Introduction
Osteosarcoma (OS) has been reported as one of the
most common aggressive types of bone tumor (1). Children and young adults
(<20-years-old) exhibit a high incidence of OS (2). In the past decades, with the
application of neoadjuvant chemotherapy using doxorubicin,
cisplatin, doxorubicin and ifosfamide in OS treatment, the 5-year
survival rate has increased to ~50–80% (3,4);
however, the prognosis of OS remains poor and the majority of
patients succumb to mortality due to metastases following surgical
resection or intensive chemotherapy (5–7).
Therefore, it is necessary to identify a sensitive molecular
biomarker and therapeutic target for the treatment of OS.
Forkhead box N3 (FOXN3), also known as checkpoint
suppressor 1, was first reported in yeast as a suppressor of check
point defects (8). FOXN3 belongs to
the forkhead box family, a novel family of transcription factors.
The FOX family has been classified into 15 subclasses, including
FOXA and FOXS (9). Numerous studies
have demonstrated the physiological roles of FOXN3 in embryonic
development (10,11). Previous investigations revealed that
FOXN3 expression was notably downregulated in numerous cancer
tissues compared with in adjacent non-cancerous tissues, including
laryngeal, oral squamous cell and hepatocellular carcinomas, and
diffuse large B-cell lymphoma (12–16).
This evidence indicates that FOXN3 may serve a key function in cell
proliferation and apoptosis within human cancer via the regulation
of gene transcription. Similar to other members of the forkhead
transcription factors, FOXN3 has also been reported to bind with
several nuclear proteins, including histone deacetylase (HDAC)1,
HDAC2 and multiple endocrine neoplasia type 1 (9). Previously, it was demonstrated that
FOXN3 regulated cell proliferation by suppressing PIM2 and protein
biosynthesis (17), or by
downregulating E2F transcription factor 5 (E2F5) in human cells to
control the cell cycle (9);
however, the function and underlying mechanisms of FOXN3 in OS is
poorly understood.
Sirtuin 6 (SIRT6) belongs to the SIRT family of
proteins and has been notably identified as a critical regulator in
a variety of physiological and pathological processes, including
life span, glucose metabolism, DNA damage repair and cancer
(18). SIRT6 has been reported to
deacetylate histone H3K9 at the promoter of numerous genes involved
in lipid metabolism and glycolysis (19). A recent study revealed that SIRT6
contributes to the migration and invasion of OS cells via the
extracellular signal-regulated kinase (ERK)1/2/matrix
metalloproteinase-9 (MMP-9) pathway (20); however, the upstream molecules of
SIRT6 signaling in OS remains unknown.
The present study demonstrated that FOXN3 was
downregulated in OS tissues and cell lines. In addition, the
expression of FOXN3 was negatively associated with tumor size,
metastasis and tumor, node and metastasis (TNM) stage. The results
of the present study suggest that FOXN3 suppressed the
proliferation, migration and invasion of OS cells. Furthermore,
FOXN3 was proposed to transcriptionally suppresses SIRT6
expression, thereby inhibiting MMP-9 secretion. Therefore, FOXN3
may serve as a prognostic predictor and a therapeutic target for
patients with OS.
Materials and methods
Cell culture
Human OS cell lines, including U2OS and MG-63, and
the osteoblast cell line, hFOB1.19, were purchased from the
American Type Culture Collection (Manassas, VA, USA). All cells
were cultured in Dulbecco's modified Eagle's medium (DMEM; HyClone;
GE Healthcare Life Sciences, Logan, UT, USA) supplemented with 100
U/ml penicillin and 100 µg/ml streptomycin, and fetal bovine serum
(10%; HyClone; GE Healthcare Life Sciences). Cell were cultured in
an incubator containing 5% CO2 humidified atmosphere at
37°C.
Clinical samples
A total of 78 pairs of clinical OS and adjacent
normal tissue specimens were obtained from patients who were
diagnosed with OS in the Department of Orthopedics, Yidu Central
Hospital of Weifang (Weifang, China) during 2010 to 2016. A total
of 45 males and 33 females aged between 12–28-years-old, with a
mean age of 18.4 years were employed in the present study. All
tissues were obtained prior to the administration of immunotherapy,
chemotherapy or radiotherapy. The majority of the obtained samples
(>86%) were collected between 2010 to 2013. The survival data of
some patients were of <5 years; however, the majority of
patients' survival data were of 5 years. All patients had provided
written informed consent. The present study was approved by the
Research Ethics Committee of Yidu Central Hospital of Weifang
(LK2017012). All specimens were stored in liquid nitrogen prior to
use.
Transfection
A total of ~5×105 U2OS or MG-63 cells
were placed into 6-well plates and incubated for 24 h at 37°C.
Subsequently, cells were transfected with 2.5 µg vector (pcDNA3.1),
pcDNA3.1-FOXN3 and/or pcDNA3.1-SIRT6 or 50 nM small interfering
(si)RNAs using Lipofectamine® 2000 reagent (Invitrogen,
Thermo Fisher Scientific, Inc., Waltham, MA, USA) according to the
manufacturer's protocols. After transfection for 48 h, successful
transfection was determined using reverse
transcription-quantitative polymerase chain reaction (RT-qPCR) and
western blot analyses. The vector (pcDNA3.1), pcDNA3.1-FOXN3 and
pcDNA3.1-SIRT6 were purchased from Vigene Biosciences, Inc.
(Rockville, MD, USA). The human FOXN3 and SIRT6 gene sequences were
retrieved from the NCBI gene bank (https://www.ncbi.nlm.nih.gov/nuccore; NM_001085471 and
NM_001193285.2, respectively) SiRNAs were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). The sequence of the
siRNAs was as follows: SiFOXN3, 5′-GUACCUUCUUCAAGAGAAAUG-3′ and
scramble siRNA (siControl), 5′-UUCUCCGAACGUGUCACGU-3′.
RT-qPCR
Total RNA was extracted from tissues or cells by
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocols.
Subsequently, 2 µg RNA was used to synthesize cDNA using
SuperScript III Reverse Transcriptase kit (Invitrogen; Thermo
Fisher Scientific, Inc.). The reverse transcription conditions were
as follows: 5 min at 25°C, 30 min at 50°C and 15 min at 70°C.
Finally, qPCR was conducted using the SYBR Fast qPCR kit (Takara
Bio, Inc., Otsu, Japan) on an ABI 7500 system (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The RT-qPCR conditions were as
follows: 5 min at 98°C, denaturation at 98°C for 30 sec, annealing
at 55°C for 30 sec and extension at 72°C for 20 sec, performed for
30 cycles. The primers were as follows: FOXN3 forward,
5′-CCCTTCTCCAAGATCCTGAC-3′, reverse, 5′-GCTGTAGTTGTGATCCTCCT-3′;
SIRT6 forward, 5′-GTTCGACACCACCTTTGAG-3′, reverse,
5′-ACGTACTGCGTCTTACAC-3′ and GAPDH forward,
5′-ATTTCCTGGTATGACAACGA-3′ and reverse, 5′-GGAGATTCAGTGTGGTGG-3′.
GAPDH was used as an internal control. Each experiment was
performed in triplicate. As presented in Table I, the mean value of FOXN3 mRNA
expression in tumor cells considered as the standard; higher values
than the standard was denoted as high expression and values lower
than the standard values were considered as low expression. The
stage of OS was according to tumor, node and metastasis (TNM)
staging system (21). The relative
gene expression was measured using 2−∆∆Cq method
(22).
 | Table I.Clinicopathological variables in 78
patients with osteosarcoma. |
Table I.
Clinicopathological variables in 78
patients with osteosarcoma.
|
|
| FOXN3 protein
expression |
|
|---|
|
|
|
|
|
|---|
| Variables | Patient no.
(n=78) | Low (n=43) | High (n=35) | P-value |
|---|
| Sex |
|
|
|
|
|
Male | 45 | 23 | 22 | 0.405 |
|
Female | 33 | 20 | 13 |
|
| Age |
|
|
|
|
|
<18 | 50 | 30 | 20 | 0.248 |
|
≥18 | 28 | 13 | 15 |
|
| Tumor size |
|
|
|
|
| <8
cm | 42 | 14 | 28 | 0.001a |
| ≥8
cm | 36 | 29 | 7 |
|
| TNM stage |
|
|
|
|
|
I–III | 44 | 19 | 25 | 0.016a |
| IV | 34 | 24 | 10 |
|
| Metastasis |
|
|
|
|
|
Yes | 42 | 30 | 12 | 0.002a |
| No | 36 | 13 | 23 |
|
| SIRT6
expression |
|
|
|
|
|
High | 42 | 33 | 9 | 0.001a |
|
Low | 36 | 10 | 26 |
|
Western blotting
Total protein was extracted from tissues and cells
using radioimmunoprecipitation assay lysis buffer (Beyotime
Institute of Biotechnology, Haimen, China). The concentration of
protein was measured via a Bicinchoninic Acid (BCA) protein assay
kit (Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China) according to the manufacturer's protocols. Protein samples
(45 µg) were loaded and then separated by 10% SDS-PAGE, and then
the protein was transferred to polyvinylidene difluoride membranes
(Merck KGaA). The membranes were blocked with 5% skimmed milk at
room temperature for 1 h and then incubated with primary antibodies
at 4°C overnight. Following washing with PBS with 0.1% Tween-20
(PBST) three times, the membranes were incubated with horseradish
peroxidase (HRP)-conjugated second antibodies at room temperature
for 1 h and washed with PBST three times. The blots were identified
using Western Blotting Luminol Reagent (Santa Cruz Biotechnology,
Inc., Dallas, TX, USA). The anti-FOXN3 polyclonal antibody
(1:2,000; ab50756) and anti-SIRT6 polyclonal antibody (1:1,000;
ab62739), HRP-conjugated anti-rabbit antibody (1:5,000; ab6721) and
HRP-conjugated anti-mouse antibody (1:5,000; ab6789) were purchased
from Abcam (Cambridge, UK). The anti-β-actin antibody (1:5,000;
A3854) was purchased from Sigma-Aldrich (Merck KGaA).
Cell counting kit-8 (CCK-8) assay
A CCK-8 assay was used to investigate the roles of
FOXN3 on cell proliferation. Briefly, ~3,000 transfected cells were
resuspended in 200 µl DMEM and placed into 96-well plates; 20 µl
CCK-8 reagent (Beyotime Institute of Biotechnology) was added to
each subset well at 0, 24, 48 and 72 h. Cells were cultured at 37°C
for 1 h. The optical density was quantitated at 450 nm using a
microplate reader. Each experiment was performed in triplicate.
Colony formation assay
A colony formation assay was performed to determine
the effects of FOXN3 on cell proliferation. In brief,
~5×103 transfected U2OS and MG-63 cells with serum-free
DMEM were plated in 6-well plates, and cultured for 14 days at
37°C. Cells were fixed with 4% paraformaldehyde solution at room
temperature for 15 min and stained with 0.5% w/v crystal violet
(Sigma-Aldrich; Merck KGaA) at room temperature for 15 min.
Colonies (>50 cells) were counted under a light microscope
(magnification, ×20). A total of 5 random fields were analyzed.
Each experiment was performed in triplicate.
Wound healing analysis
Following transfection for 48 h, cells were plated
into 24-well plates. Wound healing analysis was performed to
determine the effects of FOXN3 on the migration of U2OS and MG-63
cells. In brief, cells at 90% confluence were wounded with a 10-µl
sterile pipette tip, washed with PBS three times to remove the
detached cells, and cultured in serum-free DMEM at 37°C. Following
wounding for 48 h, the images were captured under a light
microscope (magnification, ×20). The migratory distance was
measured using Image-Pro Plus software 6.0 (Media Cybernetics,
Inc., Rockville, MD, USA). Each experiment was performed in
triplicate.
Transwell invasion assay
In brief, Transwell chambers (Corning Costar;
Corning Incorporated, Corning, NY, USA) were coated with 80 µl
Matrigel and warmed at 37°C prior to use. Transfected U2OS and
MG-63 cells (2×105) were resuspended in 500 µl
serum-free DMEM and placed in the upper chambers. The lower
chambers were filled with 500 µl DMEM supplemented with 10% FBS.
The cells were incubated at 37°C for 18 h. The cells on the surface
of the chamber were removed with a swab and then stained with 0.1%
crystal violet at room temperature for 15 min. The number of cells
on the underlayer of chamber were counted under a light microscope
(magnification, ×20); 5 random fields were analyzed. Each
experiment was performed in triplicate.
Chromatin immunoprecipitation (ChIP)
and quantitative (qChIP) assays
ChIP analysis was performed using a ChIP Assay kit
(Beyotime Institute of Biotechnology) according to manufacturer's
protocols. Briefly, U2OS and MG-63 cells were cultured to 90–100%
confluence, and washed with cold-PBS three times and chemically
cross-linked with 1% formaldehyde at 37°C for 30 min. Subsequently,
cells were lysed with 2 ml lysis buffer at 4°C for 90 min and
sonicated under 4×15 times at 4°C. FOXN3 antibody (1:200, ab50756,
Abcam) and anti-rabbit IgG (1:200, ab171870, Abcam) were added to
the lysis solution and incubated at 4°C overnight. Protein A beads
were used to isolate FOXN3- or IgG-interacted DNA fragments.
Following elution with 120 µl elution buffer (10% SDS, 10% 1 M
NaHCO3, 80% ddH2O), crosslinking was reversed
with elution buffer containing 0.2 M NaCl and 2 µl RNase A (10
mg/ml; Sigma-Aldrich; Merck KGaA) at 65°C overnight. After
incubation with 4 µl proteinase K (10 mg/ml; Sigma-Aldrich; Merck
KGaA) at 60°C for 1 h, the bound chromatin was purified using a PCR
Purification kit (Qiagen, Inc., Valencia, CA, USA) according to
manufacturer's protocols, and qPCR was performed. The qPCR
conditions were as follows: 5 min at 98°C, denaturation at 98°C for
30 sec, annealing at 55°C for 30 sec and extension at 72°C for 20
sec, performed for 30 cycles. The primer sequences were as follows:
SIRT6 forward, 5′-AATAAGAAGGGCCTGATGGC-3′, reverse,
5′-TGTATGTGGGAGAAAGAAGC3′ and E2F transcription factor 5 (E2F5)
forward, 5′-TCTTCAGCAGGATCTATTAGTGG-3′ and reverse:
5′-TGTAGTCATCTGCCGGGGTA-3′. IgG was used as internal control; E2F
was used as positive control (9).
Each experiment was performed in triplicate.
Luciferase reporter assay
The promoter region (−2000, +200) of SIRT6 was
cloned into pGL3-basic plasmid (Biofeng, Beijing, China). U2OS and
MG-63 cells (5×105) were seeded in 6-well plates and
transfected with vector or pcDNA3.1-FOXN3 (0, 0.5, 1 or 2 µg),
together with 1 µg pGL3-basic plasmid or pGL3-SIRT6 and 0.5 µg
Renilla plasmid (Promega Corporation, Madison, WI, USA)
using Lipofectamine® 2000 reagent (Invitrogen, Thermo
Fisher Scientific, Inc.). Following transfection for 24 h,
Renilla luciferase and firefly activities were determined
using a dual-luciferase reporter system (Promega Corporation)
according to the manufacturer's protocols. Renilla
luciferase was used as internal control. Each experiment was
performed in triplicate.
ELISA
Briefly, after transfection for 48 h, cells were
seeded into 6-well plates at a density of 3×105
cells/well with serum-free DMEM. Cell culture supernatants were
harvested 72 h later, and centrifuged at 2,000 × g for 15 min at
4°C. The effects of FOXN3 on the secretion of MMP-2 and MMP-9 into
the supernatant from U2OS and MG-63 cells were determined using
corresponding ELISA kits (cat. nos. CSB-E04675h and CSB-E08006h;
Cusabio, Wuhan, China) according to the manufacturer's protocols.
Each experiment was performed in triplicate.
Gelatin zymography
Secreted MMPs in conditioned medium (serum-free
DMEM) were affinity-adsorbed with gelatin-Sepharose as previously
described (23). The concentration
of protein was measure using a BCA protein assay kit (Beijing
Solarbio Science & Technology Co., Ltd.). SDS sample buffer
lacking dithiothreitol was added and proteins (10 µg/ml) were
resolved on 10% acrylamide, SDS gels contained polymerized gelatin
(0.5–2 mg/ml). MMPs were renatured via two detergent exchange
washes (2.5% Triton X-100, 50 mM Tris HCl, 5 mM CaCl2, 1
µM ZnCl2; 1% Triton X-100, 50 mM Tris HCl, 5 mM
CaCl2, 1 µM ZnCl2). Gels were incubated for
48 h at 23°C for MMP-mediated degradation of gelatin, which was
followed by staining with Coomassie brilliant blue at room
temperature for 1 h. Gels were destained with destaining solution
(40% methanol, 50% acetic acid, 10% ddH2O) at room
temperature, and scanned and inverse images were quantified using
ImageJ software 1.8.0 (National Institutes of Health, Bethesda, MD,
USA), with relative grayscale values for MMP-9 normalized to cell
lysate FOXN3/β-actin ratios.
Statistical analysis
Data were presented as the mean ± standard
deviation. Prism 5 software (GraphPad Software, Inc., La Jolla, CA,
USA) was used for statistical analysis. Comparisons between cancer
and adjacent normal tissue were performed using a paired-samples
t-test based on a bi-directional hypothesis for continuous
variables. The Kaplan-Meier method and a log-rank test were used to
analyze survival curves. A Student's t-test was applied to analyze
the differences between two groups. Differences between multiple
groups were analyzed by analysis of variance followed by a Tukey's
post hoc test. P<0.05 was considered to indicate a statistically
significant difference.
Results
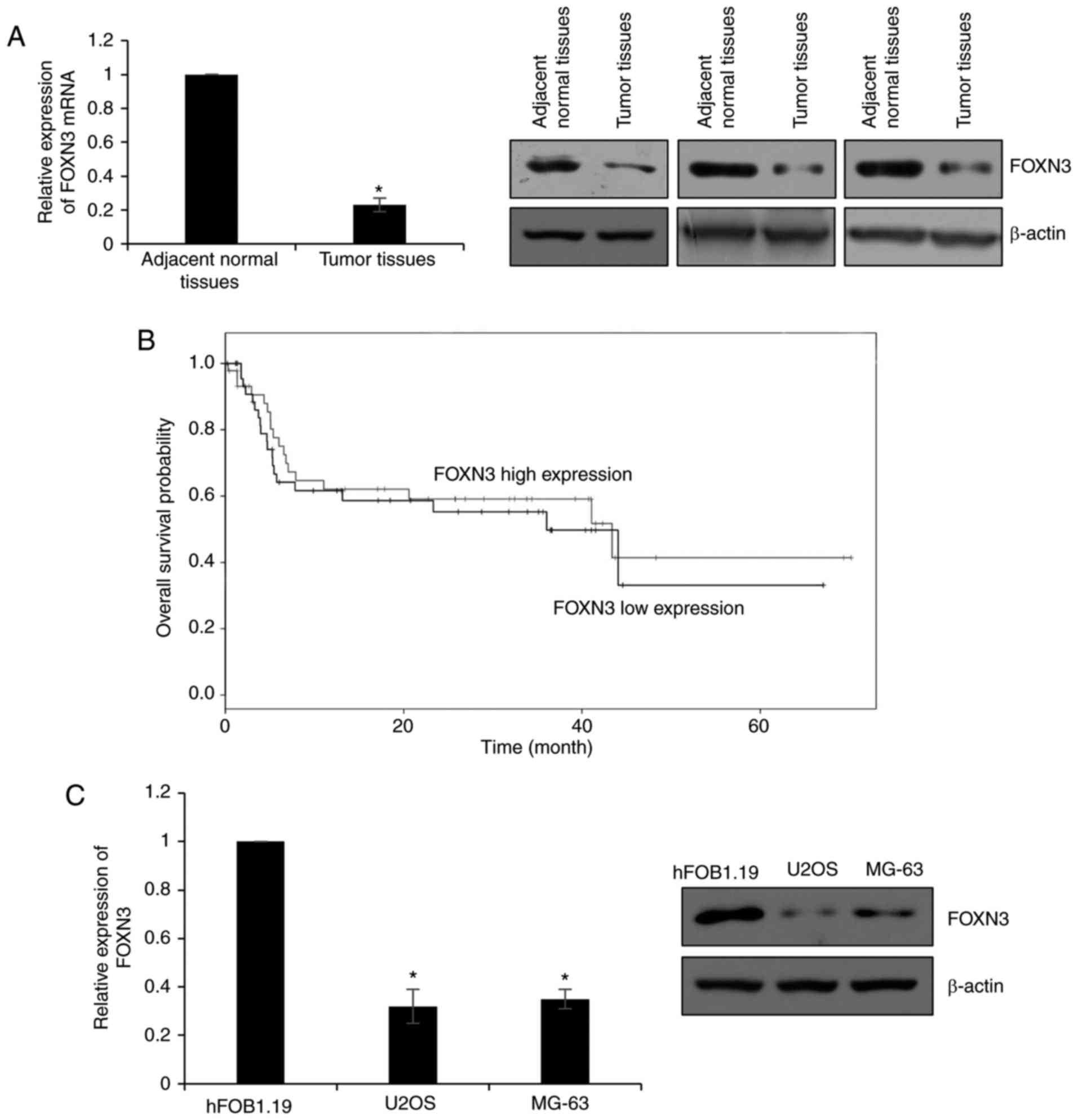
FOXN3 expression is significantly
downregulated in OS
To determine the expression profile of FOXN3 in OS,
78 pairs of OS and adjacent normal tissues were obtained to detect
the expression of FOXN3 by RT-qPCR and western blotting. The
results of RT-qPCR and western blotting revealed that the mRNA
levels of FOXN3 were significantly downregulated in tumor tissues,
compared with adjacent normal tissues; the protein expression
levels were notably downregulated in the OS tissues compared with
in the adjacent normal samples (Fig.
1A). In addition, the association between the expression of
FOXN3 and the clinical information of patients with OS was
analyzed, which demonstrated that the expression of FOXN3 was
negatively correlated with tumor size, metastasis and TNM stage
(Table I). These findings indicated
that FOXN3 may serve as a tumor suppressor in OS.
Additionally, the present study analyzed whether
FOXN3 expression was associated with the prognosis of OS via
Kaplan-Meier analysis. The results suggested that patients who
possessed high expression levels of FOXN3 exhibited better
prognosis than patients with low FOXN3 expression levels (P=0.041;
Fig. 1B). Furthermore, the
expression of FOXN3 in OS cells (U2OS and MG-63) was examined. The
osteoblast cell line hFOB1.19 was used as a control. Similar
results were observed to that of OS and adjacent normal tissues.
The expression levels of FOXN3 were significantly lower in U2OS and
MG-63 cells compared with in hFOB1.19 cells (Fig. 1C). Collectively, these data
indicated that FOXN3 expression is downregulated in OS and may
serve as a tumor suppressor.
FOXN3 inhibits the proliferation of OS
cells
To evaluate the roles of FOXN3 in the progression of
OS, FOXN3 was ectopically expressed or downregulated in U2OS and
MG-63 cells; the expression of FOXN3 was analyzed via RT-qPCR and
western blotting. The mRNA expression levels of FOXN3 in the
overexpression group were significantly upregulated compared with
in the vector-transfected group; however, cells transfected with
siFOXN3 exhibited significantly decreased expression levels
compared with in the siControl group. Additionally, the expression
levels of FOXN3 protein were notably increased and decreased in the
overexpression and downregulated groups, respectively, compared
with in the corresponding controls (Fig. 2A). As of the negative correlation
between FOXN3 expression and tumor size reported in the present
study, the effects of FOXN3 on cell proliferation were determined
via CCK-8 and colony formation assays. The results of the CCK-8
assay demonstrated that ectopic expression of FOXN3 significantly
decreased and knockdown of FOXN3 significantly increased cell
proliferation compared with in the vector and siControl groups,
respectively (Fig. 2B).
Subsequently, a colony formation assay was conducted, which
revealed that overexpression of FOXN3 significantly reduced the
number of colonies, and knockdown of FOXN3 significantly increased
the number of colonies compared with in the corresponding control
groups (Fig. 2C). The findings of
the present study suggested that FOXN3 inhibits the proliferation
of OS cells.
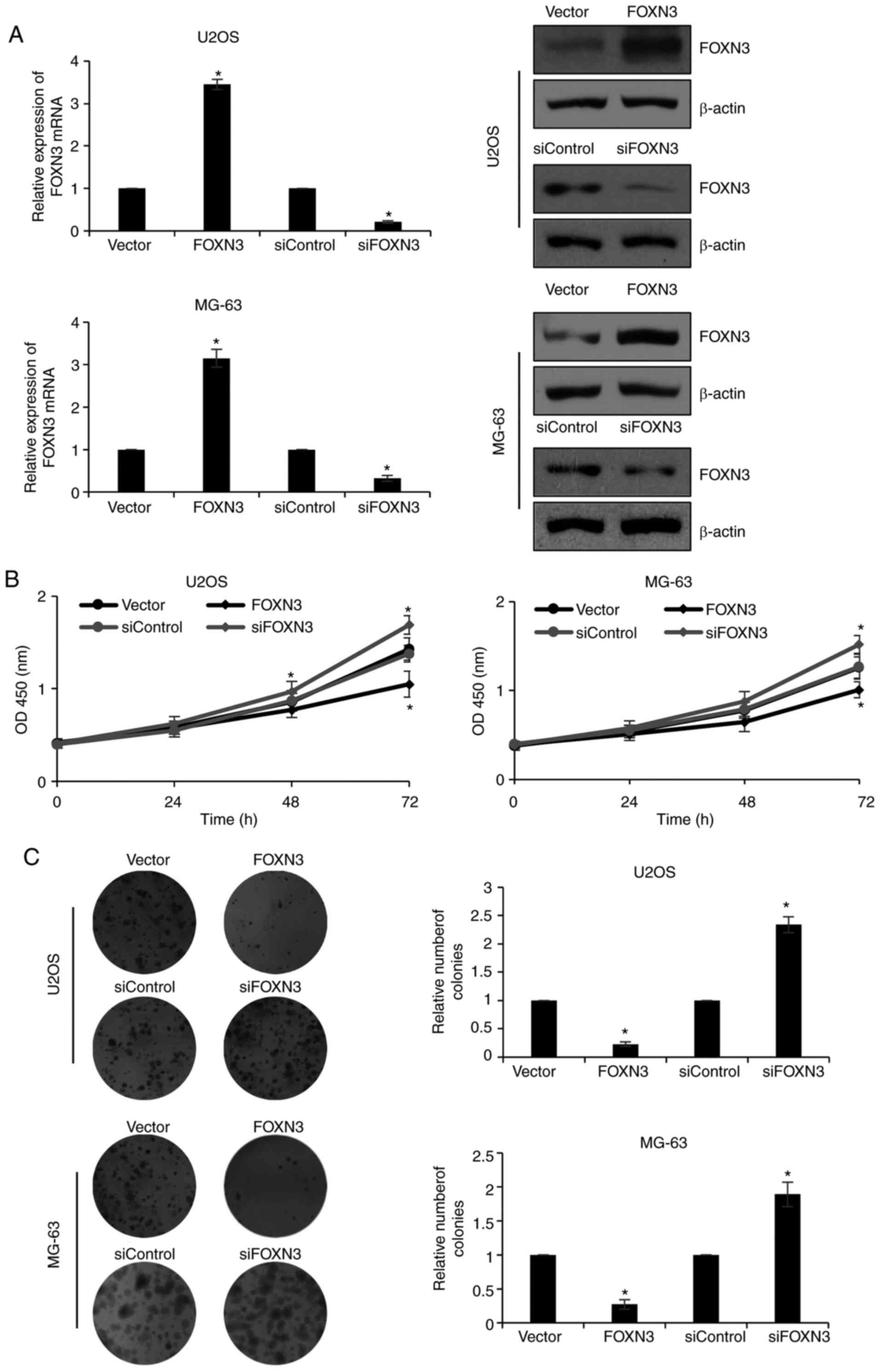 | Figure 2.FOXN3 inhibits the proliferation of
osteosarcoma cells. (A) Reverse transcription-quantitative
polymerase chain reaction and western blot analyses of FOXN3
expression in U2OS or MG-63 cells transfected with vector, FOXN3,
siControl, siFOXN3. *P<0.05 FOXN3 vs. vector, siFOXN3 vs.
siControl. (B) Effects of FOXN3 on cell proliferation was assessed
using a CCK-8 assay. *P<0.05 FOXN3 vs. vector, siFOXN3 vs.
siControl. (C) Colony formation assay was performed in U2OS or
MG-63 cells transfected with vector, FOXN3 or siControl, siFOXN3
Magnification, ×20. *P<0.05 FOXN3 vs. vector, siFOXN3 vs.
siControl. FOXN3, forkhead box N3; si, small interfering RNA. |
Downregulation of FOXN3 expression
promotes the migration and invasion of OS cells
In the present study, the roles of FOXN3 on cell
migration and invasion were analyzed. A wound healing assay was
performed to determine the effects of FOXN3 on cell migration,
which revealed that FOXN3 overexpression significantly suppressed
cell migration compared with in the vector group; however,
inhibition of FOXN3 expression significantly promoted cell
migration compared with in the siControl group (Fig. 3A). In addition, a Transwell invasion
assay was conducted to determine the effects of FOXN3 on cell
invasion, which demonstrated that FOXN3 overexpression
significantly decreased the number of invaded cells compared with
in the vector group; however, downregulation of FOXN3 increased the
number of invaded cells compared with in the siControl group
(Fig. 3B). These data indicated
that FOXN3 suppressed the migration and invasion of OS cells.
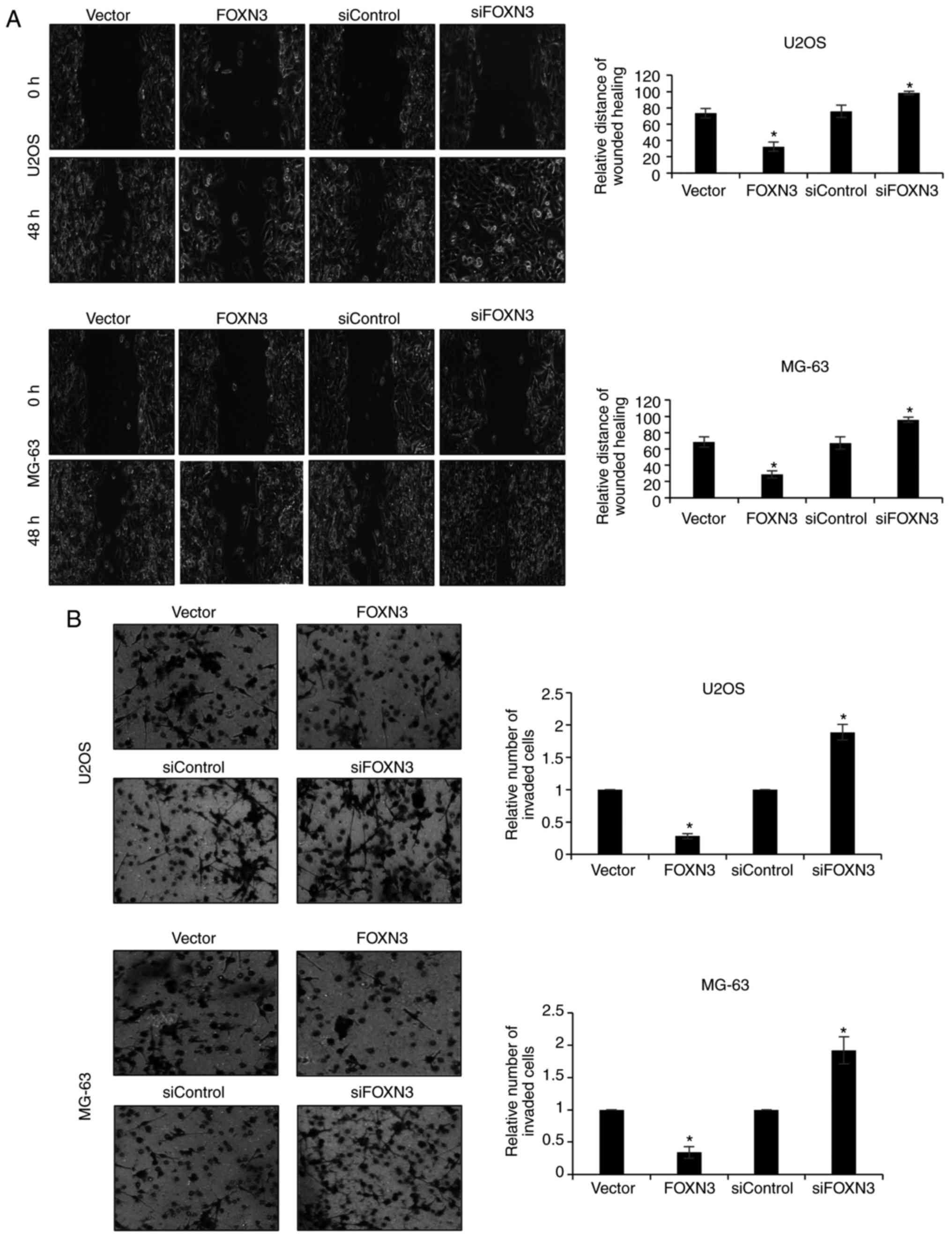 | Figure 3.Downregulation of FOXN3 expression
promotes the migration and invasion of osteosarcoma cells. (A) U2OS
or MG-63 cells were transfected with vector, FOXN3, siControl and
siFOXN3, respectively. Following 48 h post-transfection, a wound
healing assay was performed. The relative distance of wound healing
was measured under a microscope. *P<0.05 vs. FOXN3 vs. vector,
siFOXN3 vs. siControl. Magnification, ×20. (B) FOXN3 was
overexpressed or downregulated in U2OS or MG-63 cells. A Transwell
invasion assay was performed to detect the effects of FOXN3 on cell
invasion. *P<0.05 FOXN3 vs. vector, siFOXN3 vs. siControl. FOXN3
(magnification, ×20), forkhead box N3; si, small interfering
RNA. |
FOXN3 transcriptionally regulates
SIRT6 expression
A recent study reported that SIRT6 contributed to
the migration and invasion of OS cells via the ERK1/2/MMP-9 pathway
(20); however, the upstream
effectors of SIRT6 in OS remains unknown. In order to determine
whether FOXN3 regulates SIRT6 expression, FOXN3 expression was
upregulated or silenced in U2OS and MG-63 cells, RT-qPCR and
western blot analyses were conducted. The results revealed that the
expression levels of SIRT6 mRNA were significantly downregulated
when FOXN3 was overexpressed compared with in the vector group;
FOXN3 silencing significantly increased the expression of SIRT6
compared with in the siControl group (Fig. 4A). Additionally, ChIP and qChIP
assays were performed, which revealed that FOXN3 may bind the
promoter region of SIRT6 in U2OS and MG-63 cells; E2F5 was used as
a positive control (9) (Fig. 4B). Additionally, the results of the
dual luciferase reporter assay suggested that FOXN3 significantly
suppressed SIRT6 expression via transcription compared with in
vector-transfected cells (Fig. 4C).
In addition, the expression of SIRT6 in U2OS and MG-63 OS cells,
and the osteoblast cell line, hFOB1.19 were determined. The results
revealed that SIRT6 expression was significantly upregulated in
U2OS and MG-63 cells compared with in hFOB1.19 cells (Fig. 4D). Furthermore, the expression of
SIRT6 in OS tissues was negatively correlated with FOXN3 expression
(Table I; P<0.05). Collectively,
the expression of SIRT6 was proposed to be regulated by FOXN3 in
OS.
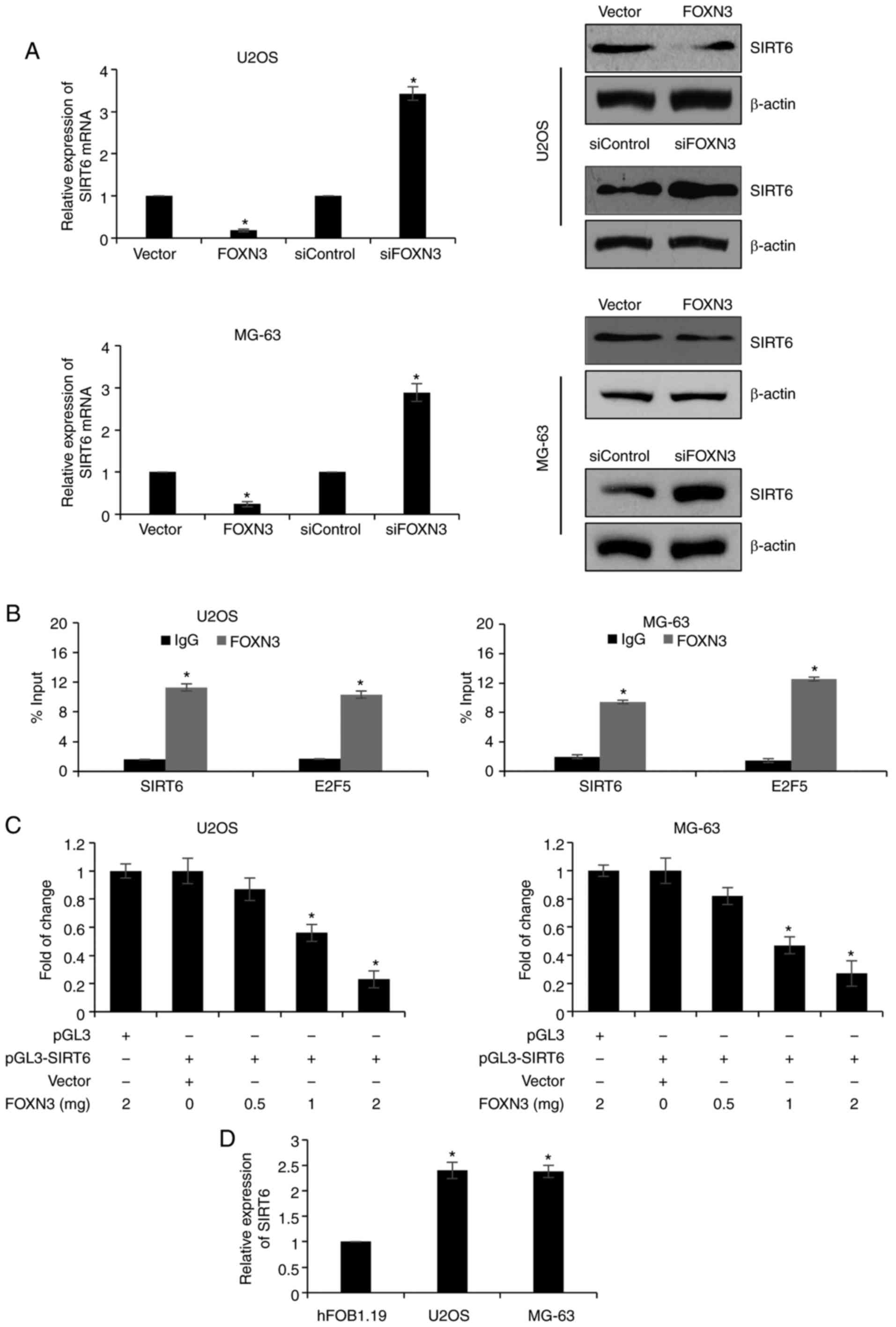 | Figure 4.FOXN3 transcriptionally regulates
SIRT6 expression. (A) U2OS or MG-63 cells were transfected with
vector, FOXN3, siControl or siFOXN3, respectively. After 48 h
post-transfection, the expression of SIRT6 was detected by RT-qPCR
and western blotting. *P<0.05 FOXN3 vs. vector, siFOXN3 vs.
siControl. (B) Binding of FOXN3 to the SIRT6 promoter in U2OS or
MG-63 cells was analyzed by ChIP and quantitative ChIP assays using
antibodies to FOXN3 and IgG, and RT-qPCR was used to analyze the
SIRT6 promoter. E2F5 was used as a positive control. *P<0.05 vs.
IgG. (C) Activity of the SIRT6 promoter following transfection of
FOXN3 into U2OS or MG-63 cells was detected via a luciferase
reporter assay. *P<0.05 vs. vector + pGL3-SIRT6. (D) RT-qPCR
analysis of SIRT6 expression in osteosarcoma cell lines and the
osteoblast cell line, hFOB1.19, which was used as a control group.
*P<0.05 vs. hFOB1.19. ChIP, chromatin immunoprecipitation;
FOXN3, forkhead box N3; RT-qPCR, reverse transcription-quantitative
polymerase chain reaction; si, small interfering RNA. |
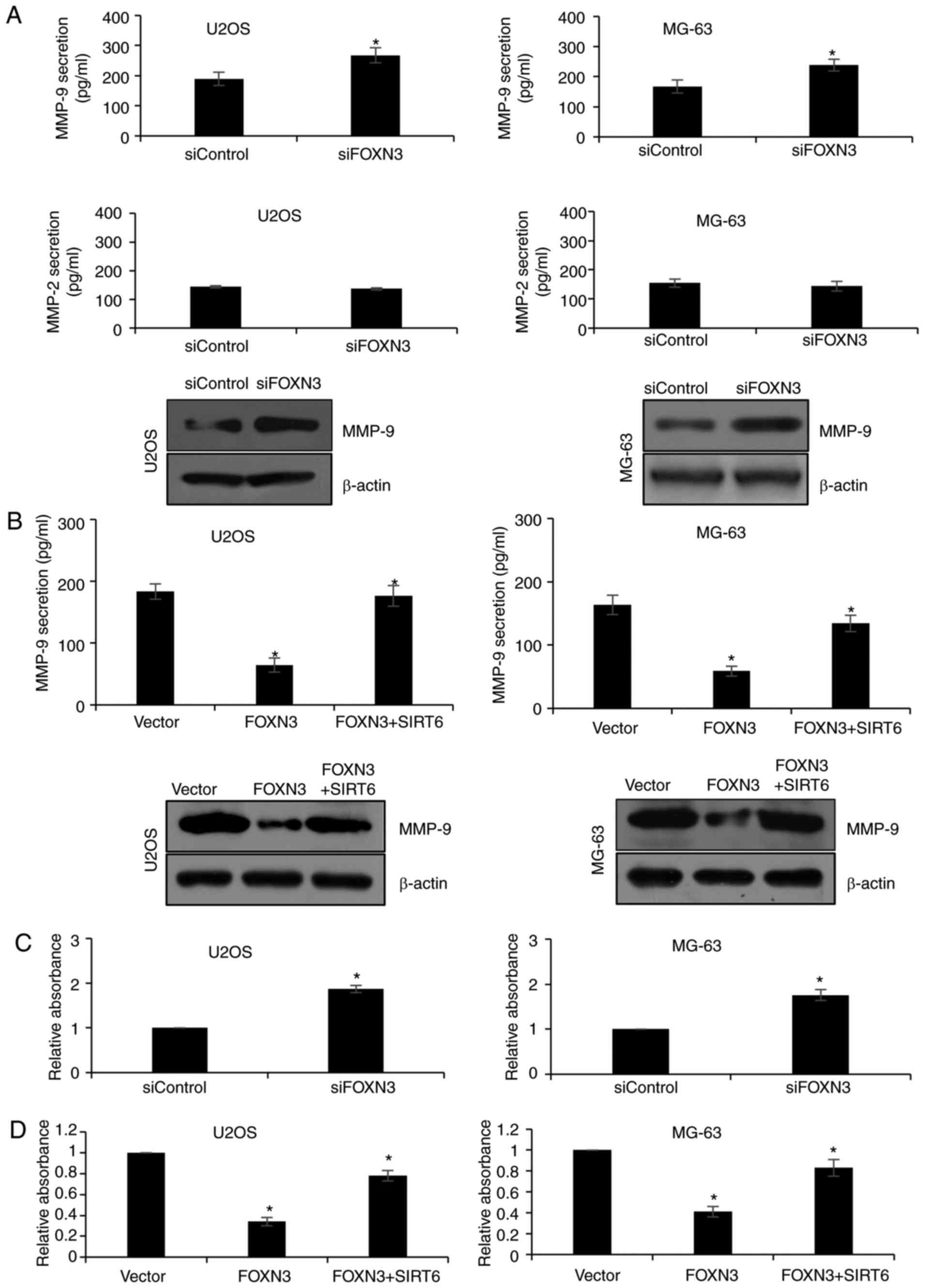
FOXN3 suppresses MMP-9 secretion via
the regulation of SIRT6 expression
MMP-9 can digest gelatins or denatured collagens
(24), and has been reported to
promote metastasis in some types of cancers (25,26).
In addition, SIRT6 was proposed to regulate MMP-9 expression in OS
and non-small cell lung cancer (27). As FOXN3 was determined to regulate
SIRT6 expression in the present study, it was hypothesized that
FOXN3 may also regulate MMP-9 secretion. The results of ELISA
demonstrated that FOXN3 knockdown significantly increased the
secretion levels of MMP-9 in U2OS and MG-63 cells compared with in
the siControl group; FOXN3 silencing was observed to have no
notable effect on MMP-2 secretion (Fig.
5A). FOXN3 overexpression significantly reduced the secretion
levels of MMP-9 in U2OS and MG-63 cells compared with in the vector
group (Fig. 5B), which were
significantly reversed when cells simultaneously overexpressed
FOXN3 and SIRT6 (Fig. 5B).
Furthermore, the gelatin zymography assay demonstrated that FOXN3
significantly suppressed MMP-9 secretion compared with in the
vector group, which was reversed following overexpression of SIRT6
(Fig. 5C and D). The findings of
the present study demonstrated that FOXN3 may suppresses MMP-9
secretion via the regulation of SIRT6.
Discussion
At present, the biological roles of FOXN3 are still
poorly understood. Numerous studies have demonstrated the
physiological roles of FOXN3 in the craniofacial development of
mouse and eye development of xenopus laevis (28,29).
FOXN3 has been reported to be downregulated in HCC and other cancer
types (9); however, the roles of
FOXN3 in OS require further investigation.
In the present study, the expression pattern of
FOXN3 in OS was analyzed. Downregulation of FOXN3 expression in OS
tissues was observed at the mRNA and protein levels, which
suggested that FOXN3 expression may be a novel diagnostic marker of
OS. Additionally, FOXN3 was also downregulated in the OS cell
lines, U2OS and MG-63, compared with in the osteoblast cell line,
hFOB1.19. The expression of FOXN3 was negatively correlated with
tumor size and metastasis, as well as TNM stage, suggesting that
FOXN3 may serve as a tumor suppressor in OS. Furthermore, low
expression of FOXN3 may predict a poor prognosis for patients with
OS.
A recent study revealed that FOXN3 inhibited the
proliferation of HCC cells (9). In
the present study, numerous functional experiments, including
colony formation and CCK-8 assays were performed, which revealed
that FOXN3 also suppressed the proliferation of OS cells. In
addition, wound healing and Transwell invasion assays also
suggested that FOXN3 inhibited the migration and invasive abilities
of OS cells.
A recent study proposed that SIRT6 was overexpressed
in OS tissues and cell lines (20).
SIRT6 facilitated the migration and invasion of OS cells (20); however, the upstream effectors of
SIRT6 in OS have not yet been reported. The present study
demonstrated that FOXN3 could regulate SIRT6 expression at the mRNA
and protein levels, which suggested that FOXN3 may
transcriptionally regulate SIRT6 expression. Subsequently, ChIP and
qChIP, as well as a luciferase reporter assay suggested that SIRT6
was transcriptionally regulated by FOXN3. In addition, the
secretion of MMP-9 was suppressed by FOXN3, which may occur via the
regulation of SIRT6.
However, there are limitations to the present study.
Firstly, the expression of FOXN3 in OS tissues should be detected
via immunohistochemistry staining. Additionally, experiments should
be performed to determine whether FOXN3 suppresses the migration
and invasion of OS cells in vivo. Furthermore, it is
necessary to determine the upstream effectors of FOXN3 in OS. A
previous report indicated that the Wnt signaling pathway could
reduce FOXN3 expression (30). In
addition, the Wnt/β-catenin signaling pathway was reported to be
activated in OS tissues and cells, and aberrant activation may
serve a central role in the tumorigenesis, metastasis and
chemotherapeutic responses of OS (31,32).
Numerous reports have revealed that microRNAs regulate the Wnt
signaling pathway in OS (33–35),
indicating that microRNAs may be the upstream molecules involved in
FOXN3 signaling.
In summary, the present study revealed that FOXN3
may serve as a tumor suppressor in OS and that FOXN3 may be
considered as a novel prognostic predictor and therapeutic target
for the treatment of OS.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
WX, XZ and WZ conceived and designed the work. WX,
LM, and ZW constructed expression plasmids, prepared proteins and
performed experiments. WX and LM analyzed the data. WX, WZ and XZ
wrote the paper. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was approved by the Research
Ethics Committee of Yidu Central Hospital of Weifang (LK2017012,
Weifang, China). All patients had provided written informed
consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bielack S, Carrle D and Casali PG; ESMO
Guidelines Working Group, : Osteosarcoma: ESMO clinical
recommendations for diagnosis, treatment and follow-up. Ann Oncol.
4 Suppl 20:S137–S139. 2009.
|
|
2
|
Damron TA, Ward WG and Stewart A:
Osteosarcoma, chondrosarcoma, and Ewing's sarcoma: National cancer
data base report. Clin Orthop Relat Res. 459:40–47. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ottaviani G and Jaffe N: The epidemiology
of osteosarcoma. Cancer Treat Res. 152:3–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tan ML, Choong PF and Dass CR:
Osteosarcoma: Conventional treatment vs. gene therapy. Cancer Biol
Ther. 8:106–117. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bacci G, Briccoli A, Rocca M, Ferrari S,
Donati D, Longhi A, Bertoni F, Bacchini P, Giacomini S, Forni C, et
al: Neoadjuvant chemotherapy for osteosarcoma of the extremities
with metastases at presentation: Recent experience at the Rizzoli
Institute in 57 patients treated with cisplatin, doxorubicin, and a
high dose of methotrexate and ifosfamide. Ann Oncol. 14:1126–1134.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rainusso N, Wang LL and Yustein JT: The
adolescent and young adult with cancer: State of the art-bone
tumors. Curr Oncol Rep. 15:296–307. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bielack SS, Kempf-Bielack B, Delling G,
Exner GU, Flege S, Helmke K, Kotz R, Salzer-Kuntschik M, Werner M,
Winkelmann W, et al: Prognostic factors in high-grade osteosarcoma
of the extremities or trunk: An analysis of 1,702 patients treated
on neoadjuvant cooperative osteosarcoma study group protocols. J
Clin Oncol. 20:776–790. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Scott KL and Plon SE: CHES1/FOXN3
interacts with Ski-interacting protein and acts as a
transcriptional repressor. Gene. 359:119–126. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sun J, Li H, Huo Q, Cui M, Ge C, Zhao F,
Tian H, Chen T, Yao M and Li J: The transcription factor FOXN3
inhibits cell proliferation by downregulating E2F5 expression in
hepatocellular carcinoma cells. Oncotarget. 7:43534–43545.
2016.PubMed/NCBI
|
|
10
|
Balciunaite G, Keller MP, Balciunaite E,
Piali L, Zuklys S, Mathieu YD, Gill J, Boyd R, Sussman DJ and
Holländer GA: Wnt glycoproteins regulate the expression of FoxN1,
the gene defective in nude mice. Nat Immunol. 3:1102–1108. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li C, Lusis AJ, Sparkes R, Tran SM and
Gaynor R: Characterization and chromosomal mapping of the gene
encoding the cellular DNA binding protein HTLF. Genomics.
13:658–664. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chang JT, Wang HM, Chang KW, Chen WH, Wen
MC, Hsu YM, Yung BY, Chen IH, Liao CT, Hsieh LL and Cheng AJ:
Identification of differentially expressed genes in oral squamous
cell carcinoma (OSCC): Overexpression of NPM, CDK1 and NDRG1 and
underexpression of CHES1. Int J Cancer. 114:942–949. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Markowski J, Tyszkiewicz T, Jarzab M,
Oczko-Wojciechowska M, Gierek T, Witkowska M, Paluch J, Kowalska M,
Wygoda Z, Lange D and Jarzab B: Metal-proteinase ADAM12, kinesin 14
and checkpoint suppressor 1 as new molecular markers of laryngeal
carcinoma. Eur Arch Otorhinolaryngol. 266:1501–1507. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li S, Mo Z, Yang X, Price SM, Shen MM and
Xiang M: Foxn4 controls the genesis of amacrine and horizontal
cells by retinal progenitors. Neuron. 43:795–807. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Katoh M: Identification and
characterization of human FOXN5 and rat Foxn5 genes in silico. Int
J Oncol. 24:1339–1344. 2004.PubMed/NCBI
|
|
16
|
Wang X, He B, Gao Y and Li Y: FOXR2
contributes to cell proliferation and malignancy in human
hepatocellular carcinoma. Tumour Biol. 37:10459–10467. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huot G, Vernier M, Bourdeau V, Doucet L,
Saint-Germain E, Gaumont-Leclerc MF, Moro A and Ferbeyre G:
CHES1/FOXN3 regulates cell proliferation by repressing PIM2 and
protein biosynthesis. Mol Biol Cell. 25:554–565. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gertler AA and Cohen HY: SIRT6, a protein
with many faces. Biogerontology. 14:629–639. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim HS, Xiao C, Wang RH, Lahusen T, Xu X,
Vassilopoulos A, Vazquez-Ortiz G, Jeong WI, Park O, Ki SH, et al:
Hepatic-specific disruption of SIRT6 in mice results in fatty liver
formation due to enhanced glycolysis and triglyceride synthesis.
Cell Metab. 12:224–236. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lin H, Hao Y, Zhao Z and Tong Y: Sirtuin 6
contributes to migration and invasion of osteosarcoma cells via the
ERK1/2/MMP9 pathway. FEBS Open Bio. 7:1291–1301. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Baumhoer D: Pathogenesis and genetics of
osteosarcoma: Current concepts and developments. Der Pathologe.
39:139–145. 2018.(In German). View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Descamps FJ, Martens E and Opdenakker G:
Analysis of gelatinases in complex biological fluids and tissue
extracts. Lab Invest. 82:1607–1608. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Padala C, Tupurani MA, Puranam K, Gantala
S, Shyamala N, Kondapalli MS, Gundapaneni KK, Mudigonda S, Galimudi
RK, Kupsal K, et al: Synergistic effect of collagenase-1 (MMP1),
stromelysin-1 (MMP3) and gelatinase-B (MMP9) gene polymorphisms in
breast cancer. PLoS One. 12:e01844482017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cai X, Zhu H and Li Y: PKCzeta, MMP2 and
MMP9 expression in lung adenocarcinoma and association with a
metastatic phenotype. Mol Med Rep. 16:8301–8306. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jia ZH, Jia Y, Guo FJ, Chen J, Zhang XW
and Cui MH: Phosphorylation of STAT3 at Tyr705 regulates MMP-9
production in epithelial ovarian cancer. PLoS One. 12:e01836222017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bai L, Lin G, Sun L, Liu Y, Huang X, Cao
C, Guo Y and Xie C: Upregulation of SIRT6 predicts poor prognosis
and promotes metastasis of non-small cell lung cancer via the
ERK1/2/MMP9 pathway. Oncotarget. 7:40377–40386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Schuff M, Rossner A, Wacker SA, Donow C,
Gessert S and Knochel W: FoxN3 is required for craniofacial and eye
development of Xenopus laevis. Dev Dyn. 236:226–239. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Samaan G, Yugo D, Rajagopalan S, Wall J,
Donnell R, Goldowitz D, Gopalakrishnan R and Venkatachalam S: Foxn3
is essential for craniofacial development in mice and a putative
candidate involved in human congenital craniofacial defects.
Biochem Biophys Res Commun. 400:60–65. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nagel S, Meyer C, Kaufmann M, Drexler HG
and MacLeod RA: Deregulated FOX genes in Hodgkin lymphoma. Genes
Chromosomes Cancer. 53:917–933. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cai Y, Mohseny AB, Karperien M, Hogendoorn
PC, Zhou G and Cleton-Jansen AM: Inactive Wnt/beta-catenin pathway
in conventional high-grade osteosarcoma. J Pathol. 220:24–33. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hendrix ND, Wu R, Kuick R, Schwartz DR,
Fearon ER and Cho KR: Fibroblast growth factor 9 has oncogenic
activity and is a downstream target of Wnt signaling in ovarian
endometrioid adenocarcinomas. Cancer Res. 66:1354–1362. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Du Z, Li F, Wang L, Huang H and Xu S:
Regulatory effects of microRNA184 on osteosarcoma via the
Wnt/β-catenin signaling pathway. Mol Med Rep. 18:1917–1924.
2018.PubMed/NCBI
|
|
34
|
Zhao X, Sun S, Xu J, Luo Y, Xin Y and Wang
Y: MicroRNA-152 inhibits cell proliferation of osteosarcoma by
directly targeting Wnt/β-catenin signaling pathway in a
DKK1-dependent manner. Oncol Rep. 40:767–774. 2018.PubMed/NCBI
|
|
35
|
Yu M, Guo D, Cao Z, Xiao L and Wang G:
Inhibitory effect of microRNA-107 on osteosarcoma malignancy
through regulation of Wnt/β-catenin signaling in vitro. Cancer
Invest. 36:175–184. 2018. View Article : Google Scholar : PubMed/NCBI
|