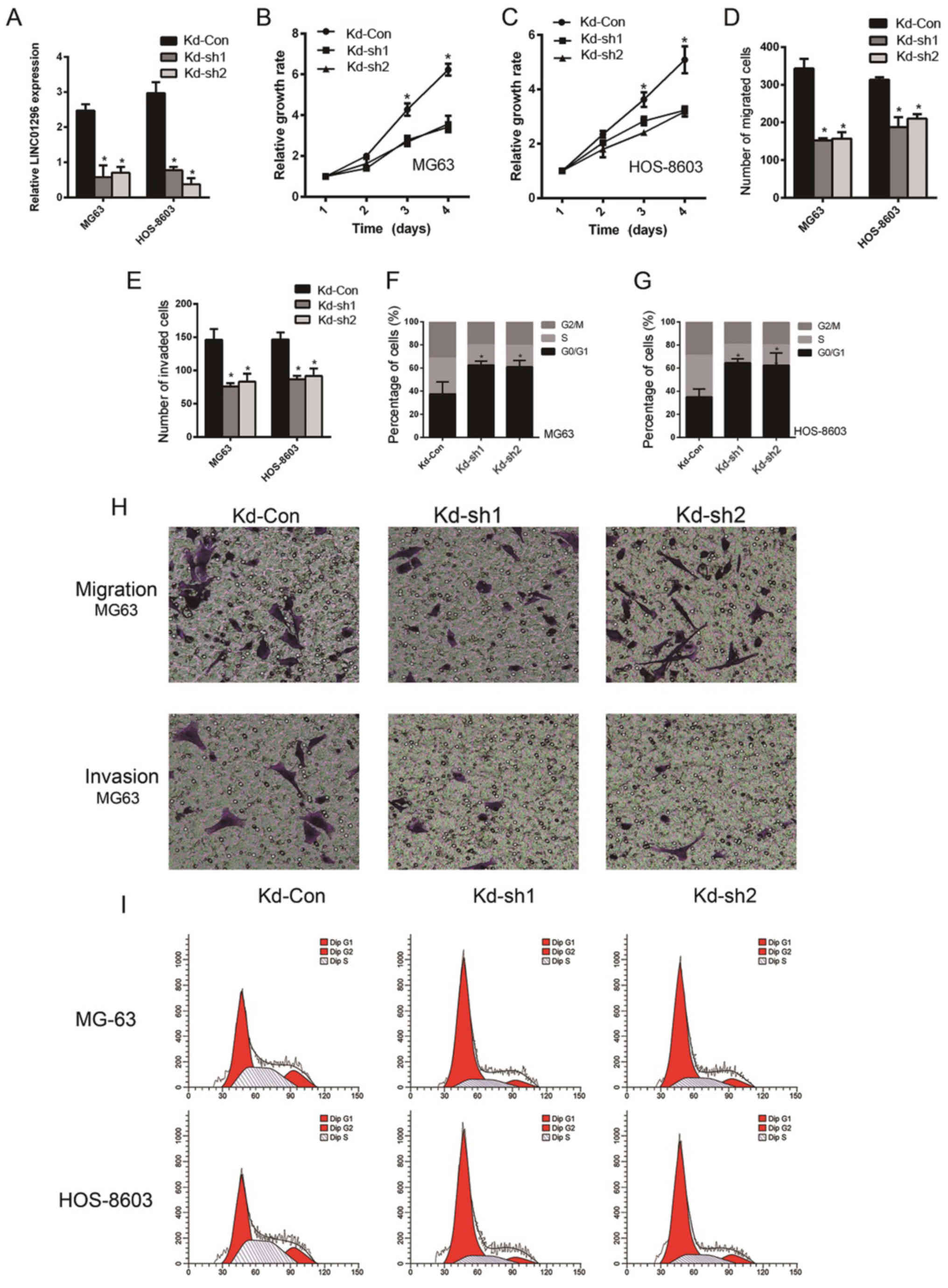
To corroborate the role of LINC01296 in promoting
osteosarcoma cell proliferation, invasion, migration and cell cycle
regulation, LINC01296 knockdown was carried out by shRNA
transfection, followed by monitoring of the phenotypical changes.
As revealed in Fig. 2A, the cells
transfected with LINC01296 specific shRNAs, Kd-sh1 and Kd-sh2,
demonstrated a significant downregulation of LINC01296, compared to
cells transfected with non-coding shRNA (P<0.05). Proliferation
of MG63 and HOS-8603 cells transfected with these shRNAs was
monitored for 4 days. A significantly lower growth rate was
observed in cells transfected with LINC01296 shRNAs (Fig. 2B and C). The migration and invasion
of cells were decreased after LINC01296 knockdown, as revealed in
Fig. 2D, E and H, respectively.
Cell cycle arrest was prominent in MG63 and HOS-8603 cells
transfected with LINC01296 shRNAs, as revealed by the increased
ratio of cells in the G0/G1 phase (Fig.
2F and G). Fig. 2I displays
representative flow cytometric graphs, from which we can summarize
the findings of Fig. 2F and G.
These data indicated that high LINC01296 expression plays an
indispensable role in enhancing the proliferation, invasion and
migration of osteosarcoma cells. Although knockdown of LINC01296 in
MG63 and HOS-8603 cell lines by shRNA is presented, we also used
siRNA for knockdown in the MG63, HOS-8603 and 143B cell lines in
pre-tests with similar results (data not shown). Due to the special
cultured condition of 143B, we only used the MG63 and HOS-8603
cells in the functional assays of this study.
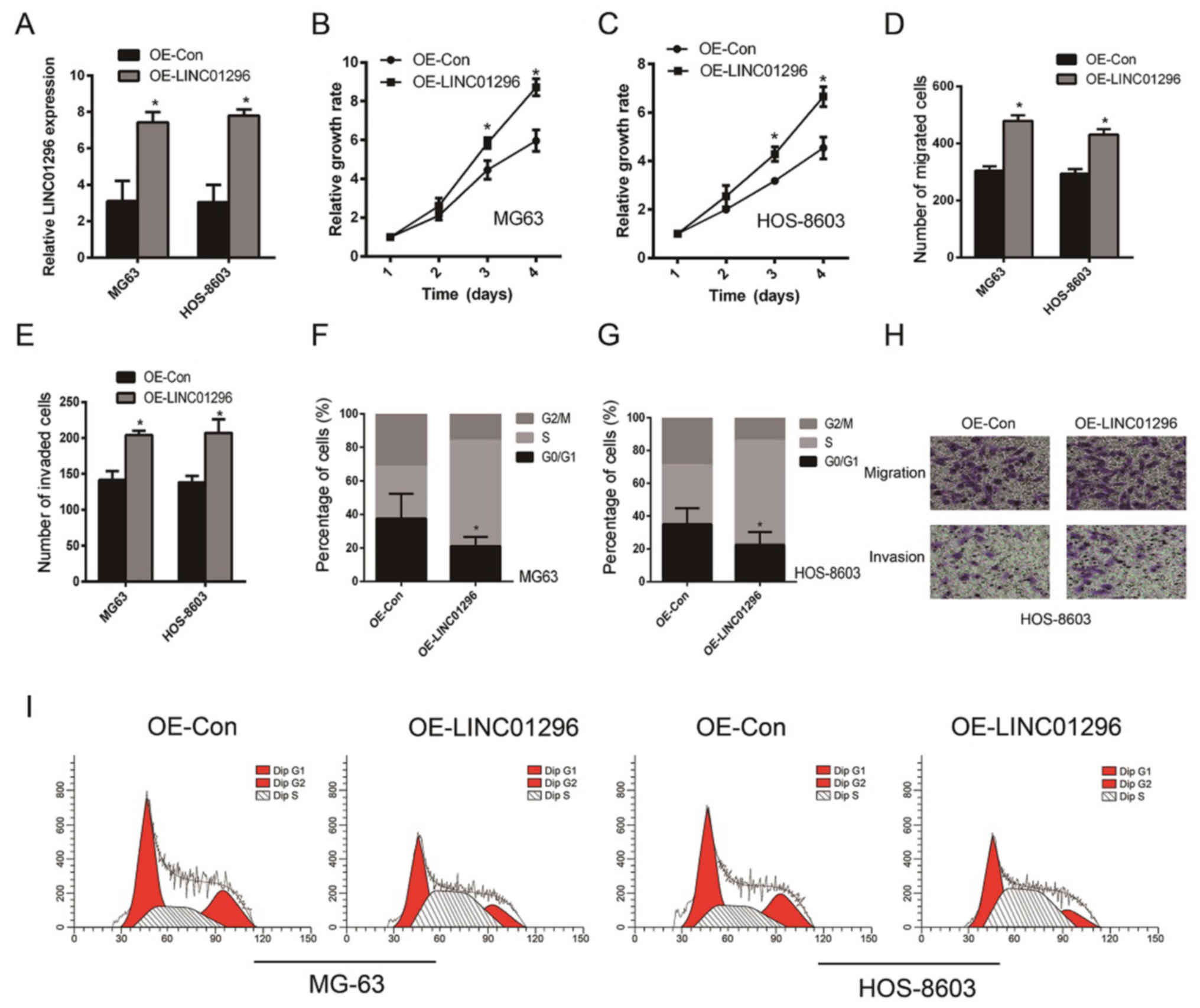
To further validate the tumor-promoting role of
LINC01296, ectopical overexpression of LINC01296 in osteosarcoma
cells was carried out. The overexpression of LINC01296 was
confirmed by qPCR analysis (Fig.
3A). The proliferation of MG63 (Fig. 3B) and HOS-8603 (Fig. 3C) was significantly increased after
LINC01296 overexpression (P<0.05), which was consistent with our
hypothesis. In addition, greater migration (Fig. 3D and H) and invasion (Fig. 3E and H) of osteosarcoma cells were
observed. Notably, the reduction of MG63 (Fig. 3F) and HOS-8603 (Fig. 3G) cells at the G0/G1 phase was quite
marked (Fig. 3I). In conclusion,
this evidence points to the pivotal role of LINC01296 in promoting
osteosarcoma progression. Nevertheless, further elucidation is
still warranted to validate the effect of LINC01296 on the
regulation of the cell cycle.
LINC01296 positively regulates cyclin
D1 expression
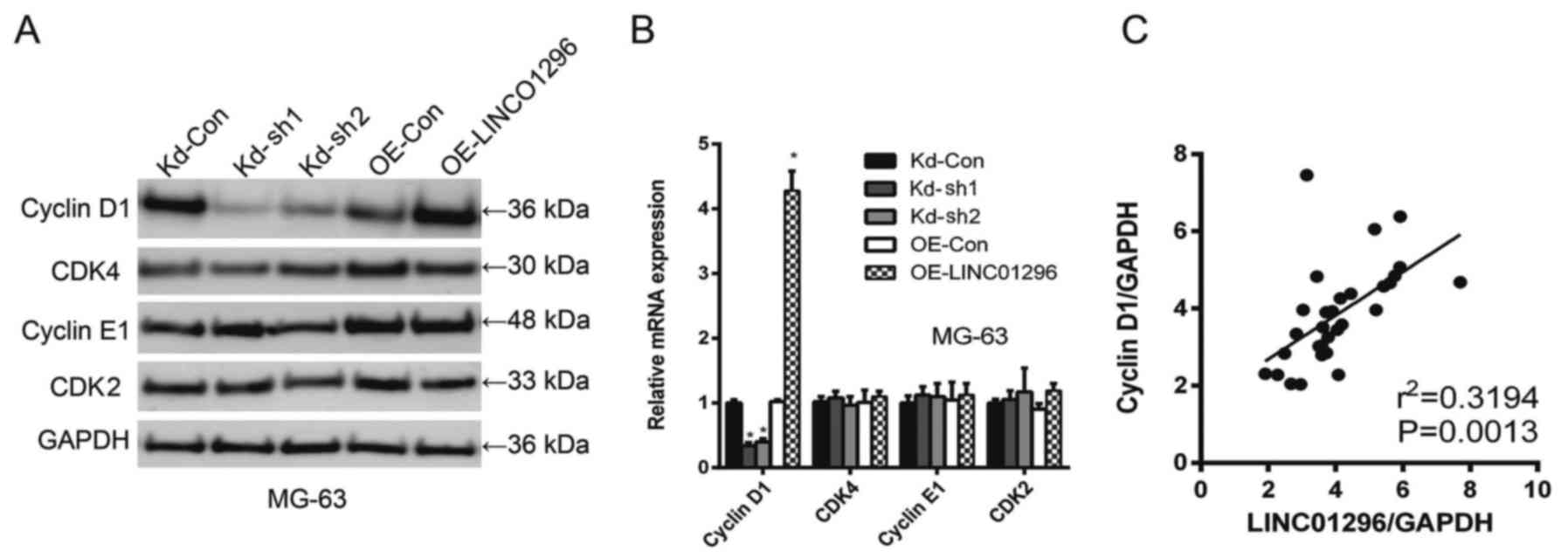
Considering the role of LINC01296 in the regulation
of the cell cycle of osteosarcoma cells, we investigated how
knockdown and overexpression of LINC01296 affected the expression
of cell cycle regulators, including cyclin D1, CDK4, cyclin E1 and
CDK2. As revealed in Fig. 4A, in
MG-63 cells, cyclin D1 expression was closely correlated with
LINC01296 levels, i.e., the knockdown of LINC01296 downregulated
cyclin D1 expression while the overexpression of LINC01296
upregulated cyclin D1. However, such a trend was not observed with
CDK4, cyclin E1 and CDK2. In qPCR analysis of cyclin D1, CDK4,
cyclin E1 and CDK2 mRNA levels, cyclin D1 was identified as a
sensitive responder to LINC01296 manipulation (Fig. 4B). Spearman's analysis indicated a
strong positive correlation between LINC01296 levels and cyclin D1
levels (Fig. 4C).
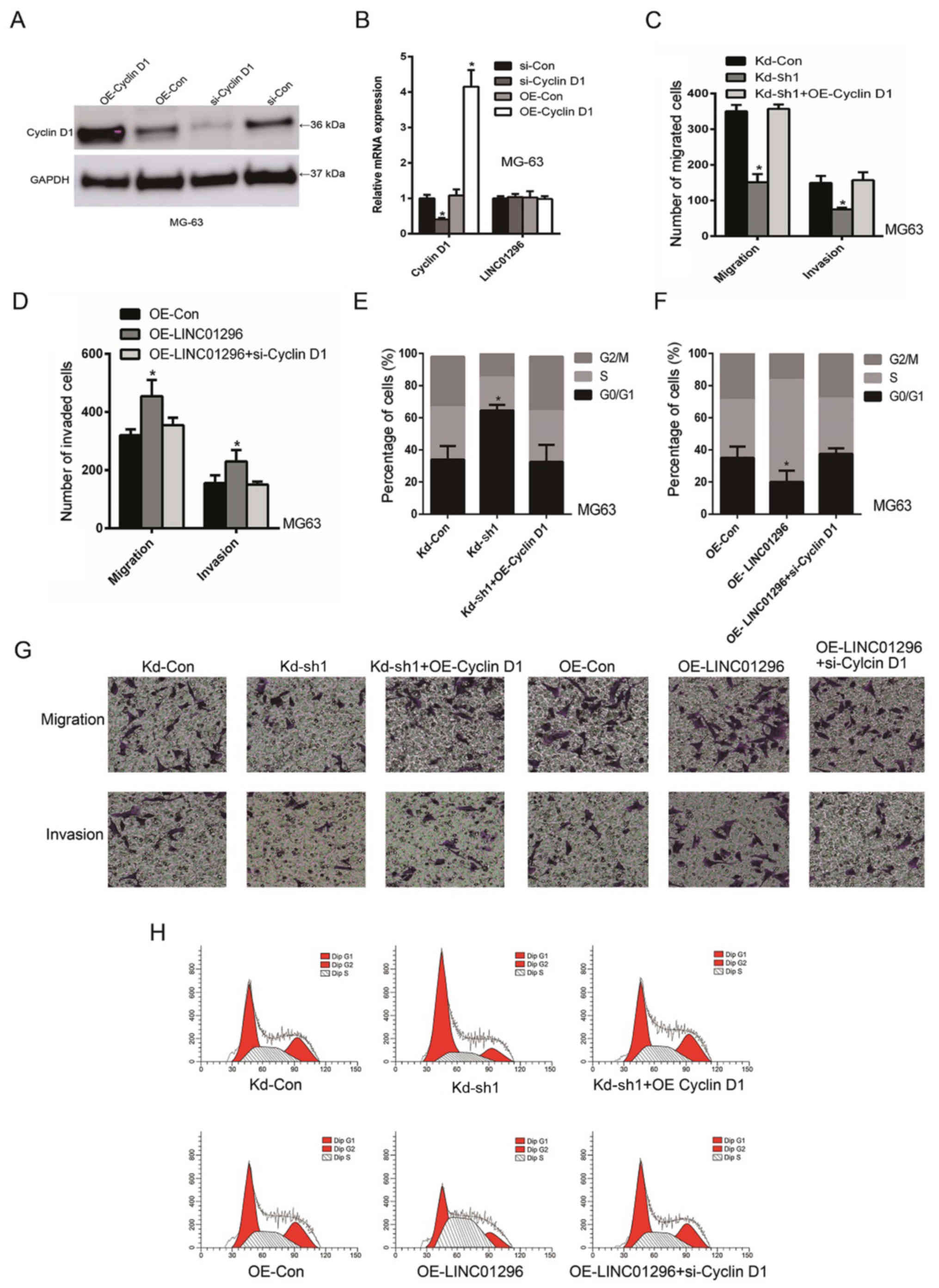
To confirm the interaction between cyclin D1 and
LINC01296, a direct knockdown or overexpression of cyclin D1 was
carried out to antagonize the effect of LINC01296 in promoting
osteosarcoma. Western blot (Fig.
5A) and qRT-PCR analyses (Fig.
5B) demonstrated successful cyclin D1 overexpression and
knockdown, respectively. In addition, as revealed in Fig. 5C and H, knockdown of LINC01296
decreased cell migration and invasion, while cyclin D1
overexpression increased cell migration and invasion. Consistently,
while overexpression of LINC01296 increased cell migration and
invasion, knockdown of cyclin D1 decreased cell migration and
invasion in MG63 cells (Fig. 5D).
Further study revealed that while knockdown of LINC01296 induced
cell cycle arrest, cyclin D1 overexpression reduced cell cycle
arrest (Fig. 5E and H).
Consistently, while overexpression of LINC01296 reduced the number
of cells at the G0/G1 phase, knockdown of cyclin D1 concurrently
led to the increase in cell cycle arrest. (Fig. 5D and H). This evidence demonstrated
that the interaction between LINC01295 and cyclin D1 was
responsible for the tumor-promoting role of LINC01296.
Discussion
Accumulating evidence has demonstrated that
non-protein coding genes are important regulators of cancer. With
the aim of unraveling the molecular mechanism of osteosarcoma, the
tumor-promoting role of LINC01296 in osteosarcoma was investigated
for the first time in the present study. The results revealed that
malignant osteosarcoma was characterized by high LINC01296
expression, which was consistent with the oncogenic function of
LINC01296 in other cancers (13,14,19).
Furthermore, it was revealed in this study that high LINC01296
expression played an indispensable role in promoting the
proliferation, migration and invasion of osteosarcoma cells.
Evidently, knockdown of LINC01296 resulted in retarded
proliferation, migration and invasion, and marked cell cycle
arrest. To elucidate the molecular mechanism of LINC01296 in
osteosarcoma, we investigated how LINC01296 expression affected
cell cycle regulators. Cyclins are key regulators of CDKs
(cyclin-dependent kinases). Distinct expression and degradation
patterns of different cyclins lead to the temporal coordination of
mitotic events. Cyclin D1 regulates CDK3 and CDK6 by forming a
complex with them, thereby governing cell cycle G1/S transition.
Cyclin D1 serves as a central regulator for cell cycle progression,
and aberrant expression of this protein is a significant
contributor to tumorigenesis (20).
We demonstrated that the expression of cyclin D1 was found to be
positively correlated with LINC01296 levels. Knockdown of LINC01296
led to marked downregulation of cyclin D1. We reviewed the
potential pathways in which LINC01296 was involved, but there was
no clear elaboration of these pathways in published research.
However, we found that there were some related micro-RNAs or
proteins including miR-21a, miR-122, miR-5095 and MMP9, and MMPs
that were reported to play an important role in the cell cycle
(19,21–23).
The potential pathways included miR-21a/p27, p38MAPK/Sp-1/p21 and
Skp2/p27/p21. However, the direct interaction between LINC01296 and
cyclin D1 was not established in this study. Our ongoing research
aims to address the clear mechanism of LINC01296 and cyclin D1. But
given the involvement of cyclin D1 in numerous oncogenic signaling
pathways (24), it can be concluded
that LINC01296 is a vital molecule for a large spectrum of
oncogenic processes.
Diagnosis and therapy of osteosarcoma still remain a
critical challenge nowadays. The upregulation of LINC01296 in
osteosarcoma makes it a promising biomarker for sensitive detection
of such a disease. In addition to the increased expression of
lncRNAs in tumors, increased lncRNA levels can also be detected in
serum, which enhances the development of lncRNA-based facile cancer
detection methods (25). Validation
of the diagnostic potential of serum LINC01296 in osteosarcoma
diagnosis is warranted. The compromised proliferation, migration
and invasion induced by LINC01296 knockdown also potentiates gene
therapy strategies based on this approach. For example, in
vivo delivery of siRNA for silencing tumor-promoting lncRNAs
has been utilized to impede cancer progression (26–28).
Our ongoing studies focus on evaluation of the therapeutic efficacy
of LINC01296 knockdown in vivo. This approach could
precisely control the expression of LINC01296, thereby suppressing
the proliferation, migration and invasion of tumors. Moreover, this
proposed approach obviates the side effects associated with
conventional cancer therapies such as chemotherapy and radiation
therapy (29). It would not be
surprising that cancer therapeutic approaches based on lncRNA
knockdown could offer substantial benefit to the clinical
management of osteosarcoma.
In summary, the present study reported that high
LINC01296 expression was found in osteosarcoma. LINC01296
expression was associated with the proliferation, migration, and
invasion of osteosarcoma cells. In this study, positive correlation
between the upregulation of cyclin D1 and high LINC01296 expression
was reported, which in part accounts for the tumor-promoting role
of LINC01296 in osteosarcoma. These data demonstrated that
LINC012967 can be a valuable target for the diagnosis and treatment
of osteosarcoma.
Acknowledgements
Not applicable.
Funding
We would like to thank the Jilin Provincial Science
and Technology Department Outstanding Youth Talent Fund (no.
20170520014JH) and the National Natural Science Foundation (no.
81201382) for their financial support.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
PL, the corresponding author of this study,
participated in every step of the design project and in specific
experiments. XY also participated in the design of this study and
most of the experiments, and was the writer of this manuscript. LP
participated in the collection of case data and cultured the cells
for this study, and provided advice for the revision of the
manuscript. TY helped in the experiments and modified the language
of the manuscript. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of Jilin University (Changchun, China). Informed
consent was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rinn JL and Chang HY: Genome regulation by
long noncoding RNAs. Annu Rev Biochem. 81:145–166. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu J, Jung C, Xu J, Wang H, Deng S,
Bernad L, Arenas-Huertero C and Chua NH: Genome-wide analysis
uncovers regulation of long intergenic noncoding RNAs in
Arabidopsis. Plant Cell. 24:4333–4345. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Schmitt AM and Chang HY: Long noncoding
RNAs in cancer pathways. Cancer Cell. 29:452–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liang WC, Fu WM, Wong CW, Wang Y, Wang WM,
Hu GX, Zhang L, Xiao LJ, Wan DC, Zhang JF, et al: The lncRNA H19
promotes epithelial to mesenchymal transition by functioning as
miRNA sponges in colorectal cancer. Oncotarget. 6:22513–22525.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li H, Yu B, Li J, Su L, Yan M, Zhu Z and
Liu B: Overexpression of lncRNA H19 enhances carcinogenesis and
metastasis of gastric cancer. Oncotarget. 5:2318–2329. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bhan A and Mandal SS: LncRNA HOTAIR: A
master regulator of chromatin dynamics and cancer. Biochim Biophys
Acta. 1856:151–164. 2015.PubMed/NCBI
|
|
7
|
Prensner JR and Chinnaiyan AM: The
emergence of lncRNAs in cancer biology. Cancer Discov. 1:391–407.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang KC and Chang HY: Molecular mechanisms
of long noncoding RNAs. Mol Cell. 43:904–914. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li W, Xie P and Ruan WH: Overexpression of
lncRNA UCA1 promotes osteosarcoma progression and correlates with
poor prognosis. J Bone Oncol. 5:80–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun XH, Yang LB, Geng XL, Wang R and Zhang
ZC: Increased expression of lncRNA HULC indicates a poor prognosis
and promotes cell metastasis in osteosarcoma. Int J Clin Exp
Pathol. 8:2994–3000. 2015.PubMed/NCBI
|
|
12
|
Zhang Q, Geng PL, Yin P, Wang XL, Jia JP
and Yao J: Down-regulation of long non-coding RNA TUG1 inhibits
osteosarcoma cell proliferation and promotes apoptosis. Asian Pac J
Cancer Prev. 14:2311–2315. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu J, Cheng G, Zhang C, Zheng Y, Xu H,
Yang H and Hua L: Long noncoding RNA LINC01296 is associated with
poor prognosis in prostate cancer and promotes cancer-cell
proliferation and metastasis. Onco Targets Ther. 10:1843–1852.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qiu JJ and Yan JB: Long non-coding RNA
LINC01296 is a potential prognostic biomarker in patients with
colorectal cancer. Tumor Biol. 36:7175–7183. 2015. View Article : Google Scholar
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Song LN: Effects of retinoic acid and
dexamethasone on proliferation, differentiation, and glucocorticoid
receptor expression in cultured human osteosarcoma cells. Oncol
Res. 6:111–118. 1994.PubMed/NCBI
|
|
17
|
Jiang C, Fang X, Zhang H, Wang X, Li M,
Jiang W, Tian F, Zhu L and Bian Z: AMD3100 combined with triptolide
inhibit proliferation, invasion and metastasis and induce apoptosis
of human U2OS osteosarcoma cells. Biomed Pharmacother. 86:677–685.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wen J, Wang L, Zhang M, Xie D and Sun L:
Repression of telomerase activity during in vitro differentiation
of osteosarcoma cells. Cancer Invest. 20:38–45. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Seitz AK, Christensen LL, Christensen E,
Faarkrog K, Ostenfeld MS, Hedegaard H, Nordentoft I, Nielsen MM,
Palmfeldt J, Thomson M, et al: Profiling of long non-coding RNAs
identifies LINC00958 and LINC01296 as candidate oncogenes in
bladder cancer. Sci Rep. 7:3952017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee Y, Dominy JE, Choi YJ, Jurczak M,
Tolliday N, Camporez JP, Chim H, Lim JH, Ruan HB, Yang X, et al:
Cyclin D1-Cdk4 controls glucose metabolism independently of cell
cycle progression. Nature. 510:547–551. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Qin QH, Yin ZQ, Li Y, Wang BG and Zhang
MF: Long intergenic noncoding RNA 01296 aggravates gastric cancer
cells progress through miR-122/MMP-9. Biomed Pharmacother.
97:450–457. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang K, Zhang M, Wang C and Ning X: Long
noncoding RNA LINC01296 harbors miR-21a to regulate colon carcinoma
proliferation and invasion. Oncol Res. Apr 19–2018.(Epub ahead of
print). View Article : Google Scholar
|
|
23
|
Zhang D, Li H, Xie J, Jiang D, Cao L, Yang
X, Xue P and Jiang X: Long noncoding RNA LINC01296 promotes tumor
growth and progression by sponging miR-5095 in human
cholangiocarcinoma. Int J Oncol. 52:1777–1786. 2018.PubMed/NCBI
|
|
24
|
Qie S and Diehl JA: Cyclin D1, cancer
progression, and opportunities in cancer treatment. J Mol Med
(Berl). 94:1313–1326. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhou X, Yin C, Dang Y, Ye F and Zhang G:
Identification of the long non-coding RNA H19 in plasma as a novel
biomarker for diagnosis of gastric cancer. Sci Rep. 5:115162015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Williams GT and Pickard MR: Long
non-coding RNAs: New opportunities and old challenges in cancer
therapy. Transl Cancer Res. 5 Suppl:S564–S566. 2016. View Article : Google Scholar
|
|
27
|
Li Han C and Chen Y: Small and long
non-coding RNAs: Novel targets in perspective cancer therapy. Curr
Genomics. 16:319–326. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fatima R, Akhade VS, Pal D and Rao SM:
Long noncoding RNAs in development and cancer: Potential biomarkers
and therapeutic targets. Mol Cell Ther. 3:52015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li S, Sun W, Wang H, Zuo D, Hua Y and Cai
Z: Research progress on the multidrug resistance mechanisms of
osteosarcoma chemotherapy and reversal. Tumor Biol. 36:1329–1338.
2015. View Article : Google Scholar
|