Introduction
Glioblastoma is the most common malignant tumor of
the nervous system; it is highly invasive, resistant to treatment
and relapses easily (1). Numerous
studies have focused on glioblastoma in relation to methylation
status of a panel of genes (2),
angiogenesis and DNA repair pathways (3,4),
epidermal growth factor receptor, p53, isocitrate dehydrogenase 1
and MDM2 mutation status (5), and
various therapeutic strategies (6,7);
however, the overall survival of patients remains poor, with a
median survival time of 12–15 months (8). Therefore, further investigations of
the mechanisms and potential therapeutic strategies for
glioblastoma are necessary.
Epsin (EPN)-3 is a member of the endocytosis adapter
protein gene family, which contains several different interaction
motifs, and is involved in clathrin-mediated endocytosis (9,10). The
Epsin protein family is conserved between yeast, Drosophila
melanogaster and mammals. In mammals, the Epsin family includes
EPN1, EPN2 and EPN3. EPN1 and EPN2 are ubiquitously expressed, to
various degrees, in different tissues, particularly in the brain
(11,12). Previous studies have reported that
EPN1 and EPN2 may be upregulated in a variety of cancer types and
are associated with tumor cell proliferation, migration and
invasion (11,12), whereas EPN3 expression is normally
limited to gastric parietal cells (13), as well as wounded or pathological
tissue, rather than normal brain tissue (14). It has been reported that EPN3 is
overexpressed in basal cell carcinoma and ulcerative colitis
(13,14). In addition, several studies have
suggested that Epsins, as adapter proteins in clathrin-mediated
endocytosis, are involved in the regulation of various signal
receptors, including receptor tyrosine kinases, Notch ligands and
receptors, and WNT receptors; defective regulation of these
signaling ligands and receptors is associated with tumorigenesis
(15–17).
Notch signaling regulates tissue renewal and
development, as well as serving various roles in cell
differentiation, proliferation and apoptosis in a tissue
context-dependent manner (18).
Aberrant loss or gain of Notch signaling components is associated
with multiple human diseases (19–21).
Activation of Notch signaling requires its receptors to bind their
specific ligands, resulting in receptor proteolysis and the release
of the active Notch intracellular domain (NICD), which translocates
to the nucleus to initiate downstream target gene transcription
(22). A previous study has
suggested that Epsins are required for Notch signaling pathway
activation (9).
WNT/β-catenin signaling is also important in tumor
development. WNT signaling is activated through endocytosis
(17,23,24),
and Epsins are required for the stability of the downstream protein
Dishevelled and WNT signaling activation in colon cancer
development (17).
Notch and WNT signaling pathways are associated with
epithelial-mesenchymal transition (EMT) (25–27).
EMT is a vital mechanism in embryonic development and tissue
repair, and is an essential mechanism of cancer cell metastasis
(28). The characteristics of EMT
include the loss of the expression or function of epithelial
(E)-cadherin with a concomitant increase in mesenchymal markers,
including Vimentin (VIM) and Neural-cadherin (29), potentially leading to the spread and
dissemination of cancer cells. EMT may be induced by several
transcription factors, including Snail1, Slug and Twist (25).
In the present study, EPN3 was revealed to be highly
expressed in high-grade glioma tissues compared with low-grade
samples. The effects of EPN3 on glioblastoma cell migration and
invasion were observed by inducing the gain and loss of EPN3
expression in U87 and U251 glioblastoma cells; the results revealed
that the overexpression of EPN3 promoted glioblastoma cell
migration and invasion, and increased the expression of NICD1,
β-catenin, VIM, Slug, Twist and zinc-finger E-box-binding homeobox
(ZEB)-1, whereas the expression level of E-cadherin was decreased.
These results suggested that EPN3 may promote glioblastoma cell
migration and invasion by inducing EMT, and that EPN3 may be a
potential therapeutic target in glioblastoma.
Materials and methods
Tumor samples and cell culture
Glioma tissues from four different World Health
Organization (WHO) grades were collected from 167 patients at the
Department of Neurosurgery at Tianjin Huanhu Hospital (Tianjin,
China), and included 71 women and 96 men (age 1–73 years; Table I); human colon carcinoma tissue was
donated by Professor Li Qi (Tianjin Medical University Cancer
Institute and Hospital). Informed consent was obtained from all
patients or their guardian, and the use of patient tissue was
approved by the ethics committee of Tianjin Huanhu Hospital
(2016–3). Glioblastoma cell lines
A172, LN229, LN308, U251 and U87 were purchased from the American
Type Culture Collection (Genetimes ExCell Technology, Inc.,
Shanghai, China). Cells were cultured in Dulbecco's modified
Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc.), and maintained in a
humidified atmosphere with 5% CO2 at 37°C.
 | Table I.Relationship between EPN3 expression
and clinicopathological features in patients with glioma. |
Table I.
Relationship between EPN3 expression
and clinicopathological features in patients with glioma.
|
|
| EPN3
expression |
|
|
|---|
|
|
|
|
|
|
|---|
| Clinicopathological
feature | n | Low | High | Positive rate
(%) |
P-valuea |
|---|
| WHO grade |
| I | 49 | 41 | 8 | 17.7 | <0.001 |
| II | 47 | 38 | 9 |
|
|
|
III | 36 | 19 | 17 | 52.1 |
|
| IV | 35 | 15 | 20 |
|
|
| Sex |
|
|
|
| 0.322 |
|
Male | 96 | 62 | 34 | 35.4 |
|
|
Female | 71 | 51 | 20 | 28.2 |
|
| Age (years) |
|
<45 | 90 | 62 | 28 | 31.1 | 0.715 |
|
≥45 | 77 | 51 | 26 | 33.8 |
|
| Tumor size
(cm) |
|
<4.5 | 121 | 82 | 39 | 32.2 | 0.963 |
|
≥4.5 | 46 | 31 | 15 | 32.6 |
|
Plasmid vectors and transfection
All plasmids were purchased from OriGene
Technologies, Inc. (Rockville, MD, USA). The pGFP-C-shLenti vector
encoding short hairpin (sh)RNA against EPN3 (shEPN3; cat. no.
TL304750; 5′-CACAACTACTCCGAGGCAGAAATCAAGGT-3′) was used to silence
EPN3 expression; Scrambled-shRNA (cat. no. TR30021;
5′-GCACTACCAGAGCTAACTCAGATAGTACT-3′) was used as negative control.
pCMV6-EPN3 (cat. no. RC212297) was used for the upregulation of
EPN3; pCMV6-Entry (cat. no. PS100001) was used as negative control.
Cells were seeded at a density of 2×105 cells/well in
6-well culture plates, and plasmid transfections (2,500 ng) were
performed using Lipofectamine® 2000 transfection reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's instructions, when cell cultures reached 90%
confluency; cells were incubated in a humidified atmosphere with 5%
CO2 at 37°C for 48 h. Gene expression levels were
evaluated by reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) and western blot analysis at 48 h
post-transfection.
RT-qPCR
Total RNA was extracted from the transfected cells
(5×106) using 1 ml TRIzol reagent (Invitrogen; Thermo
Fisher Scientific, Inc.), according to the manufacturer's protocol.
cDNA was synthesized using a cDNA Reverse Transcription Kit
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. qPCR was performed using Power SYBR Green
PCR master mix (Applied Biosystems; Thermo Fisher Scientific, Inc.)
and an ABI 7500 series Real-Time PCR machine (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The thermocycling condition was as
follows: Initial denaturation at 95°C (30 sec), followed by 40
cycles of denaturation at 95°C (20 sec), annealing at 60°C (20 sec)
and elongation at 72°C (20 sec). mRNA expression levels were
normalized to GAPDH and were calculated using the 2−ΔΔCt
method (30). The primers used for
PCR are listed in Table II.
| Table II.Primer sequences used for reverse
transcription-quantitative polymerase chain reaction. |
Table II.
Primer sequences used for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Sequence
(5′→3′) |
|---|
| EPN3 | F:
CTTGGCTGACATCTTCGTACCT |
|
| R:
TGTGTTCGGCCTAAAACCTG |
| E-cadherin | F:
CAGCACGTACACAGCCCTAA |
|
| R:
ACCTGAGGCTTTGGATTCCT |
| Vimentin | F:
AGATGGCCCTTGACATTGAG |
|
| R:
TGGAAGAGGCAGAGAAATCC |
| Notch1 | F:
ACCAATACAACCCTCTGCGG |
|
| R:
GGCCCTGGTAGGTCATCATC |
| Notch2 | F:
GGAGGCACCTGTATTGACCT |
|
| R:
ATGCCCTGGATGGAAAATGGA |
| Notch3 | F:
CTGTGGCCCTCATGGTATCT |
|
| R:
ACCGTTCAGGCATGGGTTG |
| Notch4 | F:
TTCCCAGAACCCTGTGCCAAT |
|
| R:
AACTGGCACGTCTCACCCAG |
| Snail1 | F:
CCTCCCTGTCAGATGAGGAC |
|
| R:
GTTCCTTATGGAGTCGGACC |
| Slug | F:
GGGGAGAAGCCTTTTTCTTG |
|
| R:
AGGACGTGTTTGTACTCCT |
| Twist | F:
GGAGTCCGCAGTCTTACGAG |
|
| R:
TCTGGAGGACCTGGTAGAGG |
| ZEB1 | F:
GCACAACCAAGTGCAGAAGA |
|
| R:
GCCTGGTTCAGGAGAAGATG |
| ZEB2 | F:
AAATGCACAGAGTGTGGCAAGG |
|
| R:
CTGCTGATGTGCGAACTGTAGGA |
| GAPDH | F:
CAATGACCCCTTCATTGACCT |
|
| R:
ATGACAAGCTTCCCGTTCTC |
Antibodies
Anti-EPN3 was purchased from Sigma-Aldrich (cat. no.
SAB1303019; Merck KGaA, Darmstadt, Germany); anti-GAPDH was
purchased from ProteinTech Group, Inc. (cat. no. 10494-1-AP;
Chicago, IL, USA); anti-E-cadherin (cat. no. sc-8426), anti-Slug
(cat. no. sc-166902), anti-Snail 1 (cat. no. sc-393172), anti-ZEB2
(cat. no. sc-271984) and anti-Twist (cat. no. sc-81417) were from
Santa Cruz Biotechnology, Inc. (Dallas, TX, USA); anti-VIM was from
BioLegend, Inc. (cat. no. 677801; San Diego, CA, USA); and
anti-NICD1 (cat. no. WL03097a), anti-ZEB1 (cat. no. WL01657) and
anti-β-catenin (cat. no. WL0962a) were from Wanleibio Co., Ltd.
(Shanghai, China). The horseradish peroxidase (HRP)-conjugated goat
anti-mouse (cat. no. ZB-2305) and HRP-conjugated goat anti-rabbit
(ZB-2301) secondary antibodies were purchased from ZSGB-BIO
(OriGene Technologies, Inc.).
Western blot analysis
Total protein was extracted from cells
(1×107) using Radioimmunoprecipitation Assay lysis
buffer (Beyotime Institute of Biotechnology, Shanghai, China) on
ice. Protein concentrations were determined by Bicinchoninic Acid
assay (Beijing Solarbio Science & Technology Co., Ltd.,
Beijing, China). Equal amounts of protein (30 µg) were separated
using SDS-PAGE and transferred onto polyvinylidene fluoride
membranes (EMD Millipore, Billerica, MA, USA). Membranes were
blocked with 5% skim milk in TBST (1X TBS + 0.1% Tween-20) buffer
for 2 h. Following washing 3 times for 5 min in TBST, membranes
were incubated with primary antibodies against EPN3, E-cadherin,
Slug, Twist (all at 1:500), VIM (1:5,000), NICD1 (1:500), β-catenin
(1:1,000) and GAPDH (1:10,000) overnight at 4°C. Membranes were
washed 3 times (5 min each) in TBST, and subsequently incubated
with the corresponding horseradish peroxidase-conjugated secondary
antibodies (1:1,000) for 1 h at room temperature. Protein bands
were visualized using an Enhanced Chemiluminescence Western Blot
Detection kit, and the intensity of protein bands were normalized
to GAPDH and quantified by densitometric analysis using Quantity
One software (version 4.6.9; Bio-Rad Laboratories, Inc., Hercules,
CA, USA).
Immunohistochemistry (IHC)
Human colon carcinoma tissues were used as a
positive control. Colon carcinoma and glioma tissue samples were
fixed using 10% formalin at room temperature for 2 h, embedded in
paraffin and sectioned (4 µm), slides were deparaffinized with
xylene and rehydrated in a decreasing ethanol series. Endogenous
peroxidase activity was blocked by incubating the slides in 3%
hydrogen peroxide for 15 min. Following heat-induced antigen
retrieval at 100°C for 10 min, sections were incubated with 5%
rabbit serum (ZSGB-BIO; OriGene Technologies, Inc.) for 10 min to
eliminate non-specific binding. Anti-EPN3 (1:200) was added and the
sections were incubated overnight at 4°C. The sections were washed
3 times (5 min each) using PBS, and the corresponding optimized
secondary antibodies (Bond Polymer refine detection; cat. no.
DS9800; Leica Microsystems, Ltd., Milton Keynes, UK) were added to
the sections and incubated for 1 h at room temperature, followed by
incubation with the streptavidin-HRP complex (Santa Cruz
Biotechnology, Inc.). Immunoreactivity was visualized by incubating
with 3,3′-diaminobenzidine (Sigma-Aldrich; Merck KGaA) for 3–15
min, and the sections were counterstained with hematoxylin. Two
pathologists blinded to the clinical data independently scored the
percentage of positive-staining cells and the intensity of the EPN3
staining. A three-tiered scale was used to grade the staining
intensity (1, negative or weak; 2, medium; 3, strong), whereas the
percentage of EPN3-positive tumor cells was scored as follows: 0,
≤10% positive tumor cells; 1, 11–24%; 2, 25–50%; 3, 51–75% and 4,
>75%. The product of the intensity and positive percentage score
of the cells was used as a staining index (negative, 4; positive,
≥4).
Transwell migration and invasion
assays
Migration and invasion assays were performed using
24-well Transwell chambers either with (invasion) or without
(migration) BD BioCoat Matrigel (BD Biosciences, Franklin Lakes,
NJ, USA). A total of 5×104 cells were suspended in
serum-free DMEM and seeded into the upper chamber; 500 µl DMEM with
10% FBS was added to the lower chamber. Following incubation for 24
h at 37°C, the non-migratory or non-invasive cells remaining in the
upper chambers were gently removed with a cotton swab. The cells
that had migrated or invaded to the lower surface of the membrane
were washed twice with PBS, fixed with 4% paraformaldehyde for 30
min and stained with 0.1% crystal violet for 15 min. Cells were
counted in five different fields of view under a light
microscope.
Statistical analysis
All experiments were performed in triplicate.
Normally distributed data were presented as the mean ± standard
deviation, and were analyzed using Student's t-test in SPSS version
17 (SPSS Inc., Chicago, IL, USA) and GraphPad Prism 5 (GraphPad
Software, Inc., La Jolla, CA, USA). The χ2 test was used
to analyze categorical variables. P<0.05 was considered to
indicate a statistically significant difference.
Results
EPN3 expression levels are higher in
high-grade tumor samples
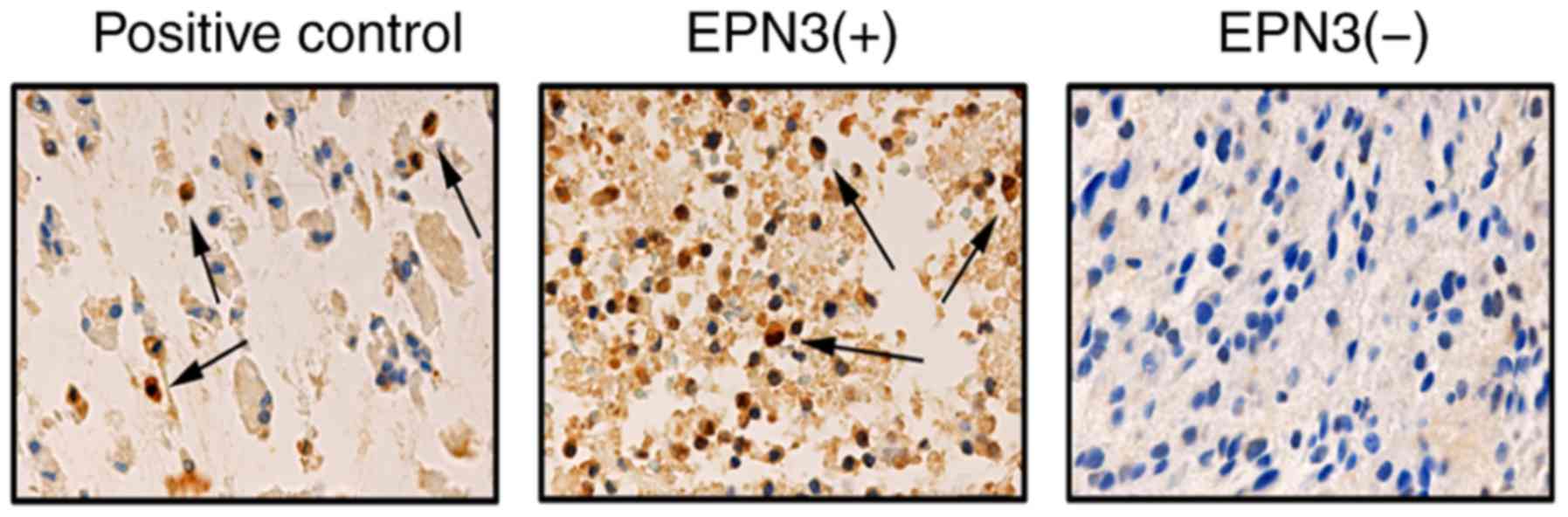
EPN3 expression was examined by IHC in tissue
samples from 167 patients with glioma, including 96 low-grade (WHO
grade I/II) and 71 high-grade (WHO grade III/IV) tumor samples
(Fig. 1; Table I). The results demonstrated that
EPN3 was highly expressed in 17 out of 96 low-grade glioma tissues
(17.7%), and 37 out of 71 high-grade glioma tissues (52.1%), and
the difference in EPN3 expression levels between low- and
high-grade glioma tissue samples was significant (P<0.001;
Table I). No significant
differences were identified in EPN3 expression based on patient
age, sex or tumor size (P>0.05; Table I).
Overexpression of EPN3 enhances
migration and invasion of U87 and U251 glioblastoma cells
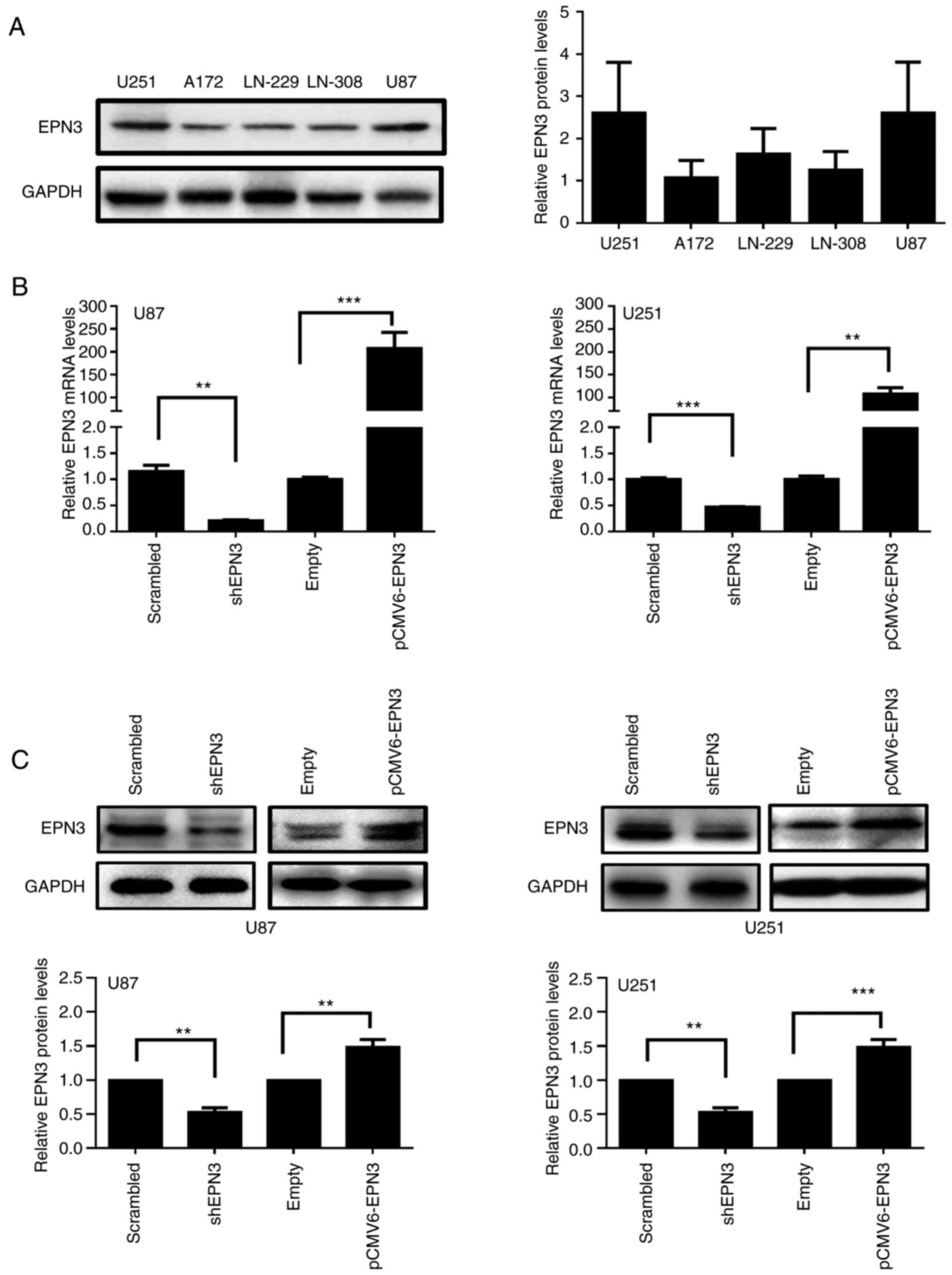
Varying EPN3 protein expression levels were detected
among the five glioblastoma cell lines examined (A172, LN229, U251,
LN308 and U87), with the highest expression levels detected in U251
and U87 cells, and the lowest level in A172 cells (Fig. 2A). U251 and U87 cells were selected
for further experiments. Notably, although U87 is different from
the original glioblastoma cells, Marie Allen et al have
reported that U87 cell line is of CNS origin and is likely to be a
bona fide human glioblastoma cell line with unknown patient origin
(31). U251 and U87 cells
transfected with shEPN3 exhibited reduced EPN3 mRNA and protein
expression levels compared with cells transfected with
Scrambled-shRNA (Fig. 2B and C). In
addition, cells transfected with EPN3 overexpression vector
exhibited increased levels of EPN3 mRNA and protein expression
compared with expression levels empty vector-transfected cells
(Fig. 2B and C).
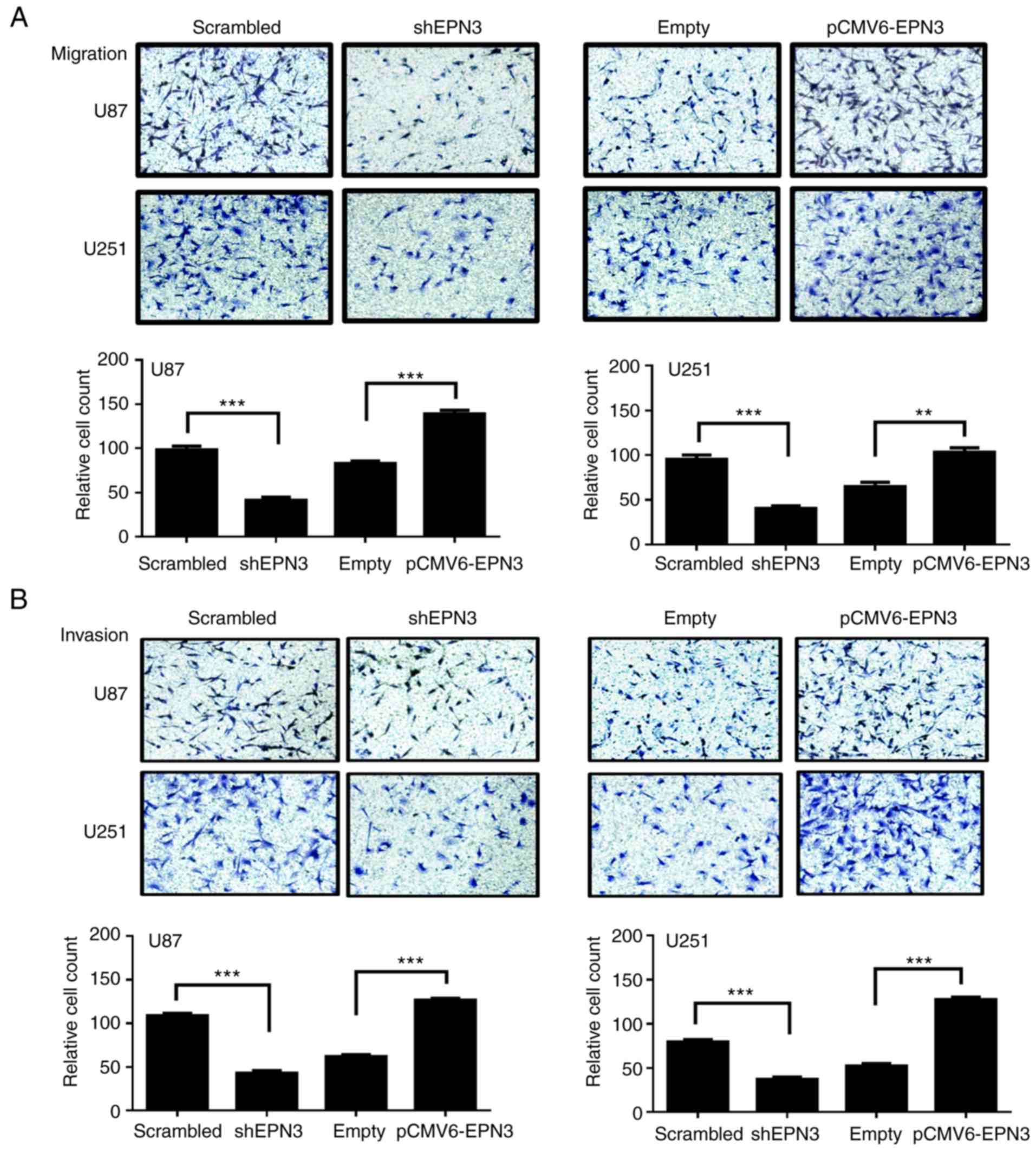
The effects of EPN3 on glioblastoma cell migration
and invasion were examined using Transwell migration and invasion
assays. The results demonstrated that the reduction of EPN3
expression significantly inhibited U87 and U251 glioblastoma cell
migration and invasion compared with the control cells (P<0.001;
Fig. 3A and B, respectively).
Conversely, the overexpression of EPN3 significantly promoted U87
and U251 cell migration and invasion compared with control cells
(P<0.01, P<0.001; in Fig. 3).
These results indicated that EPN3 overexpression enhanced
glioblastoma cell migration and invasion in vitro.
Overexpression of EPN3 induces EMT in
U87 and U251 glioblastoma cells
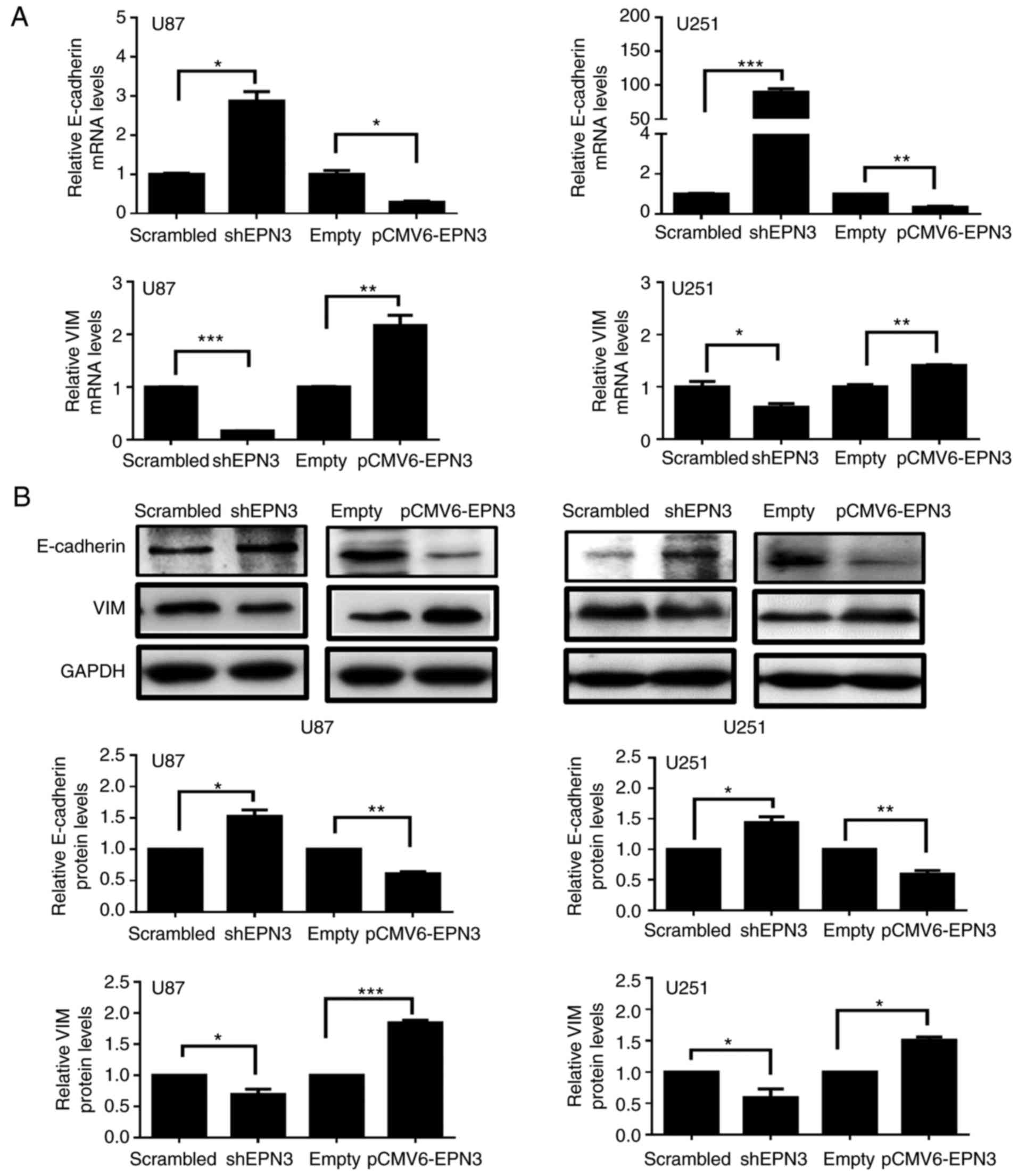
EMT serves an important role in the process of tumor
cell migration and invasion (32);
therefore, expression levels of key molecular markers of EMT,
E-cadherin and VIM, were analyzed by RT-qPCR and western blotting
in transfected U87 and U251 cells. U87 and U251 cells transfected
with shEPN3 exhibited increased levels of E-cadherin mRNA and
protein expression (Fig. 4A and B),
whereas the expression levels of VIM decreased. Conversely,
EPN3-overexpressing U87 and U251 cells exhibited decreased
E-cadherin mRNA and protein expression levels (Fig. 4A and B, respectively), whereas the
mRNA and protein expression levels of VIM were increased. These
results indicated that EPN3 may be associated with EMT, which may
contribute to the migratory and invasive ability of glioblastoma
cells.
Overexpression of EPN3 increases
Notch1 and β-catenin expression in U87 and U251 glioblastoma
cells
The Notch pathway is activated in glioblastoma cells
and promotes their migration and invasion (33,34);
EPN1 and EPN2 were reported to be adaptor proteins in
clathrin-mediated endocytosis, which is necessary for the
activation of the Notch pathway (9,35–37).
Therefore, whether EPN3 was involved in the activation of the Notch
pathway in glioblastoma cells was examined. RT-qPCR analysis was
used to identify the effects on Notch receptor expression following
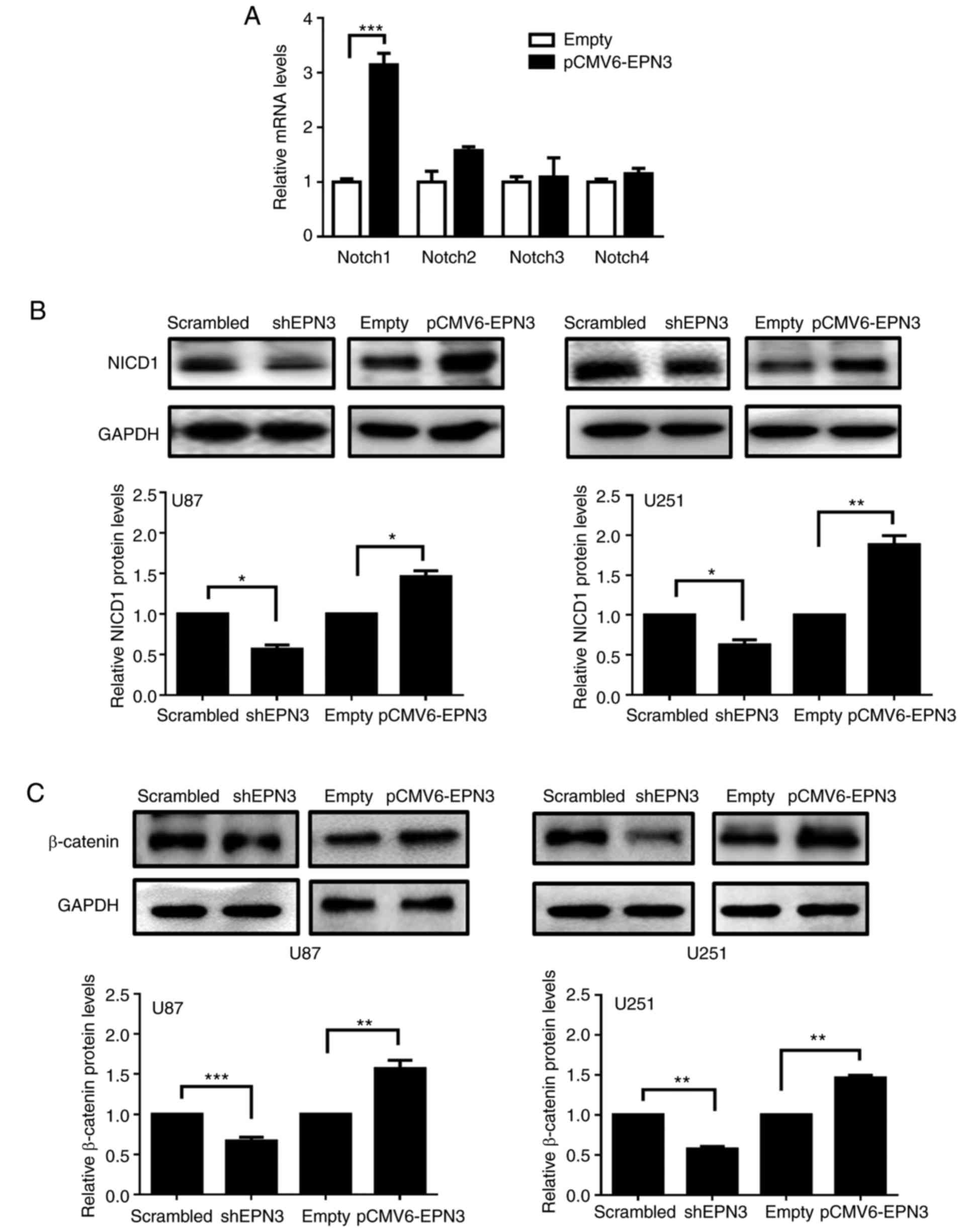
the overexpression of EPN3 in U87 cells. The results suggested that
Notch1 expression increased when EPN3 was overexpressed, whereas no
significant differences were observed in Notch2, Notch3 or Notch4
expression (Fig. 5A). As Notch1 is
expressed in primary human gliomas and glioblastoma cell lines
(18), the expression of the
activated NICD1 was examined by western blotting in transfected U87
and U251 glioblastoma cells. It was demonstrated that the
expression levels of NICD1 decreased when EPN3 was downregulated
and increased when EPN3 was overexpressed in U87 and U251 cell
lines compared with the respective control-transfected cells
(Fig. 5B). WNT/β-catenin signaling
is also activated via endocytosis (23,24),
and it was previously reported that Epsin is required for WNT
signaling activation in colon cancer development (17). In the present study, the expression
of β-catenin was decreased when EPN3 was downregulated (Fig. 5C). In conclusion, EPN3 may function
to activate Notch1 and WNT signaling pathways to promote
tumorigenesis in glioblastoma.
Overexpression of EPN3 increases Slug,
Twist and ZEB1 expression levels in U87 and U251 glioblastoma
cells
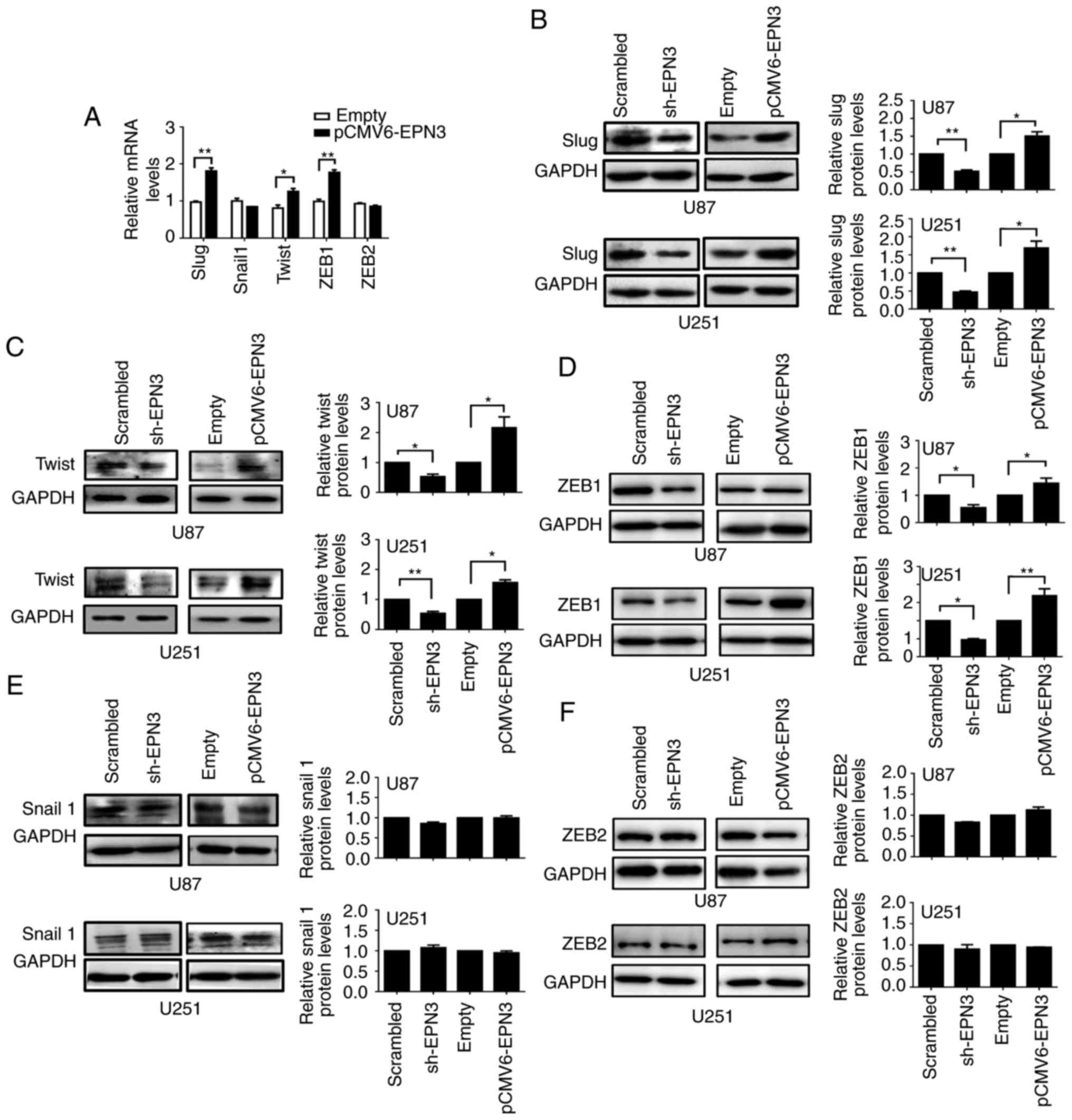
Notch activation induces EMT through the
Slug-induced repression of E-cadherin (38) and the upregulation of EMT-inducing
transcription factors, including Snail1, Slug, ZEB1, ZEB2 and Twist
(25). Whether EPN3 activated these
EMT transcription factors was assessed using RT-qPCR and western
blotting (Fig. 6). The results
demonstrated that Slug, Twist and ZEB1 mRNA expression levels were
increased in cells overexpressing EPN3 compared with the respective
levels in cells transfected with empty vector (Fig. 6A); Snail1 and ZEB2 mRNA expression
levels remained unaffected. Conversely, protein expression levels
of Slug, Twist and ZEB1 were decreased in shEPN3-transfected U87
and U251 cells, compared with Scramble shRNA-transfected cells,
whereas their expression levels were increased when EPN3 was
overexpressed (Fig. 6B-D)
Conversely, Snail1 and ZEB2 expression levels were unaltered
(Fig. 6E and F, respectively).
These results suggested that EPN3 may increase the transcription of
the EMT transcription factors Slug, Twist and ZEB1, rather than
Snail-1 and ZEB2, to induce EMT in glioblastoma cells.
Discussion
The Epsin protein family may promote oncogenesis and
cancer progression (11); EPN1 and
EPN2 have been associated with various types of tumor (39–41),
whereas studies regarding EPN3 are limited. A recent study reported
that the high expression levels of EPN3 were associated with an
increased risk of metastasis in estrogen receptor-positive breast
cancer and non-small cell lung cancer (42). Other studies have suggested that
EPN3 promotes cellular senescence and is associated with the p53
pathway (43,44). The present study demonstrated that
EPN3 was expressed in glioma tissues and glioblastoma cell lines.
The results also revealed that the overexpression of EPN3 promoted
glioblastoma cell migration and invasion, which indicated that EPN3
may serve as a potential target for glioblastoma therapy.
Relatively little is known about the role of EPN3 in
the endosome. Howe and Mobley proposed the signaling endosome
hypothesis as a cellular mechanism for distance communication
(45); they suggested that
activated Tyrosine kinase receptor A and other signaling proteins
were internalized into an endocytic organelle, which was then
retrograde-transported from the axon tip to the neuron cell body.
In addition, Vanlandingham et al reported that EPN1 serves
an important endosomal role in the efficient retrograde transport
of bone morphogenetic protein (BMP) signaling endosomes into motor
neuron nuclei, and that EPN1 may negatively regulate BMP signaling
at the plasma membrane of the neuromuscular junction, while serving
a positive role in nuclear accumulation of phosphorylated Mothers
against decapentaplegic (46).
These data suggested that EPN3, as an endocytosis adapter protein,
may serve different roles in different subcellular localization;
however, additional studies to determine these roles are
required.
Epsin family proteins are required for Notch pathway
activation (9); Notch signaling
plays vital roles in glioblastoma cell survival and proliferation
(18), migration and invasion
(38), radioresistance (21) and EMT (25,26).
Similarly, WNT/β-catenin signaling has been reported to be
implicated in the proliferation of neural stem cells (47), and the activation of canonical
WNT/β-catenin signaling enhances the migration and invasion of
glioblastoma cells via activating ZEB1 and other activators of EMT
(27). Consistently, the results of
the present study demonstrated that EPN3 enhanced the migration and
invasion of glioblastoma cells, which may have been due to the
activation of Notch and WNT/β-catenin signaling. It has been
reported that the Notch and WNT pathways interact in breast
tumorigenesis (48), and that Notch
signaling upregulates the expression of Snail 1 and Slug (49). Slug is important for Notch-mediated
EMT as it functions to repress E-cadherin expression, which in turn
induces β-catenin activation in human breast cancer (38). However, in colorectal cancer, Notch1
counteracts WNT/β-catenin signaling through chromatin modification
(50). Nevertheless, the present
study hypothesized that Notch signaling and WNT signaling may act
synergistically to promote glioblastoma cell migration and
invasion.
The present study results indicated that
overexpression of EPN3 may activate EMT in glioblastoma cells and
increase the expression of Slug, Twist and ZEB1. In addition,
previous studies have reported that E-cadherin may be removed
through clathrin-mediated endocytosis, which EPN3 is involved in,
and may lead to E-cadherin degradation and the consequent
disassembly of adherens junctions (51). Taken together, EPN3 may promote
glioblastoma cell EMT, which may be dependent or independent on
activating EMT.
In conclusion, the present study results
demonstrated the oncogenic role of EPN3 in glioblastoma and its
induction of EMT; the activation of Notch and WNT signaling
pathways may also have enhanced the migration and invasion of
glioblastoma cells induced by EPN3 overexpression. However, there
are some limitations in the present study. First, whether the
effects of EPN3 on Notch and WNT pathway are related to
EPN3-enhanced glioblastoma cells migration and invasion, as well as
EMT, remain to be investigated. Second, whether transcription
factors are involved in EPN3-induced Notch and WNT pathways also
needs further research. However, further investigations are needed
to clarify the precise molecular mechanisms of the effects on
glioblastoma migration and invasion induced by EPN3
overexpression.
Acknowledgments
We thank Dr Li Qi for donating the human colon
carcinoma tissue (Tianjin Medical University Cancer Institute and
Hospital, Tianjin, China). We also thank Dr Qiaoli Wu and Mrs.
Xiuhua Yao (Clinical Laboratory, Tianjin Key Laboratory of Cerebral
Vascular and Neurodegenerative Diseases, Tianjin Neurosurgical
Institute, Tianjin Huanhu Hospital) for their technical support and
assistance in the study.
Funding
This work was supported by a grant from The Tianjin
Health and Family Planning Commission (grant no. 14KG118) and The
National Nature Science Foundation of China (grant no.
81501035).
Availability of data and materials
The data sets used and/or analyzed during the
current study are available from the corresponding author on
reasonable request.
Authors' contributions
YW performed most experiments. WS analyzed the
patient data regarding the glioma tissues. PK, CH and ZM drafted
the manuscript. XY and QW revised the manuscript critically for
important intellectual content; BZ gave final approval of the
version to be published. These authors were also involved in the
conception of the study. Each author has participated sufficiently
in the study to take public responsibility for appropriate portions
of the content and agreed to be accountable for all aspects of the
study in ensuring that questions related to the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
Informed consent was obtained from all patients, and
the use of patient tissue was approved by the Ethics Committee of
Tianjin Huanhu Hospital (Tianjin, China; 2016-3).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
BMP
|
bone morphogenetic protein
|
|
EMT
|
epithelial-mesenchymal transition
|
|
EPN3
|
Epsin 3
|
|
NICD1
|
Notch1 intracellular domain
|
References
|
1
|
Khan MN, Sharma AM, Pitz M, Loewen SK,
Quon H, Poulin A and Essig M: High-grade glioma management and
response assessment-recent advances and current challenges. Curr
Oncol. 23:e383–e391. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Majchrzak-Celińska A, Paluszczak J,
Szalata M, Barciszewska AM, Nowak S, Kleszcz R, Sherba A and
Baer-Dubowska W: The methylation of a panel of genes differentiates
low-grade from high-grade gliomas. Tumor Biol. 36:3831–3841. 2015.
View Article : Google Scholar
|
|
3
|
Cloughesy TF, Cavenee WK and Mischel PS:
Glioblastoma: From molecular pathology to targeted treatment. Ann
Rev Pathol. 9:12014. View Article : Google Scholar
|
|
4
|
Olar A and Aldape KD: Using the molecular
classification of glioblastoma to inform personalized treatment. J
Pathol. 232:165–177. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Montgomery RM, Lde Queiroz S and Rogerio
F: EGFR, p53, IDH-1 and MDM2 immunohistochemical analysis in
glioblastoma: Therapeutic and prognostic correlation. Arq
Neuropsiquiatr. 73:561–568. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Furnari FB, Fenton T, Bachoo RM, Mukasa A,
Stommel JM, Stegh A, Hahn WC, Ligon KL, Louis DN, Brennan C, et al:
Malignant astrocytic glioma: Genetics, biology, and paths to
treatment. Genes Dev. 21:2683–2710. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xie X, Cho B and Fischer JA: Drosophila
Epsin's role in Notch ligand cells requires three Epsin protein
functions: The lipid binding function of the ENTH domain, a single
Ubiquitin interaction motif, and a subset of the C-terminal protein
binding modules. Dev Biol. 363:399–412. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen H, Fre S, Slepnev VI, Capua MR, Takei
K, Butler MH, Di Fiore PP and De Camilli P: Epsin is an
EH-domain-binding protein implicated in clathrin-mediated
endocytosis. Nature. 394:793–797. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tessneer KL, Cai X, Pasula S, Dong Y, Liu
X, Chang B, McManus J, Hahn S, Yu L and Chen H: Epsin family of
endocytic adaptor proteins as oncogenic regulators of cancer
progression. J Can Res Updates. 2:144–150. 2013.PubMed/NCBI
|
|
12
|
Rosenthal JA, Chen H, Slepnev VI,
Pellegrini L, Salcini AE, Di Fiore PP and De Camilli P: The epsins
define a family of proteins that interact with components of the
clathrin coat and contain a new protein module. J Biol Chem.
274:33959–33965. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ko G, Paradise S, Chen H, Graham M, Vecchi
M, Bianchi F, Cremona O, Di Fiore PP and De Camilli P: Selective
high-level expression of epsin 3 in gastric parietal cells, where
it is localized at endocytic sites of apical canaliculi. Proc Natl
Acad Sci USA. 107:215112010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Spradling KD, McDaniel AE, Lohi J and
Pilcher BK: Epsin 3 is a novel extracellular matrix-induced
transcript specific to wounded epithelia. J Biol Chem.
276:29257–29267. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bache KG, Slagsvold T and Stenmark H:
Defective downregulation of receptor tyrosine kinases in cancer.
EMBO J. 23:2707–2712. 2014. View Article : Google Scholar
|
|
16
|
Mcmahon HT and Boucrot E: Molecular
mechanism and physiological functions of clathrin-mediated
endocytosis. Nat Rev Mol Cell Biol. 12:517–533. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chang B, Tessneer KL, McManus J, Liu X,
Hahn S, Pasula S, Wu H, Song H, Chen Y, Cai X, et al: Epsin is
required for Dishevelled stability and Wnt signalling activation in
colon cancer development. Nat Commun. 6:63802015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Purow BW, Haque RM, Noel MW, Su Q, Burdick
MJ, Lee J, Sundaresan T, Pastorino S, Park JK, Mikolaenko I, et al:
Expression of Notch-1 and its ligands, Delta-like-1 and Jagged-1,
is critical for glioma cell survival and proliferation. Cancer Res.
65:2353–2363. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang J, Wakeman TP, Lathia JD, Hjelmeland
AB, Wang XF, White RR, Rich JN and Sullenger BA: Notch promotes
radioresistance of glioma stem cells. Stem Cells. 28:17–28.
2010.PubMed/NCBI
|
|
20
|
Korkaya H and Wicha MS: HER-2, notch, and
breast cancer stem cells: Targeting an axis of evil. Clin Cancer
Res. 15:1845–1847. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Benedito R, Roca C, Sörensen I, Adams S,
Gossler A, Fruttiger M and Adams RH: The Notch ligands Dll4 and
Jagged1 have opposing effects on angiogenesis. Cell. 137:1124–1135.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kopan R and Ilagan MX: The canonical Notch
signaling pathway: Unfolding the activation mechanism. Cell.
137:216–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Blitzer JT and Nusse R: A critical role
for endocytosis in Wnt signaling. BMC Cell Biol. 7:282006.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yamamoto H, Komekado H and Kikuchi A:
Caveolin is necessary for Wnt-3a-dependent internalization of LRP6
and accumulation of beta-catenin. Dev Cell. 11:213–223. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gonzalez DM and Medici D: Signaling
mechanisms of the epithelial-mesenchymal transition. Sci Signal.
7:re82014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zavadil J, Cermak L, Soto-Nieves N and
Böttinger EP: Integration of TGF-beta/Smad and Jagged1/Notch
signalling in epithelial-to-mesenchymal transition. EMBO J.
23:1155–1165. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kahlert UD, Maciaczyk D, Doostkam S, Orr
BA, Simons B, Bogiel T, Reithmeier T, Prinz M, Schubert J,
Niedermann G, et al: Activation of canonical WNT/β-catenin
signaling enhances in vitro motility of glioblastoma cells by
activation of ZEB1 and other activators of
epithelial-to-mesenchymal transition. Cancer Lett. 325:42–53. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Prieto-García E, Díaz-García CV,
García-Ruiz I and Agulló-Ortuño MT: Epithelial-to-mesenchymal
transition in tumor progression. Med Oncol. 34:1222017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Allen M, Bjerke M, Edlund H, Nelander S
and Westermark B: Origin of the U87MG glioma cell line: Good news
and bad news. Sci Transl Med. 8:354re32016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yang J and Weinberg RA:
Epithelial-mesenchymal transition: At the crossroads of development
and tumor metastasis. Dev Cell. 14:818–829. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Xu R, Shimizu F, Hovinga K, Beal K, Karimi
S, Droms L, Peck KK, Gutin P, Iorgulescu JB, Kaley T, et al:
Molecular and clinical effects of Notch inhibition in glioma
patients: A phase 0/I trial. Clin Cancer Res. 22:4786–4796. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cheng W, Zhang C, Ren X, Jiang Y, Han S,
Liu Y, Cai J, Li M, Wang K, Liu Y, et al: Bioinformatic analyses
reveal a distinct Notch activation induced by STAT3 phosphorylation
in the mesenchymal subtype of glioblastoma. J Neurosurg.
126:249–259. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen H, Ko G, Zatti A, Di Giacomo G, Liu
L, Raiteri E, Perucco E, Collesi C, Min W, Zeiss C, et al:
Embryonic arrest at midgestation and disruption of Notch signaling
produced by the absence of both epsin 1 and epsin 2 in mice. Proc
Natl Acad Sci USA. 106:13838–13843. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Overstreet E, Fitch E and Fischer JA: Fat
facets and Liquid facets promote Delta endocytosis and Delta
signaling in the signaling cells. Development. 131:5355–5366. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang W and Struhl G: Drosophila
Epsin mediates a select endocytic pathway that DSL ligands must
enter to activate Notch. Development. 131:5367–5380. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Leong KG, Niessen K, Kulic I, Raouf A,
Eaves C, Pollet I and Karsan A: Jagged1-mediated Notch activation
induces epithelial-to-mesenchymal transition through Slug-induced
repression of E-cadherin. J Exp Med. 204:2935–2948. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pasula S, Cai X, Dong Y, Messa M, McManus
J, Chang B, Liu X, Zhu H, Mansat RS, Yoon SJ, et al: Endothelial
epsin deficiency decreases tumor growth by enhancing VEGF
signaling. J Clin Invest. 122:4424–4438. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tessneer KL, Pasula S, Cai X, Dong Y, Liu
X, Yu L, Hahn S, McManus J, Chen Y, Chang B and Chen H: Endocytic
adaptor protein epsin is elevated in prostate cancer and required
for cancer progression. ISRN Oncol. 2013:4205972013.PubMed/NCBI
|
|
41
|
Coon BG, Burgner J, Camonis JH and Aguilar
RC: The epsin family of endocytic adaptors promotes fibrosarcoma
migration and invasion. J Biol Chem. 285:33073–33081. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hellwig B, Madjar K, Edlund K, Marchan R,
Cadenas C, Heimes AS, Almstedt K, Lebrecht A, Sicking I, Battista
MJ, et al: Epsin family member 3 and ribosome-related genes are
associated with late metastasis in estrogen receptor-positive
breast cancer and long-term survival in non-small cell lung cancer
using a genome-wide identification and validation strategy. PLoS
One. 11:e01675852016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Nagano T, Nakano M, Nakashima A, Onishi K,
Yamao S, Enari M, Kikkawa U and Kamada S: Identification of
cellular senescence-specific genes by comparative transcriptomics.
Sci Rep. 6:317582016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mori J, Tanikawa C, Ohnishi N, Funauchi Y,
Toyoshima O, Ueda K and Matsuda K: Epsin 3, a novel p53 target,
regulates the apoptotic pathway and gastric carcinogenesis1.
Neoplasia. 19:185–195. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Howe CL and Mobley WC: Signaling endosome
hypothesis: A cellular mechanism for long distance communication. J
Neurobiol. 58:207–216. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Vanlandingham PA, Fore TR, Chastain LR,
Royer SM, Bao H, Reist NE and Zhang B: Epsin 1 promotes synaptic
growth by enhancing BMP signal levels in motoneuron nuclei. PLoS
One. 8:e659972013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Nusse R: Wnt signaling and stem cell
control. Cell Res. 18:523–527. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ayyanan A, Civenni G, Ciarloni L, Morel C,
Mueller N, Lefort K, Mandinova A, Raffoul W, Fiche M, Dotto GP and
Brisken C: Increased Wnt signaling triggers oncogenic conversion of
human breast epithelial cells by a Notch-dependent mechanism. Proc
Natl Acad Sci USA. 103:3799–3804. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang Z, Li Y, Kong D, Banerjee S, Ahmad A,
Azmi AS, Ali S, Abbruzzese JL, Gallick GE and Sarkar FH:
Acquisition of epithelial-mesenchymal transition phenotype of
gemcitabine-resistant pancreatic cancer cells is linked with
activation of Notch signaling pathway. Cancer Res. 69:2400–2407.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kim HA, Koo BK, Cho JH, Kim YY, Seong J,
Chang HJ, Oh YM, Stange DE, Park JG, Hwang D and Kong YY: Notch1
counteracts WNT/β-catenin signaling through chromatin modification
in colorectal cancer. J Clin Invest. 122:3248–3259. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kohn KW, Zeeberg BM, Reinhold WC and
Pommier Y: Gene expression correlations in human cancer cell lines
define molecular interaction networks for epithelial phenotype.
PLoS One. 9:e992692014. View Article : Google Scholar : PubMed/NCBI
|