Introduction
Cervical cancer is one of the most commonly
diagnosed malignancies in the world (1). The mortality rate associated with
cervical cancer is the highest among all female reproductive tract
malignant tumors, and cervical cancer seriously threatens the
health of women worldwide (2,3).
Although improvements in preventative measures and expansion of
cervical cancer screening have significantly diminished the
incidence and mortality rate in the developed world, there is a
lack of cervical cancer screening in developing countries (4). Cervical cancer has a poor prognosis
due to local invasion and lymphatic metastasis (5). Thus, it is necessary to investigate
the biological mechanisms driving cervical cancer to provide novel
insight into the prevention and therapeutic management of this
disease.
The expression and dysfunction of homeobox (HOX)
genes play an important role in the development of various
malignant tumors, such as lung cancer, breast cancer, colon cancer,
prostate cancer and leukemia (6–9). HOXA5
is a member of HOX gene family and is a transcriptional regulator
that regulates cell growth, differentiation and apoptosis (10,11).
As the HOX genes have become a ‘hot topic’ in cancer research, more
researchers have paid close attention concerning the relationship
between HOXA5 and multiple malignancies in recent years. It has
been found that lack of HOXA5 function and other genetic damage can
lead to breast cancer development (12). It has been reported that HOXA5
induces cell apoptosis and decreases cell drug resistance in breast
cancer, lung cancer and glioblastoma (13,14).
Moreover, a reduction in HOXA5 expression was recognized to be
closely related to cell proliferation and invasion in tumors such
as non-small cell lung cancer and esophageal squamous (15,16).
However, the role of HOXA5 in cervical carcinoma development still
remains unclear. In the present study, we aimed to investigate and
acquire insight into the molecular mechanisms of HOXA5 in cervical
cancer to improve treatment of the disease.
Protein kinase B (AKT) is a homologue of v-AKT, a
proto-oncogene that exists in human chromosomes (17). A variety of hormones, growth factors
and cytokines can stimulate Akt activation, thereby regulating cell
growth, proliferation, motility, invasion, apoptosis and other
processes (18). Studies have
indicated that phosphorylation of AKT is increased in prostate,
breast and cervical cancer cells (19–21).
AKT was found to induce cell proliferation and survival in β cells,
endothelial cells, cardiomyocytes and tumor cells (21,22).
It has been reported that AKT activation can be constrained by
HOXA5 in mouse white adipocytes (13). Nevertheless, the effect of HOXA5 on
AKT phosphorylation in cervical cancer cells remains unknown. In
our study, we explored the correlation between HOXA5 and AKT
phosphorylation in cervical cancer. p27 is an important member of
the p21 family of cyclin-dependent kinase inhibitors and is a
broad-spectrum cyclin-dependent kinase (CDK) inhibitor that blocks
the cell cycle through the transition point into the S phase, and
then inhibits cell growth and proliferation, and is widely
recognized as a tumor-suppressor gene in various types of cancer,
including cervical cancer (23–25).
Moreover, p27 expression could be regulated via AKT activation
(26) and thus we hypothesized that
HOXA5 regulates cell proliferation and apoptosis via AKT/p27 in
cervical cancer.
We aimed to probe into the molecular mechanisms of
HOXA5 in cervical cancer by assessing the expression of HOXA5 in
cervical cancer cells. We investigated the cell proliferation and
apoptosis in cervical cancer cell lines after overexpressing HOXA5.
We also investigated the molecular mechanism of cell proliferation
and apoptosis following the overexpression of HOXA5 in cervical
cancer.
Materials and methods
Cell culture and tissues
The cervical cancer cell lines ME-180 (cat. no.
HTB-33™), HT-3 (cat. no. HTB-32™), HeLa (cat. no. CCL-2™) and SiHa
(cat. no. HTB-35™) (ATCC; Manassas, VA, USA) were grown in
Dulbecco's modified Eagle's medium (HyClone; GE Healthcare Life
Sciences, Logan, UT, USA) supplemented with 10% fetal bovine serum
(FBS; HyClone; GE Healthcare Life Sciences), 100 U/ml of
penicillin, and 100 µg/ml of streptomycin. Human cervical
epithelial cells (HCerEpiC; ScienCell Research Laboratories,
Carlsbad, CA, USA; cat. no. CP7060) (27), were cultured in cervical epithelial
cell medium (ScienCell Research Laboratories). Cells were cultured
in an atmosphere of 5% CO2 at 37°C.
Cervical tumor samples (n=10, 40–50 years of age)
were collected in the Fourth People's Hospital of Shaanxi from
women with the average year of 42 years undergoing hysterectomies
without having been treated with radiotherapy or chemotherapy. Two
patients had a diagnosis of squamous cell carcinoma (SCC), 6
presented with adenomatous carcinoma (ADC) and 2 patients had an
intermediate diagnosis of adenosquamous cell carcinoma (ASC).
Normal tissues (n=10, 40–50 years of age) were collected in the
Fourth People's Hospital of Shaanxi from women with the average
year of 45 years old undergoing surgery for myoma or adenomyoma.
Normal tissue samples were non-malignant and negative for human
papilloma virus and ThinPrep cytologic tests. The collections were
performed between August 2017 and September 2017 and were approved
by the Ethics Committee of The Fourth People's Hospital of Shaanxi.
Informed consent was obtained from all of the patients.
RT-qPCR assay
Total RNA of cells was extracted by using TRizol
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
reverse-transcribed into cDNA using the PrimeScript™ First Strand
cDNA Synthesis kit (Takara Biotechnology Co., Ltd., Dalian, China).
The volume of reverse-transcription polymerase chain reaction
(RT-qPCR) was 20 µl and it contained 10 µl SYBR Premix Ex Taq II
(Takara Biotechnology Co., Ltd.). The primers were as follows:
HOXA5, sense primer: 5′-AGCCACAAATCAAGGACACA-3′ and antisense
primer: 5′-GCTCGCTCACGGAACTATG-3′; GAPDH, sense primer:
5′-CGAAGGTGAAGGTCGGAGT-3′ and antisense primer:
5′-GAAGATGGTGATGGGATTTC-3′. The cycling parameters consisted of
94°C for 30 sec; 40 cycles of 94°C for 40 sec, 60°C for 30 sec and
72°C for 30 sec and 72°C for 10 min. The relative levels of gene
expression were quantified using the 2−ΔΔCt method
(28). GAPDH was the reference
gene. The experiment was repeated three times.
Western blot analysis
Cells were treated with lysate and protein was
extracted. Then, the protein was quantified with a BCA kit
(Beyotime Institute of Biotechnology, Nantong, China). After
loading 25 µg of the protein onto gel for 12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), the proteins
were separated and transferred onto polyvinylidene fluoride (PVDF)
membranes (Bio-Rad Laboratories, Hercules, CA, USA) by
electrophoretic transfer. The PVDF membranes were then incubated
with 5% skim milk for 1.5 h, followed by incubation overnight at
4°C with the primary antibodies (all from Abcam Inc., Cambridge,
MA, USA): anti-HOXA5 (dilution 1:800; cat. no. ab82645) anti-AKT
(dilution 1:500; cat. no. ab8805), anti-Akt (phospho S473)
(dilution 1:600; cat. no. ab81283), anti-p27 (dilution 1:1,000;
cat. no. ab32034) and anti-GAPDH (dilution 1:500; cat. no. ab9485).
Next, the polyvinylidene fluoride was incubated with a secondary
antibody (dilution 1:800; cat. no. ab7090) diluted in the blocking
buffer. Finally, the protein was detected using enhanced
chemiluminescence. The experiment was repeated three times.
Construction of the recombinant
plasmids
The full-length HOXA5 gene (GenBank™ accession
number NM_019102.3) was amplified using RT-PCR, followed by
insertion into the plasmid pcDNA.3.1 (Clontech Inc., Palo Alto, CA,
USA). The restriction enzymes used for the cDNA and plasmid
pcDNA.3.1 were EcoRI and BamHI. The recombinant
plasmid was then transducted into Escherichia coli DH5α
cells (Tiangen, Beijing, China), followed by amplification at 37°C.
Afterwards, the recombinant plasmids were extracted using a plasmid
DNA extraction kit (Takara Biotechnology Co., Ltd.). The
recombinant plasmid containing the correct sequence was named
pcDNA.3.1-HOXA5.
Cell transfection
The ME-180 and HT-3 cell lines were separately
cultured in 96-well plates and incubated in an incubator (Thermo
Fisher Scientific, Inc.) with 5% CO2 for 24 h. The cells
were transfected with pcDNA.3.1-HOXA5 (0.2 µg), pcDNA.3.1 (0.2 µg)
or pcDNA.3.1-p27 (0.2 µg) using Turbofect (0.5 µl; Thermo Fisher
Scientific, Inc.) and incubated at 37°C in 5% CO2 for
24, 36, 48 or 72 h. Transfection efficiency was measured by RT-qPCR
and western blot assays.
Flow cytometric analysis
Cell apoptosis was examined using an Annexin
V-fluorescein isothiocyanate (FITC) and propidium iodide (PI) kit
(Jiancheng Bioengineering Institute, Nanjing, China) in accordance
with the supplier's instructions. Briefly, cells were harvested and
centrifuged at 200 × g for 5 min. The cells (5×105
cells/ml) were resuspended in the binding buffer and mixed with 5
µl of Annexin V-FITC in 195 µl of the cell suspension for 30 min at
4°C. Afterwards, cells were incubated with PI (6 µl) for 8 min. The
apoptotic cells were quantified by FACS analyzer (Beckman Coulter,
Brea, CA, USA). The experiment was repeated three times.
Caspase-3 activity assay
Caspase-3 activity in the cells was measured in line
with the explanatory memorandum of a caspase-3 activity assay kit
(Beyotime Institute of Biotechnology). In brief, cells
(1×106) were centrifugated, washed trice with PBS and
incubated in 500 µl lysis buffer on ice for 15 min. The activity of
caspase-3 was tested by a 200 mmol/l AcDEVD-MCA fluorogenic
substrate in the assay buffer, and fluorescence intensity was
quantified with spectrofluorometry. The experiment was repeated
three times.
Cell viability
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT) assay was used to quantify cell viability according
to the specifications. Firstly, cells were incubated in a 96-well
plate in humid atmosphere of 5% CO2 at 37°C for 24 h
followed by transfection. MTT (5 g/l) diluted in phosphate-buffered
saline (PBS) was added and incubated at 37°C for 6 h. Subsequently,
dimethyl sulfoxide (DMSO) (160 µl/well) was added to dissolve the
formazan, and the absorbance (OD) values were read using an
microplate reader at 490 nm (Thermo Fisher Scientific, Inc.). The
experiment was repeated three times.
Bromodeoxyuridine (BrdU) assay
Cell proliferation was tested using a BrdU kit
(Roche Applied Science, Mannheim, Germany) on the basis of the
instructions. Cells were plated in 96-well plates and transfections
were performed. Cells were incubated with BrdU solution (10
µl/well) for 1.5 h and then denaturing solution (100 µl/well) for
25 min. Cells were then stained with anti-BrdU antibody for 1.5 h
at room temperature followed by staining with secondary antibody
solution. Next, 100 µl of tetramethyl benzidine substrate was added
and allowed to incubate for 30 min. Finally, the results were
detected at 450 nm using a SpectroFluor Plus multiwell plate reader
(Tecan Group, Ltd., Mannedorf, Switzerland). The experiment was
repeated three times.
Transwell invasion assays
Bio-Coat cell migration chambers (BD Biosciences,
San Jose, CA, USA) were used to assess cervical cancer cell
invasion. Chambers were coated with Matrigel (Becton Dickinson; BD
Biosciences), and the transfected cells were suspended using 350 µl
serum-free medium (1×105 cells) and added to the top
chamber. Then, the cells were cultured with 5% CO2 at
37°C for 48 h followed by the addition of culture medium (500 µl)
with 10% FBS to the lower chamber. Non-invading cells were gently
removed using a cotton swab, and invasive cells were fixed with 95%
ethanol and stained with trypan blue. The invasive cells were
quantified as the mean count of stained cells in six random fields
under bright field microscopy (×400 magnification) using an Olympus
IX70 inverted microscope (Olympus Corp., Tokyo, Japan). The
experiment was repeated three times.
Statistical analysis
Statistical significance was determined by the
Student's t-test between two groups and by one-way ANOVA followed
by a Bonferroni test for multiple groups. A P-value of <0.05 was
considered to indicate a statistically significant result. Data are
expressed as the mean ± standard deviation (SD). Statistical
analyses were processed with SPSS version 22.0 software (IBM Corp.,
Armonk, NY, USA).
Results
Downregulation of the expression of
HOXA5 in cervical cancer
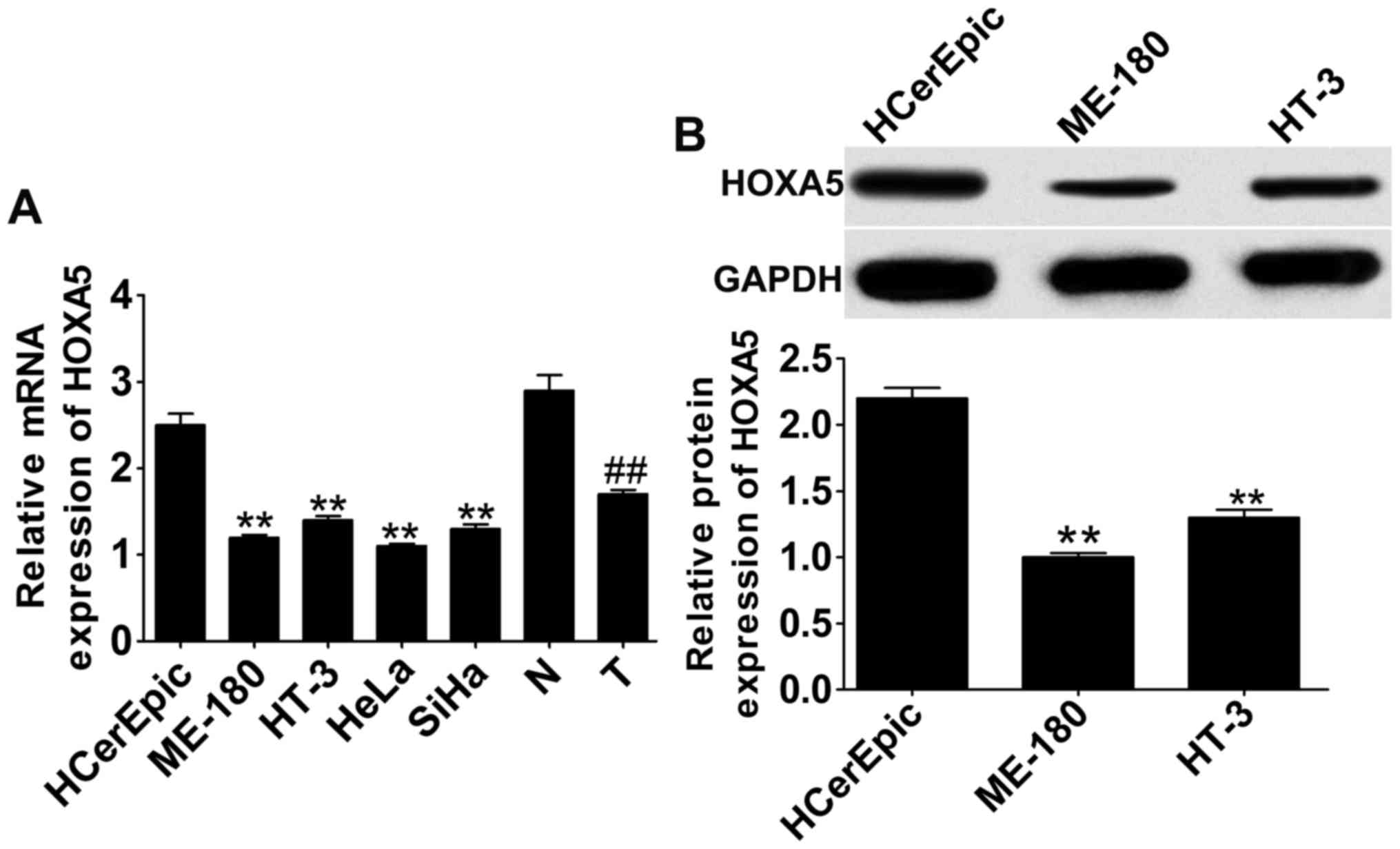
We quantified the expression of HOXA5 in cervical
cancer cell lines and tissues to explore the expression of HOXA5 in
cervical cancer. The results showed that mRNA expression (Fig. 1A) of HOXA5 as determined by RT-qPCR
in cervical cancer cell lines and tissues was significantly
decreased in both when compared with human cervical epithelial
HCerEpic cells and normal tissues. The mRNA reduction of HOXA5 in
ME-180, HT-3, HeLa and SiHa cells exhibited no significant
differences between each cell line; thus we detected the protein
expression of HOXA5 in ME-180 and HT-3 cells. The protein
expression (Fig. 1B) of HOXA5 as
determined by western blot analysis in ME-180 and HT-3 cells was
also significantly decreased when compared with the HCerEPic cell
line.
HOXA5 overexpression induces cell
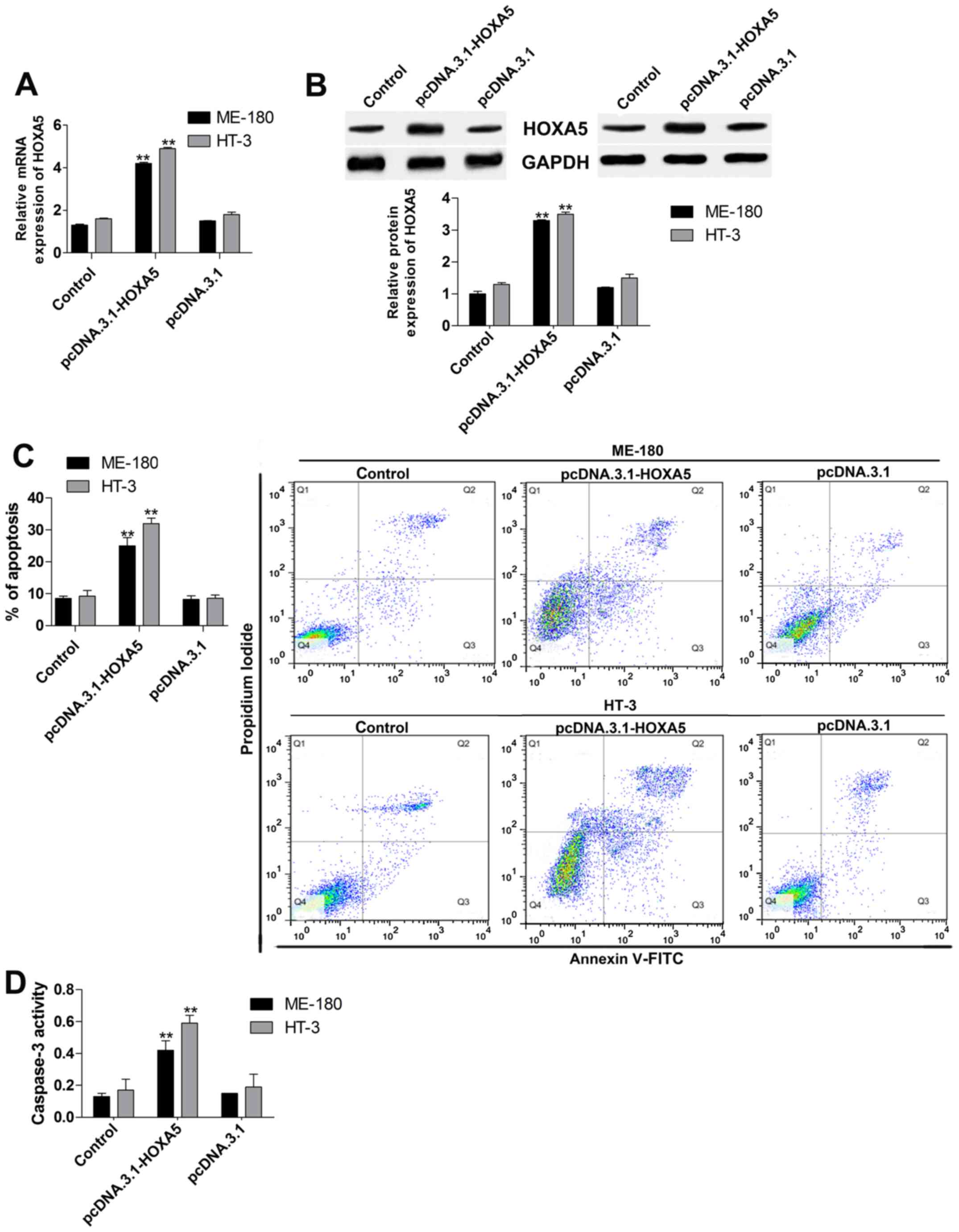
apoptosis in cervical cancer cell lines
To investigate the potential role of HOXA5 in
cervical cancer, we measured cell apoptosis by using Annexin
V-FITC/PI assay and caspase-3 activity testing after HOXA5
overexpression in ME-180 and HT-3 cells. Firstly, we overexpressed
HOXA5 by cells transfection with pcDNA.3.1-HOXA5, and the data
indicated that mRNA (Fig. 2A) and
protein (Fig. 2B) expression of
HOXA5 were both increased twice. Thus, the gain-of-function
experiment of HOXA5 was successful. Subsequently, the effect of
HOXA5 upregulation on cell apoptosis in ME-180 and HT-3 cells were
tested. In the Annexin V-FITC/PI assay (Fig. 2C), cell apoptosis was significantly
induced by upregulation of HOXA5. Moreover, caspase-3 activity
(Fig. 2D) was also significantly
facilitated after pcDNA.3.1-HOXA5 transfection.
HOXA5 overexpression suppresses cell
proliferation and invasion in cervical cancer cell lines
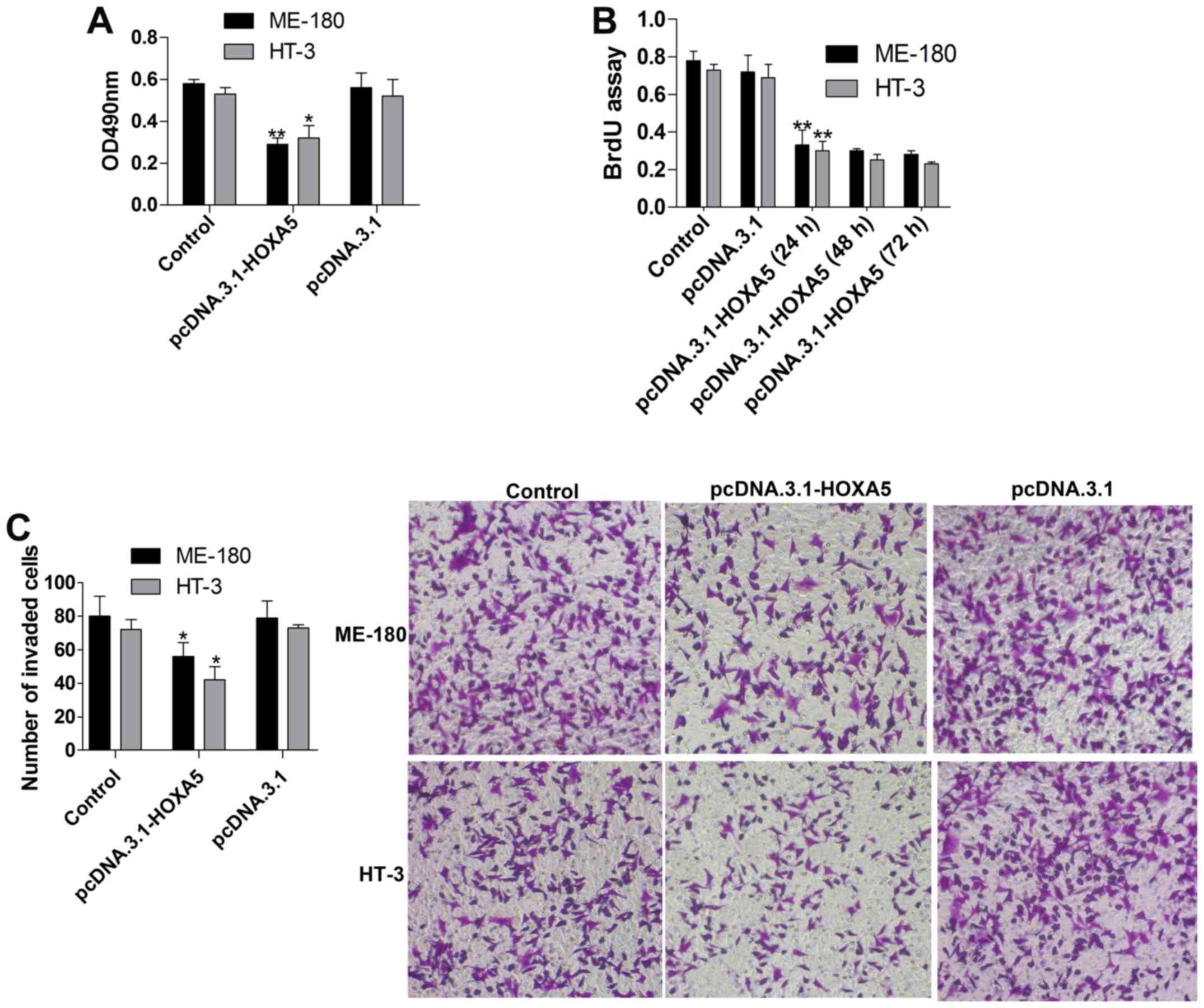
To further estimate the influence of HOXA5
overexpression on cervical cancer, we tested cell viability and
proliferation using MTT and BrdU assays, respectively. The results
revealed that cell viability (Fig.
3A) was markedly suppressed in the pcDNA.3.1-HOXA5 group when
compared to that noted in the pcDNA.3.1group. We also assessed the
proliferation (Fig. 3B) of cells
transfected with pcDNA.3.1-HOXA5 for 24, 48 and 72 h, and it was
showed that the proliferation in the pcDNA.3.1-HOXA5 (24 h) group
was significantly inhibited in contrast to the pcDNA.3.1 group, and
the suppression was slightly time-dependent. Additionally, cell
invasion (Fig. 3C) was detected by
Transwell invasion assay, and the data showed a significant
decrease in cell invasion after HOXA5 overexpression. Therefore,
HOXA5 displayed a valid inhibitory effect on cell proliferation and
invasion in cervical cancer cell lines.
Regulation of AKT activity and p27
expression by HOXA5
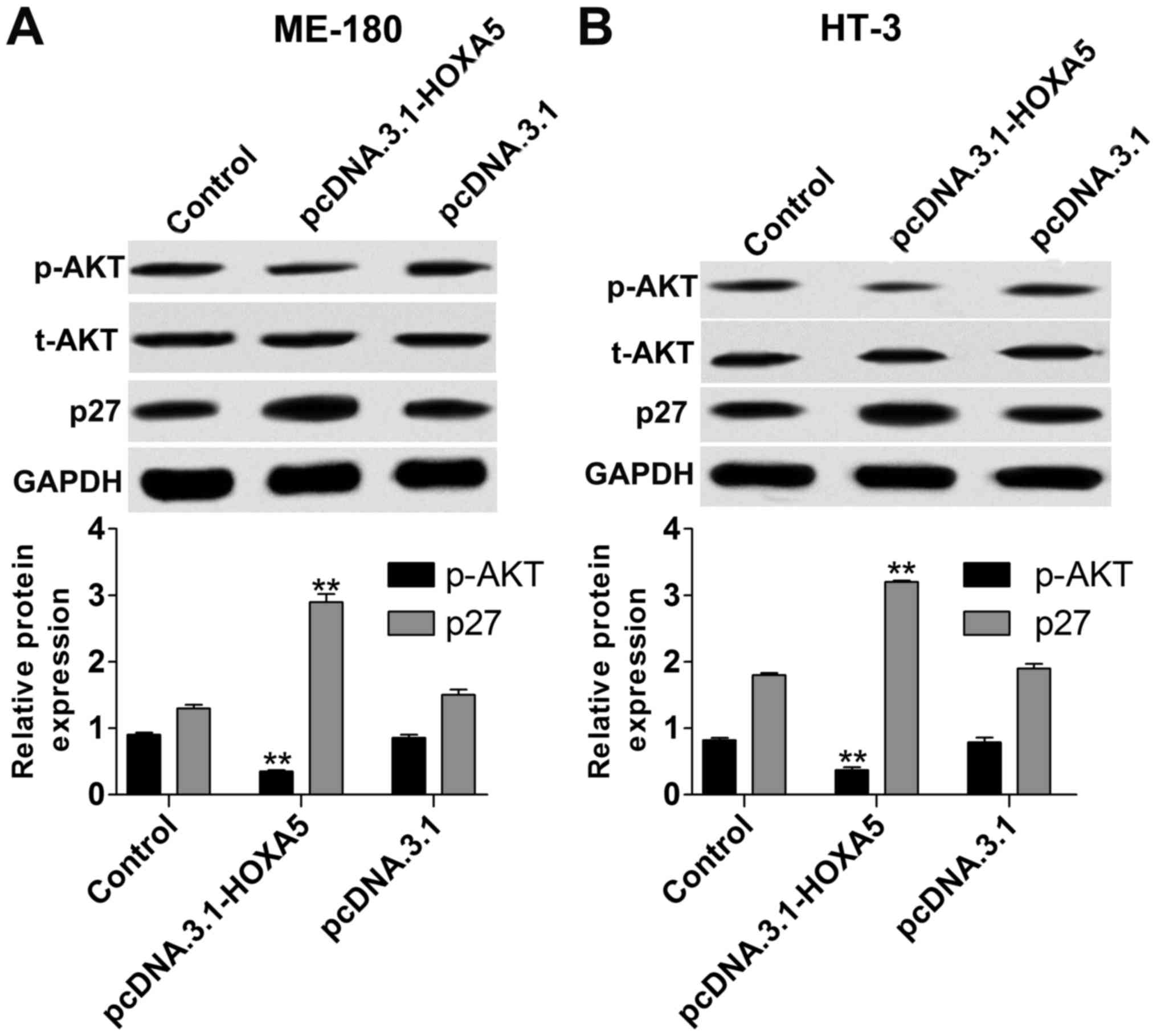
To gain insight into the molecular mechanisms of
HOXA5 in cervical cancer, we determined the protein expression of
p-AKT and p27 via western blot assay in ME-180 and HT-3 cells with
HOXA5 upregulation. The results indicated that the protein level of
p-AKT (Fig. 4) in the
pcDNA.3.1-HOXA5 group was significantly reduced in contrast to the
pcDNA.3.1 group. Meanwhile, p27 (Fig.
4) expression was increased in the pcDNA.3.1-HOXA5 group.
Moreover, we treated ME-180 and HT-3 cells with SC79 (AKT
activator, 0.1–10 µg/ml) for 1 h after HOXA5 overexpression. We
found that SC79 (1–10 µg/ml) markedly activated AKT (p-AKT
intensity increase) in both the ME-180 and HT-3 cells and the
activity on p-AKT was dose-dependent (Fig. 5A). Thus, we measured the protein
expression of p27 in ME-180 and HT-3 cells with HOXA5
overexpression and SC79 (1 µg/ml) incubation for 1 h, and the
results showed that p27 was obviously downregulated (Fig. 5B). Meanwhile, caspase-3 activity
(Fig. 5C) was inhibited and cell
viability (Fig. 5D) and invasion
(Fig. 5E) were elevated. Thus,
HOXA5 controls cervical cancer cell proliferation and apoptosis via
the regulation of p-AKT/p27.
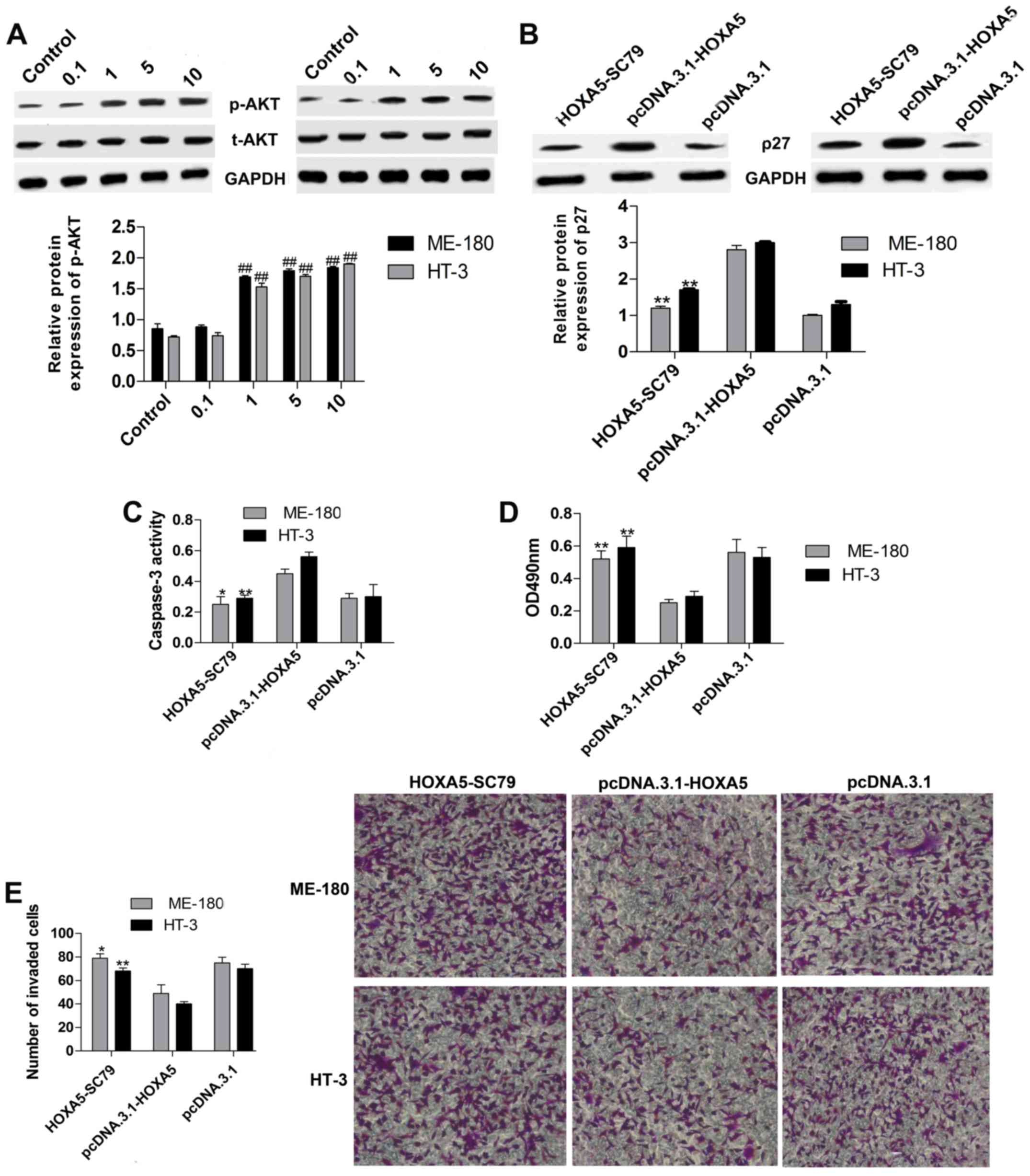 | Figure 5.HOXA5 regulates p27 expression via
controlling AKT activity. (A) The relative protein levels of p-AKT
were measured by western blot analyses in ME-180 and HT-3 cells
treated with SC79 (AKT activator, 0.1–10 µg/ml) for 1 h after HOXA5
overexpression. 0.1, 1, 5, 10: Cells treated with SC79 (0.1, 1, 5,
10 µg/ml) for 1 h after HOXA5 overexpression. n=3,
##P<0.01 vs. the control group. (B) The relative
protein levels of p27 were determined by western blot analyses in
ME-180 and HT-3 cells treated with SC79 (AKT activator, 1 µg/ml)
for 1 h after HOXA5 overexpression. (C) Caspase-3 activity was
detected by a kit in ME-180 and HT-3 cells treated with SC79 (1
µg/ml) for 1 h after HOXA5 overexpression. (D) and (E) Cell
proliferation and invasion abilities were separately measured by
the MTT and Transwell invasion assays in ME-180 and HT-3 cells
treated with SC79 (1 µg/ml) for 1 h after HOXA5 overexpression.
HOXA5-SC79: Cells treated with SC79 (1 µg/ml) for 1 h after HOXA5
overexpression. n=3, *P<0.05, **P<0.01 vs. the
pcDNA.3.1-HOXA5 group. |
Discussion
Early symptoms of cervical cancer are not obvious,
and the malignant potential in advanced stages reaches a high
degree (29). Currently, lack of
effective treatment and poor prognosis give rise to the high
mortality rate associated with cervical cancer (30). Therefore, the study of the
pathogenesis of cervical cancer and the discovery of molecular
markers for clinical diagnosis and treatment are crucial to improve
the survival of patients with cervical cancer. Studies have
demonstrated that HOXA5 plays a significant role in tumor
development. The heterotopic expression of HOXA5 was found to
promote the proliferation and differentiation of esophageal cancer
(31). Zhang et al
demonstrated that HOXA5 could constrain cell proliferation via
modulating p21 in non-small cell lung cancer (32). Wang et al demonstrated that
HOXA5 inhibition by miR-1271 facilitated non-small cell lung cancer
cell proliferation and invasion (11). Ectopic expression of HOXA5 was found
to induce apoptosis in human liposarcomas (33). Liu et al found that HOXA5
upregulation inhibits the proliferation and promotes apoptosis in
K562 leukemia cells (34). However,
the role of HOXA5 in cervical cancer is still unclear. In the
present study, we found that HOXA5 was suppressed in cervical
cancer cell lines and tissues. Overexpression of HOXA5 was achieved
by transfection with pcDNA.3.1-HOXA5, which distinctly accelerated
caspase-3 activity and cell apoptosis in cervical cancer cell
lines. Moreover, HOXA5 overexpression had inhibitory effects on
cell viability, proliferation and invasion of cervical cancer cell
lines. The role of HOXA5 in cervical cancer was found to be
consistent with that in different cancers in the literature
mentioned above.
Feng et al found that HOXA5 could restrain
AKT activity in the white adipocytes in mice (13). AKT, a proto-oncogene, has a
significant regulatory effect on cell proliferation, apoptosis and
invasion of cancer cells. Palacios et al found that AKT
activation results in chronic lymphocytic leukemia (CLL) B-cell
proliferation (35). Yang et
al demonstrated that inhibition of AKT activation by Matrine
inhibited bladder cancer cell proliferation and invasion in
vitro (36). Upregulation of
AKT phosphorylation was also found to induce cell growth and
invasion in non-small cell lung cancer (37). Targeting of the PI3K/AKT pathway by
microRNA-29b promoted the apoptosis of hepatic stellate cells in
liver fibrosis (38). Das et
al demonstrated that suppression of AKT promoted
FOXO3a-dependent apoptosis in prostate cancer (39). Moreover, Jeyamohan et al
found that inhibition of the PI3K/Akt signaling pathway could
induce cell apoptosis in cervical cancer (40). In the present study, we investigated
whether AKT plays a role in the regulation of cervical cancer
development by HOXA5. Analysis of the results showed that AKT
phosphorylation was suppressed by HOXA5 overexpression in cervical
cancer lines. Additionally, AKT activation by SC79 visibly
abolished the effect of HOXA5 overexpression on caspase-3 activity,
cell apoptosis, cell viability, proliferation and invasion in
cervical cancer cell lines. Therefore, AKT plays an important role
in the regulation of cervical cancer development by HOXA5.
p27, as a tumor suppressor, has been found to
display inhibitory effects on cervical cancer cells. miR-196a was
found to elevate cell proliferation via decreasing p27 expression
in laryngeal cancer (41). Singh
et al reported that resveratrol increases cell apoptosis via
the p27 pathway in prostate cancer cells (42). Cui et al showed that
upregulation of p27 by Snai2 inhibited the proliferation and tumor
formation of human cervical cancer cells (43). It was reported that AKT
phosphorylation could elicit p27 transcription in HeLa cells
(44). Thus, we assumed that HOXA5
could regulate p27 via controlling AKT phosphorylation in cervical
cancer cell lines. In the present study, data showed that HOXA5
overexpression increased the expression of p27, and inhibited
cervical cancer development. In addition, elevation of AKT
phosphorylation reversed the effect of HOXA5 upregulation on p27
expression. Therefore, HOXA5 overexpression inhibits cervical
cancer development via modulating p-AKT/p27.
In summary, the results of the present study
revealed that HOXA5 is down-regulated in cervical cancer cell lines
and tissues. The gain-of-function of HOXA5 markedly decreased cell
viability, proliferation and invasion. Meanwhile, HOXA5
overexpression induced caspase-3 activity and increased cell
apoptosis in cervical cancer cell lines. Furthermore,
overexpression of HOXA5 upregulated p27 expression via controlling
the activation of AKT, providing a novel target for the treatment
of cervical cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
ZW designed and prepared the experiments, performed
the experiments and wrote the manuscript. CY contributed to the
reagents/materials/analysis tools. HW wrote, modified and revised
the manuscript and was also involved in the conception of the
study. All authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
This research was approved by the Ethics Committee
of The Fourth People's Hospital of Shaanxi. Informed consent was
obtained from all of the patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
HOX
|
homeobox
|
|
AKT
|
protein kinase B
|
|
CDK
|
cyclin-dependent kinase
|
|
RT-qPCR
|
reverse-transcription polymerase chain
reaction
|
|
PVDF
|
polyvinylidene fluoride
|
|
Annexin V-FITC
|
Annexin V-fluorescein
isothiocyanate
|
|
PI
|
propidium iodide
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
|
|
PBS
|
phosphate-buffered saline
|
|
OD
|
optical density
|
|
BrdU
|
bromodeoxyuridine
|
|
SD
|
standard deviation
|
References
|
1
|
Munagala R, Aqil F, Jeyabalan J and Gupta
RC: Tanshinone IIA inhibits viral oncogene expression leading to
apoptosis and inhibition of cervical cancer. Cancer Lett.
356:536–546. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Carter P, Alifrangis C, Cereser B,
Chandrasinghe P, Del Bel Belluz L, Fotopoulou C, Frilling A, Herzog
T, Moderau N, Tabassum N, et al: Assessing tumor molecular
profiling to guide treatments for patients with advanced female
genital tract malignancy. Oncotarget. 9:6007–6014. 2017.PubMed/NCBI
|
|
3
|
Ditto A, Martinelli F, Bogani G, Fischetti
M, Di Donato V, Lorusso D and Raspagliesi F: Fertility-sparing
surgery in early-stage cervical cancer patients: Oncologic and
reproductive outcomes. Int J Gynecol Cancer. 25:493–497. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bao H, Zhang L and Wang L, Zhang M, Zhao
Z, Fang L, Cong S, Zhou M and Wang L: Significant variations in the
cervical cancer screening rate in China by individual-level and
geographical measures of socioeconomic status: A multilevel model
analysis of a nationally representative survey dataset. Cancer Med.
7:2089–2100. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Imai T, Satoh I, Matsumoto K, Asada Y,
Yamazaki T, Morita S, Saijo S, Okubo JI, Wakamori S, Saijo S and
Matsuura K: Retrospective observational study of occult cervical
lymph-node metastasis in T1N0 tongue cancer. Jpn J Clin Oncol.
47:130–136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hussain I, Bhan A, Ansari KI, Deb P,
Bobzean SA, Perrotti LI and Mandal SS: Bisphenol-A induces
expression of HOXC6, an estrogen-regulated homeobox-containing gene
associated with breast cancer. Biochim Biophys Acta. 1849:697–708.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kiehl S, Zimmermann T, Savai R,
Pullamsetti SS, Seeger W, Bartkuhn M and Dammann RH: Epigenetic
silencing of downstream genes mediated by tandem orientation in
lung cancer. Sci Rep. 7:38962017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang JL, Qi Z, Li YH, Zhao HM, Chen YG and
Fu W: TGFβ induced factor homeobox 1 promotes colorectal cancer
development through activating Wnt/β-catenin signaling. Oncotarget.
8:70214–70225. 2017.PubMed/NCBI
|
|
9
|
Beltran H, Prandi D, Mosquera JM, Benelli
M, Puca L, Cyrta J, Marotz C, Giannopoulou E, Chakravarthi BV,
Varambally S, et al: Divergent clonal evolution of
castration-resistant neuroendocrine prostate cancer. Nat Med.
22:298–305. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gao F, Liu W, Guo Q, Bai Y, Yang H and
Chen H: Physcion blocks cell cycle and induces apoptosis in human B
cell precursor acute lymphoblastic leukemia cells by downregulating
HOXA5. Biomed Pharmacother. 94:850–857. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y, Xu L and Jiang L: miR-1271
promotes non-small-cell lung cancer cell proliferation and invasion
via targeting HOXA5. Biochem Biophys Res Commun. 458:714–719. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Teo WW, Merino VF, Cho S, Korangath P,
Liang X, Wu RC, Neumann NM, Ewald AJ and Sukumar S: HOXA5
determines cell fate transition and impedes tumor initiation and
progression in breast cancer through regulation of E-cadherin and
CD24. Oncogene. 35:5539–5551. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Feng F, Ren Q, Wu S, Saeed M and Sun C:
Hoxa5 increases mitochondrial apoptosis by inhibiting
Akt/mTORC1/S6K1 pathway in mice white adipocytes. Oncotarget.
8:95332–95345. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhu Q, Lv T, Wu Y, Shi X, Liu H and Song
Y: Long non-coding RNA 00312 regulated by HOXA5 inhibits tumour
proliferation and promotes apoptosis in Non-small cell lung cancer.
J Cell Mol Med. 21:2184–2198. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ma C, Wu G, Zhu Q, Liu H, Yao Y, Yuan D,
Liu Y, Lv T and Song Y: Long intergenic noncoding RNA 00673
promotes non-small-cell lung cancer metastasis by binding with EZH2
and causing epigenetic silencing of HOXA5. Oncotarget.
8:32696–32705. 2017.PubMed/NCBI
|
|
16
|
Takahashi O, Hamada J, Abe M, Hata S,
Asano T, Takahashi Y, Tada M, Miyamoto M, Kondo S and Moriuchi T:
Dysregulated expression of HOX and ParaHOX genes in human
esophageal squamous cell carcinoma. Oncol Rep. 17:753–760.
2007.PubMed/NCBI
|
|
17
|
Paraskevopoulou MD and Tsichlis PN: A
perspective on AKT 25-plus years after its discovery. Sci Signal.
10(pii): eaan87912017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li XT, Wang HZ, Wu ZW, Yang TQ, Zhao ZH,
Chen GL, Xie XS, Li B, Wei YX, Huang YL, et al: miR-494-3p
regulates cellular proliferation, invasion, migration, and
apoptosis by PTEN/akt signaling in human glioblastoma cells. Cell
Mol Neurobiol. 35:679–687. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang YQ, Wei XL, Liang YK, Chen WL, Zhang
F, Bai JW, Qiu SQ, Du CW, Huang WH and Zhang GJ: Over-expressed
twist associates with markers of epithelial mesenchymal transition
and predicts poor prognosis in breast cancers via ERK and Akt
activation. PLoS One. 10:e01358512015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang GM, Bao CY, Wan FN, Cao DL, Qin XJ,
Zhang HL, Zhu Y, Dai B, Shi GH and Ye DW: MicroRNA-302a suppresses
tumor cell proliferation by inhibiting AKT in prostate cancer. PLoS
One. 10:e01244102015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yeung CL, Tsang TY, Yau PL and Kwok TT:
Human papillomavirus type 16 E6 suppresses microRNA-23b expression
in human cervical cancer cells through DNA methylation of the host
gene C9orf3. Oncotarget. 8:12158–12173. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhao J, Li L and Peng L: MAPK1
up-regulates the expression of MALAT1 to promote the proliferation
of cardiomyocytes through PI3K/AKT signaling pathway. Int J Clin
Exp Pathol. 8:15947–15953. 2015.PubMed/NCBI
|
|
23
|
Huang Y, Yoon MK, Otieno S, Lelli M and
Kriwacki RW: The activity and stability of the intrinsically
disordered Cip/Kip protein family are regulated by non-receptor
tyrosine kinases. J Mol Biol. 427:371–386. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wan C, Hou S, Ni R, Lv L, Ding Z, Huang X,
Hang Q, He S, Wang Y, Cheng C, et al: MIF4G domain containing
protein regulates cell cycle and hepatic carcinogenesis by
antagonizing CDK2-dependent p27 stability. Oncogene. 34:237–245.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Huang J, Zhou N, Watabe K, Lu Z, Wu F, Xu
M and Mo YY: Long non-coding RNA UCA1 promotes breast tumor growth
by suppression of p27 (Kip1). Cell Death Dis. 5:e10082014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Narita Y, Nagane M, Mishima K, Huang HJ,
Furnari FB and Cavenee WK: Mutant epidermal growth factor receptor
signaling down-regulates p27 through activation of the
phosphatidylinositol 3-kinase/Akt pathway in glioblastomas. Cancer
Res. 62:6764–6769. 2002.PubMed/NCBI
|
|
27
|
Li H, Jia Y, Cheng J, Liu G and Song F:
LncRNA NCK1-AS1 promotes proliferation and induces cell cycle
progression by crosstalk NCK1-AS1/miR-6857/CDK1 pathway. Cell Death
Dis. 9:1982018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mwaka AD, Orach CG, Were EM, Lyratzopoulos
G, Wabinga H and Roland M: Awareness of cervical cancer risk
factors and symptoms: Cross-sectional community survey in
post-conflict northern Uganda. Health Expect. 19:854–867. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cao S, Liu W, Li F, Zhao W and Qin C:
Decreased expression of lncRNA GAS5 predicts a poor prognosis in
cervical cancer. Int J Clin Exp Pathol. 7:6776–6783.
2014.PubMed/NCBI
|
|
31
|
Zhang H, Zhao JH and Suo ZM: Knockdown of
HOXA5 inhibits the tumorigenesis in esophageal squamous cell
cancer. Biomed Pharmacother. 86:149–154. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang ML, Nie FQ, Sun M, Xia R, Xie M, Lu
KH and Li W: HOXA5 indicates poor prognosis and suppresses cell
proliferation by regulating p21 expression in non small cell lung
cancer. Tumour Biol. 36:3521–3531. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee DH, Forscher C, Di Vizio D and
Koeffler HP: Induction of p53-independent apoptosis by ectopic
expression of HOXA5 in human liposarcomas. Sci Rep. 5:125802015.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu WJ, Zhang T, Guo QL, Liu CY and Bai
YQ: Effect of ATRA on the expression of HOXA5 gene in K562 cells
and its relationship with cell cycle and apoptosis. Mol Med Rep.
13:4221–4228. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Palacios F, Abreu C, Prieto D, Morande P,
Ruiz S, Fernández-Calero T, Naya H, Libisch G, Robello C, Landoni
AI, et al: Activation of the PI3K/AKT pathway by microRNA-22
results in CLL B-cell proliferation. Leukemia. 29:115–125. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yang Y, Guo JX, Shao ZQ and Gao JP:
Matrine inhibits bladder cancer cell growth and invasion in vitro
through PI3K/AKT signaling pathway: An experimental study. Asian
Pac J Trop Med. 10:515–519. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fu QF, Liu Y, Fan Y, Hua SN, Qu HY, Dong
SW, Li RL, Zhao MY, Zhen Y, Yu XL, et al: Alpha-enolase promotes
cell glycolysis, growth, migration, and invasion in non-small cell
lung cancer through FAK-mediated PI3K/AKT pathway. J Hematol Oncol.
8:222015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wang J, Chu ES, Chen HY, Man K, Go MY,
Huang XR, Lan HY, Sung JJ and Yu J: microRNA-29b prevents liver
fibrosis by attenuating hepatic stellate cell activation and
inducing apoptosis through targeting PI3K/AKT pathway. Oncotarget.
6:7325–7338. 2015.PubMed/NCBI
|
|
39
|
Das TP, Suman S, Alatassi H, Ankem MK and
Damodaran C: Inhibition of AKT promotes FOXO3a-dependent apoptosis
in prostate cancer. Cell Death Dis. 7:e21112016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Jeyamohan S, Moorthy RK, Kannan MK and
Arockiam AJ: Parthenolide induces apoptosis and autophagy through
the suppression of PI3K/Akt signaling pathway in cervical cancer.
Biotechnol Lett. 38:1251–1260. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Jin C, Zhang Y and Li J: Upregulation of
MiR-196a promotes cell proliferation by downregulating
p27kip1 in laryngeal cancer. Biol Res. 49:402016.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Singh SK, Banerjee S, Acosta EP, Lillard
JW and Singh R: Resveratrol induces cell cycle arrest and apoptosis
with docetaxel in prostate cancer cells via a p53/
p21WAF1/CIP1 and p27KIP1 pathway. Oncotarget.
8:17216–17228. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Cui N, Yang WT and Zheng PS: Slug inhibits
the proliferation and tumor formation of human cervical cancer
cells by up-regulating the p21/p27 proteins and down-regulating the
activity of the Wnt/β-catenin signaling pathway via the
trans-suppression Akt1/p-Akt1 expression. Oncotarget.
7:26152–26167. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Dan HC, Sun M, Yang L, Feldman RI, Sui XM,
Ou CC, Nellist M, Yeung RS, Halley DJ, Nicosia SV, et al:
Phosphatidylinositol 3-kinase/Akt pathway regulates tuberous
sclerosis tumor suppressor complex by phosphorylation of tuberin. J
Biol Chem. 277:35364–35370. 2002. View Article : Google Scholar : PubMed/NCBI
|