Introduction
To date, short microRNAs (miRNAs/miRs), long
non-coding RNAs (lncRNAs) and circular RNAs (circRNAs), which
together comprise non-coding RNAs (ncRNAs), have been validated
(1,2). They account for ~95% of the total RNA
in eukaryotic transcription (3,4).
CircRNAs are produced from the back-splicing of exons and/or
introns, forming exonic or intronic circRNAs, respectively
(3). As a novel type of endogenous
ncRNAs, circRNAs were first observed more than 40 years ago by
Sanger et al (4–6). Originally, they were thought to be
errors of splicing without any specific function and to be fairly
rare (7–9), as circRNAs have only been identified
in a few mammalian genes, such as ETS proto-oncogene 1
transcription factor (10) and sex
determining region Y (SRY) (11).
Subsequently, an increasing number of circRNAs have been
discovered, and their roles in cancer have been determined through
advances in high-throughput RNA sequencing technology (RNA-Seq) and
bioinformatic methods (11,12). There is also an increasing interest
in their biofunction as miRNA sponges (7,13–15),
in regulating gene expression and transcription (7,16), and
also as RNA-binding protein (RBP) sponges (17–19).
Cell proliferation, migration, invasion and metastasis are also
related to the function of circRNAs; therefore, they may
potentially be neoteric biomarkers and remedial targets for cancer
therapy (20–23). In recent years, some previous
studies have reported that circRNAs are stable, abundant and
conserved, presenting tissue- and cell-specific expression patterns
(24–26).
In the present review, we briefly delineate the
biogenesis, characteristics and biofunctions of circRNAs. We then
utilize databases such as CircNet to evaluate known circRNAs, and
how they are involved in specific processes and human diseases such
as cancer. Furthermore, we discuss how circRNAs combine with miRNA
sponges, regulating gene transcription and expression, and with RBP
sponges in different cancers, confirming the clinical value of
circRNAs in tumor diagnosis, treatment and prognosis.
Biogenesis of circRNAs
On the basis of various biogenetic patterns,
circRNAs can be divided into the following three categories: exonic
circRNAs (EcircRNAs) (7), circular
intronic RNAs (ciRNAs) (27) and
exon-intron circRNAs (EIciRNAs) (16) (Fig.
1).
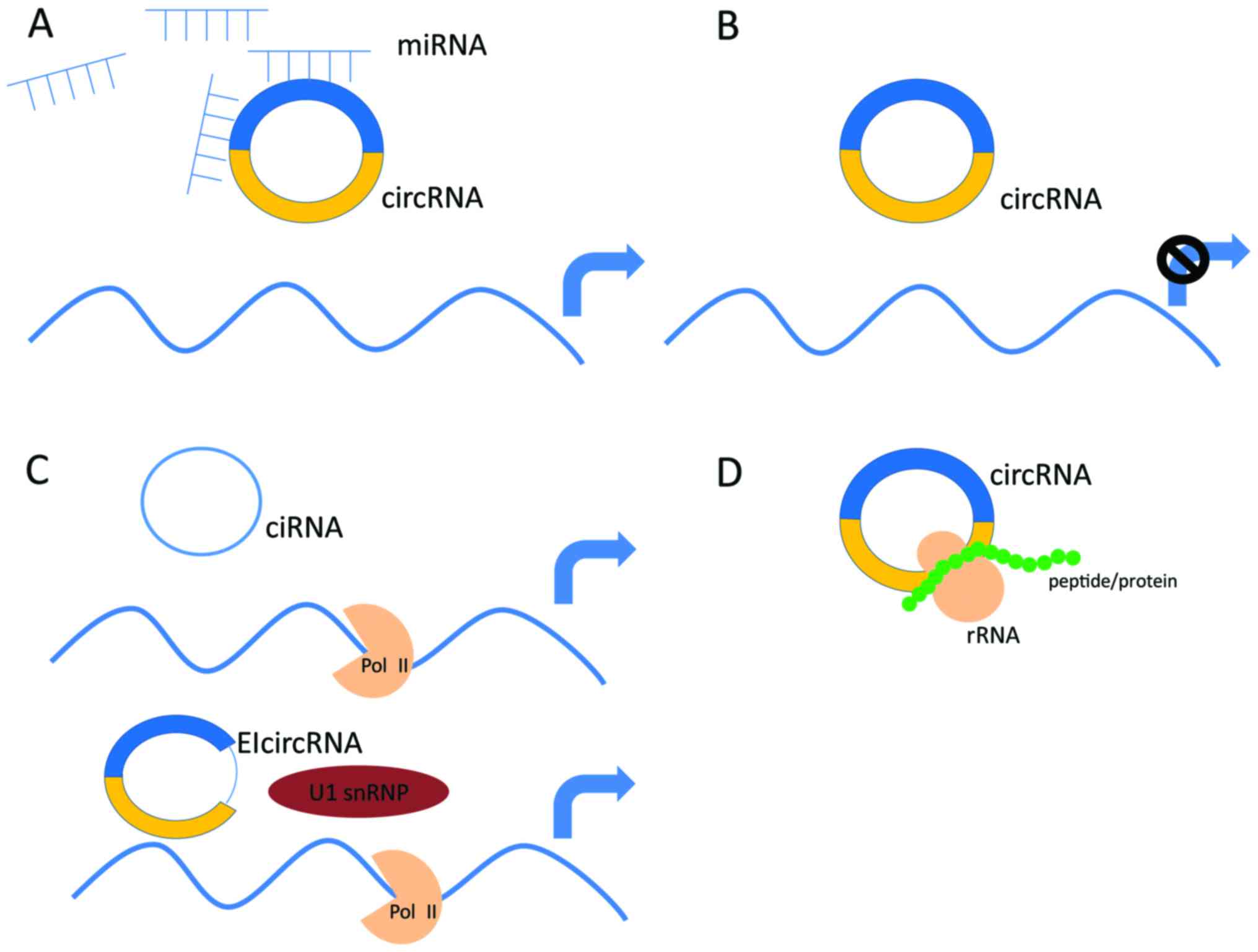
 | Figure 1.The mechanism underlying the
synthesis of four types of circRNAs. In the nucleus of eukaryotic
cells, DNA is transcribed to form hnRNA or pre-mRNA, which contain
coding exons and introns. Based on the above products, circRNAs are
generated via different splicing methods. (A) Intron-pairing-driven
circularization. Two complementary introns form a circular
structure containing several introns and exons through a
base-pairing connection. Finally, introns are removed to form
EcircRNAs, which can also lead to the formation of EIciRNAs. (B)
Lariat-driven circularization. The exon 1 at the 3′ end provides an
SD site and the 5′ front end of exon 4 provides a covalent splice
acceptor site after exon skipping. Then, a lasso-circular structure
containing exons 2 and 3 forms. EcircRNAs are produced when introns
are removed. (C) RBP-driven circularization. The binding of RBPs
acts as a vehicle that binds two non-adjacent introns. Then
circRNAs are generated after the removal of introns. (D) ciRNAs.
Introns form a lasso modality after splicing that is removed
following dissociation, thereby forming stable circular loops via
binding with the 5-splice site (orange box), which is rich in ‘C’
residues and the branch point (purple box) of ‘GU-rich’ sequences;
this generates the ciRNAs after debranching. The majority of the
aforementioned circRNAs are produced in the nucleus. Due to their
unique circular structure they can enter the cytoplasm, as well as
the intercellular substance and blood through exosomes, which are
extremely stable and are not easily degraded by RNA nucleases.
hnRNA, heterogeneous nuclear RNA; circRNA, circular RNA; ciRNA,
circular intronic RNA; pre-mRNA, precursor messenger RNA; EcircRNA,
exonic circular RNA; EIcircRNA, exon-intron circular RNA; SD,
splice donor; RBP, RNA-binding protein. |
EcircRNAs
The most plentiful circRNAs are EcircRNAs. The
majority are derived from the coding gene of pre-mRNA, but do not
involve coding proteins (7,28,29).
Commonly, there is sequential splicing in eukaryotic cells
resulting in the removal of non-coding introns by alternative
splicing of pre-mRNAs after transcription; this generates connected
exons containing protein-coding genes which form translatable
directly linear RNAs that produce the relevant protein (30). Nevertheless, exon sequences of genes
are linked end-to-end, namely the EcircRNAs shaped by
back-splicing. According to published reviews, there are generally
two models of back-splicing (7):
Lariat-driven circularization and intron-pairing-driven
circularization. The former suggests that during the transcription
of pre-mRNA, the pre-mRNA is partially folded and then segmental
splicing of the RNA [i.e. the splice donor of one exon is
covalently connected with the different exon(s), shortening the
distance between the original non-adjacent exons] gives rise to
exon skipping and the shape of a circRNA model. Potentially, each
of the exons can be skipped by transcription. Afterwards, the
EcircRNA is produced through intra-lariat splicing (31).
The intron-pairing-driven circularization pattern
has also been well established. In this pattern, exons involved in
circularization are linked to introns containing reciprocal
complementary sequences that produce spatially close and end-to-end
splice exons to generate circRNAs. However, it is worth noting that
not all circRNAs are produced by complementary sequences on either
side of an exon. Wang and Wang (32) constructed a model of a minigene
in vitro and concluded that the reciprocal complementation
between upstream or downstream exon-flanking introns was not
necessary for the production of circRNAs. In addition, in 2014, a
negative correlation between linear mRNA splicing and back-splicing
of circRNAs was identified by Ashwal-Fluss et al (33).
There is a common feature in both of these models
described. Intron-pairing introns can trigger exon circularization
(27). In addition, the human
genome contains a mass of complementary sequences whose pairing can
produce diverse circRNAs by alternative circularization (34).
Circular intronic RNAs (ciRNAs)
Intron regions account for at least 20% of the human
genome, and the majority form a lasso-like structure (28). However, several introns containing
key nucleotide sequences via the formation of intron-derived
ciRNAs, cannot debranch after splicing and replacing (34,35).
It has been well established that the formation of ciRNAs relies on
conserved sequences at both ends of the intron. These sequences
promote debranching and produce a circRNA (27). Additional protein factors that have
an impact on the formation of ciRNAs have not been defined. The
formation of ciRNAs is quite different from that of EcircRNAs.
CiRNAs are comprised of 2′-5′phospholipid-linked nucleotides, and
EcircRNAs are 3′-5′phospholipid-linked nucleotides (36). CiRNAs are located in the nuclei and
are capable of binding and affecting the expression of the original
genes (37).
EIcircRNAs
Within the back-splicing process that forms
EcircRNAs, circRNAs containing unspliced introns are present and
are stable (16). These introns
possibly act as mediators of the splicing process, or produce
specific circRNAs. By means of RNA sequencing and bioinformatics
analysis, Li et al (38)
discovered >100 circRNAs that interact with polymerase (Pol) II
by presetting a RNA Pol II CLIP sample in human cells
(immobilization of Pol II and RNA after immunoprecipitation of Pol
II antibody). Further studies have reported that these types of
circRNAs can also be formed via exon back-splicing; however, these
circRNAs simultaneously contain exons and introns that are
abundantly localized in the nuclei (39–41).
Therefore, these distinct circRNAs were called retained-intron
circRNAs.
Characteristics of circRNAs
According to the published literature, there are
several important traits of circRNAs. i) Plentifulness: Salzman
et al (29) first suggested
that circRNAs are distributed more widely than linear RNAs in
mammals. ii) Stability: As circRNAs are covalently closed loops,
and thus protected from RNase R, it has also been proposed that
they are more stable than linear mRNAs (29). iii) Conservation: circRNAs are
highly conserved across different species. A number of circRNAs
have been detected in humans, mice, fruit flies and even plants
(7,42). iv) Situation: circRNAs are primarily
composed of EcircRNAs and are predominantly located in the
cytoplasm and possibly in miRNA response elements (MREs) (13,43).
Intronic circRNAs including ciRNAs and EIciRNAs, are both mainly
located in the nuclei, and can therefore regulate gene
transcription or post-transcription (7,27). v)
Expression: circRNAs display tissue and/or development-specific
expression patterns. For example, circRNAs can be easily detected
in mammals, especially during synaptic and neuronal
differentiation, as their expression levels are markedly increased
(44). vi) Functionality: A few
circRNAs contain miRNA binding sites which can combine with miRNA
in order to weaken the endogenous miRNA-mediated mRNA binding
activities via competitive binding (7).
Biofunctions of circRNAs
CircRNAs can function as miRNA sponges, gene
transcription and expression regulators, RBP sponges, and can also
regulate gene expression levels to affect protein/peptide
translators at either the transcriptional or post-transcriptional
level (Fig. 2).
CircRNAs act as DNA replication
regulators
DNA replication is a process by which two copies of
DNA molecules are duplicated from the original template. During DNA
replication, circRNAs are widely distributed in the nuclei, and can
be paired with DNA to form DNA-RNA triple helixes, which affect DNA
replication as they bind with the corresponding DNA strand.
Recently, Schmitz et al (45) reported that pRNA can interact with
transcription termination factor-I, a target transcription factor,
to form a DNA-RNA triplex and consequently recruit DNA
methyltransferase 3β to inhibit ribosomal RNA expression.
Additionally, lnc-Ras association domain family member 1 (RASSF1)
binds with DNA to form a DNA/RNA complex at the transcription
initiation site in order to inhibit the transcription of RASSF1A
(46). Previous evidence indicates
that the interactions between DNA and nucleic circRNAs may be
similar to those of linear ncRNAs (47).
CircRNAs act as miRNA sponges
miRNAs are a class of short (~22 nt) and common
ncRNAs that regulate gene expression after transcription via the
direct base pairing of miRNA target sites (48). As regulators of competitive
endogenous RNAs (ceRNAs), circRNAs can negatively regulate the
activity of miRNAs by competing with mRNA-miRNA binding (49). For example, ciRS-7 [the circRNA
sponge for miR-7, used in the transcription of human cerebellar
degeneration-related protein 1 (CDR1)] contains over 70 selectively
preserved miRNA target sites. CiRS-7 is derived from the antisense
transcript of the CDR1 gene and can be endonucleolytically cleaved
at an miR-7 target site via close binding with miR-671 in an
Argonaute2 (AGO2)-dependent manner. Therefore, due to lysing of
ciRS-7, miR-7 can be freed. It has been identified that the
increase in the expression of miRNA target genes is as a result of
the overexpression of CDR1/ciRS-7. Furthermore, SRY and testis
specific circRNAs could act as an miR-138 sponge to regulate gene
expression (12). Currently,
serving as a miRNA sponge is thought to be the major function of
certain circRNAs.
CircRNAs act as gene expression
regulators
Previous studies have indicated that circRNAs may
serve key roles in gene expression regulation (13,50,51).
Zhang et al (27) and
Memczak et al (13)
demonstrated that circRNAs could regulate parental gene expression,
and that circRNAs are widely detected in the nuclei with a small
concentration on miRNA targets; most notably, inhibition of ciRNAs
may have led to the reduced expression of their parental genes.
Moreover, in-depth studies have revealed that EIciRNAs,
circ-eukaryotic translation initiation factor 3 subunit J and
circ-poly (A) binding protein interacting protein 2, are primarily
distributed in the nuclei, interacting with the U1 small nuclear
ribonucleoprotein particle and RNA Pol II to enhance parental gene
transcription in a cis-acting manner (38). In addition, a ciRNA, such as
ci-ankyrin repeat domain 52 (ankrd52), could interact with
elongation Pol II machinery and actively regulate Pol II
transcriptional activity via enrichment of the transcription sites
of parental genes. Thus, knockout of ci-ankrd52 may reduce parental
gene expression (27).
From these results, we hypothesize that for circRNAs
containing intronic sequences, namely EIciRNAs and ciRNAs,
transcriptional activation may be their general function; in
addition, their regulatory functions likely explain the rich
existence of EIciRNAs and ciRNAs in the cell nuclei.
CircRNAs act as RBP sponges
RBPs participate in multiple biological activities,
including cell proliferation, motility, apoptosis, senescence and
cell responses to oxidative stress via posttranscriptional
regulation such as RNA alternative splicing, conservation,
transport and translation (52,53).
Previous studies have demonstrated that circRNAs can act as sponges
for RBPs by stably binding with trinucleotide repeat containing 6A
(18), RNA quaking homolog KH
domain containing RNA binding (19), mannose binding lectin (33), AGO proteins (12), Pol II (54), and eukaryotic initiation factor
4A-III (55), to form RNA-protein
complexes (RPCs). These RPCs may control the library of RBPs or
small RNAs, and then exert mutual effects with their linear RNA
counterparts (29). In addition,
CircInteractome, a new online tool to analyze RBP binding sites on
human circRNAs, can predict the potential circRNAs binding sites
for miRNAs for a given RBP (56).
For instance, hsa_circ_0024707 has 85 predicted positions as a
sponge for AGO2, and the relatively mature hsa_circ_0000020
contains some RBP binding sites, such as circ-fragile X mental
retardation 1 which contains 10 sites and circ-ELAV like RNA
binding protein 1 which has 6 sites (56).
CircRNAs act as translation product
translators
As a number of circRNAs can bind open reading frames
(ORFs), one hypothesis is that they can be translated as protein
fragments. Perriman and Ares (57)
revealed that a simple green fluorescent protein ORF in a circRNA
could be expressed directly in Colibacillus. In addition, a
circRNA (220 nt) of a virus related to rice yellow spot virus that
could code for a 16-kDa protein was discovered by AbouHaidar et
al (58). It has also been
reported that protein/polypeptides can be coded by circRNAs both
in vitro (59) and in
vivo (57); however, only when
the RNAs contain prokaryotic ribosome-binding sites (57), or internal ribosome entry site
elements (IRES) unlike their canonical counterparts. Recently, the
circRNA database, circRNADb, which contains >32,000 human
EcircRNAs, was established (60).
It offers detailed information on the circRNAs, including their
sequence, ORF and IRES for users to predict the translatability of
certain circRNAs. However, there is no well-founded evidence to
verify how EcircRNAs that are generated from spliceosomes can act
as mRNAs.
In addition to acting as miRNA sponges and
transcriptional regulators, circRNAs can also positively act as
mRNA traps that regulate protein expression by isolating the
translation initiation point (61).
For the first time, Yang et al (62) recently reported that
N6-methyladenosine (m6A) is the most common
and abundant base modification of RNA and promotes the protein
translation of circRNAs in the human body. The group demonstrated
that m6A motifs contained many circRNAs and only one
m6A was required to drive translation initiation. Most
importantly, translation of m6A-driven circRNAs was
quite common. A large number of endogenous circRNAs have the
potential for translation, indicating that they may have a
regulatory role in the stimulation of circRNA-derived proteins in
cell responses to environmental stressors.
Metabolism of circRNAs
CircRNAs are highly stable due to their unique
structure; however, the underlying mechanisms of cell degradation
and/or detectable circRNAs have not been elucidated. A previous
study suggested that during their experiments, when multiple
circRNAs were secreted from three different types of cells, the
cells may have removed circRNAs through vesicles, such as exosomes
and microcapsules (63). This
phenomenon indicated that circRNA concentrations may exceed their
linear counterparts during extracellular vesicle (EV) preparation
when compared to the cell, implying that the mechanism underlying
the reduction in circRNAs may be via EV release from the cells into
the extracellular space.
Public databases on circRNAs
To perform classification of circRNAs, the present
review evaluated the current online databases (Table I). These databases describe circRNAs
from published articles and GenBank annotations. They list circRNAs
that have been experimentally confirmed, those that are
computational predictions and those that are based on the ORF
predicted size. Herein, we list several key features of the
databases.
 | Table I.Public circRNA databases. |
Table I.
Public circRNA databases.
| Name (ref.) | starBase v2.0
(84) | circBase (66) | circ2Traits
(56) | CircNet (67) | CircInteractome
(58) | CircBANK (84) | circRNADb (62) |
|---|
| Website | http://starbase.sysu.edu.cn | http://www.circbase.org/ | http://gyanxet-beta.com/circdb/ | http://circnet.mbc.nctu.edu.tw/ | http://circinteractome.nia.nih.gov | http://www.circbank.cn/ | http://reprod.njmu.edu.cn/circrnadb |
| circRNA disease
association | Not available | Not available | 105 diseases | Not available | Not available | Not available | Not available |
| circRNA
annotation | Not available | Not available | 1,953 human
circRNAs | 212,950
circRNAs | Not available | 140,790 human
circRNAs | 32,914 human
circRNAs of exons |
| Sequence
alignment | Not available | A web interface of
BLAST | A web interface of
BLAST | Not available | Not available | A web interface of
BLAST | Not available |
| circRNA reference
source | circBase v0.1 | Back-spliced
junction sites in animals reported in 2013 and 2015 | circRNA dataset
from Memczak et al 2013 to 2015 | Reported human
circRNAs from | CircBase, starBase
V.2.0, IRESite (6) | CircBase, reported
human circRNAs from 2013 to 2015 | CircBase,
starBaseV.2.0, IRESite |
| circRNA position on
the genome | A customized genome
browser accessible through keyword search | Linked out to UCSC
genome browser | A customised genome
browser | An integrated
genome browser synchronized with the network graphical user
interface | Linked out to UCSC
genome browser | A customized genome
browser accessible through keyword search | A customized genome
browser accessible through keyword search |
| circRNA sample
source | circBase v0.1
source samples | The samples where
the back-spliced junction sites were discovered | Not available | i. For which sample
junction sites were discovered. ii. Expression level in available
samples. iii. Clustered sample conditions | Not available | The samples where
the back-spliced junction sites were discovered | Not available |
| circRNA naming | Same as circBase,
except CDR1 antisense (CDR1as) | A serial number for
every detected back-spliced junction site | A serial number for
every detected back-spliced junction site | A systematic naming
system which provides information to the source gene and annotated
exons of circRNAs | Same as
circBase | Same as circBase
host gene; circBANK ID | A serial number for
every detected back-spliced junction site |
| circRNA expression
profiles in samples | Not available | Not available | Not available | An all-sample
expression heat-map for every circRNA and linear isoform | Not available | Not available | Not available |
| Address on miRNA
regulatory relationships | Identifies circRNA
and miRNA interactions through Chip-Seq data analysis | Not available | Identifies circRNA
and miRNA interactions | A network-driven
graphical interface shows the relationship between miRNA target
genes and circRNAs | Identifies circRNA
and miRNA interactions | Identifies circRNA
and miRNA interactions | Not available |
| RNA-binding
protein | Not available | Not available | Not available | Not available | Identifies
RNA-binding protein | RNA-binding protein
potential | Not available |
| CircRNA
isoforms | Not available | Not available | Not available | All traceable on
the integrated genome browser | Not available | Not available | Not available |
i) Circbase: Basic information analysis of circRNAs.
The latest version of the database was released in June 2017, and
it can assist with gaining a specific understanding of the
screening and verification of circRNAs; it is also possible to
directly download relevant sequence and annotation information
(64). ii) CircNet: ceRNA
functional research database. Using 464 RNA-seq sequencing
datasets, new circRNA predictions and genome annotations were
performed, and known and newly predicted circRNA expression was
calculated to construct a circRNA-miRNA-gene regulatory network
from which the above information was obtained (65). iii) Circ2Traits: A circRNA database
that generates potential associations with human diseases or traits
to construct an interaction network by predicting the interactions
between miRNAs and protein-coding genes, long non-coding genes, and
circRNAs, thereby constructing an interactive network for Gene
Ontology enrichment analysis of protein-coding genes in
miRNAs-circRNA interaction groups (54). iv) CircRNADb: circRNA-encoded
protein database. A total of 32,914 human exon circRNA records were
included, each of which included genomic location information, RNA
editing, corresponding genomic sequences, IRES sequence elements,
predicted ORFs, and the related references (60). v) CircInteractome: A predictive
binding target database. The binding sites of the known 109 RNA
binding proteins to circRNAs in circbase were predicted and the
potential binding sites for miRNAs to circRNA were predicted using
Targetscan software. This software also performs circRNA molecular
searches, circRNA binding protein prediction, polymerase chain
reaction (PCR) primer design and small interfering (si)RNA
interference sequence design. It also displays the RBP binding site
of circRNA, the possibility of analyzing circRNA as an RBP protein
‘sponge’ or ‘decoy’, and the RBP protein information that may be
bound by the junction and flanking sequences of the circRNA
(56). vi) CircBank: A total of
140,790 human circRNAs were recorded in the circBank database, and
each circRNA record contains detailed information. The information
for each circRNA mainly includes: The detailed sequence of the
circRNA, the more homologous circRNA and its corresponding sequence
in mice, the predictive analysis of miRNA binding, the ORF
predictive information, the mutations and polymorphisms recorded by
COSMIC Site summary, and the m6A modification
information (64). Notwithstanding
the cancer-associated expression datasets, we expect the circRNA
expression database, as displayed in Table I, to merge with other datasets to
become more disease-oriented.
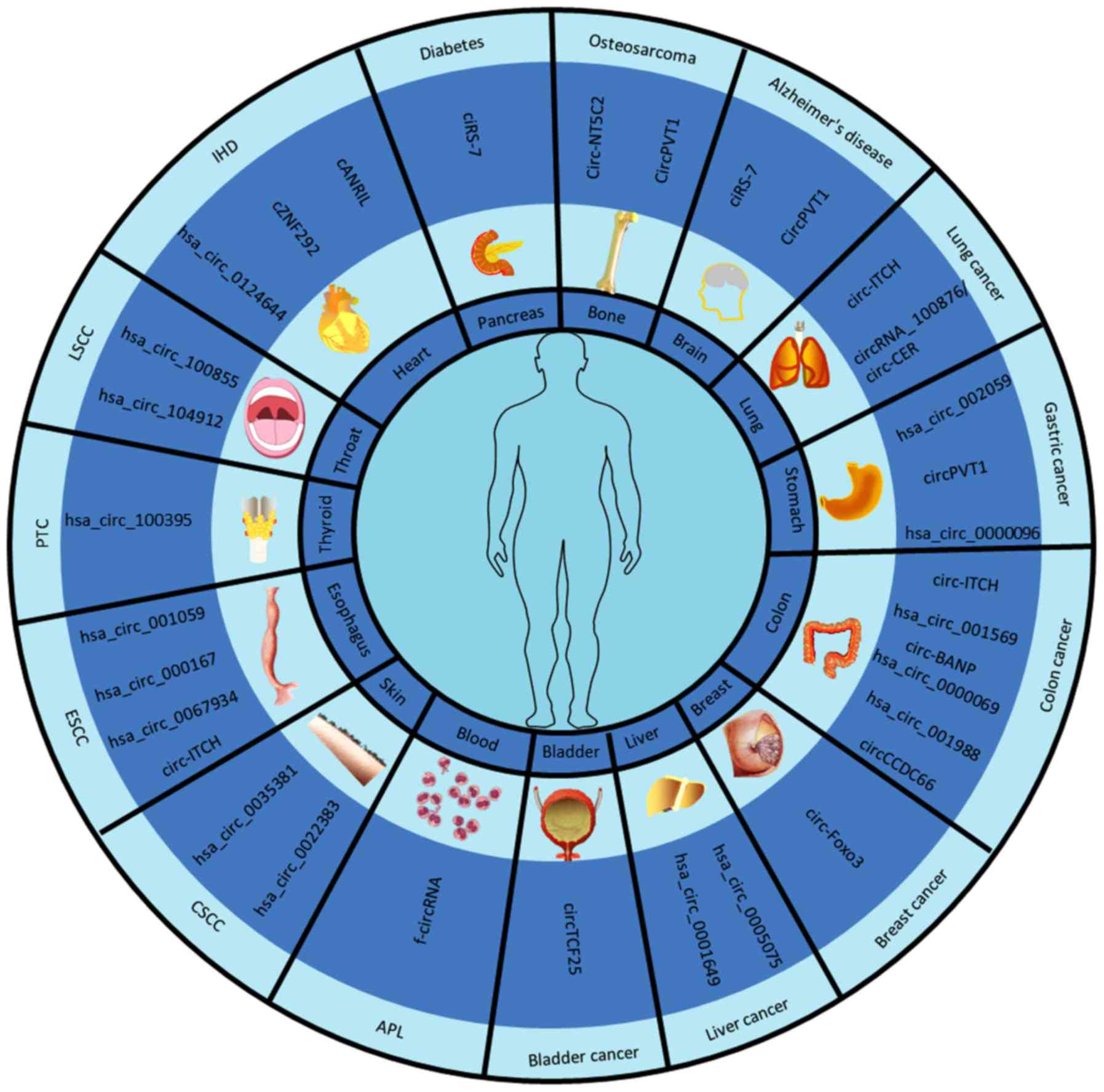
CircRNAs and human disease
Based on the pivotal role of genetic regulation, the
altered expression of circRNAs associated with specific diseases is
beginning to be increasingly reported. Altered circRNA expression
has been observed in several diseases including diabetes,
atherosclerosis, heart disease and neurological disorders (Table II).
| Table II.circRNAs identified in human
diseases. |
Table II.
circRNAs identified in human
diseases.
| Disease | circRNAs | Function | (Refs.) |
|---|
| Ischaemic heart
Disease (IHD) | cANRIL | An increased risk
of atherosclerosis by repressing of the INK4A/ARF locus | (49) |
|
| cZNF292 | Regulation of
hypoxia in endothelial cells and control of angiogenesis | (12) |
|
|
hsa_circ_0124644 | Upregulated in
coronary artery disease | (52) |
| Alzheimer's
disease | ciRS-7 | Upregulated UBE2A
that aids the clearance of amyloid peptides | (54) |
|
| circPVT1 | Cell senescence
inhibitor | (58) |
| Diabetes | ciRS-7 | Inhibits miR-7
function in islet β cells, which can inversely improve secretion
insulin | (59) |
CircRNAs and cancer
Recent studies have suggested that circRNAs may have
a pivotal role during the development and progression of different
types of cancers (66–68). CircRNAs have become a novel area of
interest in the early diagnosis and therapy of cancers due to their
abundance, high stability and notable regulatory functions. The
fact that circRNAs appear to behave as miRNA sponges has also
increased this interest (69). The
expression of an miRNA is dysregulated via different mechanisms in
humans, such as miRNA gene amplification or deletion, the abnormal
transcriptional regulation of miRNAs, and epigenetic changes in
miRNA generation (70). Zheng et
al (71) analyzed a total of
six human normal tissues (brain, heart, lung, liver, colon, and
stomach) and seven human cancers (bladder cancer, breast cancer,
hepatocellular carcinoma, gastric cancer, colorectal cancer, renal
clear cell and prostate adenocarcinoma) to obtain the sequencing
data of over 27,000 circRNA candidate genes. Compared to normal
tissues, circRNAs were usually downregulated in tumor tissues and
circRNA levels were significantly associated with clinical features
including staging, age, gender and distant metastasis, due to
errors in the back-splicing machinery of malignant tumors, the
dysregulation of miRNAs due to the degradation of circRNAs in
cancerous tissues, or the reduction in circRNAs as a result of
increasing cell proliferation (72). In addition, circRNAs are observed
widely in exosomes (73),
illustrating that a potential future strategy to diagnose cancer
may involve detecting blood plasma circRNAs. Herein, we discuss the
latest discoveries focusing on abnormal circRNAs in cancer
(Table III; Fig. 3).
| Table III.Summary of circRNAs in a series of
cancers. |
Table III.
Summary of circRNAs in a series of
cancers.
| Cancer type | Name of
circRNA | Expression
level | Scope | Sponge target
and/or pathway | Samples | (Refs.) |
|---|
| Gastric cancer |
hsa_circ_002059 | Down | Gastric cancer vs.
normal | – | Tissues
(n=101) | (67) |
|
| circPVT1 | Up | Gastric cancer vs.
normal | miR-125a/b; G0/G1
to S phase | – | (68) |
|
|
hsa_circ_0000096 | Down | Gastric cancer vs.
normal | miR-224,
miR-200a | Tissues (n=17) | (68) |
| Lung cancer | circ-ITCH | Down | LCSS vs.
normal | miR-7, miR-214;
Wnt/β-catenin signaling pathway; cell cycle progression | Tissues (n=78) | (70) |
|
| circRNA_
100876/circ-CER | Up | NSCLC vs.
normal | – | Tissues
(n=101) | (71) |
| Colorectal
cancer | circ-ITCH | Down | CRC vs. normal | miR-7, miR-20a,
miR-214 | Tissues (n=45) | (69) |
|
|
hsa_circ_001569 | Up | CRC vs. normal | miR-145 | Tissues (n=30) | (18) |
|
| circ-BANP | Up | CRC vs. normal | – | Tissues
(n=2,608) | (77) |
|
|
hsa_circ_0000069 | Up | CRC vs. normal | Cell cycle
progression | Tissues (n=30) | (78) |
|
|
hsa_circ_001988 | Down | CRC vs. normal | – | Tissues (n=31) | (79) |
|
| circCCDC66 | Up | CRC vs. normal | miR-33b,
miR-93 | Tissues (n=48) | (81) |
| Esophageal
cancer |
hsa_circ_001059 | Up | ESCC vs.
normal | miR-30c-1,
miR-30c-2, miR-122, miR-139-3p, miR-339-5p, miR-1912 | – | (82) |
|
|
hsa_circ_000167 | Down | ESCC vs.
normal | miR-181a-2,
miR-512-5p, miR-521, miR-556-5p, miR-663b, miR-1204 |
|
|
|
|
hsa_circ_0067934 | Up | ESCC vs.
normal | – | Tissues (n=51) | (83) |
|
| circ-ITCH | Down | ESCC vs.
normal | miR-7, miR-17,
miR-214; Wnt/β-catenin pathway | Tissues
(n=684) | (16) |
| Breast cancer | circ-Foxo3 | Down | Breast cancer vs.
normal | miR-22, miR-136,
miR-138, miR-149, miR-433, miR-762, miR-3614-5p, miR-3622b-5p | – | (89,90) |
| Liver cancer |
hsa_circ_0005075 | Up | Liver cancer vs.
normal | miR-23b-5p,
miR-93-3p, miR-581, miR-23a-5p | Tissues (n=61) | (92) |
|
|
hsa_circ_0001649 | Up | Liver cancer vs.
normal | miR-7 | Tissues (n=89) | (96) |
| Bladder cancer | circTCF25 | Up | Bladder cancer vs.
normal | miR-103a-3p,
miR-107; PI3K-Akt signaling, CDK6, proliferation and migration | Tissues (n=40) | (99) |
| Acute promyelocytic
leukemia | f-circRNA | – |
f-circRNA-expressing vs. normal | PI3K and MAPK
signal transduction pathways | Cells | (100) |
| Osteosarcoma
(OS) | circ-NT5C2 | Up | OS vs. normal | miR-448 | Tissues (n=4) | (103) |
|
| circPVT1 | Up |
| slicing
gene-ABCB1 | Tissues (n=80) | (104) |
| Cutaneous
squamous |
hsa_circ_0035381 | Up | cSCC vs.
normal | miR-124-5p,
miR-9-5p | Tissues (n=12) | (106) |
| cell carcinoma
(cSCC) |
hsa_circ_0022383 | Down | miR-124-5p |
|
|
|
| Papillary thyroid
cancer (PTC) |
hsa_circ_100395 | Down | PTC vs. normal | miR-141-3p,
miR-200a-3p | Tissues (n=18) | (107) |
| Laryngeal
squamous |
hsa_circ_100855 | Up | LSCC vs.
normal | – | Tissues (n=52) | (108) |
| cell cancer
(LSCC) |
hsa_circ_104912 | Down | LSCC vs.
normal | – |
|
|
CircRNAs and gastric cancer
Via RNA-seq analysis, 180 circRNAs have been
detected with diverse expression in gastric tumor tissues as
differentiated from normal tissues (74). Hsa_circ_002059, a representative
circRNA, was observed to be downregulated in 101 gastric cancer
tissues (73). The dysregulated
expression of hsa_circ_002059 was markedly correlated with sex, age
of disease onset, tumor-node-metastasis (TNM) stage and distant
metastasis (73). In conclusion,
these results suggest that circRNAs, particularly hsa_circ_002059,
may be potentially stable biomarkers for gastric carcinoma
diagnosis and staging.
Another circRNA, circPVT1, has also been confirmed
to be upregulated in gastric cancer (74). In addition, circPVT1 was found to
suppress miR-125, which is a tumor suppressor family sponge. This
circRNA also indirectly regulated the expression of the
transcription factor, E2F transcription factor 2 (E2F2), as
determined by a luciferase reporter assay, and was found to serve a
crucial role in the progression of the cell cycle and tumor
inhibitor protein functions (74).
Therefore, the abnormal expression of circPVT1 may reduce the
anticancer effect of E2F2 and miR-125b. Furthermore, circPVT1 was
shown to facilitate the colony formation of gastric cancer by
inhibiting miR-125 (74).
Expression of the circRNA, hsa_circ_0000096, also
called circHIAT1, has also been confirmed to be downregulated in
gastric cancer cell lines and cancerous gastric tissues when
compared with non-cancerous epithelial cells and normal gastric
tissues (75). Furthermore, the
knockdown of hsa_circ_0000096 by siRNA has been observed to
significantly inhibit cell proliferation and migration both in
vitro and in vivo, and it also was found to suppress the
progression of the cell cycle, arresting the S phase of cell
transition from G0/G1 in gastric cancer cells (75). In addition, knockdown of
hsa_circ_0000096 has been observed to suppress tumor growth in
xenograft nude mouse models. It is commonly thought that a high
expression of circRNAs may be observed in cancers; however,
tumor-inhibiting genes have been reported in low levels.
Nevertheless, hsa_circ_0000096 seems to be an exception to the
above. This phenomenon could be explained by its interaction with
miRNAs. The circRNA databases have revealed that hsa_circ_0000096
can bind with 17 different miRNAs. More, previous results of
reverse transcription-quantitative PCR (RT-qPCR) have confirmed
that the cluster of differentiation-40 regulator decreases the
levels of miR-224 in a number of gastric cancer cell lines and the
primary function of miR-200a is to increase E-cadherin levels,
following the knockdown of hsa_circ_0000096 (75).
These previous studies have presented the clinical
implications of hsa_circ_002059 and hsa_circ_0000096 as biomarkers,
as determing their expression level is able to distinguish between
normal gastric mucosa and gastric cancer tissue. In addition, these
circRNAs can also be used as molecular biomarkers to evaluate
gastrectomy.
CircRNAs and lung cancer
The circRNA circ-itchy E3 ubiquitin protein ligase
(ITCH) has been confirmed to play a crucial role in suppressing
lung cancer progression by functioning as a miRNA sponge in
multiple carcinogenic miRNAs. The expression of ITCH can be
enhanced by this action and in turn inhibit the Wnt/β-catenin
signaling, which is a significant pathway involved in tumor cell
proliferation and the cell migration process. Previous studies have
verified that circ-ITCH can bind to different types of miRNAs,
including, miR-7, miR-17, miR-128, miR-214, and miR-216b (12,13,76).
Wan et al (77) reported
that circ-ITCH expression was greater in cancerous tissues when
compared with that noted in adjacent non-cancerous tissue and
normal cells of 78 matched patients with lung cancer. The report
demonstrated that >70% of the lung cancer tissues had
significantly decreased expression of circ-ITCH (77). To elucidate the associations between
circ-ITCH and the miRNAs, miR-7 and miR-214, circ-ITCH plasmids
were cotransfected into the human lung cancer cell lines A549 and
H460, and a luciferase reporter of miR-7 or miR-214 was
established; the luciferase activity was then detected. After
transfection with an empty vector in both cell lines (including the
control group containing plasmid with circ-ITCH), the luciferase
activity was significantly reduced in a concentration-dependent
manner by miR-7 or miR-214 mimics. However, no obvious change in
luciferase activity was observed in cells with circ-ITCH
overexpression. Therefore, these results indicated that circ-ITCH
may suppress miR-7 and miR-214 by binding to them as a sponge in
lung cancer tissues (77).
Yao et al (78) confirmed that in tissues from 101
patients with non-small cell lung cancer (NSCLC), circRNA_100876
was significantly upregulated when compared with the corresponding
adjacent normal lung tissues. Among the estimated
clinicopathological results, the overexpression of circRNA_100876
was significantly associated with regional lymph node metastasis
and advanced stages of the tumor. Patients with NSCLC and elevated
circRNA_100876 expression exhibited notably shorter overall
survival than those with low expression (78). This therefore suggested that
circ_100876 may be a potential biomarker for tumor cell
proliferation, progression and metastasis in NSCLC (78).
CircRNAs and colorectal cancer
(CRC)
As aforementioned in lung cancer, circ-ITCH was also
observed to have reduced expression in 45 matched colorectal cancer
(CRC) tissues when compared with that noted in adjacent
non-cancerous colorectal tissues (76). In a previous study investigating the
CRC cell lines, SW480 and HCT116, circ-ITCH sponge activity was
demonstrated for miR-7, miR-20a, and miR-214 (76). These miRNAs have been revealed to
downregulate many target genes that are primarily involved in the
regulation and completion of the G1/S transitional phase, including
the proliferative target gene cyclin D1 (76). It has also been suggested that the
aberrant expression of circ-ITCH may inhibit the expression of the
target genes c-Myc and cyclin D1 of the Wnt/β-catenin signaling
pathway (76). These results
indicated that circ-ITCH may be involved in the regulation of the
Wnt/β-catenin signaling pathway, and in turn, the Wnt/β-catenin
signaling pathway may serve a significant regulatory role in cell
proliferation and migration (76).
Consistent with this, overexpression of circ-ITCH was found to
reduce HCT116 and SW480 cell proliferation. Consequently, circ-ITCH
is likely to have an anti-proliferative role in CRCs (76).
Another circRNA, hsa_circ_001569, has also been
demonstrated to exhibit high levels of expression in CRC when
compared with that noted in non-cancerous tissues (54,79,80).
miR-145 could bind hsa_circ_001569, which contains three target
genes [E2F5, B-cell lymphoma 2 associated athanogene 4 (BAG4) and
formin like 2 (FMNL2)], and may be inhibited by the 3,-untranslated
region promoter, as determined by evaluating two public databases
(StarBase v2.0 and circBase) and three bioinformatic websites
(TargetScan, Pictar and miRANDA) (54,64,81). A
previous study reported that E2F5 is a transcription factor that
can control the gene expression of proteins involved in cell cycle
control (82), that BAG4 is
involved in cancer cell invasion (83,84),
and that FMNL2 participates in cell growth, motility, invasion,
metastasis and epithelial-mesenchymal transition (EMT) (85,86).
hsa_circ_001569 has been demonstrated to promote CRC cell
proliferation and invasion by blocking the downregulation of
E2F5/BAG4/FMNL2 by miR-145 (80).
Subsequently, the group revealed that hsa_circ_001569 promotes cell
proliferation by increasing the number of S and G2/M phase cells in
the cell cycle (80). Furthermore,
hsa_circ_001569 was knocked down in SW620 and LoVo cells, which has
been indicated to reverse invasion (80). Notably, unlike the other circRNAs
that have been mentioned, hsa_circ_001569 may directly inhibit the
transcriptional activity of miR-145, rather than inhibiting its
expression (80).
Similarly, CRC tissues have also been revealed to
have high levels of circ-BTG3 associated nuclear protein (BANP)
(87). Knockout of circ-BANP using
siRNA also reduced the proliferation of the CRC cell lines HCT116
and HT29. Furthermore, the expression of phosphorylated (p)-protein
kinase B (Akt) was downregulated via this knockout, indicating that
it may be involved in the phosphoinositide-3 kinase (PI3K)/Akt
signaling pathway, which is known to have roles in controlling
cancer cell survival and the cell cycle (87).
Guo et al (88) demonstrated that hsa_circ_0000069 was
also overexpressed in CRC, as determined by unsupervised
hierarchical cluster analysis. Inhibition of hsa_circ_0000069 with
siRNA may suppress cell proliferation, migration and invasion, as
well as induce HT-29 cell arrest at the G0/G1 phase of the cell
cycle (88).
Hsa_circ_001988 was also found to be downregulated
in CRC cell lines when compared with normal samples in 31 matched
CRC tissues and non-cancerous colon mucosa (89). In addition, the results indicated
that the expression level of hsa_circ_001988 was associated with
cancer cell differentiation, and even perineural invasion. Nerve
infiltration is a clear predictor of the prognosis of CRC patients,
which is related to the survival time and recurrence of CRC
(71,90). These results indicate that
hsa_circ_001988 may be a potential new biomarker of CRC prognosis
(89). Hsiao et al (91) analyzed the expression of circRNAs
and adjacent non-tumor tissues in 48 colorectal tumors. The results
revealed that circ-cyclin B1, circ-cyclin-dependent kinase
(CDK)-13, and circ-coiled-coil domain containing 66 (CCDC66), which
is comprised of exons 8–10, were elevated in CRC tissues (91). Notably, the RT-qPCR results from
multiple cancer cell lines, including colorectal (Caco-2, HCT116,
HT-29, and LS123), mammary gland (MCF-7, MDA-MB-231, and
MDA-MB-468), pancreatic (BxPC-3 and MIA PaCa-2) and cervical cancer
(HeLa) cell lines, demonstrated that there was an abnormal
expression of circCCDC66, which suggested that this circRNA may
have important implications for the majority of oncogenes (91). In addition, circCCDC66 is thought to
regulate a variety of pathological processes, including cell
proliferation, migration and invasion in vivo and in
vitro (91). Furthermore, a
previous study identified 99 potential miRNAs that could bind exons
8–10 of circCCDC66. This evidence suggests that circCCDC66 may act
as an miR sponge to protect the miRNA activity of the oncogene,
Myc, from CRC. It's key role in CRC was confirmed in miR-33b and
miR-93 by knocking out circCCDC66 (91).
CircRNAs and esophageal cancer
Previous studies have reported that several
circRNAs, including hsa_circ_001059, hsa_circ_000167,
hsa_circ_0067934, and circ-ITCH, may be involved in
cancer-associated mortality in esophageal squamous cell carcinoma
(ESCC) (92–94). Su et al (93) identified two distinct circRNAs,
hsa_circ_001059 and hsa_circ_000167, from >3,700 human circRNAs,
that were observed with altered expression levels in the human
anti-radioresistant esophageal cancer cell line KYSE-150R, when
compared with the adjacent normal cell line KYSE-150. CircRNA-miRNA
coexpression network analysis indicated that the two circRNAs were
the key factors in the potential circRNA-miRNA network (93). These results indicated that circRNAs
may have dysregulated expression in esophageal cancer cells during
the development of radiation-resistant expression (93).
The circRNA hsa_circ_0067934 has also been examined
in ESCC tissues and was also significantly overexpressed; it is
thought to be associated with differentiation, as well T stage I–II
and TNM stage I–II (94).
Furthermore, siRNA knockdown of hsa_circ_0067934 activity was
reported to inhibit cell proliferation, migration, and invasion in
ESCC cells (94). When considering
other clinical factors, lymph node metastasis or tumor size was not
associated with the expression of hsa_circ_0067934. Notably, TNM
staging was applied to assess patient prognosis, and it is possible
that hsa_circ_0067934 may also have the potential to become a
biomarker for diagnosing ESCC.
In addition, following analysis of 684 ESCCs and
adjacent non-cancerous tissues, circ-ITCH was shown to be
downregulated in ESCC, acting as a tumor inhibitor gene for lung
cancer and CRC (92). Li et
al (92) revealed that
circ-ITCH can act as a sponge for miR-7, miR-17, and miR-214,
passively regulating the Wnt/β-catenin pathway via the upregulation
of the target gene ITCH. Therefore, this result indicates that
circ-ITCH may be a passive regulator of the Wnt/β-catenin signaling
pathway via the indirect sponge effect of miRNAs (92).
CircRNAs and breast cancer
Approximately one fifth of breast cancer cases
worldwide are diagnosed as ductal carcinoma in situ (DCIS)
via mammography (95). Generally,
DCIS is deemed to be highly curable; however, some women with DCIS
unfortunately suffer from the life-threatening type of invasive
breast cancer, invasive ductal cancer (IDC); even though the
invasive factors of progression remain unclear. This may result in
the same histological subtypes sharing similar patterns of gene
expression (96,97).
Galasso et al (97) analyzed the expression of circRNAs in
the DCIS/IDC samples of five patients and also evaluated the
invasive breast cancer cell line, MCF-7, using published RNA-seq
databases and the bioinformatics testing pipeline, CIRCexplorer.
Hsa_circ_0122662 and hsa_circ_0001358 were both identified in the
IDC samples and the MCF-7 cell line. hsa-circ-0001358 was found to
be associated with five miRNAs, miR-200c-3p, miR-200b-3p,
miR-376a-3p, miR-376b-3p and miR-429, as determined by the Starbase
human Pan Cancer tool (97).
miR-200c-3p, miR-200b-3p, and miR-429, which belong to the miR-200
family, were found to reduce the migration and invasion of breast
cancer cells (98,99). The genes known to participate in
epithelial-mesenchymal transition (EMT) that are also known targets
of the miR-200 family were detected, namely zinc finger E-box
binding homeobox 1 (ZEB1), ZEB2, vimentin, B lymphoma Mo-MLV
insertion region 1 homolog, and fibronectin 1 (97). However, no significant interactions
were identified (97). It is also
worth noting that excess circRNAs were observed to be expressed in
DCIS or IDC, indicating measurable differences in dynamic
expression. As a result, differential circRNA expression may
warrant further exploration in order to further understand the
molecular mechanisms of DCIS progression to IDC.
Circ-forkhead box O3 (Foxo3) has been shown to be
downregulated in breast cancer cell lines and breast tumor tissues
when compared with that noted in adjacent non-cancer cell lines or
non-cancerous tissues (100).
Overexpression of circ-Foxo3 was found to significantly reduce
proliferation and cell survival in the breast cancer cell line,
MDA-MB-231 (101). Du et al
(101) injected MDA-MB-231 cells
transfected with circ-Foxo3 subcutaneously in nude mice,
demonstrating that circ-Foxo3 suppressed tumor growth.
Subsequently, through tumor section TUNEL staining,
circ-Foxo3-transfected tumor cells were observed to exhibit
extensive cell death, suggesting that apoptosis occurred in the
tumor. In addition, 25 additional binding sites were examined in
circ-Foxo3 for eight miRNAs, including miR-22, miR-136, miR-138,
miR-149, miR-433, miR-762, miR-3614-5p, and miR-3622b-5p (100). It was also revealed that when
these miRNAs were transfected into MDA-MB-231 cells, they decreased
cellular apoptosis. Therefore, these studies indicated that
circ-Foxo3 may serve a crucial biological role as a miRNA
sponge.
CircRNAs and hepatocellular
carcinoma
An increasing body of evidence has revealed that
circRNAs are associated with the development of hepatocellular
carcinoma (HCC). By analyzing integrated bioinformatic data, many
circRNAs have been demonstrated to be involved in the
overexpression of miR-181a-3p and thus, the enzyme
O(6)-methylguanine-DNA methyltransferase, which is known to be
involved in DNA disruption, thereby suggesting a potential link
between circRNAs and HCC progression through miRNA regulation
(102). Shang et al
(103) in a recent and extensive
study identified that there are 61 distinctly expressed circRNAs in
HCC tissues when compared with that noted in adjacent normal
tissues. Of the three mentioned above that were further verified
through RT-qPCR, hsa_circ_0005075 was significantly associated with
a number of clinicopathological factors of HCC patients (103). Through pathway analysis and Gene
Ontology of hsa_circ_0005075, it was revealed that this circRNA was
strongly associated with cell adhesion, which is an important
process in tumor cell proliferation and metastasis (103). Furthermore, larger liver cancer
tumors were found to exhibit a higher expression level of
hsa_circ_0005075 than smaller tumors, demonstrating that
hsa_circ_0005075 may regulate tumor growth. Therefore,
hsa_circ_0005075 is likely to be a promising biomarker for HCC
(103). In addition, the four
binding sites, miR-23b-5p, miR-93-3p, miR-581, and miR-23a-5p, for
hsa_circ_0005075 were located within HCC tissues, as determined via
the predictive program Arraystar (DNASTAR, Inc., Madison, WI, USA),
which performs sequence variation analysis (103). Shang et al (103) also demonstrated that miR-23a-5p
has the largest interaction network, following analysis of the
hsa_circ_0005075 circRNA-miR-mRNA network. Interestingly,
miR-23b-5p has been observed to be downregulated in gastric and
esophageal cancer (104,105). It has been hypothesized that
hsa_circ_0005075 may act as a miRNA sponge to suppress the
expression of miR-23b-5p in cancer (103). As described above, the expression
of hsa_circ_0005075 was high in HCC tissues, thus it has been
suggested that it may be involved in tumor progression. However,
further study is required to elucidate the molecular mechanisms
underlying the development of HCC and how hsa_circ_0005075
functions as a miRNA sponge.
Hsa_circ_0001649 has also been indicated to have
decreased expression in HCC tissues, when compared with that
observed in adjacent liver tissues (106). Following the inhibition of
hsa_circ_0001649 using siRNA, the expression levels of the
pro-metastatic matrix metalloproteinases (MMPs), MMP9, MMP10 and
MMP13, were increased (106).
These results demonstrated that the expression of hsa_circ_0001649
is negatively correlated with the metastasis of HCC and thus, could
represent a potential marker of HCC prognosis (106).
In addition, ciRS-7 has also been indicated to be
significantly upregulated in HCC tissues (107). HCC cell proliferation and invasion
were significantly reduced potentially via the release of miR-7
(107,108). Thus, ciRS-7 may also be a miR-7
sponge, affecting the expression of miR-7-targeted genes through an
indirect regulatory pathway.
CircRNAs and bladder cancer
Bladder cancer is one of the most commonly
diagnosed malignancy of the genitourinary system and is also one of
the most common causes of cancer-associated mortality in men
(109). Recently, Zhong et
al (110) reported the altered
expression of six circRNAs via high throughput microarrays. Using
this method, the authors revealed that circ-transcription factor 25
(TCF25), circ-protein tyrosine kinase 2, circ-zinc finger RNA
binding protein, and circBC048201 were significantly upregulated,
and the two circRNA circ-family with sequence similarity 169 member
A and circ-tripartite motif-containing 24 were both downregulated
in bladder cancer tissues when compared with paired non-cancerous
tissues. Further study revealed that circTCF25 could bind with the
miRs, miR-103a-3p and miR-107, within the PI3K-Akt signaling
pathway and affect cancer progression (110). Abnormal overexpression of
circTCF25 was revealed to facilitate the proliferation and
migration of bladder cancer cell lines by inhibiting miR-103a-3p
and miR-107 (110). Analysis of
the miRNA sponge, circTCF25, revealed that the elevated protein
levels of CDK6 negatively regulated oncogenic factors (110). These results demonstrated that the
circTCF25-miR-103a-3p/miR-107-CDK6 network has an important role in
the development of bladder cancer (110).
CircRNAs and acute promyelocytic
leukemia
Guarnerio et al (111) demonstrated that cancer-associated
chromosomal translocations result in the fusion of circRNAs
(f-circRNAs), which are produced by the transcriptional exons of
different genes that are affected post-translation. Subsequently,
the group analyzed the acute promyelocytic leukemia (APL) cell line
NB4 and evaluated the expression of f-circRNA (either
f-circ-progesterone receptor or f-circM9). The results supported
the concept that f-circRNA, coupled with other tumor irritants, may
support the progression of leukemia in humans (111). Further research also demonstrated
that circRNAs are widespread in human thrombocytes and are
increased by 17- to 188-fold when compared with that in nucleated
tissues. As circRNAs are degraded by exonucleases, their
localization relative to that of linear RNAs can be used as a
marker of mRNA abundance in the absence of transcription (112). A previous study revealed that
neoplasm-cultured platelet levels could differ in cancer patients
with 96% accuracy when compared with healthy groups, and the
localization of the primary tumor could be accurately identified
with 71% accuracy from six cancer types by mRNA sequencing
(113). These results demonstrated
that f-circRNAs and changes in the level of blood platelets may
become potential markers for the diagnosis and treatment of
tumors.
CircRNAs and osteosarcoma
The incidence of osteosarcoma remains high in
adolescents and children, and the overall survival rate is remains
low. Identifying novel early detection markers and therapeutic
targets are required for the treatment of osteosarcoma. Liu et
al revealed that circ-5′-nucleotidase cytosolic II (NT5C2) was
abnormally expressed in osteosarcoma as determined by
high-throughput analysis; it promoted tumor cell progression and
metabolism by regulating miR-448 expression (114). In addition, a previous study
revealed that downregulation of circ-PVT1 decreased the expression
of the classical multidrug resistance-associated gene, ATP binding
cassette subfamily B member 1, suggesting that circ-PVT1 may be
more efficient than alkaline phosphatase for the diagnosis of
osteosarcoma (115). Taken
together, the results of these studies indicate that circ-NT5C2 and
circ-PVT1 may become early diagnostic markers and treatment
targets.
CircRNAs and other types of
cancers
Numerous circRNAs have been implicated in a wide
range of cancers, including gastric, lung, colorectal, esophageal,
breast, hepatic, vesical cancers, and in APL as discussed above
(Table III). However, it is clear
that circRNAs are not limited to these types of cancers. Due to the
rapid expansion of the research field, researchers are frequently
reporting the important roles of circRNAs in different types of
cancers.
A previous study identified circRNA expression
patterns in pancreatic ductal adenocarcinoma (PDAC), using an
aberrant microarray chip. The results indicated that circRNAs are
involved in the growth and metastasis of PDAC (116). Sand et al (117) revealed that 322 circRNAs are
expressed in cutaneous squamous cell carcinoma (cSCC) and a total
of 1,603 MREs were determined in the differentially expressed
circRNAs. The results demonstrated that circRNAs are expressed in
cSCC and participate in the formation of cancer through the
involvement of complementary MREs, which interfere with miRNA
sequences in the epigenetic control of cSCC-associated miRNAs
(117).
Using Arraystar Human circRNA Microarray, Peng
et al (118) validated 88
upregulated circRNAs and 10 significantly downregulated circRNAs in
papillary thyroid cancers (PTC), compared to matched metastatic
normal thyroid samples. Using these dysregulated circRNAs and
predicted MREs, a well-connected network map of circRNA-miR
interactions was established using Cytoscape software. One of the
downregulated circRNAs, hsa_circRNA_100395, showed potential for
functioning with two miRNAs associated with cancers, miR-141-3p and
miR-200a-3p (118). This result
indicates that the hsa_circRNA_100395-miR-141-3p or
hsa_circRNA_100395-miR-200a-3p interaction may be involved in the
nosogenesis of PTC tumors (118).
However, this theory requires further analysis to be proven
correct.
Analysis of laryngeal squamous cell cancers (LSCCs)
revealed that hsa_circ_100855 and hsa_circ_104912, among the 698
identified aberrant circRNAs, were markedly upregulated and
downregulated, respectively (119). The upregulation of hsa_circ_100855
was related with the T3-4 stage, cervical lymph node metastasis and
the late clinical stage of LSCC, while the downregulation of
hsa_circ_104912 was correlated with late stage, cervical lymph node
metastasis and poor differentiation. The results of this previous
study indicated that hsa_circ_100855 and hsa_circ_104912 may be
potential neoteric biomarkers for the diagnosis and progression of
LSCC (119).
Conclusions
Taken together, these results indicate that
circRNAs as mediators may be involved in the occurrence and
development of cancer. Particular circRNAs, such as circ-ITCH, may
even play crucial roles in more than one type of cancer.
Nevertheless, the majority of the previous studies mentioned above
lack clear and convincing arguments to explain the molecular
mechanisms of action, and thus, it is essential to obtain a deeper
understanding of the implications of their interactions in cancer
on a molecular level. Therefore, the clinical significance of
circRNAs as novel clinical diagnostic, metastatic and prognostic
biomarkers requires further study.
CircRNAs are a class of abundant ncRNAs that have
been demonstrated to regulate gene expression through a variety of
complicated mechanisms, and may also have the potential to encode
proteins; however, the underlying mechanisms have not been
completely elucidated. Nevertheless, circRNAs are becoming
potentially important regulators of cellular cytopathophysiology
and potential biomarkers for disease onset or progression. The
current levels of knowledge on circRNAs are relatively new and
limited; thus, further research is urgently needed to fully
understand their functions and clinical potential.
Perspectives
Recently, research into circRNAs has focused on
their biogenesis and functions as miRNA sponges and biomarkers.
Although there have been several studies on circRNAs, their
bioinformatic functions and related molecular mechanisms require
further study. In fact, most circRNA functions are still largely
unexplored, such as their specific expression during tumor cell
development, localization and metabolism. CircRNAs have good
potential as clinical diagnostic and prognostic biomarkers when
compared with miRNAs and lncRNAs, due to their stable ring
structure. Based on their location, there are three subclasses of
circRNAs: circRNAs are located in the cytoplasm and ciRNAs and
EIciRNAs are both nuclear (7,13,27),
implying that circRNAs may play epigenetic roles in cell
biology.
To date, there is no uniform nomenclature for
circRNAs, which has caused confusion regarding the terminology.
Although there are two sets of nomenclature rules, they both have
their own advantages and disadvantages. One is named according to
the nomenclature in the circBASE database, such as
‘hsa_circRNA_seven numbers’ (64),
while the other is according to their biofunctions, such as ciRS-7
(13,16), or their host genes, such as
circ-ITCH (77,92) and circMTO1 (120). Despite this, both methods have
their own drawbacks. The former has a complex form that is not
easily understood. The latter may provide the information of the
gene that generates multiple circRNAs after transcription, and
thus, this method is not used to name each circRNA based on its own
characteristics.
CircRNAs were initially cognitively untranslatable,
but recent reports have argued that some circRNAs carry ORFs, and
that some of them contain IRES elements. As more evidence emerges,
there is a growing interest in these functions and roles of
circRNAs. It is predicted that this direction and trend in research
regarding ORFs will become the new key area of interest in the
field of circRNAs.
A recent study reported, for the first time, the
presence of extensive circRNAs in exosomes (121), which are small membranous vesicles
involved in endocytosis that are secreted by most types of cells
(122). The sorting and loading of
circRNAs into exosomes could be controlled by altering the
associated miRNA levels in cells; these circRNAs may transfer to
other recipient cells through biological activity. There is no
doubt that circRNAs have potential as tumor biomarkers of clinical
diagnostic and prognostic value. The above conclusions lay the
foundations for a novel type of exosome-based cancer biomarker of
the development of circRNAs and reveal the potential biological
function of exosomal circRNAs. Research into circRNAs broadens our
understanding of the eukaryotic genome and provides a new direction
to investigate the comprehensive understanding of common diseases.
Further studies on circRNAs may reveal biocontrol-inducing factors,
the functions of different types of these molecules, whether
circRNAs can interact directly with the genome (as with lncRNAs),
and the kinetic mechanisms of circRNA degradation.
With the development of more advanced scientific
technology and more in-depth study of circRNAs, future research may
elucidate the physiological and pathological processes that
circRNAs are involved in in the human body, the role of circRNAs in
the blood as biomarkers and may further evaluate the biological
functions of more unknown circRNAs. Authors report no competing
interest in this work.
Acknowledgements
The authors thank Mr. Jia Liu for his suggestions
on the structure and language editing of the whole article.
Funding
The Department of Thoracic Surgery, Affiliated
Hospital of Qingdao University supported this study.
Availability of data and materials
Not applicable.
Authors' contributions
YW, TL, JL and WJ conceived and designed the study.
YW wrote the paper. YW and QW jointly designed the figures. YW, TL,
QW, JL and WJ reviewed and edited the manuscript. All authors read
and approved the manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
No authors report any competing interest.
Glossary
Abbreviations
Abbreviations:
|
circRNAs
|
circular RNAs
|
|
miRNAs
|
microRNAs
|
|
lncRNAs
|
long non-coding RNAs
|
|
ncRNAs
|
non-coding RNAs
|
|
RNA-Seq
|
RNA sequencing technology
|
|
RBP
|
RNA-binding protein
|
|
pre-mRNA
|
precursor messenger RNA
|
|
hnRNA
|
heterogeneous nuclear RNA
|
|
ecircRNAs
|
exonic circRNAs
|
|
ciRNAs
|
circular intronic RNAs
|
|
EIciRNAs
|
exon-intron circRNA
|
|
MREs
|
miRNA response elements
|
|
ceRNAs
|
competitive endogenous RNAs
|
|
ORFs
|
open reading frames
|
|
EV
|
extracellular vesicle
|
|
NSCLC
|
non-small cell lung cancer
|
|
CRC
|
colorectal cancer
|
|
ESCC
|
esophageal squamous cell
carcinoma
|
|
DCIS
|
ductal carcinoma in situ
|
|
HCC
|
hepatocellular carcinoma
|
|
PDAC
|
pancreatic ductal adenocarcinoma
|
|
cSCC
|
cutaneous squamous cell carcinoma
|
|
LSCC
|
laryngeal squamous cell cancer
|
|
APL
|
acute promyelocytic leukemia
|
|
PTC
|
papillary thyroid cancer
|
|
IHD
|
ischaemic heart disease
|
References
|
1
|
Amodio N, Raimondi L, Juli G, Stamato MA,
Caracciolo D, Tagliaferri P and Tassone P: MALAT1: A druggable long
non-coding RNA for targeted anti-cancer approaches. J Hematol
Oncol. 11:632018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Glasgow AMA, De Santi C and Greene CM:
Non-coding RNA in cystic fibrosis. Biochem Soc Trans. 46:619–630.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zong L, Sun Q, Zhang H, Chen Z, Deng Y, Li
D and Zhang L: Increased expression of circRNA_102231 in lung
cancer and its clinical significance. Biomed Pharmacother.
102:639–644. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Soslau G: Circular RNA (circRNA) was an
important bridge in the switch from the RNA world to the DNA world.
J Theor Biol. 447:32–40. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jiang XM, Li ZL, Li JL, Xu Y, Leng KM, Cui
YF and Sun DJ: A novel prognostic biomarker for cholangiocarcinoma:
circRNA Cdr1as. Eur Rev Med Pharmacol Sci. 22:365–371.
2018.PubMed/NCBI
|
|
6
|
Zhang HD, Jiang LH, Sun DW, Hou JC and Ji
ZL: CircRNA: A novel type of biomarker for cancer. Breast Cancer.
25:1–7. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jeck WR, Sorrentino JA, Wang K, Slevin MK,
Burd CE, Liu J, Marzluff WF and Sharpless NE: Circular RNAs are
abundant, conserved, and associated with ALU repeats. RNA.
19:141–157. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gao Y, Zhang J and Zhao F: Circular RNA
identification based on multiple seed matching. Brief Bioinform.
Feb 28–2017.(Epub ahead of print). doi: 10.1093/bib/bbx014.
|
|
9
|
Lei K, Bai H, Wei Z, Xie C, Wang J, Li J
and Chen Q: The mechanism and function of circular RNAs in human
diseases. Exp Cell Res. 368:147–158. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li D, Chen Y, Mei H, Jiao W, Song H, Ye L,
Fang E, Wang X, Yang F, Huang K, et al: Ets-1 promoter-associated
noncoding RNA regulates the NONO/ERG/Ets-1 axis to drive gastric
cancer progression. Oncogene. 37:4871–4886. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao ZJ and Shen J: Circular RNA
participates in the carcinogenesis and the malignant behavior of
cancer. RNA Biol. 14:514–521. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kulcheski FR, Christoff AP and Margis R:
Circular RNAs are miRNA sponges and can be used as a new class of
biomarker. J Biotechnol. 238:42–51. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Militello G, Weirick T, John D, Döring C,
Dimmeler S and Uchida S: Screening and validation of lncRNAs and
circRNAs as miRNA sponges. Brief Bioinform. 18:780–788.
2017.PubMed/NCBI
|
|
16
|
Salzman J, Chen RE, Olsen MN, Wang PL and
Brown PO: Cell-type specific features of circular RNA expression.
PLoS Genet. 9:e10037772013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim J, Abdelmohsen K, Yang X, De S,
Grammatikakis I, Noh JH and Gorospe M: LncRNA OIP5-AS1/cyrano
sponges RNA-binding protein HuR. Nucleic Acids Res. 44:2378–2392.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dai X, Zhang N, Cheng Y, Yang T, Chen Y,
Liu Z, Wang Z, Yang C and Jiang Y: RNA-binding protein
trinucleotide repeat-containing 6A regulates the formation of
circular RNA 0006916, with important functions in lung cancer
cells. Carcinogenesis. May 3–2018.(Epub ahead of print). doi:
10.1093/carcin/bgy061. View Article : Google Scholar
|
|
19
|
Conn SJ, Pillman KA, Toubia J, Conn VM,
Salmanidis M, Phillips CA, Roslan S, Schreiber AW, Gregory PA and
Goodall GJ: The RNA binding protein quaking regulates formation of
circRNAs. Cell. 160:1125–1134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu J, Jiang Z, Chen C, Hu Q, Fu Z, Chen J,
Wang Z, Wang Q, Li A, Marks JR, et al: CircIRAK3 sponges miR-3607
to facilitate breast cancer metastasis. Cancer Lett. 430:179–192.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yuan Y, Liu W, Zhang Y, Zhang Y and Sun S:
CircRNA circ_0026344 as a prognostic biomarker suppresses
colorectal cancer progression via microRNA-21 and microRNA-31.
Biochem Biophys Res Commun. 503:870–875. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhou J, Wang H, Chu J, Huang Q, Li G, Yan
Y, Xu T, Chen J and Wang Y: Circular RNA hsa_circ_0008344 regulates
glioblastoma cell proliferation, migration, invasion, and
apoptosis. J Clin Lab Anal. e224542018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou LH, Yang YC, Zhang RY, Wang P, Pang
MH and Liang LQ: CircRNA_0023642 promotes migration and invasion of
gastric cancer cells by regulating EMT. Eur Rev Med Pharmacol Sci.
22:2297–2303. 2018.PubMed/NCBI
|
|
24
|
Hou LD and Zhang J: Circular RNAs: An
emerging type of RNA in cancer. Int J Immunopathol Pharmacol.
30:1–6. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jakobi T and Dieterich C: Deep
computational circular RNA analytics from RNA-seq data. Methods Mol
Biol. 1724:9–25. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Chen BJ, Byrne FL, Takenaka K, Modesitt
SC, Olzomer EM, Mills JD, Farrell R, Hoehn KL and Janitz M:
Analysis of the circular RNA transcriptome in endometrial cancer.
Oncotarget. 9:5786–5796. 2018.PubMed/NCBI
|
|
27
|
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin
QF, Xing YH, Zhu S, Yang L and Chen LL: Circular intronic long
noncoding RNAs. Mol Cell. 51:792–806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guo JU, Agarwal V, Guo H and Bartel DP:
Expanded identification and characterization of mammalian circular
RNAs. Genome Biol. 15:4092014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Salzman J, Gawad C, Wang PL, Lacayo N and
Brown PO: Circular RNAs are the predominant transcript isoform from
hundreds of human genes in diverse cell types. PLoS One.
7:e307332012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Fumagalli MR, Zapperi S and La Porta CAM:
Impact of the cross-talk between circular and messenger RNAs on
cell regulation. J Theor Biol. 454:386–395. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Siede D, Rapti K, Gorska AA, Katus HA,
Altmüller J, Boeckel JN, Meder B, Maack C, Völkers M, Müller OJ, et
al: Identification of circular RNAs with host gene-independent
expression in human model systems for cardiac differentiation and
disease. J Mol Cell Cardiol. 109:48–56. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang Y and Wang Z: Efficient backsplicing
produces translatable circular mRNAs. RNA. 21:172–179. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ashwal-Fluss R, Meyer M, Pamudurti NR,
Ivanov A, Bartok O, Hanan M, Evantal N, Memczak S, Rajewsky N,
Kadener S, et al: circRNA biogenesis competes with pre-mRNA
splicing. Mol Cell. 56:55–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wilusz JE: Circular RNAs: Unexpected
outputs of many protein-coding genes. RNA Biol. 14:1007–1017. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hsiao KY, Sun HS and Tsai SJ: Circular
RNA-new member of noncoding RNA with novel functions. Exp Biol Med.
242:1136–1141. 2017. View Article : Google Scholar
|
|
36
|
Holdt LM, Kohlmaier A and Teupser D:
Molecular roles and function of circular RNAs in eukaryotic cells.
Cell Mol Life Sci. 75:1071–1098. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Petkovic S and Müller S: Synthesis and
engineering of circular RNAs. Methods Mol Biol. 1724:167–180. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li Z, Huang C, Bao C, Chen L, Lin M, Wang
X, Zhong G, Yu B, Hu W, Dai L, et al: Exon-intron circular RNAs
regulate transcription in the nucleus. Nat Struct Mol Biol.
22:256–264. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Chen L, Huang C, Wang X and Shan G:
Circular RNAs in eukaryotic cells. Curr Genomics. 16:312–318. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang Y and Wang Z: Constructing GFP-based
reporter to study back splicing and translation of circular RNA.
Methods Mol Biol. 1724:107–118. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhang XO, Dong R, Zhang Y, Zhang JL, Luo
Z, Zhang J, Chen LL and Yang L: Diverse alternative back-splicing
and alternative splicing landscape of circular RNAs. Genome Res.
26:1277–1287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang PL, Bao Y, Yee MC, Barrett SP, Hogan
GJ, Olsen MN, Dinneny JR, Brown PO and Salzman J: Circular RNA is
expressed across the eukaryotic tree of life. PLoS One.
9:e908592014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Qu S, Yang X, Li X, Wang J, Gao Y, Shang
R, Sun W, Dou K and Li H: Circular RNA: A new star of noncoding
RNAs. Cancer Lett. 365:141–148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Rybak-Wolf A, Stottmeister C, Glažar P,
Jens M, Pino N, Giusti S, Hanan M, Behm M, Bartok O, Ashwal-Fluss
R, et al: Circular RNAs in the mammalian brain are highly abundant,
conserved, and dynamically expressed. Mol Cell. 58:870–885. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Schmitz KM, Mayer C, Postepska A and
Grummt I: Interaction of noncoding RNA with the rDNA promoter
mediates recruitment of DNMT3b and silencing of rRNA genes. Genes
Dev. 24:2264–2269. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Beckedorff FC, Ayupe AC, Crocci-Souza R,
Amaral MS, Nakaya HI, Soltys DT, Menck CF, Reis EM and
Verjovski-Almeida S: The intronic long noncoding RNA ANRASSF1
recruits PRC2 to the RASSF1A promoter, reducing the expression of
RASSF1A and increasing cell proliferation. PLoS Genet.
9:e10037052013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Müller S and Appel B: In vitro
circularization of RNA. RNA Biol. 14:1018–1027. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhao F, Han Y, Liu Z, Zhao Z, Li Z and Jia
K: circFADS2 regulates lung cancer cells proliferation and invasion
via acting as a sponge of miR-498. Biosci Rep. 38:pii:
BSR201805702018. View Article : Google Scholar
|
|
49
|
Lin X and Chen Y: Identification of
potentially functional CircRNA-miRNA-mRNA regulatory network in
hepatocellular carcinoma by integrated microarray analysis. Med Sci
Monit Basic Res. 24:70–78. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang M, Jia L and Zheng Y: circRNA
expression profiles in human bone marrow stem cells undergoing
osteoblast differentiation. Stem Cell Rev. Jul 25–2018.(Epub ahead
of print). doi: 10.1007/s12015-018-9841-x. View Article : Google Scholar
|
|
51
|
Li S, Sun X, Miao S, Lu T, Wang Y, Liu J
and Jiao W: hsa_circ_0000729, a potential prognostic biomarker in
lung adenocarcinoma. Thorac Cancer. 9:924–930. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Haque S and Harries LW: Circular RNAs
(circRNAs) in health and disease. Genes. 8:E3532017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Schneider T, Hung LH, Schreiner S, Starke
S, Eckhof H, Rossbach O, Reich S, Medenbach J and Bindereif A:
CircRNA-protein complexes: IMP3 protein component defines subfamily
of circRNPs. Sci Rep. 6:313132016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ghosal S, Das S, Sen R, Basak P and
Chakrabarti J: Circ2Traits: A comprehensive database for circular
RNA potentially associated with disease and traits. Front Genet.
4:2832013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Gehring NH, Kunz JB, Neu-Yilik G, Breit S,
Viegas MH, Hentze MW and Kulozik AE: Exon-junction complex
components specify distinct routes of nonsense-mediated mRNA decay
with differential cofactor requirements. Mol Cell. 20:65–75. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Dudekula DB, Panda AC, Grammatikakis I, De
S, Abdelmohsen K and Gorospe M: CircInteractome: A web tool for
exploring circular RNAs and their interacting proteins and
microRNAs. RNA Biol. 13:34–42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Perriman R and Ares M Jr: Circular mRNA
can direct translation of extremely long repeating-sequence
proteins in vivo. RNA. 4:1047–1054. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
AbouHaidar MG, Venkataraman S, Golshani A,
Liu B and Ahmad T: Novel coding, translation, and gene expression
of a replicating covalently closed circular RNA of 220 nt. Proc
Natl Acad Sci USA. 111:14542–14547. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chen CY and Sarnow P: Initiation of
protein synthesis by the eukaryotic translational apparatus on
circular RNAs. Science. 268:415–417. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Chen X, Han P, Zhou T, Guo X, Song X and
Li Y: circRNADb: A comprehensive database for human circular RNAs
with protein-coding annotations. Sci Rep. 6:349852016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Jeck WR and Sharpless NE: Detecting and
characterizing circular RNAs. Nat Biotechnol. 32:453–461. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yang Y, Fan X, Mao M, Song X, Wu P, Zhang
Y, Jin Y, Yang Y, Chen LL, Wang Y, et al: Extensive translation of
circular RNAs driven by N6-methyladenosine. Cell Res.
27:626–641. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Lasda E and Parker R: Circular RNAs
co-precipitate with extracellular vesicles: A possible mechanism
for circRNA clearance. PLoS One. 11:e01484072016. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Glažar P, Papavasileiou P and Rajewsky N:
circBase: A database for circular RNAs. RNA. 20:1666–1670. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Liu YC, Li JR, Sun CH, Andrews E, Chao RF,
Lin FM, Weng SL, Hsu SD, Huang CC, Cheng C, et al: CircNet: A
database of circular RNAs derived from transcriptome sequencing
data. Nucleic Acids Res. 44:D209–D215. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Burd CE, Jeck WR, Liu Y, Sanoff HK, Wang Z
and Sharpless NE: Expression of linear and novel circular forms of
an INK4/ARF-associated non-coding RNA correlates with
atherosclerosis risk. PLoS Genet. 6:e10012332010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
He J, Xie Q, Xu H, Li J and Li Y: Circular
RNAs and cancer. Cancer Lett. 396:138–144. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Wang F, Nazarali AJ and Ji S: Circular
RNAs as potential biomarkers for cancer diagnosis and therapy. Am J
Cancer Res. 6:1167–1176. 2016.PubMed/NCBI
|
|
69
|
Kent OA and Mendell JT: A small piece in
the cancer puzzle: microRNAs as tumor suppressors and oncogenes.
Oncogene. 25:6188–6196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Peng Y and Croce CM: The role of MicroRNAs
in human cancer. Signal Transduct Target Ther. 1:150042016.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen
B, Luo Y, Lyu D, Li Y, Shi G, et al: Circular RNA profiling reveals
an abundant circHIPK3 that regulates cell growth by sponging
multiple miRNAs. Nat Commun. 7:112152016. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Scotti MM and Swanson MS: RNA mis-splicing
in disease. Nat Rev Genet. 17:19–32. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Li P, Chen S, Chen H, Mo X, Li T, Shao Y,
Xiao B and Guo J: Using circular RNA as a novel type of biomarker
in the screening of gastric cancer. Clin Chim Acta. 444:132–136.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Chen J, Li Y, Zheng Q, Bao C, He J, Chen
B, Lyu D, Zheng B, Xu Y, Long Z, et al: Circular RNA profile
identifies circPVT1 as a proliferative factor and prognostic marker
in gastric cancer. Cancer Lett. 388:208–219. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Li P, Chen H, Chen S, Mo X, Li T, Xiao B,
Yu R and Guo J: Circular RNA 0000096 affects cell growth and
migration in gastric cancer. Br J Cancer. 116:626–633. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Huang G, Zhu H, Shi Y, Wu W, Cai H and
Chen X: cir-ITCH plays an inhibitory role in colorectal cancer by
regulating the Wnt/β-catenin pathway. PLoS One. 10:e01312252015.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Wan L, Zhang L, Fan K, Cheng ZX, Sun QC
and Wang JJ: Circular RNA-ITCH suppresses lung cancer proliferation
via inhibiting the Wnt/β-catenin pathway. Biomed Res Int.
2016:15794902016. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Yao JT, Zhao SH, Liu QP, Lv MQ, Zhou DX,
Liao ZJ and Nan KJ: Over-expression of CircRNA_100876 in non-small
cell lung cancer and its prognostic value. Pathol Res Pract.
213:453–456. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Bachmayr-Heyda A, Reiner AT, Auer K,
Sukhbaatar N, Aust S, Bachleitner-Hofmann T, Mesteri I, Grunt TW,
Zeillinger R and Pils D: Correlation of circular RNA abundance with
proliferation - exemplified with colorectal and ovarian cancer,
idiopathic lung fibrosis, and normal human tissues. Sci Rep.
5:80572015. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Xie H, Ren X, Xin S, Lan X, Lu G, Lin Y,
Yang S, Zeng Z, Liao W, Ding YQ, et al: Emerging roles of
circRNA_001569 targeting miR-145 in the proliferation and invasion
of colorectal cancer. Oncotarget. 7:26680–26691. 2016.PubMed/NCBI
|
|
81
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Jiang Y, Yim SH, Xu HD, Jung SH, Yang SY,
Hu HJ, Jung CK and Chung YJ: A potential oncogenic role of the
commonly observed E2F5 overexpression in hepatocellular carcinoma.
World J Gastroenterol. 17:470–477. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Annunziata CM, Kleinberg L, Davidson B,
Berner A, Gius D, Tchabo N, Steinberg SM and Kohn EC: BAG-4/SODD
and associated antiapoptotic proteins are linked to aggressiveness
of epithelial ovarian cancer. Clin Cancer Res. 13:6585–6592. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Ozawa F, Friess H, Zimmermann A, Kleeff J
and Büchler MW: Enhanced expression of Silencer of death domains
(SODD/BAG-4) in pancreatic cancer. Biochem Biophys Res Commun.
271:409–413. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Li Y, Zhu X, Zeng Y, Wang J, Zhang X, Ding
YQ and Liang L: FMNL2 enhances invasion of colorectal carcinoma by
inducing epithelial-mesenchymal transition. Mol Cancer Res.
8:1579–1590. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Liang L, Li X, Zhang X, Lv Z, He G, Zhao
W, Ren X, Li Y, Bian X, Liao W, et al: MicroRNA-137, an HMGA1
target, suppresses colorectal cancer cell invasion and metastasis
in mice by directly targeting FMNL2. Gastroenterology.
144:624–635.e4. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Zhu M, Xu Y, Chen Y and Yan F: Circular
BANP, an upregulated circular RNA that modulates cell proliferation
in colorectal cancer. Biomed Pharmacother. 88:138–144. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Guo JN, Li J, Zhu CL, Feng WT, Shao JX,
Wan L, Huang MD and He JD: Comprehensive profile of differentially
expressed circular RNAs reveals that hsa_circ_0000069 is
upregulated and promotes cell proliferation, migration, and
invasion in colorectal cancer. Onco Targets Ther. 9:7451–7458.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Wang X, Zhang Y, Huang L, Zhang J, Pan F,
Li B, Yan Y, Jia B, Liu H, Li S and Zheng W: Decreased expression
of hsa_circ_001988 in colorectal cancer and its clinical
significances. Int J Clin Exp Pathol. 8:16020–16025.
2015.PubMed/NCBI
|
|
90
|
Cortés-López M and Miura P: Emerging
functions of circular RNAs. Yale J Biol Med. 89:527–537.
2016.PubMed/NCBI
|
|
91
|
Hsiao KY, Lin YC, Gupta SK, Chang N, Yen
L, Sun HS and Tsai SJ: Noncoding effects of circular RNA CCDC66
promote colon cancer growth and metastasis. Cancer Res.
77:2339–2350. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Li F, Zhang L, Li W, Deng J, Zheng J, An
M, Lu J and Zhou Y: Circular RNA ITCH has inhibitory effect on ESCC
by suppressing the Wnt/β-catenin pathway. Oncotarget. 6:6001–6013.
2015.PubMed/NCBI
|
|
93
|
Su H, Lin F, Deng X, Shen L, Fang Y, Fei
Z, Zhao L, Zhang X, Pan H, Xie D, et al: Profiling and
bioinformatics analyses reveal differential circular RNA expression
in radioresistant esophageal cancer cells. J Transl Med.
14:2252016. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Xia W, Qiu M, Chen R, Wang S, Leng X, Wang
J, Xu Y, Hu J, Dong G, Xu PL and Yin R: Circular RNA
has_circ_0067934 is upregulated in esophageal squamous cell
carcinoma and promoted proliferation. Sci Rep. 6:355762016.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Pinder SE: Ductal carcinoma in situ
(DCIS): Pathological features, differential diagnosis, prognostic
factors and specimen evaluation. Mod Pathol. 23 Suppl 2:S8–S13.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Hernandez L, Wilkerson PM, Lambros MB,
Campion-Flora A, Rodrigues DN, Gauthier A, Cabral C, Pawar V,
Mackay A, A'Hern R, et al: Genomic and mutational profiling of
ductal carcinomas in situ and matched adjacent invasive breast
cancers reveals intra-tumour genetic heterogeneity and clonal
selection. J Pathol. 227:42–52. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Galasso M, Costantino G, Pasquali L,
Minotti L, Baldassari F, Corrà F, Agnoletto C and Volinia S:
Profiling of the predicted circular RNAs in ductal in situ and
invasive breast cancer: A pilot study. Int J Genomics.
2016:45038402016. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Gregory PA, Bert AG, Paterson EL, Barry
SC, Tsykin A, Farshid G, Vadas MA, Khew-Goodall Y and Goodall GJ:
The miR-200 family and miR-205 regulate epithelial to mesenchymal
transition by targeting ZEB1 and SIP1. Nat Cell Biol. 10:593–601.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Ye ZB, Ma G, Zhao YH, Xiao Y, Zhan Y, Jing
C, Gao K, Liu ZH and Yu SJ: miR-429 inhibits migration and invasion
of breast cancer cells in vitro. Int J Oncol. 46:531–538. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Yang W, Du WW, Li X, Yee AJ and Yang BB:
Foxo3 activity promoted by non-coding effects of circular RNA and
Foxo3 pseudogene in the inhibition of tumor growth and
angiogenesis. Oncogene. 35:3919–3931. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Du WW, Ling F, Yang W, Wu N, Awan FM, Yang
Z and Yang BB: Induction of tumor apoptosis through a circular RNA
enhancing Foxo3 activity. Cell Death Differ. 24:357–370. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Caiment F, Gaj S, Claessen S and Kleinjans
J: High-throughput data integration of RNA-miRNA-circRNA reveals
novel insights into mechanisms of benzo[a]pyrene-induced
carcinogenicity. Nucleic Acids Res. 43:2525–2534. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Shang X, Li G, Liu H, Li T, Liu J, Zhao Q
and Wang C: Comprehensive circular RNA profiling reveals that
hsa_circ_0005075, a new circular rna biomarker, is involved in
hepatocellular crcinoma development. Medicine. 95:e38112016.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Dong L, Deng J, Sun ZM, Pan AP, Xiang XJ,
Zhang L, Yu F, Chen J, Sun Z, Feng M and Xiong JP: Interference
with the β-catenin gene in gastric cancer induces changes to the
miRNA expression profile. Tumour Biol. 36:6973–6983. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Warnecke-Eberz U, Chon SH, Hölscher AH,
Drebber U and Bollschweiler E: Exosomal onco-miRs from serum of
patients with adenocarcinoma of the esophagus: Comparison of miRNA
profiles of exosomes and matching tumor. Tumour Biol. 36:4643–4653.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Qin M, Liu G, Huo X, Tao X, Sun X, Ge Z,
Yang J, Fan J, Liu L and Qin W: Hsa_circ_0001649: A circular RNA
and potential novel biomarker for hepatocellular carcinoma. Cancer
Biomark. 16:161–169. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Yu L, Gong X, Sun L, Zhou Q, Lu B and Zhu
L: The circular RNA Cdr1as act as an oncogene in hepatocellular
carcinoma through targeting miR-7 expression. PLoS One.
11:e01583472016. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Xu L, Zhang M, Zheng X, Yi P, Lan C and Xu
M: The circular RNA ciRS-7 (Cdr1as) acts as a risk factor of
hepatic microvascular invasion in hepatocellular carcinoma. J
Cancer Res Clin Oncol. 143:17–27. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Zhong Z, Lv M and Chen J: Screening
differential circular RNA expression profiles reveals the
regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in
bladder carcinoma. Sci Rep. 6:309192016. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Guarnerio J, Bezzi M, Jeong JC, Paffenholz
SV, Berry K, Naldini MM, Lo-Coco F, Tay Y, Beck AH and Pandolfi PP:
Oncogenic role of fusion-circrnas derived from cancer-associated
chromosomal translocations. Cell. 165:289–302. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Alhasan AA, Izuogu OG, Al-Balool HH, Steyn
JS, Evans A, Colzani M, Ghevaert C, Mountford JC, Marenah L,
Elliott DJ, et al: Circular RNA enrichment in platelets is a
signature of transcriptome degradation. Blood. 127:e1–e11. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Best MG, Sol N, Kooi I, Tannous J,
Westerman BA, Rustenburg F, Schellen P, Verschueren H, Post E,
Koster J, et al: RNA-Seq of tumor-educated platelets enables
blood-based pan-cancer, multiclass, and molecular pathway cancer
diagnostics. Cancer Cell. 28:666–676. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Liu X, Zhong Y, Li J and Shan A: Circular
RNA circ-NT5C2 acts as an oncogene in osteosarcoma proliferation
and metastasis through targeting miR-448. Oncotarget.
8:114829–114838. 2017.PubMed/NCBI
|
|
115
|
Kun-Peng Z, Xiao-Long M and Chun-Lin Z:
Overexpressed circPVT1, a potential new circular RNA biomarker,
contributes to doxorubicin and cisplatin resistance of osteosarcoma
cells by regulating ABCB1. Int J Biol Sci. 14:321–330. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Qu S, Song W, Yang X, Wang J, Zhang R,
Zhang Z, Zhang H and Li H: Microarray expression profile of
circular RNAs in human pancreatic ductal adenocarcinoma. Genom
Data. 5:385–387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Sand M, Bechara FG, Gambichler T, Sand D,
Bromba M, Hahn SA, Stockfleth E and Hessam S: Circular RNA
expression in cutaneous squamous cell carcinoma. J Dermatol Sci.
83:210–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Peng N, Shi L, Zhang Q, Hu Y, Wang N and
Ye H: Microarray profiling of circular RNAs in human papillary
thyroid carcinoma. PLoS One. 12:e01702872017. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Xuan L, Qu L, Zhou H, Wang P, Yu H, Wu T,
Wang X, Li Q, Tian L, Liu M and Sun Y: Circular RNA: A novel
biomarker for progressive laryngeal cancer. Am J Transl Res.
8:932–939. 2016.PubMed/NCBI
|
|
120
|
Han D, Li J, Wang H, Su X, Hou J, Gu Y,
Qian C, Lin Y, Liu X, Huang M, et al: Circular RNA circMTO1 acts as
the sponge of microRNA-9 to suppress hepatocellular carcinoma
progression. Hepatology. 66:1151–1164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J,
Chen D, Gu J, He X and Huang S: Circular RNA is enriched and stable
in exosomes: A promising biomarker for cancer diagnosis. Cell Res.
25:981–984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Melo SA, Luecke LB, Kahlert C, Fernandez
AF, Gammon ST, Kaye J, LeBleu VS, Mittendorf EA, Weitz J, Rahbari
N, et al: Glypican-1 identifies cancer exosomes and detects early
pancreatic cancer. Nature. 523:177–182. 2015. View Article : Google Scholar : PubMed/NCBI
|