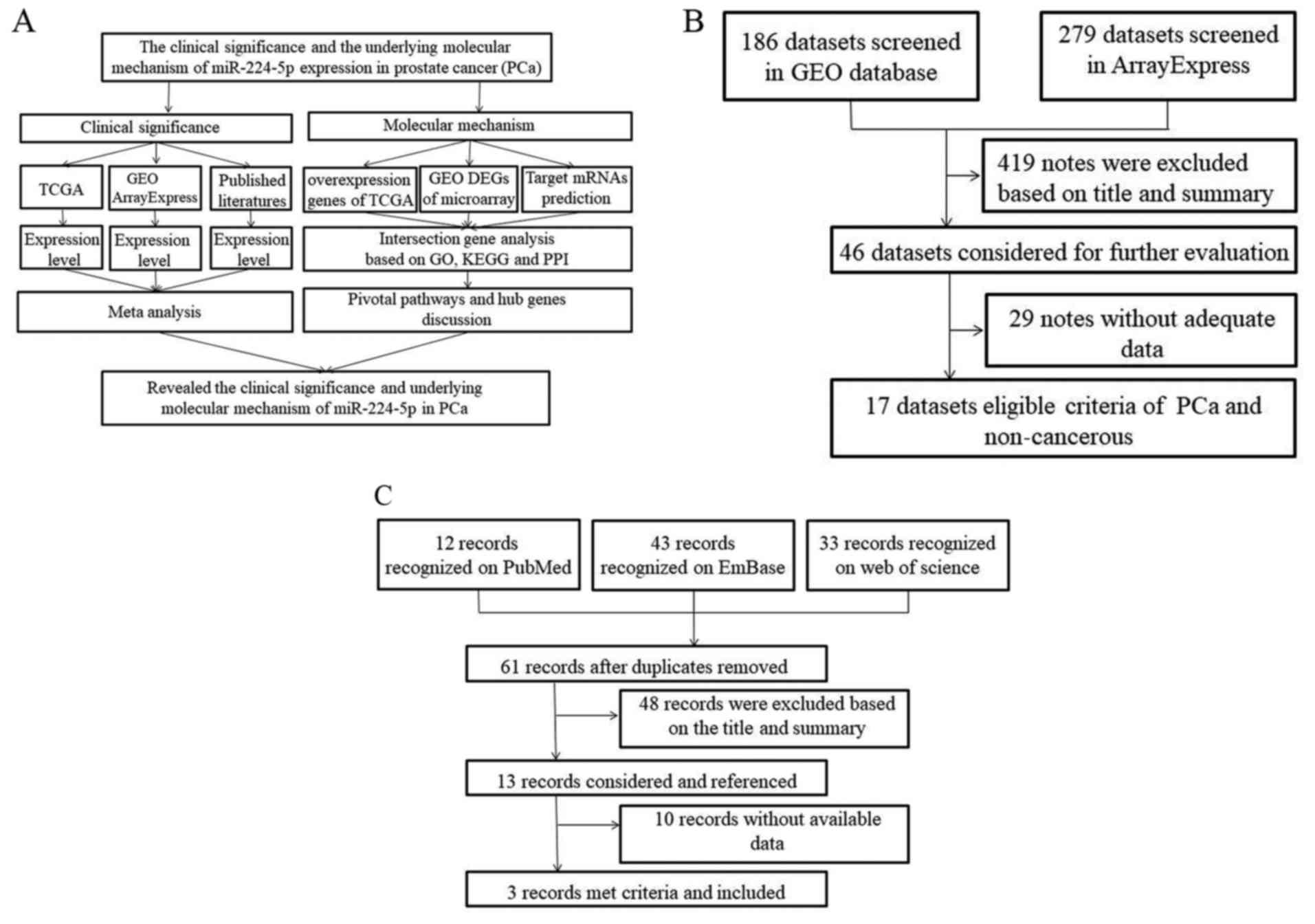
Introduction
According to cancer statistics from 2017, prostate
adenocarcinoma (PCa) was ranked highest for newly diagnosed cases
of cancer in men in the USA, and was considered the 3rd leading
cause of cancer-associated mortality in men (1). Although the rates of incidence and
mortality vary in different nations and regions, PCa poses a health
risk to men due to its high incidence and mortality rates (2–4). PCa
is a heterogeneous type of cancer; patients with PCa are prone to a
relapse and metastasis, and the prognosis of PCa is associated with
the age of patients (5,6). The majority of patients are diagnosed
with PCa in its intermediate or terminal stage, which presents
challenges to treatment and recovery of patients. Fortunately, the
diagnosis and treatment of PCa have been modified owing to medical
advances. For example, screening prostate-specific antigen in the
early stage is considered an effective approach for early diagnosis
and immediate treatment of PCa (7).
Despite this, the incidence and mortality rates of PCa remain high,
therefore, further investigations are urgently required to clarify
the onset and mechanism of progression of PCa (8).
Based on previous studies, in addition to obesity,
gene expression is closely associated with the onset and
development of PCa (9,10). MicroRNAs (miRNAs) are small,
non-coding RNA molecules of ~ 22 nucleotides, which perform
functions in the post-transcriptional regulation of gene expression
(11–15). miRNAs act as oncogenes or
antioncogenes in tumorigenesis, thus influencing the onset and
development of tumors (16–20). Previous studies have shown that the
downregulated expression of miRNA may be essential in the onset and
development of PCa (21–23). In addition, increased expressed of
miRNA was likely to regulate the expression of target genes and
exert its influences on the progression of PCa (24,25).
Currently, investigations on the role of
miRNA-224-5p (miR-224-5p) in PCa have been limited. Mavridis et
al performed a case-control study involving 73 cases of PCa and
66 cases of benign prostatic hyperplasia, demonstrating that the
expression of miR-224-5p was downregulated in PCa tissues and, with
disease progression, the expression of miR-224-5p was decreased
further; patients with the lower expression of miR-224-5p were also
more likely to have a poorer prognosis (26). Fu et al investigated the
targeting association between miR-224-5p and
calcium/calmodulin-dependent protein (CAMKK2) in tissues from 20
cases of PCa and non-cancerous counterpart tissues; it was found
that the expression of miR-224-5p was markedly lower in PCa
tissues, and patients with a lower expression of miR-224-5p tended
to have a poorer survival rate; additionally, miR-224-5p inhibited
the proliferation of tumor cells by targeting CAMKK2 (27). Having applied reverse
transcription-quantitative polymerase chain reaction analysis to
examine tissues from 36 cases of PCa and 14 non-cancerous tissues,
Kristensen et al confirmed that the expression of miR-224-5p
was decreased in PCa tissues and predicted an unsatisfactory
outcome for patients (28). These
previous studies offer important insights into the effects of
miR-224-5p on PCa tissues, however, they were performed with a
small sample size (n<80) and the results were not confirmed with
a larger sample size, which may reduce the convincingness of the
results. Furthermore, the studies mentioned above failed to perform
bioinformatics analyses, which may assist in identifying more
prospective target genes of miR-224-5p in PCa. Therefore, further
verification is required of the expression of miR-224-5p and its
clinical significance in PCa. In addition, the mining of multiple
databases and bioinformatics analyses is required to examine the
prospective molecular mechanism underlying the role of miR-224-5p
in PCa.
The present study aimed to verify the expression of
miR-224-5p in PCa using data from different databases, including
The Cancer Genome Atlas (TCGA), Gene Expression Omnibus (GEO),
ArrayExpress, and previous literature. In addition, prospective
target genes of miR-224-5p were collected using online prediction
tools and differentially expressed genes in TCGA and GEO.
Bioinformatics analyses were also used to further examine the
signaling pathways of miR-224-5p in PCa (Fig. 1A).
Materials and methods
Collection of PCa data from TCGA
Data of Illumina HiSeq level 3 were acquired from
the Launch ata portal of TCGA (https://cancergenome.nih.gov/). Following summarizing
of the raw count data, raw data on miRNA expression in tissues of
500 cases of PCa and 52 normal prostatic tissues were obtained,
from which reads per million data of pre-miR224 were extracted,
including tissues from 498 cases of PCa and 51 normal PCa tissues.
In these 500 cases, the age of the patients ranged between 41 and
78 years, with a mean age of 61 years. Additional
clinicopathological parameters are listed in Table I.
 | Table I.Association between the expression of
miR-224-5p between prostate cancer tissue and non-cancerous tissue
based on The Cancer Genome Atlas data. |
Table I.
Association between the expression of
miR-224-5p between prostate cancer tissue and non-cancerous tissue
based on The Cancer Genome Atlas data.
|
|
| miR-224
expression | T-test |
|---|
|
|
|
|
|
|---|
| Clinicopathological
parameter | Cases | Mean | SD | FC | T-value | P-value |
|---|
| Group |
| Normal
adjacent | 51 | 5.4773 | 0.86449 | 1.0000 | −3.413 | 0.001 |
|
Cancer | 498 | 5.0255 | 1.19453 | 0.9175 |
|
|
| Age (years) |
|
<60 | 204 | 4.9880 | 1.36730 | 1.0000 | −0.086 | 0.931 |
|
≥60 | 296 | 4.9982 | 1.26126 | 1.0020 |
|
|
| Pathological T
stage |
|
T1-T2 | 188 | 5.1306 | 1.21866 | 1.0000 | 1.880 | 0.061 |
|
T3-T4 | 305 | 4.9052 | 1.33624 | 0.9561 |
|
|
| N stage |
| N0 | 348 | 5.0312 | 1.28350 | 1.0000 | 1.347 | 0.179 |
| N1 | 79 | 4.8182 | 1.19763 | 0.9577 |
|
|
| M stage |
| M0 | 456 | 4.9999 | 1.25252 | 1.0000 | 1.715 | 0.087 |
| M1 | 4 | 3.9179 | 1.76728 | 0.7836 |
|
|
| Gleason score |
| ≤7 | 291 | 5.1057 | 1.13197 | 1.0000 | 1.143 | 0.254 |
| 8≥ | 203 | 4.9815 | 1.26629 | 0.9757 |
|
|
| Gleason grade |
| 2 | 1 | 4.8074 |
|
| 1.048 | 0.371 |
| 3 | 197 | 5.1616 | 1.12858 |
|
|
|
| 4 | 248 | 5.0052 | 1.23926 |
|
|
|
| 5 | 48 | 4.8763 | 1.16617 |
|
|
|
| Recurrence |
| No | 373 | 5.0491 | 1.30796 | 1.0000 | 1.008 | 0.314 |
|
Yes | 58 | 4.8636 | 1.27528 | 0.9633 |
|
|
| Clinical T
stage |
|
T1-T2 | 351 | 5.0312 | 1.31167 | 1.0000 | −0.338 | 0.735 |
|
T3-T4 | 55 | 5.0945 | 1.14167 | 1.0126 |
|
|
Screening of differentially expressed
microarrays of miRNAs in PCa
In the microarray GEO (https://www.ncbi.nlm.nih.gov/gds/) and ArrayExpress
(https://www.ebi.ac.uk/arrayexpress/)
databases, a search was performed using the following key words:
(parastata OR prostatic gland OR prostate gland OR prostat* AND
(cancer OR carcinoma OR adenocarcinoma OR tumor OR malignan* OR
neoplas*) AND (miR OR miRNA OR microRNA OR miR OR miRNA OR microRNA
OR ‘miR’ OR ‘miRNA’ OR ‘microRNA’). Subsequently, the
differentially expressed microarrays of miRNAs in PCa were screened
and downloaded. Finally, studies were included which fulfilled the
following criteria: i) compared the cancerous tissues with the
controls; ii) contained microarrays of miRNA expression in PCa
tissues, biofluids and cell lines; and iii) had more than 3 samples
in each microarray. The procedures for the search are shown in
Fig. 1B.
Literature search
From the PubMed (https://www.ncbi.nlm.nih.gov/pubmed), Embase
(https://embase.com/) and Web of Science
(http://apps.webofknowledge.com/UA_General
Search_input.do?product=UA&search_mode=GeneralSearch&SID=8ErJvGamhqk4GR7zNPu&preferencesSaved=)
databases, studies on differentially expressed miR-224-5p in PCa
tissues and in non-cancerous controls were collected. The key words
used for the search included the following: (parastata OR prostatic
gland OR prostate gland OR prostat*) and (cancer OR carcinoma OR
adenocarcinoma OR tumor OR malignan* OR neoplas*) and (miR-224 OR
miRNA-224 OR microRNA-224 OR miR224 OR miRNA224 OR microRNA224 OR
‘miR 224’ OR ‘miRNA 224’ OR ‘microRNA 224’ OR miR-224-5p OR
miRNA-224-5p OR microRNA-224-5p). The studies were included if they
met the following standards: i) involved the comparison of PCa with
non-cancerous controls in tissues, biofluids and cell lines; and
ii) provided the mean ± standard deviation (SD) or diagrams from
which data extraction was possible. The procedures for the
literature screening are shown in Fig.
1C.
Collection of differentially expressed
genes of PCa in TCGA
Gene expression profiling interactive analysis
(GEPIA; http://gepia.cancer-pku.cn/), a
visualized website based on TCGA database developed by Peking
University (Beijing, China), contains various functional analyses,
including the comparison of differentially expressed genes in
cancerous and non-cancerous tissues (29). Data on the differential genes of PCa
on GEPIA were retrieved, and differentially expressed genes
calculated using the Linear Models for Microarray Data (LIMMA)
package (http://gepia.cancer-pku.cn/detail.php?gene=) were
downloaded. The genes were selected for further analysis if log2
fold change (FC)>1.
Selection of microarrays of
differentially expressed genes of PCa
In the GEO and ArrayExpress databases, a search was
performed using the aforementioned key words for prostate and
cancer. The relevant microarrays were included in the study if they
conformed to the following criteria: i) investigation of
differentially expressed mRNA based on post-transcriptional
miR-224-5p; and ii) comparison of PCa cell lines and normal cell
lines. If there existed numerous similar samples in one microarray,
the intersections were obtained. When dealing with different
microarrays, the unions were obtained. Further analyses were
performed on all results.
Prediction of potential target genes
of miR-224-5p
The microRNA-mRNA prediction was performed with the
miRWalk2.0 (http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/)
online prediction tools involving 12 prediction tools, namely,
DIANA microT v4, RNA22, Pictar2, miRWalk, miRNAMap, RNAhybrid,
mirBridge, TargetScan, miRMap, miRanda, PITA and miRDB. Genes that
were predicted by three tools qualified for the present study. In
order to acquire the potential target genes with accuracy, the
overexpressed genes in TCGA, mRNAs expressed at low levels
following miR-224-5p transcription and the predicted microarrays
were combined, and the unions were obtained. Bioinformatics
analysis was performed on these results.
In silico analysis
Gene Ontology (GO) and Kyoto Encyclopedia of Genes
and Genomes (KEGG) analyses were performed on the genes that
appeared in TCGA, GEO and target genes prediction tools on Database
for Annotation, Visualization and Integrated Discovery (DAVID;
http://david.abcc.ncifcrf.gov/). Bingo
on Cytoscape 3.5.0 (http://www.cytoscape.org/) was applied to construct
network analysis of GO terms, and the ClueGO and CluePedia plugins
were used to establish the KEGG network. For genes enriched in
significant pathways, the data in TCGA were used to verify their
expression levels in PCa. In addition, protein-protein interaction
(PPI) analysis was performed, a PPI network was constructed and the
interactive associations between proteins were confirmed on the
Search Tool for the Retrieval of Interacting Genes 0.5 (https://string-db.org/cgi/input.pl?sessionId=Ce1Dx9pYDluc&input_page_show_search=on)
(30–37) database. Based on TCGA data, the mRNA
expression of key genes in the PPI network were also confirmed, and
the mechanism of miR-224-5p in PCa was further examined.
Statistical analysis
SPSS 23.0 (IBM Corp., Armonk, NY, USA) was used for
statistical analysis of the expression of miR-224-5p in PCa. An
independent t-test was applied to evaluate the differentials of
miR-224-5p between PCa tissues and non-cancerous tissues, and the
results are presented as the mean ± SD. The standardized mean
difference (SMD) was used to combine all the included studies on
STATA 2.0 (StataCorp, College Station, TX, USA), and to calculate
the expression trend of miR-224-5p in PCa. In addition, a receiver
operating characteristic (ROC) was used to analyze the sensitivity
and specificity of each study, and their cut-off value was
calculated. Subsequently, the cut-off value was applied to identify
the true positive, false positive, false negative and true negative
of each study, following which a diagnostic test four-fold table
was produced. STATA 12.0 was then used to confirm the expression of
miR-224-5p in PCa, and the summary ROC (sROC) was used to measure
its credibility. In addition, in order to examine the expression
trend in each study, scatter diagrams were produced to show the
expression of miR-224-5p in PCa tissues and adjacent tissues via
GraphPad Prism 5.0 (GraphPad Software, Inc., La Jolla, CA, USA).
P<0.05 was considered to indicate statistically significant
difference.
Results
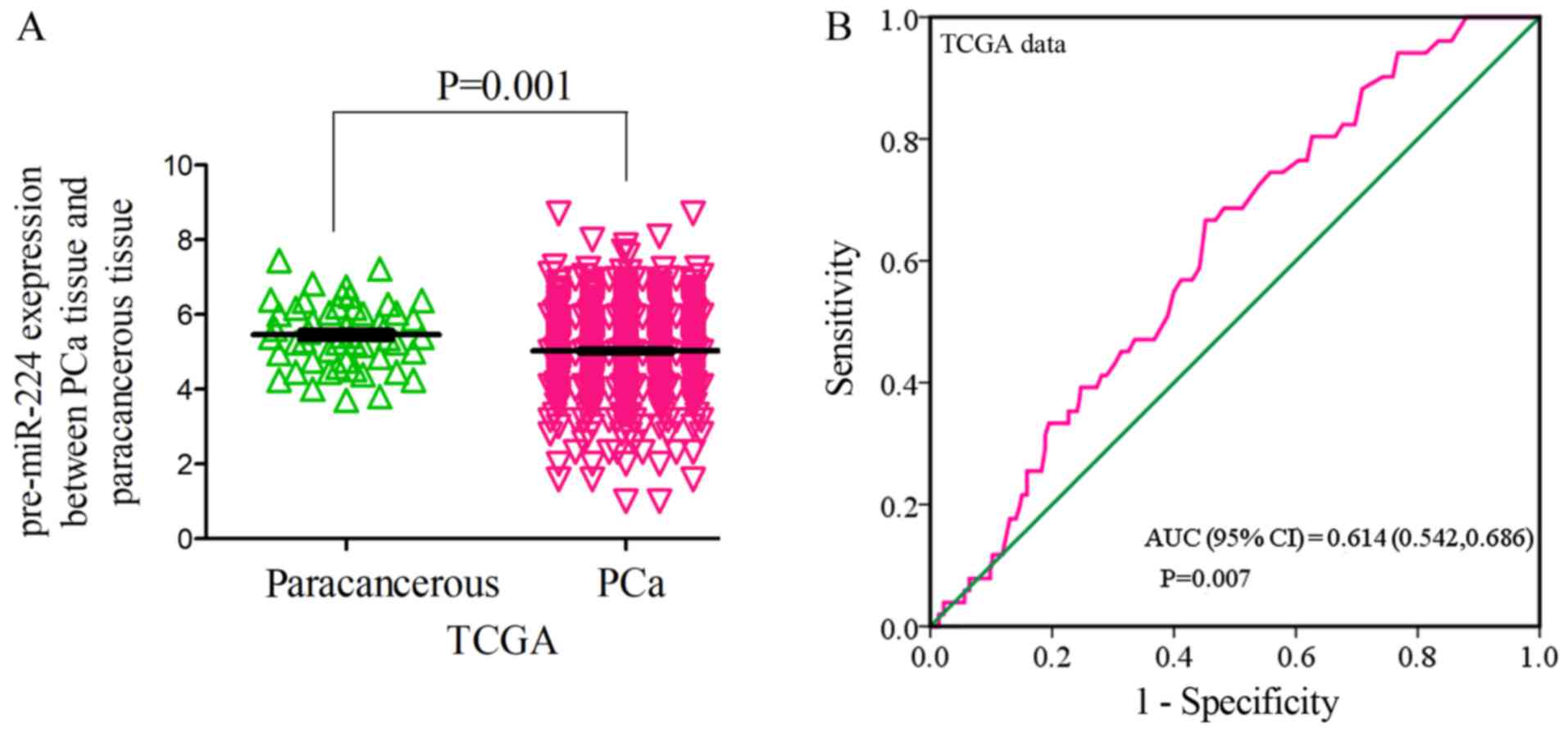
Expression of miR-224-5p in PCa
According to the data in TCGA, the expression of
pre-miR-224 in PCa tissues was 5.0255±1.1945 (PCa, vs. normal, 498,
vs. 52), which was significantly lower than that of normal adjacent
tissues (5.4773±0.8645, FC=0.9175, P=0.001; Fig. 2A). The area under the curve (AUC) of
the downregulated pre-miR-224 in PCa tissues was 0.614 (95% CI,
0.542, 0.686. P=0.007; Fig. 2B). It
was found that pre-miR-224 tended to exhibit lower expression with
the progression of clinical staging by comparing T3-4 and T1-2
(4.9052±1.3362 vs. 5.1306±1.2187, FC=0.9561, P=0.061), M1 and M0
(3.9179±1.7677 vs. 4.9999±1.2525, FC=0.7836, P=0.087; Table I). However, no clear associations
were found between its expression and prognosis or other types of
staging.
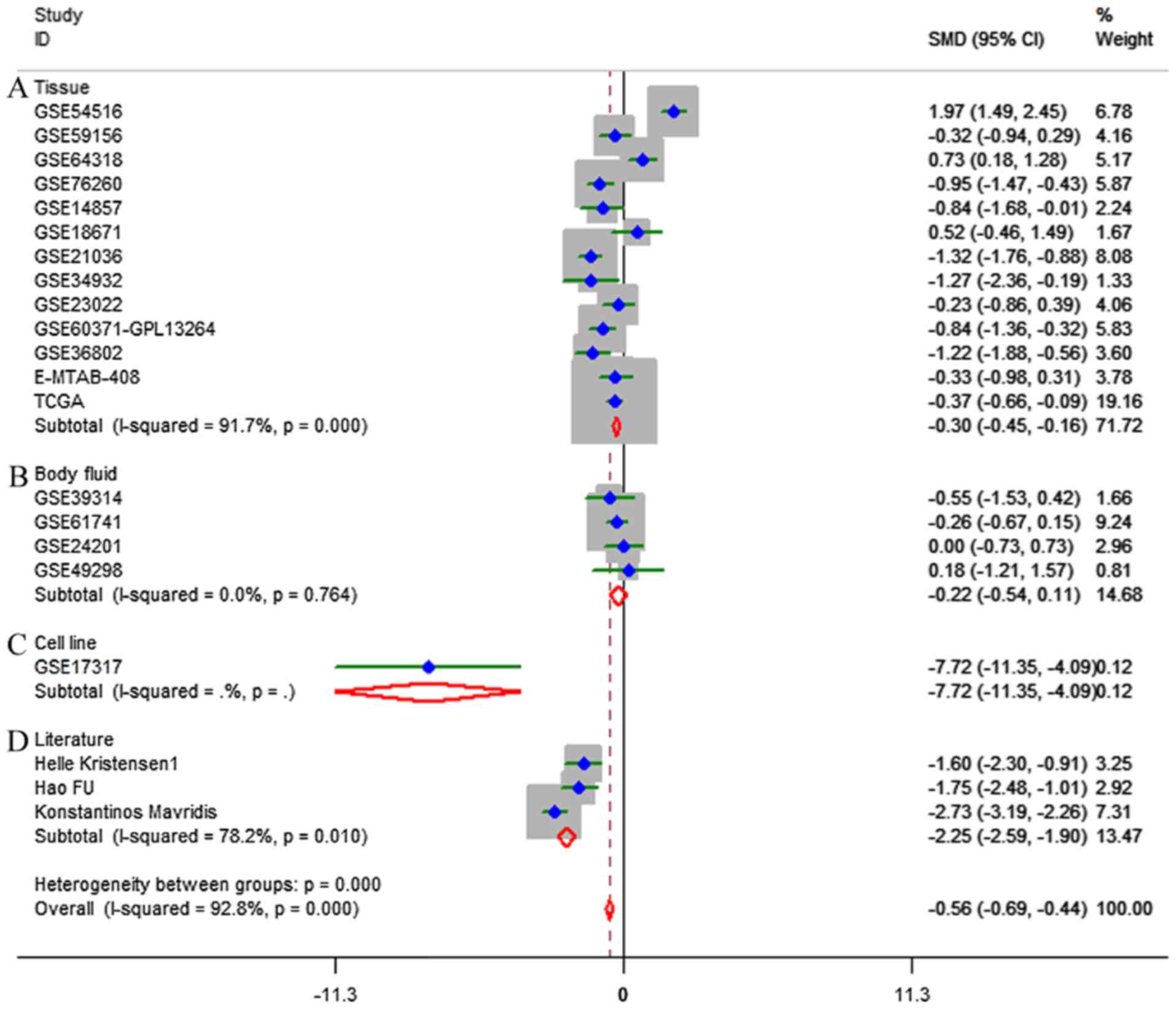
Based on the inclusion criteria, a total of 17
microarrays were eventually considered eligible for the present
study, which were categorized into three subtypes: Tissues,
biofluids and cell lines (Fig. 3).
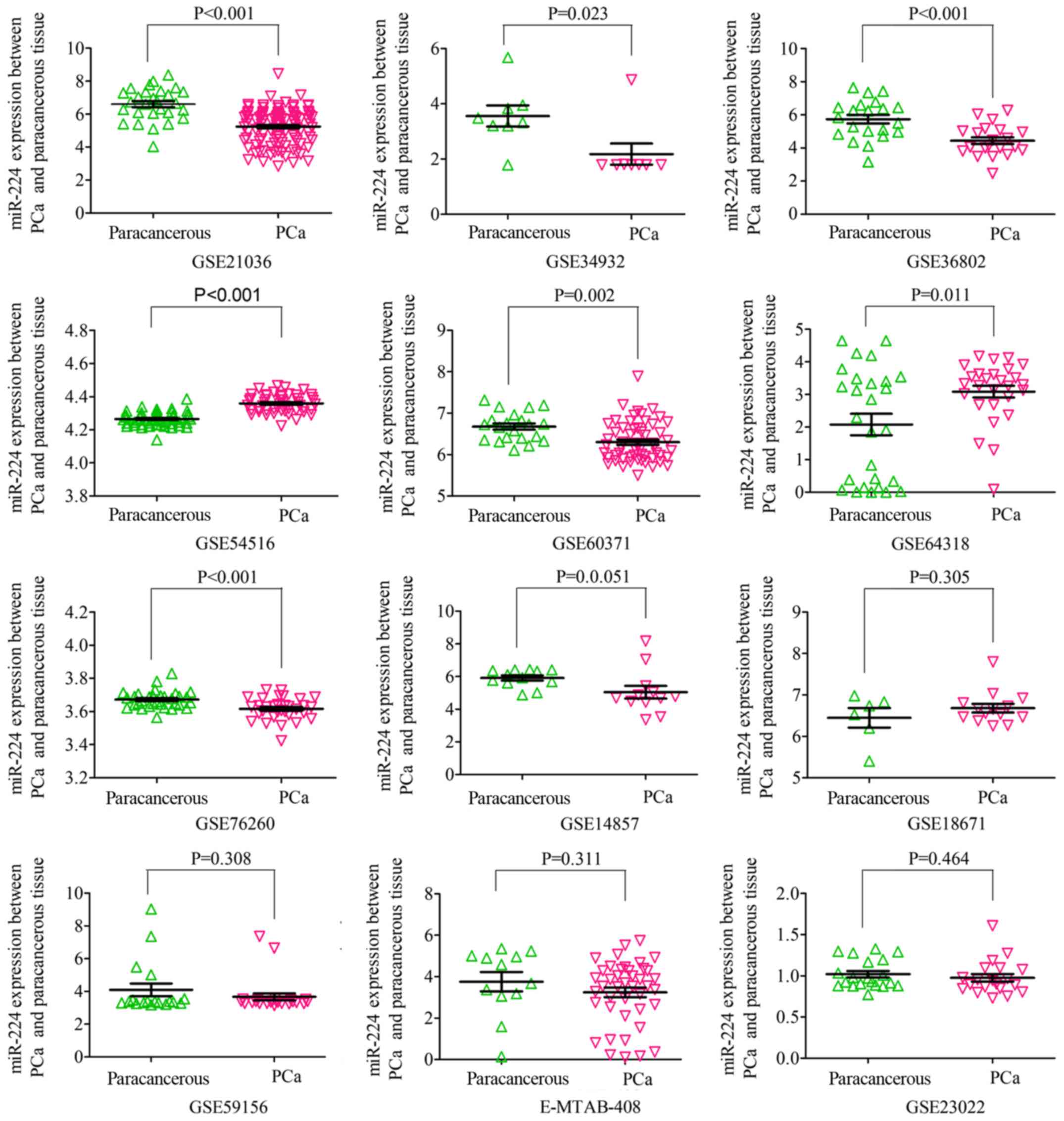
In the subgroup of tissues, 12 microarrays were included, among
which the expression of miR-224-5p was notably lower in PCa tissues
in GSE76260, GSE21036, GSE34932, GSE60371 and GSE36802. In GSE54516
and GSE64318, the expression of miR-224-5p was upregulated. In
terms of SMD, a low expression of miR-224-5p was identified in PCa
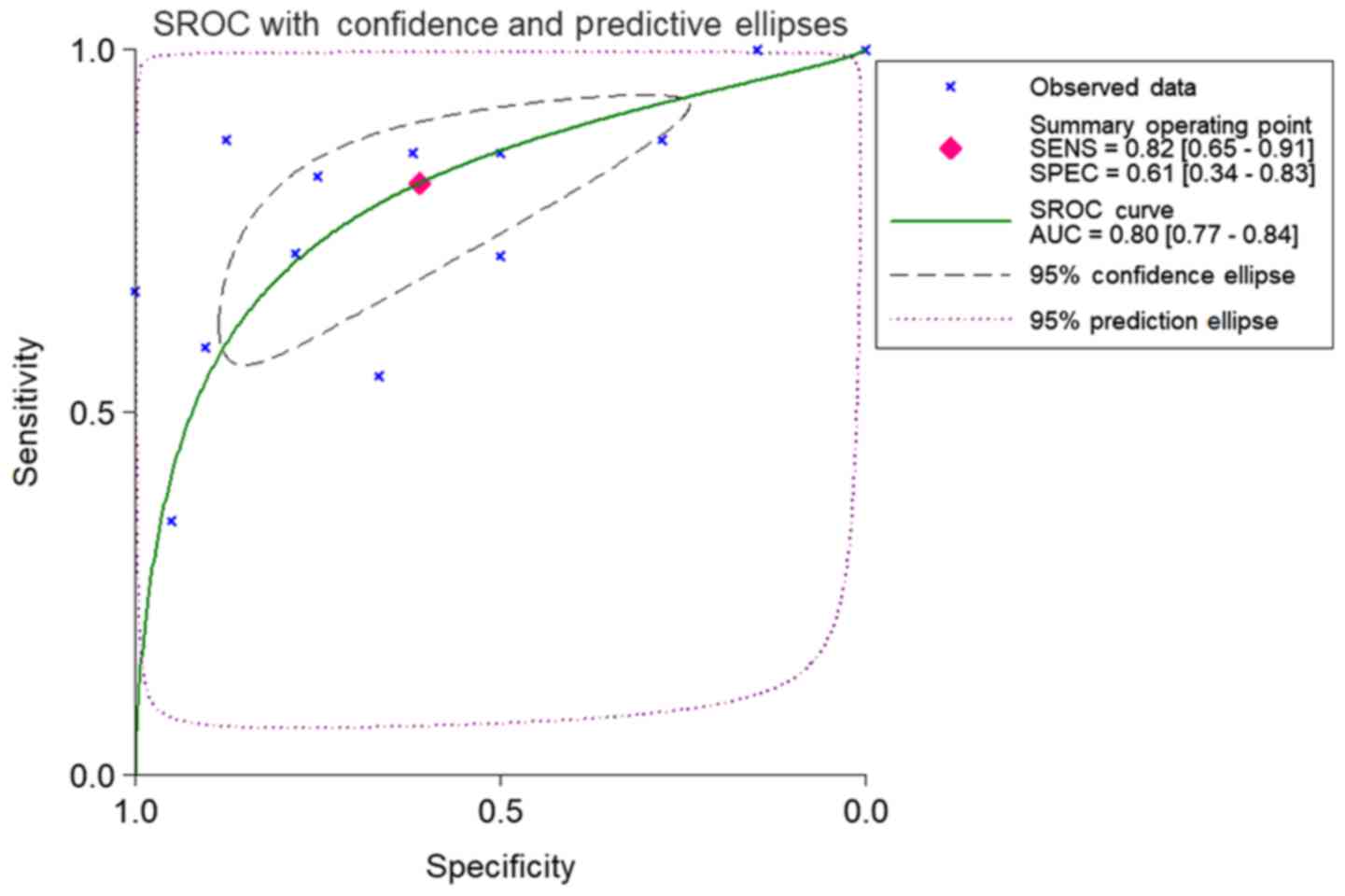
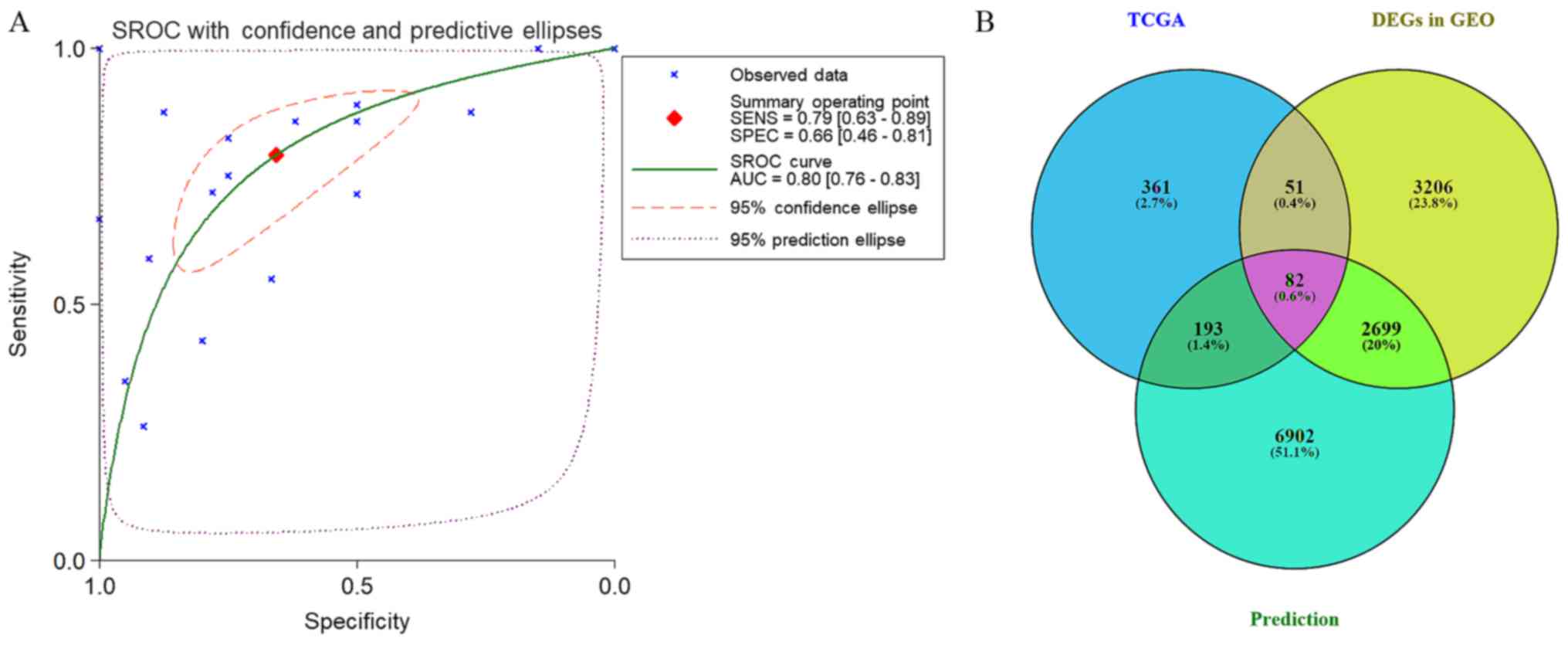
tissues: Sub-SMD (95% CI)=−0.304 (−0.452, −0.156) (Table II and Fig. 3A; P<0.001) PCa, vs. normal=421,
vs. 253); sROC AUC (95% CI)=0.80 (0.77, 0.84) (Fig. 4). The optimum sensitivity and
specificity were 0.82 (95% CI: 0.65, 0.91) and 0.61 (95% CI: 0.34,
0.83, Fig. 4), respectively. The
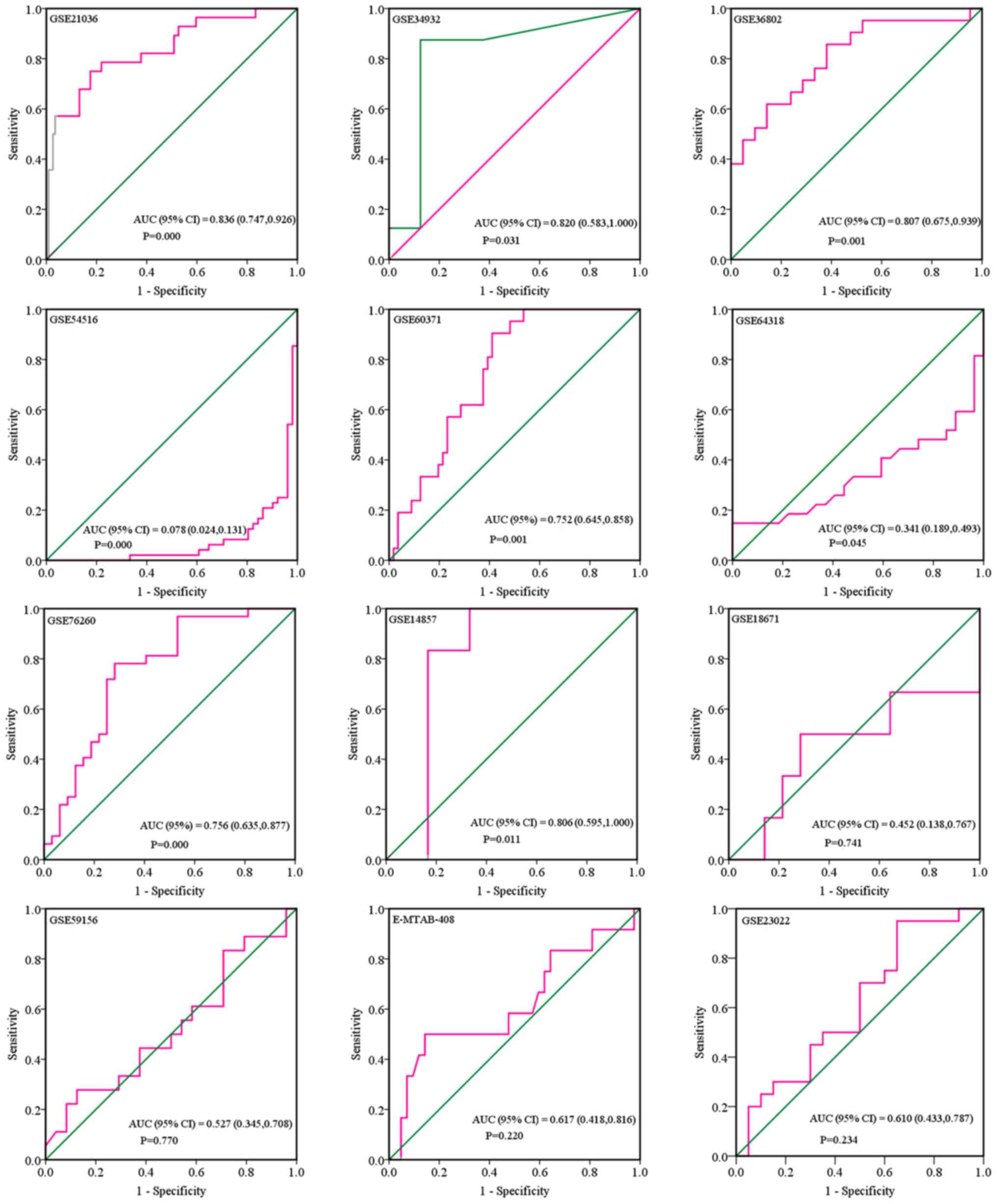
scatter diagram and ROC curve of expression of each microarray are
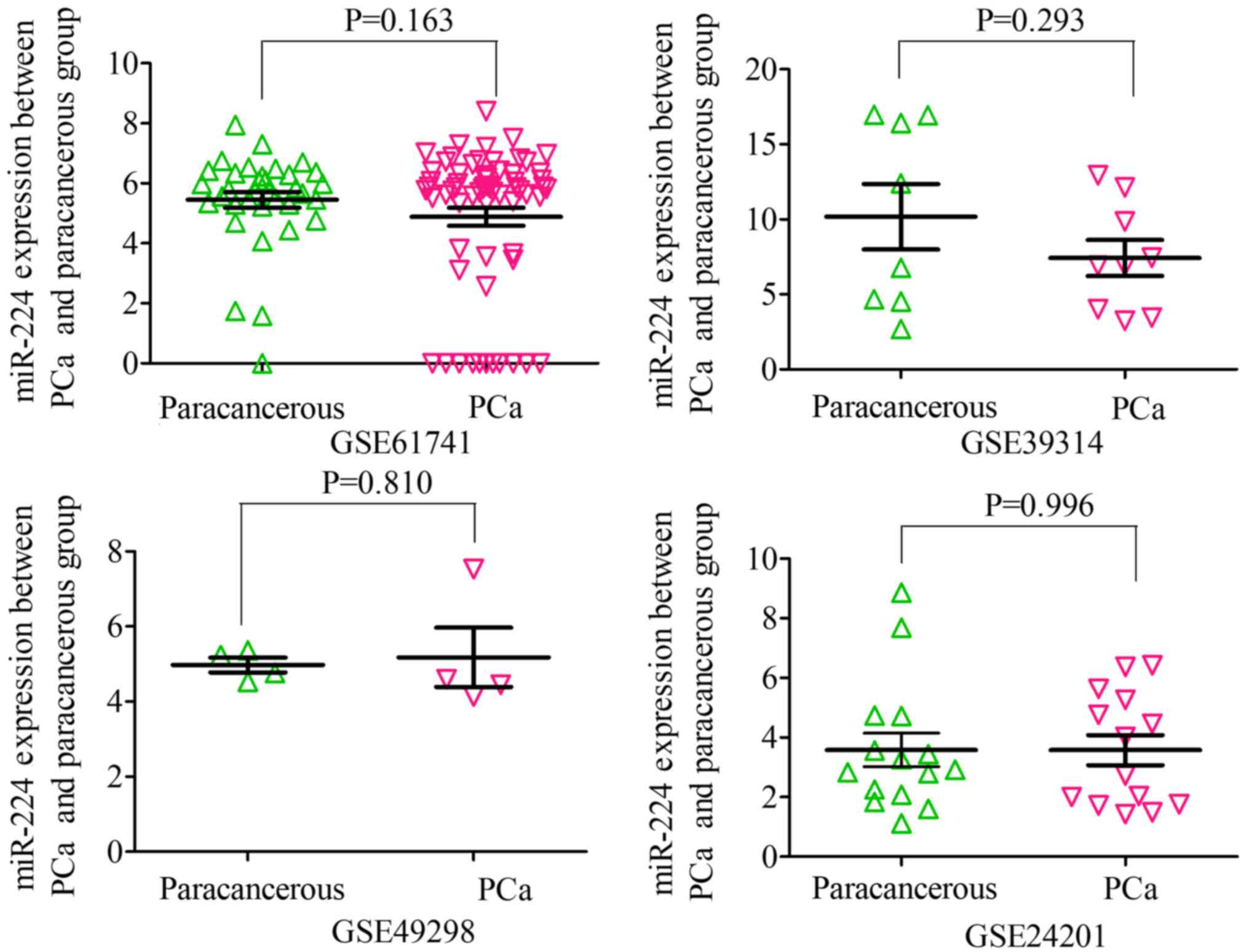
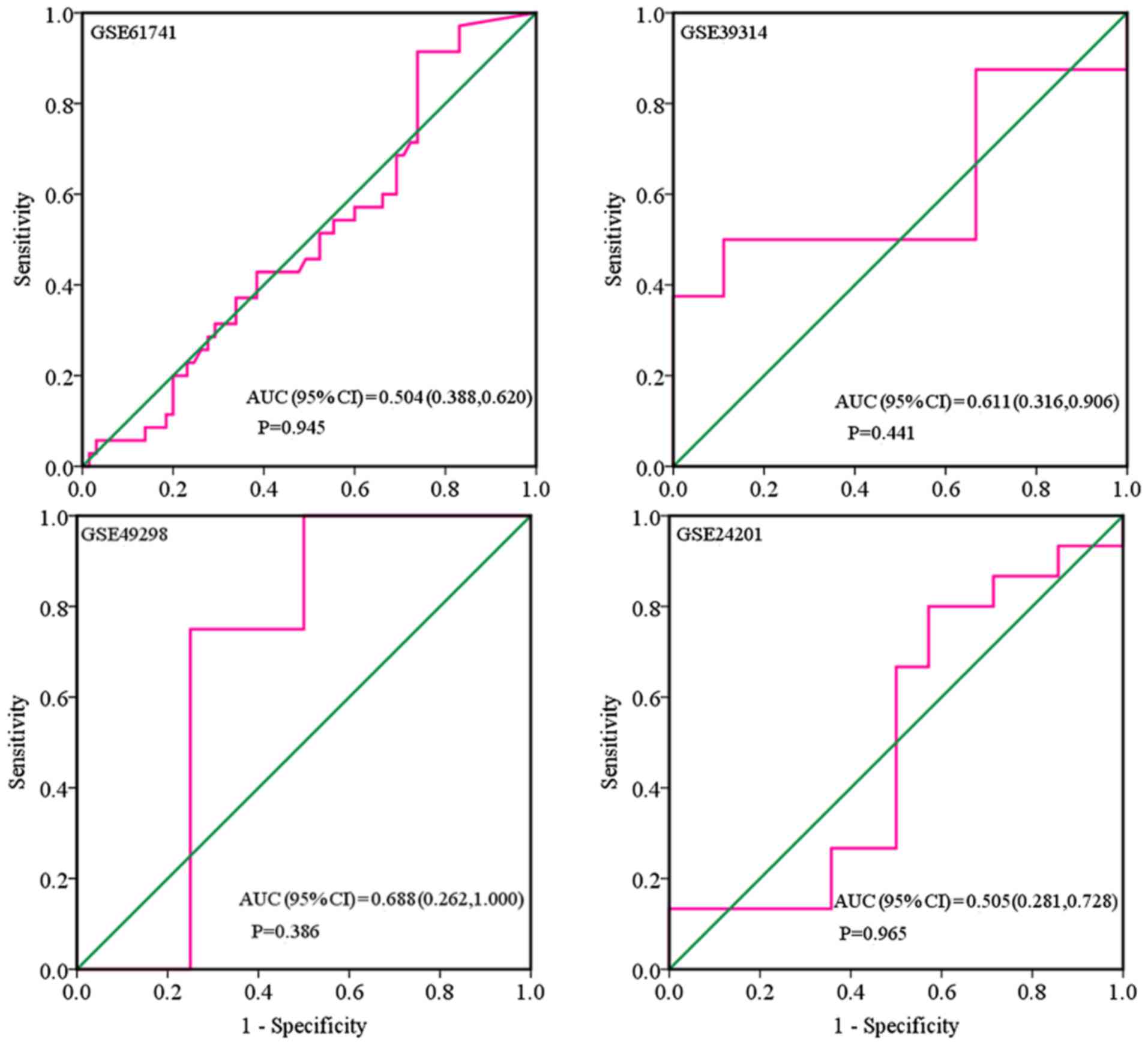
shown in Figs. 5 and 6. When analyzing the biofluids subgroup,
four microarrays were included (GSE39314, GSE61741, GSE24201 and
GSE49298). The SMD result indicated that Sub-SMD (95% CI)=−0.216
(−0.544, −0.111), P=0.195 (Table
III and Fig. 3B) PCa, vs.
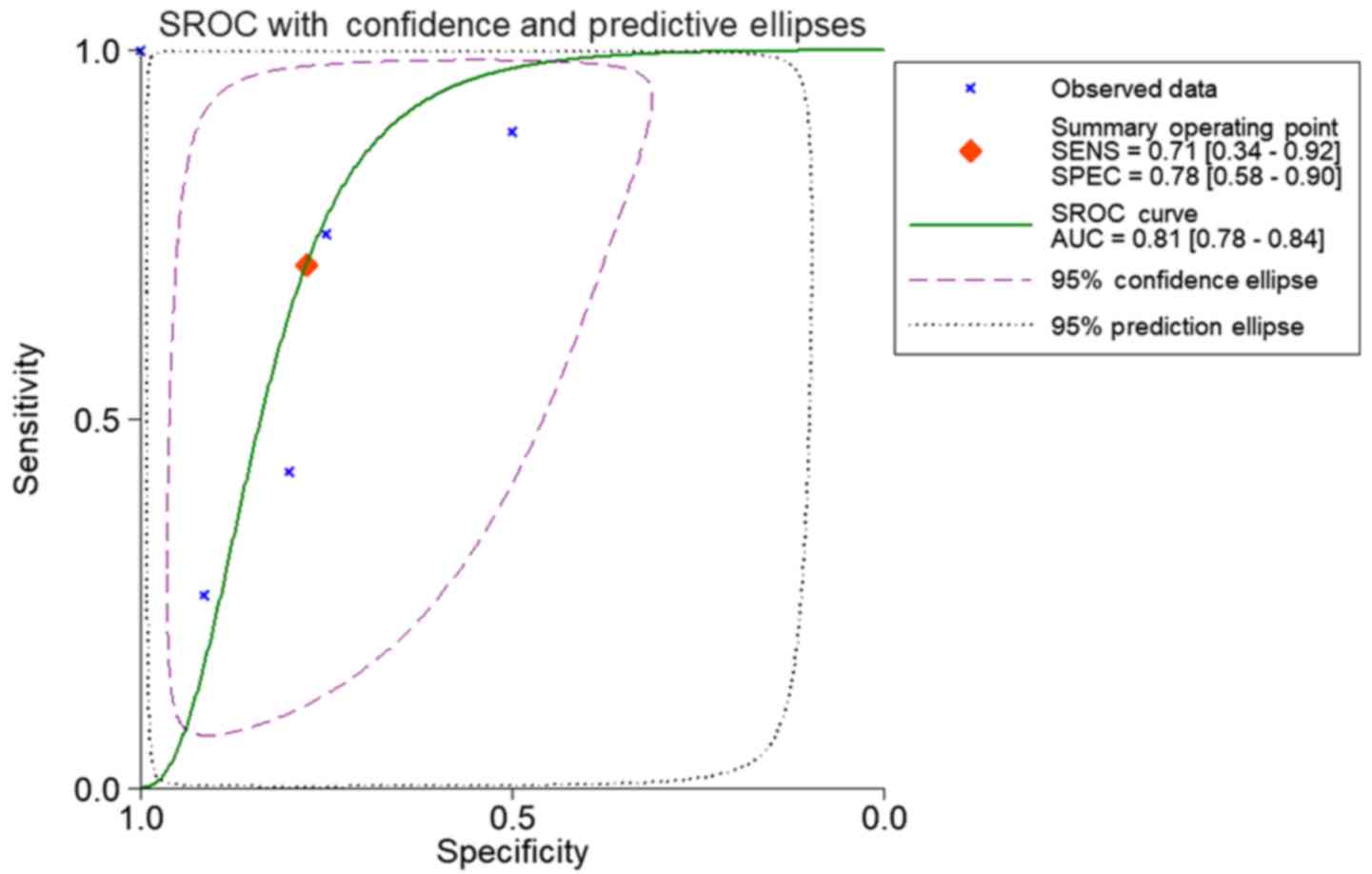
normal, 92, vs 62. sROC analysis revealed the following: AUC (95%
CI)=0.81 (0.78, 0.84), sensitivity (SENS; 95% CI)=0.71 (0.34,
0.92), specificity (SPEC; 95% CI)=0.78 (0.58, 0.90) (Fig. 7). The scatter diagrams and ROC
curves are shown in Figs. 8 and
9. The expression of miR-224-5p was
markedly lower in PCa biofluids despite no clear significance. In
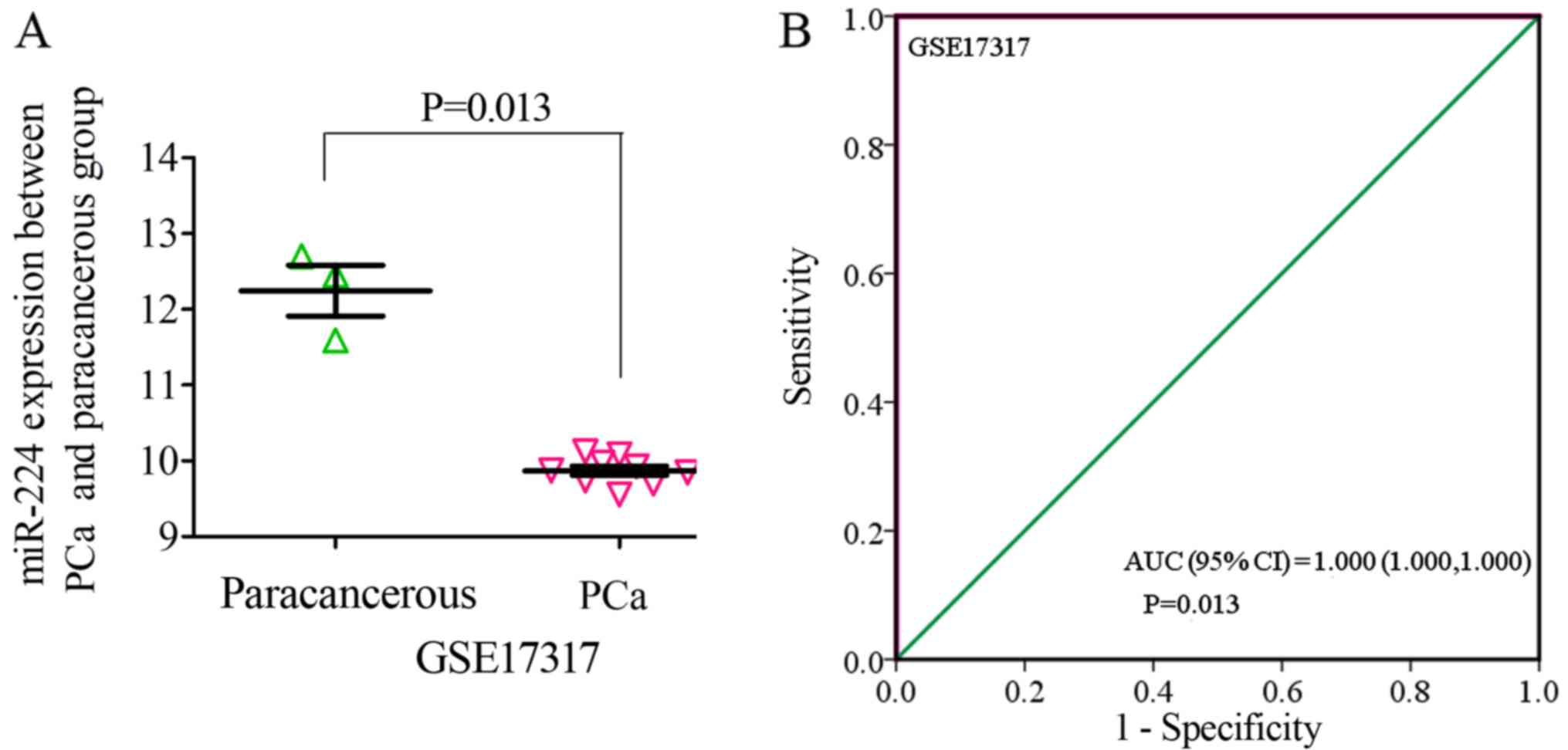
the subgroup of cell lines, only one microarray was included: Mean
± SD: 9.8705±0.1835 (P=0.013, PCa, vs. normal, 9, vs. 3; ROC AUC
(95% CI)=1.000 (1.000, 1.000), P=0.013 (Fig. 10). The expression of miR-224-5p was
significantly lower in the PCa cell lines, and exhibited higher
specificity.
| Table II.Characteristics of the expression of
microRNA-224-5p between studies of prostate adenocarcinoma and
non-cancerous tissue based on Gene Expression Omnibus, ArrayExpress
and TCGA datasets. |
Table II.
Characteristics of the expression of
microRNA-224-5p between studies of prostate adenocarcinoma and
non-cancerous tissue based on Gene Expression Omnibus, ArrayExpress
and TCGA datasets.
|
| Patients | Controls |
|
|
|---|
|
|
|
|
|
|
|---|
| Study | N | Mean | SD | N | Mean | SD | T-value | P-value |
|---|
| GSE54516 | 51 | 4.3588 | 0.0525 | 48 | 4.2649 | 0.0420 | 9.858 | <0.001 |
| GSE59156 | 24 | 3.6686 | 1.0369 | 18 | 4.0963 | 1.6407 | −1.033 | 0.308 |
| GSE64318 | 27 | 3.0859 | 0.9412 | 27 | 2.0776 | 1.7120 | 2.682 | 0.011 |
| GSE76260 | 32 | 3.6162 | 0.0662 | 32 | 3.6723 | 0.0512 | −3.794 | <0.001 |
| GSE14857 | 12 | 5.0417 | 1.3522 | 12 | 5.9110 | 0.5401 | −2.068 | 0.051 |
| GSE18671 | 14 | 6.6809 | 0.3943 | 6 | 6.4473 | 0.5782 | 1.057 | 0.305 |
| GSE21036 | 114 | 5.2423 | 1.0451 | 28 | 6.6041 | 0.9873 | −6.243 | <0.001 |
| GSE34932 | 8 | 2.1788 | 1.0875 | 8 | 3.5600 | 1.0803 | −2.549 | 0.023 |
| GSE23022 | 20 | 0.9755 | 0.2065 | 20 | 1.0201 | 0.1732 | −0.740 | 0.464 |
| GSE60371 | 56 | 6.3003 | 0.4778 | 21 | 6.6752 | 0.3432 | −3.285 | 0.002 |
| GSE36802 | 21 | 4.4416 | 0.9294 | 21 | 5.7362 | 1.1766 | −3.957 | <0.001 |
| E-MTAB-408 | 42 | 3.2419 | 1.5142 | 12 | 3.7560 | 1.6093 | −1.023 | 0.311 |
| TCGA | 498 | 5.0164 | 1.2154 | 52 | 5.4589 | 0.8661 | −3.413 | 0.001 |
| Total |
| Standardized mean
difference (95% CI)=−0.304 (−0.452, −0.156) P<0.001 |
|
| Table III.Characteristics of the expression of
microRNA-224-5p between prostate adenocarcinoma and non-cancerous
body fluid studies based on the Gene Expression Omnibus
dataset. |
Table III.
Characteristics of the expression of
microRNA-224-5p between prostate adenocarcinoma and non-cancerous
body fluid studies based on the Gene Expression Omnibus
dataset.
|
| Patients | Controls |
|
|
|---|
|
|
|
|
|
|
|---|
| Study | N | Mean | SD | N | Mean | SD | T-value | P-value |
|---|
| GSE39314 | 9 | 7.4306 | 3.6046 | 8 | 10.1763 | 6.1584 | −1.104 | 0.293 |
| GSE61741 | 65 | 4.8860 | 2.4333 | 35 | 5.4507 | 1.5656 | −1.407 | 0.163 |
| GSE24201 | 14 | 3.5766 | 1.8929 | 15 | 3.5814 | 2.1733 | −0.006 | 0.996 |
| GSE49298 | 4 | 5.1792 | 1.5787 | 4 | 4.9743 | 0.3945 | 0.252 | 0.810 |
| Total |
| Standardized mean
difference (95% CI)=−0.216 (−0.544, 0.111) P=0.195 |
|
Furthermore, in the literature search, three studies
were retrieved providing a mean ± SD (Fig. 1C) (15–17).
The results also suggested that the expression of miR-224-5p was
downregulated in PCa tissues: SMD (95% CI)=−2.245 (−2.587, −1.904),
P<0.001 (Table IV and Fig. 3D; PCa, vs. normal, 129, vs. 100).
The results of all studies provided the following results: Overall
SMD (95% CI)=−0.562 (−0.687, −0.436; P<0.001); overall sROC AUC
(95% CI)=0.80 (0.76, 0.83); SENS (95% CI) 0.79 (0.63, 0.89); SPEC
(95% CI)=0.66 (0.46, 0.81) (Table V
and Figs. 3 and 11A). This provided a more reliable
conclusion that the expression of miR-224-5p was reduced in
PCa.
| Table IV.Characteristics of three studies
selected from previous literature. |
Table IV.
Characteristics of three studies
selected from previous literature.
|
|
| Control | Prostate
adenocarcinoma |
|
|
|---|
|
|
|
|
|
|
|
|---|
| Author, date | Country | N | Mean ± SD | N | Mean ± SD | Detection
method | Refs:PMID |
|---|
| Fu et al
(27) | China | 20 | 2.171±0.564 | 20 | 1.344±0.358 | RT-qPCR | 25394900 |
| Mavridis et
al (26) | Athens | 66 | 8.470±2.090 | 73 | 4.228±0.809 | RT-qPCR | 23136246 |
| Kristensen et
al (28) | Denmark | 14 | 2.929±0.147 | 36 | 2.562±0.252 | RT-qPCR | 24737792 |
| Total | Standardized mean
difference (95% CI)=−2.245 (−2.587, −1.904) P=0.195,
P<0.001 |
|
| Table V.Characteristics of the expression of
microRNA-224-5p between prostate adenocarcinoma and non-cancerous
studies. |
Table V.
Characteristics of the expression of
microRNA-224-5p between prostate adenocarcinoma and non-cancerous
studies.
|
| Patients | Controls |
|
|
|---|
|
|
|
|
|
|
|---|
| Study | N | Mean | SD | N | Mean | SD | T-value | P-value |
|---|
| GSE54516 | 51 | 4.3588 | 0.0525 | 48 | 4.2649 | 0.0420 | 9.858 | <0.001 |
| GSE59156 | 24 | 3.6686 | 1.0369 | 18 | 4.0963 | 1.6407 | −1.033 | 0.308 |
| GSE64318 | 27 | 3.0859 | 0.9412 | 27 | 2.0776 | 1.7120 | 2.682 | 0.011 |
| GSE76260 | 32 | 3.6162 | 0.0662 | 32 | 3.6723 | 0.0512 | −3.794 | <0.001 |
| GSE14857 | 12 | 5.0417 | 1.3522 | 12 | 5.9110 | 0.5401 | −2.068 | 0.051 |
| GSE18671 | 14 | 6.6809 | 0.3943 | 6 | 6.4473 | 0.5782 | 1.057 | 0.305 |
| GSE21036 | 114 | 5.2423 | 1.0451 | 28 | 6.6041 | 0.9873 | −6.243 | <0.001 |
| GSE34932 | 8 | 2.1788 | 1.0875 | 8 | 3.5600 | 1.0803 | −2.549 | 0.023 |
| GSE23022 | 20 | 0.9755 | 0.2065 | 20 | 1.0201 | 0.1732 | −0.740 | 0.464 |
| GSE60371 | 56 | 6.3003 | 0.4778 | 21 | 6.6752 | 0.3432 | −3.285 | 0.002 |
| GSE36802 | 21 | 4.4416 | 0.9294 | 21 | 5.7362 | 1.1766 | −3.957 | <0.001 |
| E-MTAB-408 | 42 | 3.2419 | 1.5142 | 12 | 3.7560 | 1.6093 | −1.023 | 0.311 |
| TCGA | 498 | 5.0164 | 1.2154 | 52 | 5.4589 | 0.8661 | −3.413 | 0.001 |
| GSE39314 | 9 | 7.4306 | 3.6046 | 8 | 10.1763 | 6.1584 | −1.104 | 0.293 |
| GSE61741 | 65 | 4.8860 | 2.4333 | 35 | 5.4507 | 1.5656 | −1.407 | 0.163 |
| GSE24201 | 14 | 3.5766 | 1.8929 | 15 | 3.5814 | 2.1733 | −0.006 | 0.996 |
| GSE49298 | 4 | 5.1792 | 1.5787 | 4 | 4.9743 | 0.3945 | 0.252 | 0.810 |
| GSE17317 | 9 | 9.8705 | 0.1835 | 3 | 12.2457 | 0.5820 | −7.559 | 0.013 |
| Kristensen et
al (28) | 36 | 2.5623 | 0.2523 | 14 | 2.9291 | 0.1469 |
| <0.001 |
| Fu et al
(27) | 20 | 1.3444 | 0.3578 | 20 | 2.1705 | 0.5643 |
| <0.001 |
| Mavridis et
al (26) | 73 | 4.2280 | 0.8090 | 66 | 8.4700 | 2.0900 |
| <0.001 |
| Total |
| Standardized mean
difference (95% CI)=−0.562 (−0.687, −0.436) P<0.001 |
|
Prospective target genes of miR-224-5p
in PCa
The prospective target genes of moR-224-5p in PCa
were determined based on the results of TCGA, GEO and prediction
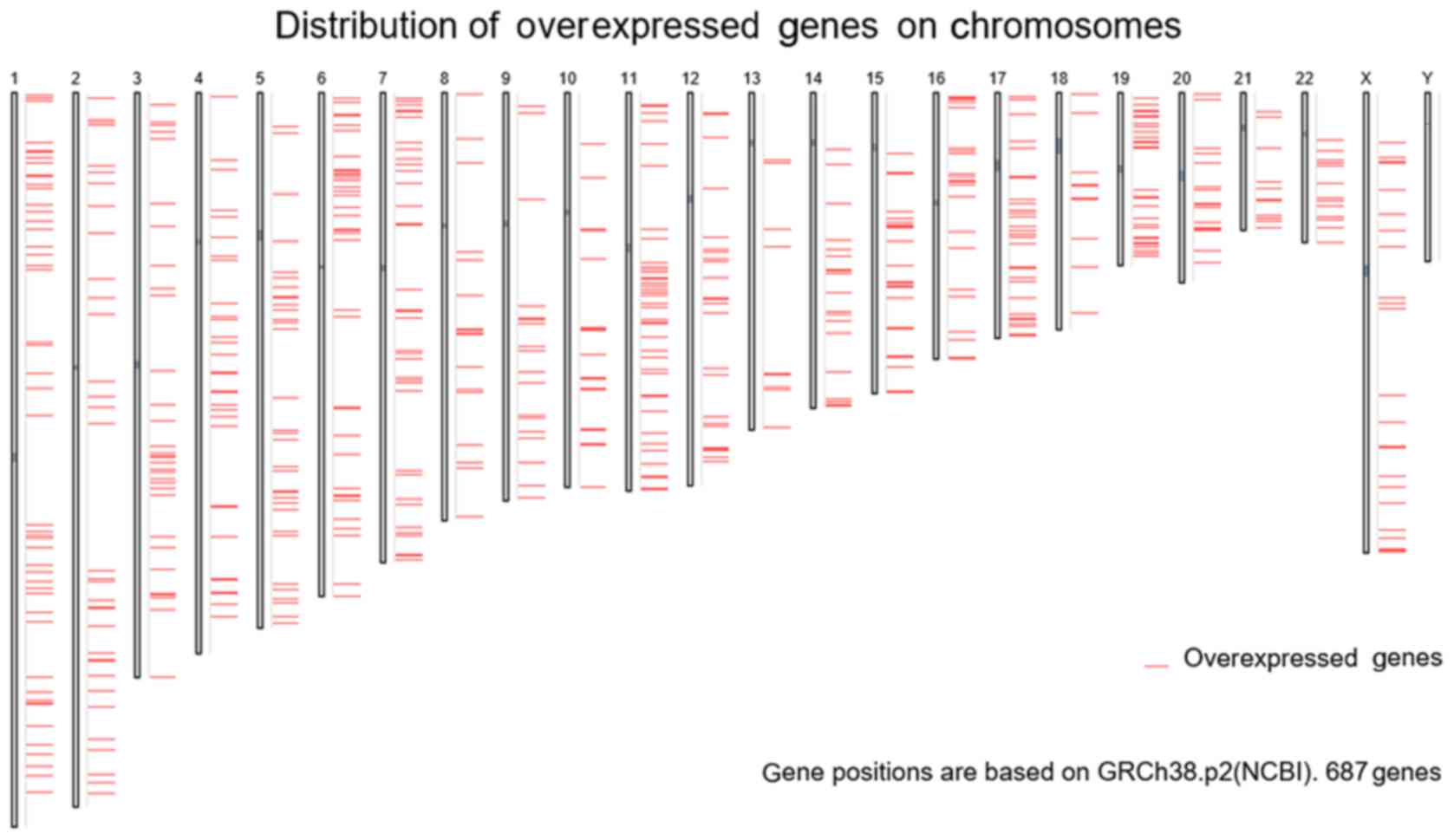
tools. Based on TCGA data, 3,019 differentially expressed genes in
PCa tissues were acquired from GEPIA, and the FC was used as a
measure (log2FC>1.0). Finally, 687 overexpressed genes were
obtained from the PCa tissues (Fig.
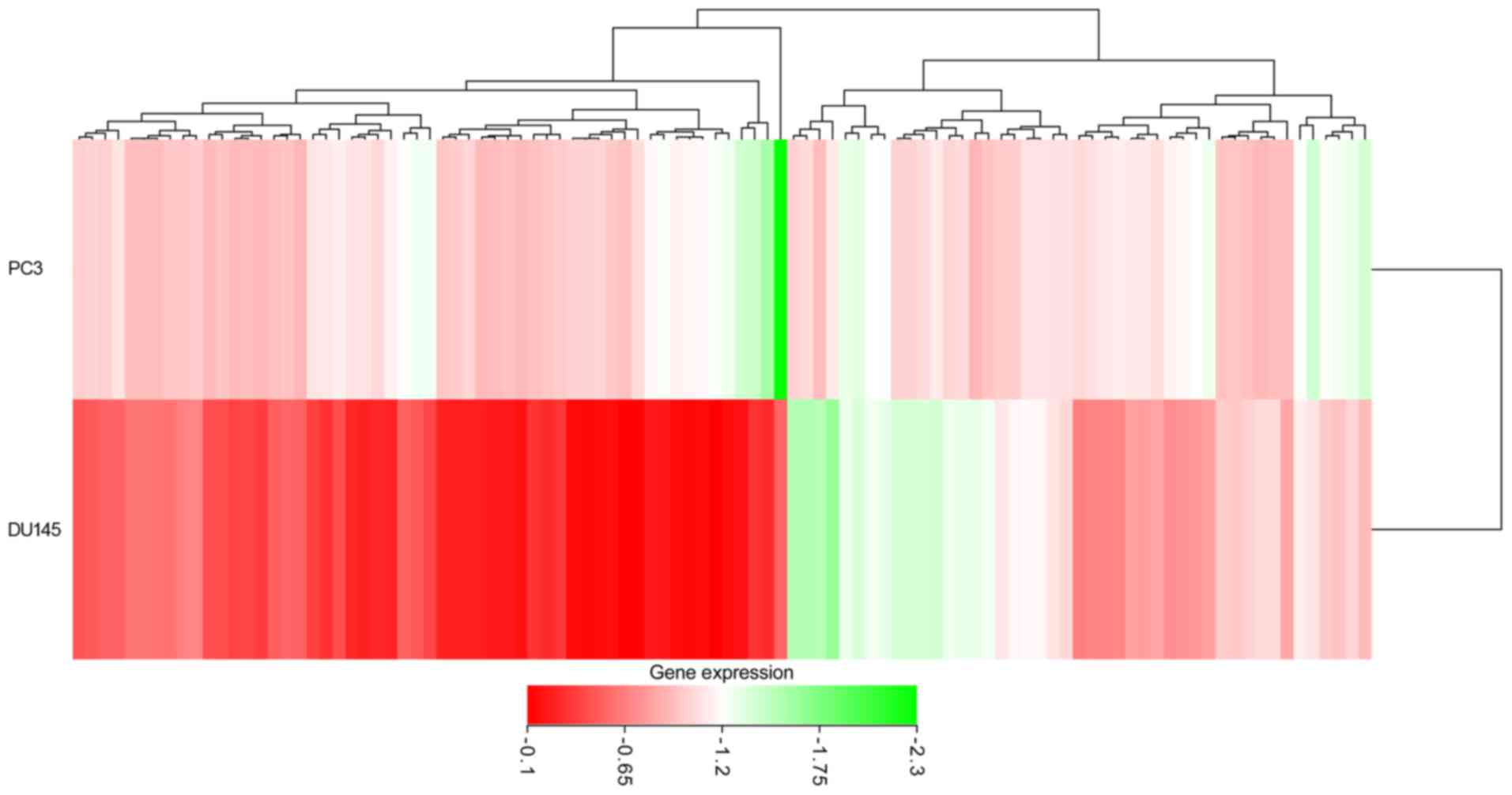
12). In addition, from GEO two cell lines were retrieved that
were transfected with PC3 and DU145, and the microarrays of
differentially expressed mRNA (GSE51053 and GSE56243) were
examined. According to the unions in which the gene expression
value was below −0.1, a total of 3,616 (GSE51053) and 3,326
(GSE56243) genes with low expression were obtained. Following
combining the results of two microarrays and eliminating the
duplicates, 6,038 genes with low expression that had been
transfected with miR-224-5p were obtained. The top 100 genes are
shown in the heat-map in Fig. 13.
In addition, 102,240 potential target genes of miR-224-5p were
accumulated via miRWalk2.0. Genes that appeared in at least three
prediction tools were selected, with 9,876 genes acquired. A total
of 82 overlapped mRNA genes qualified for further analysis
following combining the results of TCGA, GEO and prediction tools
(Fig. 11B).
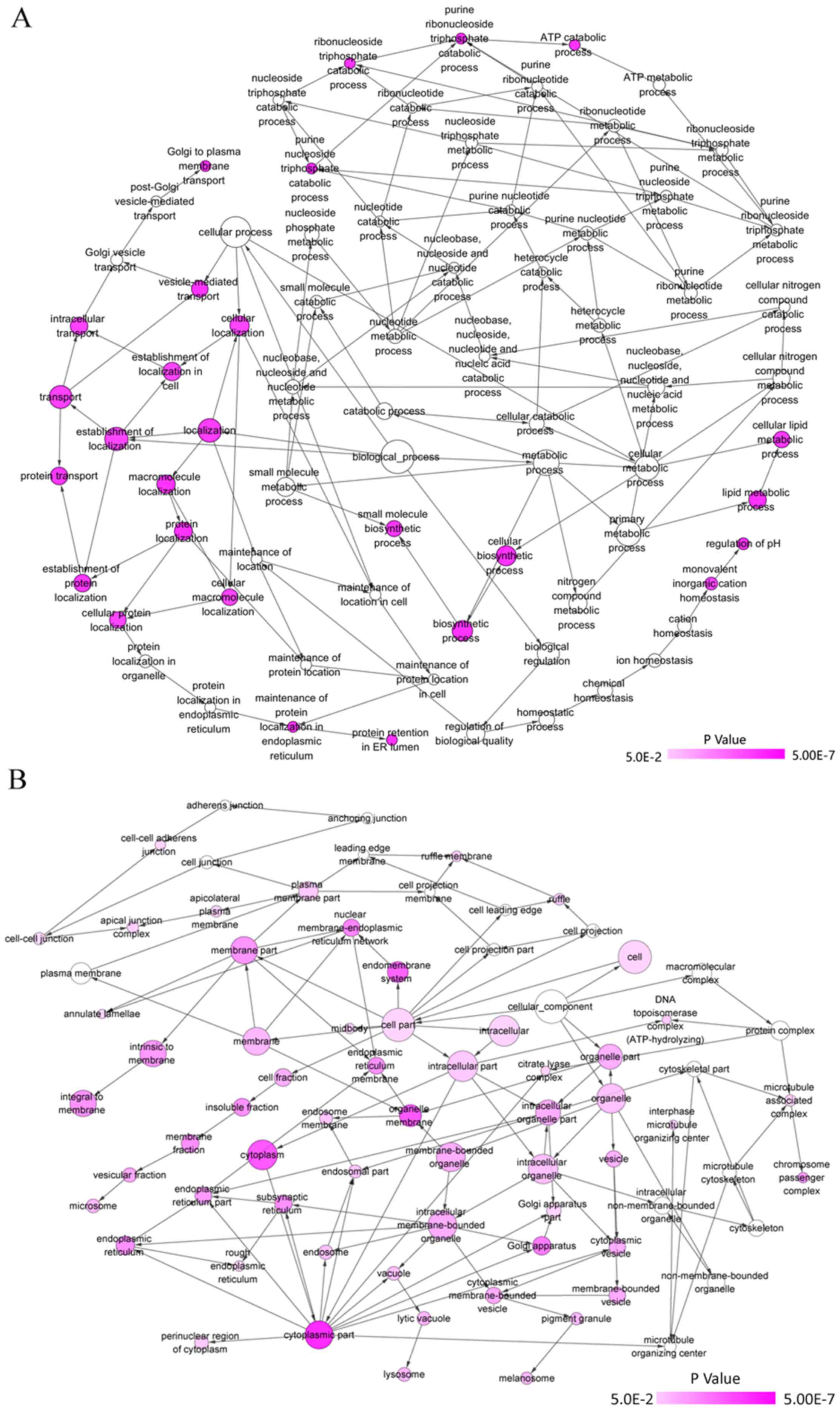
In silico analysis
GO and KEGG analyses were performed for the 82
overlapped target genes using DAVID, which indicated that the GO
terms were enriched in the following pathways: Intracellular
transport, vesicle-mediated transport, protein transport of
biological processes; Golgi apparatus, membrane fraction, insoluble
fraction of cellular components; protein homodimerization activity,
protein dimerization activity, identical protein binding of
molecular functions (Table VI and
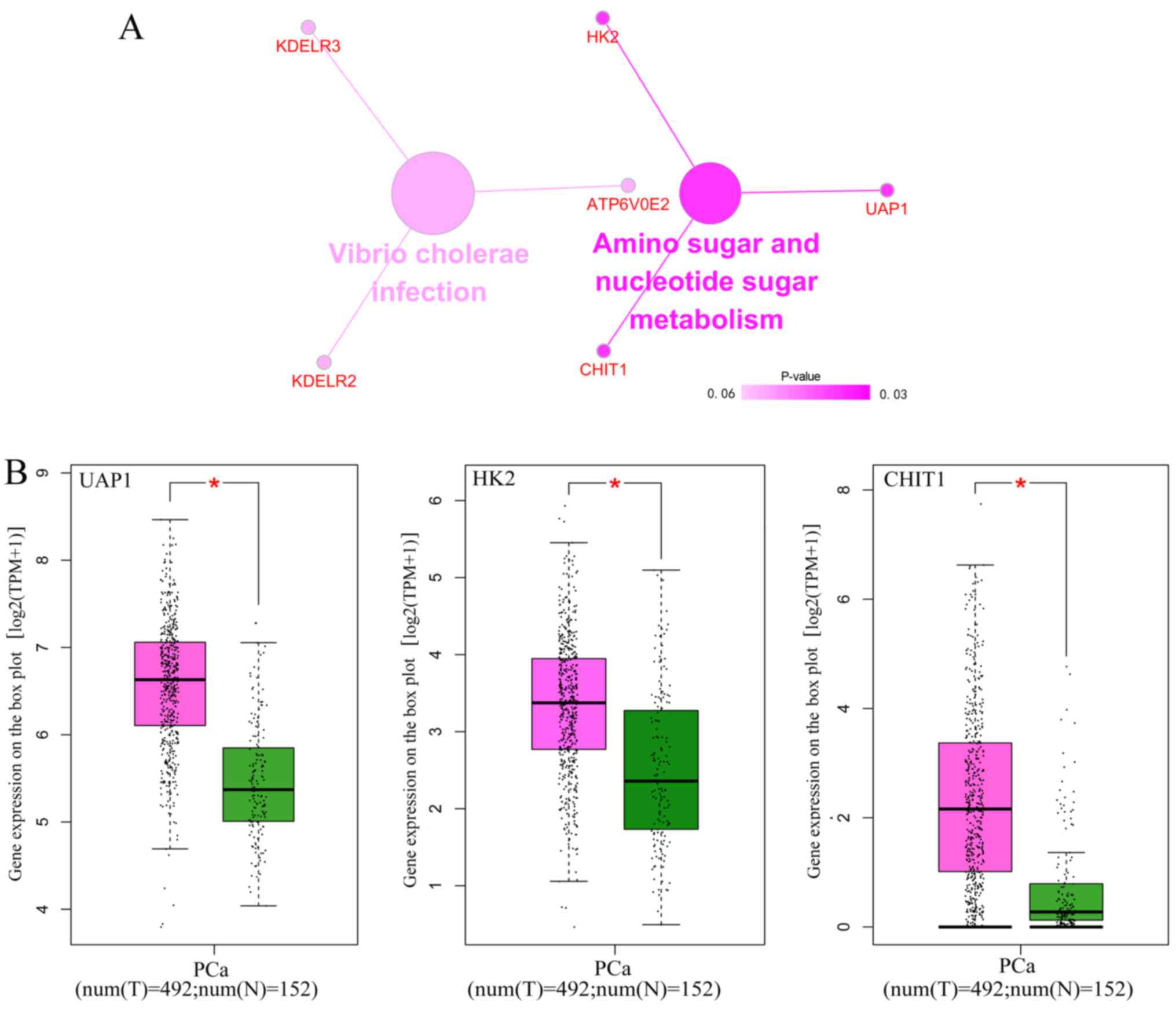
Figs. 14 and 15). KEGG analysis demonstrated that these
82 genes were simply enriched in two pathways: Amino sugar and
nucleotide sugar metabolism, and Vibrio cholerae infection.
Significance was only found in the amino sugar and nucleotide sugar
metabolism pathway (P<0.05; Table
VI and Fig. 16A), in which
UDP-N-acetylglucosamine pyrophosphorylase 1 (UAP1), hexokinase 2
(HK2) and chitinase 1 (CHIT1) were notably upregulated in PCa
tissues (P<0.05; Fig. 16B). The
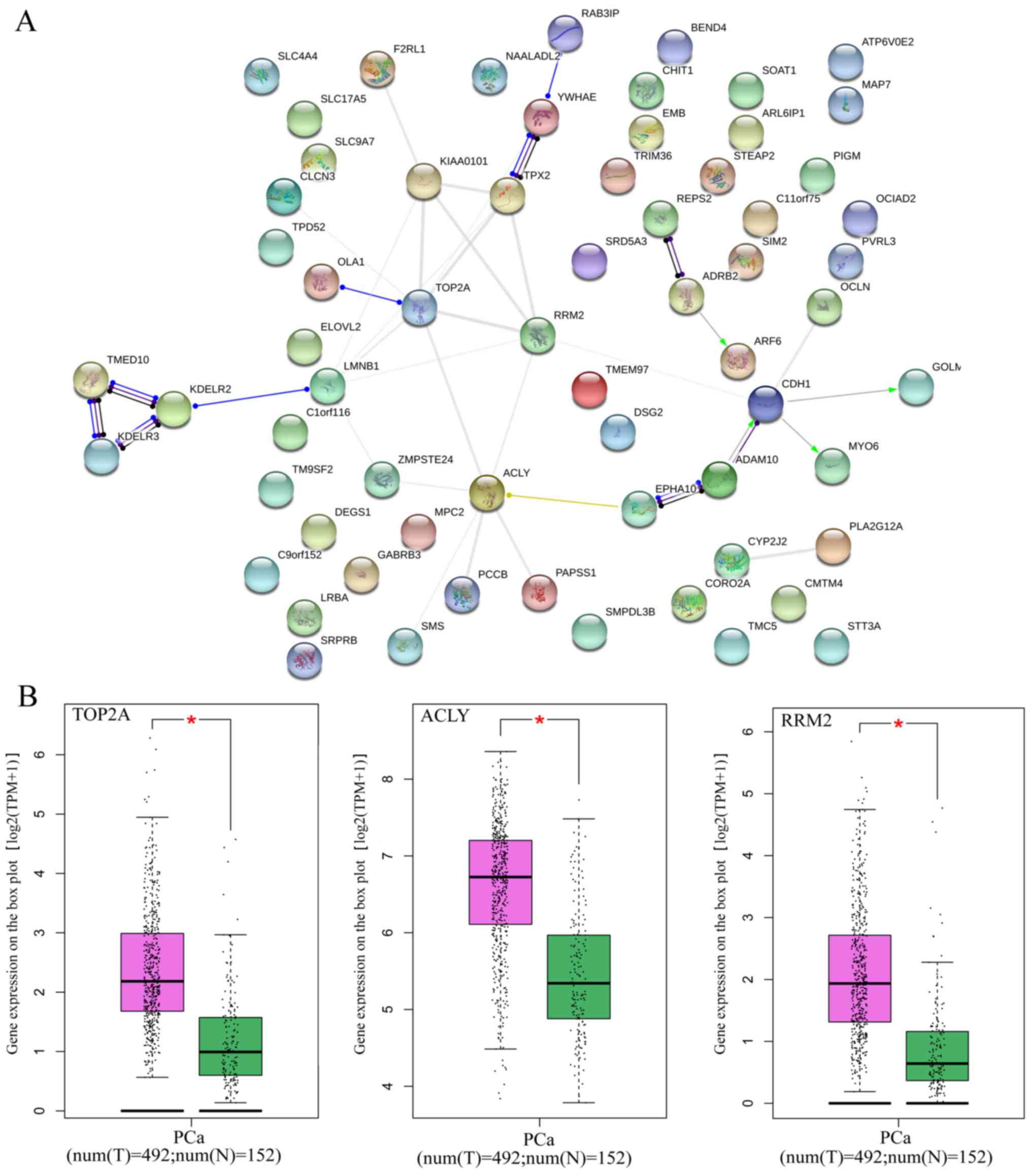
PPI analysis suggested that DNA topoisomerase 2-α (TOP2A), ATP
citrate lyase (ACLY) and ribonucleotide reductase regulatory
subunit M2 (RRM2) were key genes for protein interaction, which had
higher expression levels in PCa tissues (P<0.05; Table VII and Fig. 17). miR-224-5p may have negative
axial associations with the expression of UAP1, HK2, CHIT1, TOP2A,
ACLY and RRM2 in PCa, which requires further experiments for
confirmation (Fig. 18).
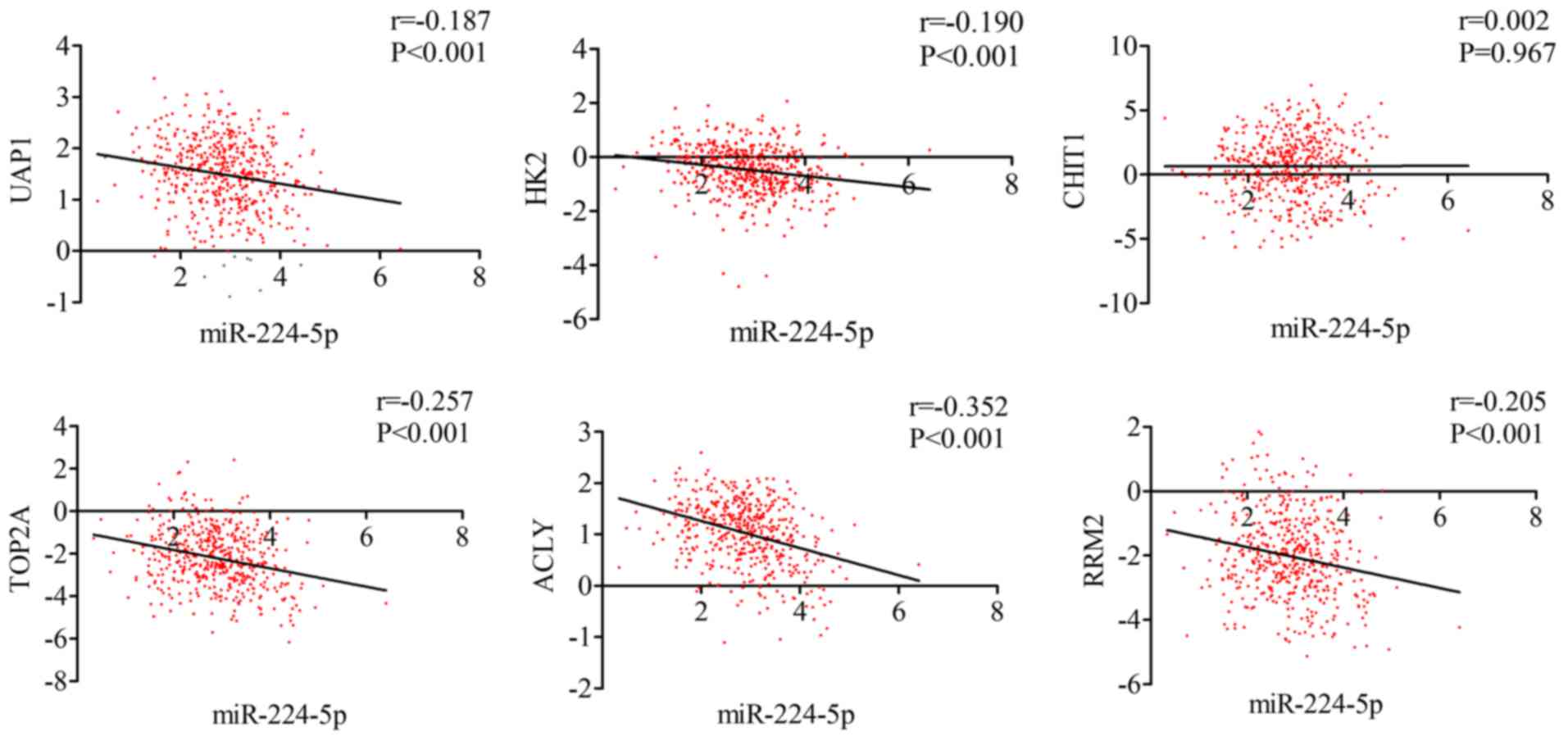 | Figure 18.Correlation between the targets
(UAP1, HK2, CHIT1, TOP2A, ACLY, RRM2) and miR-224-5p in prostate
adenocarcinoma. miR, microRNA; UAP1, UDP-N-acetylglucosamine
pyrophosphorylase 1; HK2, hexokinase 2; CHIT1, chitinase 1; TOP2A,
DNA topoisomerase 2-α; ACLY, ATP citrate lyase; RRM2,
ribonucleotide reductase regulatory subunit M2. |
| Table VI.Top 10 most significant GO terms of
the potential target genes of microRNA-224-5-p in prostate
adenocarcinoma. |
Table VI.
Top 10 most significant GO terms of
the potential target genes of microRNA-224-5-p in prostate
adenocarcinoma.
| Category | Term | Count | P-value | FDR |
|---|
| Biological
process |
|
GOTERM_BP_FAT |
GO:0046907~intracellular transport | 10 | 0.002874 | 4.327707 |
|
GOTERM_BP_FAT |
GO:0016192~vesicle-mediated transport | 9 | 0.004584 | 6.818167 |
|
GOTERM_BP_FAT | GO:0015031~protein
transport | 10 | 0.007548 | 10.992314 |
|
GOTERM_BP_FAT |
GO:0045184~establishment of protein
localization | 10 | 0.007995 | 11.607428 |
|
GOTERM_BP_FAT | GO:0006163~purine
nucleotide metabolic process | 5 | 0.010383 | 14.821117 |
|
GOTERM_BP_FAT |
GO:0006885~regulation of pH | 3 | 0.010551 | 15.043556 |
|
GOTERM_BP_FAT | GO:0034613~cellular
protein localization | 7 | 0.011114 | 15.783945 |
|
GOTERM_BP_FAT | GO:0070727~cellular
macromolecule localization | 7 | 0.011491 | 16.275859 |
|
GOTERM_BP_FAT | GO:0008610~lipid
biosynthetic process | 6 | 0.016047 | 22.014591 |
|
GOTERM_BP_FAT | GO:0006605~protein
targeting | 5 | 0.016883 | 23.026892 |
| Cellular
component |
|
GOTERM_CC_FAT | GO:0005794~Golgi
apparatus | 15 | 0.000193 | 0.241372 |
|
GOTERM_CC_FAT | GO:0005624~membrane
fraction | 14 | 0.000337 | 0.419916 |
|
GOTERM_CC_FAT |
GO:0005626~insoluble fraction | 14 | 0.000477 | 0.595260 |
|
GOTERM_CC_FAT |
GO:0005783~endoplasmic reticulum | 15 | 0.000517 | 0.644979 |
|
GOTERM_CC_FAT |
GO:0042598~vesicular fraction | 7 | 0.001999 | 2.470347 |
|
GOTERM_CC_FAT | GO:0000267~cell
fraction | 14 | 0.004767 | 5.799806 |
|
GOTERM_CC_FAT |
GO:0031090~organelle membrane | 14 | 0.005271 | 6.394020 |
|
GOTERM_CC_FAT |
GO:0005768~endosome | 7 | 0.006966 | 8.368649 |
|
GOTERM_CC_FAT | GO:0016021~integral
to membrane | 40 | 0.008019 | 9.576264 |
|
GOTERM_CC_FAT |
GO:0005792~microsome | 6 | 0.009016 | 10.705916 |
| Molecular
function |
|
GOTERM_MF_FAT | GO:0042803~protein
homodimerization activity | 8 | 0.000927 | 1.226713 |
|
GOTERM_MF_FAT | GO:0046983~protein
dimerization activity | 9 | 0.003674 | 4.779822 |
|
GOTERM_MF_FAT |
GO:0042802~identical protein binding | 9 | 0.009776 | 12.255478 |
|
GOTERM_MF_FAT | GO:0043176~amine
binding | 4 | 0.014922 | 18.132938 |
|
GOTERM_MF_FAT | GO:0046982~protein
heterodimerization activity | 5 | 0.016408 | 19.761183 |
|
GOTERM_MF_FAT | GO:0046923~ER
retention sequence binding | 2 | 0.018664 | 22.176377 |
|
GOTERM_MF_FAT |
GO:0031406~carboxylic acid binding | 4 | 0.030136 | 33.449456 |
|
GOTERM_MF_FAT | GO:0016597~amino
acid binding | 3 | 0.032286 | 35.386583 |
|
GOTERM_MF_FAT | GO:0019899~enzyme
binding | 7 | 0.035546 | 38.224143 |
|
GOTERM_MF_FAT |
GO:0015297~antiporter activity | 3 | 0.040576 | 42.376479 |
| KEGG category | Term | Count | P-value | Genes |
|
KEGG_PATHWAY | hsa00520: Amino
sugar and nucleotide sugar metabolism | 3 | 0.038278 | UAP1, HK2,
CHIT1 |
|
KEGG_PATHWAY | hsa05110: Vibrio
cholerae infection | 3 | 0.059125 | KDELR3, KDELR2
ATP6V0E2 |
| Table VII.Top 10 genes with combined scores in
the protein-protein interaction network of potential target genes
of microRNA-224-5p in prostate adenocarcinoma. |
Table VII.
Top 10 genes with combined scores in
the protein-protein interaction network of potential target genes
of microRNA-224-5p in prostate adenocarcinoma.
| Node 1 | Node 2 | Node 1 string
internal ID | Co-expression | Experimentally
determined interaction | Database
annotated | Automated text
mining | Combined score |
|---|
| TOP2A | RRM2 | 1860923 | 0.880 | 0.000 | 0.000 | 0.673 | 0.960 |
| MYO6 | CDH1 | 1855148 | 0.000 | 0.360 | 0.900 | 0.214 | 0.945 |
| CDH1 | ADAM10 | 1844988 | 0.050 | 0.091 | 0.900 | 0.409 | 0.942 |
| TMED10 | KDELR2 | 1848494 | 0.310 | 0.137 | 0.900 | 0.105 | 0.939 |
| KDELR3 | TMED10 | 1859221 | 0.270 | 0.137 | 0.900 | 0.105 | 0.936 |
| TOP2A | KIAA0101 | 1860923 | 0.926 | 0.000 | 0.000 | 0.145 | 0.934 |
| RAB3IP | YWHAE | 1862399 | 0.000 | 0.346 | 0.900 | 0.000 | 0.931 |
| TOP2A | TPX2 | 1860923 | 0.879 | 0.000 | 0.000 | 0.340 | 0.916 |
| EPHA10 | ADAM10 | 1855819 | 0.053 | 0.000 | 0.900 | 0.111 | 0.908 |
| ADRB2 | ARF6 | 1848724 | 0.000 | 0.000 | 0.900 | 0.112 | 0.907 |
| TOP2A | RRM2 | 1860923 | 0.880 | 0.000 | 0.000 | 0.673 | 0.960 |
| MYO6 | CDH1 | 1855148 | 0.000 | 0.360 | 0.900 | 0.214 | 0.945 |
| CDH1 | ADAM10 | 1844988 | 0.050 | 0.091 | 0.900 | 0.409 | 0.942 |
| TMED10 | KDELR2 | 1848494 | 0.310 | 0.137 | 0.900 | 0.105 | 0.939 |
| KDELR3 | TMED10 | 1859221 | 0.270 | 0.137 | 0.900 | 0.105 | 0.936 |
Discussion
In the present study, referring to data in the TCGA,
GEO, ArrayExpress and previous literature, it was confirmed that
the expression of miR-224-5p was notably downregulated in PCa
tissues, biofluids and cell lines, and its downregulated expression
may be associated with the progression of PCa. In addition,
prospective target genes of miR-224-5p in PCa were identified via
prediction tools and microarrays of differentially expressed mRNAs
in TCGA and GEO. Bioinformatics analysis was subsequently performed
on these potential genes. GO and KEGG analyses revealed that
miR-224-5p mediated the expression of UAP1, HK2 and CHIT1 in order
to regulate the amino sugar and nucleotide sugar metabolism
signaling pathway, thus exerting its effects in PCa. miR-224-5p may
also be vital in PCa by being involved in the protein interaction
through TOP2A, ACLY and RRM2.
Previous studies have demonstrated that the abnormal
expression of miR-224-5p was a crucial factor in the initiation and
progress of tumors. For example, the high expression of miR-224-5p
was likely to be involved in the onset of digestive tract
malignancy (38); the expression of
miR-224-5p was significantly reduced in mucinous breast cancer
(39); and cell experiments and
histologic examination performed by Zheng et al revealed
that miR-224-5p exhibited lower expression in uveal melanoma
(40). However, studies on the
expression of miR-224-5p in PCa have been limited. Only a small
number of studies with small sample sizes have found that
miR-224-5p was expressed at a low level in prostatic cancer
(26–28), and this was not confirmed by a
second study involving a larger sample size. Therefore, these
findings may, to a certain extent, contain errors. The present
study, using data from TCGA, GEO and ArrayExpress databases, and
previous literature, investigated the expression of miR-224-5p in
PCa from PCa tissues, biofluids and cell lines. Taking advantage of
the public data, it was confirmed that the expression of miR-224-5p
was low in PCa. The results of a study by Fu et al indicated
that downregulated miR-224-5p was closely associated with terminal
clinical staging and metastasis (27). The present study, based on analysis
of TCGA data, revealed that the expression of pre-miR-224 was
decreased only in the T stage and M stage of PCa, although
additional investigations are required to confirm its expression in
other stages. Furthermore, when investigating the association
between miR-224-5p and prognosis, Lin et al found that the
low expression of miR-224-5p was associated with a poorer RFS rate
(41). Wan et al examined
the recurrence rate of patients with PCa following surgery, and
found that downregulated miR-224-5p was closely associated with RFS
rate (42). Mavridis et al
investigated the survival rate of 58 patients with PCa, and
confirmed that the lower the expression of miR-224-5p, the poorer
the RFS rate of the patients (26).
Fu et al examined 20 patients of PCa and demonstrated that
those patients with upregulated miR-224-5p tended to have improved
OS rates (27). In the present
study, data on the survival rate of 500 patients with PCa was
acquired from TCGA. No significant association was found between
the expression of pre-miR224 and the OS rates of patients, with
additional follow-up investigations required to confirm this.
Bioinformatics analysis was used in the present
study to examine the molecular mechanism of miR-224-5p in PCa. KEGG
analysis indicated that potential target mRNAs were significantly
enriched in the amino sugar and nucleotide sugar metabolism
signaling pathway. Previous studies have demonstrated that this
pathway may be involved in carbohydrate metabolism in tumor
tissues, affecting the onset and development of cancer (43,44).
Genes enriched in this pathway included UAP1, HK2 and CHIT1, all of
which exhibited notably high expression in PCa tissues. It was
demonstrated that higher expression of UAP1 in PCa tissues
accelerated the growth of cancer cells (45). In addition, a high expression of HK2
in PCa is essential in cell proliferation, apoptosis and
carbohydrate metabolism (46–48).
Although experiments have shown that the differentially expressed
CHIT1 is correlated with colon carcinoma (49), its association with PCa has not been
elucidated. Additionally, analysis of three key genes in the PPI
network, TOP2A, ACLY and RRM2, revealed that the expression levels
of these three genes were markedly elevated. A study by
Schaeferklein et al in 2015 showed that the increased
expression of TOP2A in PCa stimulated the growth and proliferation
of PCa cells via androgen receptor (50). Similarly, a study by Shah et
al in 2016 further elucidated the correlations of ACLY with
androgen receptor gene expression, and the proliferation and
apoptosis of PCa cells. Xin et al found that ACLY inhibited
the generation of fat in PCa tissues via a targeting association,
which may suppress the growth and metastasis of PCA cells (51,52).
RRM2 has been shown to convert ribonucleotide into deoxynucleotide
and be involved in multiple biological processes, including the
synthesis of DNA and cell growth. In vitro experiments have
demonstrated that upregulated RRM2 promotes the proliferation and
metastasis of tumor cells in PCa (53,54).
Several studies have examined miR-224-5p targeting mRNA. Wan et
al confirmed that miR-224-5p targeted and upregulated APLN, and
these were involved in the onset and progression of PCa (42). Fu et al identified CAMKK2 as
the target gene of miR-224-5p, and noted the negative axial
regulatory associations between them; these 2 genes were also found
to collaborate to promote the progression of PCa (27). According to the results of the
present study, the target genes of miR-224-5p in PCa may include
UAP1, HK2, CHIT1, TOP2A, ACLY and RRM2. miR-224-5p is likely to
regulate the expression of these genes, therefore performing its
functions in the initiation and development of PCa. However, these
results based on theoretical analysis require additional in
vivo and in vitro experiments for confirmation.
In conclusion, the present study, based on data from
TCGA, GEO and ArrayExpress databases, and previous literature,
examined the expression of miR-224-5p in PCa and its clinical
significance. It was confirmed that the expression of miR-224-5p
was low in PCa and was associated with its clinical progression. In
addition, miR-224-5p may exert its effects in PCa by mediating
UAP1, HK2, CHIT1, TOP2A, ACLY and RRM2. The present study provides
a theoretical foundation for future investigations on the
oncogenesis of PCa.
Acknowledgements
The authors would like to thank all members of the
Molecular Oncology Group of the First Affiliated Hospital of
Guangxi Medical University (Nanning, Guangxi Zhuang Autonomous
Region 530021, China) for their professional suggestions. FCM, JM
and JCZ reviewed and edited the manuscript.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
BLG and XHH and GC conceived and designed the study.
BLG, LJZ, LG and RQH performed the experiments. BLG and GC wrote
the paper. BLG, LJZ, LG, RQH, GC and XHH reviewed and edited the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
This study does not involve animal or human related
experiments.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
An H, Tao N, Li J, Guan Y, Wang W, Wang Y
and Wang F: detection of prostate cancer metastasis by whole body
magnetic resonance imaging combined with bone scintigraphy and PSA
Levels. Cell Physiol Biochem. 40:1052–1062. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chen SL, Wang SC, Ho CJ, Kao YL, Hsieh TY,
Chen WJ, Wu PR, Ko JL, Lee H and Sung WW: Prostate cancer
mortality-to-incidence ratios are associated with cancer care
disparities in 35 countries. Sci Rep. 7:4000032017.
|
|
4
|
Pan XW, Gan SS, Ye JQ, Fan YH, Hong Υ, Chu
CM, Gao Y, Li L, Liu X, Chen L, et al: SMC1A promotes growth and
migration of prostate cancer in vitro and in vivo. Int J Oncol.
49:1963–1972. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ost P, Bossi A, Decaestecker K, De
Meerleer G, Giannarini G, Karnes RJ, Roach M III and Briganti A:
Metastasis-directed therapy of regional and distant recurrences
after curative treatment of prostate cancer: A systematic review of
the literature. Eur Urol. 67:852–863. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pettersson A, Robinson D, Garmo H,
Holmberg L and Stattin P: Age at diagnosis and prostate cancer
treatment and prognosis: A population-based cohort study. Ann
Oncol. 29:377–385. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jemal A, Fedewa SA, Ma J, Siegel R, Lin
CC, Brawley O and Ward EM: prostate cancer incidence and PSA
testing patterns in relation to USPSTF screening recommendations.
JAMA. 314:2054–2061. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu J, Chen Z, Wang T, Liu L, Zhao L, Guo
G and Wang D: influence of four radiotracers in PET/CT on
diagnostic accuracy for prostate cancer: A Bivariate Random-Effects
Meta-Analysis. Cell Physiol Biochem. 39:467–480. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Barrington WE, Schenk JM, Etzioni R,
Arnold KB, Neuhouser ML, Thompson IM Jr, Lucia MS and Kristal AR:
Difference in association of obesity with prostate cancer risk
between US African American and Non-Hispanic White Men in the
Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA
Oncol. 1:342–349. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vidal AC, Howard LE, Sun SX, Cooperberg
MR, Kane CJ, Aronson WJ, Terris MK, Amling CL and Freedland SJ:
Obesity and prostate cancer-specific mortality after radical
prostatectomy: Results from the Shared Equal Access Regional Cancer
Hospital (SEARCH) database. Prostate Cancer Prostatic Dis.
20:72–78. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gambari R, Brognara E, Spandidos DA and
Fabbri E: Targeting oncomiRNAs and mimicking tumor suppressor
miRNAs: Νew trends in the development of miRNA therapeutic
strategies in oncology (Review). Int J Oncol. 49:1–32. 2016.
View Article : Google Scholar
|
|
12
|
Gao Y, Feng B, Han S, Lu L, Chen Y, Chu X,
Wang R and Chen L: MicroRNA-129 in human cancers: From
tumorigenesis to clinical treatment. Cell Physiol Biochem.
39:2186–2202. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Paul P, Chakraborty A, Sarkar D, Langthasa
M, Rahman M, Bari M, Singha RS, Malakar AK and Chakraborty S:
Interplay between miRNAs and Human Diseases: A Review. J Cell
Physiol. 233:2007–2018. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xie T, Huang M, Wang Y, Wang L, Chen C and
Chu X: MicroRNAs as regulators, biomarkers and therapeutic targets
in the drug resistance of colorectal cancer. Cell Physiol Biochem.
40:62–76. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang X, Tang W, Li R, He R, Gan T, Luo Y,
Chen G and Rong M: Downregulation of microRNA-132 indicates
progression in hepatocellular carcinoma. Exp Ther Med.
12:2095–2101. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Huang WT, Wang HL, Yang H, Ren FH, Luo YH,
Huang CQ, Liang YY, Liang HW, Chen G and Dang YW: Lower expressed
miR-198 and its potential targets in hepatocellular carcinoma: A
clinicopathological and in silico study. OncoTargets Ther.
9:5163–5180. 2016. View Article : Google Scholar
|
|
17
|
Pang C, Liu M, Fang W, Guo J, Zhang Z, Wu
P, Zhang Y and Wang J: MiR-139-5p is increased in the peripheral
blood of patients with prostate cancer. Cell Physiol Biochem.
39:1111–1117. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
van Beijnum JR, Giovannetti E, Poel D,
Nowak-Sliwinska P and Griffioen AW: miRNAs: Micro-managers of
anticancer combination therapies. Angiogenesis. 20:269–285. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu C, Zhuang Y, Jiang S, Liu S, Zhou J, Wu
J, Teng Y, Xia B, Wang R and Zou X: Interaction between
Wnt/β-catenin pathway and microRNAs regulates
epithelial-mesenchymal transition in gastric cancer (Review). Int J
Oncol. 48:2236–2246. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang X, Pang YY, He RQ, Lin P, Cen JM,
Yang H, Ma J and Chen G: Diagnostic value of strand-specific
miRNA-101-3p and miRNA-101-5p for hepatocellular carcinoma and a
bioinformatic analysis of their possible mechanism of action. FEBS
Open Bio. 8:64–84. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Feng Q, Huang X and Urology DO: Expression
of miRNA-32, −196a,-218,-128 and let7i in serum of patients with
prostate cancer and its clinical significance. Guangdong Yixue.
2016.
|
|
22
|
Pan Y, Gan Q, Jun LU, Huang T and Mang KE:
Serological determination and its clinical significance of
miRNA-129 and miRNA-21 in prostate cancer patients. Chinese Journal
of Health Laboratory Technology. 2016.
|
|
23
|
Yao CH, Yuan XC, Liu C and Deng JP:
Expression and diagnostic value of miRNA-15a in prostate cancer.
Chin J Immunol. 2016.
|
|
24
|
Lu S, Wang MS, Chen PJ, Ren Q and Bai P:
miRNA-186 inhibits prostate cancer cell proliferation and tumor
growth by targeting YY1 and CDK6. Exp Ther Med. 13:3309–3314. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tian XM, Luo YZ, He P, Li J, Ma ZW and An
Y: Inhibition of invasion and migration of prostate cancer cells by
miRNA-509-5p via targeting MDM2. Genet Mol Res. 16:doi:
10.4238/gmr16019195.
|
|
26
|
Mavridis K, Stravodimos K and Scorilas A:
Downregulation and prognostic performance of microRNA 224
expression in prostate cancer. Clin Chem. 59:261–269. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fu H, He HC, Han ZD, Wan YP, Luo HW, Huang
YQ, Cai C, Liang YX, Dai QS, Jiang FN, et al: MicroRNA-224 and its
target CAMKK2 synergistically influence tumor progression and
patient prognosis in prostate cancer. Tumour Biol. 36:1983–1991.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kristensen H, Haldrup C, Strand S,
Mundbjerg K, Mortensen MM, Thorsen K, Ostenfeld MS, Wild PJ, Arsov
C, Goering W, et al: Hypermethylation of the GABRE~miR-452~miR-224
promoter in prostate cancer predicts biochemical recurrence after
radical prostatectomy. Clin Cancer Res. 20:2169–2181. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen M, Wang J, Luo Y, Huang K, Shi X, Liu
Y, Li J, Lai Z, Xue S, Gao H, et al: Identify Down syndrome
transcriptome associations using integrative analysis of microarray
database and correlation-interaction network. Hum Genomics.
12:22018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
He X, Zhang C, Shi C and Lu Q:
Meta-analysis of mRNA expression profiles to identify
differentially expressed genes in lung adenocarcinoma tissue from
smokers and non-smokers. Oncol Rep. 39:929–938. 2018.PubMed/NCBI
|
|
32
|
Li F, Shi W, Wan Y, Wang Q, Feng W, Yan X,
Wang J, Chai L, Zhang Q and Li M: Prediction of target genes for
miR-140-5p in pulmonary arterial hypertension using bioinformatics
methods. FEBS Open Bio. 7:1880–1890. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liang L, Zeng JH, Wang JY, He RQ, Ma J,
Chen G, Cai XY and Hu XH: Down-regulation of miR-26a-5p in
hepatocellular carcinoma: A qRT-PCR and bioinformatics study.
Pathol Res Pract. 213:1494–1509. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mi B, Liu G, Zhou W, Lv H, Liu Y and Liu
J: Identification of genes and pathways in the synovia of women
with osteoarthritis by bioinformatics analysis. Mol Med Rep.
17:4467–4473. 2018.PubMed/NCBI
|
|
35
|
Wang L, Peng Z, Wang K, Qi Y, Yang Y,
Zhang Y, An X, Luo S and Zheng J: NDUFA4L2 is associated with clear
cell renal cell carcinoma malignancy and is regulated by ELK1.
PeerJ. 5:e40652017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wei D: A multigene support vector machine
predictor for metastasis of cutaneous melanoma. Mol Med Rep.
17:2907–2914. 2018.PubMed/NCBI
|
|
37
|
Zhang H, Liu J, Fu X and Yang A:
Identification of key genes and pathways in tongue squamous cell
carcinoma using bioinformatics analysis. Med Sci Monit.
23:5924–5932. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang L, Huang LS, Chen G and Feng ZB:
Potential targets and clinical value of miR-224-5p in cancers of
the digestive tract. Cell Physiol Biochem. 44:682–700. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhou F, Li S, Meng HM, Qi LQ and Gu L:
MicroRNA and histopathological characterization of pure mucinous
breast carcinoma. Cancer Biol Med. 10:22–27. 2013.PubMed/NCBI
|
|
40
|
Zheng X, Tang H, Zhao X, Sun Y, Jiang Y
and Liu Y: Long non-coding RNA FTH1P3 facilitates uveal melanoma
cell growth and invasion through miR-224-5p. PLoS One.
12:e01847462017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lin ZY, Huang YQ, Zhang YQ, Han ZD, He HC,
Ling XH, Fu X, Dai QS, Cai C, Chen JH, et al: MicroRNA-224 inhibits
progression of human prostate cancer by downregulating TRIB1. Int J
Cancer. 135:541–550. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wan Y, Zeng ZC, Xi M, Wan S, Hua W, Liu
YL, Zhou YL, Luo HW, Jiang FN and Zhong WD: Dysregulated
microRNA-224/apelin axis associated with aggressive progression and
poor prognosis in patients with prostate cancer. Hum Pathol.
46:295–303. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cao J, Lu XX, Li Y, Zhu LQ, Yang C, Ou C
and Tang YP: Applying gene set enrichment analysis and
meta-analysis to screen key genes controlling the development and
progression of hepatic carcinoma. World Chin J Digestology.
20:7542012.(In Chinese). View Article : Google Scholar
|
|
44
|
Li Z, Li BQ, Jiang M, Chen L, Zhang J, Liu
L and Huang T: Prediction and analysis of retinoblastoma related
genes through gene ontology and KEGG. Biomed Res Int.
2013:3040292013.PubMed/NCBI
|
|
45
|
Itkonen HM, Engedal N, Babaie E, Luhr M,
Guldvik IJ, Minner S, Hohloch J, Tsourlakis MC, Schlomm T and Mills
IG: UAP1 is overexpressed in prostate cancer and is protective
against inhibitors of N-linked glycosylation. Oncogene.
34:3744–3750. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Kudryavtseva AV, Nyushko KM, Zaretsky AR,
et al: Expression of HK2 gene is deregulated in Prostate cancer.
Asian J Pharm. 11:S158–S161. 2017.
|
|
47
|
Tao T, Chen M, Jiang R, Guan H, Huang Y,
Su H, Hu Q, Han X and Xiao J: Involvement of EZH2 in aerobic
glycolysis of prostate cancer through miR-181b/HK2 axis. Oncol Rep.
37:1430–1436. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tao T, Xiang P and Huang T: Expression of
HK2 in prostate cancer and its effect on malignant phenotype of
prostate cancer cells. Chinese Journal of Clinical &
Experimental Pathology. 2017.
|
|
49
|
Li FF, Yan P, Zhao ZX, Liu Z, Song DW,
Zhao XW, Wang XS, Wang GY and Liu SL: Polymorphisms in the CHIT1
gene: Associations with colorectal cancer. Oncotarget.
7:39572–39581. 2016.PubMed/NCBI
|
|
50
|
Schaefer-Klein JL, Murphy SJ, Johnson SH,
Vasmatzis G and Kovtun IV: Topoisomerase 2 alpha cooperates with
androgen receptor to contribute to prostate cancer progression.
PLoS One. 10:e01423272015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Shah S, Carriveau WJ, Li J, Campbell SL,
Kopinski PK, Lim HW, Daurio N, Trefely S, Won KJ, Wallace DC, et
al: Targeting ACLY sensitizes castration-resistant prostate cancer
cells to AR antagonism by impinging on an ACLY-AMPK-AR feedback
mechanism. Oncotarget. 7:43713–43730. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xin M, Qiao Z, Li J, Liu J, Song S, Zhao
X, Miao P, Tang T, Wang L, Liu W, et al: miR-22 inhibits tumor
growth and metastasis by targeting ATP citrate lyase: Evidence in
osteosarcoma, prostate cancer, cervical cancer and lung cancer.
Oncotarget. 7:44252–44265. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Afrasiabi A, Fiuji F, Mirhafez R and Avan
A: RRM2 (ribonucleotide reductase M2). Atlas Genet Cytogenet Oncol
Haematol. 19:p32–37. 2015.
|
|
54
|
Huang Y, Liu X, Wang YH, Yeh SD, Chen CL,
Nelson RA, Chu P, Wilson T and Yen Y: The prognostic value of
ribonucleotide reductase small subunit M2 in predicting recurrence
for prostate cancers. Urol Oncol. 32:51.e9–51.e19. 2014. View Article : Google Scholar
|