Introduction
Cervical cancer is the fourth most common cause of
fatality that threatens women's health excessively in developing
counties (1). Cervical cancer cases
were associated with 265,700 fatalities in 2012 (2). Approximately half a million new cases
occur worldwide each year (2).
Although 80% of early-stage cervical cancer cases can be treated
with surgery, radiotherapy or chemotherapy, there is still a large
number of advanced-stage patients with poor prognoses (3). The association between human
papillomaviruses (HPV) and cervical cancer has been strongly
verified (4); however, the specific
mechanisms remain to be fully elucidated. Therefore, further
studies on the underlying mechanism of tumor initiation and
development are necessary.
Non-coding (nc)RNA is a functional RNA molecule that
is transcribed from DNA but not translated into proteins (5). Micro(mi)RNAs are a vital component of
the endogenous ncRNA family, which are ~19–25 nucleotides in
length. The majority of miRNAs are highly conserved in sequence and
are involved in multiple cellular functions via the
post-transcriptional regulation of gene transcription (6). According to previous studies, several
miRNAs (7–11) have been consistently reported to be
involved in the development of cervical cancer, which suggests that
miRNAs are highly correlated with the pathogenesis of cervical
cancer.
Competing endogenous RNAs (ceRNAs) are transcripts
with the same miRNA response element (MRE) that act as miRNA
sequestering molecules and compete to bind to miRNAs to regulate
their target genes, thereby affecting the biological behavior of
tumors (12). Circular (circ)RNAs
are a novel class of the ceRNA and a distinctive type of ncRNA that
were identified in plant viruses in 1976 (13). Unlike traditional linear RNA,
circRNAs are structured as a distinct closed loop. Therefore,
circRNAs are not easily degraded and stably expressed in various
organisms (14). Following the
binding to miRNA by MREs, circRNAs can also serve an important role
in the occurrence and development of tumors as tumor suppressors or
proto-oncogenes (15). Recently,
various studies have indicated that circRNAs are involved in the
initiation and development of multiple diseases, including gastric
cancer (16), osteoarthritis
(17) and hepatocellular carcinoma
(18).
As the regulatory gene for miRNAs, more studies have
suggested that circRNAs may serve key roles in the carcinogenesis
of multiple types of cancer (16,18,19).
However, research regarding cervical cancer is limited. Wang et
al (20) revealed that
hsa-circ-0101996 combined with hsa-circ-0101119 in peripheral whole
blood was identified as the potential biomarkers for human cervical
squamous cell cancer. Gao et al (21) reported that hsa-circ-0018289 was
upregulated in cervical cancer and promotes proliferation,
migration and invasion of tumor cells. Furthermore, Ma et al
(22) indicated that activated
has-circ-000284 promotes cell proliferation and invasion in
cervical cancer. This evidence supports that circRNA is likely to
participate in the development of cervical cancer, and probably by
indirectly regulating the expression of target genes through
affecting miRNAs.
The molecular mechanism underlying the role of
ncRNAs in the carcinogenesis and progression of cervical cancer
remains unknown. Therefore, in order to further understand the
potential role of ncRNAs in cervical cancer, the differentially
expressed RNAs (including circRNA, miRNA and mRNA) were identified
by microarray and databases and a regulatory circRNA-miRNA-mRNA
network was constructed (Fig. 1).
The present study may illuminate the underlying mechanisms of
cervical cancer pathogenesis and provide novel biomarkers and
targets for cervical cancer.
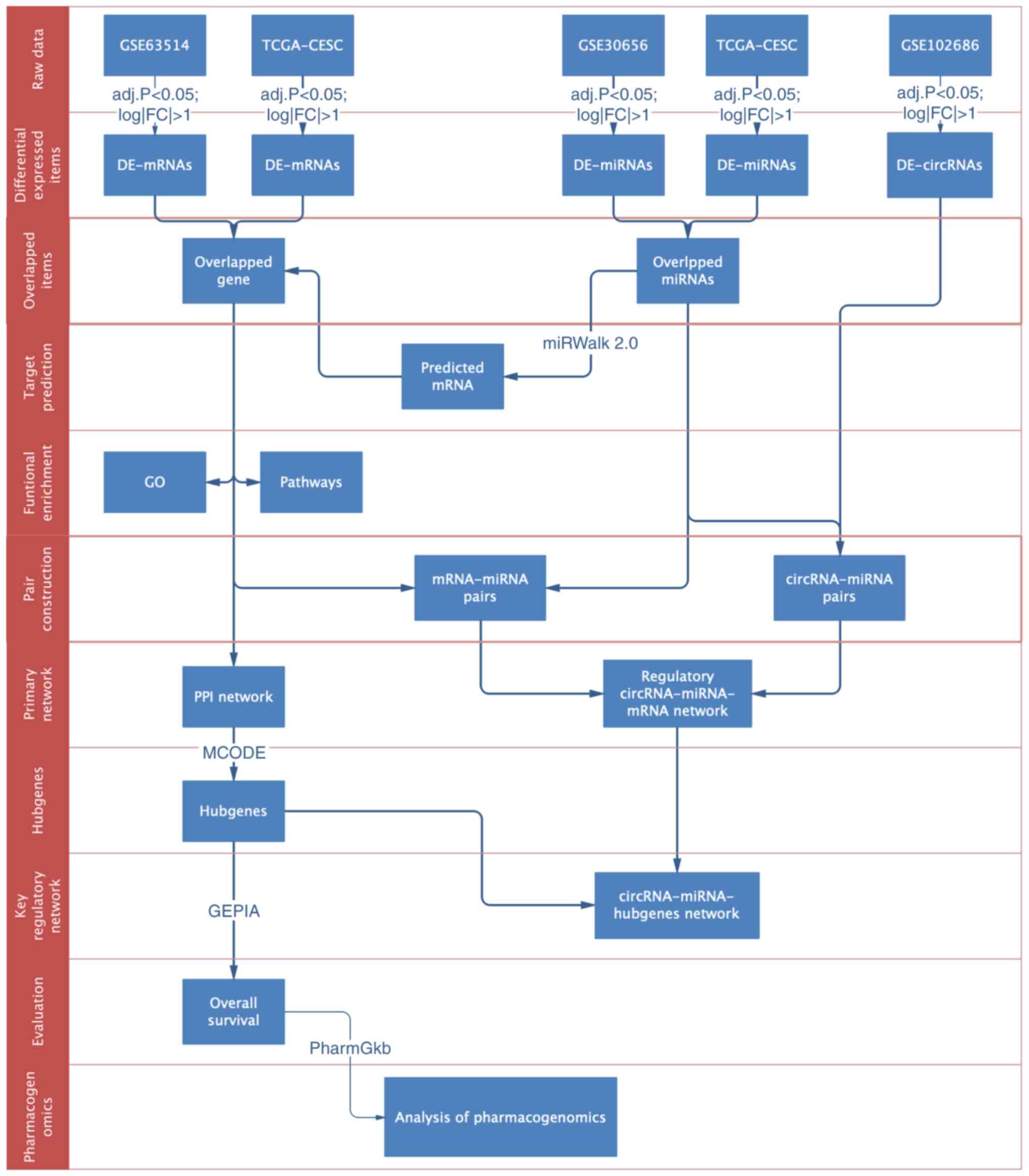 | Figure 1.Flowchart of the construction and
clinical evaluation of circRNA-miRNA-mRNA network. TCGA, The Cancer
Genome Atlas; CESC, cervical squamous cell carcinoma and
endocervical adenocarcinoma; adj.p, adjusted P-value; FC,
fold-change; DE, differentially expressed; GO, Gene Ontology; PPI,
protein-protein interaction network; GEPIA, the database of Gene
Expression Profiling Interactive Analysis; circRNA, circular RNA;
miRNA, microRNA. |
Materials and methods
Raw data
Gene Expression Omnibus (GEO; http://www.ncbi.nlm.nih.gov/geo/) is an international
public repository for high-throughput microarray and sequence-based
data (23). GSE102686 circRNA
microarray, GSE30656 miRNA profiles and GSE63514 mRNA datasets for
cervical cancer were downloaded from GEO, respectively. The
fundamental information for these three profiles is summarized in
Table I. In addition, the miRNA and
mRNA expression datasets for cervical squamous cell carcinoma and
endocervical adenocarcinoma (CESC), which contained 309 cervical
cancer samples with 3 normal samples, were also downloaded from The
Cancer Genome Atlas (TCGA) by cBioPortal (http://www.cbioportal.org/) (24).
 | Table I.Basic information of the three
microarray datasets from Gene Expression Omnibus. |
Table I.
Basic information of the three
microarray datasets from Gene Expression Omnibus.
| Profile | RNA type | Platform | Organism | Experiment
type | Sample size
(T/N) | Region | Year |
|---|
| GSE102686 | circRNA | GPL19978 | Homo
sapiens | Non-coding RNA
profiling by array | 5/5 | China | 2017 |
| GSE30656 | miRNA | GPL6955 | Homo
sapiens | Non-coding RNA
profiling by array | 19/10 | Netherlands | 2012 |
| GSE63514 | mRNA | GPL570 | Homo
sapiens | Expression
profiling by array | 28/24 | USA | 2015 |
Identification of differentially
expressed (DE)-miRNAs, DE-circRNAs and DE-mRNAs
The expression difference between the normal and
cervical cancer groups was utilized to determine the DE-circRNAs
DE-miRNAs and DE-mRNAs. The adjusted P-value (adj.p) and the
absolute log value of fold-change (log|FC|) were calculated using R
software with limma package (25).
The criteria of adj.p <0.05 and log|FC| >1 were adopted to
select the DE-circRNAs, DE-miRNAs and DE-mRNAs.
Construction of miRNA-mRNA pairs
Targeted mRNAs of the collected miRNAs were
predicted using miRWalk (Version 2.0; http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/)
(26), which included 12 databases
(Microt4, miRWalk, mir-bridge, miRanda, miRDB, miRMap, Pictar2,
PITA, miRNAMap, RNAhybrid, RNA22, and TargetScan). To increase the
accuracy of prediction, the targeted genes were select using the
criteria as follows: i) The targeted genes should be predicted by
TargetScan (27) and miRanda
(28); and ii) the targeted genes
should be overlapped in at least 8/12 databases. The selected
targeted mRNAs were merged with DE-mRNAs of CESC and GSE63514. The
overlapped gene sets were analyzed with UpSetR (29) and Venn Plot, and the pairs of miRNAs
and mRNAs were subsequently constructed.
Construction of circRNA-miRNA
pairs
To further predict the target circRNAs, the
sequences of the candidate DE-miRNAs and DE-circRNAs were
downloaded from miRbase (30) and
GSE102686, respectively. Following this, the miRNA targets of
circRNAs were predicted, and the minimum free energy of
circRNA-miRNA duplexes was calculated using the RNAhybrid program
(https://bibiserv2.cebitec.uni-bielefeld.de/rnahybrid)
(31). miRNA target binding sites
on the whole circRNA sequences were predicted. To obtain
high-quality circRNAs acting as miRNA targets and distinguish those
circRNAs acting as miRNA decoys, the circRNAs that had perfect
nucleotide pairing between the 2nd and 8th positions of the 5′ end
of miRNA sequences were selected (32) and the circRNA-miRNA pairs were
identified simultaneously.
Reconstruction of the
circRNA-miRNA-mRNA network
The preliminary circRNA-miRNA-mRNA network was
reconstructed by combining the pairs of miRNA-mRNA and
miRNA-circRNA. The nodes that could not form a circRNA-miRNA-mRNA
axis were removed. The circRNA-miRNA-mRNA network was visualized
using Cytoscape software (version 3.6.1) (33).
Gene Ontology (GO) and pathway
enrichment analysis
GO and pathway enrichment analyses were performed
using The Database for Annotation, Visualization and Integrated
Discovery (https://david.ncifcrf.gov/) (34) on the differentially expressed genes
(DEGs) in the preliminary circRNA-miRNA-mRNA network. The
significantly enriched biological items for biological process
(BP), cellular components (CC), and molecular functions (MF) were
identified with P<0.05. Pathways with P<0.05 were considered
as the significantly enriched pathways. The significant GO items
and pathways were visualized using Goplot package (35).
Protein-protein interaction (PPI)
network analysis
The Search Tool for the Retrieval of Interacting
Genes/Proteins (STRING) database (https://string-db.org/) (36) was used to predict the association
between target genes in regulatory network analysis. To obtain more
accurate results, nodes with interaction score <0.7 and that
were not connected to the major network were removed. The Molecular
Complex Detection (MCODE) (37)
plug-in in Cytoscape was used to analyze PPI network modules, and
MCODE scores >5 with >5 nodes were set as cut-off criteria
with the default parameters (Degree cut-off ≥2, node score cut-off
≥0.2, K-core ≥2 and max depth = 100). The genes in the cluster were
considered hubgenes.
Reconstruction of the
circRNA-miRNA-hubgene network
In order to identify the association between
DE-circRNAs, DE-miRNAs and hubgenes, the aforementioned hubgenes
were mapped into the preliminary circRNA-miRNA-mRNA network and the
relevant DE-circRNAs and DE-miRNAs were also extracted.
Subsequently, the subnetwork that was identified as the
circRNA-miRNA-hubgene network was identified. The structure of
circRNAs and the secondary stem-loop structure of miRNAs were also
predicted by the cancer-specific circRNA database (CSCD) (38) and Vienna RNA (39), respectively.
Evaluation of overall survival (OS)
for hubgenes
To identify the effect of the hubgenes on survival,
the Gene Expression Profiling Interactive Analysis (GEPIA) database
(40) was utilized to explore the
association between these hubgenes and OS. The genes with P<0.05
were considered as critical genes.
Pharmacogenomics analysis for critical
genes
Critical genes were analyzed using the database of
PharmGkb for the potential single nucleotide polymorphisms (SNPs)
and applicable medications, as well as the pharmacogenomics
pathway. Specifically, information on clinical and SNP annotations
for the critical genes were extracted, including genes names, SNPs,
associated medications, efficiency, significance, P-values,
genotype association with medications, references and pathways.
Results
Identification of DE-circRNAs,
DE-miRNAs and DE-mRNAs
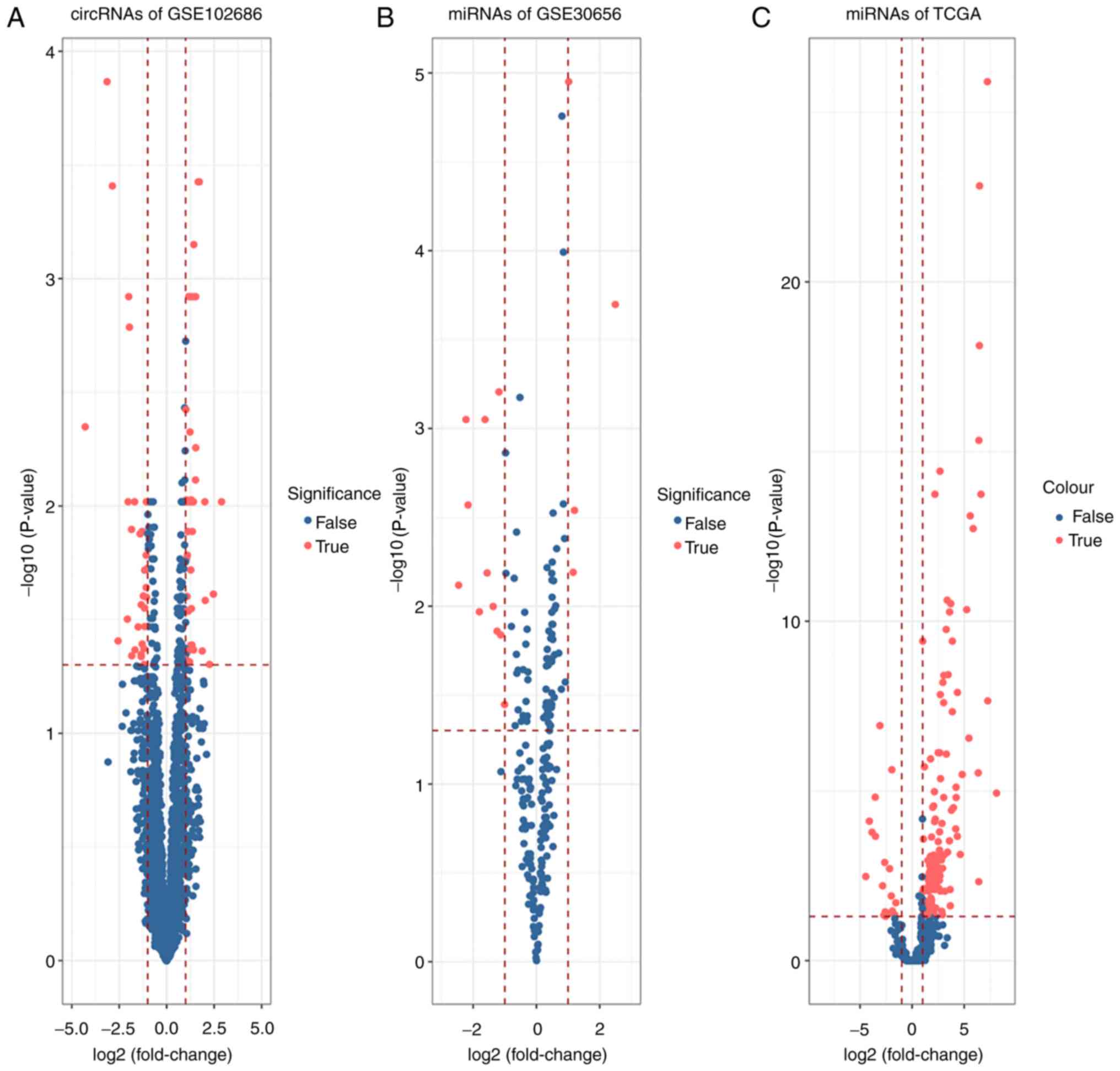
With the criteria of adj.p <0.05 and log|FC|
>1 a DE-circRNA dataset consisting of 75 DE-circRNAs, with 43
upregulated and 32 downregulated circRNAs, was identified from
GSE102686 (Fig. 2A). A DE-miRNAs
dataset, which consisted of 16 and 156 DE-miRNAs, was extracted
from GSE30656 and TCGA. By merging these two screening results, 5
miRNAs (hsa-miR-21, hsa-miR-99a, hsa-miR-106b, hsa-miR-15b and
hsa-miR-203) were obtained. hsa-miR-106b, hsa-miR-15b and
hsa-miR-21 were upregulated whereas hsa-miR-99a was downregulated
in CESC and GSE30656. However, hsa-miR-203 was upregulated in CESC
but downregulated in GSE30656 (Fig. 2B
and C). A DE-mRNA dataset, which consisted of 5,321 and 4,076
DE-mRNAs from GSE63514 and CESC, was determined (Fig. 2D and E). In addition, 3,400 targeted
genes predicted by miRWalk with 12 databases were identified. The
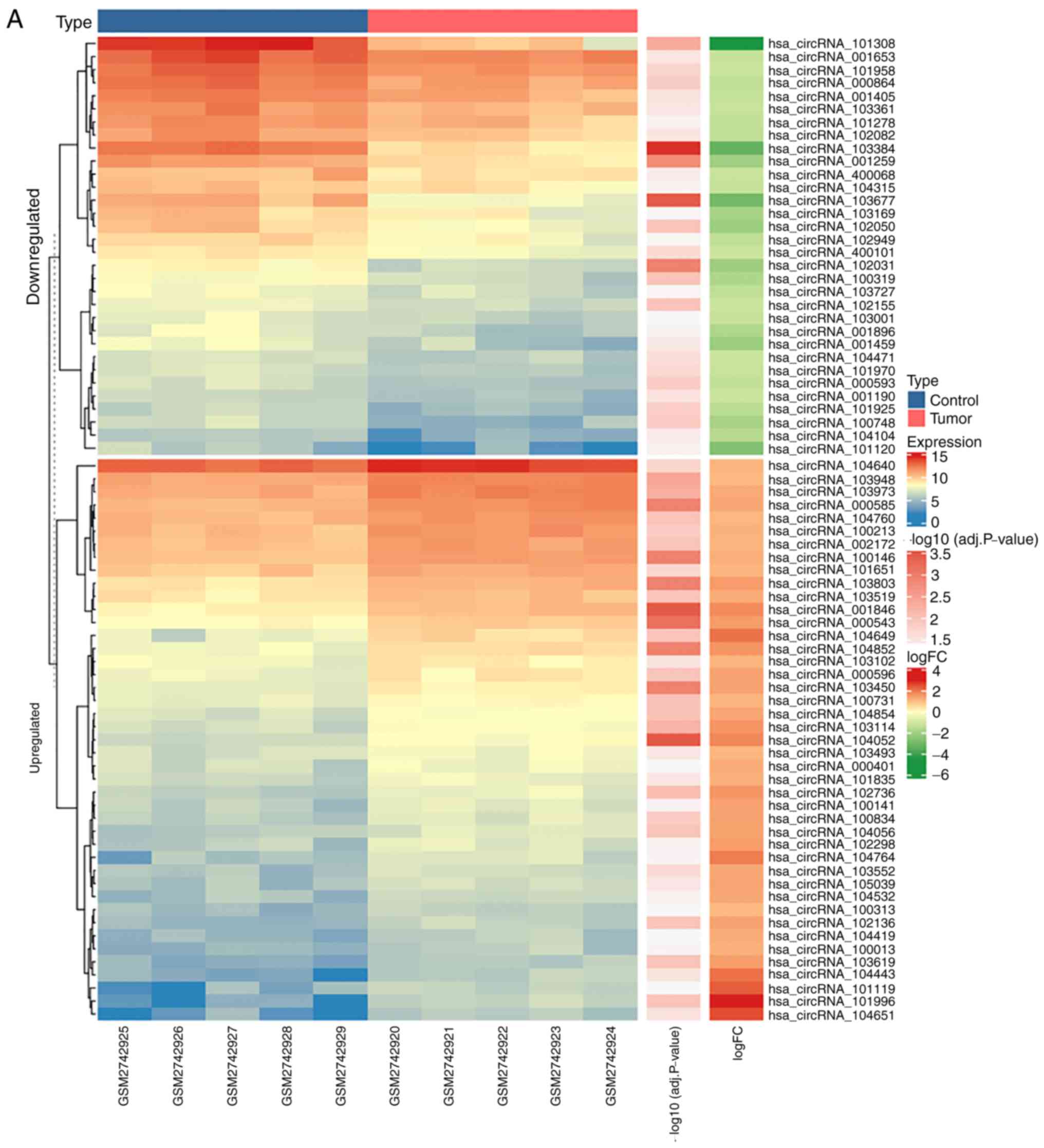
expression of the 75 DE-circRNAs is indicated in Fig. 3A, and the expression of 5 miRNAs in
GSE30656 is indicated in Fig. 3B.
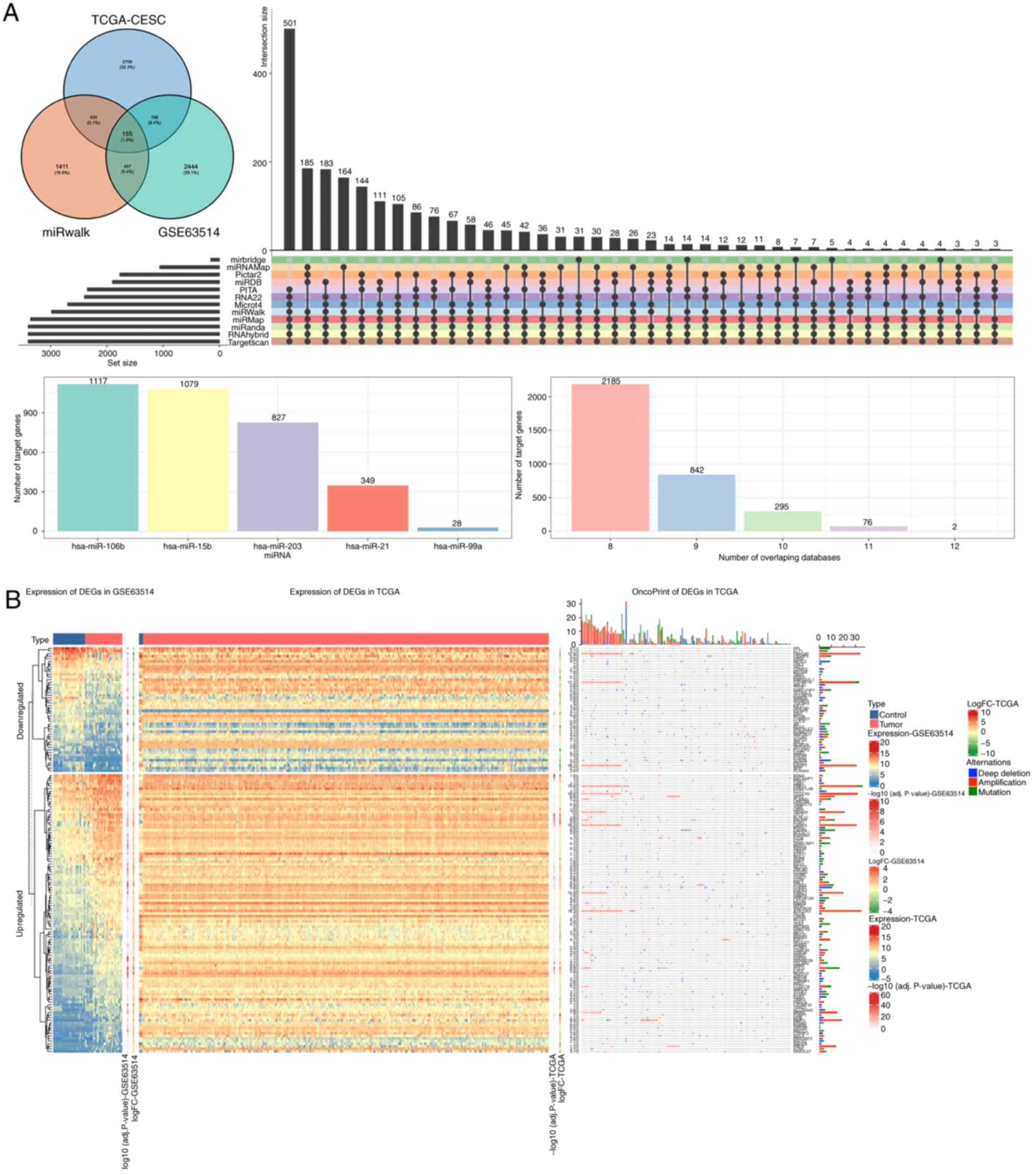
After integrating the DE-mRNAs from GSE63514 and CESC with the
targeted genes from miRWalk, 155 DE-mRNAs were identified (Fig. 4A). The expression and oncoprint of
these 155 genes in GSE63514 and TCGA is indicated in Fig. 4B.
Construction of miRNA-mRNA pairs
By merging the DE-mRNAs of GSE63514 and TCGA with
the targeted mRNAs of 5 DE-miRNAs from miRWalk, 266 miRNA-mRNA
pairs were selected. Specifically, 72 mRNAs of hsa-miR-15b, 16
mRNAs of hsa-miR-21, 2 mRNAs of hsa-miR-99a, 78 mRNAs of
hsa-miR-106b and 58 mRNAs of hsa-miR-203 were obtained.
Construction of circRNA-miRNA
pairs
Using RNAhybrid with the criterion that the circRNAs
had perfect nucleotide pairing between the 2nd and 8th positions of
the 5′ end of miRNA sequences, 12 circRNA-miRNA pairs were
obtained.
Reconstruction of the
circRNA-miRNA-mRNA network
As indicated in Fig.
5A, by combining the pairs of miRNA-mRNA and miRNA-circRNA, a
preliminary circRNA-miRNA-mRNA network was constructed. The network
was composed of 11 circRNA nodes, 4 miRNA nodes, 153 mRNA nodes and
203 edges. The network presented an initial perception of the
association between the 11 DECs (hsa_circRNA_103102,
hsa_circRNA_002172, hsa_circRNA_101835, hsa_circRNA_102155,
hsa_circRNA_102050, hsa_circRNA_103384, hsa_circRNA_104315,
hsa_circRNA_400068, hsa_circRNA_000596, hsa_circRNA_101958 and
hsa_circRNA_103519), the 4 miRNAs (hsa-miR-21, hsa-miR-203,
hsa-miR-15b and hsa-miR-106b) and the 153 mRNAs.
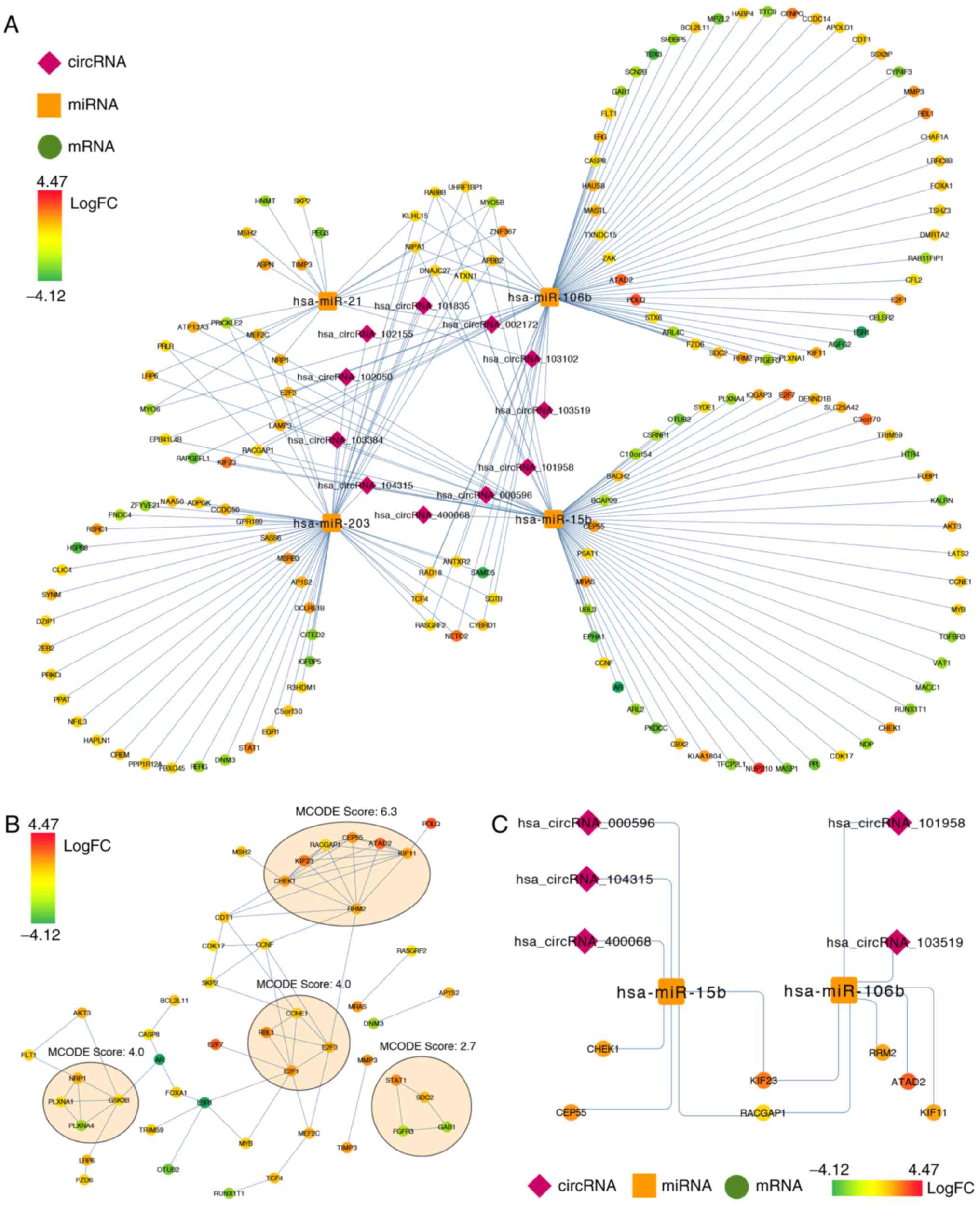 | Figure 5.Preliminary circRNA-miRNA-mRNA,
protein-protein interaction and circRNA-miRNA-hubgenes network for
the 11 DECs (hsa_circRNA_103102, hsa_circRNA_002172,
hsa_circRNA_101835, hsa_circRNA_102155, hsa_circRNA_102050,
hsa_circRNA_103384, hsa_circRNA_104315, hsa_circRNA_400068,
hsa_circRNA_000596, hsa_circRNA_101958 and hsa_circRNA_103519), the
4 miRNAs (hsa-miR-21, hsa-miR-203, hsa-miR-15b and hsa-miR-106b)
and the 155 mRNAs. (A) Preliminary circRNA-miRNA-mRNA network the
11 DECs, the 4 miRNAs and the 155 mRNAs. (B) Protein-protein
interaction network for the 155 overlapped genes. Each ellipses
represent the cluster identified by MCODE algorithm. (C) The
circRNA-miRNA-hubgenes network for the 5 circRNAs
(hsa_circRNA_000596, hsa_circRNA_104315, hsa_circRNA_400068,
hsa_circRNA_101958 and hsa_circRNA_103519), the 2 mRNAs
(hsa-miR-15b and hsa-miR-106b) and the 7 mRNAs (RRM2, CEP55, CHEK1,
KIF23, RACGAP1, ATAD2 and KIF11). circRNA, circular RNA; miRNA/miR,
microRNA; MCODE, Molecular Complex Detection. |
PPI network analysis
The STRING database was used to construct a PPI
network based on the 155 overlapped mRNAs. The original network
contained 47 nodes and 72 edges. By utilizing the algorithm of
MCODE, four clusters were identified (Fig. 5B). Using the criteria of MCODE
scores >5 and >5 nodes, one cluster was selected. There were
7 genes (RRM2, CEP55, CHEK1, KIF23, RACGAP1, ATAD2 and KIF11) in
this cluster, and these 7 genes were identified as hubgenes.
Construction of the
circRNA-miRNA-hubgenes network
By mapping the 7 hubgenes into the preliminary
circRNA-miRNA-mRNA network, and also extracting relevant circRNAs
and miRNAs, a subnetwork considered as circRNA-miRNA-hubgenes was
constructed. This subnetwork consisted of 5 circRNAs
(hsa_circRNA_000596, hsa_circRNA_104315, hsa_circRNA_400068,
hsa_circRNA_101958 and hsa_circRNA_103519), 2 mRNAs (hsa-miR-15b
and hsa-miR-106b) and 7 mRNAs (RRM2, CEP55, CHEK1, KIF23, RACGAP1,
ATAD2 and KIF11) with 14 edges (Fig.
5C). There were 22 circRNA-miRNA-mRNA regulatory axes
identified in the subnetwork (Table
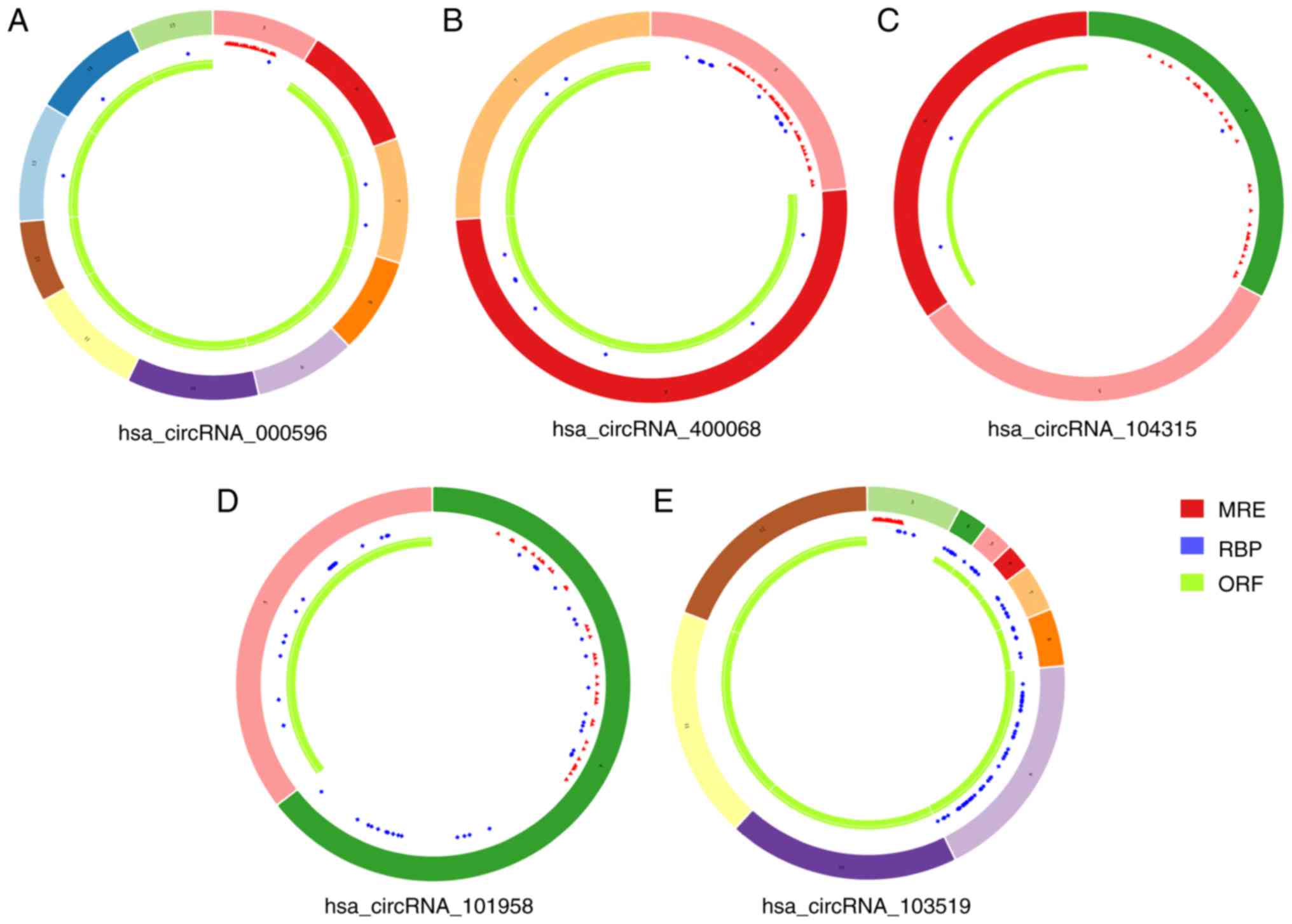
II). The structure and the primary characteristics of the
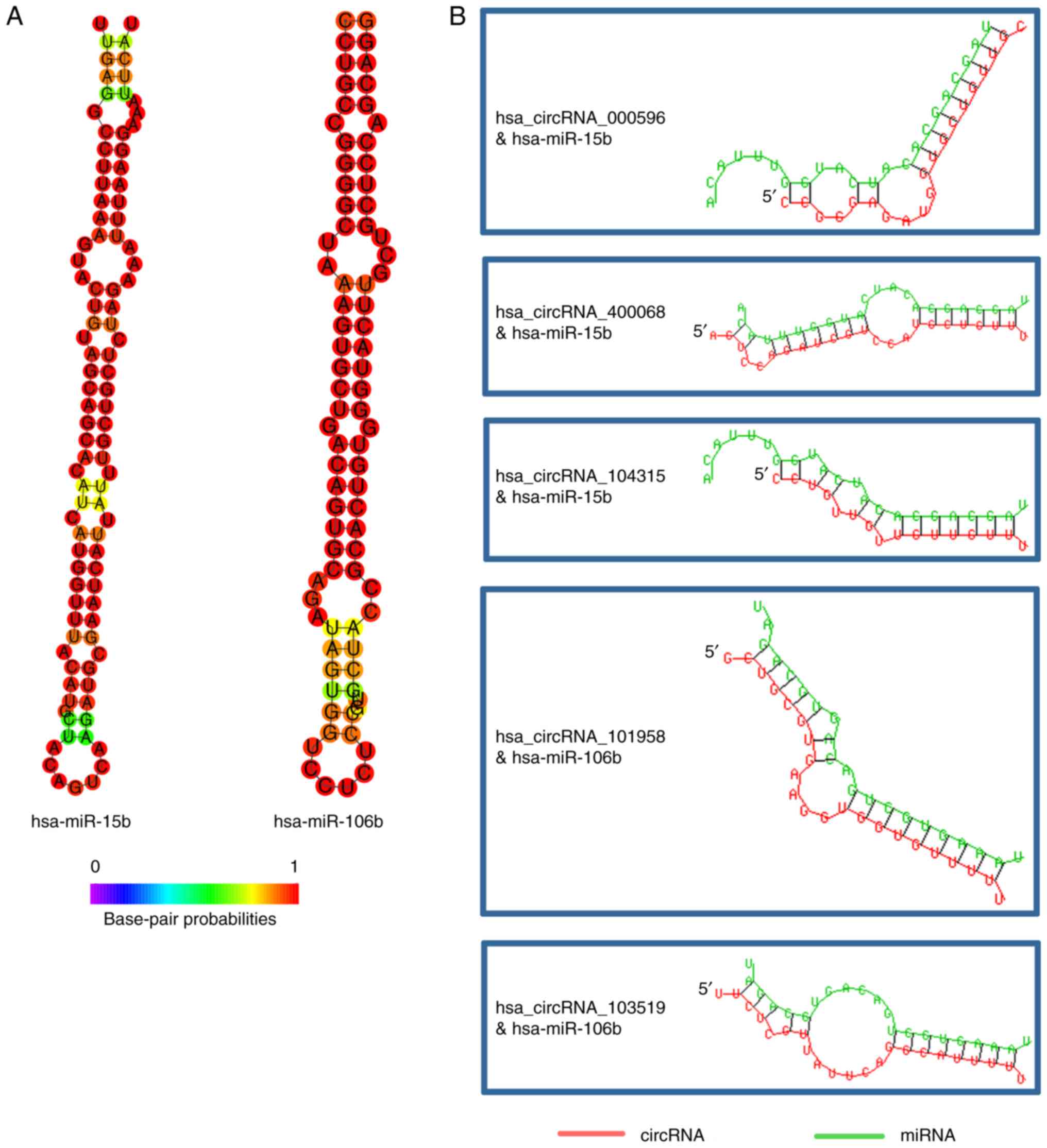
circRNAs in the subnetwork were indicated in Fig. 6 and Table III. In addition, the stem-loop
structure of hsa-miR-15b and hsa-miR-106b was revealed (Fig. 7A). The binding site to the circRNAs
was also indicated (Fig. 7B).
| Table II.Regulatory axis identified from the
circRNA-miRNA-hubgene network. |
Table II.
Regulatory axis identified from the
circRNA-miRNA-hubgene network.
| Regulatory
axis | circRNA | miRNA | Hubgene |
|---|
| 1 |
hsa_circRNA_000596 | hsa-miR-15b | CHEK1 |
| 2 |
hsa_circRNA_000596 | hsa-miR-15b | CEP55 |
| 3 |
hsa_circRNA_104315 | hsa-miR-15b | CHEK1 |
| 4 |
hsa_circRNA_104315 | hsa-miR-15b | CEP55 |
| 5 |
hsa_circRNA_400068 | hsa-miR-15b | CHEK1 |
| 6 |
hsa_circRNA_400068 | hsa-miR-15b | CEP55 |
| 7 |
hsa_circRNA_000596 | hsa-miR-15b | KIF23 |
| 8 |
hsa_circRNA_000596 | hsa-miR-15b | RACGAP1 |
| 9 |
hsa_circRNA_104315 | hsa-miR-15b | KIF23 |
| 10 |
hsa_circRNA_104315 | hsa-miR-15b | RACGAP1 |
| 11 |
hsa_circRNA_400068 | hsa-miR-15b | KIF23 |
| 12 |
hsa_circRNA_400068 | hsa-miR-15b | RACGAP1 |
| 13 |
hsa_circRNA_101958 | hsa-miR-106b | KIF23 |
| 14 |
hsa_circRNA_101958 | hsa-miR-106b | RACGAP1 |
| 15 |
hsa_circRNA_103519 | hsa-miR-106b | KIF23 |
| 16 |
hsa_circRNA_103519 | hsa-miR-106b | RACGAP1 |
| 17 |
hsa_circRNA_101958 | hsa-miR-106b | RRM2 |
| 18 |
hsa_circRNA_103519 | hsa-miR-106b | RRM2 |
| 19 |
hsa_circRNA_101958 | hsa-miR-106b | ATAD2 |
| 20 |
hsa_circRNA_103519 | hsa-miR-106b | ATAD2 |
| 21 |
hsa_circRNA_101958 | hsa-miR-106b | KIF11 |
| 22 |
hsa_circRNA_103519 | hsa-miR-106b | KIF11 |
| Table III.Primary characteristics of the 5
circRNAs identified from the circRNA-miRNA-hubgene network. |
Table III.
Primary characteristics of the 5
circRNAs identified from the circRNA-miRNA-hubgene network.
| circRNA | Chromosome | Start position | End position | Strand | Location | Gene symbol | Regulation |
|---|
|
hsa_circRNA_000596 | Chr15 | 100589061 | 100739628 | + | Exon | ADAMTS17 | Upregulated |
|
hsa_circRNA_400068 | Chr22 | 20109884 | 20114047 | + | Exon | RANBP1 | Downregulated |
|
hsa_circRNA_104315 | Chr7 | 16298014 | 16330476 | − | Exon | ISPD | Downregulated |
|
hsa_circRNA_101958 | Chr17 | 4192524 | 4207021 | − | Exon | UBE2G1 | Downregulated |
|
hsa_circRNA_103519 | Chr3 | 179042885 | 179051613 | + | Exon | ZNF639 | Upregulated |
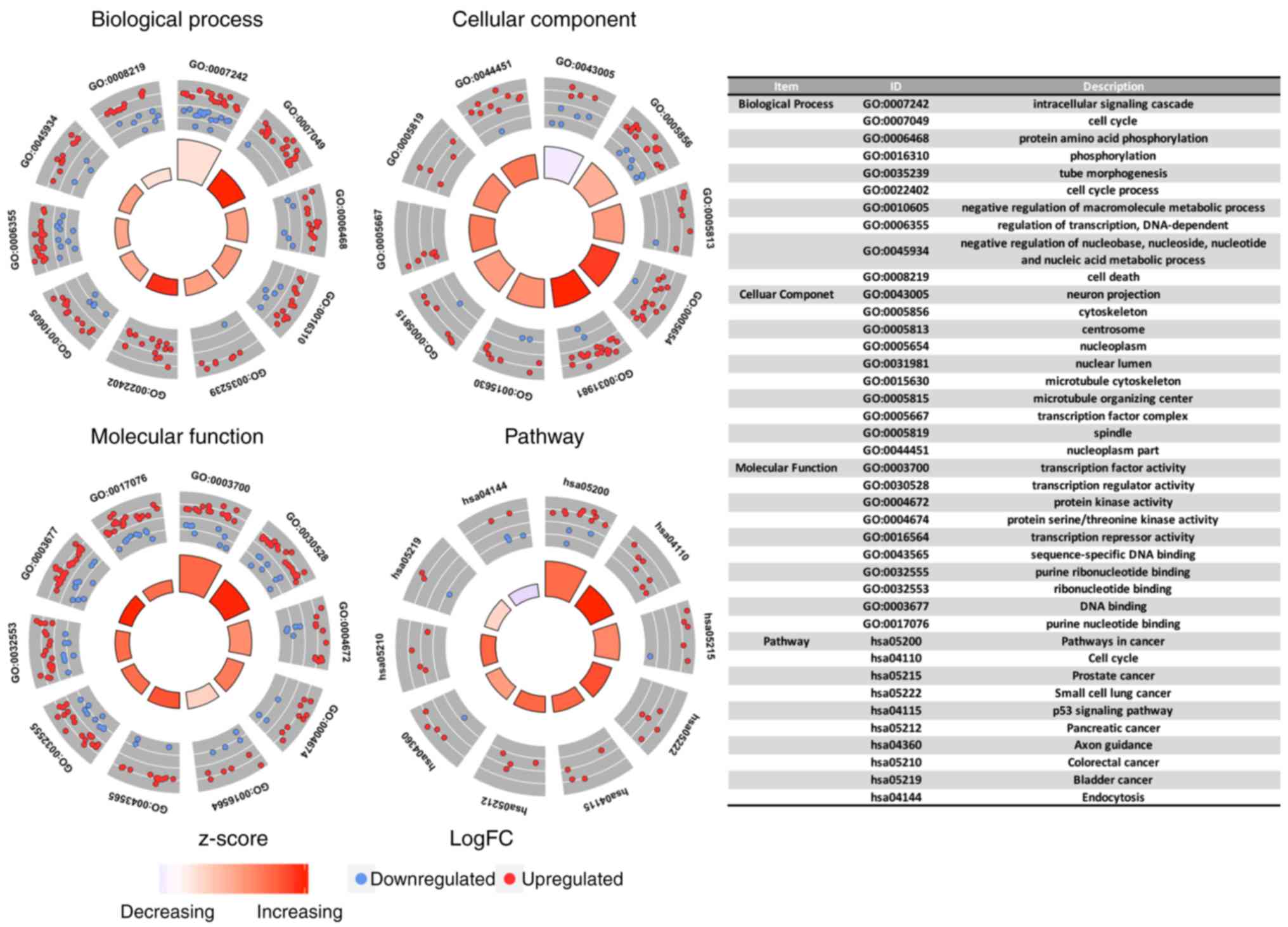
GO and pathway enrichment
analysis
The 155 overlapped genes were utilized for GO and
pathway enrichment analysis using DAVID. For GO analysis, when
considering BPs, the DEGs were enriched in intracellular signaling
cascade, cell cycle and protein amino acid phosphorylation. With
regards to CC, the top three enriched items were cell death, neuron
projection and cytoskeleton. In terms of MF, transcription factor
activity, transcription regulator activity and protein kinase
activity were enriched for the first three places. Pathway
enrichment analysis indicated that the top three significantly
enriched pathways were pathways in cancer, cell cycle and prostate
cancer (Fig. 8).
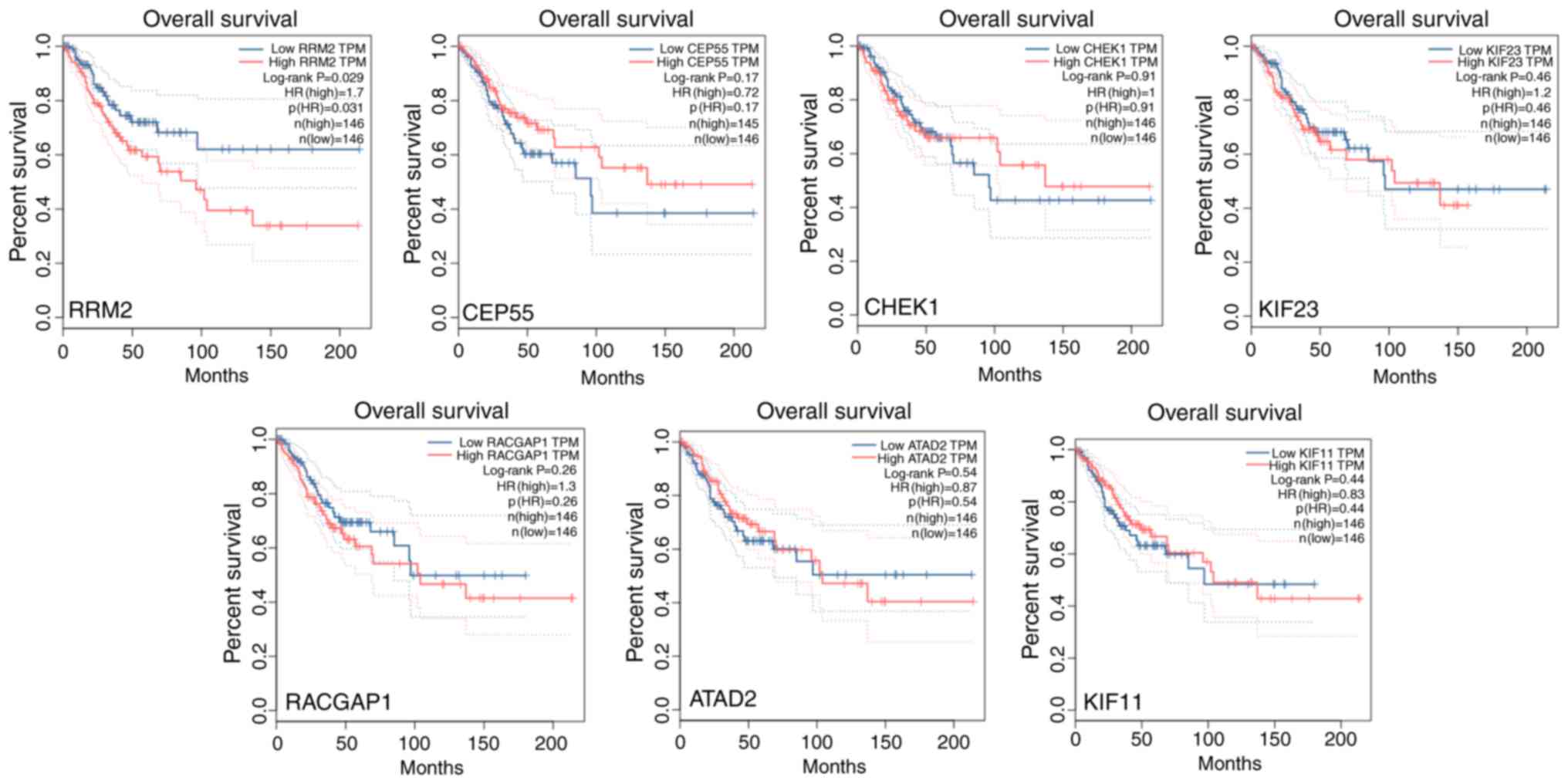
Evaluation of OS for hubgenes
GEPIAT was used to assess the OS for the 7 hubgenes.
Notably, higher expression of RRM2 revealed a significantly poorer
OS (hazard ratio = 1.7, P=0.029). However, no significant effect
was indicated for the remaining 6 hubgenes regarding OS (Fig. 9).
Pharmacogenomics analysis for
RRM2
As RRM2 significantly impacted OS, this gene was
selected for the analysis of pharmacogenomics. Using PharmGkb, the
SNPs of rs5030743 and rs1130609 were identified, which can be
treated with cladribine and cytarabine (Table IV). Notably, cladribine is used for
hairy cell leukemia, acute myeloid leukemia and chronic lymphocytic
leukemia (41–43) while cytarabine is a chemotherapeutic
drug utilized for various types of cancer, such as breast and lung
cancer (44,45). Cytarabine, similar to gemcitabine,
is activated and metabolized by deoxycytidine kinase and cytidine
deaminase (46). In addition, their
primary metabolite can also incorporate DNA into cells (47). Hence, the gemcitabine pathway was
selected to investigate the mechanism of action of cytarabine
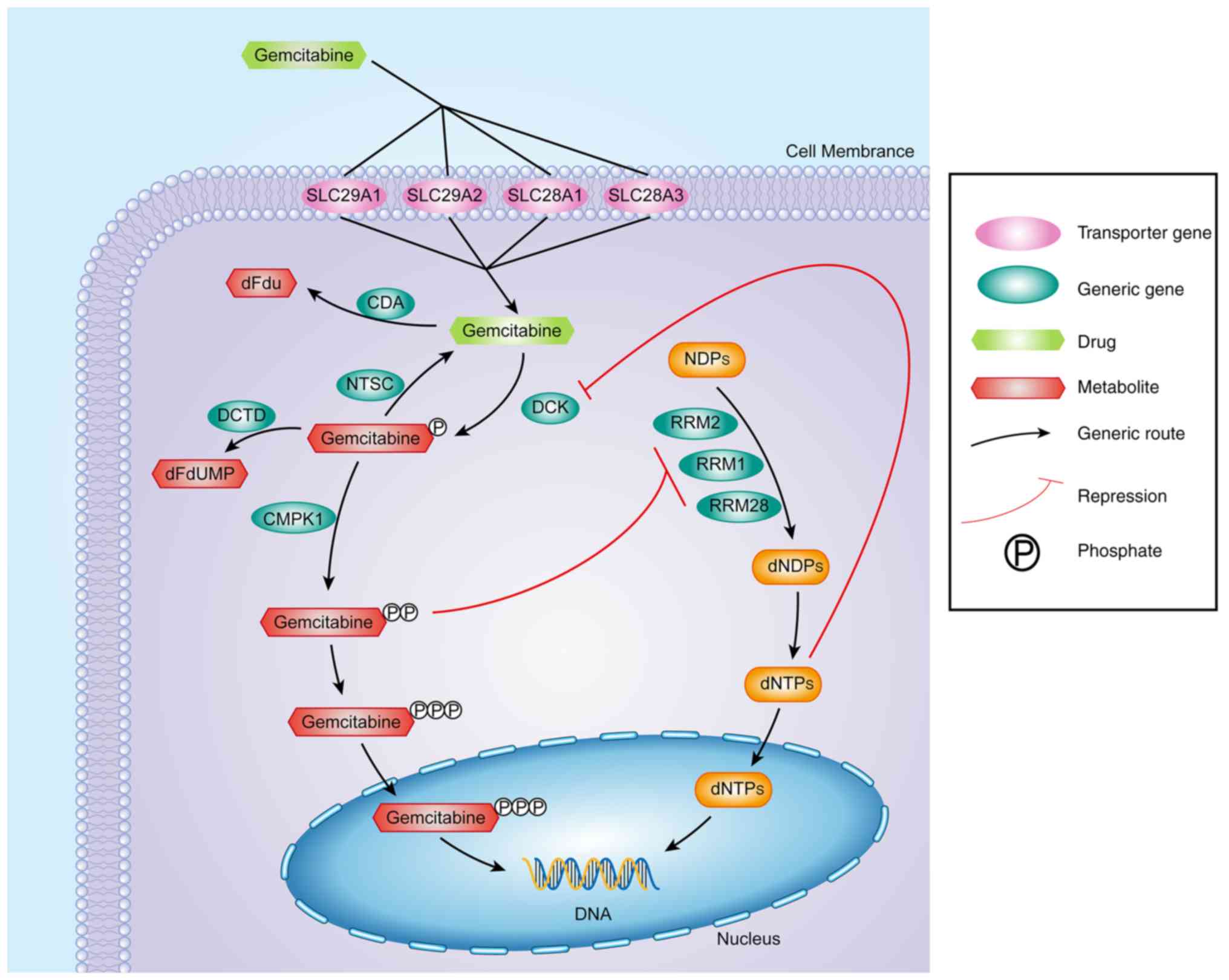
through RRM2. As indicated in Fig.
10, gemcitabine produces its therapeutic effect by acting on
the enzymes of RRM1, RRM2 and RRM2B, while the RRM1, RRM2 and RRM2B
genes also strictly regulate the activity of these enzymes.
| Table IV.Information of SNPs and clinical data
for RRM2. |
Table IV.
Information of SNPs and clinical data
for RRM2.
| Genes | SNP | Drug | Efficiency | Significant | P-value | Association | Reference | Pathways |
|---|
| RRM2 | rs5030743 | Cladribine;
Cytarabine | Effective | Yes | 0.013 | Genotypes CG + GG
is associated with decreased response to cladribine and cytarabine
in children with leukemia, myeloid, acute as compared to genotype
CC | PMID: 24024897 | Thiopurine pathway;
Fluoropyrimidine pathway; Gemcitabine pathway |
| RRM2 | rs1130609 | Cladribine;
Cytarabine | Efficacy | Yes | 0.020 | Genotypes GT + TT
is associated with decreased response to cladribine and cytarabine
in children with leukemia, myeloid, acute as compared to genotype
GG | PMID: 24024897 | Thiopurine pathway;
Fluoropyrimidine pathway; Gemcitabine pathway |
Discussion
During the past few decades, cervical cancer
research has developed rapidly, particularly with regard to the
discovery of the etiological factors. Although the key etiological
role of HPV in the development of cervical cancer has been reported
(48), the specific molecular
mechanism remains unclear. Recently, more data has suggested that
circRNAs participate in various biological processes (49–52).
Dysregulated circRNA expression has been observed in the
progression of complex diseases, including cervical cancer
(16,18,19,21).
circRNAs are a novel class of competing endogenous ncRNAs that are
widely expressed in eukaryotic cytoplasm (53). Unlike the RNAs formed by linear
splicing, circRNAs have a closed loop structure with no 5′ end cap
and 3′ end tail. The majority of known circRNAs are backspliced
from exons, which are not easily degraded by exonuclease and have
the feature of high abundance, structural stability and tissue
specificity (54). With the rapid
development of high-throughput RNA sequencing technology
accompanied with the extensive data analysis by bioinformatics,
circRNAs have been identified to possess the function of miRNA
sponges (54–56), gene transcription regulation
(54,57) and RNA-binding proteins (58–60) as
well as their translation (52,61,62).
In addition, studies have confirmed that circRNA serves an
important role in tumor cells, so may potentially serve as a novel
biomarker and therapeutic target for cancer therapy (63–65).
However, more circRNAs need to be uncovered. In the present study,
the circRNA microarray profile of GSE102686 for cervical cancer was
screened to identify DE-circRNAs. Using the criteria of FDR
<0.05 and log|FC| >1 calculated by R with limma package, 75
DE-circRNAs were selected for further analysis.
Previous studies have indicated that circRNA is a
type of high-efficiency ceRNA (66,67).
It can inhibit the binding of miRNAs to target genes and regulate
the expression level of target genes by exerting a miRNA
sequestering effect (68). To
determine whether the above 75 circRNAs function as ceRNAs in
cervical cancer, the targeted miRNAs with the sequence of
DE-circRNAs and DE-miRNAs were predicted with RNAhybrid.
Furthermore, 12 circRNA-miRNA pairs were identified. hsa-miR-106b,
hsa-miR-15b and hsa-miR-21 were identified to be upregulated in
CESC and GSE30656, and these results were consistent with previous
findings (69–73). Notably, hsa-miR-203 was upregulated
in CESC but downregulated in GSE30656. Previous studies have
suggested that the expression of miR-203 is typically downregulated
in cervical cancer tumors and cell lines (74–76).
This finding is in line with the expression profile of GSE30656 but
contradicts to TCGA. Reshmi and Pillai (77) also identified that miR-203 can be
overexpressed or underexpressed in cervical cancer cell lines.
Furthermore, Zhao et al (78) indicated that the expression level of
miR-203 in serum of patients with cervical cancer was significantly
upregulated; however, miR-203 downregulation was correlated with
lymph nodes metastasis. Hence, it was possible that upregulation
and downregulation of miR-203 could promote the progression of
cervical cancer but serves different functions. Upregulation of
miR-203 could also trigger the occurrence of cervical cancer
whereas downregulation may enhance the metastatic capacity of lymph
nodes. However, the specific mechanism requires further
investigation.
Once the 155 overlapping genes with the DE-circRNAs
and DE-miRNAs were collected, a circRNA-miRNA-mRNA regulatory
network was reconstructed. The 11 circRNAs could bind to
hsa-miR-21, hsa-miR-203, hsa-miR-106b or hsa-miR-15b as ceRNAs to
regulate the expression of 155 genes. The present results provide
evidence for the ceRNA regulatory mechanisms of 11 circRNAs in
cervical cancer. The GO and pathway enrichment analysis indicated
that 155 genes were involved in various important biological
functions and metabolic pathways associated with tumors, including
‘Pathways in cancer’, ‘Cell cycle’, ‘Prostate cancer’, ‘Small cell
lung cancer’, ‘p53 signaling pathway’ and ‘Pancreatic cancer’. It
also indicated that several type of cancer, such as prostate
cancer, lung cancer and pancreatic cancer, share the same pathways
with cervical cancer, and provided evidence for the research of
pan-cancer. To further understand the functional mechanism of the
ceRNA-miRNA-mRNA network, the PPI network was constructed and 7
hubgenes (RRM2, CEP55, CHEK1, KIF23, RACGAP1, ATAD2 and KIF11) were
identified from the PPI network. The crucial mechanisms of RRM2,
CEP55, CHEK1, ATAD2 and KIF11 in cervical cancer have been
previously studied (79–82). However, to the best of our
knowledge, KIF23 and RACGAP1 have not yet been investigated. The 22
circRNA-miRNA-mRNA axes, which were identified from the
circRNA-miRNA-hubgenes, revealed the competing regulatory
associations between the 5 circRNAs and the 2 mRNAs with the 7
hubgenes in cervical cancer. However, as the present study was an
in silico research, a further experiment of these 22
regulatory axes is required for validation.
The OS for the 7 hubgenes was evaluated in the
present study and it was revealed that RRM2 had a significant
effect on OS. Subsequently, RRM2 was investigated using the
database of PharmGkb. It was revealed that cytarabine, which is
similar to gemcitabine, may produce its therapeutic effect by
targeting to the SNPs of rs5030743 and rs1130609 of RRM2 through
the gemcitabine pathway. To date, gemcitabine has been indicated to
be feasible and effective on cervical cancer (83–85).
However, cytarabine has not yet been investigated in cervical
cancer, nor the rs5030743 and rs1130609 of RRM2. Although
cytarabine differs from gemcitabine in several important respects
(86), for instance, gemcitabine
differs structurally from cytarabine by a fluorine group
substituted at position 2′ on the furanose ring (87), gemcitabine (88–90)
and cytarabine (91,92) can be a treatment for breast cancer.
Hence, cytarabine may produce a similar effect to gemcitabine and
could be an optional chemotherapeutic drug for treating cervical
cancer by targeting rs5030743 and rs1130609 or other similar SNPs.
However, further study is necessary to validate the hypothesis.
To conclude, the present study constructed and
analyzed a circRNA-miRNA-mRNA network based on the ceRNA theory via
comprehensive bioinformatics analysis, which may provide some
evidence to future studies focused on the molecular mechanisms of
cervical cancer. The 5 circRNAs (hsa_circRNA_000596,
hsa_circRNA_104315, hsa_circRNA_400068, hsa_circRNA_101958 and
hsa_circRNA_103519) may function as ceRNAs to serve critical roles
in cervical cancer. In addition, cytarabine may produce a similar
effect to gemcitabine and could be an optional chemotherapeutic
drug for cervical cancer by targeting rs5030743 and rs1130609 or
other similar SNPs. However, the specific mechanism of action
should be confirmed by further study.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YY and YL established the concept of the present
study and drafted the paper. WW and KW performed the literature
research, data extraction and statistical analysis. WZ contributed
in the performing the literature research, data extraction and
analysis, and reviewed and approved the final version of the paper.
All authors have read and approved the manuscript.
Ethics approval and consent to
participate
The study protocol was approved by the Ethics
Committee of Zhongnan Hospital of Wuhan University. Informed
consent forms were signed by all participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Marth C, Landoni F, Mahner S, McCormack M,
Gonzalez-Martin A and Colombo N; ESMO Guidelines Committee, :
Cervical cancer: ESMO Clinical Practice Guidelines for diagnosis,
treatment and follow-up. Ann Oncol. 28 (Suppl 4):iv72–iv83. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
GuYu Z, YiMin Z, ChongDong L, GuangMing C,
Ran C and ZhenYu Z: Current status and future of targeted therapy
for patients with local advanced cervical cancer. Chinese J Pract
Gynecol Obstet. 34:1216–1220. 2018.
|
|
4
|
Fleming ND, Frumovitz M, Schmeler KM, dos
Reis R, Munsell MF, Eifel PJ, Soliman PT, Nick AM, Westin SN and
Ramirez PT: Significance of lymph node ratio in defining risk
category in node-positive early stage cervical cancer. Gynecol
Oncol. 136:48–53. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kapranov P, Cawley SE, Drenkow J,
Bekiranov S, Strausberg RL, Fodor SP and Gingeras TR: Large-scale
transcriptional activity in chromosomes 21 and 22. Science.
296:916–919. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kogo R, How C, Chaudary N, Bruce J, Shi W,
Hill RP, Zahedi P, Yip KW and Liu FF: The microRNA-218~Survivin
axis regulates migration, invasion, and lymph node metastasis in
cervical cancer. Oncotarget. 6:1090–1100. 2014.
|
|
8
|
Hou T, Ou J, Zhao X, Huang X, Huang Y and
Zhang Y: MicroRNA-196a promotes cervical cancer proliferation
through the regulation of FOXO1 and p27Kip1. Br J
Cancer. 110:1260–1268. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang Q, Qin J, Chen A, Zhou J, Liu J,
Cheng J, Qiu J and Zhang J: Downregulation of microRNA-145 is
associated with aggressive progression and poor prognosis in human
cervical cancer. Tumor Biol. 36:3703–3708. 2015. View Article : Google Scholar
|
|
10
|
Fang H, Shuang D, Yi Z, Sheng H and Liu Y:
Up-regulated microRNA-155 expression is associated with poor
prognosis in cervical cancer patients. Biomed Pharmacother.
83:64–69. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bumrungthai S, Ekalaksananan T, Evans MF,
Chopjitt P, Tangsiriwatthana T, Patarapadungkit N, Kleebkaow P,
Luanratanakorn S, Kongyingyoes B, Worawichawong S, et al:
Up-regulation of miR-21 is associated with cervicitis and human
papillomavirus infection in cervical tissues. PLoS One.
10:e01271092015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden RNA
language? Cell. 146:353–8. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sanger HL, Klotz G, Riesner D, Gross HJ
and Kleinschmidt AK: Viroids are single-stranded covalently closed
circular RNA molecules existing as highly base-paired rod-like
structures. Proc Natl Acad Sci USA. 73:3852–3856. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tian M, Chen R, Li T and Xiao B: Reduced
expression of circRNA hsa_circ_0003159 in gastric cancer and its
clinical significance. J Clin Lab Anal. 32:2018. View Article : Google Scholar
|
|
17
|
Liu Q, Zhang X, Hu X, Yuan L, Cheng J,
Jiang Y and Ao Y: Emerging roles of circRNA related to the
mechanical stress in human cartilage degradation of osteoarthritis.
Mol Ther Nucleic Acids. 7:223–230. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang XY, Huang ZL, Xu YH, Zheng Q, Chen
Z, Song W, Zhou J, Tang ZY and Huang XY: Comprehensive circular RNA
profiling reveals the regulatory role of the
circRNA-100338/MIR-141-3p pathway in hepatitis B-related
hepatocellular carcinoma. Sci Rep. 7:54282017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hsiao KY, Lin YC, Gupta SK, Chang N, Yen
L, Sun HS and Tsai SJ: Noncoding effects of circular RNA CCDC66
promote colon cancer growth and metastasis. Cancer Res.
77:2339–2350. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang Y, Huang L, Li D, Shao J, Xiong S,
Wang C and Lu S: Hsa_circ_0101996 combined with hsa_circ_0101119 in
peripheral whole blood can serve as the potential biomarkers for
human cervical squamous cell carcinoma. Int J Clin Exp Pathol.
10:11924–11931. 2017.
|
|
21
|
Gao YL, Zhang MY, Xu B, Han LJ, Lan SF,
Chen J, Dong YJ and Cao LL: Circular RNA expression profiles reveal
that hsa_circ_0018289 is up-regulated in cervical cancer and
promotes the tumorigenesis. Oncotarget. 8:86625–86633. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ma HB, Yao YN, Yu JJ, Chen XX and Li HF:
Extensive profiling of circular RNAs and the potential regulatory
role of circRNA-000284 in cell proliferation and invasion of
cervical cancer via sponging miR-506. Am J Transl Res. 10:592–604.
2018.PubMed/NCBI
|
|
23
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res. 41:D991–D995. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gao J, Aksoy BA, Dogrusoz U, Dresdner G,
Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, et al:
Integrative analysis of complex cancer genomics and clinical
profiles using the cBioPortal. Sci Signal. 6:pl12013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression
analyses for RNA-sequencing and microarray studies. Nucleic Acids
Res. 43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dweep H and Gretz N: MiRWalk2.0: A
comprehensive atlas of microRNA-target interactions. Nat Methods.
12:6972015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Agarwal V, Bell GW, Nam JW and Bartel DP:
Predicting effective microRNA target sites in mammalian mRNAs.
Elife. 4:2015. View Article : Google Scholar
|
|
28
|
Pasquinelli AE: MicroRNAs and their
targets: Recognition, regulation and an emerging reciprocal
relationship. Nat Rev Genet. 13:271–282. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Conway JR, Lex A and Gehlenborg N: UpSetR:
An R package for the visualization of intersecting sets and their
properties. Bioinformatics. 33:2938–2940. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kozomara A and Griffiths-Jones S: MiRBase:
Annotating high confidence microRNAs using deep sequencing data.
Nucleic Acids Res 42 (Database Issue). D68–D73. 2014. View Article : Google Scholar
|
|
31
|
Krüger J and Rehmsmeier M: RNAhybrid:
MicroRNA target prediction easy, fast and flexible. Nucleic Acids
Res. 34:W451–W454. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jiang H, Ma R, Zou S, Wang Y, Li Z and Li
W: Reconstruction and analysis of the lncRNA-miRNA-mRNA network
based on competitive endogenous RNA reveal functional lncRNAs in
rheumatoid arthritis. Mol Biosyst. 13:1182–1192. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jiao X, Sherman BT, Huang da W, Stephens
R, Baseler MW, Lane HC and Lempicki RA: DAVID-WS: A stateful web
service to facilitate gene/protein list analysis. Bioinformatics.
28:1805–1806. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Walter W, Sánchez-Cabo F and Ricote M:
GOplot: An R package for visually combining expression data with
functional analysis. Bioinformatics. 31:2912–2914. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Szklarczyk D, Franceschini A, Wyder S,
Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos
A, Tsafou KP, et al: STRING v10: Protein-protein interaction
networks, integrated over the tree of life. Nucleic Acids Res.
43:D447–D452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Xia S, Feng J, Chen K, Ma Y, Gong J, Cai
F, Jin Y, Gao Y, Xia L, Chang H, et al: CSCD: A database for
cancer-specific circular RNAs. Nucleic Acids Res. 46:D925–D929.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kerpedjiev P, Hammer S and Hofacker IL:
Forna (force-directed RNA): Simple and effective online RNA
secondary structure diagrams. Bioinformatics. 31:3377–3379. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Schoen MW, Woelich SK, Braun JT, Reddy DV,
Fesler MJ, Petruska PJ, Freter CE and Lionberger JM: Acute myeloid
leukemia induction with cladribine: Outcomes by age and leukemia
risk. Leuk Res. 68:72–78. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Johnston JB: Mechanism of action of
pentostatin and cladribine in hairy cell leukemia. Leuk Lymphoma.
52 (Suppl 2):S43–S45. 2011. View Article : Google Scholar
|
|
43
|
Mulligan SP, Karlsson K, Strömberg M,
Jønsson V, Gill D, Hammerström J, Hertzberg M, McLennan R, Uggla B,
Norman J, et al: Cladribine prolongs progression-free survival and
time to second treatment compared to fludarabine and high-dose
chlorambucil in chronic lymphocytic leukemia. Leuk Lymphoma.
55:2769–2777. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Mego M, Sycova-Mila Z, Obertova J, Rajec
J, Liskova S, Palacka P, Porsok S and Mardiak J: Intrathecal
administration of trastuzumab with cytarabine and methotrexate in
breast cancer patients with leptomeningeal carcinomatosis. Breast.
20:478–480. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Rusch VW, Figlin R, Godwin D and
Piantadosi S: Intrapleural cisplatin and cytarabine in the
management of malignant pleural effusions: A Lung Cancer Study
Group trial. J Clin Oncol. 9:313–319. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Baker JAR, Wickremsinhe ER, Li CH,
Oluyedun OA, Dantzig AH, Hall SD, Qian YW, Ring BJ, Wrighton SA and
Guo Y: Pharmacogenomics of gemcitabine metabolism: Functional
analysis of genetic variants in cytidine deaminase and
deoxycytidine kinase. Drug Metab Dispos. 41:541–545. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lamba JK: Genetic factors influencing
cytarabine therapy. Pharmacogenomics. 10:1657–1674. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Walboomers JM, Jacobs MV, Manos MM, Bosch
FX, Kummer JA, Shah KV, Snijders PJ, Peto J, Meijer CJ and Muñoz N:
Human papillomavirus is a necessary cause of invasive cervical
cancer worldwide. J Pathol. 189:12–19. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Rong D, Sun H, Li Z, Liu S, Dong C, Fu K,
Tang W and Cao H: An emerging function of circRNA-miRNAs-mRNA axis
in human diseases. Oncotarget. 8:73271–73281. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Du WW, Zhang C, Yang W, Yong T, Awan FM
and Yang BB: Identifying and characterizing circRNA-protein
interaction. Theranostics. 7:4183–4191. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ebbesen KK, Hansen TB and Kjems J:
Insights into circular RNA biology. RNA Biol. 14:1035–1045. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pamudurti NR, Bartok O, Jens M,
Ashwal-Fluss R, Stottmeister C, Ruhe L, Hanan M, Wyler E,
Perez-Hernandez D, Ramberger E, et al: Translation of CircRNAs. Mol
Cell. 66:9–21.e7. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Holdt LM, Kohlmaier A and Teupser D:
Molecular roles and function of circular RNAs in eukaryotic cells.
Cell Mol Life Sci. 75:1071–1098. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Jeck WR, Sorrentino JA, Wang K, Slevin MK,
Burd CE, Liu J, Marzluff WF and Sharpless NE: Circular RNAs are
abundant, conserved, and associated with ALU repeats. RNA.
19:141–57. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Xie B, Zhao Z, Liu Q, Wang X, Ma Z and Li
H: CircRNA has_circ_0078710 acts as the sponge of microRNA-31
involved in hepatocellular carcinoma progression. Gene.
683:253–261. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Xiong D, Dang Y, Lin P, Wen DY, He RQ, Luo
DZ, Feng ZB and Chen G: A circRNA-miRNA-mRNA network identification
for exploring underlying pathogenesis and therapy strategy of
hepatocellular carcinoma. J Transl Med. 16:2202018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Salzman J, Chen RE, Olsen MN, Wang PL and
Brown PO: Cell-type specific features of circular RNA expression.
PLoS Genet. 9:e10037772013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin
QF, Xing YH, Zhu S, Yang L and Chen LL: Circular intronic long
noncoding RNAs. Mol Cell. 51:792–806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Li Z, Huang C, Bao C, Chen L, Lin M, Wang
X, Zhong G, Yu B, Hu W, Dai L, et al: Exon-intron circular RNAs
regulate transcription in the nucleus. Nat Struct Mol Biol.
22:256–264. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
You X, Vlatkovic I, Babic A, Will T,
Epstein I, Tushev G, Akbalik G, Wang M, Glock C, Quedenau C, et al:
Neural circular RNAs are derived from synaptic genes and regulated
by development and plasticity. Nat Neurosci. 18:603–610. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Legnini I, Di Timoteo G, Rossi F, Morlando
M, Briganti F, Sthandier O, Fatica A, Santini T, Andronache A, Wade
M, et al: Circ-ZNF609 is a circular RNA that can be translated and
functions in myogenesis. Mol Cell. 66:22–37.e9. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Yang Y, Fan X, Mao M, Song X, Wu P, Zhang
Y, Jin Y, Yang Y, Chen LL, Wang Y, et al: Extensive translation of
circular RNAs driven by N6-methyladenosine. Cell Res. 27:626–641.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li F, Zhang L, Li W, Deng J, Zheng J, An
M, Lu J and Zhou Y: Circular RNA ITCH has inhibitory effect on ESCC
by suppressing the Wnt/β-catenin pathway. Oncotarget. 6:6001–6013.
2015.PubMed/NCBI
|
|
64
|
Xie H, Ren X, Xin S, Lan X, Lu G, Lin Y,
Yang S, Zeng Z, Liao W, Ding YQ and Liang L: Emerging roles of
circRNA_001569 targeting miR-145 in the proliferation and invasion
of colorectal cancer. Oncotarget. 7:26680–26691. 2016.PubMed/NCBI
|
|
65
|
Li P, Chen S, Chen H, Mo X, Li T, Shao Y,
Xiao B and Guo J: Using circular RNA as a novel type of biomarker
in the screening of gastric cancer. Clin Chim Acta. 444:132–136.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Mitra A, Pfeifer K and Park KS: Circular
RNAs and competing endogenous RNA (ceRNA) networks. Transl Cancer
Res. 7 (Suppl 5):S624–S628. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Holdt LM, Kohlmaier A and Teupser D:
Molecular functions and specific roles of circRNAs in the
cardiovascular system. Non-coding RNA Res. 3:75–98. 2018.
View Article : Google Scholar
|
|
68
|
Liu Q, Zhang X, Hu X, Dai L, Fu X, Zhang J
and Ao Y: Circular RNA related to the chondrocyte ECM regulates
MMP13 expression by functioning as a miR-136 ‘Sponge’ in human
cartilage degradation. Sci Rep. 6:225722016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Wang X, Tang S, Le SY, Lu R, Rader JS,
Meyers C and Zheng ZM: Aberrant expression of oncogenic and
tumor-suppressive microRNAs in cervical cancer is required for
cancer cell growth. PLoS One. 3:e25572008. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Li Y, Wang F, Xu J, Ye F, Shen Y, Zhou J,
Lu W, Wan X, Ma D and Xie X: Progressive miRNA expression profiles
in cervical carcinogenesis and identification of HPV-related target
genes for miR-29. J Pathol. 224:484–495. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Yi Y, Liu Y, Wu W, Wu K and Zhang W: The
role of miR-106p-5p in cervical cancer: From expression to
molecular mechanism. Cell death Discov. 4:362018.PubMed/NCBI
|
|
72
|
Park S, Eom K, Kim J, Bang H, Wang HY, Ahn
S, Kim G, Jang H, Kim S, Lee D, et al: MiR-9, miR-21, and miR-155
as potential biomarkers for HPV positive and negative cervical
cancer. BMC Cancer. 17:6582017. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Xu L, Xu Q, Li X and Zhang X: MicroRNA-21
regulates the proliferation and apoptosis of cervical cancer cells
via tumor necrosis factor-α. Mol Med Rep. 16:4659–4663. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Zhu X, Er K, Mao C, Yan Q, Xu H, Zhang Y,
Zhu J, Cui F, Zhao W and Shi H: miR-203 suppresses tumor growth and
angiogenesis by targeting VEGFA in cervical cancer. Cell Physiol
Biochem. 32:64–73. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Melar-New M and Laimins LA: Human
papillomaviruses modulate expression of MicroRNA 203 upon
epithelial differentiation to control levels of p63 proteins. J
Virol. 84:5212–5221. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Mao L, Zhang Y, Mo W, Yu Y and Lu H:
BANF1 is downregulated by IRF1-regulated microRNA-203 in
cervical cancer. PLoS One. 10:e01170352015. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Reshmi G and Pillai MR: Beyond HPV:
Oncomirs as new players in cervical cancer. FEBS Lett.
582:4113–4116. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Zhao S, Yao D, Chen J and Ding N:
Circulating miRNA-20a and miRNA-203 for screening
lymph node metastasis in early stage cervical cancer. Genet Test
Mol Biomarkers. 17:631–636. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Wang N, Li Y and Zhou J: Downregulation of
ribonucleotide reductase subunits M2 induces apoptosis and G1
arrest of cervical cancer cells. Oncol Lett. 15:3719–3725.
2018.PubMed/NCBI
|
|
80
|
Mazumder Indra D, Mitra S, Singh RK, Dutta
S, Roy A, Mondal RK, Basu PS, Roychoudhury S and Panda CK:
Inactivation of CHEK1 and EI24 is associated with the
development of invasive cervical carcinoma: Clinical and prognostic
implications. Int J cancer. 129:1859–1871. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Zheng L, Li T, Zhang Y, Guo Y, Yao J, Dou
L and Guo K: Oncogene ATAD2 promotes cell proliferation, invasion
and migration in cervical cancer. Oncol Rep. 33:2337–2344. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Cheng J, Lu X, Wang J, Zhang H, Duan P and
Li C: Interactome analysis of gene expression profiles of cervical
cancer reveals dysregulated mitotic gene clusters. Am J Transl Res.
9:3048–3059. 2017.PubMed/NCBI
|
|
83
|
Mutch DG and Bloss JD: Gemcitabine in
cervical cancer. Gynecol Oncol. 90:S8–S15. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Roy S, Devleena, Maji T, Chaudhuri P,
Lahiri D and Biswas J: Addition of gemcitabine to standard therapy
in locally advanced cervical cancer: A randomized comparative
study. Indian J Med Paediatr Oncol. 32:133–138. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Kalaghchi B, Abdi R, Amouzegar-Hashemi F,
Esmati E and Alikhasi A: Concurrent chemoradiation with weekly
paclitaxel and cisplatin for locally advanced cervical cancer.
Asian Pac J Cancer Prev. 17:287–291. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Seidman AD: Gemcitabine as single-agent
therapy in the management of advanced breast cancer. Oncology 15 (2
Suppl 3). S11–S14. 2001.
|
|
87
|
Wang E, Gulbis A, Hart JW and Nieto Y: The
emerging role of gemcitabine in conditioning regimens for
hematopoietic stem cell transplantation. Biol Blood Marrow
Transplant. 20:1382–1389. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Hu XC, Zhang J, Xu BH, Cai L, Ragaz J,
Wang ZH, Wang BY, Teng YE, Tong ZS, Pan YY, et al: Cisplatin plus
gemcitabine versus paclitaxel plus gemcitabine as first-line
therapy for metastatic triple-negative breast cancer (CBCSG006): A
randomised, open-label, multicentre, phase 3 trial. Lancet Oncol.
16:436–446. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Telli ML, Jensen KC, Vinayak S, Kurian AW,
Lipson JA, Flaherty PJ, Timms K, Abkevich V, Schackmann EA, Wapnir
IL, et al: Phase II study of gemcitabine, carboplatin, and iniparib
as neoadjuvant therapy for triple-negative and BRCA1/2
mutation-associated breast cancer with assessment of a tumor-based
measure of genomic instability: PrECOG 0105. J Clin Oncol.
33:1895–1901. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Park YH, Im SA, Kim SB, Sohn JH, Lee KS,
Chae YS, Lee KH, Kim JH, Im YH, Kim JY, et al: Phase II,
multicentre, randomised trial of eribulin plus gemcitabine versus
paclitaxel plus gemcitabine as first-line chemotherapy in patients
with HER2-negative metastatic breast cancer. Eur J Cancer.
86:385–393. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Niwińska A, Rudnicka H and Murawska M:
Breast cancer leptomeningeal metastasis: The results of combined
treatment and the comparison of methotrexate and liposomal
cytarabine as intra-cerebrospinal fluid chemotherapy. Clin Breast
Cancer. 15:66–72. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Laakmann E, Witzel I and Müller V:
Efficacy of Liposomal Cytarabine in the treatment of leptomeningeal
metastasis of breast cancer. Breast Care. 12:165–167. 2017.
View Article : Google Scholar : PubMed/NCBI
|