Introduction
Gliomas are the most common tumors in the central
nervous system (CNS), which are associated with a poor prognosis.
Due to the ability of gliomas to locally invade the surrounding
brain parenchyma, the median survival time is only 12–21 months,
even in patients that have received appropriate diagnosis and
treatment, such as surgery, radiation and chemotherapy (1,2).
Although considerable effort and progress has been made to
coordinate effective therapies, the heterogeneity of gliomas, with
regards to cytology and gene expression, makes it a complex task
(3,4). Therefore, understanding the molecular
mechanism and identifying specific glioma-associated genes as
pharmacological targets for treatment is critically important.
It has previously been reported that the
evolutionarily conserved Hippo pathway serves an important role in
cellular proliferation, transformation and tumorigenesis. The core
of the Hippo pathway is a kinase cascade involving the
macrophage-stimulating (MST)1/2 kinases (also known as serine
threonine kinase 4/3) of the STE-20 family, and their downstream
kinases, large tumor suppressor kinase (LATS)1/2 of the AGC family
(5). Inhibition of the Hippo
pathway results in hyperactivation of the main downstream effector
yes-associated protein (YAP)/tafazzin (TAZ)/TEA domain
transcription factor (TEAD), which in turn promotes the expression
of cell proliferation-, epithelial-mesenchymal transition (EMT)-
and anti-apoptosis-associated genes, leading to accelerated tumor
development and malignancy (6).
Therefore, in cancer, various signaling cascades that suppress the
Hippo pathway and activate YAP/TAZ/TEAD have been described.
Notably, dysregulation of the Hippo pathway has been revealed to
participate in various types of cancers, including liver, lung,
colorectal, ovarian and prostate cancers (7). In glioma, previous studies have
reported that YAP and TAZ are highly expressed, thus increasing
tumor initiation and invasion (8,9). Gene
Expression Omnibus (GSE15824) data revealed the downregulation of
LATS2 in glioma (10); however,
whether downregulation of LATS2 mediates tumorigenesis through
activation of YAP/TAZ in glioma remains unknown.
The present study confirmed that LATS2 expression
was downregulated in glioma clinical specimens. Notably, in
vitro overexpression of LATS2 suppressed proliferation,
migration and invasion of U-372 MG cells, via increasing the levels
of phosphorylated (p)-YAP/TAZ, resulting in their cytoplasmic
retention. Taken together, these findings indicated the role of
LATS2-mediated phosphorylation of YAP/TAZ in glioma
tumorigenesis.
Materials and methods
Tissue specimens and clinical
data
The present study was approved by the Ethics
Committee of The Second Hospital of Hebei Medical University
(Shijiazhuang, China), and all patients provided written informed
consent. The patients included 40 men and 28 women, with a mean age
of 39.5 years (age range, 8–73 years), who were recruited to the
study between January 2016 and December 2017. A total of 80 tissue
samples (19 astrocytoma, 24 oligodendroglioma, 25 glioblastoma
samples and 12 normal samples) were obtained from the Department of
Pathology of The Second Hospital of Hebei Medical University, and
were used in accordance with the guidelines approved by The Second
Hospital of Hebei Medical University Ethics Committee. No patients
underwent radiation or chemotherapy prior to surgery. The 12 normal
cortex tissue specimens were obtained from individuals who died in
traffic accidents; their families provided informed consent for the
use of these tissues. The use of these 12 normal cortex tissue
specimens was also approved by the Ethics Committee of The Second
Hospital of Hebei Medical University.
Cell culture
The U-372 MG (TPBT001265C; Tongpai Biological
Technology, Shanghai, China), LN-229 (BNCC341218; BeNa Culture
Collection, Kunshan, China), U-251 MG (CBP60300; BeNa Culture
Collection) and A172 human glioma cell lines (CRL-1620; American
Type Culture Collection, Manassas, VA, USA), and the HEB human
normal glial cell line (BeNa Culture Collection) were cultured in
Dulbecco's modified Eagle's medium with high glucose (cat. no.
11965–092; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
supplemented with glutamine, penicillin/streptomycin (cat. no.
15070063; Gibco; Thermo Fisher Scientific, Inc.) and 10% fetal
bovine serum (cat. no. 12483020; Gibco; Thermo Fisher Scientific,
Inc.) at 37°C in an incubator containing 5% CO2.
Plasmids (3 µg/106 cells) or small interfering (si)RNA
(30 pmol/106 cells) were transfected into cells using
Lipofectamine® 3000 (cat. no. L3000015; Invitrogen;
Thermo Fisher Scientific, Inc.) or Lipofectamine®
RNAiMAX (cat. no. 13778150; Invitrogen; Thermo Fisher Scientific,
Inc.) respectively, according to the manufacturer's protocols.
Cells were cultured for another 48 h, or for the indicated
durations, prior to further analysis.
Plasmids and siRNA
The entire human LATS2 (Gene ID, 26524; size, 3,267
bp) and YAP (Gene ID, 10413; size, 1,515 bp) genes were amplified
by polymerase chain reaction (PCR) using KOD DNA polymerase
(Toyobo Life Science, Osaka, Japan), according to the
manufacturer's protocol. The fragments were then cloned into
pcDNA3.1 plasmids (cat. no. V79020; Invitrogen; Thermo Fisher
Scientific, Inc.) by restriction enzyme cutting and ligation, and
were sequenced by GENEWIZ, Inc. (Suzhou, China) through Sanger
sequencing for validation; the empty pcDNA3.1 vector was used as a
control plasmid. The primers used for PCR are listed in Table I, and restriction enzyme recognition
sites are underlined. Site-directed mutagenesis for YAP S127A was
introduced using the Site-Directed Mutagenesis kit (Beijing SBS
Genetech Co., Ltd., Beijing, China), according to the
manufacturer's protocol. The sequences of LATS2 siRNA and
scramble siRNA, which was used as a negative control, are described
in a previous study (11) and were
purchased from Invitrogen; Thermo Fisher Scientific, Inc.
 | Table I.Polymerase chain reaction primers and
siRNA sequences. |
Table I.
Polymerase chain reaction primers and
siRNA sequences.
| Primers/siRNAs | Sequences
(5′-3′) |
|---|
| LATS2-forward | CGGGATCCATGAGGCCAAAGACTTTTCCTGCC |
| LATS2-reverse | CCGCTCGAGCTACACGTACACAGGCTGGCAGCC |
| YAP-forward | CGGAATTCATGGATCCCGGGCAGCAG |
| YAP-reverse | CGTCTAGACTATAACCATGTAAGAAAGCTTTCTTTATCTAGC |
| siLAST2 |
UACCAUAAAUACAAUCUUCTT |
| Scramble |
UUCUCCGAACGUGUCACGUTT |
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated using TRIzol®
reagent (Invitrogen; Thermo Fisher Scientific, Inc.), and the
concentration of total RNA was measured using NanoDrop 2000
(NanoDrop; Thermo Fisher Scientific, Inc., Wilmington, DE, USA).
Subsequently, RNA (1 µg) was reverse transcribed using the
PrimeScript™ RT reagent kit with gDNA Eraser (Takara Biotechnology
Co., Ltd., Dalian, China), according to the manufacturer's
protocol. RT-qPCR was conducted using SYBR Premix Ex Taq (Takara
Biotechnology Co., Ltd.) on an Applied Biosystems 7500 Real-Time
PCR System (Applied Biosystems; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA). The thermal cycling conditions were as follows:
Pre-denaturation at 95°C for 30 sec, followed by 40 cycles at 95°C
for 5 sec and 60°C for 30 sec. Nucleotide sequences of the primers
used for RT-qPCR were as follows (5′-3′): LATS2, forward
TGGCACCTACTCCCACAG, and reverse CCAAGGGCTTTCTTCATCT; GAPDH, forward
TCATGGGTGTGAACCATGAGAA; and reverse GGCATGGACTGTGGTCATGAG. The
relative mRNA expression levels of LATS2 were normalized to GAPDH
and were calculated as 2ΔCq as described in previous
study, where ΔCq = CqLATS2 - CqGAPDH
(12).
Immunohistochemistry
Tissues were fixed in 10% neutral formalin for ≤24 h
at 25°C, and were then embedded in paraffin. Paraffin-embedded
tissue sections (4 µm) were cut, and were then dewaxed in xylene
and rehydrated in graded ethanol. After rinsing with water, the
sections were heated in antigen retrieval solution (cat. no. P0081;
Beyotime Institute of Biotechnology, Shanghai, China) for 10 min,
incubated with 3% hydrogen peroxide for 5 min at 25°C to inactivate
endogenous peroxidase and blocked with 3% bovine serum albumin
(BSA; cat. no. ST023-200g; Beyotime Institute of Biotechnology) at
25°C for 30 min. Subsequently, samples were incubated with LATS2
antibody (1:100; cat. no. ab110780; Abcam, Cambridge, MA, USA) at
4°C overnight. After three 5-min washes in Tris-buffered
saline-0.1% Tween-20 (TBST), the sections were incubated with
biotin-conjugated goat anti-rabbit immunoglobulin G (IgG) (1:500;
cat. no. SA00004-2; Wuhan Sanying Biotechnology, Wuhan, Hubei,
China) at 25°C for 45 min, and were subsequently incubated with
streptavidin-biotin complex containing horseradish peroxidase (HRP)
(cat. no. P0603; Beyotime Institute of Biotechnology) at 25°C for
30 min. Finally, sections were incubated with DAB until color
developed and images were captured under a bright field microscope.
Finally, a semi-quantitative assessment of LATS2 relative
expression was conducted using ImageJ software (ImageJ bundled with
64-bit Java 1.8.0_112; National Institutes of Health, Bethesda, MD,
USA). Briefly, 10 visual fields were randomly observed and 200
cells were counted in each field. The positively stained cells were
calculated for each section.
Western blot analysis
Cells were collected and lysed in
radioimmunoprecipitation assay buffer (cat. no. 9806; Cell
Signaling Technology, Inc., Danvers, MA, USA), and total protein
concentration was determined using the bicinchoninic acid assay.
Protein samples (20 µg) were loaded, separated by 10% SDS-PAGE and
transferred to polyvinylidene difluoride membranes. Blots were
blocked in 5% (w/v) dry skimmed milk at 25°C for 60 min and were
then incubated with the following primary antibodies: Anti-LATS2
(1:1,000; cat. no. ab110780; Abcam), anti-YAP (1:5,000; cat. no.
ab52771; Abcam), anti-p-YAP S127 (1:5,000; cat. no. ab76252;
Abcam), anti-TAZ (1:1,000; cat. no. ab84927; Abcam), anti-p-TAZ
Ser89 (1:1,000; cat. no. 75275; Cell Signaling Technology, Inc.),
anti-Cyclin D1 (1:200; cat. no. ab16663; Abcam),
anti-cyclin-dependent kinase (CDK)4 (1:1,000; cat. no. ab108357;
Abcam), anti-CDK6 (1:5,000; cat. no. ab124821; Abcam) anti-β-actin
(1:5,000; cat. no. ab8226; Abcam) at 4°C overnight. After washing
three times in TBST (10 min/wash), blots were incubated with
HRP-conjugated goat anti-mouse IgG (1:5,000; cat. no. SA00001-1;
Wuhan Sanying Biotechnology) or goat anti-rabbit IgG (1:5,000; cat.
no. SA00001-2; Wuhan Sanying Biotechnology) for 45 min at room
temperature. The blots were then developed using enhanced
chemiluminescence substrate (cat. no. 32209; Pierce; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Cell proliferation analysis
Cell growth curves were plotted using the Cell
Counting Kit-8 (CCK-8) assay (Dojindo Molecular Technologies, Inc.,
Kumamoto, Japan), according to the manufacturer's protocol. A total
of 104 cells were seeded in 96-well plates with six
replicates for each condition. Following incubation with the CCK-8
reagent, absorbance was measured at 450 nm. Each experiment was
performed in triplicate.
Cell cycle analysis
Cells were collected, washed twice in cold PBS and
fixed with 75% ethanol at −20°C. A total of 5×106 fixed
cells were then incubated with propidium iodide (PI) staining
solution containing 0.5% Triton X-100, 20 µg/ml PI (cat. no. p4170;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and 0.2 mg/ml RNase
A (Sigma-Aldrich; Merck KGaA) in PBS at 4°C for 30 min. Cells were
analyzed by FACSCalibur flow cytometry (BD Biosciences, Franklin
Lakes, NJ, USA), and the DNA content of labeled cells was
calculated. Each experiment was performed in triplicate.
Scratch wound healing assay
Cells were cultured on 35-mm glass base dishes as
~90% confluent monolayers. Subsequently, the cells were scraped
with a 200-µl pipette tip and the cells were washed twice with PBS
to remove cell debris. To analyze scratch wound closure, optical
images were captured at 0 and 24 h time points using a microscope
(Olympus IX71 microscope; Olympus Corporation, Tokyo, Japan) and
were analyzed with ImageJ (ImageJ bundled with 64-bit Java
1.8.0_112; National Institutes of Health).
Invasion assay
In vitro invasion assay was performed in
transwell inserts (pore size, 8-µm pore; Costar; Corning
Incorporated, Corning, NY, USA). Cell suspensions (1×106
cells/ml; 200 µl) were seeded on fibronectin-coated polycarbonate
membranes precoated with 50 µl Matrigel (1 mg/ml; BD Biosciences).
After 18 h, the cells on the lower side of the membrane were fixed
with pre-chilled 100% methanol at 4°C for 30 min and stained with
Giemsa at 25°C for 10 min. The stained cells were captured under a
microscope (Olympus IX71 microscope; Olympus Corporation) at ×200
magnification and were counted. Experiments were performed in
triplicate.
Immunofluorescence
Cells transfected with LATS2 plasmids were grown on
glass coverslips. After gentle washing with PBS, cells were fixed
in pre-chilled 4% paraformaldehyde for 30 min at 4°C. Subsequently,
fixed cells were permeabilized with 0.05% NP-40 in PBS containing
1% BSA for 30 min at 4°C, and blocked in 3% BSA and 10% normal goat
serum (cat. no. C0265; Beyotime Institute of Biotechnology) for 30
min at 25°C. Cells were incubated with anti-YAP (1:200; cat. no.
ab52771; Abcam) and anti-TAZ (1:200; cat. no. ab84927; Abcam) at
4°C overnight. After washing three times in 0.1% PBS-Tween-20 (3
min/wash), the coverslips were incubated with Alexa
Fluor® 488-conjugated goat anti-rabbit IgG (1:500; cat.
no. ab150077; Abcam) for 30 min at room temperature. Subsequently,
slides were incubated with DAPI (cat. no. C1005; Beyotime Institute
of Biotechnology) for 5 min at 25°C and were washed three times in
PBS (5 min/wash). Fluorescence was examined under an Olympus
fluorescence microscope (Olympus Corporation).
Statistical analyses
All statistical analyses were performed using SPSS
17.0 (SPSS, Inc., Chicago, IL, USA) software. Data are presented as
the means ± standard error of the mean, and differences between
groups were evaluated using Student's t-test for two groups, or
one-way analysis of variance followed by Tukey's multiple
comparisons test for three or more groups. P<0.05 was considered
to indicate a statistically significant difference.
Results
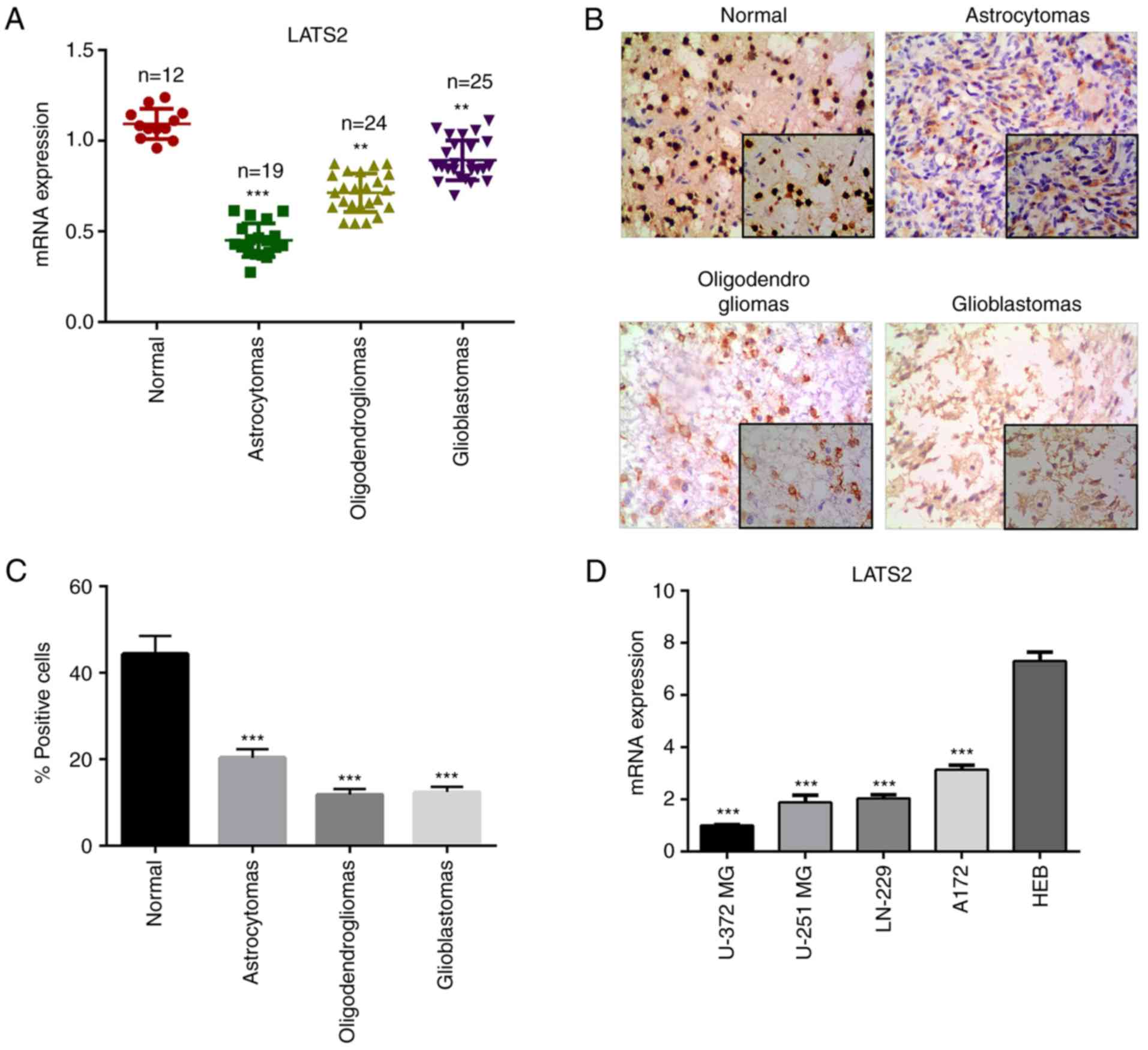
LATS2 is downregulated in glioma
To investigate the function of LATS2 in glioma, the
present study determined whether the relative expression levels of
LATS2 were altered in glioma tissues (astrocytomas,
oligodendrogliomas and glioblastomas) compared with in normal
tissue samples using RT-qPCR. The results revealed that the mRNA
expression levels of LATS2 were significantly reduced in
astrocytomas, oligodendrogliomas and glioblastomas compared with in
normal tissue samples (Fig. 1A).
For further validation, immunohistochemistry was performed to
examine the expression levels of LATS2 in clinical specimens.
Consistent with the RT-qPCR results, a lower signal was detected in
glioma samples compared with in normal tissues (Fig. 1B and C). These findings suggested
that LATS2 is downregulated in glioma, and prompted further
investigation into the biological functions of LATS2 in glioma. In
order to assess the function of LATS2 in vitro, the
expression levels of LATS2 were analyzed in a panel of glioma cell
lines, including U-372 MG, LN-229, U-251 MG and A172, and a normal
glial cell line (HEB). Of all the glioma cell lines, U-372 MG had
the lowest LATS2 expression, as revealed using RT-qPCR analysis;
this cell line was chosen for subsequent functional analysis
(Fig. 1D).
LATS2 suppresses cell proliferation,
migration and invasion in vitro
Since LATS2 functions as a core kinase in the Hippo
pathway regulating cell proliferation and EMT, the present study
assessed whether manipulating the expression levels of LATS2, by
plasmid or siRNA transfection, affected cell proliferation,
migration and invasion of U-372 MG cells. Initially, the
transfection efficiency of LATS2 plasmids and LATS2 siRNA was
confirmed. The mRNA and protein expression levels of LATS2 were
significantly upregulated in cells transfected with LATS2 plasmids
and were downregulated in cells transfected with siLATS2, compared
with in cells transfected with an empty vector or scramble siRNA
(Fig. 2A). To determine the effects
of LATS2 on cell proliferation, cell growth was measured using a
CCK-8 assay. The growth curves revealed that LATS2 overexpression
significantly inhibited cell growth, whereas LATS2 knockdown
resulted in opposite effects (Fig.
2B). Subsequently, the effects of LATS2 on cell cycle
progression were determined through flow cytometry using PI
staining. As shown in Fig. 2C,
overexpression of LATS2 increased the number of cells in the
G1 phase and decreased the number of cells in the S
phase, whereas LATS2 knockdown exhibited the opposite effect;
neither of them had any effect on the number of cells in the
G2/M phase (Fig. 2C).
These results indicated that expression of LATS2 delayed cell cycle
progression in the G1 phase. Given the known functions
of cyclin D1, CDK4 and CDK6 in G1/S transition, the
present study measured the expression levels of these proteins by
western blotting. Notably, overexpression of LATS2 decreased the
expression levels of cyclin D1, CDK4 and CDK6, where LATS2
knockdown had the opposite effect; these proteins are required for
progression through the G1 phase of the cell cycle
(Fig. 2D).
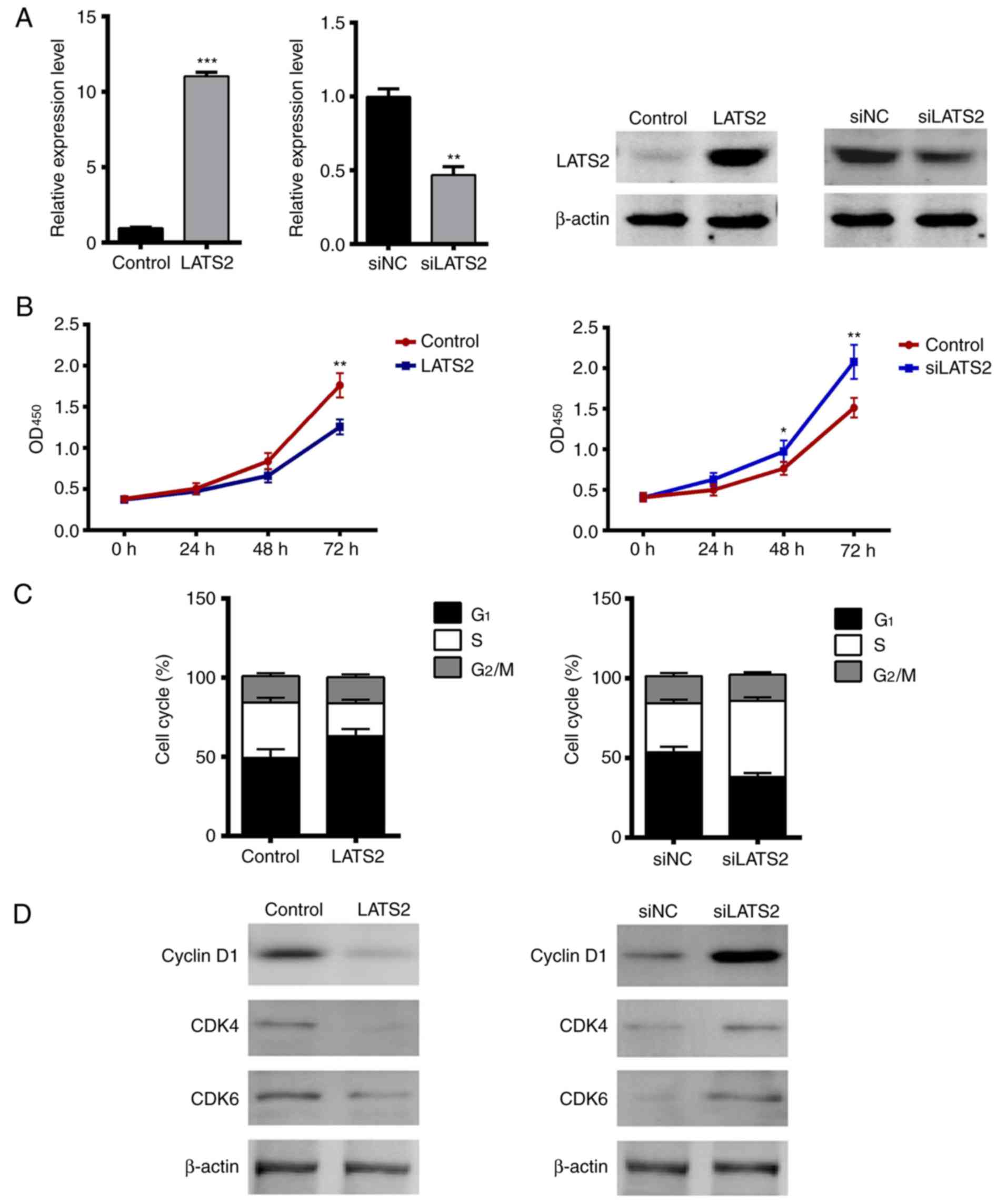 | Figure 2.Overexpression of LATS2 suppresses
cell proliferation by delaying G1/S transition in U-372
MG cells. (A) Efficiency of LATS2 plasmid and siLATS2 transfection
was validated in U-372 MG cells. The mRNA and protein expression
levels of LATS2 were significantly upregulated or downregulated in
cells transfected with plasmids or siRNA, respectively. Empty
vector or NC siRNA were used as controls. (B) Effects of LATS2 on
cell growth were determined by Cell Counting Kit-8 assay.
OD450 was measured at 0, 24, 48 and 72 h
post-transfection. Overexpression of LATS2 decreased cell growth
(left), whereas LATS2 knockdown increased cell growth (right). (C)
Effects of LATS2 on cell cycle progression were measured by flow
cytometry using propidium iodide. Overexpression of LATS2 increased
the percentage of cells in the G1 phase and decreased
the number of cells in the S phase compared with the control,
whereas knockdown of LATS2 had the opposite effects;
G2/M phase was not affected. (D) Protein expression
levels of cyclin D, CDK4 and CDK6 were determined by western
blotting and β-actin was used as an internal reference. Expression
levels of these proteins were decreased in LATS2-overexpressed
cells and were increased in LATS2 knockdown cells. All data are
presented as the means ± standard deviation (n=3). *P<0.05;
**P<0.01; ***P<0.001 compared with control or siNC groups.
CDK, cyclin-dependent kinase; LATS2, large tumor suppressor kinase
2; OD, optical density; NC, negative control; si/siRNA, small
interfering RNA. |
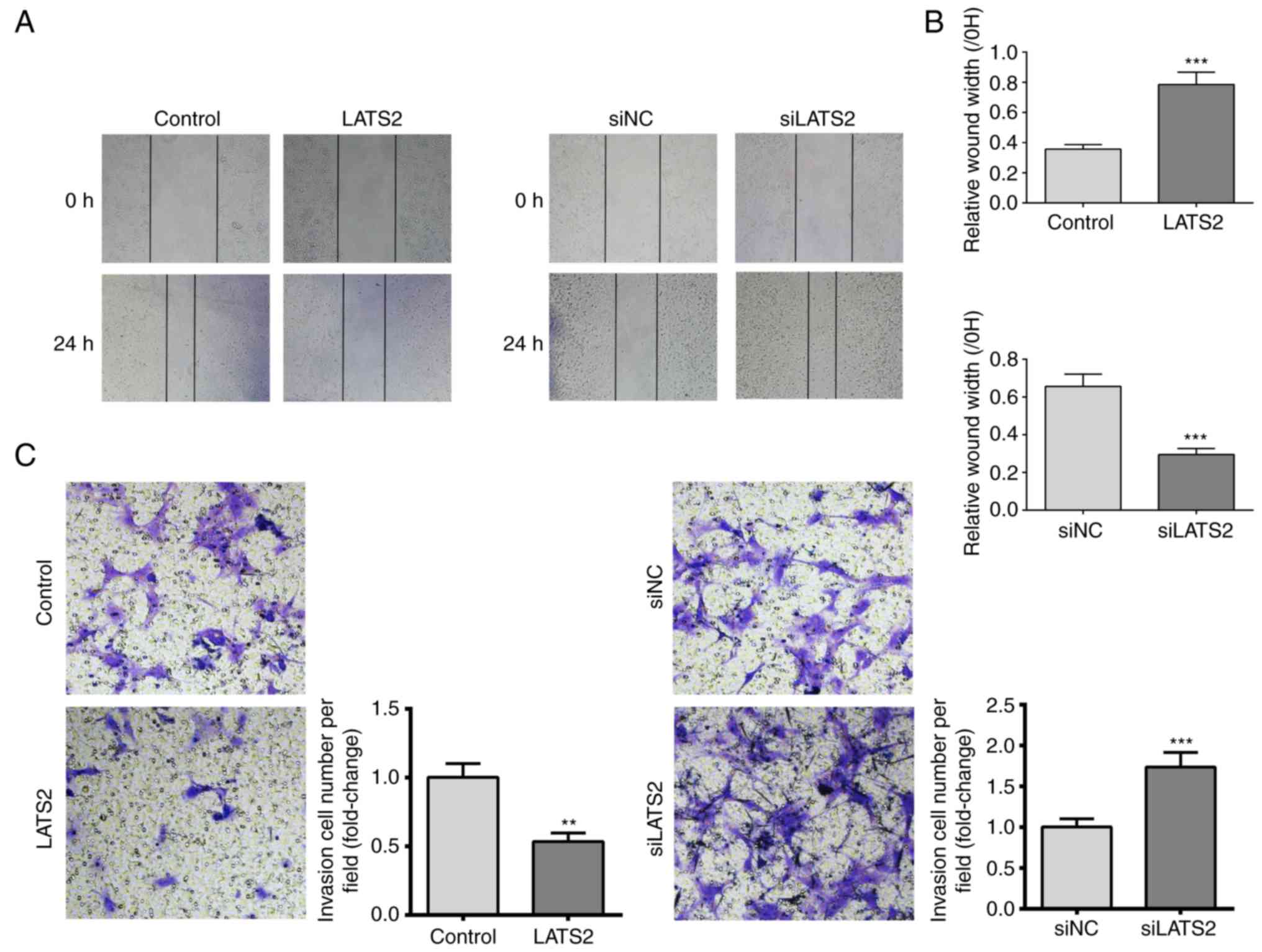
Since increased cell migration and invasion are
features of glioma, the present study investigated the effects of
LATS2 expression on cell migration and invasion using scratch wound
healing and transwell invasion assays, respectively. A U-372 MG
cell monolayer was used for the wound healing assay. The results
revealed that cells with LATS2 overexpression exhibited
significantly slower wound repair, whereas in cells with LATS2
knockdown, cell migration was increased compared with the control
group after 24 h (Fig. 3A and B).
Furthermore, transwell invasion assay revealed a reduction in the
number of invasive cells when LATS2 was overexpressed compared with
cells transfected with an empty vector. Consistently, LATS2
knockdown elevated the number of invasive cells (Fig. 3C). Taken together, these results
indicated that LATS2 dysregulation may affect growth, migration and
invasion of U-372 MG cells in vitro.
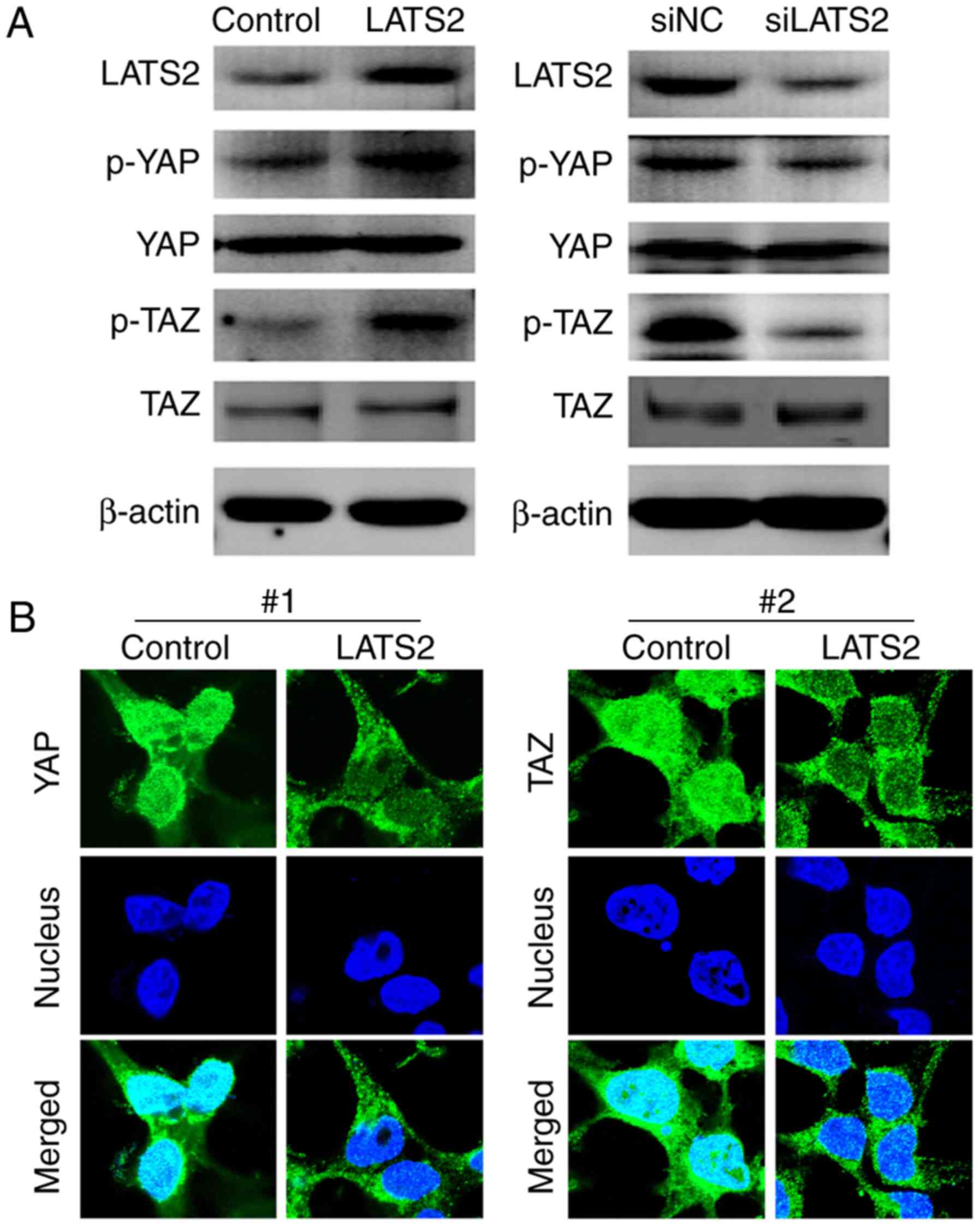
LATS2 regulates phosphorylation of YAP
and TAZ, and their subcellular localization
Since YAP and TAZ are well recognized as downstream
effectors of the Hippo pathway, the present study aimed to
determine whether LATS2 functions through phosphorylating YAP and
TAZ in U-372 MG cells. The phosphorylated and total protein
expression levels of YAP and TAZ were detected in cells transfected
with LATS2 plasmids or LATS2 siRNA. Notably, although alterations
in LATS2 expression had no effect on the total expression levels of
YAP and TAZ, p-YAP and p-TAZ were increased in cells transfected
with LATS2 plasmids and decreased in cells transfected with LATS2
siRNA (Fig. 4A). These findings
indicated that expression of LATS2 may regulate the phosphorylation
of YAP and TAZ. Since it has been reported that YAP and TAZ
phosphorylation promotes their translocation from the nucleus to
the cytoplasm (13), this study
also investigated whether LATS2-induced YAP and TAZ phosphorylation
altered their subcellular localization. Immunostaining revealed
that overexpression of LATS2 induced nuclear to cytoplasm
translocation of p-YAP and p-TAZ (Fig.
4B).
YAP S127A mutant abolishes the
inhibitory effects of LATS2 on cell proliferation, migration and
invasion
Although YAP and TAZ are the main effectors of the
Hippo pathway, LATS2 can operate through other signaling pathways,
such as Wnt signaling (14). In
order to confirm that LATS2 inhibited cell proliferation, migration
and invasion through the YAP/TAZ signaling pathway, a YAP S127A
mutant, which lacks the phosphorylation site recognized by LATS2
(15), was transfected into cells,
which were analyzed by CCK-8, scratch wound healing and transwell
invasion assays. These studies aimed to investigate whether
disrupting YAP phosphorylation abolished the inhibitory effects of
LATS2 on cell proliferation, migration and invasion. Initially, the
present study validated that YAP S127A transfection increased the
total protein expression levels of YAP, but not p-YAP in U-372 MG
cells (Fig. 5A). Compared with
transfection with LATS2 alone, cotransfection with YAP S127A and
LATS2 decreased the expression levels of p-YAP (Fig. 5B). The results of CCK-8, scratch
wound healing and transwell invasion assays consistently revealed
that YAP S127A relieved the suppressive effects of LATS2 on cell
proliferation, migration and invasion (Fig. 5C-E). These findings are consistent
with the hypothesis that LATS2 regulated cell proliferation,
migration and invasion via YAP/TAZ signaling.
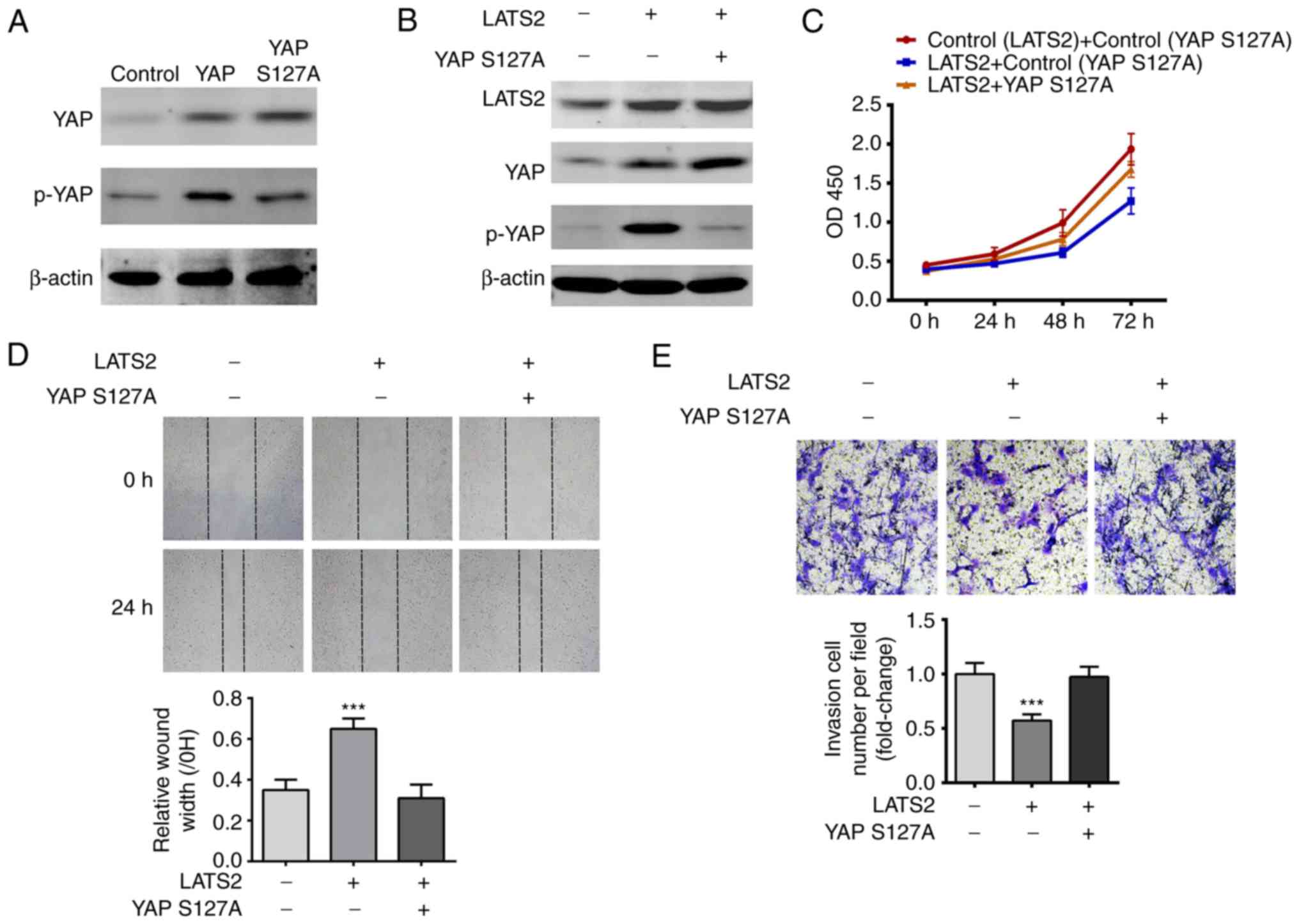 | Figure 5.YAP S127A mutant abolished the effects
of LATS2 on cell proliferation, migration and invasion. (A)
Expression levels of p- and total YAP were determined in wild-type
YAP- and YAP S127A-transfected cells by western blotting. β-actin
was used as an internal reference. Total levels of YAP were
increased in both wild-type YAP- and YAP S127A-transfected cells
compared with the control; however, p-YAP was only increased in
wild-type YAP-transfected cells. (B) YAP S127A abolished
LATS2-induced p-YAP upregulation. Expression levels of LATS2, total
YAP and p-YAP were determined by western blotting. β-actin was used
as an internal reference. (C) YAP S127A abolished LATS2-induced
suppression of cell proliferation. Growth curves were plotted with
the results of the Cell Counting Kit-8 assay, and OD450
was measured at 24, 48 and 72 h. Cell growth was restored by
cotransfection of cells with LATS2 and YAP S127A. (D) YAP S127A
restored the migration of the LATS2-transfected cells. Cell
migration in each group was determined using the scratch wound
healing assay; representative images (magnification, ×200) are
shown. Following cotransfection with YAP S127A and LATS2, cells had
a similar wound repair rate to the control group, whereas
transfection with LATS2 alone resulted in a significantly lower
repair rate compared with cells cotransfected with LATS2 and YAP
S127A. (E) Overexpression of YAP S127A restored the invasive
ability of LATS2-transfected cells. Cell invasion in each group was
determined by transwell invasion assay and the number of invasive
cells was counted (magnification, ×800). Cells cotransfected with
YAP S127A and LATS2 had a similar number of invasive cells to the
control group, whereas transfection with LATS2 alone resulted in
significantly fewer invasive cells compared with the control group.
All data are presented as the means ± standard deviation (n=3).
***P<0.001 compared with cells cotransfected with LATS2 and YAP
S127A. Each assay was performed in triplicate. LATS2, large tumor
suppressor kinase 2; p, phosphorylated; si/siRNA, small interfering
RNA; YAP, yes-associated protein. |
Discussion
The present study revealed the LATS2 was
downregulated in glioma samples compared with in normal tissue
samples, as determined by RT-qPCR and immunohistochemistry.
Subsequently, the effects of LATS2 on cell proliferation, migration
and invasion were detected in vitro using U-372 MG glioma
cells. The results indicated that LATS2 suppressed cell
proliferation, likely by decreasing the expression levels of cyclin
D1, CDK4 and CDK6, which are necessary for G1/S
transition. This result supported previous findings by Li et
al, which reported that LATS2 suppressed cell proliferation by
inhibiting G1/S transition through downregulating cyclin
E/CDK2 kinase in NIH3T3/v-ras cells (16). Notably, dysregulation of the
aforementioned proteins have also been implicated in other types of
human cancer (17), thus suggesting
that LATS2 may inhibit cell proliferation through several pathways.
Increased cell migration and invasion are two important features in
development and oncogenic transformation, which may be associated
with the high invasive ability of gliomas. The present study
demonstrated using scratch wound healing and transwell invasion
assays, that LATS2 also suppressed cell migration and invasion of
U-372 MG cells. Therefore, reduced LATS2 expression in glioma
samples may explain the increased cell migration and
invasiveness.
The present study aimed to determine the underlying
pathway that mediates the effects of LATS2 on cell proliferation,
migration and invasion. The Hippo pathway has been well recognized
as a key player in cellular proliferation, transformation and
tumorigenesis (18), and YAP/TAZ
act as downstream effectors. The present results revealed that
LATS2 knockdown decreased the expression levels of p-YAP/TAZ. This
finding is consistent with other findings, which reported that
downregulation of LATS2 alters phosphorylation of YAP/TAZ, and in
turn increases YAP and TAZ nuclear accumulation; these proteins are
pervasively activated in human malignancies (6). These findings suggested that LATS2 may
act as a tumor suppressor in glioma, as well as in other cancers
(14,19).
Glioma is a frequent primary CNS tumor, with an
average annual age-adjusted incidence rate of 6.0/100,000 in the
United States (20). Due to its
highly invasive ability, it is considered an intractable disease in
the clinic with a very short median survival time (20). Therefore, there is an urgent
requirement to further study the mechanism of tumorigenesis and
develop novel treatment strategies. Targeted therapy that inhibits
oncogenic signaling pathways and tumor-associated angiogenesis
exerts impressive effects in the clinic (21). It has previously been reported that
dysregulation of the Hippo pathway is heavily involved in the
initiation and progression of various types of cancer (22). The Hippo pathway is an
evolutionarily conserved mechanism, which serves a crucial role in
cell proliferation, apoptosis, differentiation and development. The
core component of the mammalian Hippo pathway is the MST1/2 and
LATS1/2 kinase cascade. Activation of the kinase cascade induces
translocation of YAP and TAZ from the nucleus to the cytoplasm and
degradation, which suppresses the expression of genes that promote
proliferation and inhibit apoptosis. Therefore, LATS2 is a
potential tumor suppressor that may be able to regulate several
oncogenes. Low expression of LATS2, or LATS2 copy number loss has
been identified in breast, ovarian, hepatocellular and lung cancer
(23). The present study confirmed
the downregulation of LATS2 expression in glioma. The functions and
downregulation of LATS2 in various cancers indicates that it may
serve an important role in glioma formation and progression.
Notably, studies also revealed that low expression of LATS2
predicts a better outcome for patients with cancer (24,25).
One possible reason for the contradictory function of LATS2 may be
the crosstalk of LATS2 with other signaling pathways. The roles of
LATS2 in tumorigenesis remain unclear; LATS2 may potentially
function as a tumor suppressor, not only by regulating YAP and TAZ
phosphorylation as indicated in this study, but also by suppressing
oncogenic Wnt signaling through disrupting β-catenin/B cell
CLL/lymphoma 9 interaction, as shown in a previous study (12); conversely, LATS2 has been reported
to promote the stability of p53 via inactivation of MDM2
proto-oncogene (26). However, p53
mutations have been detected in the majority of cancers; in this
case, lower expression of LATS2 may decrease the transcriptional
activity of mutant p53 and further decrease the proliferation,
invasion and migration of the cancer cells. Therefore,
heterogeneity in cytology and gene expression should be taken into
account when exploring tumorigenic mechanisms.
The present results confirmed that LATS2 was
downregulated in glioma samples. It was also revealed that
overexpression of LATS2 suppressed proliferation, migration and
invasion of U-372 MG cells by altering YAP/TAZ phosphorylation and
nuclear translocation. These results highlight the potential
function of LATS2 as a tumor suppressor in glioma, and its reduced
expression in glioma may be a contributing factor to cancer
progression.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CG and CL were involved in conception and design of
the study. JY acquired the data. HH, XL and BF analyzed the data
and drafted the manuscript. CL gave final approval of the version
to be published. All authors revised, read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of The Second Affiliated Hospital of Hebei Medical
University, and all patients provided written informed consent.
Patient consent for publication
All patients provided written informed consent.
Competing interests
There is no conflict of interest to declare.
References
|
1
|
Dellaretti M, Reyns N, Touzet G, Dubois F,
Gusmao S, Pereira JL and Blond S: Diffuse brainstem glioma:
Prognostic factors. J Neurosurg. 117:810–814. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Grossman SA, Ye X, Piantadosi S, Desideri
S, Nabors LB, Rosenfeld M and Fisher J; NABTT CNS Consortium, :
Survival of patients with newly diagnosed glioblastoma treated with
radiation and temozolomide in research studies in the United
States. Clin Cancer Res. 16:2443–2449. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang Y and Jiang T: Understanding high
grade glioma: Molecular mechanism, therapy and comprehensive
management. Cancer Lett. 331:139–146. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Baker GJ, Yadav VN, Motsch S, Koschmann C,
Calinescu AA, Mineharu Y, Camelopiragua S, Orringer D, Bannykh S,
Nichols WS, et al: Mechanisms of glioma formation: Iterative
perivascular glioma growth and invasion leads to tumor progression,
VEGF-independent vascularization, and resistance to antiangiogenic
therapy. Neoplasia. 16:543–561. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Meng Z, Moroishi T and Guan KL: Mechanisms
of Hippo pathway regulation. Genes Dev. 30:1–17. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zanconato F, Cordenonsi M and Piccolo S:
YAP/TAZ at the roots of cancer. Cancer Cell. 29:783–803. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mo J, Park HW and Guan KL: The Hippo
signaling pathway in stem cell biology and cancer. EMBO Rep.
15:642–656. 2014.PubMed/NCBI
|
|
8
|
Wang Y, Pan P, Wang Z, Zhang Y, Xie P,
Geng D, Jiang Y, Yu R and Zhou X: β-catenin-mediated YAP signaling
promotes human glioma growth. J Exp Clin Cancer Res. 36:1362017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bhat KP, Salazar KL, Balasubramaniyan V,
Wani K, Heathcock L, Hollingsworth F, James J, Gumin J, Diefes K,
Kin SH, et al: The transcriptional coactivator TAZ regulates
mesenchymal differentiation in malignant glioma. Genes Dev.
25:2594–2609. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Grzmil M, Morin P Jr, Lino MM, Merlo A,
Frank S, Wang Y, Moncayo G and Hemmings BA: MAP kinase-interacting
kinase 1 regulates SMAD2-dependent TGF-β signaling pathway in human
glioblastoma. Cancer Res. 71:2392–2402. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guo C, Wang X and Liang L: LATS2-mediated
YAP1 phosphorylation is involved in HCC tumorigenesis. Int J Clin
Exp Pathol. 8:1690–1697. 2015.PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Piccolo S, Dupont S and Cordenonsi M: The
Biology of YAP/TAZ: Hippo signaling and beyond. Physiol Rev.
94:1287–1312. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li J, Chen X, Ding X, Cheng Y, Zhao B, Lai
ZC, Al-Hezaimi K, Hakem R, Guan KL and Wang CY: LATS2 suppresses
oncogenic Wnt signaling by disrupting β-catenin/BCL9 interaction.
Cell Rep. 5:1650–1663. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhao B, Wei X, Li W, Udan RS, Yang Q, Kim
J, Xie J, Ikenoue T, Yu J, Li L, et al: Inactivation of YAP
oncoprotein by the Hippo pathway is involved in cell contact
inhibition and tissue growth control. Genes Dev. 21:2747–2761.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li Y, Pei J, Xia H, Ke H, Wang H and Tao
W: Lats2, a putative tumor suppressor, inhibits G1/S transition.
Oncogene. 22:4398–4405. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bonelli P, Tuccillo FM, Borrelli A,
Schiattarella A and Buonaguro FM: CDK/CCN and CDKI alterations for
cancer prognosis and therapeutic predictivity. Biomed Res Int.
2014:3610202014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yu FX, Zhao B and Guan KL: Hippo pathway
in organ size control, tissue homeostasis, and cancer. Cell.
163:811–828. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Visser S and Yang X: LATS tumor
suppressor: A new governor of cellular homeostasis. Cell Cycle.
9:3892–3903. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ostrom QT, Kinnersley B, Wrensch MR,
Eckel-Passow JE, Armstrong G, Rice T, Chen YW, Wiencke J, McCoy LS,
Hansen HM, et al: Sex-specific glioma genome-wide association study
identifies new risk locus at 3p21.31 in females, and finds
sex-differences in risk at 8q24.21. Sci Rep. 8:73522018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hughes PE, Caenepeel S and Wu LC: Targeted
therapy and checkpoint immunotherapy combinations for the treatment
of cancer. Trends Immunol. 37:462–476. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Moroishi T, Hansen CG and Guan KL: The
emerging roles of YAP and TAZ in cancer. Nat Rev Cancer. 15:73–79.
2015. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Furth N and Aylon Y: The LATS1 and LATS2
tumor suppressors: Beyond the Hippo pathway. Cell Death Differ.
24:1488–1501. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gao Y, Yi J, Zhang K, Bai F, Feng B, Wang
R, Chu X, Chen L and Song H: Downregulation of MiR-31 stimulates
expression of LATS2 via the hippo pathway and promotes
epithelial-mesenchymal transition in esophageal squamous cell
carcinoma. J Exp Clin Cancer Res. 36:1612017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang Y, Hu CF, Chen J, Yan LX, Zeng YX
and Shao JY: LATS2 is de-methylated and overexpressed in
nasopharyngeal carcinoma and predicts poor prognosis. BMC Cancer.
10:5382010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Aylon Y, Michael D, Shmueli A, Yabuta N,
Nojima H and Oren M: A positive feedback loop between the p53 and
Lats2 tumor suppressors prevents tetraploidization. Genes Dev.
20:2687–2700. 2006. View Article : Google Scholar : PubMed/NCBI
|