Introduction
Osteosarcoma (OS) is one of the commonest malignant
solid tumors in children and adolescents, which exhibits a
predilection to occur in the metaphysis of long bones, and most
commonly occurs in the distal femur (43%), proximal tibia (23%), or
humerus (10%) (1). Treatment for
the disease is multimodal, incorporating surgery and chemotherapy
(2). Among the various treatment
strategies, chemotherapy is the most important in view of its
multifunctionality, including reduction of the tumor size pre- or
post-operation, and inhibition of distant metastases, also
preventing their recurrence. Furthermore, large cooperative group
studies and international collaborative efforts have demonstrated
that the combination of high-dose methotrexate, doxorubicin, and
cisplatin (MAP) provides the most effective regimen (3). However, the various drawbacks of
chemotherapy, namely, metastasis, recurrence, side-effects induced
by the anticancer drugs, and chemoresistance, have yet to be
satisfactorily resolved (4).
Although ~30 years have passed since key anticancer drugs were
introduced, the development of novel therapeutic drugs for OS has
stagnated (5). Therefore, the
development of novel drugs and treatments with higher efficacies
and fewer side-effects is urgently required.
Protein homeostasis, which serves an important role
in maintaining normal cellular metabolism, is precisely regulated
according to complex processes. Proteasomes have a critical role in
regulating this process. Although essential for normal cell
regulation, proteasomal activity, in particular, is pivotal for the
proliferation of cancer cells. It has been reported that the level
and activity of proteasomes are >90% higher in primary tumors
compared with normal tissues (6),
and it is widely accepted that high levels of proteasomal activity
provide survival benefits for tumor cells in continuous
proliferation (7,8). In recent years, emerging evidence has
suggested that proteasome inhibitors may contribute towards
promising anticancer strategies (9), and a large number of them have been
used in clinical practice (10–12).
However, only a few of these proteasome inhibitors have been
demonstrated to inhibit the growth of OS cells (13,14);
furthermore, none of them have been approved by the Food and Drug
Administration (FDA) for the treatment of OS due to the high
therapeutic doses required and serious side-effects. Additionally,
all of them are covalent, and covalent inhibitors have highly
reactive and unstable chemical groups, which render them less
suitable as inhibitors (15).
PI-1840, which is a novel non-covalent proteasome inhibitor, was
first synthesized and reported in 2013 (16). Subsequently, it was revealed that
PI-1840 was able to inhibit the growth of several types of tumor
cells, and to act non-covalently. In addition, PI-1840 was revealed
to be more active against solid tumors (17); however, few systematic studies have
been published on the effects of non-covalent proteasome inhibitors
on OS cells. In the present study, two cell lines, MG-63 and U2-OS,
were selected to investigate the possible molecular mechanisms
involved in the antitumor effects of PI-1840 in vitro. It
was revealed that PI-1840 functioned well in the two cell lines
owing to its antitumor effects, including inhibition of growth and
metastasis, as well as inducing cell cycle arrest and increasing
the levels of apoptosis and autophagy. Collectively, these findings
indicated that PI-1840 may be a promising anticancer agent against
OS.
Materials and methods
Reagents and antibodies
PI-1840 was purchased from Selleck Chemicals
(Houston, TX, USA). HyClone™ Dulbecco's modified Eagle's medium
(DMEM) was purchased from GE Healthcare (Chicago, IL, USA). McCoy's
5A medium was purchased from Wuhan Boster Biological Technology,
Ltd., Wuhan, China). Fetal bovine serum (FBS) was purchased from
Roya Bio-Technology Co., Ltd. (Lanzhou, China). Dimethyl sulfoxide
(DMSO) and the Cell Counting Kit-8 (CCK-8) assay were obtained from
MedChem Express (Monmouth Junction, NJ, USA). The
Invitrogen® LIVE/DEAD® Viability/Cytotoxicity
Assay kit was purchased from Thermo Fisher Scientific, Inc.
(Waltham, MA, USA). The Annexin V-Fluorescein Isothiocyanate (FITC)
Apoptosis Detection and Cell Cycle and Apoptosis Analysis kits
(C1062 and C1052, respectively) were acquired from Beyotime
Institute of Biotechnology (Shanghai, China). Antibodies against
p21 (dilution 1:500; cat. no. WL0362), p27 (dilution 1:500; cat.
no. WL01769), Bax (dilution 1:1,000; cat. no. WL01637), B-cell
lymphoma-2 (Bcl-2) (dilution 1:500; cat. no. WL01556), caspase-8
(dilution 1:500; cat. no. WL02434), cleaved caspase-8 (dilution
1:500; cat. no. WL0153), cleaved poly ADP-ribose polymerase (PARP)
(dilution 1:500; cat. no. WL01932), cytochrome c (Cyto
c) (dilution 1:500; cat. no. WL01571), Beclin1 (dilution
1:500; cat. no. WL02508), microtubule-associated protein light
chain 3 (LC3) II/I (Ic3α/β; dilution 1:500; cat. no. WL01506),
matrix metalloproteinase 2 (MMP2) (dilution 1:500; cat. no.
WL1579), MMP9 (dilution 1:500; cat. no. WL01580), β-actin (dilution
1:1,000; cat. no. WL01845) and cytochrome c oxidase (COX IV)
(dilution 1:1,000; cat. no. WL02203) were all obtained from
Wanleibio Co., Ltd. (Shenyang, China). Antibodies against GAPDH
(dilution 1:1,000; cat. no. 5174), β-tubulin (dilution 1:1,000;
cat. no. 2146), phosphorylated (p)-p65 (dilution 1:1,000; cat. no.
8242), p65 (dilution 1:1,000; cat. no. 3033), IκBα (dilution
1:1,000; cat. no. 4812), p-IκBα (dilution 1:1,000; cat. no. 2859),
and the tyrosine kinase, WEE1 (dilution 1:1,000; cat. no. 4936),
were purchased from Cell Signaling Technology, Inc. (Danvers, MA,
USA). The antibody against cleaved caspase-9 (dilution 1:500; cat.
no. ab2324) and caspase-9 (dilution 1:500; cat. no. ab2013) were
obtained from Abcam (Cambridge, MA, USA). Transwell chambers were
obtained from Corning, Incorporated (Corning, NY, USA).
SignalSilence® IκBα siRNA I (cat. no. 6327) and
SignalSilence® Control siRNA (unconjugated; cat. no.
6568) were obtained from Cell Signaling Technology, Inc. The
mRFP-GFP-LC3 adenovirus (cat. no. HB-AP2100001) was purchased from
HanBio Biotechnology Co. Ltd. (Shanghai, China). z-VAD-fmk and
chloroquine (Selleck Chemicals) were kindly provided by Dr Zou
Jilong (Department of Orthopedic Surgery, The First Affiliated
Hospital of Harbin Medical University, Harbin, China). All other
reagents and experimental materials were purchased from common
commercial sources.
Cell culture and treatment
The MG-63 and U2-OS OS cell lines were purchased
from the Type Culture Collection of the Chinese Academy of
Sciences, Shanghai Institute of Cell Biology (Shanghai, China). The
MG-63 and U2-OS cells were separately cultured in DMEM or McCoy's
5A medium with 10% FBS and 1% penicillin/streptomycin antibiotics.
All the cells were maintained at 37°C in a humidified atmosphere of
95% air and 5% CO2. The cells were treated with
indicated concentrations of PI-1840 and DMSO (vehicle used as a
control; the concentration of DMSO was <0.1%, which would not
have affected the physiological status of the OS cells). The
concentration of the stock solution of PI-1840 used was 40 mM (in
DMSO).
Observation of morphological
changes
The MG-63 and U2-OS cells were seeded into 6-well
plates at a density of 1×106 cells/well, and
subsequently allowed to attach to the wells at 37°C for 12 h. The
cultures were then treated with the different concentrations of
PI-1840 (MG-63: 0, 30 and 60 µM; U2-OS: 0, 20 and 40 µM), and
incubated at 37°C for 48 h. Morphology of the cells was then
observed under an inverted microscope (Olympus Corp., Tokyo,
Japan), and images were captured at a magnification of ×100.
Cell viability assay
The inhibitory effects of PI-1840 on the OS cells
were evaluated using CCK-8 assays. A cell suspension was produced
from OS cells, and the cells were counted. Subsequently, the cells
were seeded into a 96-well plate at a density of 5×103
cells/well, and incubated for 12 h to allow attachment of the cells
to take place. The cells were then treated with various
concentrations of PI-1840 (0, 5, 10, 20, 40, 80 and 160 µM) for 24
or 48 h. CCK-8 solution was mixed with serum-free medium at a
concentration of 10%, and subsequently replaced with the former
medium without any bubbles during the procedure. The plate was then
incubated for 1–4 h in the incubator at 37°C. Finally, the
absorbance was measured at 450 nm using a microplate reader (BioTek
China, Beijing, China).
Live/dead viability/cytotoxicity
assay
This assay was performed using a
LIVE/DEAD® Viability/Cytotoxicity Assay kit (Thermo
Fisher Scientific, Inc.). Following the manufacturer's protocol,
MG-63 and U2-OS cells were separately suspended and seeded into
6-well plates at a density of 1×106/well, after having
incubated the cells at 37°C for 12 h to allow them to adhere to the
plate. The cells were subsequently treated with different
concentrations of PI-1840 (MG-63: 0, 30 and 60 µM; U2-OS: 0, 20 and
40 µM) for 48 h at 37°C. At the end of this incubation period, the
cells were washed twice with phosphate-buffered saline (PBS) and
incubated for a further 15 min at 37°C; during this procedure, the
cells were incubated with DMEM or McCoy's 5A solution containing
2.5 µM Calcein AM and 4 µM ethidium homodimer-1 in the dark.
Finally, the live and the dead cells (determined from the green or
the red fluorescence, respectively) were observed and counted with
a fluorescence microscope (Olympus Corp.).
Cell cycle analysis
OS cells were seeded into 6-well plates at a density
of 1×106 cells/well, and subsequently incubated for 12 h
at 37°C. After attachment, MG-63 and U2-OS cells were separately
treated with PI-1840 (MG-63, 60 µM; U2-OS, 40 µM) for 24 or 48 h at
37°C. The cells were harvested after centrifugation (1,000 × g for
5 min), and washed with PBS twice. Subsequently, the cells were
fixed with 70% ethanol at 37°C overnight, centrifuged (1,000 × g
for 5 min) and washed again; the supernatant was subsequently
discarded. Staining buffer (500 µl) was added to each sample, and
then the samples were slowly resuspended. Propidium iodide (PI) (25
µl) and ribonuclease (RNase) A (10 µl) were added into each sample.
Avoiding light, the samples were incubated for 30 min at 37°C, and
subsequently stored on ice. All the samples were analyzed using a
flow cytometer (Becton, Dickinson and Company, Franklin Lakes, NJ,
USA).
Cell apoptosis analysis
The apoptotic effects of PI-1840 on MG-63 and U2-OS
cells were evaluated using the Annexin V-FITC/PI double staining
assay (Beyotime Institute of Biotechnology). Briefly, OS cells were
seeded into 6-well plates at a density of 1×106
cells/well, and subsequently incubated for 12 h at 37°C to allow
adherence to take place. Next, cells were treated with different
concentrations (MG-63: 0, 15, 30 and 60 µM; U2-OS: 0, 10, 20 and 40
µM) of PI-1840 for 48 h in an incubator at 37°C. Following the
PI-1840 treatment, cells in each group were harvested and
centrifuged for 5 min at 1,000 × g at room temperature. The
supernatant was discarded, and subsequently 195 µl Annexin V-FITC
binding solution was added to each sample to resuspend the cells.
Cells were then stained with 5 µl Annexin V-FITC and 5 µl PI,
following the manufacturer's protocol. Subsequently, the samples
were incubated for 15 min in the dark. Finally, the samples were
analyzed using a flow cytometer (Becton, Dickinson and
Company).
Transwell assay
Matrigel™ (BD Biosciences, San Jose, CA, USA) was
diluted with cold, filtered distilled water, and subsequently was
applied to the upper chamber of the polycarbonate Transwell filters
(Corning, Inc.). OS cells were seeded into 6-well plates at a
density of 1×106 cells/well, and incubated for 12 h at
37°C to allow adherence to take place. Subsequently, cells were
treated with PI-1840 at the concentration giving rise to
half-maximal inhibition (MG-63, 60 µM; U2-OS, 40 µM) for 48 h.
Cells were digested using trypsin, and then were resuspended and
counted. Cell suspension (200 µl) was added into the upper chamber
at a density of 2×104 cells/well, and subsequently 800
µl medium containing 30% FBS was added into the lower chamber of
each well of the 24-well plates. Cells were subsequently incubated
for 48 h. The Transwell filters were washed with PBS twice, and
cells were fixed at room temperature using 4% paraformaldehyde
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 15 min. After
drying, cells were stained using 0.5% crystal violet (Amresco, LLC,
Solon, OH, USA) for 5 min, and subsequently rinsed with distilled
water. Finally, images were captured at a magnification of ×200 by
an inverted microscope (Motic, AE31; Motic Incorporation, Ltd.,
Causeway Bay, Hong Kong), and the cells were counted.
Wound healing assay
OS cells were seeded at a density of
2×106 cells/well in 6-well plates and incubated for 12 h
at 37°C to allow adherence to take place. The cells were wounded
using a 10-µl white tip, and subsequently treated with various
concentrations of PI-1840 (MG-63: 0, 60 µM; U2-OS: 0, 40 µM). The
distances between the two edges were observed using a microscope
(Nikon Corp., Tokyo, Japan) at different time-points (0, 12 and 24
h), and the area of closure was assessed using ImageJ (1.46r)
software (National Institutes of Health, Bethesda, MD, USA).
Western blotting
OS cells were seeded into 6-well plates at a density
of 1×106 cells/well, and incubated for 12 h at 37°C to
allow adherence to take place. Subsequently, the medium was
replaced by medium mixed with different amounts of PI-1840 (MG-63:
15, 30 and 60 µM; U2-OS: 10, 20 and 40 µM) or vehicle (DMSO), and
the cells were incubated at 37°C for 24 or 48 h. After incubation
with the drug, the cells were washed with PBS three times, and then
lysed with cold radioimmunoprecipitation (RIPA) buffer (Wanleibio
Co., Ltd., WLA019) containing 1% phenylmethanesulfonyl fluoride and
10% phosphatase inhibitor, after which the cell lysates were
sonicated. The supernatants were collected after centrifugation at
12,000 × g for 15 min at 4°C, and subsequently the protein
concentration was measured using a bicinchoninic acid (BCA) protein
assay kit (Beyotime Institute of Biotechnology). As far as the
mitochondrial proteins were concerned, a cell mitochondria
isolation kit (Beyotime Institute of Biotechnology) was used prior
to lysing with cold RIPA buffer. The proteins were separated by
SDS-PAGE (concentration gel 5%, separation gel 12%), and then
transferred to polyvinylidene fluoride membranes. After blocking
with 5% non-fat milk for 2 h at room temperature, the membranes
were incubated with the primary antibodies (diluted in TBS) of
interest overnight at 4°C (see the Reagents and antibodies
section aforementioned for full details of the applied antibodies).
Then, the membranes were placed into TBST, and shaken for 5 min at
room temperature, and this step was repeated four times.
Subsequently, the appropriate secondary antibody (goat anti-rabbit
IgG-HRP; dilution 1:5,000; cat. no. WLA023; Wanleibio Co., Ltd.)
was added and incubated for 1 h at 37°C without light, and enhanced
chemiluminescence (ECL) substrate (Wanleibio Co., Ltd.) was used to
visualize the protein bands. The films were scanned using a
WD-9413B Scanner (Beijing Liuyi Culture Co., Ltd, Beijing,
China).
Evaluation of fluorescent LC3
puncta
The MG-63 and U2-OS cells were seeded into 24-well
plates at a density of 5×104 cells/well and incubated
for 12 h at 37°C. The cells were subsequently transfected with
mRFP-GFP-LC3 adenovirus (at a ratio of adenovirus number to cell
number of 100:1) for 24 h at 37°C, according to the manufacturer's
protocol, prior to treatment with the various concentrations of
PI-1840 (MG-63: 0, 60 µM; U2-OS: 0, 40 µM). Following treatment
with PI-1840 for 48 h, the cells were observed under a confocal
fluorescence microscope (Olympus Corp.), and images were
captured.
RNA interference knockdown of
IκBα
OS cells were seeded into 6-well plates at the
density of 5×105/well and incubated for 12 h to allow
adherence to take place at the density of 60–70% confluence, and
subsequently the cells were transfected with 100 nM IκBα siRNA I or
control siRNA with vehicle, and then incubated for 24 h at 37°C.
Subsequently, the cells were treated with or without PI-1840
(MG-63: 0, 60 µM; U2-OS: 0, 40 µM) and incubated at 37°C for 48 h
for subsequent assays. For the transfection procedure,
Lipofectamine™ RNAiMAX Transfection reagent (Thermo Fisher
Scientific, Inc.) was used, and the manufacturer's protocol was
followed.
Statistical analysis
All data values are expressed as the mean ± standard
deviation for three or more independent experiments. The
IC50 values were calculated using GraphPad Prism
software, version 7.0.4 (GraphPad Software, Inc., La Jolla, CA,
USA). Other statistical analyses were also performed using GraphPad
Prism version 7.0.4. Differences were assessed using one-way
analysis of variance, followed by Tukey's multiple-comparison test.
P<0.05 was considered to indicate a statistically significant
value.
Results
PI-1840 inhibits the proliferation of
MG-63 and U2-OS cells
To assess the effect of PI-1840 on the proliferation
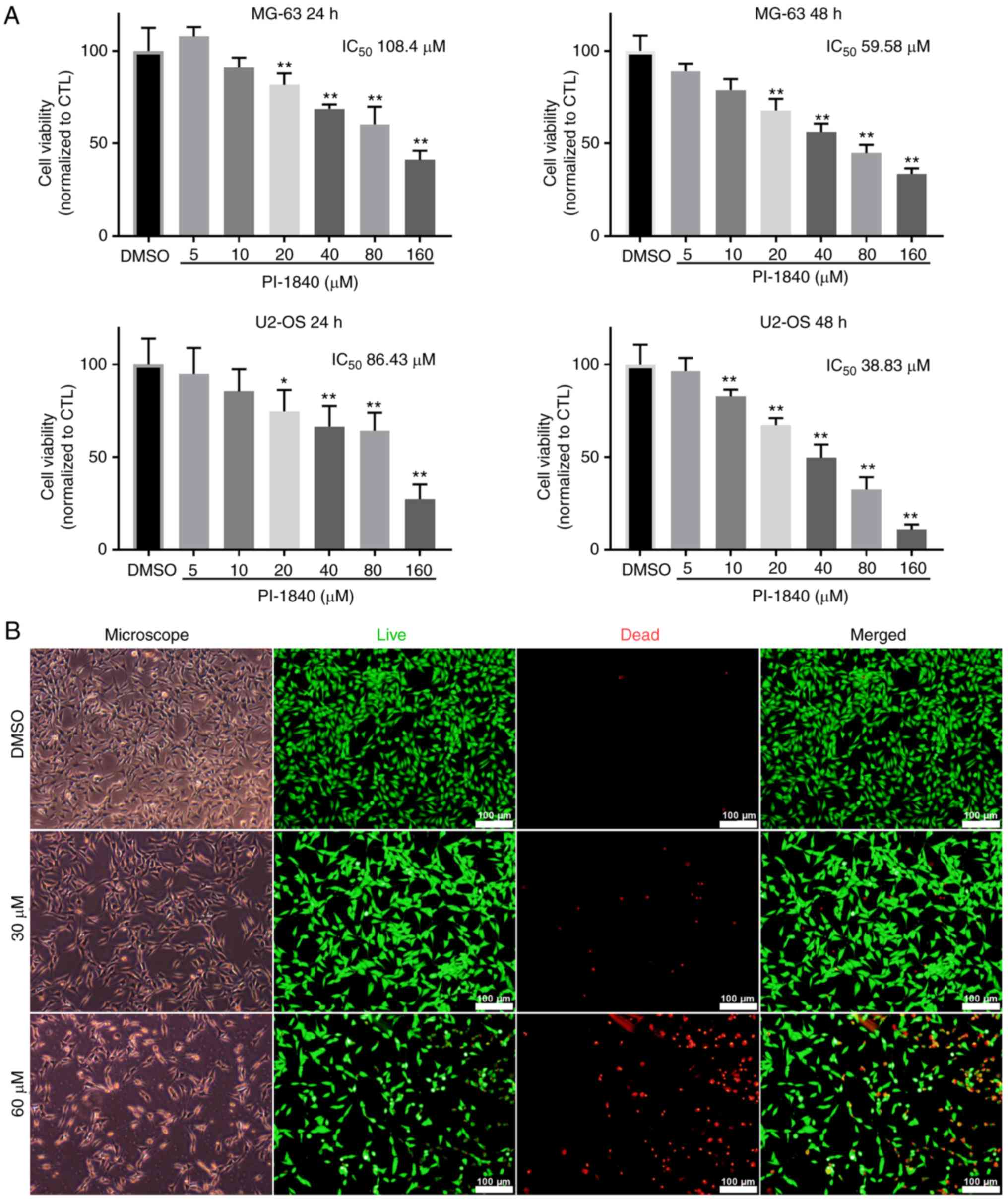
of OS cells, a CCK-8 assay was used. As revealed in Fig. 1A, cells were treated with PI-1840 at
different concentrations (5–160 µM) for 24 or 48 h. Cell growth was
inhibited in a dose-dependent manner following treatment with
PI-1840 for 48 h, and the IC50 values of the two cell
lines at 24 or 48 h were calculated to be 108.40 µM (MG-63, 24 h),
59.58 µM (MG-63, 48 h), 86.43 µM (U2-OS, 24 h), and 38.83 µM
(U2-OS, 48 h), respectively. Morphological observation and a
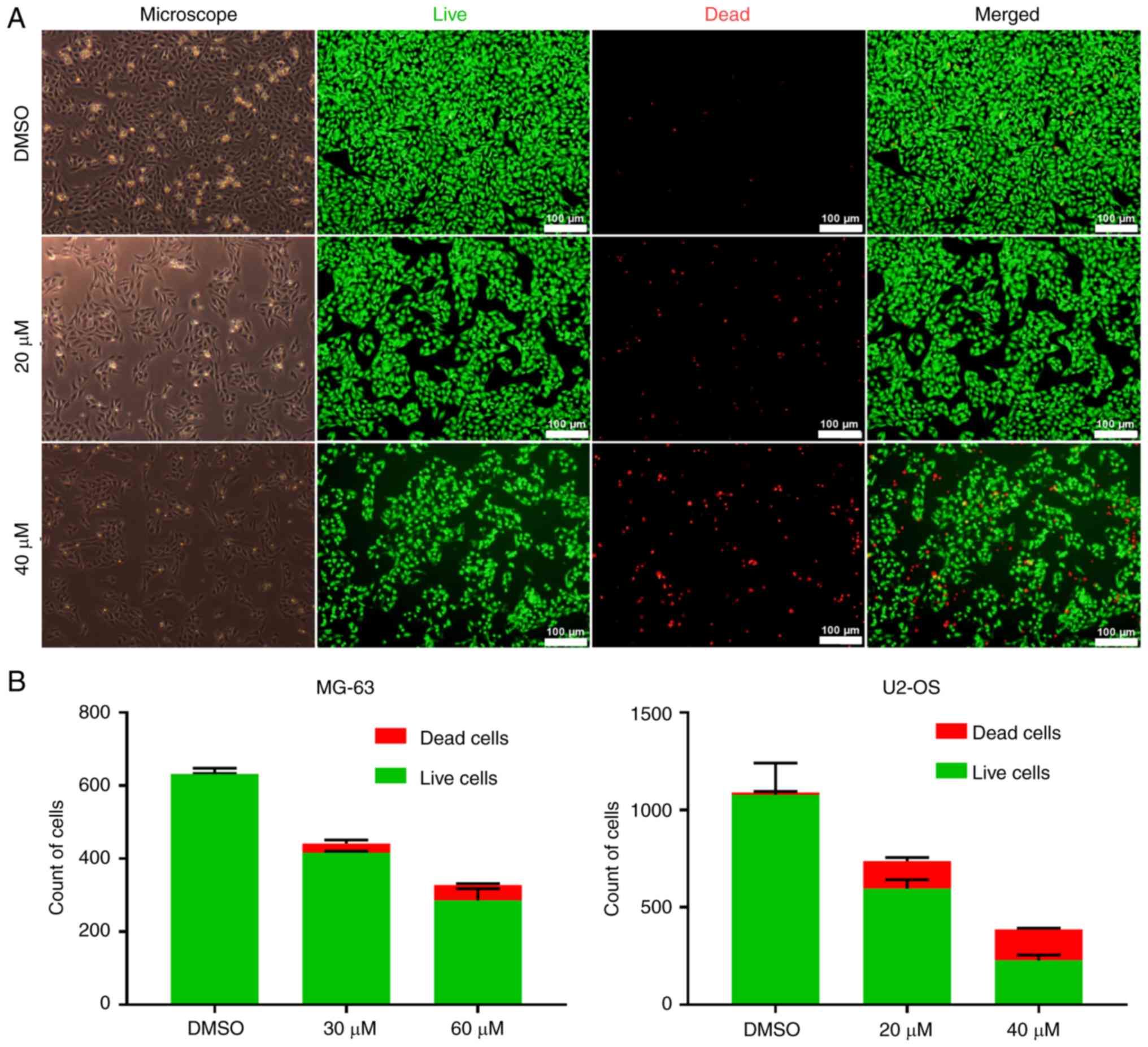
LIVE/DEAD® viability/cytotoxicity assay was also
employed to confirm the cytotoxicity of PI-1840 on OS cells. As
revealed in Figs. 1B and 2A, the cell density and the number of
cells with a normal distribution pattern were decreased, and the
number of atrophic or floating cells was increased, upon
administration of an increasing concentration of PI-1840. As the
concentration of the drug was increased, the density of live cells
(determined by the green fluorescence) was reduced, whereas the
dead cells (shown by the red fluorescence) exhibited the opposite
trend (Figs. 1B, and 2A and B).
PI-1840 induces cell cycle arrest at
the G2/M phase
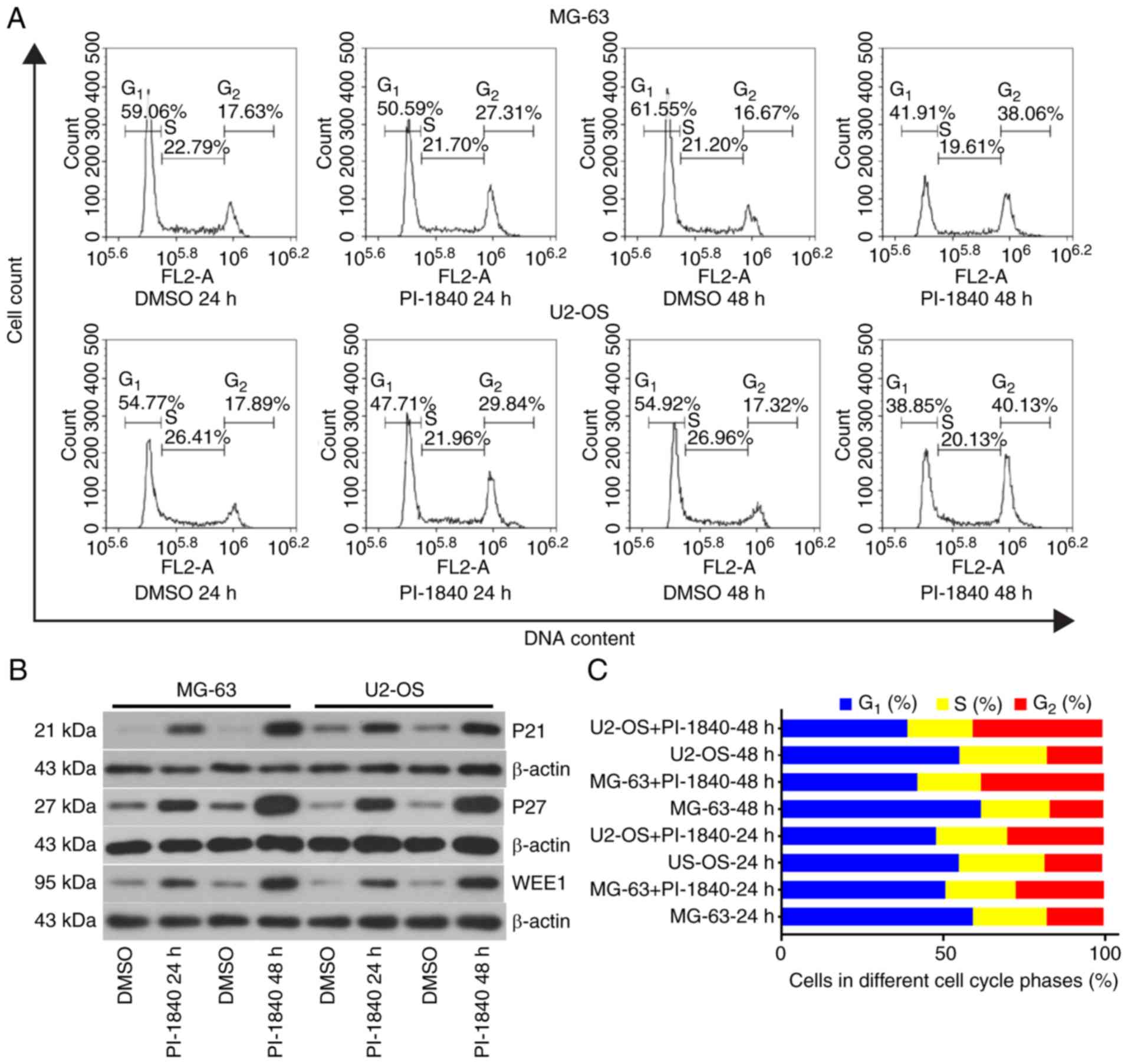
To determine the mechanism of inhibition of
proliferation, cell cycle distributions of the OS cells were
analyzed using flow cytometric analysis. As revealed in Fig. 3A and C, a marked increase in the
G2/M phase cell population was observed for the two cell
lines after 24 or 48 h treatment of PI-1840 at the indicated
concentrations. For further confirmation of cell cycle arrest,
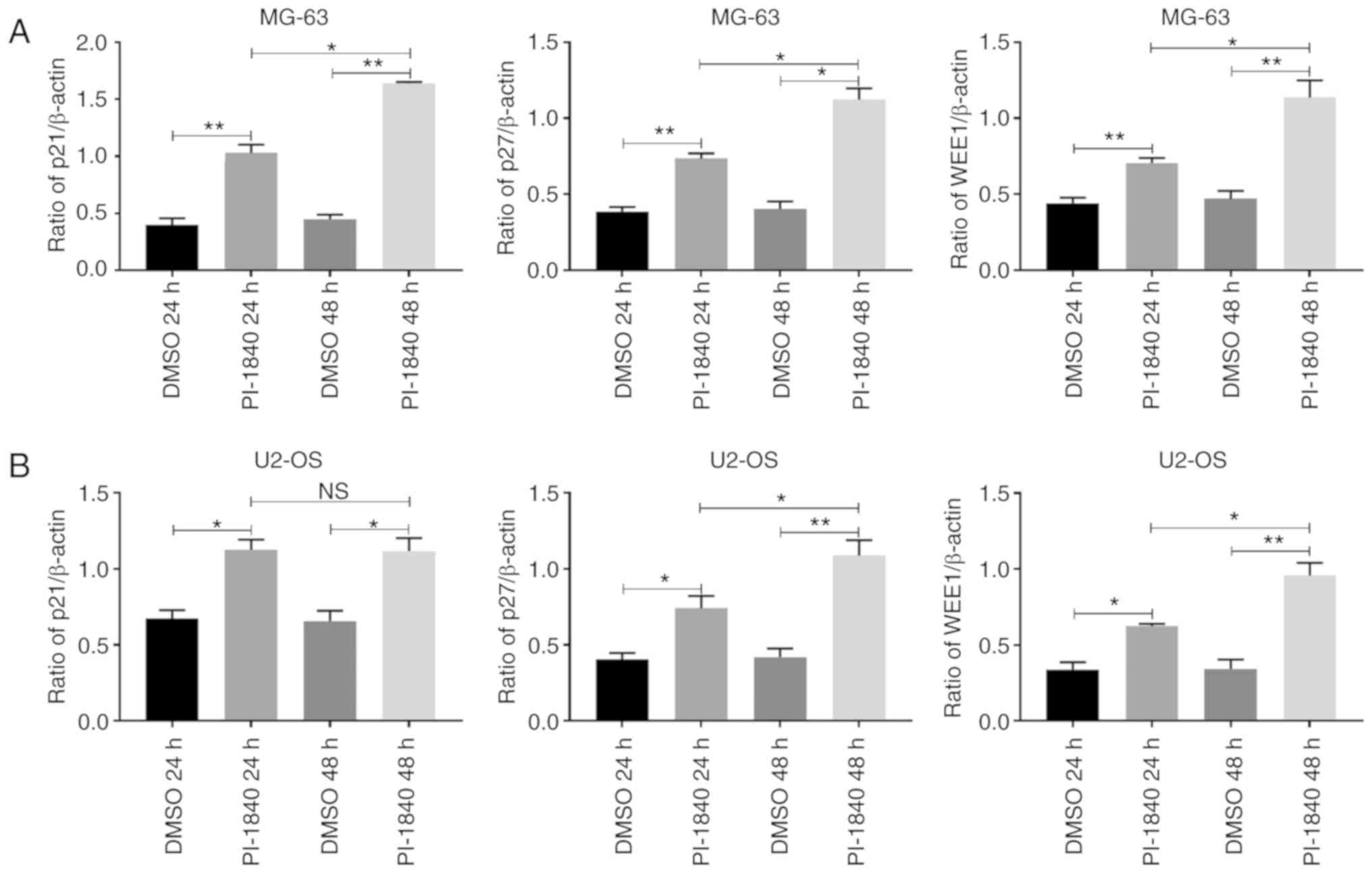
western blot assays were also performed. As revealed in Figs. 3B, and 4A and B, the cell cycle
regulation-associated proteins, p21, p27 and WEE1, accumulated in a
time-dependent manner following treatment with PI-1840 at the
indicated time-points.
PI-1840 induces apoptosis in MG-63 and
U2-OS cells
To further investigate the death of the OS cells,
Annexin V-FITC/PI double staining assay was performed. As revealed
in Fig. 5A, LL, LR, UR and UL
represent the live cells, early apoptotic cells, late apoptotic
cells, and cellular debris, respectively. The apoptotic rates
(Fig. 5B) of the two cell lines
were increased by PI-1840 treatment in a dose-dependent manner.
Subsequently, to investigate the underlying mechanism, IκBα,
p-IκBα, p65, p-p65, and apoptosis-associated proteins were analyzed
using western blot analysis. As revealed in Fig. 5C-E, following treatment with PI-1840
for 48 h, the ratio of the expression level of
(p-IκBα/control)/(IκBα/control) increased, and that of
(p-p65/control)/(p65/control) decreased, in a dose-dependent
manner, whereas the expression level of IκBα did not change
significantly. These results indicated that the NF-κB pathway was
inhibited by the administration of PI-1840. In order to further
explore the downstream apoptotic pathway, apoptosis-associated
proteins (including those of the extrinsic and intrinsic pathways)
were analyzed by western blot assay. As revealed in Fig. 6A-F, as the concentration of PI-1840
increased, the expression level of Bcl-2, which helps cells to
avoid apoptosis, was decreased in a dose-dependent manner.
Furthermore, the expression level of the mitochondrial proteins
Cyto c was decreased, while the expression levels of Bax,
and the ratios of (cleaved caspase-3/caspase-3, cleaved PARP/PARP,
cleaved caspase-8/caspase-8 and cleaved caspase-9/caspase-9 were
increased dose-dependently. The aforementioned results suggested
that, in the two cell lines, apoptosis was induced, and the
extrinsic and the intrinsic pathways were involved in this process.
To evaluate the degree of apoptosis in terms of the reduction of
cell viability, pan-caspase inhibitor (z-VAD-fmk) and CCK-8 assays
were performed. As revealed in Fig. 7A
and B, following co-treatment of the cells with z-VAD-fmk, the
percentage of living cells and cell viability were increased
significantly, suggesting that PI-1840-induced cell death was
caspase-dependent. To explore the association between inhibition of
the NF-κB pathway and the execution of apoptosis,
SignalSilence® IκBα siRNA I was applied. As revealed in
Fig. 7C and D, in the OS cells
pretreated with IκBα siRNA and subsequently treated with PI-1840,
the PI-1840-induced downregulation of cell viability was attenuated
compared with the cells co-treated with SignalSilence®
control siRNA and PI-1840. However, the cell viability of the group
co-treated with IκBα siRNA and PI-1840 remained higher compared
with that of the PI-1840 group (co-treated with
SignalSilence® control siRNA and PI-1840). In addition,
as revealed in Fig. 7E, activation
of the NF-κB pathway through inhibition of the expression of IκBα
led to lower expression levels of cleaved caspase-3, which
indicated that the caspase-dependent apoptosis pathway was
attenuated. Fig. 7F showed that
SignalSilence® IκBα siRNA I successfully suppressed the
expression of IκBα in both of the two cell lines. Collectively,
these results demonstrated that PI-1840 induced caspase-dependent
apoptosis, and that this phenomenon may be attributed, at least in
part, to the inhibition of NF-κB pathway.
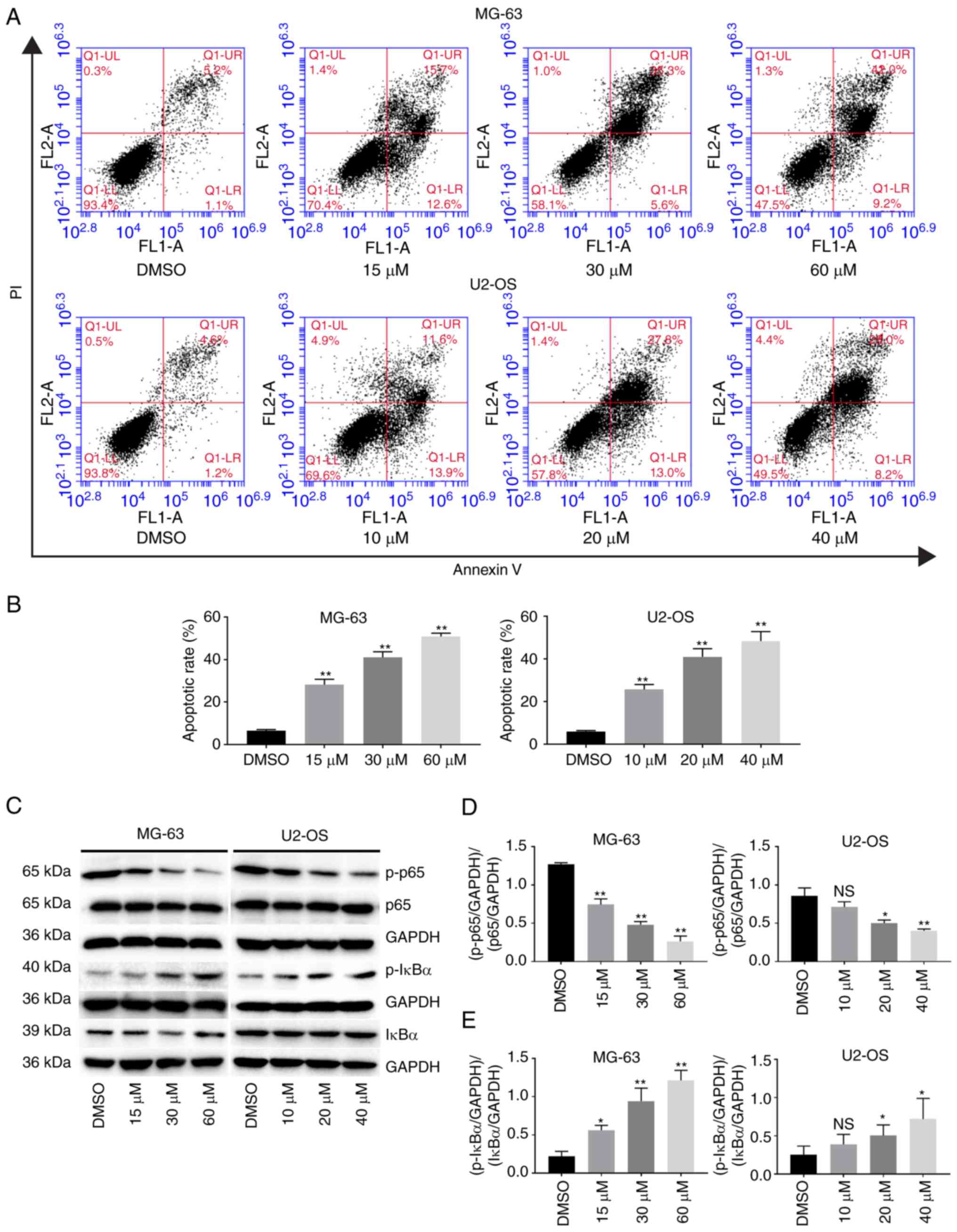 | Figure 5.PI-1840 induces the apoptosis of OS
cells. (A) PI-1840-induced apoptosis was confirmed using Annexin
V-FITC/PI double staining. The cells were exposed to PI-1840 for 48
h at the indicated concentrations. LL, LR, UR, and UL represent
live cells, early apoptotic cells, late apoptotic cells, and
cellular debris, respectively. (B) Summary plot for the apoptosis
rates (apoptosis rates=early apoptotic cell rates + late apoptotic
cell rates). (C-E) PI-1840 suppressed the nuclear factor-κB pathway
in a dose-dependent manner when cells were exposed to PI-1840 for
48 h at the indicated concentrations (n=3). *P<0.05; **P<0.01
compared with the DMSO group. ns, no significance; OS,
osteosarcoma; DMSO, dimethyl sulfoxide. |
 | Figure 6.PI-1840 induces the apoptosis of OS
cells. (A) Western blot analyses of caspase-3/cleaved caspase-3,
caspase-9/cleaved caspase-9, caspase-8/cleaved caspase-8,
PARP/cleaved-PARP, Bax, Bcl-2 and Cyto c. (B-F) Western blot
statistical analysis of caspase-3/cleaved caspase-3,
caspase-9/cleaved caspase-9, caspase-8/cleaved caspase-8,
PARP/cleaved-PARP, Bax, Bcl-2 and Cyto c (exposed to PI-1840
for 48 h; n=3). For caspase-3, −8 and −9 and PARP, the results were
presented using ‘(cleaved-protein/control)/(total
protein/control)’, *P<0.05; **P<0.01 compared with the DMSO
group. ns, no significance; CTL, control; OS, osteosarcoma; DMSO,
dimethyl sulfoxide; Bcl-2, B-cell lymphoma-2; Cyto c,
cytochrome c; PARP, poly(ADP-ribose) polymerase. |
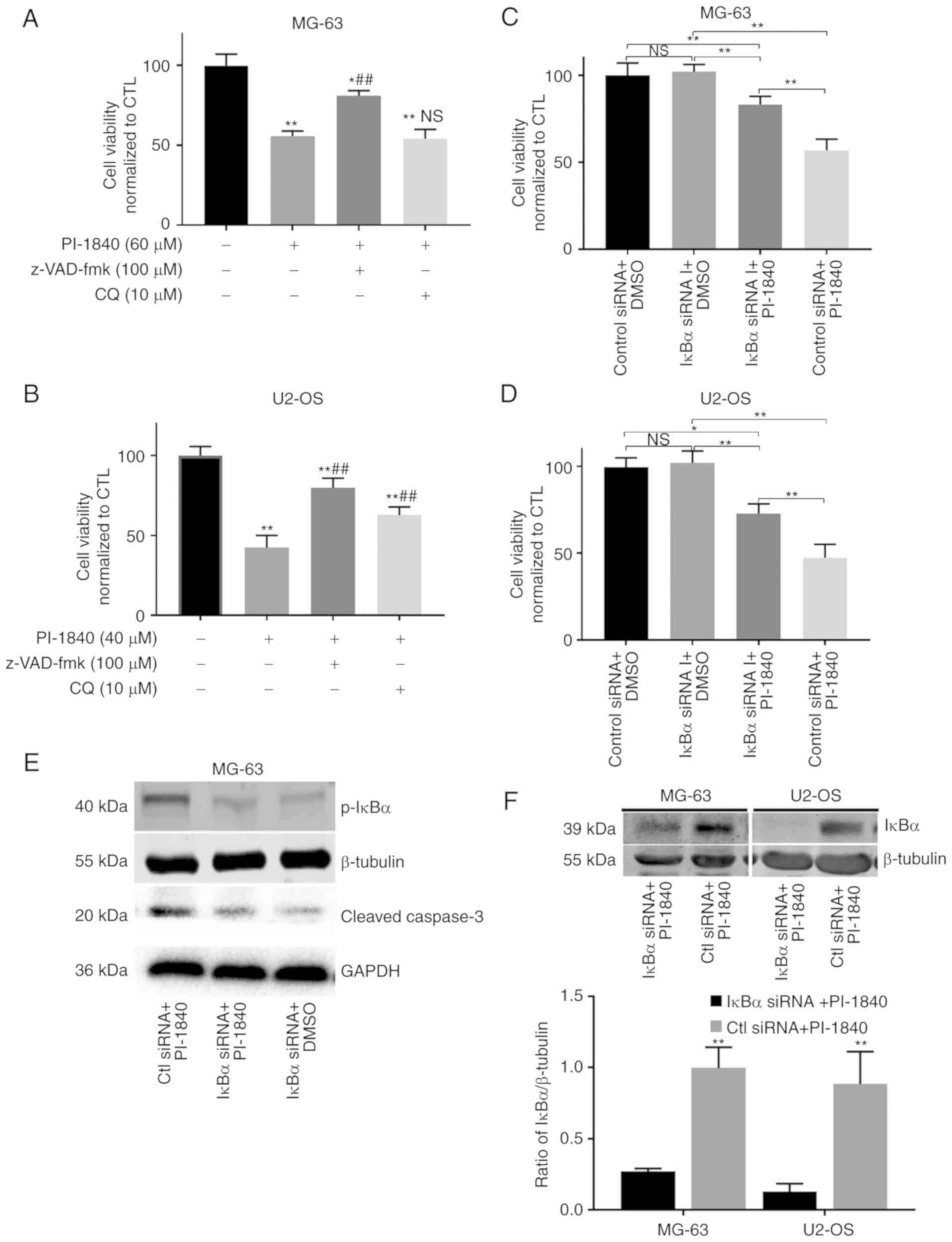 | Figure 7.(A and B) With the Cell Counting
Kit-8 assays, following treatment with PI-1840 and co-treatment
with the pan-caspase inhibitor, z-VAD-fmk, the cell viability
increased markedly compared with PI-1840 alone (48 h), although it
was still lower compared with that of DMSO group. Concurrently, the
results also revealed, following exposure to PI-1840 and
co-treatment with CQ (48 h), that the cell viability was not
significantly altered in the MG-63 cells, but was increased in the
U2-OS cells (n=6). *P<0.05; **P<0.01 compared with the DMSO
group; ##P<0.01 compared with the PI-1840 alone
group. (C and D) Blockade of protein IκBα with siRNA increased the
cell viability compared with PI-1840 alone (48 h treatment) group.
(n=6). *P<0.05; **P<0.01 compared with the control. (E) The
expression level of phospho-IκBα was markedly downregulated by
siRNA, and cleaved caspase-3 levels decreased subsequently (cells
were exposed to the IC50 concentration of PI-1840 for 48 h). (F)
The expression of IκBα was suppressed by SignalSilence®
IκBα siRNA I (cat. no. 6327) successfully in both of the two cell
lines (proteins were obtained after transfection with siRNA for 24
h). (n=3). **P<0.01 compared with the control. CTL, control;
DMSO, dimethyl sulfoxide; OS, osteosarcoma; NS, no significance;
CQ, chloroquine. |
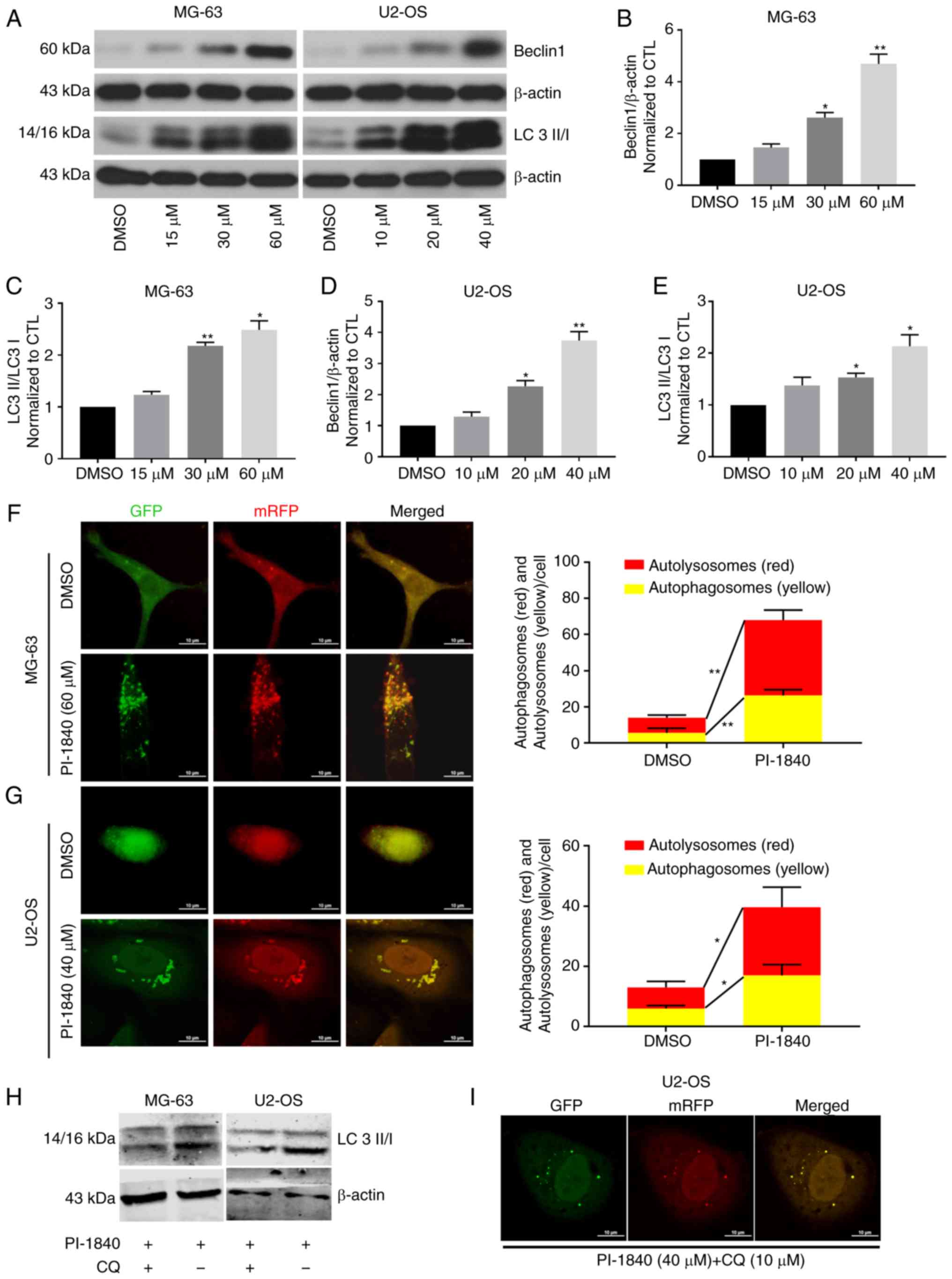
PI-1840 induces autophagy in MG-63 and
U2-OS cells
Previous research has shown that autophagy may
affect the survival of tumor cells. After having been treated with
proteasome inhibitors, the balance of protein metabolism is
disrupted, and this probably induces autophagy. Therefore, in the
present study, autophagy-associated proteins were also analyzed
using western blotting. As revealed in Fig. 8A-E, with an increasing dose of
PI-1840, the ratio of LC3 II to LC 3 I increased, and the
expression level of Beclin1 was also enhanced. To confirm the
occurrence of autophagic flux, mRFP-GFP-LC3 adenovirus transfection
was performed. As revealed in Fig. 8F
and G, the number of autophagosomes (denoted by the yellow dots
in cells) and autolysosomes (red dots in cells) increased
significantly following treatment with PI-1840 for 48 h.
Subsequently, CCK-8 assays were performed to assess whether the
occurrence of PI-1840-induced autophagy was able to affect the cell
viability of the two cell lines. As revealed in Fig. 7A, after the inhibition of autophagy
with chloroquine, for the MG-63 cell line, the cell viability was
not markedly altered compared with the group treated with PI-1840
alone. However, for the U2-OS cells (Fig. 7B), inhibiting autophagy did lead to
an attenuation of the effect of PI-1840-induced proliferation
inhibition, which suggested that PI-1840-induced autophagy was
detrimental to the U2-OS cells. In Fig.
8H and I, western blotting and fluorescence confocal images
showed that CQ could abolish the PI-1840 induced autophagy of the
OS cells.
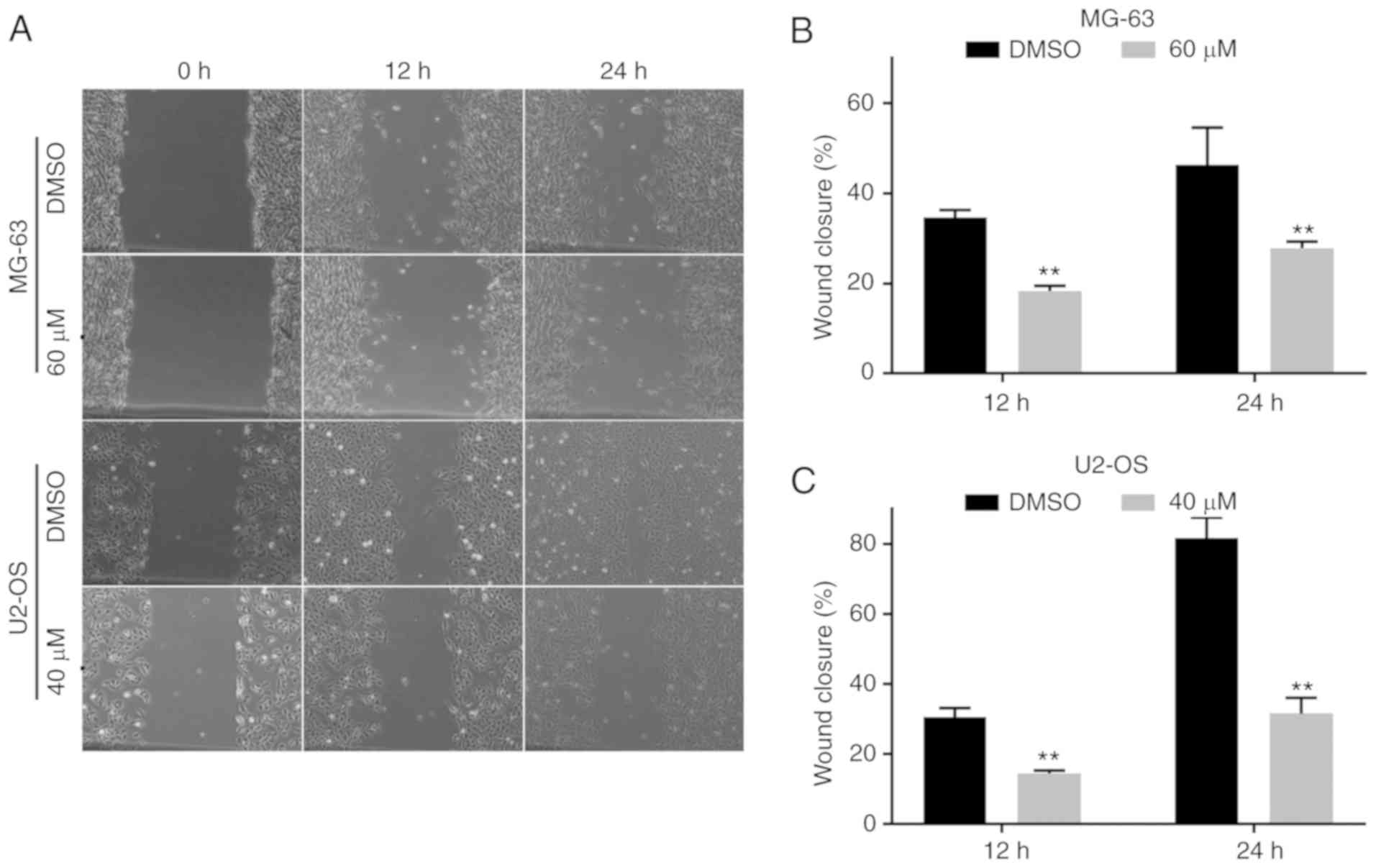
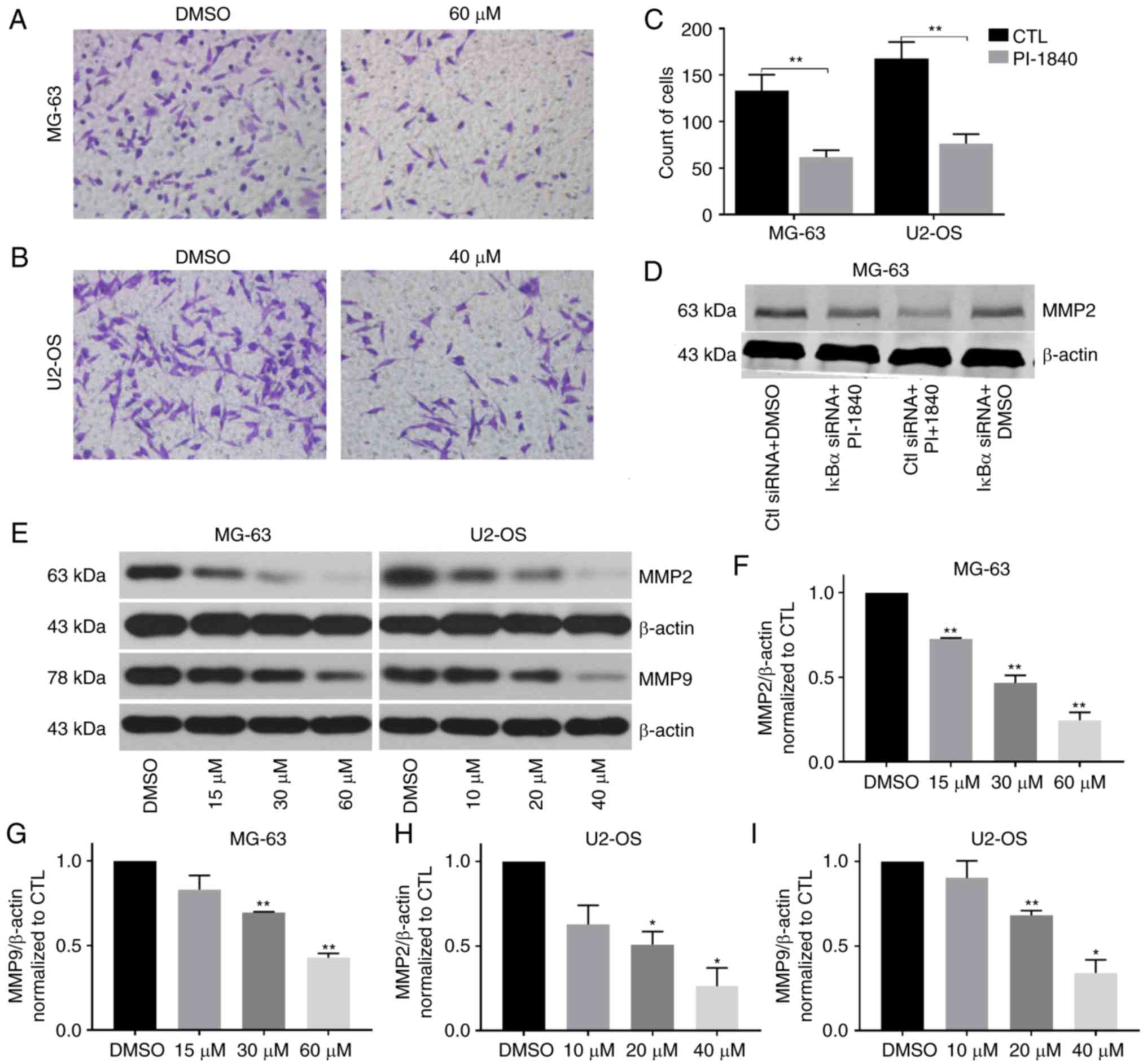
PI-1840 inhibits the ability of
migration and invasion of OS cells
To determine the effect of PI-1840 treatment on cell
migration, a wound healing assay was employed. As revealed in
Fig. 9A-C, the area between the two
edges was markedly decreased following treatment with PI-1840 for
12 and 24 h. To investigate the inhibition of invasion, Transwell
assays and western blotting were performed. As revealed in Fig. 10A-C, the number of OS cells
decreased significantly in the lower chamber compared with the
control group. Furthermore, the expression levels of the
investigated MMPs were decreased with an increasing concentration
of PI-1840 (Fig. 10E-I). In
contrast, after application of IκBα siRNA I, the expression level
of MMP2 was increased compared with the group both treated with
PI-1840 and control siRNA (Fig.
10D). These findings indicated that PI-1840 was able to
interfere with the migration and invasive capabilities of the OS
cells via inhibition of the NF-κB pathway.
Discussion
Protein metabolism has a pivotal role in cell
function, particularly in tumor cells. The disruption of the
protein metabolism process in tumor cells may contribute to
catastrophic levels of cell function disorder, affecting the
processes of proliferation, apoptosis and metastasis. Protein
degradation is predominantly executed through the proteasome (90%
of the total) and the lysosome (the remaining 10%). Therefore,
targeting the proteasome with proteasome inhibitors provides a
promising and feasible approach to suppress cancer. At present,
numerous proteasome inhibitors have been developed, and these have
continued to draw increasing attention for research purposes.
Although bortezomib, carfilzomib, and ixazomib have been approved
by the FDA for the treatment of certain types of cancer, the
side-effects of these drugs are not negligible. Furthermore, the
chemical structure of a large number of already developed
proteasome inhibitors, which determines that they bind covalently
to the proteasomes, is a major obstacle that hinders them from
being applied safely and widely in the clinic. PI-1840 is a novel
proteasome inhibitor that has fewer shortcomings compared with the
former ones owing to its unique chemical structure. It operates
with a non-covalent mode of action, is more selective, and exerts
milder effects on normal cells. However, only a small number of
systematic studies (17) have been
published to date on the anticancer effects and mechanisms of
non-covalent proteasome inhibitors, particularly with regard to OS
cell lines.
In the present study, the effects of PI-1840 on the
OS cell lines, MG-63 and U2-OS, were systematically evaluated,
including an assessment of the cell cycle, apoptosis, metastasis
and autophagy. Our analyses disclosed that PI-1840 was able to
suppress OS cells in vitro by various means. To the best of
our knowledge, our study is the first to demonstrate: i) That a
non-covalent proteasome inhibitor was able to cause the cell cycle
arrest of tumor cells; ii) non-covalent proteasome
inhibitor-induced autophagy, and its effect on the survival of OS
cells; and iii) an inhibition of the invasive capability of OS cell
lines.
In the evaluation of the inhibitory effect on
proliferation, live/dead viability assays were used. The results
revealed that the total number of cells decreased in the treated
cells compared with the DMSO cells, which was consistent with our
previous study that tested deoxyelephantopin (18). Hereby, we speculated that there are
two main reasons which led to the decrease in the total number of
cells: Firstly, many dead cells which did not attach to the plate
were washed away in the step where the cells were rinsed twice with
PBS buffer, while some dead or dying cells still attached to the
6-well plates; and thus, as the concentration of the drug
increased, the dead cells increased, and more cells washed away.
Thus, the total cells decreased as the concentration of the drug
increased. Secondly, since the cells were seeded into the 6-well
plates at the same density, after treatment with PI-1840 for 48 h,
with the concentration of PI-1840 increasing, the inhibitory effect
was stronger, thus the less total number of cells.
A previous study demonstrated that the cell cycle
period, and expression levels of cell cycle-associated proteins,
may be altered by proteasome inhibitors (19). Therefore, in the present study, the
cell cycle period and the cell cycle-associated proteins, p21, p27
and WEE1, were analyzed using flow cytometric analysis and western
blotting. p21 and WEE1 are two key cell cycle regulators that block
the G2/M transition by inhibiting the activity of
cyclin-dependent kinase 1 (CDK1)/cyclin B (20). It is noteworthy that, in our study,
the expression level of p27, which restricts the G1-S
transition by promoting the formation of the CDK2-cyclin E complex
to restrain its activity, was also increased. However, the results
of the FCM experiments indicated that the G1-S
transition was not blocked, which implied that, during the
PI-1840-induced cycle arrest, the accumulation of p27 and its
restriction on G1-S transition exerted a less pronounced
effect on cell cycle arrest compared with the overexpression of
G2/M checkpoint restrictors.
An aberrant activation of the NF-κB pathway is
common in a variety of types of human cancer, and this also holds
true for OS cells (21). A previous
study demonstrated that proteasome inhibitors may inhibit the NF-κB
pathway in intrahepatic cholangiocarcinoma (22), and the NF-κB pathway is one of the
upstream pathways of apoptosis (23). In the present study, it was
confirmed that PI-1840 could suppress the NF-κB pathway, and
subsequently induce OS cells to apoptosis in a caspase-dependent
manner. It was revealed that, in PI-1840-induced apoptosis, the
NF-κB pathway was an important activatory factor, but not the only
one, as determined by the findings of the downregulation of cleaved
caspase-3 and the increased cell viability, partly after
application of IκBα siRNA. Several key proteins lying further
downstream in the apoptosis signaling pathway were also
investigated. The level of Bcl-2, an anti-apoptotic protein that
promotes cell survival in response to a wide range of apoptotic
stimuli through inhibition of mitochondrial Cyto c release
(24), decreased as the
concentration of PI-1840 was increased. Furthermore, the level of
Cyto c in mitochondria decreased, which confirmed the
presence of apoptosis of the OS cells at the mitochondrial level
(25). In addition, the enhanced
expression of cleaved caspase-8 and cleaved caspase-9 suggested
that the PI-1840-induced apoptosis was associated with the
intrinsic and the extrinsic apoptotic pathways (26,27).
Previous studies have revealed that proteasome
inhibitors are able to inhibit the metastasis of tumor cells in
vitro and in vivo (28,29),
although none of the tested proteasome inhibitors were
non-covalent. The aforementioned results revealed that PI-1840
suppresses the NF-κB pathway, and that inhibiting the NF-κB pathway
may suppress the metastasis capability of the tumor cells (30–32).
In the present study, using western blot analysis and Transwell
assay, it was confirmed that PI-1840 was able to attenuate the
metastatic ability of the OS cells through inhibition of the NF-κB
pathway and downregulation of the expression level of MMPs.
Unlike apoptosis, the role of autophagy in cell
death and antitumor function remains controversial (33–35).
Autophagy may increase the survival rate of cells when they are
confronted with starvation, toxic substances or infection (36,37).
In contrast, autophagy is detrimental to cells under certain
circumstances, particularly tumor cells (38,39).
Furthermore, a complex situation exists between metabolic disorders
of proteins and autophagy (40),
and PI-1840 may disturb protein metabolism. Hence, in the present
study, the autophagic response of OS cells was investigated by
western blot analysis and mRFP-GFP-LC3 adenovirus assay. Beclin1, a
protein whose levels are positively associated with the level of
autophagy, was overexpressed in the two cell lines in a
dose-dependent manner following treatment with PI-1840, and
knowledge of its anticancer activity has been emerging in recent
years (41). To inhibit autophagy
of the OS cells, chloroquine was used, and subsequently a CCK-8
assay was performed to determine whether the PI-1840-induced
autophagy could influence the survival rate. Our results indicated
that, in the MG-63 cell line, autophagy did not affect the survival
rate of OS cells, and these results were consistent with a previous
study published by our group (18).
However, in the U2-OS cell line, the inhibition of autophagy
enhanced the survival rate of cells, which implied that the
function of PI-1840-induced autophagy is dependent on the type of
cell under consideration.
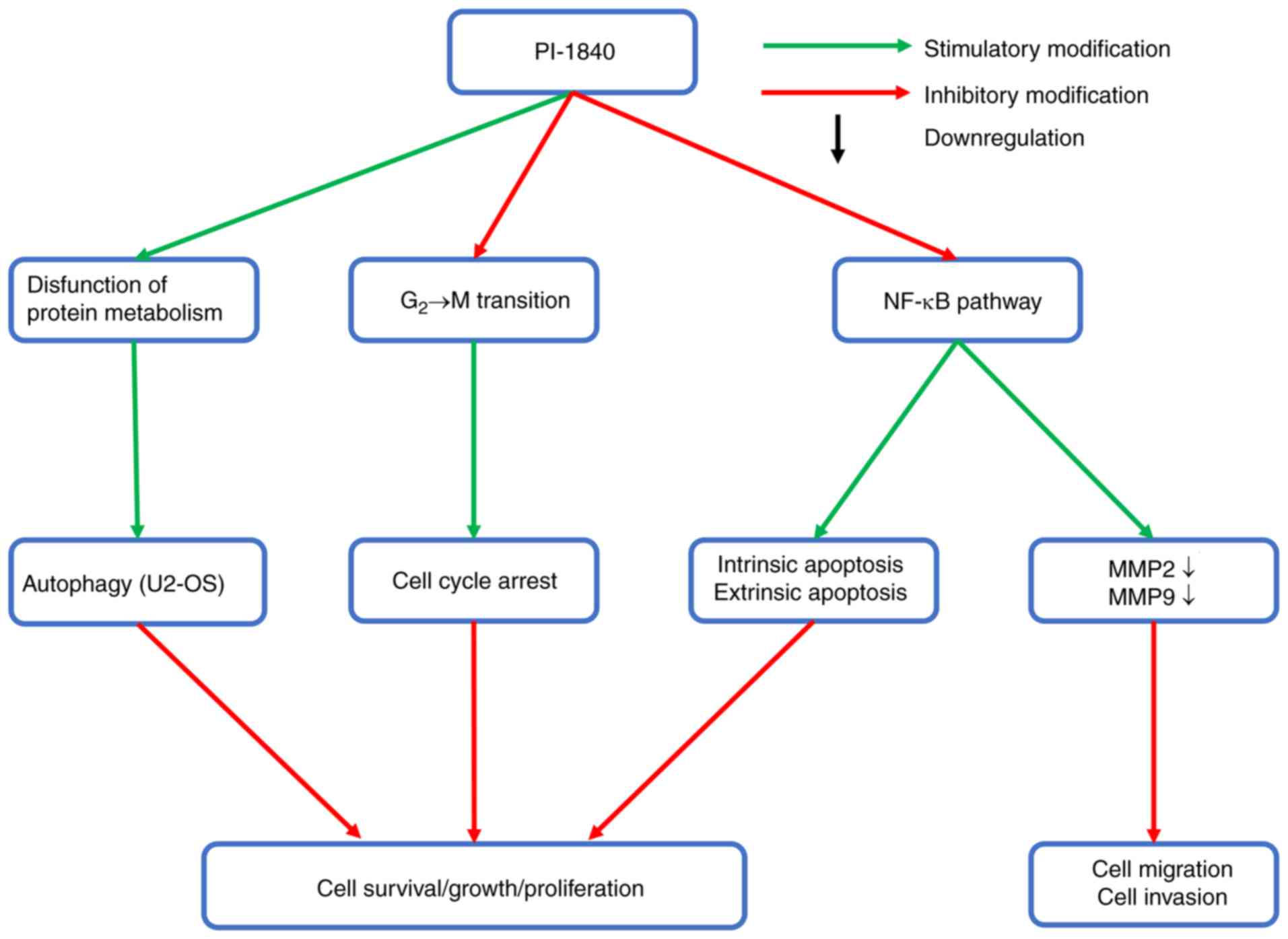
In conclusion, the present study has systematically
demonstrated the anticancer activity of a non-covalent proteasome
inhibitor, PI-1840, in the OS cell lines, MG-63 and U2-OS, by means
of inhibiting proliferation and metastasis, and inducing cell cycle
arrest, apoptosis and autophagy. A schematic representation of
these findings is presented in Fig.
11. These results provide a foundation for PI-1840 to become a
potential antitumor agent in OS treatment. However, further studies
are required to analyze the complex cross-talk that exists among
the NF-κB, apoptosis and autophagy signaling pathways (42,43). A
further limitation was that we were unable to conduct assays to
observe changes in tumor weight and metastasis in vivo after
treatment with PI-1840 in an experimental animal model, due to a
lack of available funds. Furthermore, in view of its different
anticancer mechanism with respect to the already employed drugs for
OS treatment, it would be interesting to explore the effects of
PI-1840 in combination with other first-line drugs, such as
doxorubicin or cisplatin, on OS.
Acknowledgements
The authors are grateful to staff from the High-Tech
Zone Laboratory of Public Test and Analysis Service (Shenyang,
China) for their assistance in these experiments.
Funding
Training funds for doctoral students were provided
by The First Affiliated Hospital of Harbin Medical University.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
ZB and YC conceived and designed the experiments.
YC, HX, HC, SY and CG performed the experiments. YC, HC, HX, SY and
LY analyzed the data. ZB and YC wrote the manuscript. All authors
read and approved the final manuscript and agree to be accountable
for all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bielack SS, Kempf-Bielack B, Delling G,
Exner GU, Flege S, Helmke K, Kotz R, Salzer-Kuntschik M, Werner M,
Winkelmann W, et al: Prognostic factors in high-grade osteosarcoma
of the extremities or trunk: An analysis of 1,702 patients treated
on neoadjuvant cooperative osteosarcoma study group protocols. J
Clin Oncol. 20:776–790. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Harrison DJ, Geller DS, Gill JD, Lewis VO
and Gorlick R: Current and future therapeutic approaches for
osteosarcoma. Expert Rev Anticancer Ther. 18:39–50. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Isakoff MS, Bielack SS, Meltzer P and
Gorlick R: Osteosarcoma: Current treatment and a collaborative
pathway to success. J Clin Oncol. 33:3029–3035. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Smith MA, Seibel NL, Altekruse SF, Ries
LA, Melbert DL, O'Leary M, Smith FO and Reaman GH: Outcomes for
children and adolescents with cancer: Challenges for the
twenty-first century. J Clin Oncol. 28:2625–2634. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yamamoto N and Tsuchiya H: Chemotherapy
for osteosarcoma-where does it come from? What is it? Where is it
going? Expert Opin Pharmacother. 14:2183–2193. 2013.PubMed/NCBI
|
|
6
|
Chen L and Madura K: Increased proteasome
activity, ubiquitin-conjugating enzymes, and eEF1A translation
factor detected in breast cancer tissue. Cancer Res. 65:5599–5606.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cenci S, Oliva L, Cerruti F, Milan E,
Bianchi G, Raule M, Mezghrani A, Pasqualetto E, Sitia R and Cascio
P: Pivotal advance: Protein synthesis modulates responsiveness of
differentiating and malignant plasma cells to proteasome
inhibitors. J Leukoc Biol. 92:921–931. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Voutsadakis IA: Proteasome expression and
activity in cancer and cancer stem cells. Tumor Biol.
39:10104283176922482017. View Article : Google Scholar
|
|
9
|
Teicher BA and Tomaszewski JE: Proteasome
inhibitors. Biochem Pharmacol. 96:1–9. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Moreau P, Masszi T, Grzasko N, Bahlis NJ,
Hansson M, Pour L, Sandhu I, Ganly P, Baker BW, Jackson SR, et al:
Oral ixazomib, lenalidomide, and dexamethasone for multiple
myeloma. N Engl J Med. 374:1621–1634. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Palumbo A, Chanan-Khan A, Weisel K, Nooka
AK, Masszi T, Beksac M, Spicka I, Hungria V, Munder M, Mateos MV,
et al: Daratumumab, bortezomib, and dexamethasone for multiple
myeloma. N Engl J Med. 375:754–766. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
McBride A, Klaus JO and Stockerl-Goldstein
K: Carfilzomib: A second-generation proteasome inhibitor for the
treatment of multiple myeloma. Am J Heal Pharm. 72:353–360. 2015.
View Article : Google Scholar
|
|
13
|
Lou Z, Ren T, Peng X, Sun Y, Jiao G, Lu Q,
Zhang S, Lu X and Guo W: Bortezomib induces apoptosis and autophagy
in osteosarcoma cells through mitogen-activated protein kinase
pathway in vitro. J Int Med Res. 41:1505–1519. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu R, Fu C, Sun J, Wang X, Geng S, Wang
X, Zou J, Bi Z and Yang C: A new perspective for osteosarcoma
therapy: Proteasome inhibition by MLN9708/2238 successfully induces
apoptosis and cell cycle arrest and attenuates the invasion ability
of osteosarcoma cells in vitro. Cell Physiol Biochem. 41:451–465.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Singh J, Petter RC, Baillie TA and Whitty
A: The resurgence of covalent drugs. Nat Rev Drug Discov.
10:307–317. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ozcan S, Kazi A, Marsilio F, Fang B, Guida
WC, Koomen J, Lawrence HR and Sebti SM: Oxadiazole-isopropylamides
as potent and noncovalent proteasome inhibitors. J Med Chem.
56:3783–3805. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kazi A, Ozcan S, Tecleab A, Sun Y,
Lawrence HR and Sebti SM: Discovery of PI-1840, a novel noncovalent
and rapidly reversible proteasome inhibitor with anti-tumor
activity. J Biol Chem. 289:11906–11915. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zou J, Zhang Y, Sun J, Wang X, Tu H, Geng
S, Liu R, Chen Y and Bi Z: Deoxyelephantopin induces reactive
oxygen species-mediated apoptosis and autophagy in human
osteosarcoma cells. Cell Physiol Biochem. 42:1812–1821. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gu JJ, Hernandez-Ilizaliturri FJ, Kaufman
GP, Czuczman NM, Mavis C, Skitzki JJ and Czuczman MS: The novel
proteasome inhibitor carfilzomib induces cell cycle arrest,
apoptosis and potentiates the anti-tumour activity of chemotherapy
in rituximab-resistant lymphoma. Br J Haematol. 162:657–669. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Choi YH and Yoo YH: Taxol-induced growth
arrest and apoptosis is associated with the upregulation of the Cdk
inhibitor, p21WAF1/CIP1, in human breast cancer
cells. Oncol Rep. 28:2163–2169. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Di W, Khan M, Rasul A, Sun M, Sui Y, Zhong
L, Yang L, Zhu Q, Feng L and Ma T: Isoalantolactone inhibits
constitutive NF-κB activation and induces reactive oxygen
species-mediated apoptosis in osteosarcoma U2OS cells through
mitochondrial dysfunction. Oncol Rep. 32:1585–1593. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yu GY, Wang X, Zheng SS, Gao XM, Jia QA,
Zhu WW, Lu L, Jia HL, Chen JH, Dong QZ, et al: RA190, a proteasome
subunit ADRM1 inhibitor, suppresses intrahepatic cholangiocarcinoma
by inducing NF-κB-mediated cell apoptosis. Cell Physiol Biochem.
47:1152–1166. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang CL, Wang C, Yan WJ, Gao R, Li YH and
Zhou XH: Knockdown of TNFAIP1 inhibits growth and induces apoptosis
in osteosarcoma cells through inhibition of the nuclear factor-κB
pathway. Oncol Rep. 32:1149–1155. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Murphy KM, Ranganathan V, Farnsworth ML,
Kavallaris M and Lock RB: Bcl-2 inhibits Bax translocation from
cytosol to mitochondria during drug-induced apoptosis of human
tumor cells. Cell Death Differ. 7:102–111. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhu Y, Li M, Wang X, Jin H, Liu S, Xu J
and Chen Q: Caspase cleavage of cytochrome c1 disrupts
mitochondrial function and enhances cytochrome c release.
Cell Res. 22:127–141. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Djiadeu P, Farmakovski N, Azzouz D, Kotra
LP, Sweezey N and Palaniyar N: Surfactant protein D regulates
caspase-8-mediated cascade of the intrinsic pathway of apoptosis
while promoting bleb formation. Mol Immunol. 92:190–198. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Aral K, Aral CA and Kapila Y: The role of
caspase-8, caspase-9, and apoptosis inducing factor in periodontal
disease. J Periodontol. 90:288–294. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chauhan D, Tian Z, Zhou B, Kuhn D,
Orlowski R, Raje N, Richardson P and Anderson KC: In vitro and in
vivo selective antitumor activity of a novel orally bioavailable
proteasome inhibitor MLN9708 against multiple myeloma cells. Clin
Cancer Res. 17:5311–5321. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bao X, Ren T, Huang Y, Ren C, Yang K,
Zhang H and Guo W: Bortezomib induces apoptosis and suppresses cell
growth and metastasis by inactivation of Stat3 signaling in
chondrosarcoma. Int J Oncol. 50:477–486. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yin X, Feng C, Han L, Ma Y, Jiao Y, Wang
J, Jia L, Jing F, Gao X, Zhang Y, et al: Diallyl disulfide inhibits
the metastasis of type II esophageal-gastric junction
adenocarcinoma cells via NF-κB and PI3K/AKT signaling pathways
in vitro. Oncol Rep. 39:784–794. 2018.PubMed/NCBI
|
|
31
|
Liao D, Zhong L, Duan T, Zhang RH, Wang X,
Wang G, Hu K, Lv X and Kang T: Aspirin suppresses the growth and
metastasis of osteosarcoma through the NF-κB pathway. Clin Cancer
Res. 21:5349–5359. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jiang Q, Pan Y, Cheng Y and Li H, Liu D
and Li H: Lunasin suppresses the migration and invasion of breast
cancer cells by inhibiting matrix metalloproteinase-2/-9 via the
FAK/Akt/ERK and NF-κB signaling pathways. Oncol Rep. 36:253–262.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang H: Targeting autophagy in lymphomas:
A double-edged sword? Int J Hematol. 107:502–512. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mehrpour M, Esclatine A, Beau I and
Codogno P: Autophagy in health and disease. 1. Regulation and
significance of autophagy: An overview. Am J Physiol Cell Physiol.
298:C776–C785. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Gewirtz DA: The four faces of autophagy:
Implications for cancer therapy. Cancer Res. 74:647–651. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Karsli-Uzunbas G, Guo JY, Price S, Teng X,
Laddha SV, Khor S, Kalaany NY, Jacks T, Chan CS, Rabinowitz JD and
White E: Autophagy is required for glucose homeostasis and lung
tumor maintenance. Cancer Discov. 4:914–927. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bronietzki AW, Schuster M and Schmitz I:
Autophagy in T-cell development, activation and differentiation.
Immunol Cell Biol. 93:25–34. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wu J, Gao F, Xu T, Deng X, Wang C, Yang X,
Hu Z, Long Y, He X, Liang G, et al: miR-503 suppresses the
proliferation and metastasis of esophageal squamous cell carcinoma
by triggering autophagy via PKA/mTOR signaling. Int J Oncol. Mar
16–2018.(Epub ahead of print). View Article : Google Scholar
|
|
39
|
Zhao H, Jia H, Han Q and Zhang J: Homeobox
containing 1 inhibits liver cancer progression by promoting
autophagy as well as inhibiting stemness and immune escape. Oncol
Rep. 40:1657–1665. 2018.PubMed/NCBI
|
|
40
|
White E, Mehnert JM and Chan CS:
Autophagy, metabolism, and cancer. Clin Cancer Res. 21:5037–5046.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhu J, Cai Y, Xu K, Ren X, Sun J, Lu S,
Chen J and Xu P: Beclin1 overexpression suppresses tumor cell
proliferation and survival via an autophagy-dependent pathway in
human synovial sarcoma cells. Oncol Rep. 40:1927–1936.
2018.PubMed/NCBI
|
|
42
|
Su J, Liu F, Xia M, Xu Y, Li X, Kang J, Li
Y and Sun L: p62 participates in the inhibition of NF-κB signaling
and apoptosis induced by sulfasalazine in human glioma U251 cells.
Oncol Rep. 34:235–243. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Qian HR, Shi ZQ, Zhu HP, Gu LH, Wang XF
and Yang Y: Interplay between apoptosis and autophagy in colorectal
cancer. Oncotarget. 8:62759–62768. 2017. View Article : Google Scholar : PubMed/NCBI
|