Introduction
Pancreatic cancer (PaCa) is one of the most highly
malignant carcinomas. PaCa was the fourth leading cause of
cancer-related death in Japan in 2017 (National Cancer Center
Japan. Center for Cancer Control and Information Services, 2018.
Cancer Statistics in Japan. https://ganjoho.jp/en/professional/statistics/table_download.html.)
and the third leading cause of cancer-related death in the US. In
2018, there were an estimated 55,440 newly diagnosed patients with
PaCa and 44,330 PaCa-related deaths in the US (1). The overall 5-year survival rate is ~6%
(range, 2–9%) worldwide (2). The
poor prognosis of PaCa is attributed to its typical characteristics
of late presentation, aggressive local invasion, early metastasis,
and poor response to chemotherapy (3). Most PaCa patients are already at an
advanced stage at the time of diagnosis. Approximately 10–20% of
PaCa patients are eligible for tumor resection, whereas the
remaining patients are eligible only for adjuvant therapies
(4–6). Therefore, the development of novel
target therapies is expected to improve the outcome of PaCa
treatment.
In 1997, a phase III trial comparing fluorouracil
(5-FU) monotherapy, which was the standard treatment at that time,
and gemcitabine (Gem) monotherapy for patients with PaCa was
conducted. That trial demonstrated the advantages of Gem over 5-FU
not only in terms of a clinical benefit (e.g., pain relief) but
also overall survival (5.7 vs. 4.4 months) (7). Consequently, Gem was approved in 1997
as a first-line chemotherapeutic drug for patients with locally
advanced or metastatic PaCa, and Gem remains the standard treatment
for PaCa patients (4). However, the
clinical efficacy of Gem therapy is poor, and there is little
improvement in the survival of PaCa patients who receive this
therapy (6). Clinical experience
has shown that Gem has transient effects on PaCa after beginning
chemotherapy, with the effects readily decreasing thereafter. One
of the reasons is acquisition of Gem resistance (Gem-R), the
mechanism of which is still unclear. Thus, it is reasonable to
expect that elucidation of Gem-R mechanisms will improve PaCa
treatment.
Recently, there has been increasing evidence that
chemokines play a role in tumor biology (8,9). These
studies have demonstrated that chemokines may influence tumor
growth, invasion and metastasis. Interleukin-8 (IL-8/CXCL8) is a
CXC chemokine containing a Glu-Leu-Arg motif and is secreted by
leukocytes and tumor cells. It was initially named
neutrophil-activating peptide-1 for its potent chemotactic activity
on granulocytes in inflammatory and immune diseases (10). IL-8 has been shown to play important
roles in cancer invasion (11),
angiogenesis (12) and metastasis
(13). Based on these results,
previous reports indicate that IL-8 is a potential therapeutic
target in breast (14,15), gastric (13,16),
colon (17), cervical cancer
(18), and melanoma (19). Furthermore, we previously
demonstrated a significant role of PaCa-induced IL-8 in tumor
angiogenesis (20,21). Moreover, blocking CXCR2, the main
receptor of IL-8, significantly suppressed the increased PaCa
angiogenesis and tumorigenesis (20).
The aim of this study was to determine the role of
IL-8 in the acquisition of Gem-R in PaCa. Initially, we established
Gem-R PaCa cell lines. We confirmed that IL-8 expression was
increased in accordance with the acquisition of Gem-R. Furthermore,
we found that an anti-CXCR2 Ab inhibited the angiogenic activity
induced by Gem-R in PaCa cell lines. To the best of our knowledge,
this is the first report to demonstrate the mechanisms of
Gem-R-induced tumor angiogenesis in relationship to the chemokine
network. Since regulation of the IL-8/CXCR2 axis reduced
Gem-R-induced angiogenesis, this axis may be a new therapeutic
target for Gem-R PaCa.
Materials and methods
Cell culture and treatments
The PaCa cell lines BxPC-3, AsPC-1, MIA PaCa-2,
Panc-1, and SW 1990 were obtained from the American Type Culture
Collection (ATCC; Rockville, MD, USA). BxPC-3 and AsPC-1 cells were
maintained in RPMI-1640 medium (Sigma Chemical Co.; Merck KGaA,
Darmstadt, Germany) supplemented with 10% fetal bovine serum (FBS).
MIA PaCa-2, Panc-1 and SW 1990 cells were maintained in Dulbecco's
modified Eagle's medium (DMEM) (Sigma Chemical Co.; Merck KGaA)
containing high glucose and 10% FBS. All cells were incubated at
37°C in a humidified atmosphere of 5% CO2 in air.
Establishment of Gem-R PaCa cell
lines
First, we determined the half maximal inhibitory
concentration (IC50) of Gem (Toronto Research Chemicals,
North York, Ontario, Canada) for PaCa cells using the Premix WST-1
Cell Proliferation Assay System (Takara Bio, Inc., Shiga, Japan)
according to the manufacturer's instructions. Briefly, PaCa cells
(AsPC-1, MIA PaCa-2, BxPC-3, Panc-1 and SW 1990) were seeded at
2×103/100 µl in 96-well plates and allowed to adhere
overnight. Then, the cultures were provided with fresh medium
containing various concentrations of Gem. After 72 h of incubation,
the absorbance was measured at 450 nm in each well using the
SpectraMax 340 spectrophotometer (Molecular Devices, LLC,
Sunnyvale, CA, USA). The IC50 of Gem for each PaCa cell
line was determined by constructing dose-response curves. Each PaCa
cell line was passaged at the respective Gem IC50
concentration for 2–3 weeks. After passage, the IC50
value of Gem was again determined for each cell line and each cell
line was passaged with the re-determined IC50
concentration for 2–3 weeks. The process was repeated at increasing
doses of Gem until the cell lines demonstrated at least a 50-fold
greater Gem IC50 value than that of the respective
parental cell line. The resulting cell lines were resistant to 20
µM Gem.
Cell proliferation assay
Proliferation assays were conducted using the Premix
WST-1 Cell Proliferation Assay System (Takara Bio, Inc.) according
to the manufacturer's instructions. Briefly, Gem-R and
Gem-sensitive (Gem-S) AsPC-1 or MIA PaCa-2 cells were seeded at
2×103/100 µl in 96-well plates and allowed to adhere
overnight. Then, cultures were provided with fresh medium
containing various concentrations (0–1,000 µM) of Gem. After 72 h
of incubation, the absorbance was measured at 450 nm in each well
using the SpectraMax 340 spectrophotometer (Molecular Devices,
LLC). To compare the proliferation of Gem-R and Gem-S PaCa cells in
the absence of Gem treatment, 2×104 cells were seeded in
6-well plates and allowed to adhere overnight. After 24, 48, 72 and
96 h of incubation, the cells were isolated after treatment with
0.25% Trypsin-EDTA (Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) and counted in four microscopic fields (×100) using a
compound light microscope. This assay was also performed to
investigate the effect of IL-8 on the proliferation of Gem-R and
Gem-S PaCa cells. First, Gem-S MIA PaCa-2 cells were incubated with
various concentrations (0–100 µM) of Gem and with or without 100
ng/ml recombinant human CXCL8/IL-8 (R&D Systems, Minneapolis,
MN, USA) for 72 h. Second, Gem-R and Gem-S PaCa cells were treated
with IL-8 small-interfering RNA (siRNA) or negative control siRNA
(as described below) to assess the effect of siRNA-mediated
knockdown of IL-8. In both cases, the cells were subjected to WST-1
proliferation assays as described above.
siRNA-mediated IL-8 knockdown
We performed siRNA-mediated knockdown of IL-8 in
Gem-R PaCa cells using IL-8 siRNA(h) (cat. no. sc-39631; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA), Silencer Select Negative
Control No. 1 siRNA (4390843) (Invitrogen; Thermo Fisher
Scientific, Inc.), Opti-MEM I reduced serum medium (Invitrogen;
Thermo Fisher Scientific, Inc.), and Lipofectamine®
RNAiMAX transfection reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturers' instructions. Briefly, Gem-R
MIA PaCa-2 cells were seeded in 6-well plates and allowed to adhere
overnight. Then, the cells were incubated with transfection
mixtures containing 10 nM IL-8 siRNA or negative control siRNA for
72 h. IL-8 expression was confirmed in PaCa cells treated with IL-8
siRNA or negative control siRNA by reverse-transcription
quantitative polymerase chain reaction (RT-qPCR) as described
below. The IL-8 siRNA (cat. no. sc-39631) oligonucleotides were
5′-GGGUGCAGAGGGUUGUGGAGAtt-3′ (sense), and
5′-UCUCCACAACCCUCUGCACCCtt-3′ (antisense).
Total mRNA microarray analysis
Total mRNA from Gem-R and Gem-S MIA PaCa-2 cells
were isolated using the RNeasy Plus Mini Kit (Qiagen, Inc.,
Valencia, CA, USA) according to the manufacturer's instructions.
The mRNA microarray experiments were performed at the Takara Bio
Dragon Genomics Center (Yokkaichi, Mie, Japan). Transcripts
amplified from the total mRNA were hybridized to the Affymetrix
Human Genome U133 Plus 2.0 Array (Affymetrix/Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. The
results were analyzed using the Affymetrix GeneChip™ Command
Console Software and Affymetrix Expression Console Software
(Affymetrix; Thermo Fisher Scientific, Inc.).
RT-qPCR
Total RNA was isolated from PaCa cells using the
RNeasy Plus Mini kit (Qiagen, Inc.), according to the
manufacturer's instructions, and quantitated using the NanoDrop
1000 (Thermo Fisher Scientific, Inc.). Total RNA (1 µg) was
reverse-transcribed using the SuperScript III Platinum Two-Step
qRT-PCR Kit (Invitrogen, Thermo Fisher Scientific, Inc.) according
to the manufacturer's instructions, and 1 µl of the product was
used as a template for PCR. RT-qPCR was carried out using TaqMan
Universal Master Mix and TaqMan Gene Expression Assays for IL-8
(Hs01553824_g1) and GAPDH (Hs99999905_m1) (Applied Biosystems;
Thermo Fisher Scientific, Inc.) with Chrome4 (Bio-Rad, Hercules,
CA, USA). The RT-qPCR conditions were as follows: an initial
incubation at 50°C for 2 min, followed by denaturation at 95°C for
10 min and 50 cycles of 95°C for 15 sec and 60°C for 1 min. The
relative expression levels of IL-8 were normalized to the
expression of GAPDH in each sample using standard curve method.
Cytokine array
The Human Cytokine Array Panel Array kit (R&D
Systems) was utilized according to the manufacturer's instructions.
Gem-R and Gem-S AsPC-1 cells were seeded at 1×106 in
60-mm dishes and allowed to adhere overnight. Then, the culture
medium was replaced with 1 ml RPMI-1640 containing 2% FBS. After 24
h, the cell culture supernatant was centrifuged to remove
particulates, and 500 µl was used for each array. Images were
captured using the LAS-3000 imaging system (Fujifilm, Shizuoka,
Japan), with an exposure time of 10 min. Signal analysis was
performed using Multi Gauge software (ver3.0; Fujifilm, Tokyo,
Japan), with each signal normalized to the positive controls.
Enzyme-linked immunosorbent assay
(ELISA)
All cell lines were seeded at 2×105/ml
into a 24-well plate containing medium supplemented with 10% FBS
and cultured overnight. Medium was exchanged the next day, and
cells were cultured for 48 h. The culture media were then collected
and centrifuged at 400 × g for 5 min to remove particulates and
frozen at −80°C until used for ELISA. The concentration of IL-8 was
measured using the Human CXCL8/IL-8 Quantikine ELISA kit (R&D
Systems) according to the manufacturer's instructions. The minimum
detectable dose of this kit ranges from 1.5 to 7.5 pg/ml.
Angiogenesis assay
To investigate the influence of Gem-R PaCa cells on
tube formation by human umbilical vein endothelial cells (HUVECs),
PaCa cell lines (Gem-R AsPC-1, Gem-S AsPC-1, Gem-R MIA PaCa-2, or
Gem-S MIA PaCa-2), HUVECs and fibroblasts were co-cultured using a
double chamber method in 24-well plates, and angiogenic activity
was measured using an angiogenesis kit (cat. no. KZ-1000) (Kurabo
Co., Osaka, Japan) according to the manufacturer's protocols
(21–23). Gem-R or Gem-S PaCa cells
(1×104 cells) were seeded into Transwell chambers,
consisting of polycarbonate membranes with 0.45-µm pores, and
allowed to adhere overnight. Transwell chambers were then placed
into the HUVEC/fibroblast co-culture system and exchanged on days
4, 7 and 10. Cells were cultured for 11 days, after which the
HUVECs were stained with an anti-CD31 antibody (Ab) using a tubule
staining kit (KZ-1225) (Kurabo Co.) according to the manufacturer's
protocol. Briefly, we fixed cells by iced 70% ethanol for 30 min at
room temperature. We used phosphate-buffered salts (−) with 1%
bovine serum albumin as blocking reagent. We added mouse anti-human
CD31 antibody in the kit diluted 4,000 times by blocking reagent as
the primary Ab and incubated for 60 min at 37°C. After incubation,
we washed plates by blocking reagent and added goat anti-mouse IgG
AlkP conjugate included in the kit diluted 500 times by blocking
reagent as the secondary Ab and incubated for 60 min at 37°C. The
area of tube formation was measured quantitatively in 15 different
fields for each condition using an image analyzer (Kurabo Co.). The
assay allowed quantitative evaluation of angiogenesis and
examination of tumor-stromal interactions. Using this same method,
the effects of an anti-human neutralizing CXCR2 Ab (10 µg/ml;
R&D Systems) on HUVEC tube formation in the presence of PaCa
cells were also assessed.
Statistical analysis
Differences between the means of two samples were
analyzed by unpaired t-tests. Multiple group comparisons were
performed by one-way analysis of variance (ANOVA) with a post hoc
Bonferroni test for subsequent comparisons of individual groups. A
P-value <0.05 was considered statistically significant. Mean
values and standard deviations (SDs) were calculated for
experiments performed in at least triplicate.
Results
IC50 values of Gem for PaCa
cell lines
To determine the concentrations of Gem needed to
establish Gem-R PaCa cell lines, we performed cell proliferation
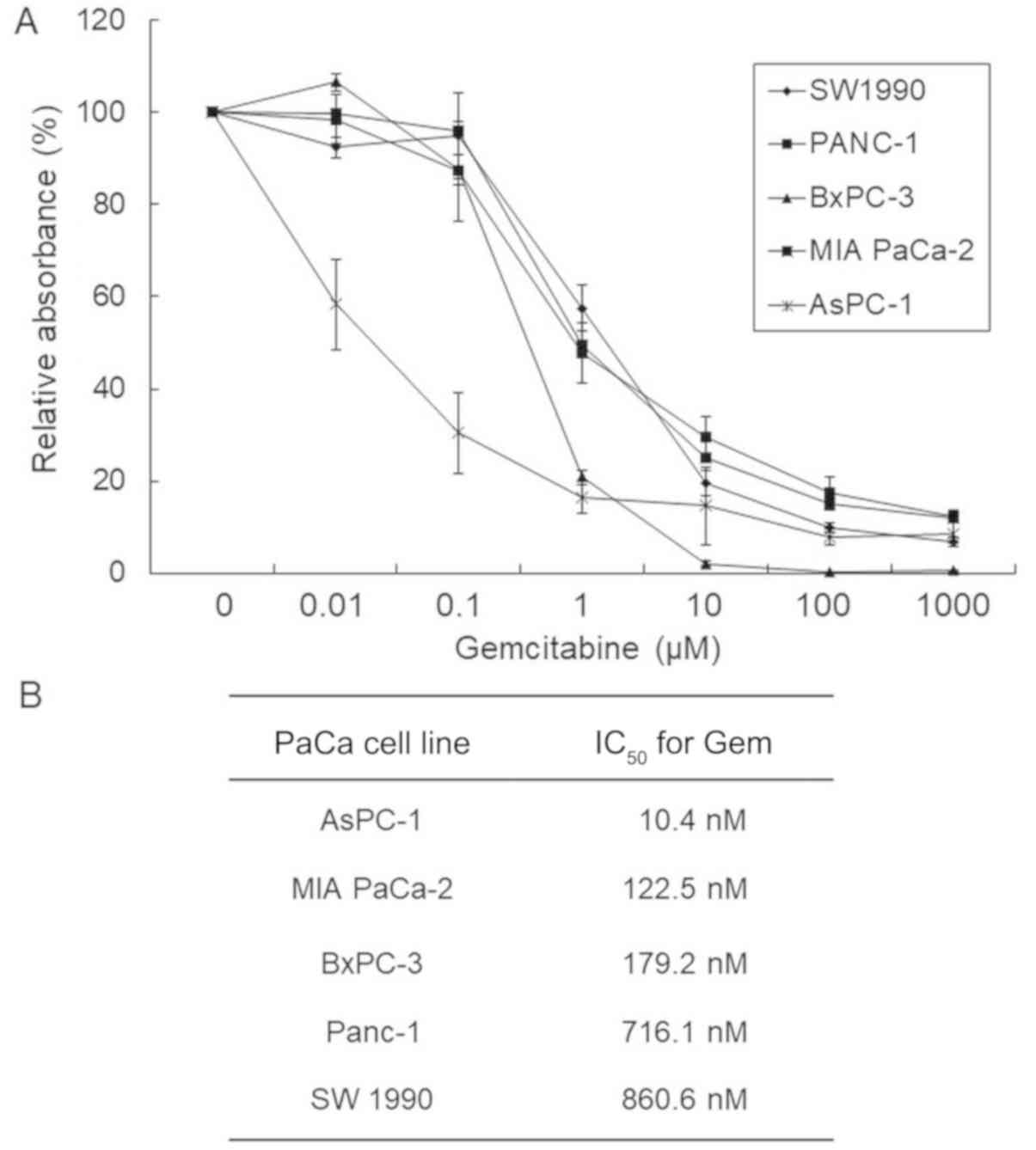
assays. The IC50 of Gem for each PaCa line evaluated was
determined by constructing dose-response curves (Fig. 1A). The IC50 values of Gem
after 72 h of treatment were 10.4, 179.2, 122.5, 716.1 and 850.6 nM
for AsPC-1, BxPC-3, MIA PaCa-2, Panc-1 and SW 1990 cells,
respectively (Fig. 1B).
Establishment of Gem-R PaCa cell
lines
We incubated the five PaCa cell lines with
IC50 values of Gem and successfully established two
Gem-R PaCa cell lines from AsPC-1 and MIA PaCa-2 cells. The
IC50 values were 7,122.3 and 10.4 nM for Gem-R and Gem-S
AsPC-1 cells, respectively, and 12,614.3 and 122.5 nM for Gem-R and
Gem-S MIA PaCa-2 cells, respectively (Fig. 2). We investigated the biological
differences between the Gem-S and Gem-R cell lines in subsequent
experiments.
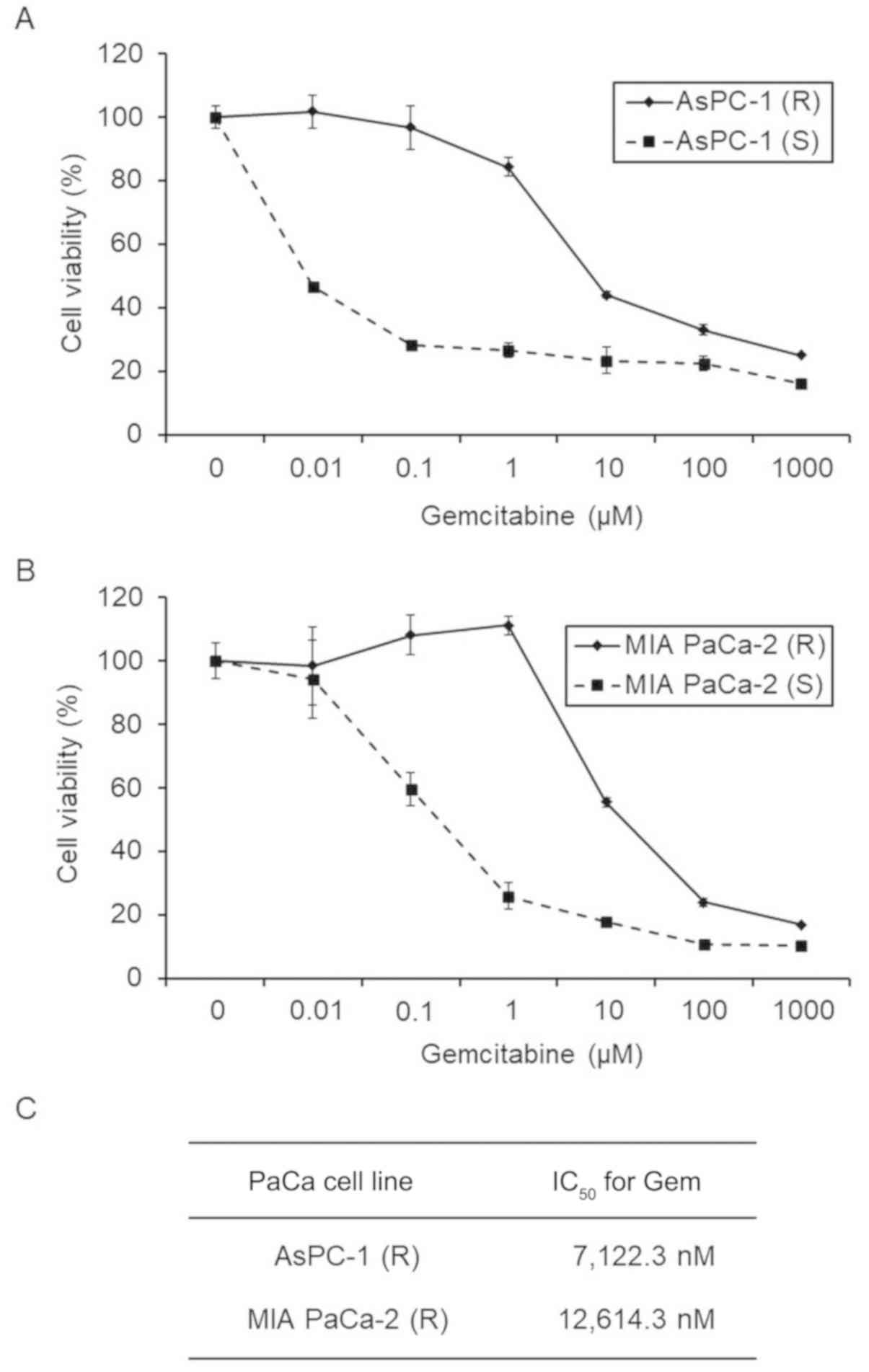 | Figure 2.Effect of Gem on the proliferation of
Gem-R and Gem-S PaCa cell lines. (A and B) Gem-R and Gem-S AsPC-1
(A) and MIA PaCa-2 (B) cells were treated with Gem at the indicated
concentrations for 72 h, and the proliferation of each cell line
was determined using WST-1 assays. Values are expressed as means ±
SD. (C) The IC50 values of Gem in the Gem-R PaCa cell
lines. AsPC-1 (R), Gem-R AsPC-1 cells; AsPC-1 (S), Gem-S AsPC-1
cells; MIA PaCa-2 (R), Gem-R MIA PaCa-2 cells; MIAPaCa-2 (S), Gem-S
MIA PaCa-2 cells; Gem, gemcitabine; PaCa, pancreatic cancer; Gem-R,
gemcitabine resistant; Gem-S, gemcitabine sensitive;
IC50, half maximal inhibitory concentration. |
Proliferation of Gem-S and Gem-R PaCa
cell lines
To compare cell proliferation between the Gem-R and
Gem-S PaCa cell lines, we performed proliferation assays. There was
no significant difference in proliferation between the Gem-R and
Gem-S cells of either line after 24–96 h of culture (Fig. 3).
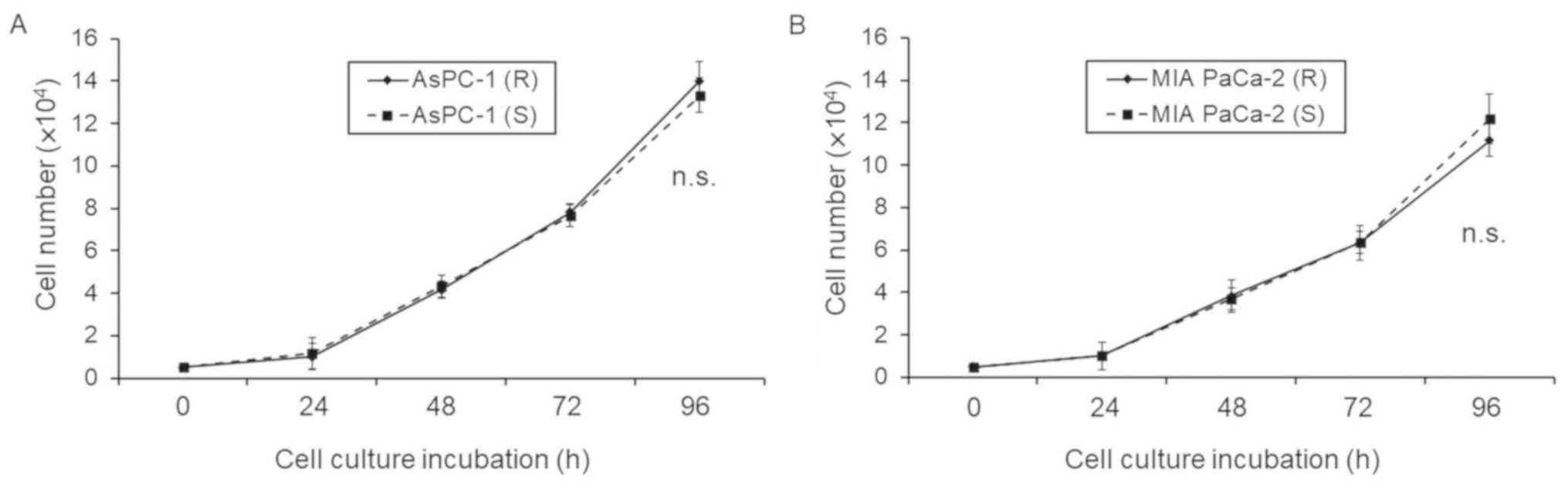 | Figure 3.Differences in proliferation between
Gem-R and Gem-S cells. (A and B) Gem-R and Gem-S AsPC-1 (A) and (B)
MIA PaCa-2 (B) cells were seeded (2×104 cells) in 6-well
plates and then collected after 0–96 h using 0.25% Trypsin-EDTA and
counted in four microscopic fields (×100) using a compound light
microscope. Values are expressed as means ± SD. NS, not
significant. There were no significant differences in cell numbers
between the Gem-R and Gem-S cells of each cell line. AsPC-1 (R),
Gem-R AsPC-1 cells; AsPC-1 (S), Gem-S AsPC-1 cells; MIA PaCa-2 (R),
Gem-R MIA PaCa-2 cells; MIAPaCa-2 (S), Gem-S MIA PaCa-2 cells; Gem,
gemcitabine; PaCa, pancreatic cancer; Gem-R, gemcitabine resistant;
Gem-S, gemcitabine sensitive; IC50, half maximal
inhibitory concentration; n.s., not significant. |
cDNA microarray analysis of Gem-S and
Gem-R MIA PaCa-2 cells
To investigate comprehensive differences in mRNA
expression between Gem-S and Gem-R MIA PaCa-2 cells, we used a cDNA
microarray containing 54,765 probe sets. Of these probes, 1,206
showed higher expression (cut-off value, 2-fold) and 3,157 lower
expression (cut-off value, 0.5-fold) in Gem-R compared with Gem-S
MIA PaCa-2 cells (Table I). Among
the genes with higher expression in the Gem-R cells, we focused on
IL-8 as we previously reported an important role of IL-8 in PaCa
angiogenesis (24).
 | Table I.cDNA microarray (54,675 genes). |
Table I.
cDNA microarray (54,675 genes).
| Differentially
expressed genes in Gem-R cells | No. of genes |
|---|
| Upregulated genes
(>2-fold) | 1,206 |
| Downregulated genes
(<0.5-fold) | 3,157 |
RT-qPCR analysis of IL-8 mRNA
expression in Gem-S and Gem-R PaCa cell lines
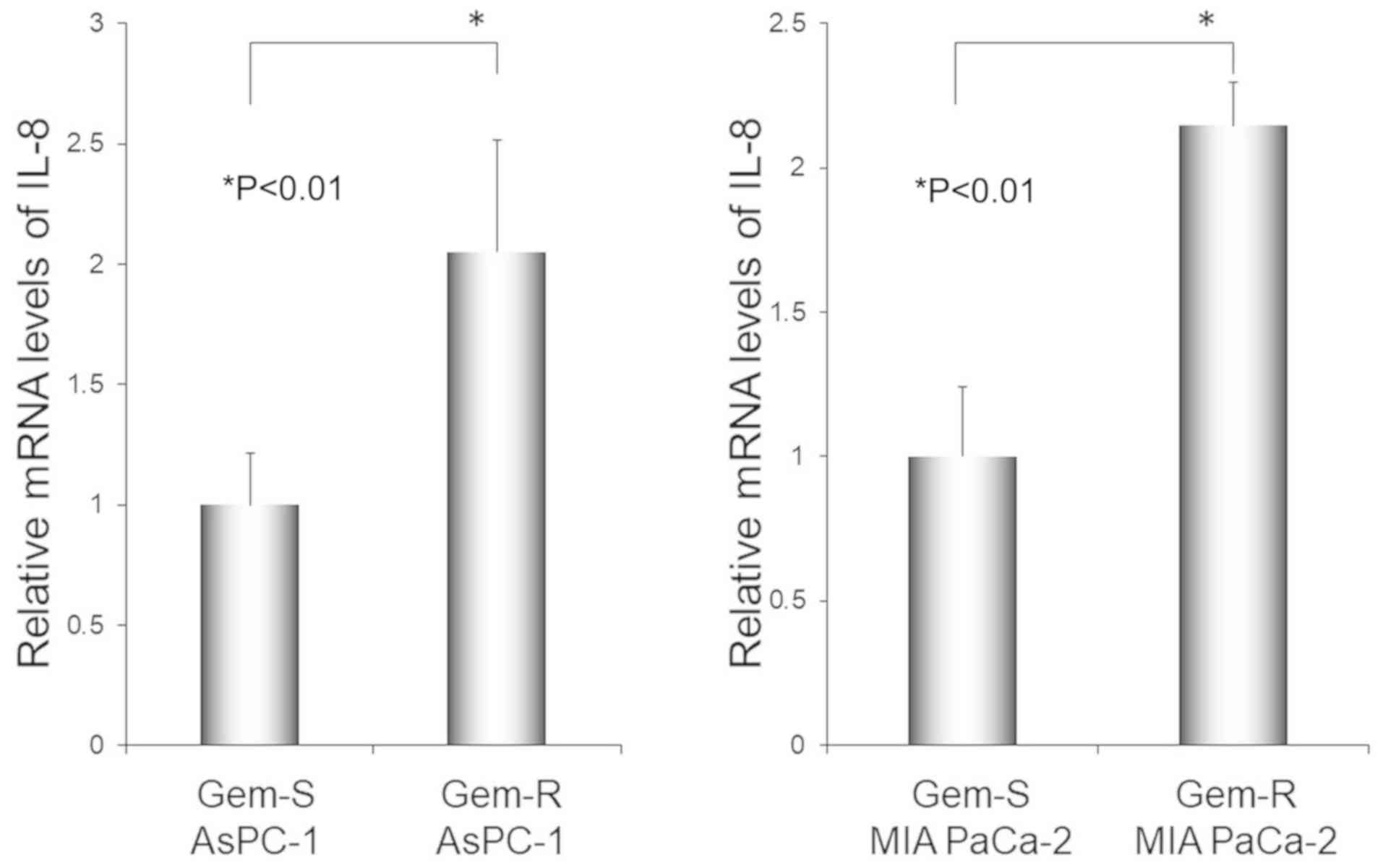
RT-qPCR revealed higher expression of IL-8 mRNA in
Gem-R when compared with Gem-S cells in both PaCa cell lines
evaluated (P<0.01 in both AsPC-1 cells and MIA PaCa-2 cells)
(Fig. 4). The RT-qPCR results were
consistent with the cDNA microarray results.
Upregulated IL-8 secretion by Gem-R
compared with Gem-S PaCa cell lines
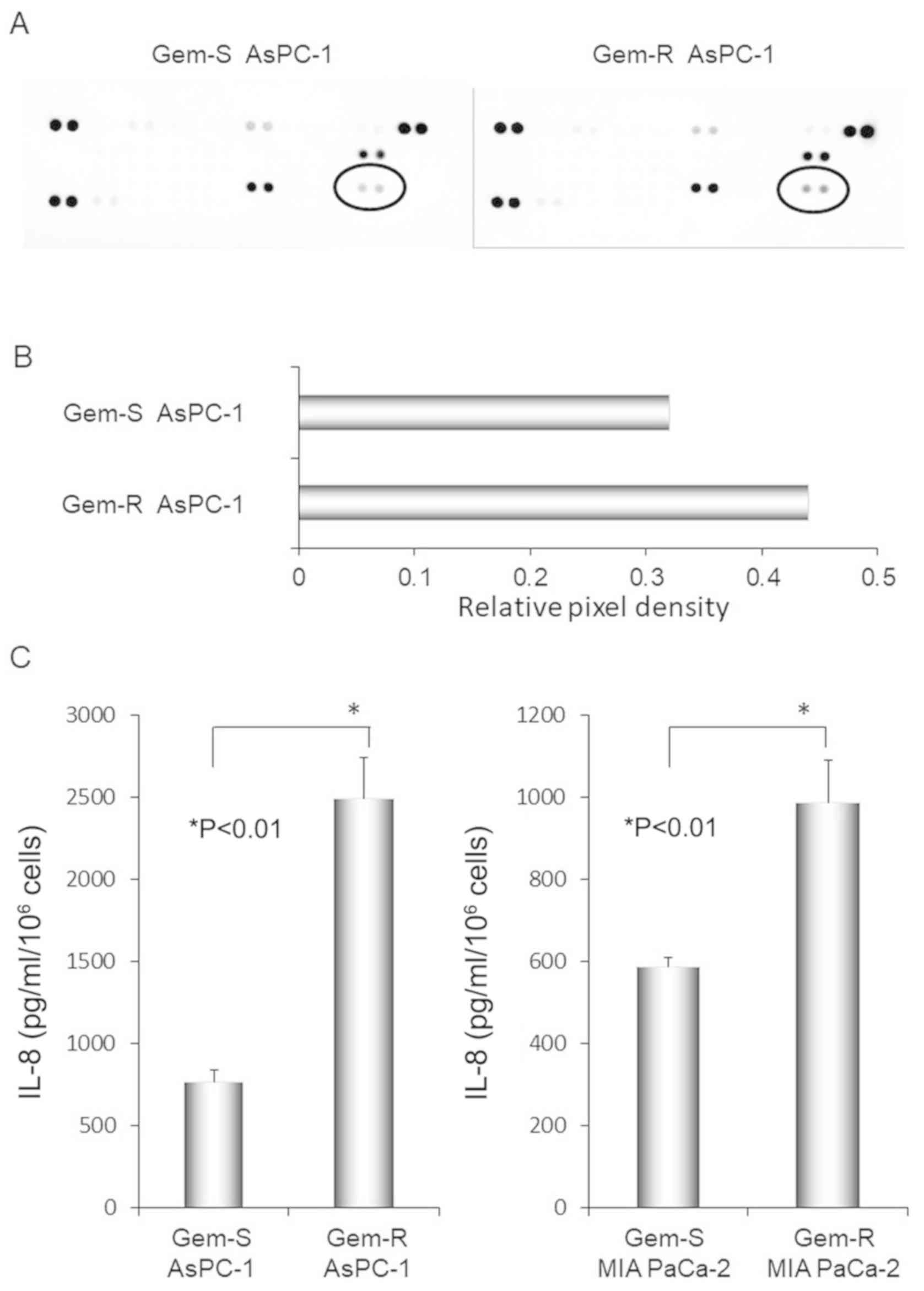
We examined differences in chemokine secretion by
Gem-R and Gem-S AsPC-1 cells using cytokine arrays (Fig. 5A). The level of IL-8 was elevated in
the culture supernatants of Gem-R compared with Gem-S cells, with
signal intensity values of 0.44 and 0.32, respectively (Fig. 5B). We next examined IL-8 protein
secretion by PaCa cell lines using ELISA (Fig. 5C). In both MIA PaCa-2 and AsPC-1
cell lines, IL-8 secretion was significantly enhanced in the Gem-R
compared with the Gem-S cell lines (P<0.01 for both cell
lines).
Effect of IL-8 administration on the
proliferation of PaCa cell lines
Gem-S PaCa cells were treated with or without IL-8
and various concentrations of Gem and then assessed by WST-1
proliferation assay. There was no significant difference in the
proliferation of Gem-S MIA PaCa-2 cells treated with vs. not
treated with IL-8 administration at any Gem concentration evaluated
(Fig. S1).
Effect of siRNA-mediated IL-8
knockdown on the proliferation of Gem-R PaCa cells
RT-qPCR confirmed successful siRNA-mediated
knockdown of IL-8 in Gem-R MIA PaCa-2 cells (P<0.01) (Fig. S2A). The effect of siRNA-mediated
knockdown of IL-8 in Gem-R MIA PaCA-2 cells was assessed by WST-1
proliferation assays. There was no significant difference in the
proliferation of Gem-R MIA PaCa-2 cells with vs. without IL-8
siRNA-mediated knockdown at any Gem concentration evaluated
(Fig. S2B).
Effects of Gem-R development and an
anti-CXCR2 Ab on HUVEC tube formation
To estimate the effect of Gem-R on angiogenesis, we
performed an in vitro angiogenesis assay. HUVECs and
fibroblasts were co-cultured with PaCa cell lines, and the effects
of an anti-CXCR2 Ab were examined. Tube formation by HUVECs was
significantly enhanced by co-culture with Gem-R compared with Gem-S
PaCa cells (P<0.01 for both AsPC-1 and MIA PaCa-2 cells).
Moreover, the enhanced tube formation by PaCa cells was inhibited
by treatment with (+) the anti-CXCR2 Ab (P<0.05 for Gem-R MIA
PaCa-2 cells and P<0.01 for Gem-R and Gem-S AsPC-1 cells and
Gem-R MIA PaCa-2 cells) (Fig.
6).
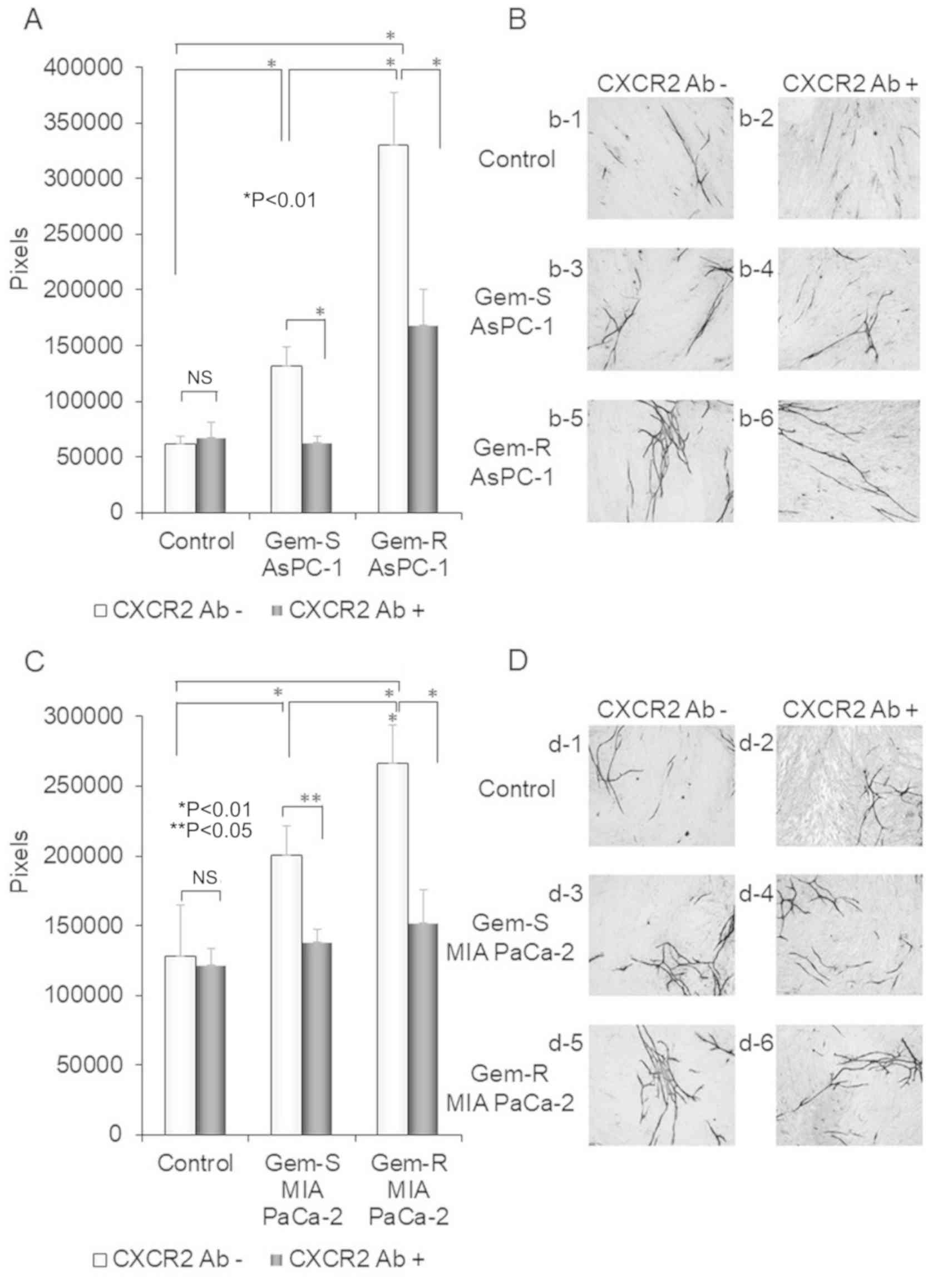 | Figure 6.Differences in tube formation by
HUVECs co-cultured with Gem-R and Gem-S PaCa cells and the effect
of an anti-CXCR2 Ab. (A-D) After incubation of HUVECs, fibroblasts
and the PaCa cell lines [Gem-R and Gem-S AsPC-1 cells (A and B) and
MIA PaCa-2 cells (C and D)] for 11 days using the double-chamber
method, HUVECs were stained with an anti-CD31 Ab. The tube
formation area was measured using an image analyzer (magnification
×40). Controls were co-cultured without PaCa cells. Values are
expressed as means ± SD. Multiple group comparisons were performed
by one-way analysis of variance (ANOVA) with a post hoc Bonferroni
test. *P<0.01, **P<0.05; NS, not significant. (B) (b-1)
co-cultured without PaCa cells and CXCR2 Ab (−); (b-2) co-cultured
without PaCa cells but with CXCR2 Ab (+); (b-3) co-cultured with
Gem-S AsPC-1 cells but without CXCR2 Ab (−); (b-4) co-cultured with
Gem-S AsPC-1 cells and CXCR2 Ab (+); (b-5) co-cultured with Gem-R
AsPC-1 cells but without CXCR2 Ab (−); (b-6) co-cultured with Gem-R
AsPC-1 cells and CXCR2 Ab (+). (D) (d-1) co-cultured without PaCa
cells and CXCR2 Ab (−); (d-2) co-cultured without PaCa cells but
with CXCR2 Ab (+); (d-3) co-cultured with Gem-S MIA PaCa-2 cells
but without CXCR2 Ab (−); (d-4) co-cultured with Gem-S MIA PaCa-2
cells and CXCR2 Ab (+); (d-5) co-cultured with Gem-R MIA PaCa-2
cells but without CXCR2 Ab (−); (d-6) co-cultured with Gem-R MIA
PaCa-2 cells and CXCR2 Ab (+). CXCR2 Ab -, not treated with CXCR2
antibody; CXCR2 Ab (+), treated with CXCR2 Antibody. Gem,
gemcitabine; PaCa, pancreatic cancer; Gem-R, gemcitabine resistant;
Gem-S, gemcitabine sensitive; CXCR2, C-X-C motif chemokine receptor
2; HUVECs, human umbilical vein endothelial cells; Ab, antibody. (B
and D) Original magnification, ×40. |
Discussion
In the present study, two main findings were
confirmed. First, the resistance of pancreatic cancer (PaCa) cells
to gemcitabine (Gem) was associated with increased interleukin
(IL)-8 production from PaCa cells. Second, an anti-CXCR2 antibody
(Ab), a neutralizing Ab for the main receptor of IL-8, inhibited
the increase in angiogenesis induced upon acquisition of Gem
resistance (Gem-R) in PaCa cells. Therefore, regulation of the
IL-8/CXCR2 axis modulates the angiogenic activity and tumorgenicity
of Gem-R PaCa cells.
Although chemotherapeutic strategies for PaCa are
advancing, the therapeutic effects are still insufficient.
Gem-based chemotherapy is one of the standard treatments for
patients with advanced PaCa. However, clinicians often find that
Gem-based treatment usually has good initial effects, but that
these effects often weaken shortly thereafter. One of the reasons
for this is acquisition of Gem-R. To improve the effect of Gem
chemotherapy on patient prognosis, a better understanding of the
mechanism by which PaCa acquires Gem-R and development of new
therapeutic strategies to overcome this resistance are
required.
Previous studies have suggested several mechanisms
of Gem-R. Gem-R can be acquired via molecular and cellular changes,
including dysregulation of Gem metabolic pathways [e.g.,
deoxycytidine kinase and ribonucleotide reductase subunits M1 and
M2 (25,26)] and key signaling pathways [e.g.,
NF-κB (27,28), PI3K/Akt (28), MAPK, ERK1/2 (29), HIF-1α (30)], increased expression of drug efflux
pumps, activation of cancer stem cells (31), and epithelial-to-mesenchymal
transition (32). However, a
breakthrough clarifying the mechanism of Gem-R or identifying the
appropriate secondary treatments have not been realized to
date.
IL-8 is a pro-inflammatory factor belonging to the
CXC chemokine family that is secreted by various cells to activate
and recruit leukocytes to sites of infection and injury. Recently,
several studies have reported that IL-8 plays important roles in
cancer progression, angiogenesis and metastasis (33–35).
Moreover, we previously demonstrated that IL-8 is an important
component of the tumor microenvironment in PaCa, and that PaCa
cells expressing high IL-8 levels have greater liver metastatic
potential and angiogenic ability compared with PaCa cells
expressing low IL-8 levels (21,24).
These results suggest that IL-8 produced by PaCa cells regulates
cancer progression and metastasis and may be an important
therapeutic target. We also revealed that blockade of the
IL-8/CXCR2 pathway significantly inhibited PaCa tumor growth by
preventing angiogenesis, in vitro and in vivo
(20). IL-8 activity occurs mainly
via interaction with its specific cell-surface G protein-coupled
receptor, CXCR2 (33,36,37).
In our previous study, to inhibit the IL-8/CXCR2 pathway, we used
both an anti-CXCR2 neutralizing Ab and a small-molecule inhibitor
of CXCR2 (SB225002) (38); both had
similar effects.
A previous study showed that IL-8 is related to
Gem-R and enhances the invasiveness of Gem-R PaCa (39). However, the specific role of IL-8 in
Gem-R PaCa has not been sufficiently elucidated, particularly with
respect to tumor angiogenesis. We reported that IL-8 plays an
important role in tumor angiogenesis to affect PaCa metastasis and
progression (40). In addition, we
successfully established Gem-R PaCa cell lines and conducted
comprehensive gene expression analyses. Among the altered genes, we
found that the expression of IL-8, which was focused on in the
present study, was increased in the Gem-R cells. Therefore, it is
reasonable to expect that elucidating the mechanism of IL-8
involvement in the acquisition of Gem-R in PaCa will be important
for the future development of PaCa treatments. Based on our
previous and present research findings, IL-8 was selected as the
focus of the present study.
CXCR2 is a cell-surface chemokine G protein-coupled
receptor with seven transmembrane domains. CXCR2 has been found on
the surfaces of endothelial, epithelial, and multiple tumor cells.
Chemokines containing a Glu-Leu-Arg motif, such as CXCL1, CXCL2,
CXCL3, CXCL5, CXCL6, and CXCL7 and IL-8, bind to CXCR2, and their
interaction with CXCR2 tends to enhance tumor angiogenesis
(37). High expression of CXCR2
showed an association with poor prognosis in laryngeal squamous
cell carcinoma, lung cancer, clear-cell renal cell carcinoma,
hepatocellular carcinoma, and pancreatic ductal carcinoma (41). There is abundant evidence on Gem-R
and IL-8 (39,42,43),
but little is known concerning the effect of Gem-R on IL-8-induced
angiogenesis in PaCa. The mechanisms underlying how Gem-R
acquisition induces IL-8 production in PaCa cells are not clear,
but it was reported that Gem promotes NF-κB activity in PaCa
(42,44). As NF-κB regulates IL-8 production
(40), we speculate that IL-8
production in Gem-R PaCa cells is associated with NF-κB activity,
which is activated by Gem.
In the present study, to analyze the mechanism of
Gem-R, we successfully established two Gem-R PaCa cell lines. Based
on a comprehensive gene expression analysis, we found that the
expression of IL-8 was elevated in the Gem-R PaCa cell lines. There
was no significant change in proliferation between Gem-R and Gem-S
cells. However, our in vitro angiogenesis assay indicated a
significant increase in angiogenic activity in Gem-R cells. In this
assay, we used a HUVEC/fibroblast co-culture system to assess the
interactions between tumor cells and the tumor microenvironment and
clearly demonstrated that an anti-CXCR2 Ab prevented the angiogenic
activity induced by Gem-R. These results suggest that upregulation
of IL-8 in Gem-R PaCa enhances angiogenesis via interaction with
CXCR2, and blockade of the IL-8/CXCR2 signaling axis effectively
reduces angiogenesis. These findings are consistent with previous
studies that suggest that IL-8 expression is enhanced by Gem
treatment in response to chemotherapy (42).
To determine whether the increase in IL-8 production
is the result vs. the initial cause of Gem-R acquisition, we
examined the effect of suppression or overexpression of IL-8 on
Gem-R. As neither recombinant IL-8 nor IL-8 siRNA treatment
affected the proliferation of Gem-R cells compared with Gem-S
cells, we believe that the increase in IL-8 production may be the
result, rather than the cause, of Gem-R.
In conclusion, we showed that acquisition of Gem-R
increased IL-8 production. This increase in IL-8 production
enhanced tumor angiogenesis in a co-culture environment consisting
of PaCa and stromal cells. Importantly, we also demonstrated that
blocking the IL-8/CXCR2 signaling axis using an anti-CXCR2 Ab
affected Gem-R PaCa angiogenic activity. Based on these results,
the IL-8/CXCR2 axis may be a potential novel therapeutic target for
Gem-R PaCa.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the JSPS KAKENHI
(grant no. 25462123).
Availability of data and materials
The datasets relevant to this study are included
within the article.
Authors' contributions
Conception and design of the study were conducted by
HI, YM and SK. Development of the study methodology was carried out
by HI, YM, GU, KO, YH, KS, KT, MM, SK, RO, MH, HT and ST.
Acquisition of the data was conducted by HI, YM and SK. Analysis
and interpretation of the data were carried out by HI, YM and SK.
Writing, review, and/or revision of the manuscript was carried out
by HI, YM and SK. Writing of the manuscript in regards to the
methods section by GU, KO, YH, KS, KT, MM, RO and HT.
Administrative, technical, or material support was accomplished by
YM and ST. Study supervision was carried out by YM. All authors
read and approved the final manuscript and agree to be accountable
for all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
Gem
|
gemcitabine
|
|
PaCa
|
pancreatic cancer
|
|
Gem-R
|
gemcitabine resistance
|
|
Gem-S
|
gemcitabine sensitive
|
|
IL-8
|
interleukin-8
|
|
CXCR2
|
C-X-C motif chemokine receptor 2
|
|
FBS
|
fetal bovine serum
|
|
IC50
|
half maximal inhibitory
concentration
|
|
RT-qPCR
|
reverse transcription-quantitative
polymerase chain reaction
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
HUVECs
|
human umbilical vein endothelial
cells
|
|
Ab
|
antibody
|
|
SD
|
standard deviation
|
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ilic M and Ilic I: Epidemiology of
pancreatic cancer. World J Gastroenterol. 22:9694–9705. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Niedergethmann M, Alves F, Neff JK,
Heidrich B, Aramin N, Li L, Pilarsky C, Grützmann R, Allgayer H,
Post S, et al: Gene expression profiling of liver metastases and
tumour invasion in pancreatic cancer using an orthotopic SCID mouse
model. Br J Cancer. 97:1432–1440. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kamisawa T, Wood LD, Itoi T and Takaori K:
Pancreatic cancer. Lancet. 388:73–85. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gillen S, Schuster T, Meyer Zum
Büschenfelde C, Friess H and Kleeff J: Preoperative/neoadjuvant
therapy in pancreatic cancer: A systematic review and meta-analysis
of response and resection percentages. PLoS Med. 7:e10002672010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wong HH and Lemoine NR: Pancreatic cancer:
Molecular pathogenesis and new therapeutic targets. Nat Rev
Gastroenterol Hepatol. 6:412–422. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Burris HA III, Moore MJ, Andersen J, Green
MR, Rothenberg ML, Modiano MR, Cripps MC, Portenoy RK, Storniolo
AM, Tarassoff P, et al: Improvements in survival and clinical
benefit with gemcitabine as first-line therapy for patients with
advanced pancreas cancer: A randomized trial. J Clin Oncol.
15:2403–2413. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Raman D, Baugher PJ, Thu YM and Richmond
A: Role of chemokines in tumor growth. Cancer Lett. 256:137–165.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kulbe H, Levinson NR, Balkwill F and
Wilson JL: The chemokine network in cancer-much more than directing
cell movement. Int J Dev Biol. 48:489–496. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Baggiolini M, Walz A and Kunkel SL:
Neutrophil-activating peptide-1/interleukin 8, a novel cytokine
that activates neutrophils. J Clin Invest. 84:1045–1049. 1989.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kuwada Y, Sasaki T, Morinaka K, Kitadai Y,
Mukaida N and Chayama K: Potential involvement of IL-8 and its
receptors in the invasiveness of pancreatic cancer cells. Int J
Oncol. 22:765–771. 2003.PubMed/NCBI
|
|
12
|
Brat DJ, Bellail AC and Van Meir EG: The
role of interleukin-8 and its receptors in gliomagenesis and
tumoral angiogenesis. Neuro Oncol. 7:122–133. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Konno H, Ohta M, Baba M, Suzuki S and
Nakamura S: The role of circulating IL-8 and VEGF protein in the
progression of gastric cancer. Cancer Sci. 94:735–740. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Todorović-Raković N and Milovanović J:
Interleukin-8 in breast cancer progression. J Interferon Cytokine
Res. 33:563–570. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lin Y, Huang R, Chen L, Li S, Shi Q,
Jordan C and Huang RP: Identification of interleukin-8 as estrogen
receptor-regulated factor involved in breast cancer invasion and
angiogenesis by protein arrays. Int J Cancer. 109:507–515. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shi J and Wei PK: Interleukin-8: A potent
promoter of angiogenesis in gastric cancer. Oncol Lett.
11:1043–1050. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ning Y, Manegold PC, Hong YK, Zhang W,
Pohl A, Lurje G, Winder T, Yang D, LaBonte MJ, Wilson PM, et al:
Interleukin-8 is associated with proliferation, migration,
angiogenesis and chemosensitivity in vitro and in vivo in colon
cancer cell line models. Int J Cancer. 128:2038–2049. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jia L, Li F, Shao M, Zhang W, Zhang C,
Zhao X, Luan H, Qi Y, Zhang P, Liang L, et al: IL-8 is upregulated
in cervical cancer tissues and is associated with the proliferation
and migration of HeLa cervical cancer cells. Oncol Lett.
15:1350–1356. 2018.PubMed/NCBI
|
|
19
|
Srivastava SK, Bhardwaj A, Arora S, Tyagi
N, Singh AP, Carter JE, Scammell JG, Fodstad Ø and Singh S:
Interleukin-8 is a key mediator of FKBP51-induced melanoma growth,
angiogenesis and metastasis. Br J Cancer. 112:1772–1781. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Matsuo Y, Raimondo M, Woodward TA, Wallace
MB, Gill KR, Tong Z, Burdick MD, Yang Z, Strieter RM, Hoffman RM
and Guha S: CXC-chemokine/CXCR2 biological axis promotes
angiogenesis in vitro and in vivo in pancreatic cancer. Int J
Cancer. 125:1027–1037. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Matsuo Y, Sawai H, Funahashi H, Takahashi
H, Sakamoto M, Yamamoto M, Okada Y, Hayakawa T and Manabe T:
Enhanced angiogenesis due to inflammatory cytokines from pancreatic
cancer cell lines and relation to metastatic potential. Pancreas.
28:344–352. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Osugi T, Oshima Y, Fujio Y, Funamoto M,
Yamashita A, Negoro S, Kunisada K, Izumi M, Nakaoka Y, Hirota H, et
al: Cardiac-specific activation of signal transducer and activator
of transcription 3 promotes vascular formation in the heart. J Biol
Chem. 277:6676–6681. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bishop ET, Bell GT, Bloor S, Broom IJ,
Hendry NF and Wheatley DN: An in vitro model of angiogenesis: Basic
features. Angiogenesis. 3:335–344. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Matsuo Y, Ochi N, Sawai H, Yasuda A,
Takahashi H, Funahashi H, Takeyama H, Tong Z and Guha S: CXCL8/IL-8
and CXCL12/SDF-1alpha co-operatively promote invasiveness and
angiogenesis in pancreatic cancer. Int J Cancer. 124:853–861. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ohhashi S, Ohuchida K, Mizumoto K, Fujita
H, Egami T, Yu J, Toma H, Sadatomi S, Nagai E and Tanaka M:
Down-regulation of deoxycytidine kinase enhances acquired
resistance to gemcitabine in pancreatic cancer. Anticancer Res.
28:2205–2212. 2008.PubMed/NCBI
|
|
26
|
Minami K, Shinsato Y, Yamamoto M,
Takahashi H, Zhang S, Nishizawa Y, Tabata S, Ikeda R, Kawahara K,
Tsujikawa K, et al: Ribonucleotide reductase is an effective target
to overcome gemcitabine resistance in gemcitabine-resistant
pancreatic cancer cells with dual resistant factors. J Pharmacol
Sci. 127:319–325. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Waters JA, Matos J, Yip-Schneider M,
Aguilar-Saavedra JR, Crean CD, Beane JD, Dumas RP, Suvannasankha A
and Schmidt CM: Targeted nuclear factor-kappaB suppression enhances
gemcitabine response in human pancreatic tumor cell line murine
xenografts. Surgery. 158:881–889. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Arlt A, Gehrz A, Müerköster S, Vorndamm J,
Kruse ML, Fölsch UR and Schäfer H: Role of NF-kappaB and Akt/PI3K
in the resistance of pancreatic carcinoma cell lines against
gemcitabine-induced cell death. Oncogene. 22:3243–3251. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zheng C, Jiao X, Jiang Y and Sun S: ERK1/2
activity contributes to gemcitabine resistance in pancreatic cancer
cells. J Int Med Res. 41:300–306. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang R, Cheng L, Xia J and Wang Z, Wu Q
and Wang Z: Gemcitabine resistance is associated with
epithelial-mesenchymal transition and induction of HIF-1α in
pancreatic cancer cells. Curr Cancer Drug Targets. 14:407–417.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen M, Xue X, Wang F, An Y, Tang D, Xu Y,
Wang H, Yuan Z, Gao W, Wei J, et al: Expression and promoter
methylation analysis of ATP-binding cassette genes in pancreatic
cancer. Oncol Rep. 27:265–269. 2012.PubMed/NCBI
|
|
32
|
Quint K, Tonigold M, Di Fazio P,
Montalbano R, Lingelbach S, Rückert F, Alinger B, Ocker M and
Neureiter D: Pancreatic cancer cells surviving gemcitabine
treatment express markers of stem cell differentiation and
epithelial-mesenchymal transition. Int J Oncol. 41:2093–2102. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu Q, Li A, Tian Y, Wu JD, Liu Y, Li T,
Chen Y, Han X and Wu K: The CXCL8-CXCR1/2 pathways in cancer.
Cytokine Growth Factor Rev. 31:61–71. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li A, Dubey S, Varney ML, Dave BJ and
Singh RK: IL-8 directly enhanced endothelial cell survival,
proliferation, and matrix metalloproteinases production and
regulated angiogenesis. J Immunol. 170:3369–3376. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Heidemann J, Ogawa H, Dwinell MB, Rafiee
P, Maaser C, Gockel HR, Otterson MF, Ota DM, Lugering N, Domschke W
and Binion DG: Angiogenic effects of interleukin 8 (CXCL8) in human
intestinal microvascular endothelial cells are mediated by CXCR2. J
Biol Chem. 278:8508–8515. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Olson TS and Ley K: Chemokines and
chemokine receptors in leukocyte trafficking. Am J Physiol Regul
Integr Comp Physiol. 283:R7–R28. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hertzer KM, Donald GW and Hines OJ: CXCR2:
A target for pancreatic cancer treatment? Expert Opin Ther Targets.
17:667–680. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Matsuo Y, Campbell PM, Brekken RA, Sung B,
Ouellette MM, Fleming JB, Aggarwal BB, Der CJ and Guha S: K-Ras
promotes angiogenesis mediated by immortalized human pancreatic
epithelial cells through mitogen-activated protein kinase signaling
pathways. Mol Cancer Res. 7:799–808. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pan MR, Hsu MC, Luo CW, Chen LT, Shan YS
and Hung WC: The histone methyltransferase G9a as a therapeutic
target to override gemcitabine resistance in pancreatic cancer.
Oncotarget. 7:61136–61151. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Matsuo Y, Sawai H, Ochi N, Yasuda A,
Sakamoto M, Takahashi H, Funahashi H, Takeyama H and Guha S:
Proteasome inhibitor MG132 inhibits angiogenesis in pancreatic
cancer by blocking NF-kappaB activity. Dig Dis Sci. 55:1167–1176.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Qiao B, Luo W, Liu Y, Wang J, Liu C, Liu
Z, Chen S, Gu J, Qi X and Wu T: The prognostic value of CXC
chemokine receptor 2 (CXCR2) in cancers: A meta-analysis.
Oncotarget. 9:15068–15076. 2017.PubMed/NCBI
|
|
42
|
Song Y, Baba T, Li YY, Furukawa K, Tanabe
Y, Matsugo S, Sasaki S and Mukaida N: Gemcitabine-induced CXCL8
expression counteracts its actions by inducing tumor
neovascularization. Biochem Biophys Res Commun. 458:341–346. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Khan MA, Srivastava SK, Bhardwaj A, Singh
S, Arora S, Zubair H, Carter JE and Singh AP: Gemcitabine triggers
angiogenesis-promoting molecular signals in pancreatic cancer
cells: Therapeutic implications. Oncotarget. 6:39140–39150. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Maliandi MV, Mato-Berciano A, Sobrevals L,
Roué G, José A and Fillat C: AduPARE1A and gemcitabine combined
treatment trigger synergistic antitumor effects in pancreatic
cancer through NF-κB mediated uPAR activation. Mol Cancer.
14:1462015. View Article : Google Scholar : PubMed/NCBI
|