Introduction
Gastric cancer is the third most lethal malignancy
worldwide (1). Currently, the main
option for treating gastric cancer is surgical resection combined
with chemotherapy and/or radiotherapy. In recent years, due to the
fast development of enterescopy and surgical techniques, the 5-year
mortality for early gastric cancer has been significantly reduced.
However, most patients with gastric cancer are diagnosed at an
advanced stage and miss the optimal timing for surgical
intervention. Even when curative resection is performed
successfully, ~40–65% of patients may still experience a recurrence
of the disease. Therefore the 5-year mortality still remains high
(2). Although extensive studies
have been carried out to explore the possible mechanisms that
underlie the development of gastric cancer, the exact mechanism
remains ambiguous. Better understanding of the related mechanisms
is important in order to establish new efficient targets and to
develop novel treatments to manage gastric cancer.
Dysregulated metabolism frequently occurs in various
cancer cells, and is considered as one of the important hallmarks
of cancer (3). The famous ‘Warburg
effect’ states that glucose consumption becomes a priority for
cancer cells even in the presence of oxygen (4,5).
Blocking cancer glycolysis is considered to be a promising
therapeutic strategy for cancer treatment. Pyruvate links
glycolysis and the TCA cycle, thus enzymes involved in the pyruvate
reaction play important roles in the metabolic node (6). Pyruvate dehydrogenase α 1 (PDHA1) is a
key component of the pyruvate dehydrogenase complex which catalyzes
pyruvate decarboxylation and serves as a gate-keeper enzyme link
between glycolysis and the mitochondrial citric acid cycle. The
inhibition of pyruvate dehydrogenase in cancer cells increases the
Warburg effect in cancer cells and renders cancer cells more
malignant (7,8). PDHA1 dysregulation has also been
revealed to be associated with metabolic reprogramming in cancer
cells (9,10). It has been reported that decreased
expression of PDHA1 predicted an unfavorable prognosis in ovarian
carcinoma (11), and low PDHA1
protein expression may indicate the aggressive features of clear
cell carcinoma (12). However, in
gastric cancer, the role of PDHA1 has not been studied.
MicroRNAs (miRNAs) are one of the main reasons for
protein level dysregulation. miRNAs are a class of small non-coding
RNAs that are generally 19–25 nucleotides in length (13,14).
miRNAs regulate gene expression by binding to the 3′untranslated
region (3′UTR) of the target mRNA, leading to inhibition of protein
translation or cleavage of the target mRNA (15,16).
Accumulating evidence has revealed that miRNAs are involved in
various biological process, including development, apoptosis and
differentiation (17–20). Research has also revealed that
dysregulation of miRNAs is associated with many diseases, including
tumors (21).
In the present study, we found that PDHA1 was
downregulated in gastric cancer, and PDHA1 downregulation promoted
glycolysis and gastric cancer cell progression. PDHA1 was directly
regulated by miR-21-5p. In addition, miR-21-5p was upregulated in
gastric cancer, and promoted gastric cancer progression by
regulating PDHA1. Our results indicated the role of the
miR-21-5p/PDHA1 axis in regulating gastric cancer metabolism and
progression, and suggests their potential value in gastric cancer
treatment.
Materials and methods
Clinical specimens
Gastric cancer and matched adjacent normal tissues
were collected from gastric cancer patients at China-Japan Union
Hospital of Jilin University. The tissue samples from 46 patients
(31 males and 15 females with age ranging from 43 to 71 years) were
collected from November 2016 to September 2017. All carcinoma
samples were obtained during surgery, snap-frozen in liquid
nitrogen and stored at −80°C until further analysis. In the present
study, all the cases of gastric tumors were divided into low-grade
(WHO I and II) or high-grade (WHO III and IV) for statistical
analysis. This study was approved by the Ethics Committee of
China-Japan Union Hospital of Jilin University. Written informed
consent was obtained from all the patients or their guardians in
accordance with the ethical committee standards.
RNA extraction and real-time
quantitative polymerase chain reaction
Total RNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Total RNA (500 ng) was used for cDNA synthesis using a High
Capacity RNA-to-cDNA Kit (Applied Biosystems; Thermo Fisher
Scientific, Inc.). miScript Reverse Transcription kit (Qiagen GmbH,
Hilden, Germany) was used for reverse transcription of miRNAs.
Quantitative real-time polymerase chain reaction (PCR) was carried
out on an ABI PRISM 7500 Sequence Detection System (Applied
Biosystems, Foster City, CA, USA) using SYBR Premix Ex Taq
(Takara Biotechnology Co., Ltd., Dalian, China) according to the
manufacturer's instructions. The miRNA sequence-specific reverse
transcription (RT)-PCR for miR-21-5p and endogenous control U6 was
performed using Hairpin-it™ miRNAs qPCR quantitation kit and U6
snRNA real-time PCR normalization kit (Shanghai GenePharma Co.,
Ltd., Shanghai, China). The primers for PDHA1 are: forward,
5′-GAGCTGAGCAGCTGTGTAAC-3′ and reverse,
5′-TGCCAATCGTTACAGGTATTACAG-3′. The thermocycling conditions for
miR-21-5p were: 94°C for 3 min (hold), 40 cycles at 94°C for 15 sec
(denaturation); 55°C for 25 sec (annealing); 72°C for 25 sec
(elongation). The thermocycling conditions for PDHA1 were: 95°C for
20 sec (enzyme activation), 40 cycles at 95°C, for 3 sec
(denaturation); 60°C for 30 sec (annealing/elongation). All
reactions were performed in triplicate.
Immunohistochemistry and in situ
hybridization
Tumor or matched normal tissues were fixed with 4%
paraformaldehyde, and then the samples were embedded in paraffin
and sectioned. Following overnight incubation at 4°C with the PDHA1
antibody (1:100; cat. no. 3205; Cell Signaling Technology, Inc.,
Danvers, MA, USA), the bound antibodies were detected with the
biotin-streptavidin-peroxidase system (Vector Laboratories, Inc.,
Burlingame, CA, USA) using diaminobenzidine (Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany) as chromogen.
For the in situ hybridization assay,
DIG-labeled locked nucleic acid (LNA)-based probe specific for
miR-21-5p (Exiqon A/S, Vedbaek, Denmark) was introduced. In
addition, the in situ hybridization signal was detected by
overnight incubation at 4°C with horseradish peroxidase
(HRP)-conjugated anti-digoxigenin 21H8 (1:200; cat. no. ab420;
Abcam, Cambridge, MA, USA).
Transfection
The miR-21-5p mimics, mimics negative control,
inhibitor, inhibitor negative control (inhibitor control), the
small interfering RNA targeting PDHA1 (siPDHA1) and control-siRNA
(siCon) were purchased from Shanghai GenePharma Co., Ltd. Cells
were seeded at a density of 2–3×105 cells/well in 6-well
plates for 24 h. The cells were then transfected with
oligonucleotides or plasmid, using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) following the manufacturer's
instructions.
Construction and preparation of the
PDHA1 lentivirus
PDHA1 cDNA was amplified from an adult gastric
tissue cDNA library. The recombinant lentiviruses were packaged
using the pMagic 4.0 lentivirus pLV expression system (Invitrogen;
Thermo Fisher Scientific, Inc.). Briefly, recombination was
produced by co-transfection of 293T cells with the lentivirus
plasmid (pLV- PDHA1) and packaging plasmids (pLV1.0, pLV2.0 and
pLV3.0) using the Lipofectamine 2000 transfection reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol.
Cell proliferation assay
Cell proliferation was assessed using the MTT assay.
MTT was diluted in phosphate-buffered saline (PBS) to a final
concentration of 5 mg/ml and sterile filtered. Cells were incubated
with a final concentration of 0.5 mg/ml MTT at 37°C for 4 h. Cell
culture supernatants were carefully removed, and dimethyl sulfoxide
(DMSO) was added. The absorbance values were determined using a
microplate reader (Pharmacia Biotech, Uppsala, Sweden) at a
wavelength of 570 nm. The experiments were performed 3 times.
Colony formation assay
The cells were transfected with PDHA siRNA or
control siRNA and seeded into a 6-well plate. Following ~2 weeks of
culture, the cells were fixed in methanol, stained with 0.1% (w/v)
crystal violet and washed with water. The cell colonies were scored
and the representative light images were photographed using a Canon
camera.
Glucose and lactate assessments
Glucose and lactate contents in culture medium were
evaluated using the BS-200 Chemistry Analyzer (Mindray Bio-Medical
Electronics Co., Ltd., Shenzhen, China) and EnzyChrom™ D-Lactate
Assay kit (BioAssay Systems, Hayward, CA, USA), respectively. Data
were normalized to the cell number in each well. For glucose uptake
assays, the cells were maintained under normal conditions for 24 h
and 10 µM 2-NBDG (Life Technologies; Thermo Fisher Scientific,
Inc.) was added to the medium for 30 min in the dark at 37°C. After
being washed with PBS twice, the labeled cells were collected as
single cell suspensions and the fluorescence intensities were
determined by flow cytometry.
PDHA1 targeting miRNAs prediction
The putative miRNAs targeting PDHA1 were analyzed
using online programs including DIANA TOOLS (http://diana.imis.athena-innovation.gr/DianaTools/index.php?r=microT_CDS/index),
TargetScan 7.2 (http://www.targetscan.org/vert_72/) and miRDB
(http://mirdb.org/miRDB/index.html).
Dual-Luciferase Reporter Assay
The 3′untranslated region (3′UTR) of PDHA1
containing the miR-21-5p binding sites was amplified by PCR from
human genomic DNA. The wild-type 3′UTR of PDHA1 as well as the
mutant 3′UTR with nucleotide substitutions in the putative binding
sites corresponding to the seed sequence of miR-21-5p were inserted
into the psiCHECK-2 vector immediately downstream of the stop codon
of luciferase to develop psiCHECK2-PDHA1-3′UTR and
psiCHECK-PDHA1-mut-3′UTR, respectively. Either of these vectors was
co-transfected with miRNAs into cells using Lipofectamine 2000
according to the manufacturer's protocols. Luciferase activity was
assessed 48 h after transfection using a Dual-Luciferase Reporter
Assay kit (Promega Corp., Madison, WI, USA) and Victor Luminometer
(PerkinElmer, Inc., Waltham, MA, USA). The firefly luciferase
activity was normalized using co-transfected Renilla
luciferase for transfection efficiency. Three independent
experiments were performed and the data are presented as the mean ±
standard deviation (SD).
Protein extraction and western blot
analysis
Total protein from cells was extracted with RIPA
Lysis Buffer (Cell Signaling Technology, Inc.) and total protein
was assessed using the Bradford protein assay (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Protein (30 mg) were
separated by 10% SDS-PAGE and transferred onto polyvinylidene
fluoride (PVDF) membranes (EMD Millipore, Billerica, MA, USA).
After blocking for non-specific binding with 5% skimmed milk
dissolved in Tris-buffered saline plus Tween-20 (TBS-T; 0.1%
Tween-20; pH 8.3) at room temperature for 1 h, the membranes were
incubated with antibodies against PDHA1 (1:1,000 dilution; cat. no.
3205) and β-actin (1:2,000 dilution; cat. no. 4970) (both from Cell
Signaling Technology, Inc.) at 4°C overnight. After incubation with
a secondary antibody conjugated to horseradish peroxidase (cat. no.
7074; Cell Signaling Technology) at room temperature for 1 h, the
protein bands were visualized using an enhanced chemiluminescence
detection kit (EMD Millipore). ImageJ-1.51k software (developed by
National Institutes of Health, Bethesda, MD, USA) was used for
density measurement.
Statistical analysis
Data were analyzed using SPSS software 15.0 (SPSS,
Inc., Chicago, IL, USA). Differences of PDHA1 and miR-21-5p
expression between tumor tissues and adjacent non-tumor tissues
were analyzed by the Wilcoxon matched-pairs test. The relationship
between the expression of PDHA1 or miR-21-5p and the
clinicopathologic grade was assessed by the χ2 test.
Quantitative data are presented as the mean ± the standard
deviation. Statistical differences between groups were determined
using the Student's t-test. Differences were considered significant
when P<0.05.
Results
PDHA1 is downregulated in human
gastric cancer
Pyruvate dehydrogenase (PDH) is a multienzyme
complex that functions as a gatekeeper in glucose metabolism by
oxidatively decarboxylating pyruvate to produce acetyl-CoA for the
TCA cycle, and therefore is closely related to increased glycolysis
in cancer. PDHA1 is the main component of PDH, which catalyzes
pyruvate decarboxylation and serves as a gate-keeper enzyme link
between glycolysis and the mitochondrial citric acid cycle. To
determine the role of PDHA1 in gastric cancer, we determined
whether PDHA1 is dysregulated in gastric cancer patient samples. By
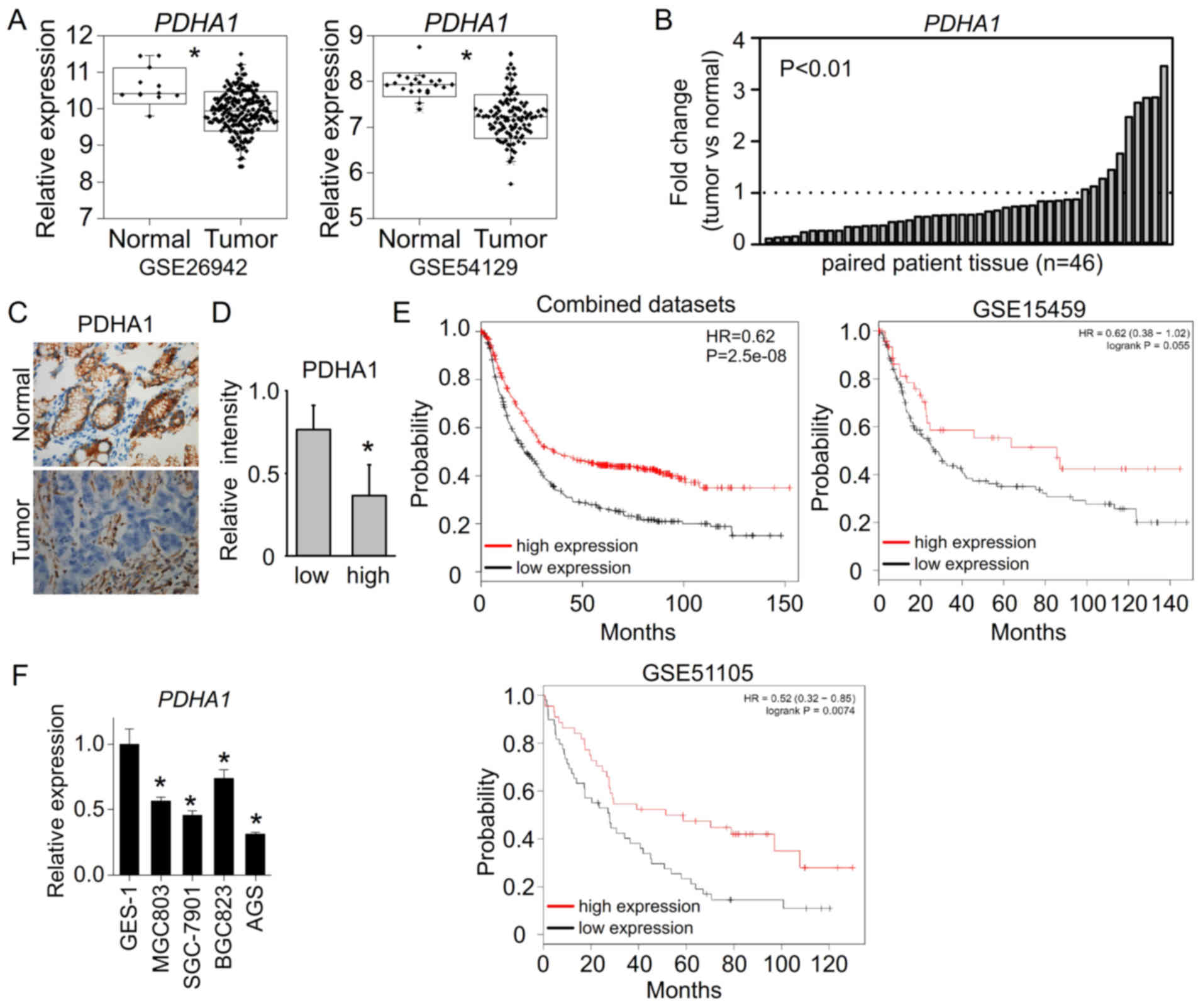
analyzing the gastric cancer patient datasets, we found that PDHA1
expression was significantly downregulated in gastric cancer
patient samples compared to normal gastric tissues (GSE26942 and
GSE54120) (Fig. 1A). In our paired
gastric cancer samples, the mRNA levels of PDHA1 were also
downregulated in 36 out of 46 gastric cancer tissues compared to
their adjacent non-tumor tissues (Fig.
1B). Moreover, we further assessed the protein levels of PDHA1
in gastric cancer tissues. The similar results revealed that PDHA1
protein levels were also downregulated in gastric cancer tissues
compared to their adjacent normal tissues (Fig. 1C). In addition, the PDHA1 levels
were less expressed in high grade gastric cancer tissues (Fig. 1D), suggesting that the
downregulation of PDHA1 may be correlated with gastric cancer
progression. The survival analysis of patients further revealed
that low PDHA1 was associated with poor survival (Fig. 1E). We further assessed the
expression levels of PDHA1in gastric cancer cell lines. The results
revealed that PDHA1 expression was significantly reduced in gastric
cancer cell lines compared to normal gastric cell line GES-1
(Fig. 1F). These data indicated
that PDHA1 was downregulated in gastric cancer and involved in
gastric cancer development.
PDHA1 downregulation promotes
glycolysis and cancer progression
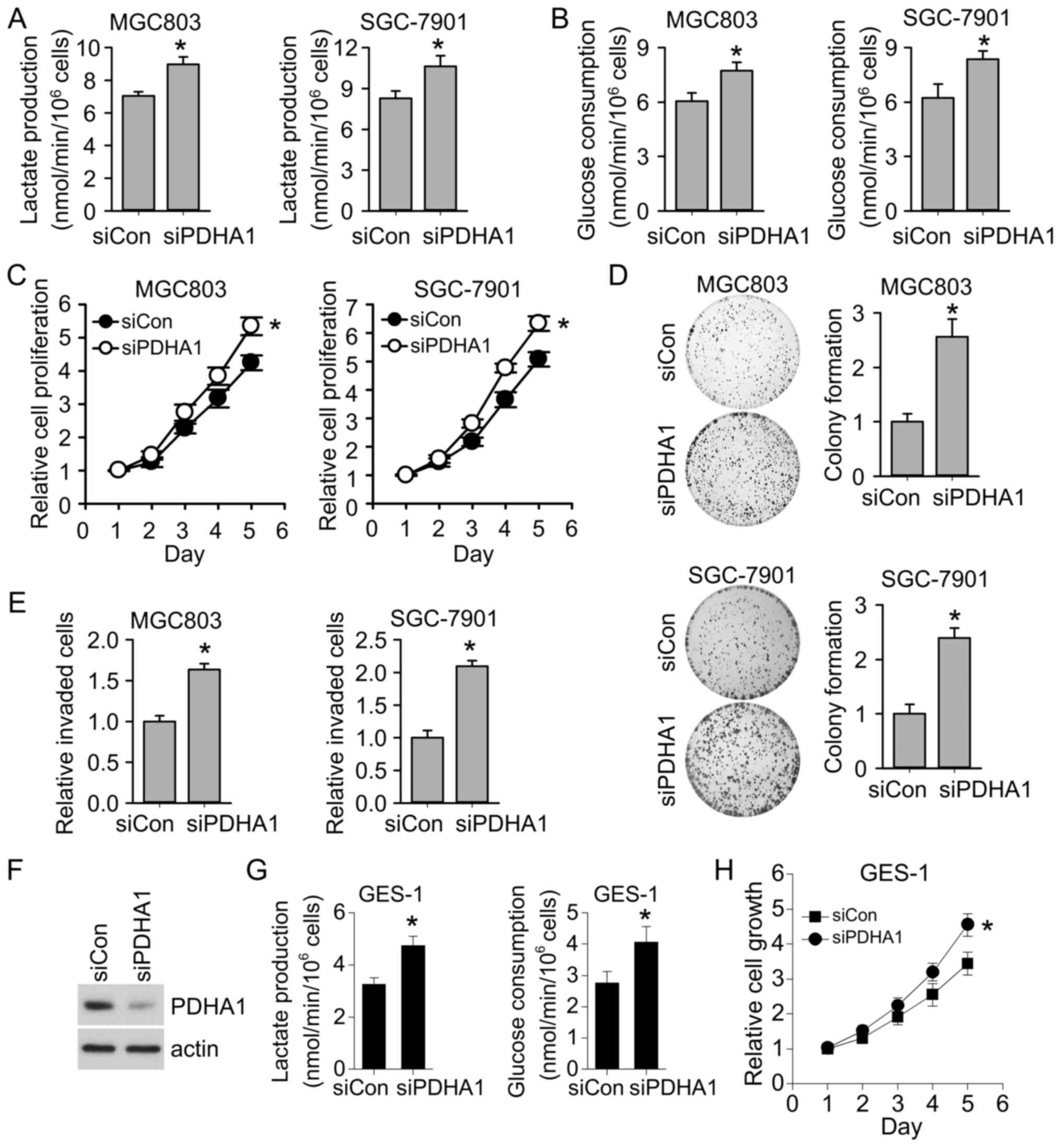
To examine the role of PDHA1 downregulation in
gastric cancer progression, we knocked down PDHA1 in gastric cancer
cell lines MGC803 and SGC-7901. Consistent with the role of PDHA1
in regulating glycolysis, our results revealed that knockdown of
PDHA1 promoted glycolysis. Lactate production and glucose
consumption were significantly increased following PDHA1 knockdown
(Fig. 2A and B). Moreover, we found
that PDHA1 knockdown significantly promoted cell proliferation
(Fig. 2C). Consistently, the colony
formation ability of gastric cancer cells was also enhanced after
PDHA1 knockdown (Fig. 2D). Our
results also revealed that PDHA1 knockdown promoted gastric cancer
invasion (Fig. 2E). All these
results indicated that PDHA1 downregulation in gastric cancer cell
enhanced glycolysis and promoted gastric cancer progression.
Notably, we found that knockdown of PDHA1 in normal gastric cells
(Fig. 2F) also resulted in
increased glycolysis and cell growth (Fig. 2G and H), further supporting that
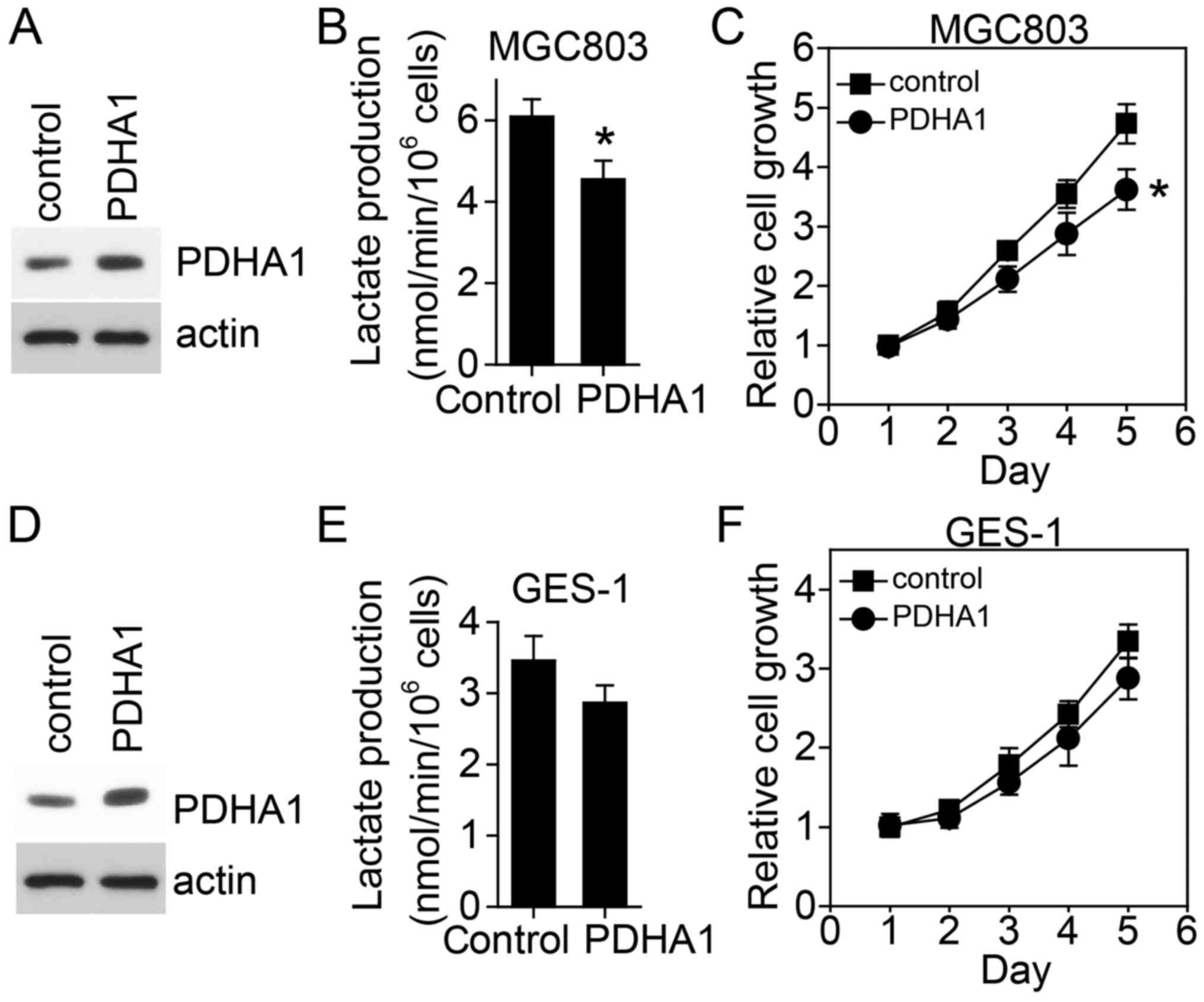
PDHA1 downregulation plays a tumor promoting role. In contrast,
overexpression of PDHA1 in gastric cancer cells reduced glycolysis
and resulted in cell growth inhibition (Fig. 3A-C). Even in normal gastric cells,
overexpression of PDHA1 also inhibited glycolysis and cell growth,
however, not significantly (Fig.
3D-F).
miR-21-5p targets PDHA1 in gastric
cancer cells
miRNAs target transcript mRNAs to regulate gene
expression and play important roles in cancer. To investigate the
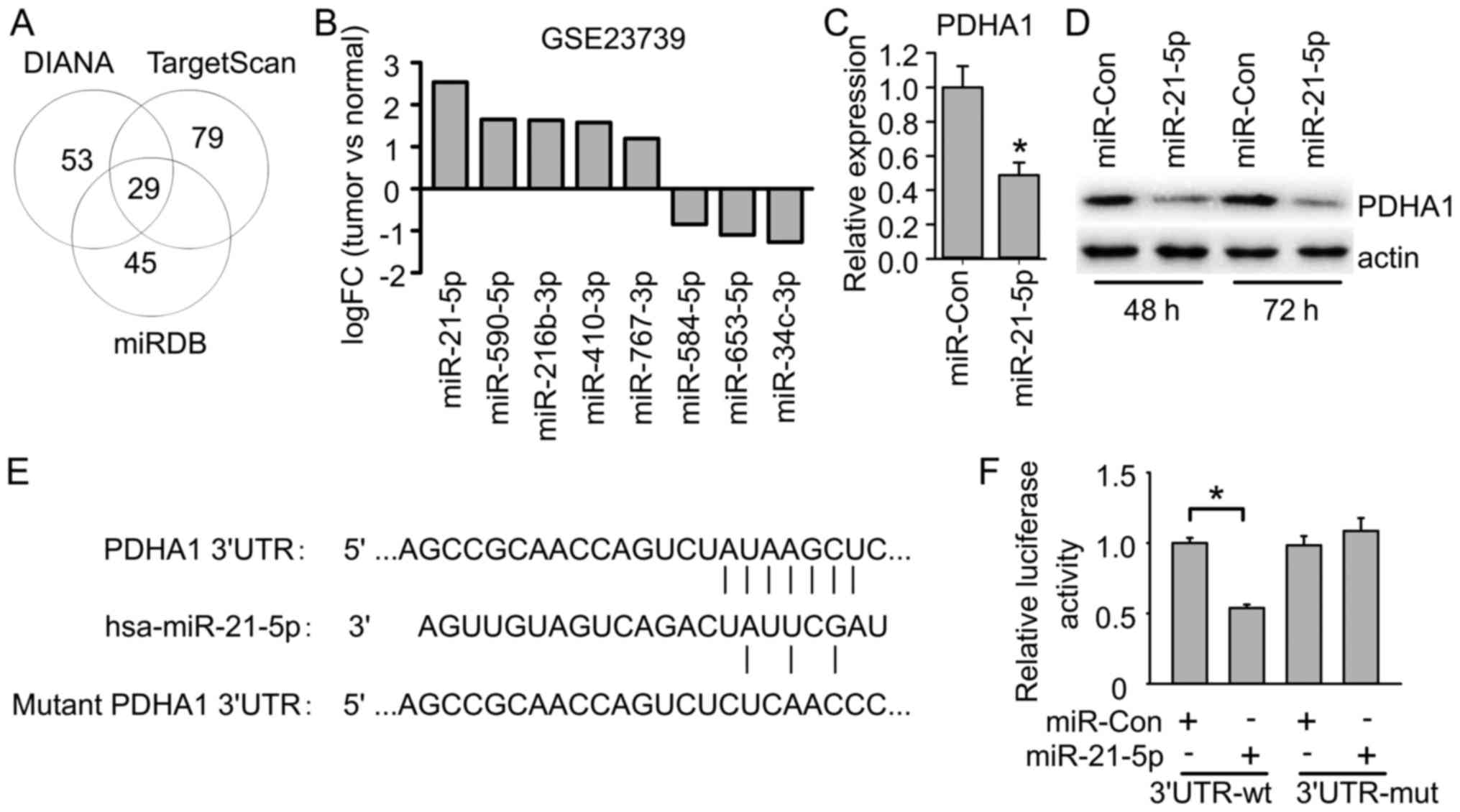
miRNA which regulates PDHA1 in gastric cancer, we searched putative
miRNAs targeting PDHA1 using online programs including DIANA,
TargetScan and miRDB. DIANA revealed 82 candidate miRNAs targeting
PDHA1, TargetScan revealed 108 candidate miRNAs, and miRDB revealed
74 candidate miRNAs. By comparing the common miRNAs, we found 29
miRNAs targeting PDHA1 predicted by all these three programs
(Fig. 4A), suggesting they likely
target PDHA1. To further investigate which one might target PDHA1
in gastric cancer, we first compared their expression in clinical
gastric cancer samples. Several of these candidates exhibited
dysregulation in gastric cancer samples compared to normal gastric
samples. Among them, miR-21-5p exhibited the most significant
upregulation in gastric cancer samples (Fig. 4B), which was consistent with PDHA1
downregulation in gastric cancer. We further examined whether
miR-21-5p targets PDHA1 in gastric cancer cells. miR-21-5p mimics
were transfected into MGC-803 cells, and the mRNA and protein
levels of PDHA1 were examined. The results revealed that miR-21-5p
transfection significantly reduced PDHA1 mRNA and protein levels in
gastric cancer cells (Fig. 4C and
D). Consistently, miR-21-5p had a conserved binding site on the
3′UTR of PDHA1 mRNA (Fig. 4E). We
further used a luciferase assay to confirm PDHA1 as a target of
miR-21-5p. The results revealed that miR-21-5p reduced the PDHA1
3′UTR-derived luciferase activity, but did not affect luciferase
activity derived by PDHA1 3′UTR with the mutant miR-21-5p binding
site (Fig. 4F). These data
indicated that miR-21-5p directly targeted PDHA1 mRNA to suppress
PDHA1 expression.
miR-21-5p is upregulated in gastric
cancer
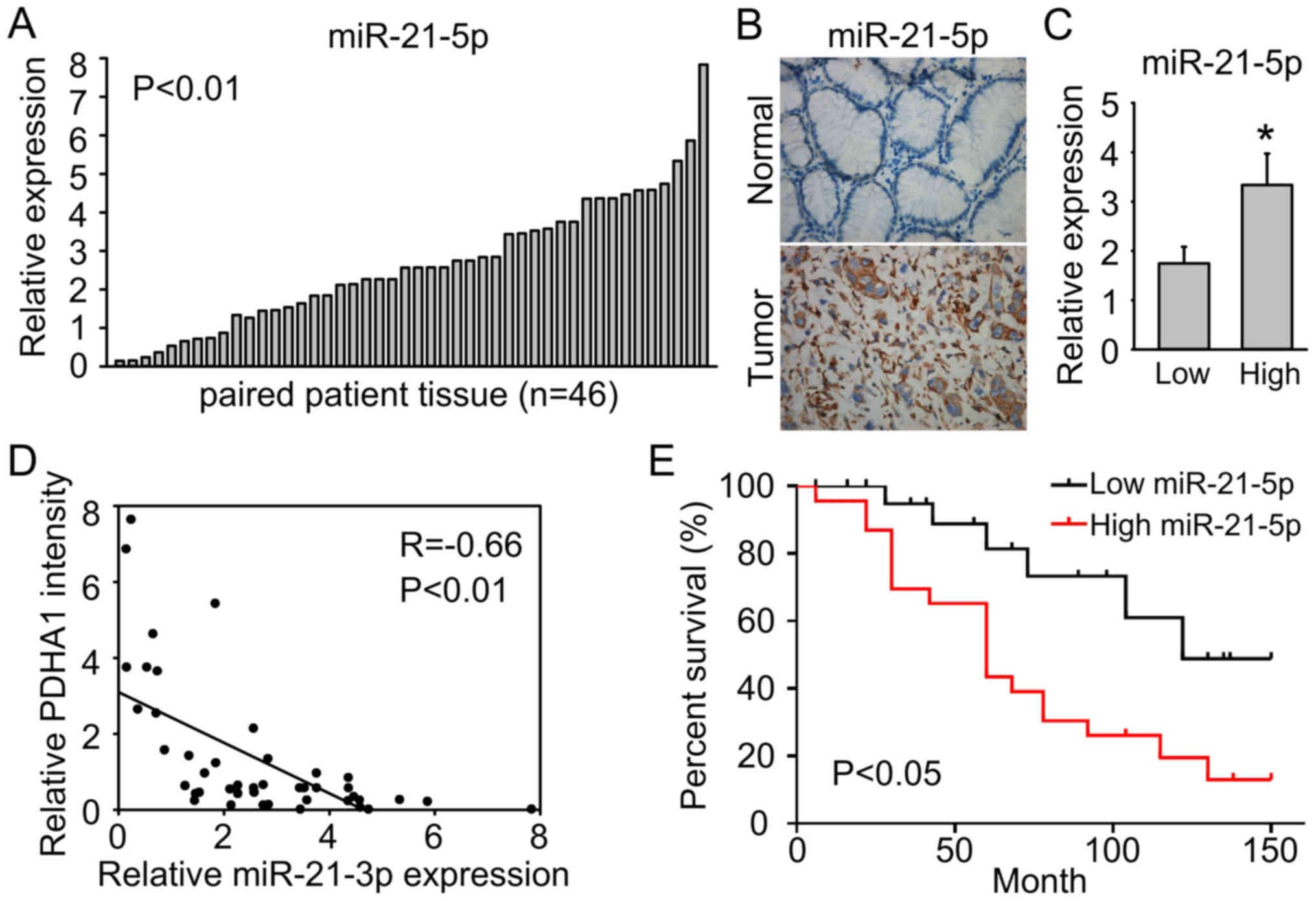
We further confirmed miR-21-5p upregulation in our
gastric cancer samples. We assessed the expression levels of
miR-21-5p in 46 pairs of gastric cancer tissue samples by q-PCR
analysis. As shown in Fig. 5A,
miR-21-5p was significantly upregulated in gastric cancer tissues
compared to adjacent non-tumor tissues (P<0.01). The results of
in situ hybridization analysis also revealed that miR-21-5p
was upregulated in gastric cancer tissues (Fig. 5B). Notably, miR-21-5p was
significantly upregulated in high grade gastric cancer compared to
low grade tissues (Fig. 5C).
Moreover, PDHA1 was negatively associated with miR-21-5p in gastric
cancer samples (Fig. 5D), further
supporting that miR-21-5p targets PDHA1 in gastric cancer. In
addition, high miR-21-5p expression was associated with poor
survival (Fig. 5E).
miR-21-5p regulates glycolysis and
inhibits gastric cancer cell growth
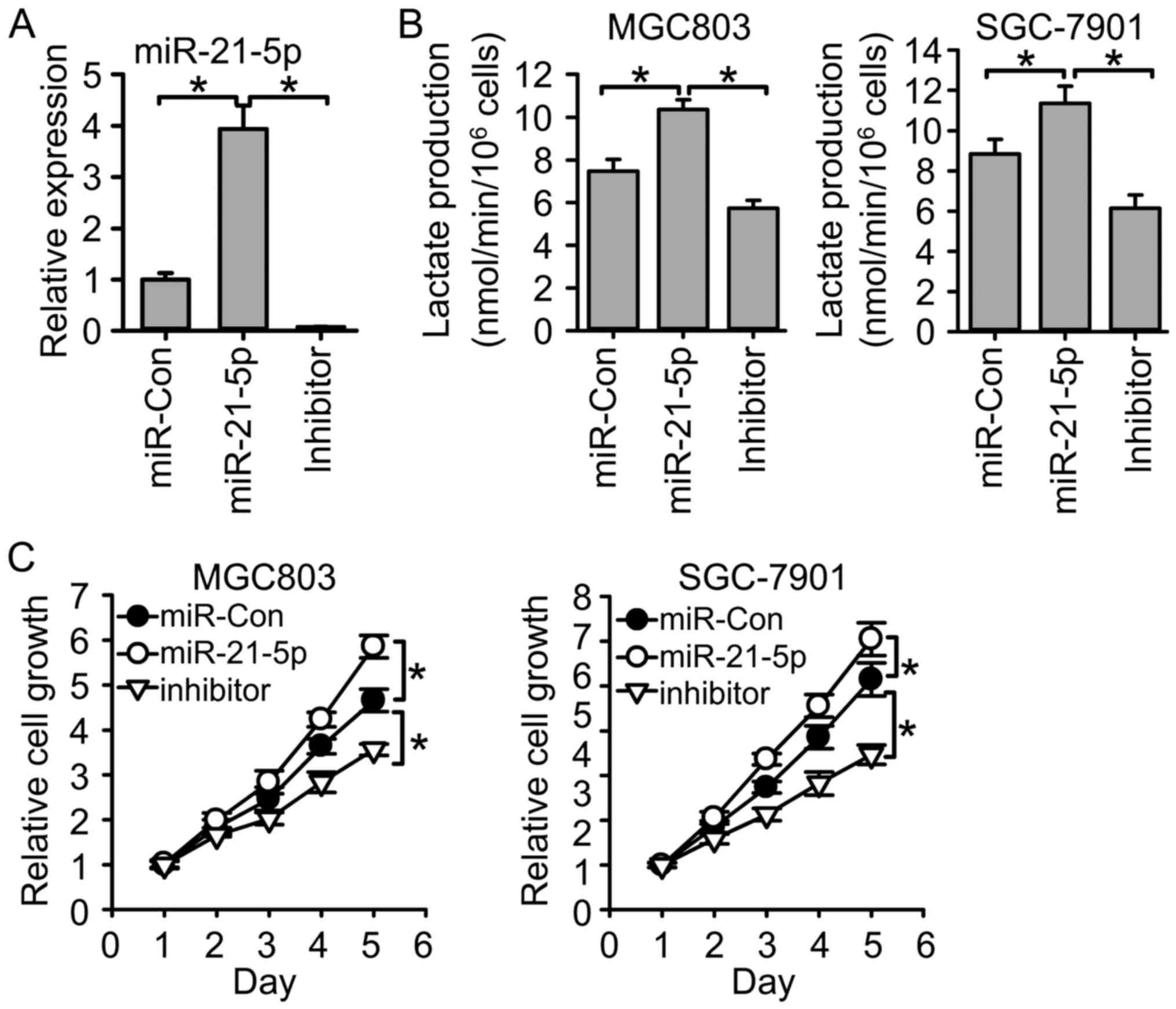
Since miR-21-5p targeted PDHA1 in gastric cancer, we
next examined whether miR-21-5p also regulated glycolysis and cell
growth. We adjusted miR-21-5p expression through transfection of
miR-21-5p mimics and its inhibitor (Fig. 6A). The results revealed that
transfection of miR-21-5p mimics increased lactate production,
while transfection of miR-21-5p inhibitor reduced lactate
production (Fig. 6B), suggesting
that miR-21-5p promotes glycolysis in gastric cancer. In addition,
miR-21-5p overexpression promoted gastric cancer cell growth, while
miR-21-5p inhibition reduced gastric cancer cell growth (Fig. 6C). These results indicated the tumor
promoting role in gastric cancer.
miR-21-5p targets PDHA1 to regulate
glycolysis and gastric cancer cell growth
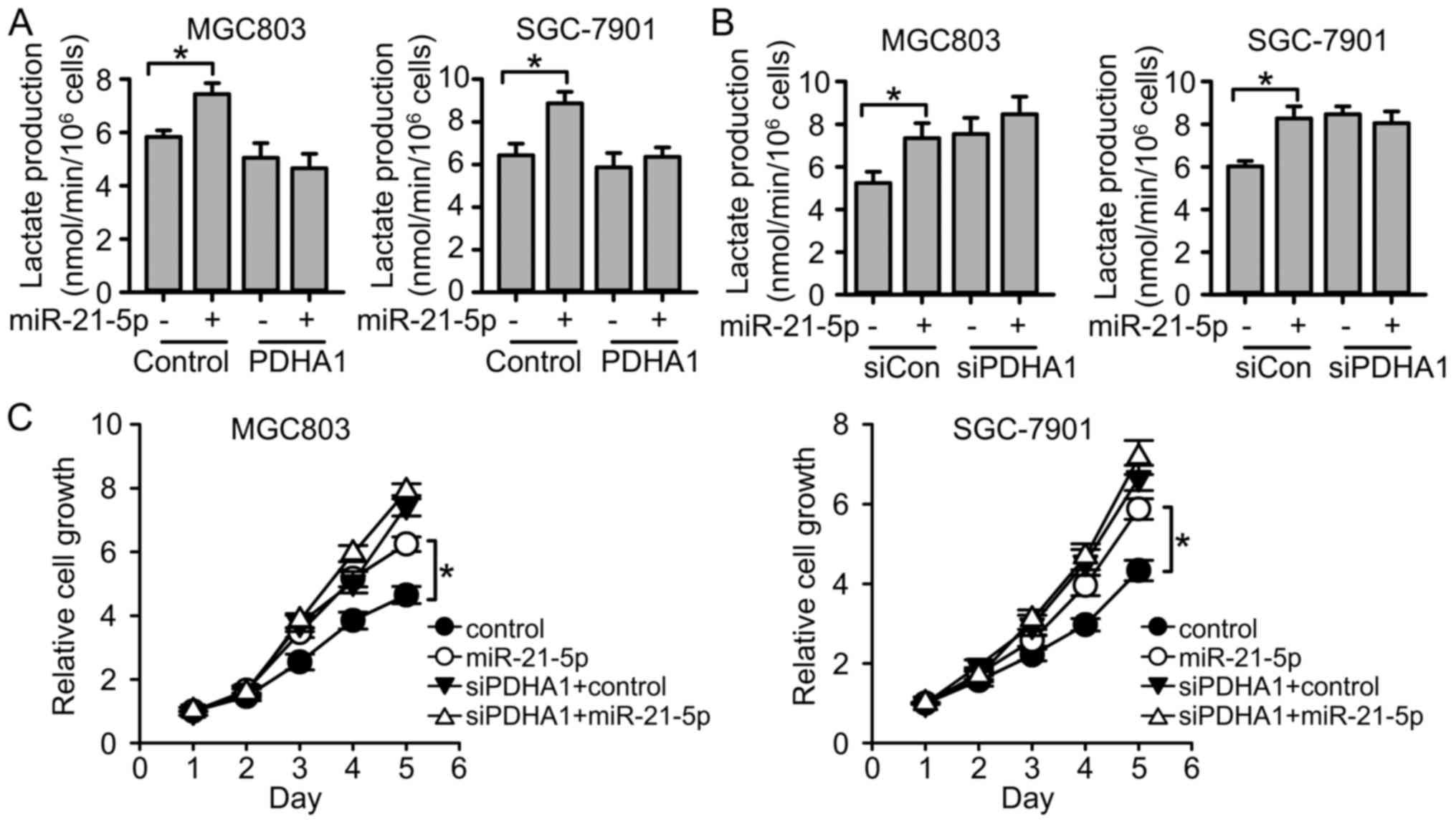
Next, we investigated whether miR-21-5p regulated
glycolysis and cell growth through PDHA1. miR-21-5p and PDHA1 were
both overexpressed in gastric cancer cell lines. We revealed that
miR-21-5p promoted glycolysis in control cells, but failed to
promote glycolysis in PDHA1-overexpressed cells (Fig. 7A). Consistently, when we suppressed
PDHA1 expression, miR-21-5p failed to further increase the levels
of glycolysis (Fig. 7B). The
results indicated that miR-21-5p regulated glycolysis through
PDHA1. Moreover, we found that miR-21-5p promoted cell
proliferation, while PDHA1 knockdown impaired the effect of
miR-21-5p on cell proliferation (Fig.
7C). All the results indicated that miR-21-5p targets PDHA1 to
regulate glycolysis and gastric cancer cell proliferation.
Collectively, we revealed that PDHA1 was
downregulated in gastric cancer, and PDHA1 downregulation promoted
glycolysis and cancer progression. miR-21-5p upregulation
contributed to PDHA1 downregulation and regulated gastric cancer
cell progression. Our study indicated that miR-21-5p and PDHA1 are
involved in gastric cancer progression, and suggests that we could
target miR-21-5p to modulate PDHA1 expression and benefit gastric
cancer treatment.
Discussion
In the present study, we demonstrated that PDHA1 was
downregulated in gastric cancer cells, and PDHA1 downregulation
promoted glycolysis and resulted in enhanced cell proliferation,
colony formation and invasion of gastric cancer cells. Further
study revealed that miR-21-5p was upregulated in gastric cancer,
and targeted PDHA1 to suppress PDHA1 expression. miR-21-5p
functioned through PDHA1 to regulate glycolysis and gastric cancer
progression. Our results indicated that the promotion of glycolysis
in gastric cancer was dependent on the downregulation of PDHA1,
resulting from the upregulation of miR-21-5p.
Cancer cells have a distinct metabolic pattern with
enhanced glycolysis levels, that is known as the famous ‘Warburg
effect’ (4,5). This phenomenon is considered as one of
the important hallmarks of cancer (3), and has been taken advantage to benefit
cancer diagnosis and therapy. Clinically, high glucose uptake is
used to diagnose or monitor the treatment responses of cancers by
imaging the uptake of 2–18 F-deoxyglucose with PET-CT (3). Many metabolic enzymes have been found
to serve as therapeutic targets for cancer treatment. Pyruvate
metabolic enzymes regulate pyruvate transferring from the
glycolytic pathway to the TCA cycle, and thus play essential roles
in cancer cell metabolism and cancer progression. Pyruvate
dehydrogenase (PDH) is a multienzyme complex consisting of multiple
copies of E1, E2 and E3 subunits, along with an E3 binding protein
(E3BP), which serves to bind E3 to the complex. PDH functions as a
gatekeeper in glucose metabolism by oxidatively decarboxylating
pyruvate to produce acetyl-CoA for the TCA cycle. Therefore, this
enzyme plays an important role in the metabolic node (6). PDHA1 is the main component of PDH,
which catalyzes pyruvate decarboxylation and serves as a
gate-keeper enzyme link between glycolysis and the mitochondrial
citric acid cycle. In the present study, we revealed that
downregulation of PDHA1 in gastric cancer cells was associated with
poor survival, and led to increased glycolysis and promoted cancer
progression. These results were consistent with the phenotype
caused by dysregulation of other pyruvate-associated enzymes such
as pyruvate dehydrogenase kinase 1 (PDK1) and mitochondrial
pyruvate carrier (MPC). Consistent with the tumor-suppressive role
of PDHA1, we revealed that PDHA1 was downregulated in gastric
cancer tissues, particularly in high-grade tumors.
Considering the tumor-suppressive role of PDHA1 in
cancer, it is important to readjust PDHA1 expression to modulate
cancer metabolism and cell growth. Thus, it is necessary to
investigate how PDHA1 expression is regulated. A miRNA can regulate
gene expression by binding to the 3′UTR of the target mRNA. miRNAs
play important roles in most cellular processes by regulating the
protein expression of target genes. It has been reported that
miRNAs are dysregulated in various types of human cancers. The
dysregulation of miRNAs leads to altered expression of target genes
including tumor suppressors and oncogenes, and regulates tumor
promotion and progression. To determine which miRNA regulates PDHA1
in gastric cancer, we first analyzed the common miRNAs targeting
PDHA1 in three well known miRNA prediction programs, and then
analyzed which miRNAs were upregulated in gastric cancer. These
miRNAs were likely to target PDHA1 in gastric cancer. Among these
candidates, miR-21-5p was the most highly upregulated miRNA in
gastric cancer, and we further confirmed that it directly targeted
PDHA1 expression. We demonstrated that overexpressed miR-21-5p
significantly suppressed PDHA1 expression, and more importantly,
miR-21-5p expression was negatively associated with PDHA1
expression in clinical gastric cancer samples.
miR-21 has been frequently reported to be aberrantly
overexpressed in diverse tumors, including glioblastoma, breast
cancer and malignant cholangiocytes (22,23).
Recently, miR-21-5p was also reported to be overexpressed in
gastric cancer (24). Previous
studies demonstrated that miR-21-5p plays a tumor-promoting role in
many types of cancer. It functions by regulating several oncogenic
pathways such as PI3K/AKT and modulating matrix metalloproteases
(25,26). Consistently, we found that miR-21-5p
was upregulated in gastric cancer tissues and miR-21-5p expression
was associated with tumor grade, and exhibited high expression in
high-grade tumors and low expression in low-grade tumors.
Inhibition of miR-21-5p led to reduced cell growth. The results
revealed that miR-21-5p serves as an oncogene in gastric cancer.
However, the role of miR-21-5p in cancer metabolism has not been
studied. We revealed for the first time that miR-21-5p
overexpression promoted glycolysis and cell proliferation in
gastric cancer cells; miR-21-5p targeted PDHA1 to regulate
glycolysis and cancer progression. Either forced overexpression or
knockdown of PDHA1 could abrogate miR-21-5p-induced glycolysis.
This suggests that miR-21-5p regulated glycolysis in gastric cancer
mainly through PDHA1.
In the present study, we demonstrated that PDHA1,
the essential pyruvate metabolism enzyme, is downregulated in
gastric cancer cells, and PDHA1 downregulation promoted glycolysis
and cancer cell growth. We further revealed that PDHA1 was a direct
target of miR-21-5p. miR-21-5p was upregulated in gastric cancer,
and suppressed PDHA1 expression to promote glycolysis and cancer
cell growth. In summary, our study illustrated that the
miR-21-5p/PDHA1 axis was involved in gastric cancer glycolysis and
progression, suggesting their potential benefit in gastric cancer
treatment.
Acknowledgements
Not applicable.
Funding
The present study was supported in part by a grant
from the Jilin Programs for Science and Technology Development (no.
20160101034JC).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
ZL, TM and DW conceived and designed the study. ZL,
MY, BF and XF performed the experiments. ZL wrote the manuscript.
ZL, XF, TM and DW reviewed and edited the manuscript. All authors
read and approved the manuscript and agree to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Ethics Committee of China-Japan Union Hospital of Jilin University.
Written informed consent was obtained from all the patients or
their guardians in accordance with the ethical committee
standards.
Patient consent for publication
Not applicable.
Competing interests
The authors state that they have no competing
interests.
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hamashima C, Shabana M, Okada K, Okamoto M
and Osaki Y: Mortality reduction from gastric cancer by endoscopic
and radiographic screening. Cancer Sci. 106:1744–1749. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Warburg O: On the origin of cancer cells.
Science. 123:309–314. 1956. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Warburg O: On respiratory impairment in
cancer cells. Science. 124:269–270. 1956.PubMed/NCBI
|
|
6
|
Bresters TW, de Kok A and Veeger C: The
pyruvate-dehydrogenase complex from Azotobacter vinelandii.
2. Regulation of the activity. Eur J Biochem. 59:347–353. 1975.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim JW, Tchernyshyov I, Semenza GL and
Dang CV: HIF-1-mediated expression of pyruvate dehydrogenase
kinase: A metabolic switch required for cellular adaptation to
hypoxia. Cell Metab. 3:177–185. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dupuy F, Tabaries S, Andrzejewski S, Dong
Z, Blagih J, Annis MG, Omeroglu A, Gao D, Leung S, Amir E, et al:
PDK1-dependent metabolic reprogramming dictates metastatic
potential in breast cancer. Cell Metab. 22:577–589. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li Y, Li X, Zhong Y, Ji Y, Yu D, Zhang M,
Wen JG, Zhang H, Goscinski MA, Nesland JM, et al: PDHA1 gene
knockout in prostate cancer cells results in metabolic
reprogramming towards greater glutamine dependence. Oncotarget.
7:53837–53852. 2016.PubMed/NCBI
|
|
10
|
Liu F, Zhang W, You X, Liu Y, Li Y, Wang
Z, Wang Y, Zhang X and Ye L: The oncoprotein HBXIP promotes glucose
metabolism reprogramming via downregulating SCO2 and PDHA1 in
breast cancer. Oncotarget. 6:27199–27213. 2015.PubMed/NCBI
|
|
11
|
Li Y, Huang R, Li X, Li X, Yu D, Zhang M,
Wen J, Goscinski MA, Trope CG, Nesland JM, et al: Decreased
expression of pyruvate dehydrogenase A1 predicts an unfavorable
prognosis in ovarian carcinoma. Am J Cancer Res. 6:2076–2087.
2016.PubMed/NCBI
|
|
12
|
Lin CS, Lee HT, Lee MH, Pan SC, Ke CY,
Chiu AW and Wei YH: Role of mitochondrial DNA copy number
alteration in human renal cell carcinoma. Int J Mol Sci.
17:E8142016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Filipowicz W, Bhattacharyya SN and
Sonenberg N: Mechanisms of post-transcriptional regulation by
microRNAs: Are the answers in sight? Nat Rev Genet. 9:102–114.
2008. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hao NB, He YF, Li XQ, Wang K and Wang RL:
The role of miRNA and lncRNA in gastric cancer. Oncotarget.
8:81572–81582. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bartels CL and Tsongalis GJ: MicroRNAs:
Novel biomarkers for human cancer. Clin Chem. 55:623–631. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lai WF and Siu PM: MicroRNAs as regulators
of cutaneous wound healing. J Biosci. 39:519–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Farh KK, Grimson A, Jan C, Lewis BP,
Johnston WK, Lim LP, Burge CB and Bartel DP: The widespread impact
of mammalian MicroRNAs on mRNA repression and evolution. Science.
310:1817–1821. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yi R, O'Carroll D, Pasolli HA, Zhang Z,
Dietrich FS, Tarakhovsky A and Fuchs E: Morphogenesis in skin is
governed by discrete sets of differentially expressed microRNAs.
Nat Genet. 38:356–362. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kloosterman WP and Plasterk RH: The
diverse functions of microRNAs in animal development and disease.
Dev Cell. 11:441–450. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Masoudi MS, Mehrabian E and Mirzaei H:
MiR-21: A key player in glioblastoma pathogenesis. J Cell Biochem.
119:1285–1290. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Han JG, Jiang YD, Zhang CH, Yang YM, Pang
D, Song YN and Zhang GQ: A novel panel of serum
miR-21/miR-155/miR-365 as a potential diagnostic biomarker for
breast cancer. Ann Surg Treat Res. 92:55–66. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sekar D, Krishnan R, Thirugnanasambantham
K, Rajasekaran B, Islam VI and Sekar P: Significance of microRNA 21
in gastric cancer. Clin Res Hepatol Gastroenterol. 40:538–545.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang P, Guan Q, Zhou D, Yu Z, Song Y and
Qiu W: miR-21 inhibitors modulate biological functions of gastric
cancer cells via PTEN/PI3K/mTOR Pathway. DNA Cell Biol. 37:38–45.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shen KH, Hung JH, Chang CW, Weng YT, Wu MJ
and Chen PS: Solasodine inhibits invasion of human lung cancer cell
through downregulation of miR-21 and MMPs expression. Chem Biol
Interact. 268:129–135. 2017. View Article : Google Scholar : PubMed/NCBI
|