Introduction
Gastric cancer (GC) is the fourth most common
diagnosed cancer worldwide and ranks second as the cause of
cancer-related death (1). The
etiology of GC is complicated and remains an active focus of
research. In addition to environmental risk factors, genetics may
play a role in the development and progression of the disease
(2,3). Many studies have explored the genetic
mechanisms underlying GC, including the direct effects of
protein-coding genes, e.g. TP53, HOXB5, TGF-β, SK and
THBS1 (4–8), and the indirect effects of non-coding
RNAs (ncRNAs), e.g., let-7a, miR-21 and SNGH8 (9,10).
Recently, a new and largest class (11) of non-coding small RNAs, i.e.,
piwi-interacting RNAs (piRNAs), which interact with a subset of
Argonaute proteins related to Piwi, has become a focus of intense
research (12). piRNAs are
non-coding RNAs with 24–32 nucleotides (nt). Although structurally
only slightly longer than microRNAs (miRNAs; 19–25 nt), piRNAs are
different in expression pattern and quantity, as well as in genomic
organization (13–15). piRNAs are initially discovered in
germline cells. Germline and certain ‘lower’ eukaryotic organisms
are ‘non-aging’ biological systems that display an indefinite
capacity for renewal and a lifespan-stable genome integrity, and
are potentially immortal (16).
piRNA is a shared feature of all non-aging biological systems
(16). Interestingly, cancer stem
cells also display an indefinite capacity of renewal and
proliferation and are potentially immortal (16). An increasing number of studies have
shown aberrant piRNA expression as a molecular signature across
multiple tumor types (17,18), including renal cancer (19), breast cancer (14,20),
multiple myeloma (21), and
hepatocellular carcinoma (22),
suggesting that piRNAs may be involved in the pathogenesis of
cancer (12). For instance,
upregulated expression of piR-36743, piR-20365, piR-20485, piR-4987
and piR-932 was observed in breast cancer as compared to matched
non-cancerous tissues (23,24); and the expression of piRABC was
increased in bladder cancer, as compared to their corresponding
adjacent non-cancerous tissues (25). These findings also suggest that
piRNAs are a shared feature of this non-aging biological system,
cancers. In an earlier work, we reported associations between
piRNAs and Alzheimer's disease (26), a clinical condition strongly related
to aging. Cancers are just opposite to the aging diseases in
renewal and proliferation capacity, which also suggests a potential
role of piRNAs in cancers. Recently, a study reported that 156
piRNAs were significantly deregulated in GC (27).
The sequences of piRNAs primarily complement the
transposable elements (TEs). The most widely recognized and
well-characterized function of piRNAs is to suppress the activities
of TEs at genomic and epigenetic levels (17). TEs are capable of moving from one
genomic locus to another, thereby causing insertional mutations
(28,29). There are two major classes of TEs:
RNA transposons (or retrotransposons) and DNA transposons, as
distinguished by their mechanism of transposition (28). Retrotransposons include long
terminal repeats (LTRs), long interspersed nuclear element (LINE),
and short interspersed nuclear elements (SINE). As the primary
determinants of genomic instability (16), TEs exert pathological effects via
controlling the transcription of neighboring genes (30). The downregulation of piRNA pathways
would increase the repeats of retrotransposon and cause DNA damage,
which represents the primary genetic determinant of aging. The
upregulation of piRNAs suppresses the activity of TEs, which
protects against TE-mediated mutagenesis, maintains genomic
integrity, and self-renewal and proliferation capacity, and thereby
preserves the non-aging features of cancer cells (16). Additionally, piRNA may also be
involved in a TE-independent mechanism in cancer risk; that is, it
may regulate the transcription of certain key cancer-related genes
via non-TE sequence complementarity (16,31) or
modulation of chromatin organization (16). In the present study, we aimed to
identify the piRNAs in association with GC across the transcriptome
using a piRNA profile. Further, we explored whether these
GC-associated piRNAs are expressed in other tissues, and whether
their nearest protein-coding genes were expressed in the stomach
and perhaps even related to gastrointestinal cancers.
For many years investigators have scanned the whole
genome to search for risk DNA variants for GC. In reviewing all
published genome-wide association studies (GWASs) and whole
genome/exome sequencing studies of GC, we found that only three
variants were genome-wide significant in association with GC and
replicated across at least two independent studies at single-point
level. The three variants are rs4072037 at MUC1 (on Chr1)
(32–35), rs13361707 at PRKAA1 (on Chr5)
(34,36) and rs2294008 at PSCA (on Chr8)
(34,35). Numerous candidate gene studies
supported these GWAS findings. However, the mechanisms underlying
these SNP-GC associations are not clear. Here we examined whether
the GC-related piRNAs might mediate the SNP-GC associations, in
order to explore the potential roles of piRNAs in the pathogenesis
of GC.
Materials and methods
Subjects
The primary cohort including eight gastric
cancer(GC) tissues and adjacent normal tissues extracted from eight
male patients (aged 49–79 years old) was examined in a piRNA
microarray study. The clinical characteristics of the GC patients
are shown in Table I. Fresh gastric
tissue samples were collected from GC patients who underwent
surgical resection at the Fujian Provincial Cancer Hospital. A
second cohort of 20 paired GC and adjacent normal tissues extracted
from 15 male and 5 female patients (aged 32–78 years old) was also
obtained from the same hospital for validating the findings
obtained from the first cohort. A third cohort of 181 paired GC and
adjacent normal tissues extracted from 134 male and 47 female
patients (aged 28–73 years old) from the same hospital was tested
for piwi protein expression in the stomach. These cohorts were
recruited between June 2015 and February 2018. None of the patients
received chemotherapy before surgery and tissues were obtained
immediately after tumor resection. Medical diagnoses were confirmed
histopathologically, and clinical pathological data were obtained.
Histological classification of GC was assessed according to the
Lauren's criteria (37). Patients
were staged according to the 7th Edition of the TNM staging system
of the American Joint Commission on Cancer (AJCC) (38). The study was approved by the
Research Ethics Committee of the Fujian Provincial Cancer Hospital,
China. Informed consents from all patients were obtained prior to
the study. All experiments were performed in accordance with
relevant guidelines and regulations.
 | Table I.Pathological features of the subjects
with gastric carcinoma. |
Table I.
Pathological features of the subjects
with gastric carcinoma.
| Parameters | Discovery samples
(n) (for sequencing) | Validation samples
(n) (for qPCR) |
|---|
| Sex |
|
|
|
Male | 8 | 15 |
|
Female | 0 | 5 |
| Age (years) |
|
|
|
<60 | 4 | 9 |
|
≥60 | 4 | 11 |
| Location |
|
|
|
EGJ | 4 | 6 |
|
Non-EGJ | 4 | 14 |
| Depth of
invasion |
|
|
|
<T2 | 0 | 3 |
|
≥T2 | 8 | 17 |
| Lauren's type |
|
|
|
Intestinal-type | 4 | 6 |
|
Diffuse-type | 4 | 14 |
| Tumor size
(cm) |
|
|
|
<5 | 4 | 10 |
| ≥5 | 4 | 10 |
| TNM stage |
|
|
|
I+II | 3 | 6 |
|
III+IV | 5 | 14 |
| LN metastasis |
|
|
|
Absent | 2 | 6 |
|
Present | 6 | 14 |
piRNA microarray experiment
The ArrayStar HG19 piRNA array was used to profile
piRNA expression in humans (ArrayStar; Agilent Technologies, Santa
Clara, CA, USA). This array contains probes for 23,677 piRNAs
obtained from the National Center for Biotechnology Information
(NCBI) database (https://www.ncbi.nlm.nih.gov/nuccore/?term=piRNA)
and landmark publications (15,39).
Human piRNAs were mapped to the HG19 genome sequence using UCSC
Blat (https://genome.ucsc.edu/cgi-bin/hgBlat?command=start).
Total RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad,
CA, USA) according to manufacturer's instructions. Sample labeling
was performed using an RNA ligase method (40). The labeled samples were hybridized
onto Arraystar Human piRNA Array in Agilent's SureHyb Hybridization
Chambers according to manufacturer's standard protocols (Agilent
Technologies). After slides were scanned, data were extracted using
Agilent Feature Extraction software. Raw signal intensities were
normalized in quantiles by GeneSpring GX v11.5.1 (Agilent
Technologies), and low intensity piRNAs were filtered. Agilent
Feature Extraction software (version 11.0.1.1) was used to analyze
the acquired array images (40).
Normalized intensity values were then
log2-transformed.
Quality assessment of piRNA data after
normalization and filtering
We used a Box Plot to visualize the distributions of
a dataset. We compared the distributions of the intensities from
all samples and showed that. after normalization, the distributions
of log2-ratios among the samples were nearly the same (Fig. S1). We used a Scatter Plot to assess
the piRNA expression variation (or reproducibility) between arrays
(Fig. S2). This plot showed good
reproducibility of our array data.
The distribution of all of the observed P-values
from the differential expression analysis (see below) was fitted to
the expected P-values using a QQ plot. A transcriptomic inflation
factor (λ) was computed from these P-values and defined as the
ratio of the median of the empirically observed distribution of the
test statistic to the expected median, thus quantifying the extent
of the bulk inflation and the excess false positive rate. It was
computed for the genomic control analysis to reflect the maximum
possible inflation factor if the associations were affected by
population stratification. A λ with a departure of less than 0.1
from a value of 1.0 is considered an indicator of very good quality
and robust associations.
Differential expression analysis
The normalized and log2-transformed intensities were
compared between GC cases and normal controls by paired Student's
t-test. Fold-changes (FCs) between the two groups were calculated
by ‘normalized intensity values in Group 1 divided by Group 2’.
Significantly differentially expressed piRNAs, which were
associated with GC, were identified through Volcano Plot filtering
(FC≥1.5 and P≤0.05). The expression level of each piRNA in each
individual was also shown in a Heat Map, and clustered using
Hierarchical clustering analysis to show the relationships among
the expression levels of samples.
Quantitative reverse
transcription-polymerase chain reaction (qRT-PCR) experiment
We selected some candidate piRNAs from the top list
of the significantly differentially expressed piRNAs to be examined
by qPCR in an independent cohort (20 cases vs. 20 controls), in
order to control the quality of microarray experiment and validate
piRNA-GC associations as derived from the differential expression
analysis on the array data. The selection criteria for these
candidate piRNAs include that: i) their expression in gastric
cancer tissues should have top intensities >1,500 in the array
data; ii) their expression in gastric cancer tissues should be
higher than adjacent tissues with top FCs ≥2.4; and iii) they are
located at or close to those genes that have been reported by
literature to be cancer-related. After this selection, only two
piRNAs, i.e., DQ579739 and DQ600515, were included (Table II). Both piRNAs were abundantly
expressed in gastric cancer tissues [log2(normalized intensity)
>10], and differentially expressed between cases and controls in
the array data (P<0.05).
| Table II.Top piRNAs significantly
differentially expressed between the cases and controls. |
Table II.
Top piRNAs significantly
differentially expressed between the cases and controls.
|
|
| Normalized
intensity |
| Cancer vs.
control |
|---|
|
|
|
|
|
|
|---|
| piRNA | Length (nt) | Chr | Cancer | Control | FC | P-value | Gene |
|---|
| With top
intensities in cancer (intensities >1,500) |
|
DQ588779a | 29 | chr1 | 14,194 | 6,844 | 2.1 | 0.037 | SKI |
|
DQ595533a | 29 | chr3 | 13,314 | 6,148 | 2.2 | 0.035 | COLQ |
| DQ571813 | 31 | chr16 | 12,123 | 5,710 | 2.1 | 0.046 | VAC14 |
| DQ590386 | 29 | chr16 | 11,953 | 5,766 | 2.1 | 0.046 | VAC14 |
| DQ595534 | 30 | chr16 | 11,307 | 5,429 | 2.1 | 0.038 | VAC14 |
|
DQ600670a | 30 | chr11 | 4,410 | 1,805 | 2.4 | 0.032 | EED |
|
DQ600515a | 29 | chr1 | 4,071 | 1,668 | 2.4 | 0.041 | ATAD3B |
|
DQ580665a | 26 | chr7 | 2,349 | 5,133 | 2.2 | 0.041 | POLR2J |
|
DQ579739a | 30 | chr19 | 1,844 | 774 | 2.4 | 0.029 | CXCL17 |
| DQ578739 | 31 | chr2 | 1,709 | 3,422 | 2.0 | 0.007 | to
LOC100507334 |
| DQ583443 | 30 | chr9 | 1,582 | 3,683 | 2.3 | 0.015 | FAM225A |
| With top FC
between cases and controls (FC≥2.4) |
|
DQ590922a | 29 | chr9 | 32 | 10 | 3.1 | 0.036 | to TLE4 |
|
DQ572848a | 30 | chr9 | 22 | 8 | 2.7 | 0.024 | to TLE4 |
|
DQ600670a | 30 | chr11 | 4,410 | 1,805 | 2.4 | 0.032 | EED |
|
DQ600515a | 29 | chr1 | 4,071 | 1,668 | 2.4 | 0.041 | ATAD3B |
|
DQ579739a | 30 | chr19 | 1,844 | 774 | 2.4 | 0.029 | CXCL17 |
| DQ571419 | 28 | chr1 | 1,443 | 605 | 2.4 | 0.032 | RPS8 |
| With top FC ↓
between cases and controls (FC≥3.8) |
| DQ574148 | 30 | chr22 | 25 | 110 | 4.4 | 0.012 | IGLL1 |
| DQ583043 | 30 | chr19 | 32 | 135 | 4.2 | 0.015 | to
LOC100652909 |
| DQ574145 | 30 | chr22 | 18 | 75 | 4.2 | 0.01 | IGLL1 |
| DQ574146 | 30 | chr22 | 49 | 197 | 4.1 | 0.012 | IGLL1 |
| DQ598137 | 28 | chr10 | 15 | 58 | 4.0 | 0.017 | NUTM2A |
| DQ570015 | 30 | chr15 | 429 | 1,642 | 3.8 | 0.036 | GOLGA8A |
| DQ574147 | 31 | chr22 | 55 | 208 | 3.8 | 0.010 | IGLL1 |
| With lowest
P-values between cases and controls and FC ↑ (P≤0.025) |
|
DQ570687a | 28 | chr22 | 910 | 448 | 2.0 | 0.012 | RPL3 |
| DQ580689 | 28 | chr1 | 180 | 90 | 2.0 | 0.016 | NBPF3 |
|
DQ575884a | 32 | chr2 | 1,393 | 691 | 2.0 | 0.017 | to
EPCAM |
|
DQ572848a | 30 | chr9 | 22 | 8 | 2.7 | 0.024 | to TLE4 |
| With lowest
P-values between cases and controls and FC ↓ (P≤0.005) |
| DQ577345 | 31 | chr19 | 383 | 969 | 2.5 | 0.003 | to
LOC400685 |
| DQ587262 | 31 | chr6 | 97 | 233 | 2.4 | 0.003 | to
LOC100507584 |
| DQ575557 | 29 | chr3 | 25 | 83 | 3.4 | 0.003 | to
IQSEC1 |
| DQ594511 | 29 | chr8 | 22 | 45 | 2.0 | 0.004 | to
LOC100287846 |
| DQ594440 | 31 | chr3 | 60 | 164 | 2.7 | 0.004 | to
IQSEC1 |
| DQ576821 | 28 | chr19 | 28 | 77 | 2.7 | 0.004 | ZNF490 |
| DQ570858 | 31 | chr15 | 30 | 106 | 3.5 | 0.005 | HERC2P7 |
| With locations
at or close to cancer-related genes |
|
DQ600515a | 29 | chr1 | 4,071 | 1,668 | 2.4 | 0.041 | ATAD3B |
| DQ594797 | 26 | chr2 | 14 | 7 | 2.1 | 0.042 | to
TEKT4 |
| DQ580529 | 30 | chr22 | 13 | 30 | 2.3 | 0.022 | to
WBP2NL |
|
DQ579739a | 30 | chr19 | 1,844 | 774 | 2.4 | 0.029 | CXCL17 |
|
DQ570687a | 28 | chr22 | 910 | 448 | 2.0 | 0.012 | RPL3 |
|
DQ600670a | 30 | chr11 | 4,410 | 1,805 | 2.4 | 0.032 | EED |
|
DQ590922a | 29 | chr9 | 32 | 10 | 3.1 | 0.036 | to TLE4 |
|
DQ595533a | 29 | chr3 | 13,314 | 6,148 | 2.2 | 0.035 | COLQ |
|
DQ588779a | 29 | chr1 | 14,194 | 6,844 | 2.1 | 0.037 | SKI |
| DQ588880 | 30 | chr17 | 30 | 67 | 2.2 | 0.032 | to
HOXB5 |
|
DQ580665a | 26 | chr7 | 2,349 | 5,133 | 2.2 | 0.041 | POLR2J |
| DQ590827 | 30 | chr16 | 179 | 367 | 2.0 | 0.037 | DHODH |
|
DQ575884a | 32 | chr2 | 1,393 | 691 | 2.0 | 0.017 | to
EPCAM |
| DQ580779 | 28 | chr9 | 61 | 184 | 3.0 | 0.035 | TMOD1 |
| DQ600254 | 29 | chr14 | 144 | 354 | 2.5 | 0.027 | PNMA1 |
To measure the piRNA expression levels, a miScript
Reverse Transcription (RT) kit (Guangzhou RiboBio Co., Ltd.,
Guangzhou, China) was used to generate cDNA, according to the
manufacturer's instructions (40).
Next, qRT-PCR was performed in a 20 µl reaction volume with the
following reagents (Guangzhou RiboBio Co., Ltd): SYBR-Green Mix [10
µl of SYBR-Green I mix, 0.5 µl of miDETECTATrack™ miRNA forward and
reverse primers (10 µM), and 2 µl of cDNA template] on the ABI7500
system (Applied Biosystems; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) with the following protocol: 95°C for 3 min and
40 cycles of 95°C for 10 sec, 60°C for 20 sec, and 70°C for 1
sec.
Using the comparative 2−ΔΔCq method
(41) and an example gastric
carcinoma sample as a calibrator, the relative expression levels in
all gastric carcinoma samples and adjacent non-tumorous stomach
tissues were quantified. Hs_5S_miScript expression was used to
normalize piRNA expression levels between different samples.
Risk gene expression in the
stomach
The primary function of piRNAs is to suppress TEs
that will then control the transcription of nearby genes. piRNAs
affect the phenotypes primarily via TE suppression. The target TEs
that are potentially regulated by the risk piRNAs were searched
from UCSC Genome Browser (http://genome.ucsc.edu; genome assembly, hg19; track,
RepeatMasker). We hypothesized that, without prior knowledge, the
nearest genes to which the sequences of the piRNAs are
complementary should be the most likely targets for TE regulation
(30,31). These genes were thus carefully
examined in this study. The relationship between the genes and GC
or related cancers was searched in PubMed (Table SI). The mRNA expression of these
genes in stomach was examined in two independent cohorts without
cancers, including an American cohort (n=262) (42), and a UK European cohort (n=176)
(43). The density of the proteins
encoded by these genes was also examined in stomach of a Germany
cohort (44). If a gene had a
normalized intensity >36 (by RNA microarray), a TPM >1 (by
RNA-Seq) or a value >10 ppm (by mass spectrometry-based
proteomics microarray) in the expression levels in human stomach of
these three cohorts or had been reported to be expressed in human
stomach in the literature, it was marked ‘E: Expressed’ (Table SI). The detailed demographic data
of these three cohorts and the expression values of the top four
genes are shown in Table SII.
Overall, the genes expressed in stomach are more likely to be
implicated in stomach diseases than non-stomach genes. Of note, the
expression of these genes was not examined in the primary cohort,
which represented a limitation of the study.
The piwi protein expression in the
stomach
piRNAs regulate cellular activities by interacting
with piwi proteins to form a complex (17). The golden criterion to confirm the
presence of piRNAs is the existence of piwi proteins in the same
tissue. To test the expression of piwi in stomach, the mRNA and
protein expression of piwi, including PIWIL1, PIWIL2, PIWIL3 and
PIWIL4 was examined in the above three independent cohorts,
including the American (42), UK
European (43) and Germany cohorts
(44). The most abundant piwi
protein (here, PIWIL4) was further tested in our own samples (181
GC and 181 adjacent tissues), using microarray and
immunohistochemistry (IHC) technologies (see Supplementary
Methods).
Expression quantitative trait locus
(eQTL) analysis
The 3 genome-wide significant
(P<5×10−8) risk variants for GC were genotyped in the
primary cohort of 16 subjects (8 cases and 8 controls). The
associations between these risk variants and GC have been
replicated by at least two independent genome-wide association
studies (GWASs) (Table SIII). To
examine whether the GC-associated piRNA expression was controlled
by any of these GC-risk SNPs, we performed eQTL analysis. Usually,
a piRNA controlled by DNA variants is more likely to be functional
than those uncontrolled. Associations between the genotypes and
expression level of each GC-associated piRNA were analyzed using
t-test or one-way ANOVA. This correlation is assumed to potentially
underlie SNP-disease associations. The design of the whole study
was based on a regulation pathway as illustrated in Fig. S3.
Results
Differential expression of piRNAs
between cases and controls (Figs. S4,
1 and 2, and Tables II and
SI)
Among the 23,677 piRNAs, 8,759 (37.0%) were detected
in the human stomach. The expression levels of these 8,759 piRNAs
in each individual are shown in the HeatMap (Fig. S4). A total of 8,759 P-values were
obtained from the 8,759 differential expression analyses. The
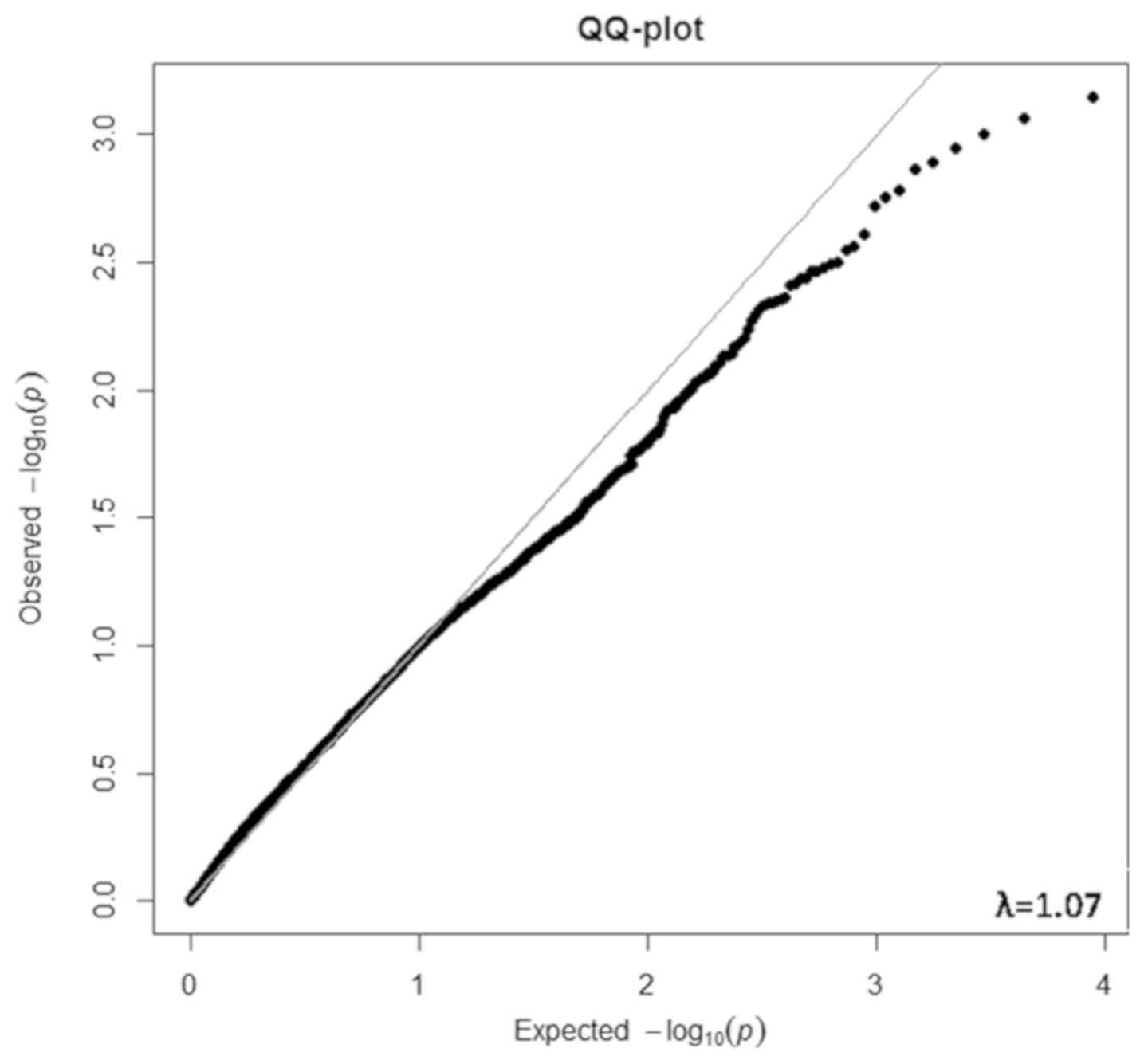
observed and expected P-values fitted very well (see QQ plot in
Fig. 1). The λ was 1.07, indicating
that the array data were of high quality, the differential
expression analysis was robust, and transcriptome-wide inflation
and excess false positive rates in the differential expression
analysis were very low. That is, the associations were less likely
to be affected by population stratification.
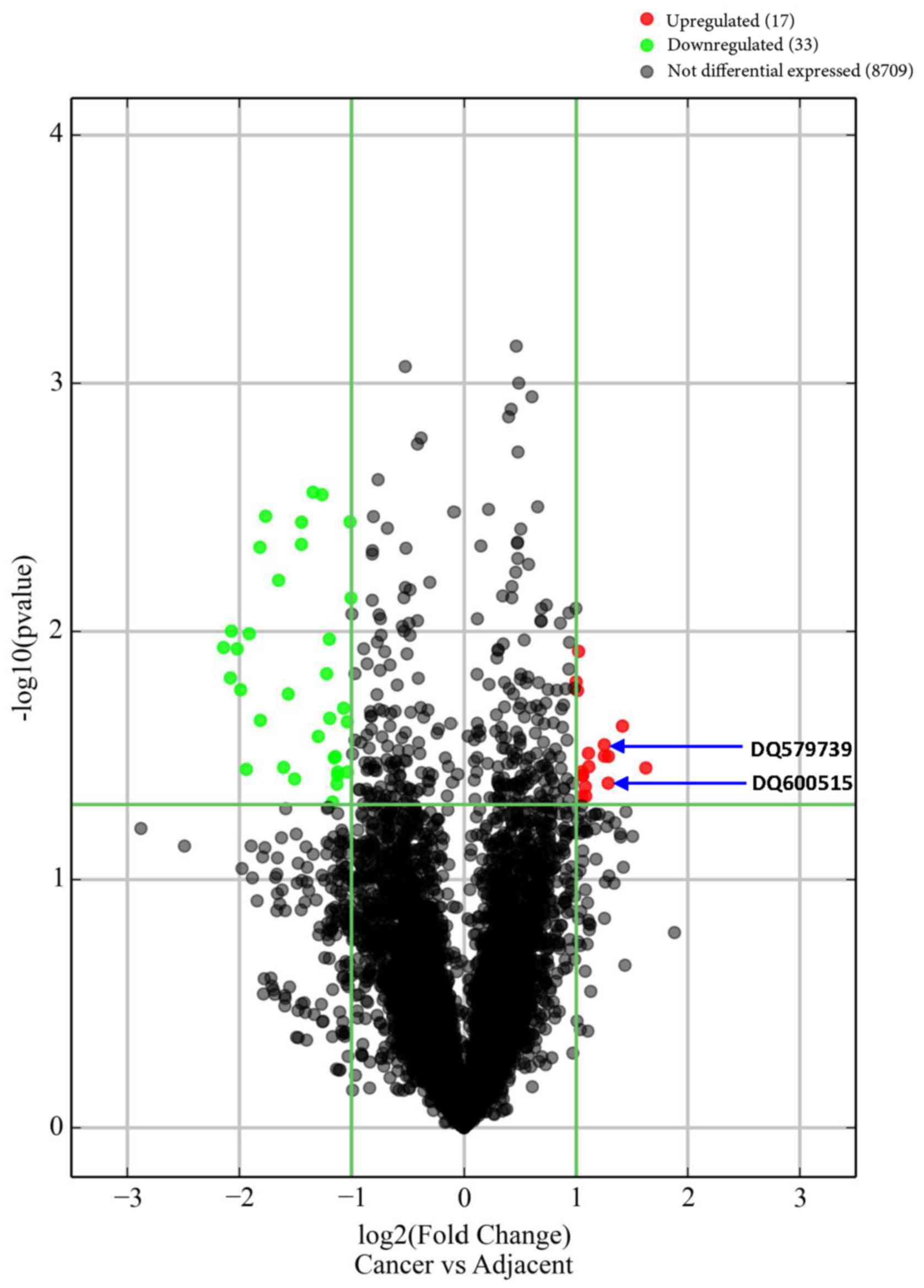
Fifty piRNAs with length of 26–32 nt were nominally
differentially expressed between cases and controls (FC>2.0;
P<0.05; without Bonferroni correction) (Fig. 2; Table
SI). The mean log2-transformed normalized intensity of these 50
piRNAs was 7.46±3.34 (3.21–13.79) in GC cases, and 8.02±2.97
(2.75–12.74) in controls. Among the 50 risk/protective piRNAs, 17
were upregulated and 33 were downregulated in cases in contrast to
controls. Among the 50 piRNAs, 41 risk piRNAs potentially targeted
at 41 TEs, including 1 DNA transposon, 20 SINEs, 12 LINEs and 8
LTRs (Table SI). Thirty piRNAs
mapped within 23 protein-coding genes, 13 mapped close to 11
protein-coding genes, and the other 7 mapped close to 5 ncRNAs.
Twenty-two of these 34 (23+11) protein-coding genes (64.7%) were
expressed in stomach, and 18 piRNAs were located in 15 piRNA
clusters, 10 of which were located in intergenic regions. Seventeen
(77.2%) of these 22 protein-coding stomach genes have been related
to various human cancers in the literature, including gastric
cancer (6–8), colon cancer (45–50),
colorectal cancer (51–53), pancreatic cancer (54–56),
hepatocellular carcinoma (57–59)
and bladder cancer (60) (Table SI).
Thirteen piRNAs had log2-transformed normalized
intensities >10 (i.e., >1,000 before transformation). The top
five piRNAs with highest intensities in cases were DQ588779 at
SKI (SKI proto-oncogene) (on chr1), DQ595533 at COLQ
(on chr3), and DQ571813, DQ590386 and DQ595534 at VAC14 (on
chr16) (Table II).
Two piRNAs, i.e., DQ590922 and DQ572848 close to
TLE4 (on chr9), were expressed with >2.5 FCs in cases compared
to controls. Nine piRNAs were expressed with >3.5 FCs in
controls compared to cases; and the top five of them were DQ574148,
DQ574145 and DQ574146 at IGLL1 (on chr22), DQ598137 at
NUTM2A (on chr10), and DQ583043 close to LOC100652909
(on chr19) (Table II).
Ten piRNAs were significantly differentially
expressed between cases and controls with P<0.01. The four most
significant ones with higher FCs in cases were DQ570687 at
RPL3 (on chr22), DQ580689 at NBPF3 (on chr1),
DQ575884 close to EPCAM (on chr2) and DQ572848 close to
TLE4 (on chr9) (0.012≤P≤0.024). The five most significant
ones with lower FCs in cases were all intergenic, including
DQ577345 close to LOC400685 (chr19), DQ587262 close to
LOC100507584 (chr6), DQ594511 close to LOC100287846
(chr8), and DQ575557 and DQ594440 close to IQSEC1 (chr3)
(0.003≤P≤0.004) (Table
II).
Some genes within or near which the GC-associated
piRNAs are located have been reported to be associated with
gastrointestinal cancers. They are CXCL17, EPCAM, EED, TLE4,
SKI, RPL3 and HOXB5 (Table
SI).
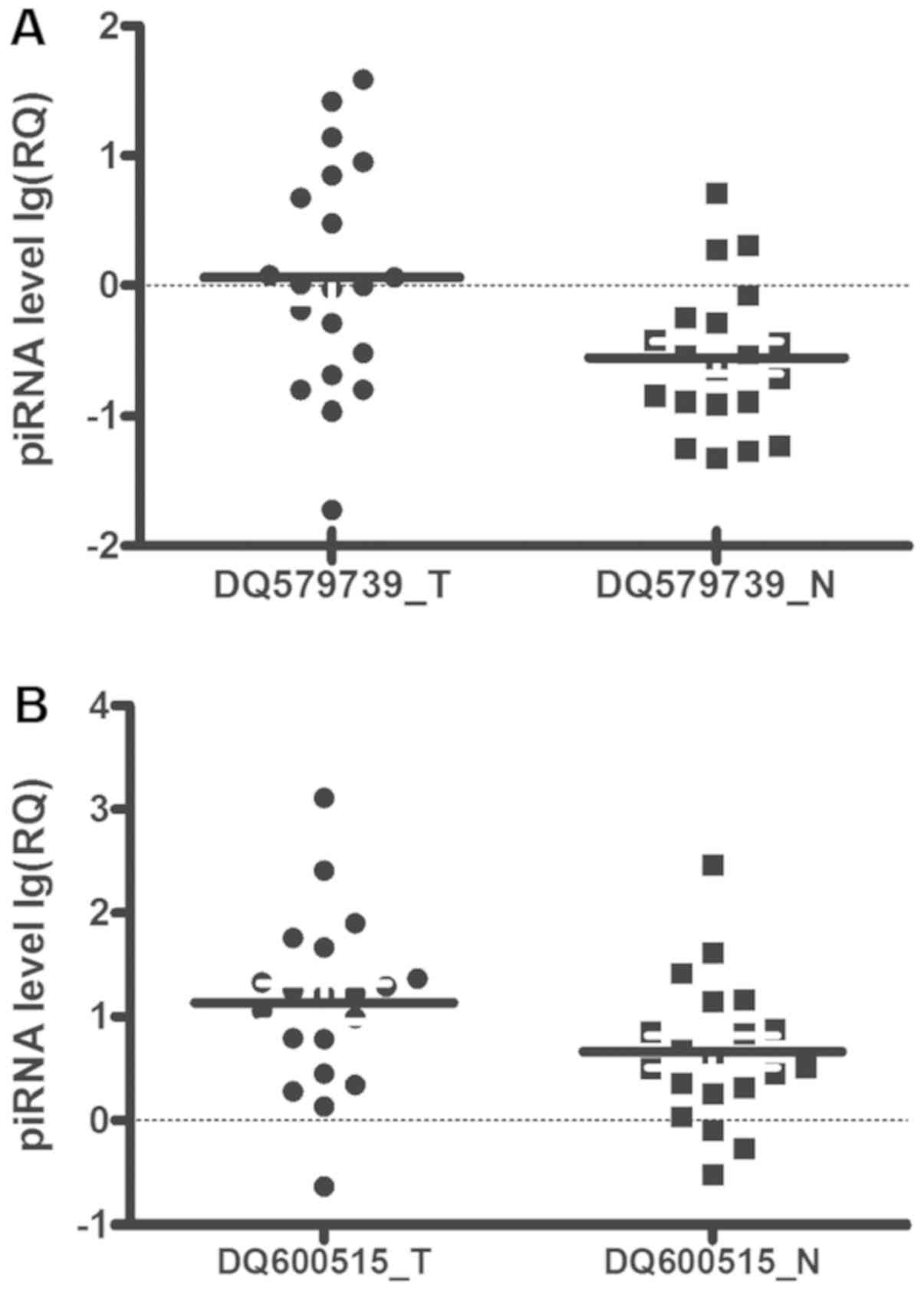
qPCR validation
In a second cohort of 20 patients with gastric
cancer, qPCR analysis on two piRNAs selected from the top list
verified the quality of microarray experiment and validated the
piRNA-GC associations in the primary cohort. DQ579739 and DQ600515
expression in the second cohort was concordant with the microarray
data in the primary cohort. Notably, DQ579739 (P=0.001) and
DQ600515 (P=0.005) expression in GC in the second cohort was
significantly higher than in adjacent tissues (Fig. 3), consistent with the differential
analysis on the array data in the primary cohort (Fig. 2) and in the same direction of
association (i.e., upregulated).
Gene expression in the stomach
Among the 50 GC-associated piRNAs, 43 were located
within or close to 34 protein-coding genes. Twenty-two (64.7%) of
the 34 genes within or close to which the 26 GC-associated piRNAs
(60.5%) were located were expressed in human stomach (Table SI). The top four risk genes most
abundant in stomach were CXCL17, EPCAM, RPL3 and RPS8
(Table SII).
piRNA expression is correlated with
SNPs
eQTL analysis showed that many GC-associated piRNAs
were nominally correlated with the genome-wide significant risk
SNPs (1.6×10−4≤P<0.05; Table III). After Bonferroni correction
(α=0.001), the correlations of rs4072037 at MUC1 with
DQ595533, DQ590386, DQ595534 and DQ571813 in the whole sample (N+T)
(P=3.6×10−4, 2.6×10−4, 1.6×10−4
and 2.6×10−4, respectively) and with DQ573237 in control
group (P=9.3×10−4) remained significant. The results are
also illustrated in Fig. S3.
| Table III.The risk piRNAs correlated with the
genome-wide significant replicated risk SNPs. |
Table III.
The risk piRNAs correlated with the
genome-wide significant replicated risk SNPs.
|
|
|
| Chr1: MUC1
rs4072037 | Chr5: PRKAA1
rs13361707 | Chr8: PSCA
rs2294008 |
|---|
|
|
|
|
|
|
|
|---|
| piRNA | Chr | Gene | N | T | N+T | N | T | N+T | N | N+T |
|---|
| DQ575884 | chr2 | to
EPCAM | 0.026 | 0.048 | 0.006 | 0.044 |
| 0.023 |
|
|
| DQ578739 | chr2 | to
LOC100507334 |
| 0.038 |
|
|
|
|
|
|
| DQ571556 | chr2 | to
SPATA31C1 | 0.027 |
|
|
|
|
|
|
|
| DQ595533 | chr3 | COLQ | 0.003 | 0.023 |
3.6×10−4 | 0.033 |
| 0.005 |
|
|
| DQ593293 | chr5 | to
C5orf52 | 0.018 |
|
|
|
|
|
|
|
| DQ573237 | chr6 | SAYSD1 |
9.3×10−4 | 0.039 | 0.005 | 0.016 | 0.035 | 0.003 |
|
|
| DQ580665 | chr7 | POLR2J | 0.044 |
| 0.039 | 0.025 |
|
|
|
|
| DQ583443 | chr9 | FAM225A | 0.042 |
|
|
|
|
|
|
|
| DQ571073 | chr12 | to
CLLU1 |
| 0.039 |
|
|
|
|
|
|
| DQ600254 | chr14 | PNMA1 | 0.008 | 0.045 | 0.002 | 0.032 |
| 0.010 |
|
|
| DQ572562 | chr15 | DNM1P46 |
|
| 0.016 |
|
|
|
|
|
| DQ590386 | chr16 | VAC14 | 0.003 | 0.022 |
2.6×10−4 | 0.032 |
| 0.004 |
|
|
| DQ595534 | chr16 | VAC14 | 0.002 | 0.015 |
1.6×10−4 | 0.026 |
| 0.003 |
|
|
| DQ571813 | chr16 | VAC14 | 0.002 | 0.025 |
2.6×10−4 | 0.024 |
| 0.004 |
|
|
| DQ577343 | chr19 | to
LOC400685 | 0.002 |
| 0.023 |
|
|
|
|
|
| DQ577344 | chr19 | to
LOC400685 | 0.003 |
| 0.015 |
|
|
|
|
|
| DQ577345 | chr19 | to
LOC400685 | 0.005 |
| 0.013 |
|
|
|
|
|
| DQ583043 | chr19 | to
LOC100652909 |
|
|
|
|
|
| 0.029 |
|
| DQ574146 | chr22 | IGLL1 |
|
|
|
|
|
|
| 0.045 |
| DQ574147 | chr22 | IGLL1 |
|
|
|
|
|
|
| 0.042 |
| DQ580529 | chr22 | to
WBP2NL | 0.004 |
|
|
|
|
|
|
|
Piwi proteins are expressed in the
stomach
Among the four piwi proteins, PIWIL4 was most
abundant in stomach in the American cohort (TPM=3.9; GTEx). It was
also expressed in our stomach samples of both 80 GC and 127
adjacent tissues (all expression scores ≥2+). There was
significant difference in the weighted expression scores between GC
and adjacent tissues (P=2.3×10−7; Table IV; Fig. S5).
| Table IV.Piwi protein expression in 181
gastric cancer and adjacent tissues. |
Table IV.
Piwi protein expression in 181
gastric cancer and adjacent tissues.
|
| Weighted expression
scores |
|
|---|
|
|
|
|
|---|
|
| − | 1+ | 2+ | 3+ | P-value |
|---|
| Cancer | 49 | 52 | 57 | 23 |
|
| Normal | 21 | 33 | 110 | 17 |
2.3×10−7 |
Discussion
In the present study, we found that piRNAs are
abundant in the human stomach, with 50 of them significantly
associated with GC. Many GC-associated piRNAs were correlated with
the genome-wide significant risk SNPs for GC and potentially
regulated the transcription of cancer-related genes in the stomach
via TE-suppression or TE-independent mechanisms. These findings
support the proposition that piRNAs might play a role in
determining GC risk.
These 50 piRNAs might impact GC risk via targeting
the TEs and then regulating the expression of the proximate
protein-coding genes, by sequence complementarity. Of them, 17
piRNAs, were upregulated in GC. This upregulation might suppress
the activity of TEs, which maintains the self-renewal and
proliferation capacity of cancer cells. The other 33 piRNAs were
significantly downregulated in GC. This downregulation would
increase the repeats of retrotransposon and cause DNA damage. We
illustrate these potential mechanisms underlying the pathogenicity
of piRNAs in Fig. S3.
In the present study, 8,759 piRNAs were detected in
the human stomach. Across the transcriptome, an average of 5% of
the piRNAs mapped within the protein-coding genes (61). However, in the present study, of the
50 differentially expressed piRNAs, 30 (60%) mapped within
protein-coding genes, suggesting a strong relationship between
these piRNAs and GC. Among the 34 protein-coding genes where the
GC-associated piRNAs are located, 50% were expressed in stomach and
associated with cancers, again in support of a strong relationship
between the piRNAs and GC. Thus, the GC-associated piRNAs
contribute to the pathogenesis of GC possibly via regulating their
nearest genes based on sequence complementarity (31).
Two piRNAs, i.e., DQ579739 and DQ588779, are
particularly worthy of attention (1). In our study, DQ579739 expression in GC
was higher than in adjacent tissues in both microarray and qPCR
validation data across two independent cohorts. The sequence of
DQ579739 is complementary to AluSc8, a SINE-type TE, within the
CXCL17 gene. CXCL17 mRNA was the most abundant in
human stomach among all protein-coding genes examined in our study.
As a novel acid CXC chemokine, CXCL17 is significantly upregulated
in colon cancer (45),
hepatocellular carcinoma (57),
pancreatic carcinogenesis (56) and
lung macrophages (62), and is
associated with carcinogenesis, tumor proliferation, tumor
progression (63), and angiogenesis
(64,65). CXCL17 might act as a chemokine that
accelerates tumor progression. The changes in individual piRNA
level could influence both auto-regulatory gene expression and the
expression of the gene to which the piRNA is complementary
(66). Our finding suggests that
this piRNA may be involved in GC development by regulating its host
gene and its interacting proteins via AluSc8 (2). Among all piRNAs examined, the most
abundant one in human stomach is DQ588779 at SKI (SKI
proto-oncogene) (on chr1). This SKI proto-oncogene encodes the
nuclear proto-oncogene protein homolog of avian sarcoma viral
(v-ski) oncogene. It is expressed abundantly in the stomach, and
has been associated with many types of cancers including GC
(6,7). DQ588779 might play important roles in
the development of GC by targeting AluSz6, a SINE-type TE, which
may regulate the expression of stomach SKI gene.
Many GC-associated piRNAs were nominally correlated
with the genome-wide significant risk SNPs, suggesting that these
GC-associated piRNAs may be biologically functional. Several piRNAs
complementary to COLQ, SAYSD1 and VAC14 were
significantly correlated with rs4072037 on MUC1, suggesting
a potential trans-acting regulation that may underlie SNP-GC
associations.
To conclude, piRNAs are abundant in the human
stomach and may play important roles in the pathogenesis of gastric
cancer, and underlie the SNP-cancer associations as have been
identified by GWASs. Overall, the current findings support piRNAs
as potential biomarkers for GC diagnosis and prognosis. It is
warranted to replicate these findings in larger sample in future
work.
Supplementary Material
Supporting Data
Acknowledgements
We thank Dr Chiang-Shan Li of Yale University for
thorough editing of the manuscript.
Funding
The present study was supported by the National
Clinical Key Specialty Construction Program of China, research
grants from the Ministry of Health, China (WKJ2016-2-05), the
Fujian Provincial Health Technology Project (2015-CX-7,
2013-ZQN-JC-8, 2015-ZQN-JC-7), the Natural Science Foundation of
Fujian Province (nos. 2016J01513, 2016J01508 and 2019J01196), the
Joint Funds for the Innovation of Science and Technology, Fujian
province (no. 2017Y9082), the Science and Technology Program of
Fujian Province (no. 2018Y2003) and the National Institute on
Alcohol Abuse and Alcoholism (NIAAA) grants R21 AA021380, R21
AA020319 and R21 AA023237. To note, the study sponsors only
provided the funds, but did not participated in the study.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
XLi, XZ, JX, QM, HHC and XLu contributed to the
formulation of the research goals and aims and the writing,
reviewing, and editing of the article. XLi, YX, DH, HeZ, ZY, CL,
GC, FL, WZ, YS, HuZ, JZ, TS, JX, XZ and XLu provided the patient
resources and searched and collected the data. XLi, YX, DH, QM,
HeZ, ZY, CL, GC, FL WZ, YS, HuZ, JZ, TS, HHC and XLu conducted the
study and performed the analysis. XLi, XZ and XLu secured funding
for the present study. XLi and XLu are responsible for the overall
content as guarantors. All authors read and approved the manuscript
and agree to be accountable for all aspects of the research in
ensuring that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
The study was approved by the Research Ethics
Committee of the Fujian Provincial Cancer Hospital, China. Informed
consents from all patients were obtained prior to the study. All
experiments were performed in accordance with relevant guidelines
and regulations.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lin X, Hu D, Chen G, Shi Y, Zhang H, Wang
X, Guo X, Lu L, Black D, Zheng XW and Luo X: Associations of THBS2
and THBS4 polymorphisms to gastric cancer in a Southeast Chinese
population. Cancer Genet. 209:215–222. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Berger H, Marques MS, Zietlow R, Meyer TF,
Machado JC and Figueiredo C: Gastric cancer pathogenesis.
Helicobacter. 21 (Suppl 1):S34–S38. 2016. View Article : Google Scholar
|
|
4
|
Kim J, Yum S, Kang C and Kang SJ:
Gene-gene interactions in gastrointestinal cancer susceptibility.
Oncotarget. 7:67612–67625. 2016.PubMed/NCBI
|
|
5
|
Lin XD, Chen SQ, Qi YL, Zhu JW, Tang Y and
Lin JY: Polymorphism of THBS1 rs1478604 A>G in 5-untranslated
region is associated with lymph node metastasis of gastric cancer
in a Southeast Chinese population. DNA Cell Biol. 31:511–519. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Nakao T, Kurita N, Komatsu M, Yoshikawa K,
Iwata T, Utsunomiya T and Shimada M: Expression of thrombospondin-1
and Ski are prognostic factors in advanced gastric cancer. Int J
Clin Oncol. 16:145–152. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takahata M, Inoue Y, Tsuda H, Imoto I,
Koinuma D, Hayashi M, Ichikura T, Yamori T, Nagasaki K, Yoshida M,
et al: SKI and MEL1 cooperate to inhibit transforming growth
factor-beta signal in gastric cancer cells. J Biol Chem.
284:3334–3344. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hong CS, Jeong O, Piao Z, Guo C, Jung MR,
Choi C and Park YK: HOXB5 induces invasion and migration through
direct transcriptional up-regulation of β-catenin in human gastric
carcinoma. Biochem J. 472:393–403. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xie SS, Jin J, Xu X, Zhuo W and Zhou TH:
Emerging roles of non-coding RNAs in gastric cancer: Pathogenesis
and clinical implications. World J Gastroenterol. 22:1213–1223.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang T, Ji Y, Hu D, Chen B, Zhang H, Li
C, Chen G, Luo X, Zheng XW and Lin X: SNHG8 is identified as a key
regulator of epstein-barr virus(EBV)-associated gastric cancer by
an integrative analysis of lncRNA and mRNA expression. Oncotarget.
7:80990–81002. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fu A, Jacobs DI, Hoffman AE, Zheng T and
Zhu Y: PIWI-interacting RNA 021285 is involved in breast
tumorigenesis possibly by remodeling the cancer epigenome.
Carcinogenesis. 36:1094–1102. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cheng J, Guo JM, Xiao BX, Miao Y, Jiang Z,
Zhou H and Li QN: piRNA, the new non-coding RNA, is aberrantly
expressed in human cancer cells. Clin Chim Acta. 412:1621–1625.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zuo L, Wang Z, Tan Y, Chen X and Luo X:
piRNAs and their functions in the brain. Int J Hum Genet. 16:53–60.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ross RJ, Weiner MM and Lin H: PIWI
proteins and PIWI-interacting RNAs in the soma. Nature.
505:353–359. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aravin A, Gaidatzis D, Pfeffer S,
Lagos-Quintana M, Landgraf P, Iovino N, Morris P, Brownstein MJ,
Kuramochi-Miyagawa S, Nakano T, et al: A novel class of small RNAs
bind to MILI protein in mouse testes. Nature. 442:203–207. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sturm Á, Perczel A, Ivics Z and Vellai T:
The Piwi-piRNA pathway: Road to immortality. Aging Cell.
16:906–911. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mei Y, Clark D and Mao L: Novel dimensions
of piRNAs in cancer. Cancer Lett. 336:46–52. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ng KW, Anderson C, Marshall EA, Minatel
BC, Enfield KS, Saprunoff HL, Lam WL and Martinez VD:
Piwi-interacting RNAs in cancer: Emerging functions and clinical
utility. Mol Cancer. 15:52016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Busch J, Ralla B, Jung M, Wotschofsky Z,
Trujillo-Arribas E, Schwabe P, Kilic E, Fendler A and Jung K:
Piwi-interacting RNAs as novel prognostic markers in clear cell
renal cell carcinomas. J Exp Clin Cancer Res. 34:612015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hashim A, Rizzo F, Marchese G, Ravo M,
Tarallo R, Nassa G, Giurato G, Santamaria G, Cordella A, Cantarella
C and Weisz A: RNA sequencing identifies specific PIWI-interacting
small non-coding RNA expression patterns in breast cancer.
Oncotarget. 5:9901–9910. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yan H, Wu QL, Sun CY, Ai LS, Deng J, Zhang
L, Chen L, Chu ZB, Tang B, Wang K, et al: piRNA-823 contributes to
tumorigenesis by regulating de novo DNA methylation and
angiogenesis in multiple myeloma. Leukemia. 29:196–206. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Law PT, Qin H, Ching AK, Lai KP, Co NN, He
M, Lung RW, Chan AW, Chan TF and Wong N: Deep sequencing of small
RNA transcriptome reveals novel non-coding RNAs in hepatocellular
carcinoma. J Hepatol. 58:1165–1173. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang G, Hu H, Xue X, Shen S, Gao E, Guo
G, Shen X and Zhang X: Altered expression of piRNAs and their
relation with clinicopathologic features of breast cancer. Clin
Transl Oncol. 15:563–568. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang H, Ren Y, Xu H, Pang D, Duan C and
Liu C: The expression of stem cell protein Piwil2 and piR-932 in
breast cancer. Surg Oncol. 22:217–223. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chu H, Hui G, Yuan L, Shi D, Wang Y, Du M,
Zhong D, Ma L, Tong N, Qin C, et al: Identification of novel piRNAs
in bladder cancer. Cancer Lett. 356:561–567. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Qiu W, Guo X, Lin X, Yang Q, Zhang W,
Zhang Y, Zuo L, Zhu Y, Li CR, Ma C and Luo X: Transcriptome-wide
piRNA profiling in human brains of Alzheimer's disease. Neurobiol
Aging. 57:170–177. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Martinez VD, Enfield KSS, Rowbotham DA and
Lam WL: An atlas of gastric PIWI-interacting RNA transcriptomes and
their utility for identifying signatures of gastric cancer
recurrence. Gastric Cancer. 19:660–665. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Levin HL and Moran JV: Dynamic
interactions between transposable elements and their hosts. Nat Rev
Genet. 12:615–627. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Malone CD and Hannon GJ: Small RNAs as
guardians of the genome. Cell. 136:656–668. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Reilly MT, Faulkner GJ, Dubnau J,
Ponomarev I and Gage FH: The role of transposable elements in
health and diseases of the central nervous system. J Neurosci.
33:17577–17586. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Roy J, Sarkar A, Parida S, Ghosh Z and
Mallick B: Small RNA sequencing revealed dysregulated piRNAs in
Alzheimer's disease and their probable role in pathogenesis. Mol
Biosyst. 13:565–576. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hu N, Wang Z, Song X, Wei L, Kim BS,
Freedman ND, Baek J, Burdette L, Chang J, Chung C, et al:
Genome-wide association study of gastric adenocarcinoma in Asia: A
comparison of associations between cardia and non-cardia tumours.
Gut. 65:1611–1618. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Abnet CC, Freedman ND, Hu N, Wang Z, Yu K,
Shu XO, Yuan JM, Zheng W, Dawsey SM, Dong LM, et al: A shared
susceptibility locus in PLCE1 at 10q23 for gastric adenocarcinoma
and esophageal squamous cell carcinoma. Nat Genet. 42:764–767.
2010. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Z, Dai J, Hu N, Miao X, Abnet CC,
Yang M, Freedman ND, Chen J, Burdette L, Zhu X, et al:
Identification of new susceptibility loci for gastric non-cardia
adenocarcinoma: Pooled results from two Chinese genome-wide
association studies. Gut. 66:581–587. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Helgason H, Rafnar T, Olafsdottir HS,
Jonasson JG, Sigurdsson A, Stacey SN, Jonasdottir A, Tryggvadottir
L, Alexiusdottir K, Haraldsson A, et al: Loss-of-function variants
in ATM confer risk of gastric cancer. Nat Genet. 47:906–910. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shi Y, Hu Z, Wu C, Dai J, Li H, Dong J,
Wang M, Miao X, Zhou Y, Lu F, et al: A genome-wide association
study identifies new susceptibility loci for non-cardia gastric
cancer at 3q13.31 and 5p13.1. Nat Genet. 43:1215–1218. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chen YC, Fang WL, Wang RF, Liu CA, Yang
MH, Lo SS, Wu CW, Li AF, Shyr YM and Huang KH: Clinicopathological
variation of lauren classification in gastric cancer. Pathol Oncol
Res. 22:197–202. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Edge SB and Compton CC: The American joint
committee on cancer: The 7th edition of the AJCC cancer staging
manual and the future of TNM. Ann Surg Oncol. 17:1471–1474. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lau NC, Seto AG, Kim J, Kuramochi-Miyagawa
S, Nakano T, Bartel DP and Kingston RE: Characterization of the
piRNA complex from rat testes. Science. 313:363–367. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
He QQ, Xiong LL, Liu F, He X, Feng GY,
Shang FF, Xia QJ, Wang YC, Qiu DL, Luo CZ, et al: MicroRNA-127
targeting of mitoNEET inhibits neurite outgrowth, induces cell
apoptosis and contributes to physiological dysfunction after spinal
cord transection. Sci Rep. 6:352052016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Yuan JS, Reed A, Chen F and Stewart CN Jr:
Statistical analysis of real-time PCR data. BMC Bioinformatics.
7:852006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
GTEx Consortium: The genotype-tissue
expression (GTEx) project. Nat Genet. 45:580–585. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wu C, Orozco C, Boyer J, Leglise M,
Goodale J, Batalov S, Hodge CL, Haase J, Janes J, Huss JW III and
Su AI: BioGPS: An extensible and customizable portal for querying
and organizing gene annotation resources. Genome Biol. 10:R1302009.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wilhelm M, Schlegl J, Hahne H, Gholami AM,
Lieberenz M, Savitski MM, Ziegler E, Butzmann L, Gessulat S, Marx
H, et al: Mass-spectrometry-based draft of the human proteome.
Nature. 509:582–587. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ohlsson L, Hammarström ML, Lindmark G,
Hammarström S and Sitohy B: Ectopic expression of the chemokine
CXCL17 in colon cancer cells. Br J Cancer. 114:697–703. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Oosterhoff D, Overmeer RM, de Graaf M, van
der Meulen IH, Giaccone G, van Beusechem VW, Haisma HJ, Pinedo HM
and Gerritsen WR: Adenoviral vector-mediated expression of a gene
encoding secreted, EpCAM-targeted carboxylesterase-2 sensitises
colon cancer spheroids to CPT-11. Br J Cancer. 92:882–887. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Xie X, Li F, Zhang H, Lu Y, Lian S, Lin H,
Gao Y and Jia L: EpCAM aptamer-functionalized mesoporous silica
nanoparticles for efficient colon cancer cell-targeted drug
delivery. Eur J Pharm Sci. 83:28–35. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Russo A, Maiolino S, Pagliara V, Ungaro F,
Tatangelo F, Leone A, Scalia G, Budillon A, Quaglia F and Russo G:
Enhancement of 5-FU sensitivity by the proapoptotic rpL3 gene in
p53 null colon cancer cells through combined polymer nanoparticles.
Oncotarget. 7:79670–79687. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Pagliara V, Saide A, Mitidieri E,
d'Emmanuele di Villa Bianca R, Sorrentino R, Russo G and Russo A:
5-FU targets rpL3 to induce mitochondrial apoptosis via
cystathionine-β-synthase in colon cancer cells lacking p53.
Oncotarget. 7:50333–50348. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Russo A, Pagliara V, Albano F, Esposito D,
Sagar V, Loreni F, Irace C, Santamaria R and Russo G: Regulatory
role of rpL3 in cell response to nucleolar stress induced by Act D
in tumor cells lacking functional p53. Cell Cycle. 15:41–51. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wang SY, Gao K, Deng DL, Cai JJ, Xiao ZY,
He LQ, Jiao HL, Ye YP, Yang RW, Li TT, et al: TLE4 promotes
colorectal cancer progression through activation of JNK/c-Jun
signaling pathway. Oncotarget. 7:2878–2888. 2016.PubMed/NCBI
|
|
52
|
Liu YL, Gao X, Jiang Y, Zhang G, Sun ZC,
Cui BB and Yang YM: Expression and clinicopathological significance
of EED, SUZ12 and EZH2 mRNA in colorectal cancer. J Cancer Res Clin
Oncol. 141:661–669. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Seo GS, Yu JI, Chae SC, Park WC, Shin SR,
Yoo ST, Choi SC and Lee SH: EED gene polymorphism in patients with
colorectal cancer. Int J Biol Markers. 28:274–279. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Song L, Chen X, Gao S, Zhang C, Qu C, Wang
P and Liu L: Ski modulate the characteristics of pancreatic cancer
stem cells via regulating sonic hedgehog signaling pathway. Tumour
Biol. Oct 12–2016.(Epub ahead of print). View Article : Google Scholar :
|
|
55
|
Jiang SH, He P, Ma MZ, Wang Y, Li RK, Fang
F, Fu Y, Tian GA, Qin WX and Zhang ZG: PNMA1 promotes cell growth
in human pancreatic ductal adenocarcinoma. Int J Clin Exp Pathol.
7:3827–3835. 2014.PubMed/NCBI
|
|
56
|
Hiraoka N, Yamazaki-Itoh R, Ino Y,
Mizuguchi Y, Yamada T, Hirohashi S and Kanai Y: CXCL17 and ICAM2
are associated with a potential anti-tumor immune response in early
intraepithelial stages of human pancreatic carcinogenesis.
Gastroenterology. 140:310–321. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Li L, Yan J, Xu J, Liu CQ, Zhen ZJ, Chen
HW, Ji Y, Wu ZP, Hu JY, Zheng L and Lau WY: CXCL17 expression
predicts poor prognosis and correlates with adverse immune
infiltration in hepatocellular carcinoma. PLoS One. 9:e1100642014.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Chan AW, Tong JH, Chan SL, Lai PB and To
KF: Expression of stemness markers (CD133 and EpCAM) in
prognostication of hepatocellular carcinoma. Histopathology.
64:935–950. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Kang JS: Increased expression of
epithelial cell adhesion molecule (EpCAM) in rat hepatic tumors
induced by diethylnitrosamine. Asian Pac J Cancer Prev.
13:3627–3630. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Luo J, Cai Q, Wang W, Huang H, Zeng H, He
W, Deng W, Yu H, Chan E, Ng CF, et al: A microRNA-7 binding site
polymorphism in HOXB5 leads to differential gene expression in
bladder cancer. PLoS One. 7:e401272012. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Brennecke J, Aravin AA, Stark A, Dus M,
Kellis M, Sachidanandam R and Hannon GJ: Discrete small
RNA-generating loci as master regulators of transposon activity in
Drosophila. Cell. 128:1089–1103. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Burkhardt AM, Maravillas-Montero JL,
Carnevale CD, Vilches-Cisneros N, Flores JP, Hevezi PA and Zlotnik
A: CXCL17 is a major chemotactic factor for lung macrophages. J
Immunol. 193:1468–1474. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Matsui A, Yokoo H, Negishi Y,
Endo-Takahashi Y, Chun NA, Kadouchi I, Suzuki R, Maruyama K,
Aramaki Y, Semba K, et al: CXCL17 expression by tumor cells
recruits CD11b+Gr1 high F4/80- cells and promotes tumor
progression. PLoS One. 7:e440802012. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Weinstein EJ, Head R, Griggs DW, Sun D,
Evans RJ, Swearingen ML, Westlin MM and Mazzarella R: VCC-1, a
novel chemokine, promotes tumor growth. Biochem Biophys Res Commun.
350:74–81. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Mu X, Chen Y, Wang S, Huang X, Pan H and
Li M: Overexpression of VCC-1 gene in human hepatocellular
carcinoma cells promotes cell proliferation and invasion. Acta
Biochim Biophys Sin (Shanghai). 41:631–637. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Esposito T, Magliocca S, Formicola D and
Gianfrancesco F: piR_015520 belongs to Piwi-associated RNAs
regulates expression of the human melatonin receptor 1A gene. PLoS
One. 6:e227272011. View Article : Google Scholar : PubMed/NCBI
|