Introduction
In this decade, the study of cancer
microenvironments has garnered attention as it may help to
elucidate novel anticancer mechanisms, which have not been
exploited with the use of conventional cytotoxic anticancer drugs
(1). Various mesenchymal cells
observed in tumors have been hypothesized to serve passive roles in
tumor growth (1). However, studies
have revealed that these mesenchymal cells instead serve an active
and irreplaceable role in cancer cell growth in terms of
proliferation, invasion and angiogenesis (2,3).
Angiogenesis is an essential mechanism for the metastasis and
proliferation of cancer cells (4).
Thus, anti-angiogenetic drugs, including bevacizumab and
aflibercept, are regarded as important for colorectal cancer
therapy (5). It has been
hypothesized that angiogenesis in cancer is caused by cancer cells
(5). However, recent studies
revealed that stromal cells in cancer cause angiogenesis (6,7).
Additional studies are needed to clarify the mechanism of
angiogenesis caused by stromal cells. An increasing number of
studies have focused on the interaction between tumor cells and
stromal cells (cancer-stromal interaction, CSI), which serves an
important role in the growth of several cancers (1,8,9). Among
them, a previous study by our group demonstrated that inflammation
causes VEGF secretion from cancer-associated fibroblasts (CAFs) in
tumors via interleukin (IL)-6, which is secreted by CAFs and not by
cancer cells (10). IL-6 may be the
key cytokine that promotes VEGF secretion from CAFs and thus VEGF
levels may be decreased by a drug that decreases IL-6 levels.
Eicosapentaenoic acid (EPA), a polyunsaturated fatty
acid (PUFA), is a well-known fish oil; EPA has been reported to
decease systemic inflammation, which is caused by IL-6 (11,12).
EPA has also been demonstrated to decrease inflammation caused by
cancer (13). It has reported that
EPA decreased IL-6 secretion in esophageal cancer cell lines
(11). Nevertheless, a previous
study by our group has revealed that a greater amount of IL-6 was
released by cancer stromal cells than by cancer cells (10). At present, various mechanisms of EPA
that effect cancer cells have been clarified (14–19).
Certain PUFAs have also been revealed to inhibit VEGF expression in
colon cancer cells (20,21). However, how EPA influences cancer
stromal cells and the CSI has not yet been demonstrated. Thus, in
the present study, the authors investigated the in vitro
effects of EPA on cancer stromal cells, with regards to
angiogenesis.
Materials and methods
Agents
Sodium salts of 5-, 8-, 11-, 14-, and 17-EPA were
procured from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). EPA
(5 mg) was added to 154 µl of 100% ethanol to prepare the stock
solution, which was then diluted to yield a final ethanol
concentration of 0.03%. Lipopolysaccharide (LPS, from E.
coli O111:B4; 10 mg; Sigma-Aldrich; Merck KGaA) was
concentrated to yield a final concentration of 50 µg/ml. Anti-IL-6
myeloma receptor antibodies (MRA) were kindly provided for free by
Chugai Pharmaceutical Co., Ltd. (Tokyo, Japan). To certify the
established cell lines as fibroblasts, anti-α-smooth muscle actin
(α-SMA) monoclonal (1A4; cat. no. ab7817), anti-IL-6 (cat. no.
ab9324) and anti-VEGF (cat. no. ab52917) antibodies were purchased
from Abcam (Cambridge, MA, USA) and used for immunofluorescence
staining. Additionally, anti-vimentin (V9; cat. no. M072529),
anti-cytokeratin (AE1/AE3; cat. no. M351529; both Dako; Agilent
Technologies, Inc., Santa Clara, CA, USA) and anti-cluster of
differentiation (CD) 90 (5E10; cat. no. BD-555593; BD Pharmingen;
BD Biosciences, San Jose, CA, USA) mouse monoclonal antibodies were
purchased. To confirm the effect of EPA on fibroblasts, p44/42
mitogen-activated protein kinase (MAPK, also known as ERK1/2 or
ERK) monoclonal (cat. no. 4695) and phosphorylated ERK
(Thr202/Tyr204; p-ERK; cat. no. 4377; both Cell Signaling
Technology, Inc., Danvers, MA, USA) antibodies were used.
Anti-GAPDH antibodies were purchased from Abcam (Cambridge, MA,
USA; cat. no. ab9485). U0126, a dual specificity mitogen-activated
protein kinase kinase 1 (MEK)/ERK inhibitor, was purchased from
Cell Signaling Technology, Inc.
Isolation and culture of human colon
fibroblasts
The authors of the present study established
fibroblast cell lines from a specimen resected from a 73-year-old
Japanese female patient with well-differentiated colon cancer at
Department of Gastroenterological Surgery, Nagoya City University
(Nagoya, Japan) in January 2016. The technical procedure was
described in a previous report (22) and was performed as described
previously (10). Following
receiving written informed consent, tissues were retrieved from two
separate regions: Colon carcinoma tissue and non-malignant colon
tissue. To avoid contamination, these tissues were collected from
the serosal side, taking special care not to pierce the mucosa.
Following fragmenting with scissors, the tissue was incubated in
Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich; Merck
KGaA) containing 1,000 U/ml Dispase® (Godo Shusei Co.,
Ltd., Tokyo, Japan) for 2 h. Then, the fragments were cultivated in
DMEM containing 5% fetal bovine serum (FBS; Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) and 1%
antibiotic-antimycotic solution, and incubated at 37°C in air
containing 5% CO2. To confirm that the cultivated cells
were fibroblasts, the cells were examined using immunofluorescence
to determine whether they expressed α-SMA, IL-6 and VEGF.
Fibroblasts isolated from malignant regions or benign regions were
termed as CAFs or normal fibroblasts (NFs), respectively.
Fibroblasts in the third to sixth passage were used in subsequent
experiments. Patient 1 was a 42-year-old Japanese male patient with
well-differentiated colon cancer. Patient 2 was a 74-year-old
Japanese male patient with poorly-differentiated colon cancer. The
protocols used for sample collection and fibroblast examinations of
Patient 1 and 2 were the same as described above.
Immunofluorescence staining
Immunofluorescence staining of the cultured cells
was performed according to a method described previously (10). CAFs and NFs were grown in chamber
slides, fixed with 10% formalin for 10 min at room temperature,
treated with 0.2% Triton X-100 for 10 min at room temperature, and
blocked with 1% bovine serum albumin (BSA; cat. no. WDH4409; Wako,
Osaka, Japan) in PBS for 1 h at room temperature. Then, the slides
were treated with anti-α-SMA (1:100) or anti-IL-6 (1:1,000) mouse
monoclonal antibodies for 2 h at room temperature, and then with
Alexa Fluor 488®-conjugated donkey anti-mouse
immunoglobulin (Ig)G secondary antibodies (1:200; cat. no. ab15019;
Invitrogen; Thermo Fisher Scientific, Inc.) for 30 min at room
temperature. The membranes were also incubated with anti-VEGF
rabbit monoclonal antibodies (1:500) for 2 h at room temperature
and then with Alexa Fluor 488®-conjugated goat
anti-rabbit IgG secondary antibodies (cat. no. ab150077; Abcam) for
1 h at room temperature. These slides were mounted with ProLong
Gold Antifade Reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
and observed under a BZ-X700 fluorescence microscope (Keyence
Corporation, Osaka, Japan) equipped with a charge-coupled device
camera. The magnification was ×200.
Immunostaining
To confirm the phenotypic characterization of CAFs
and NFs, anti-vimentin (V9), anti-cytokeratin (AE1/AE3) and
anti-CD90 (5E10) mouse monoclonal antibodies were used as described
by a previous study by our group (10). Briefly, CAFs and NFs were fixed with
10% formalin for 10 min at room temperature to cross-link the
proteins, then blocked with 3% BSA in PBS for 1 h at room
temperature. Resin was not used for this protocol. Then, the
primary antibodies (vimentin, 1:80; CD90, 1:100; cytokeratin, 1:80)
were applied for 60 min at room temperature. Horseradish peroxidase
(HRP)-conjugated secondary antibodies (cat. no. K4001; Dako;
Agilent Technologies, Inc.) was applied for 60 min at room
temperature. These slides were observed under a BX51 light
microscope (Olympus Corporation, Tokyo, Japan) at a magnification
of ×100.
Cell lines
Human umbilical vein endothelial cells (HUVECs) were
purchased from PromoCell GmbH (Heidelberg, Germany). They were
cultivated in Endothelial Cell Growth Medium (EGM) kit (PromoCell
GmbH) and incubated at 37°C in air containing 5% CO2.
EGM contained 2% fetal calf serum, 0.4% endothelial cell growth
supplement, epidermal growth factor (0.1 ng/ml), basic fibroblast
growth factor (1 ng/ml), heparin (90 µg/ml) and hydrocortisone (1
µg/ml; all PromoCell GmbH).
Cell viability assay
The viability assay was performed using Premix WST-1
Cell Viability Assay System (Takara Bio, Inc., Otsu, Japan)
according to the manufacturer's protocol. Each assay was conducted
at least three times (n=5). Fibroblasts (3×103/well) and
HUVECs (5×103/well) were seeded into 96-well plates and
incubated overnight. Next, the cells were treated with various
concentrations (0, 10, 30, 50, 70 and 90 µM) of EPA in serum-free
DMEM and incubated at 37°C in air containing 5% CO2.
After 48 h, the medium was removed and replaced with fresh medium
(90 µl/well) containing Premix WST-1 (10 µl/well). The cells were
then incubated at 37°C for 2 h. Absorbance was measured at 450 nm
using a SpectraMax 340 spectrophotometer (Molecular Devices, LLC,
Sunnyvale, CA, USA). The control cells were untreated.
Furthermore, WST-1 assays were conducted on the
fibroblasts after 0, 24,48 and 72 h following the incubated with 0,
10 or 30 µM EPA. Absorbance was measured at 450 nm. The absorbance
values of each EPA concentration was compared with the value at 0
h.
ELISA
Fibroblasts were seeded into 6-well plates at a
density of 3×104 cells/ml with 5% FBS in DMEM. Following
overnight incubation, the fibroblasts were divided into four groups
and cultured in serum-free DMEM; the groups were as follows: i)
Untreated, ii) 30 µM EPA, iii) 50 µg/ml LPS, and iv) 50 µg/ml LPS
and 30 µM EPA (9.7 µg/ml) Additionally, CAFs were divided into
several groups and cultured in serum-free DMEM; the groups were as
follows: i) Untreated, ii) 50 µg/ml LPS, iii) 50 µg/ml LPS and 10
µg/ml MRA (Actemra®; Chugai Pharmaceutical Co., Ltd.,
Tokyo, Japan), iv) LPS (0, 0.1, 1, 10 or 100 µg/ml) with or without
30 µM EPA, v) IL-6 (0, 1 or 10 µg/ml; cat. no. 550071; BD
Pharmingen; Franklin Lakes, NJ, USA) with or without 30 µM EPA, vi)
30 µM EPA, and vii) 10 µM U0126 (3.8 µg/ml). The untreated cells
were incubated with concentrated 0.03% ethanol in serum-free DMEM
and were used as the controls. After 48 h, the culture supernatants
were collected and centrifuged (12 × g, 4°C, 3 min) to pellet any
detached cells. Their VEGF and IL-6 expression levels were
quantified from the supernatants using VEGF (cat. no. DVE00) and
IL-6 (cat. no. D6050) ELISA kits (both R&D Systems Inc.,
Minneapolis, MN, USA), respectively. The ELISAs were performed
according to the manufacturer's protocol. All experiments were
performed three times in triplicate. A lot of the in vivo
cells possessed so few IL-6 receptors on their membranes; therefore
the authors of the present study incubated these cells with soluble
receptors. IL-6 was supplemented with soluble 40 ng/ml IL-6
receptor (cat. no. 200-26R; Human recombinant soluble IL-6 receptor
was purchased from Peprotech; Rocky Hill, NJ, USA).
Western blotting
CAFs (1.0×105/ml) were seeded into 10-cm
dishes with 5% FBS in DMEM and incubated at 37°C in a
CO2. incubator until fibroblast growth was
semi-confluent. After the cells were cultured in serum-free DMEM
for 3 h for starvation, the medium was changed. The cells were
cultured for an additional 3 h without treatment or in the presence
of EPA (10 or 30 µM) or 10 µM U0126. Untreated cells were incubated
with concentrated 0.03% ethanol in serum-free DMEM and were used as
the controls.
Protein samples were extracted from CAFs using
radioimmunoprecipitation lysis buffer with Protease Inhibitor
Single Use Cocktail and Phosphatase Inhibitor Cocktail (all Thermo
Fisher Scientific, Inc.), as described previously (23). Protein concentrations were measured
using a BCA protein assay kit (Thermo Fisher Scientific, Inc.).
Equal amounts of the protein extract were denatured by boiling at
90°C for 5 min. Proteins (20 µg/lane) were fractionated on 4–15 %
Mini-PROTEAN TGX gels and the protein bands were transferred onto
nitrocellulose membranes (both Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The primary and secondary antibody reactions
were performed using an iBind Flex Western System (Thermo Fisher
Scientific, Inc.). The membranes were incubated with iBind Flex
Solution (iBind Flex Buffer, iBind Flex Additive and distilled
water) for 10 min at room temperature to block nonspecific binding.
The membranes were incubated with anti-ERK, -p-ERK (both 1:1,000)
and -GAPDH (1:2,500) primary antibodies, and then HRP-conjugated
goat anti-rabbit polyclonal secondary antibodies (1:2,000; cat. no.
P0448; Dako; Agilent Technologies, Inc.). These primary antibodies
and secondary antibodies reactions were performed at room
temperature for 2.5 h. Protein-antibody complexes were visualized
using SuperSignal West Pico Chemiluminescent Substrate or
SuperSignal West Femto Chemiluminescent Substrate (Thermo Fisher
Scientific, Inc.). The immunoreactive protein bands were detected
and the band densities were quantified by densitometry using an
Amersham Imager 600 with Amersham Imager 600 analysis software
(version 1.2; GE Healthcare Life Sciences, Little Chalfont, UK).
Experiments were performed at least three times.
Angiogenic effects of EPA
To study the angiogenic effect of EPA, the
capillary-like structure formation (CSF) of the HUVECs was
evaluated using the angiogenesis assay on Matrigel (BD Biosciences)
with or 30 µM without EPA. For the reconstitution of a basement
membrane matrix, Matrigel was diluted to 50% concentration with
cold serum-free DMEM and added to a 24-well tissue culture plate
(250 µl/well) at 4°C. The 24-well plate was incubated for >2 h
at 37°C to allow the Matrigel to solidify, as described previously
(24).
HUVECs were trypsinized, counted and resuspended in
EGM, and added to the surface of the reconstructed basement
membrane (5×104 cells/well). The medium of cultured CAFs
were removed and termed conditioned medium (CM), which was diluted
to 50% concentration with EGM. HUVECs were divided into three
groups and cultured for 16 h to allow the CSF to take place; the
groups were as follows: i) Serum-free DMEM and untreated, ii)
serum-free DMEM and 30 µM EPA, iii) serum-free DMEM and 500 pg/ml
VEGF (Recombinant VEGF165; cat. no. 890220; R&D Systems, Inc.),
iv) CM and untreated, ii) CM and 30 µM EPA, iii) CM, 30 µM EPA and
500 pg/ml VEGF. These endotubes were quantified by counting nine
fields (three regions per plate) at a magnification of ×40, with
each condition being assessed in triplicate. These were observed
with inverted phase contrast fluorescence microscopy using a
BZ-X700 fluorescence microscope. Untreated cells were incubated
with concentrated 0.03% ethanol in serum-free DMEM and were used as
the controls.
Statistical analysis
All statistical analyses were performed using EZR
software (Easy R) version 1.27 (Saitama Medical Center, Jichi
Medical University, Saitama, Japan). Data are presented as mean ±
standard error of the mean. Differences between groups were
compared by Student's t-test or a one-way analysis of variance
followed by Tukey's test. P<0.05 indicated that the difference
between groups was statistically significant.
Results
IL-6, VEGF and α-SMA expression is
elevated in isolated CAFs
Cells with large cytoplasm and spindle-shaped cells
were observed in samples from normal and cancerous colon stroma.
The immunofluorescence staining revealed that the two cell types
from colon stroma (CAFs and NFs) secreted α-SMA, IL-6 and VEGF
(Fig. 1A). In addition,
immunostaining demonstrated that CAFs and NFs were positive for
expression of CD90 and vimentin, and negative for cytokeratin. Due
to the high expression α-SMA and IL-6, the cells from human colon
stroma were regarded as fibroblasts; as the cells from the colon
cancer stroma exhibited a high expression of α-SMA, an indicator of
CAFs (10), they were clarified as
CAFs. It should be noted that CAFs have properties of
myofibroblasts and express α-SMA (25). The expression level of IL-6 was
higher in CAFs compared with that of NFs, whereas the expression of
VEGF and α-SMA in NFs and CAFs was almost the same. IL-6 and VEGF
secretions from CAFs and NFs were also evaluated using ELISA
(Fig. 1B). The expression of IL-6
(P<0.001) and VEGF (P=0.001) cultured in 5% FBS in DMEM was
significantly higher in CAFs compared with NFs; IL-6 and VEGF
expression was 5.0- and 1.5-fold higher, respectively.
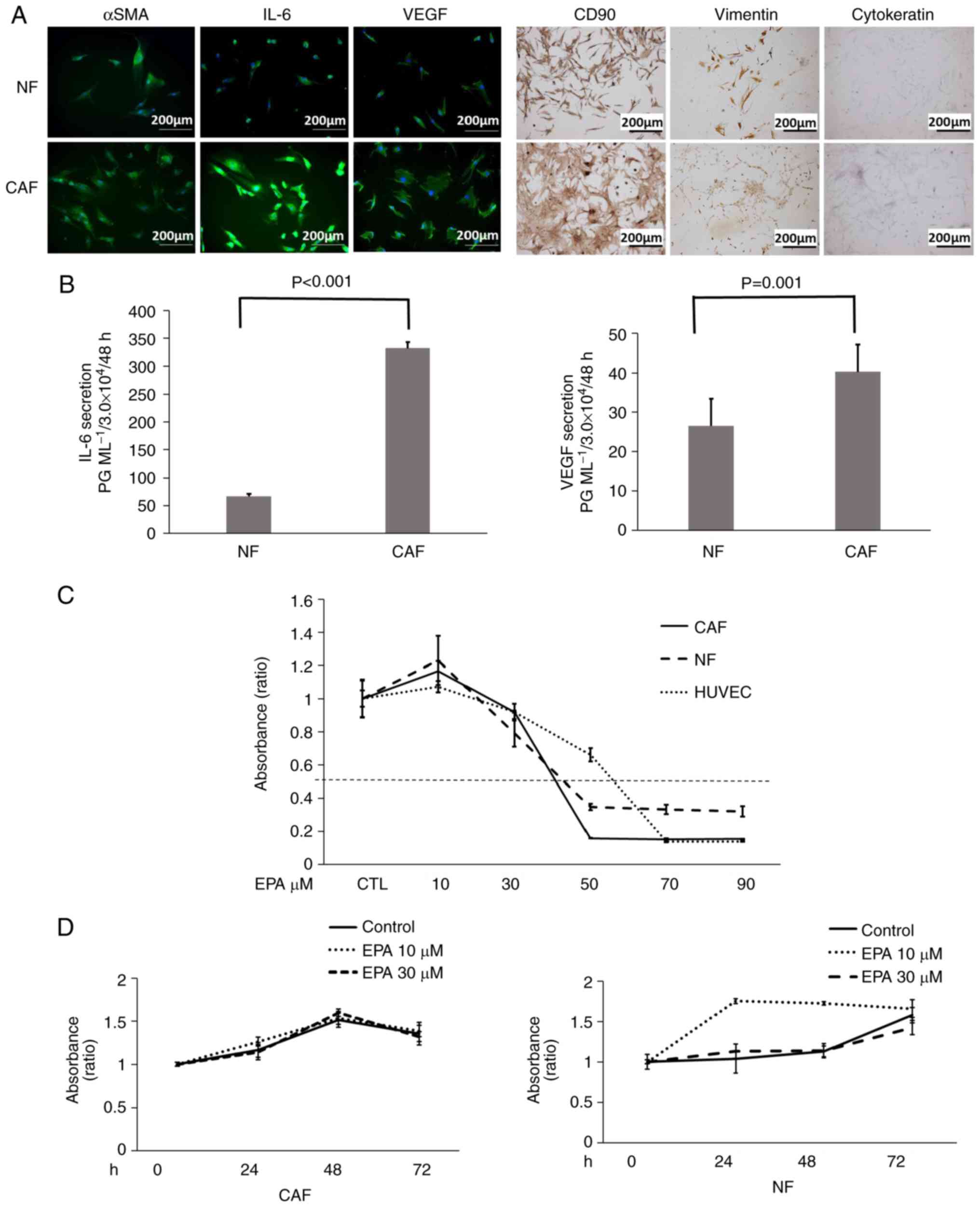 | Figure 1.IL-6, VEGF and α-SMA expression is
elevated in isolated CAFs, and NFs, CAFs and HUVECs remain viable
with 30 µM EPA treatment. (A) Immunofluorescence staining of
proteins in NFs and CAFs. (B) Concentration of IL-6 and VEGF
secreted in NFs and CAFs, analyzed by ELISA. (C) The WST-1 assay
was used to assess cell viability in NFs, CAFs and HUVECs following
EPA treatment. The WST-1 assay was used to assess cell viability in
(D) CAFs and (D) NFs with and without EPA treatment. Data are
representative of three independent experiments and are presented
as mean ± standard error of the mean. IL, interleukin; VEGF,
vascular endothelial growth factor; SMA, smooth muscle actin; CD,
cluster of differentiation; CAF, cancer-associated fibroblast; NF,
normal fibroblast; HUVEC, human umbilical vein endothelial cell;
EPA, eicosapentaenoic acid. |
Human colon fibroblasts and HUVECs
remain viable with 30 µM EPA treatment
The WST-1 assay demonstrated that EPA suppressed the
viability of CAFs, NFs and HUVECs in what appeared to be a
concentration-dependent manner (Fig
1C). At >50 µM EPA, fibroblast viability reached a plateau.
However, the viability of CAFs, NFs and HUVECs were not suppressed
by treatment with <30 µM EPA, with ~80% viability observed
compared with the control samples. The viability of CAFs, NFs and
HUVECs after treatment with 30 µM EPA was 95.1, 79.2 and 92.1%,
respectively; no significant difference in the viability of these
cells was identified compared with their respective controls.
Furthermore, no significant decrease in the viability of CAFs and
NFs was identified when the cells were incubated with 30 µM EPA for
24, 48 or 72 h (Fig. 1D). In NFs,
it is still unknown why the viability increased with 10 µM EPA.
Thus, in the present study, the authors used EPA at a concentration
of 30 µM to evaluate the suppression of VEGF and IL-6 secretion
without viability suppression.
EPA reduces VEGF and IL-6 secretion in
NFs and CAFs with or without LPS-stimulation, and reduces
IL-6-induced VEGF secretion in CAFs
ELISA examination demonstrated that IL-6 secretion
from CAFs was high without stimulation; IL-6 secretion from NFs was
markedly lower. IL-6 secretion from CAFs and NFs increased markedly
following LPS stimulation. EPA significantly reduced the
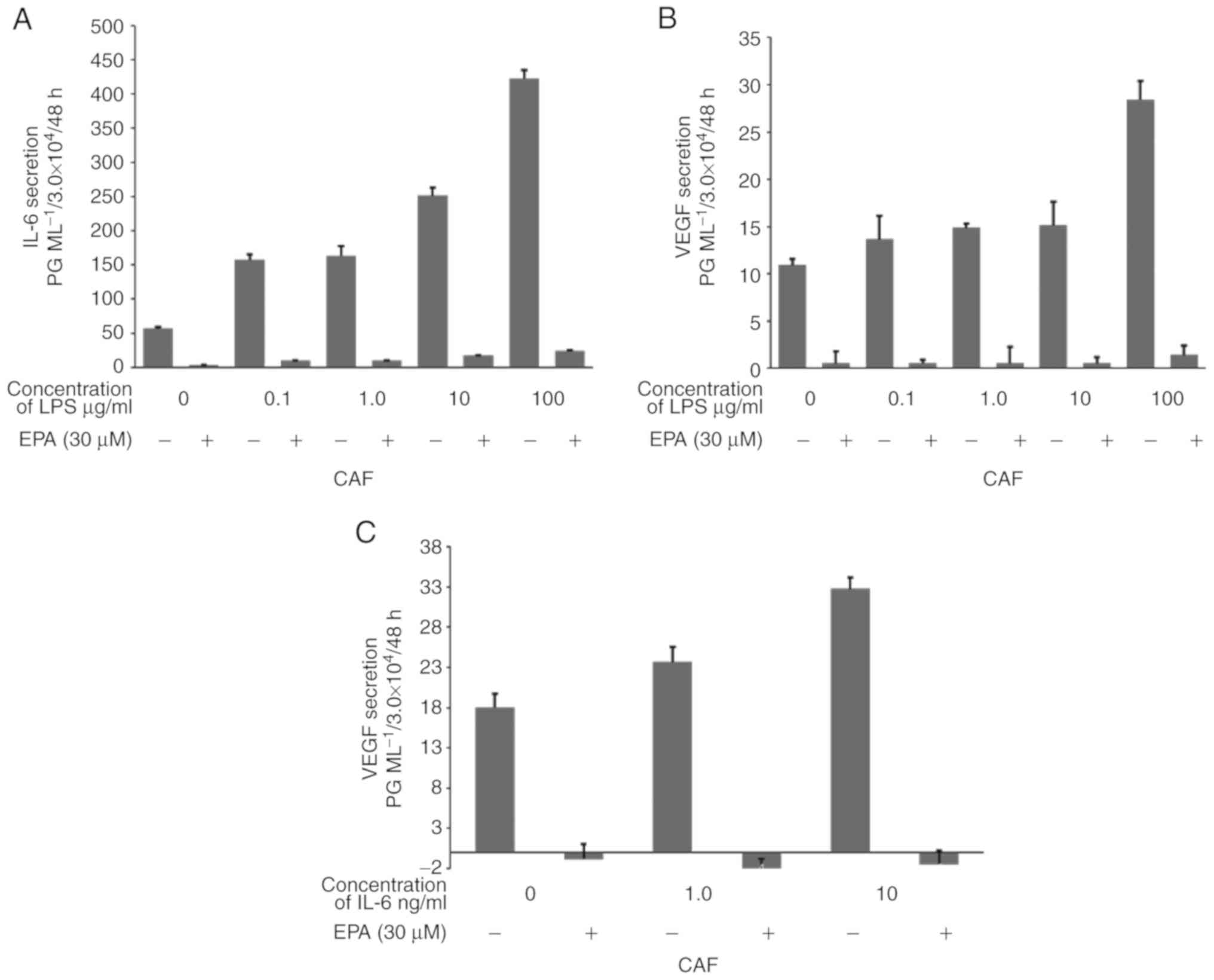
IL-6-induced increase in secretion in CAFs and NFs (Fig 2A and B). As presented in Fig 2C and D, LPS (50 µg/ml) also increased
the VEGF secretion from the two types of human colon fibroblasts.
CAFs and NFs secreted higher levels of IL-6 (CAFs, 476.5 pg/ml;
NFs, 260.7 pg/ml) and VEGF (54.1 and 60.2 pg/ml) with LPS
stimulation compared with the control cells that did not have LPS
stimulation (IL-6: CAF, 276.3 pg/ml; NF, 15.2 pg/ml; VEGF: CAF,
19.5 pg/ml; NF, 16.0 pg/ml; Fig.
2A-D). Additionally, EPA (30 µM) significantly suppressed the
secretion of IL-6 (CAF, 30.9 pg/ml; NF, 8.2 pg/ml) and VEGF (CAF, 0
pg/ml; NF, 0 pg/ml; all P<0.01) compared with the control group;
additionally, EPA (30 µM) significantly suppressed the secretion of
both cytokines despite LPS stimulation (IL-6: CAF, 171.4 pg/ml; NF,
57.2 pg/ml; VEGF: CAF, 0 pg/ml; NF, 0 pg/ml; all P<0.001) when
compared with the LPS stimulated cells. While EPA suppressed VEGF
secretion completely even after LPS stimulation (Fig. 2C and D), it did not completely
suppress IL-6 secretion even without LPS stimulation (Fig. 2A and B). On the other hand, MRA, an
anti-IL-6 receptor antibody, suppressed the LPS-induced promotion
of VEGF secretion. However, the VEGF secretion level from CAFs
following LPS stimulation and treatment with 10 µg/ml MRA was not
at the same level as that of the control, and no significant
difference was identified (Fig.
2E). Whereas MRA significantly suppressed VEGF secretion from
NFs to a level that was almost the same as that in the control
(P=0.023; Fig. 2F).
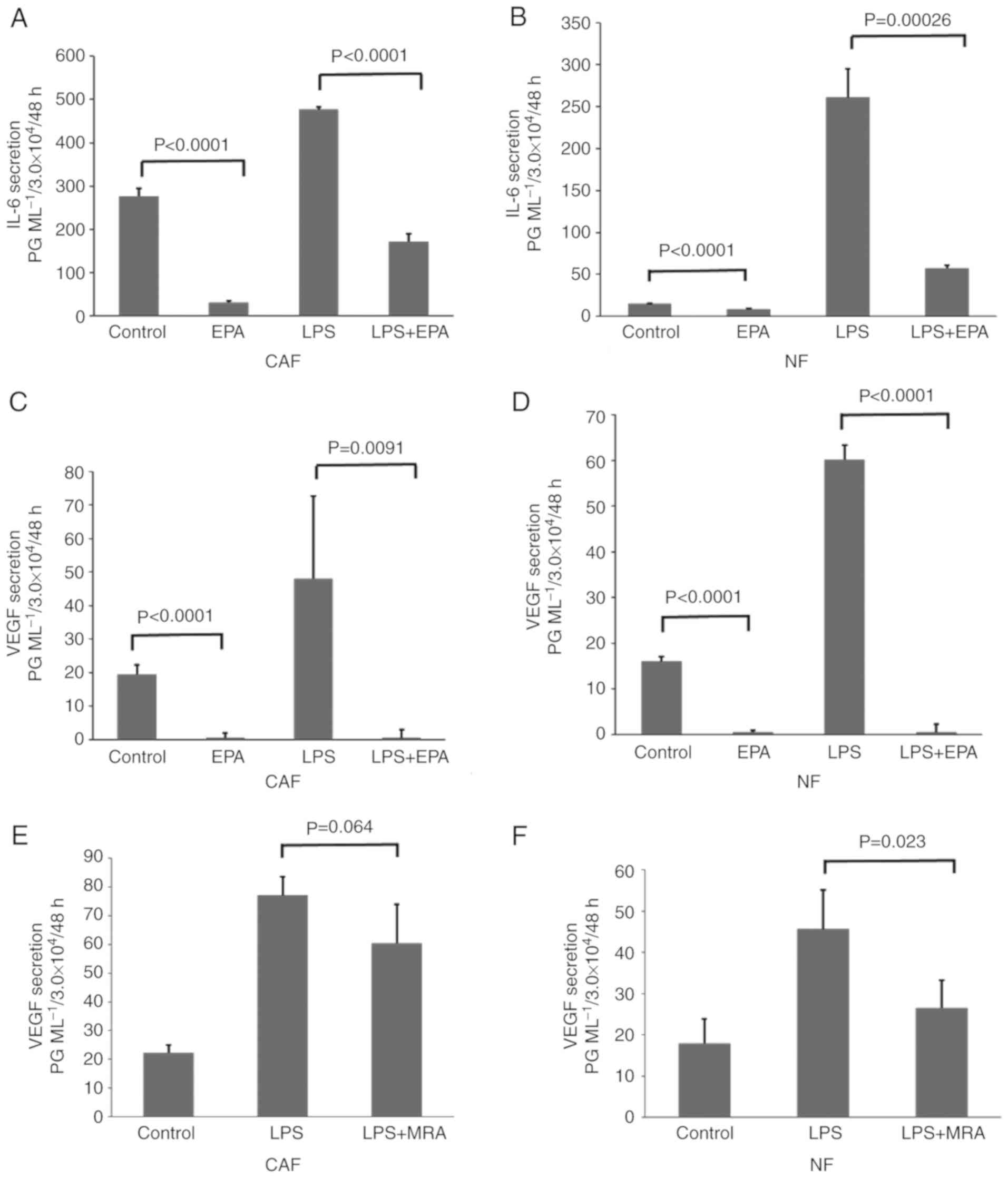 | Figure 2.EPA reduces VEGF and IL-6 secretion
in NFs and CAFs with or with LPS-stimulation. IL-6 and VEGF protein
concentrations were detected by ELISA. (A-D) CAFs and NFs were
either untreated, or treated with 30 µM EPA, 50 µg/ml LPS or LPS
and EPA. The concentration of IL-6 secreted from (A) CAFs and (B)
NFs. The concentration of VEGF secreted from (C) CAFs and (D) NFs.
(E and F) CAFs and NFs were either untreated, or treated with 50
µg/ml LPS or LPS and 10 µg/ml MRA. The concentration of VEGF
secreted from (E) CAFs and (F) NFs. Data are representative of
three independent experiments and are presented as mean ± standard
error of the mean. IL, interleukin; VEGF, vascular endothelial
growth factor; LPS, lipopolysaccharide; CAF, cancer-associated
fibroblast; NF, normal fibroblast; EPA, eicosapentaenoic acid; MRA,
anti-IL-6 myeloma receptor antibodies. |
Then, to evaluate the association between these
secretion levels and inflammation, IL-6 and VEGF secretion levels
from CAFs treated with various concentrations of LPS with or
without EPA treatment were quantified using ELISA. While LPS
promoted IL-6 and VEGF secretion from CAFs in what appeared to be a
dose-dependent manner, the addition of 30 µM EPA suppressed these
secretions completely, even with increasing LPS concentrations
(Fig. 3A and B). Furthermore, the
administration of IL-6 to CAFs promoted VEGF secretion in what
appeared to be a dose-dependent manner, while the addition of 30 µM
EPA suppressed VEGF secretion even with the addition of 10 ng/ml of
IL-6 (Fig. 3C).
EPA (30 µM) and 10 µM U0126 reduce
p-ERK expression, and IL-6 and VEGF secretion from CAFs
The influence of EPA on VEGF administration in CAFs,
in terms of ERK phosphorylation in fibroblast cells, was examined
by western blotting (Fig. 4A). ERK
phosphorylation, and IL-6 and VEGF secretion from CAFs without EPA
treatment were compared with those from CAFs treated with EPA or
U0126, a MEK inhibitor. EPA inhibited p-ERK expression in what
appeared to be a dose-dependent manner (Fig. 4A). Although no significant
difference was identified following 10 µM EPA administration, 30 µM
EPA significantly inhibited the p-ERK/GAPDH expression ratio
compared with the control group (P=0.0012; Fig. 4B). U0126 also significantly
decreased p-ERK expression compared with the control group
(P<0.0001). In contrast, EPA and U0126 treatment did not
significantly affect the expression of total ERK (Fig. 4C). The expression ratio of pERK/ERK
was inhibited with U0126 (P=0.0074) and 30 µM EPA (P=0.028;
Fig. 4D). Additionally, IL-6 and
VEGF secretion from CAFs was significantly inhibited by EPA and
U0126 compared with the control group (both P<0.002; Fig. 4E and F).
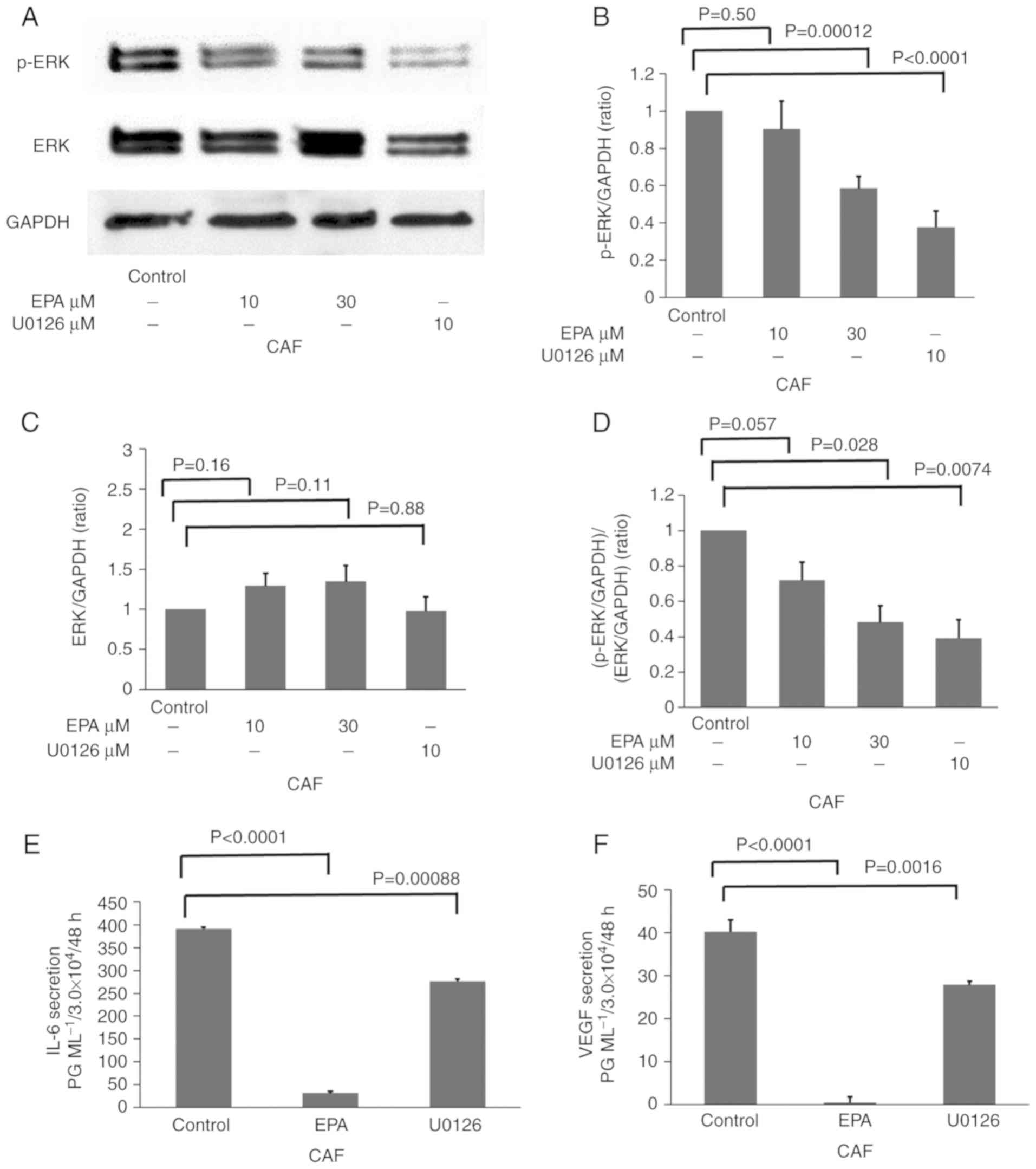 | Figure 4.EPA (30 µM) and 10 µM U0126 reduce
p-ERK expression, and IL-6 and VEGF secretion from CAFs. CAFs were
either untreated, or treated with 10 µM EPA, 30 µM EPA or 10 µM
U0126, a dual specificity mitogen-activated protein kinase kinase
1/ERK inhibitor. (A) ERK and p-ERK signals were evaluated by
western blotting. The densitometric analysis of (B) p-ERK, (C) ERK
and (D) p-ERK/ERK as ratios of GAPDH. The concentration of (E) IL-6
and (F) VEGF secreted from CAFs was analyzed by ELISA. Data are
representative of three independent experiments and are presented
as mean ± standard error of the mean. ERK, mitogen-activated
protein kinase; p-, phosphorylated; IL, interleukin; VEGF, vascular
endothelial growth factor; CAF, cancer-associated fibroblast; EPA,
eicosapentaenoic acid. |
EPA reduces CSF in HUVECs cultured in
CM and serum-free DMEM via the inhibition of VEGF secretion
The angiogenesis assay on Matrigel used to study CSF
by HUVECs (Fig. 5) reflects
angiogenesis in vivo. CSF by HUVECs was observed in the
control group (Fig. 5A). EPA
suppressed CSF in HUVECs cultured in serum-free DMEM (Fig. 5B). VEGF, which is an angiogenic
factor (26), markedly promoted CSF
in HUVECs at a concentration of 500 pg/ml (Fig. 5C). The supernatant obtained from a
CAF culture also markedly promoted CSF in HUVECs (Fig. 5D). EPA also suppressed CSF in HUVECs
cultured with the CAF supernatant (Fig.
5E). To evaluate whether this CSF suppression by EPA is caused
by its direct effect on HUVECs or by VEGF suppression, 500 pg/ml
VEGF was added to the HUVECs cultured in the CAF supernatant with
EPA (Fig. 5F). The count of tube
formation was evaluated in Fig. 5G.
The results demonstrated that EPA and VEGF significantly increased
CSF in HUVECs compared with the control cells, and that EPA, and
EPA and VEGF combined significantly increased CSF in HUVECs treated
with CM. These results indicated that CSF decreased due to EPA
administration may be caused by the inhibition of VEGF secretion
and not by the direct inhibition of HUVEC growth.
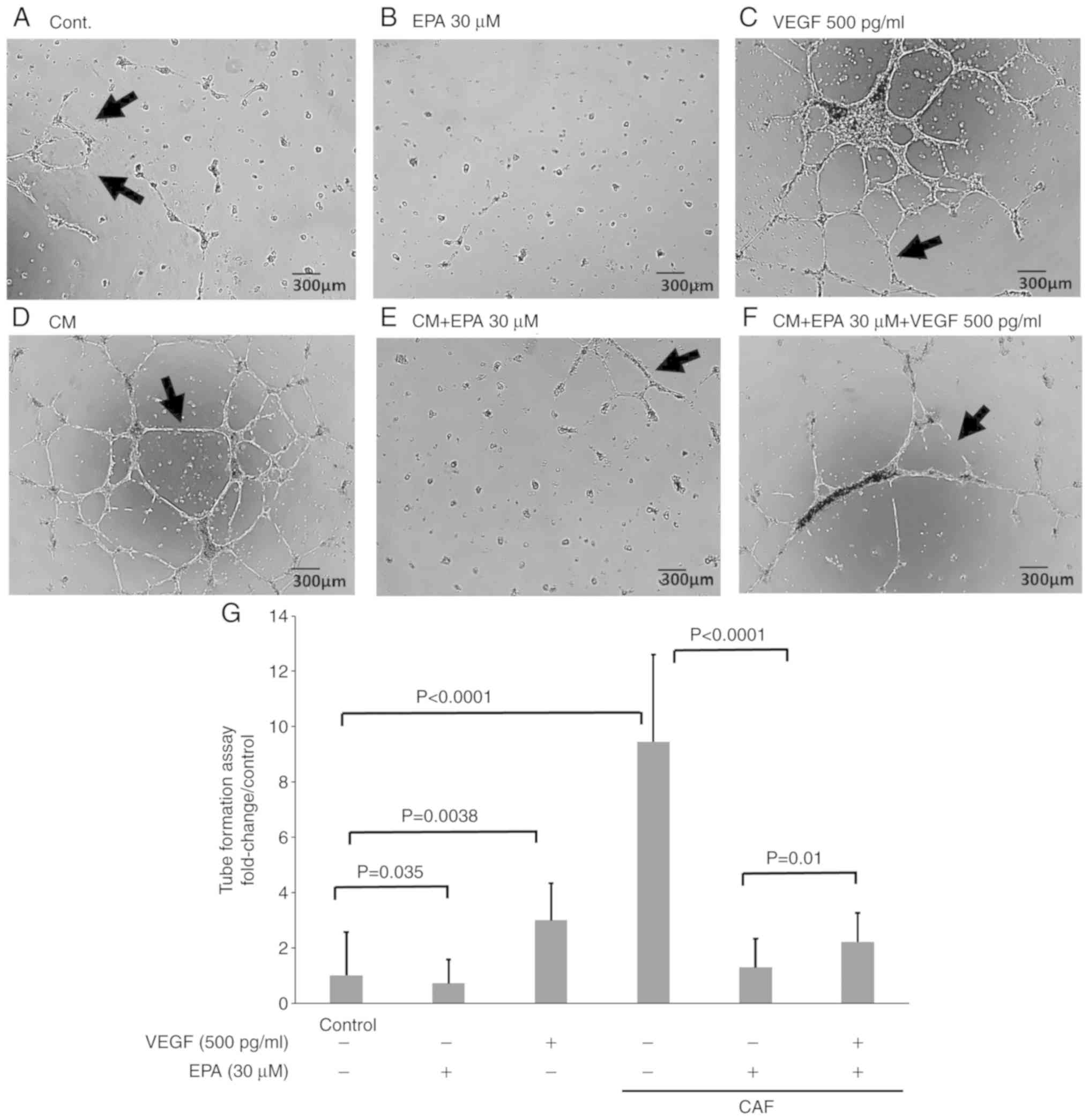 | Figure 5.EPA reduces CSF in HUVECs cultured in
CM and serum-free DMEM via the inhibition of VEGF secretion. The
angiogenesis assay on Matrigel was conducted on HUVECs. CSF (arrow)
was counted in nine fields. HUVECs were cultured in (A) serum-free
DMEM alone, (B) serum-free DMEM with 30 µM EPA, (C) serum-free DMEM
with 500 pg/ml VEGF, (D) CM alone, (E) CM with 30 µM EPA, and (F)
CM with 30 µM EPA and 500 pg/ml VEGF. (G) Quantification of CSF.
Data are representative of three independent experiments and are
presented as mean ± standard error of the mean. HUVEC, human
umbilical vein endothelial cell; CSF, capillary-like structure
formation; VEGF, vascular endothelial growth factor; DMEM,
Dulbecco's modified Eagle's medium; CM, supernatant from
cancer-associated fibroblasts; EPA, eicosapentaenoic acid. |
EPA decreases VEGF secretion from CAFs
by suppressing ERK phosphorylation
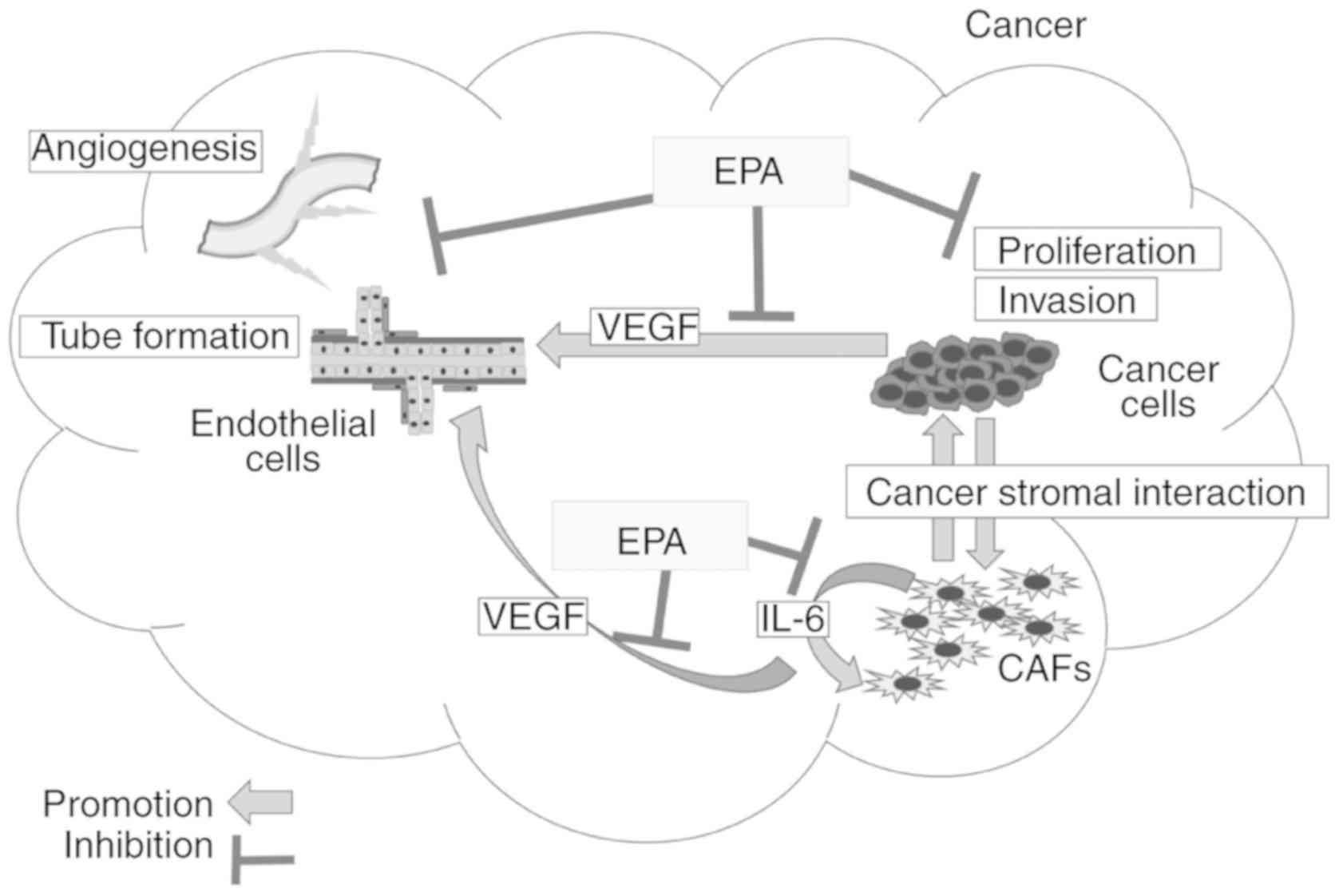
From the results in Figs. 1–5,
it is clarified that EPA suppresses the secretion of IL-6 and VEGF
from CAFs, which causes angiogenesis in cancer (Fig 6). Previous studies revealed that the
anticancer mechanisms of EPA are as follows: i) Reduction of IL-6
secretion from cancer cells causes the suppression of tumor
proliferation and invasion (11,16,27),
which may be caused by the suppression of ERK phosphorylation in
cancer cells (14,20,28,29),
and ii) anti-angiogenesis effects are caused by the direct
suppression of endothelial cell proliferation mediated by
cyclooxygenases (30). The results
of the present study demonstrated novel anti-angiogenesis
mechanisms of EPA, involving the decrease of VEGF secretion from
CAFs by the suppression of ERK phosphorylation.
CAFs from other colorectal cancer
patients remain viable with 30 µM EPA treatment, and EPA reduces
VEGF and IL-6 secretion in CAFs from other colorectal cancer
patients with or without LPS-stimulation
Examinations of CAFs from other colorectal cancer
patients were also performed (Fig.
S1). Patient 1 was a 42-year-old Japanese male patient with
well-differentiated colon cancer. Patient 2 was a 74-year-old
Japanese male patient with poorly-differentiated colon cancer. No
significant differences in the cell viability of all fibroblasts
were identified in patients treated with or without 30 µM EPA
(Fig. S1A and B). LPS (50 µg/ml)
stimulation increased IL-6 secretion from NFs and CAFs retrieved
from the patients, and EPA (30 µM) suppressed the secretion of IL-6
whether LPS stimulation occurred or not (Fig. S1C and D). LPS also increased VEGF
secretion from CAFs, while EPA suppressed the secretion of VEGF in
CAFs with and without LPS stimulation (Fig. S1E and F).
Discussion
The present study has revealed a great deal of the
mechanism behind the anticancer effects of EPA. One effect of EPA
is anti-inflammatory via the suppression of IL-6 secretion. This
may be caused by the inhibition of the ERK signal by EPA. IL-6
causes various types of tumor progression, including tumor invasion
and tumor proliferation (1). Greten
et al (31) and Grivennikov
et al (32) reported that
IL-6 also causes tumor angiogenesis via nuclear factor NF-κB.
Furthermore, IL-6 has been demonstrated to cause systemic
metabolism, which causes cachexia in patients with advanced cancer
(13). Thus, it was hypothesized
that the inhibition of IL-6 secretion by EPA suppresses tumor
progression via several mechanisms (33). For a long time, it was been
hypothesized that this mechanism is affected by IL-6 secreted from
cancer cells (11). However, a
previous study by our group demonstrated that more IL-6 is released
from the cancer stroma compared with cancer cells (10). Thus, it remains necessary to clarify
whether EPA decreases IL-6 secretion from cancer stromal cells to a
level that is same as that from cancer cells.
Another anticancer effect of EPA is its
anti-angiogenic effect, which occurs via the direct suppression of
endothelial cell growth and tubular formation (30). Szymczak et al (30) reported that EPA modulates
angiogenesis via cyclooxygenases. The results of the present study
demonstrated that EPA addition suppressed CSF in HUVECs in
vitro. The authors of the present study hypothesize that this
is mechanism may also cause an anticancer effect.
In the present study, the authors demonstrated a
novel anticancer mechanism of EPA, which involves an
anti-angiogenic effect associated with CSI caused by CAFs and
cancer cells. This is caused by the EPA-mediated suppression of
VEGF secretion from CAFs. CAFs secreted more IL-6 compared with
NFs. Furthermore, LPS stimulation increased IL-6 secretion from
CAFs. Additionally, IL-6 increased VEGF secretion from CAFs in what
appeared to be a dose-dependent manner. This indicates that
inflammation caused by bacterial infection or cancer itself may
promote IL-6 secretion from CAFs, which may promote the autocrine
secretion of VEGF from CAFs. In contrast, the present study
determined that EPA entirely decreased VEGF secretion from
CAFs.
Nevertheless, from the results of the present study,
two hypotheses remain for the decrease of VEGF secretion from CAFs.
One involves the inhibition of IL-6 secretion from CAFs by EPA,
which results in the suppression of VEGF secretion from CAFs, and
the other states that EPA suppresses VEGF secretion from CAFs
directly. EPA suppressed VEGF secretion from CAFs completely
despite the increasing levels of IL-6. The results of the present
study do not support the theory that EPA decreases the VEGF
secretion from CAFs indirectly by decreasing IL-6 secretion from
CAFs. The authors of the present study concluded that EPA affects
the VEGF secretion mechanism of CAFs directly by inhibiting ERK
phosphorylation. The western blotting examination of the present
study revealed that ERK phosphorylation reduced in CAFs treated
with EPA or U0126, a MEK/ERK inhibitor. It was also revealed that
CAFs treated with EPA or U0126 exhibited decreased IL-6 and VEGF
secretion. These results demonstrated that the mechanism elucidated
in the present study is similar to the mechanism revealed in
another study, which demonstrated that EPA decreased IL-6 secretion
from cancer cells (15). It was
reported that, in colon cancer cells, ERK activation is a key step
in the upregulation of VEGF induced by serum starvation (34). In addition, it was also reported
that EPA was able to markedly inhibit ERK-1 and −2 phosphorylation
(14,20,28,29).
The origin of CAFs has been discussed for a long
time. Several studies demonstrated that insisted stromal
fibroblasts transform into CAFs (1,35);
however, adipocytes, epithelial cells (through
epithelial-mesenchymal transition), endothelial cells (through
endothelial-mesenchymal transition), bone marrow-derived
mesenchymal stem cells and hematopoietic stem cells are also
considered as origins of CAFs (1,36–38).
The present study revealed that similar fibroblasts obtained from
different regions of the colon had different characters. The
present study demonstrated that while it was necessary to stimulate
fibroblasts from the normal colon region to secrete IL-6, those
from the cancer colon secreted IL-6 without any stimulation.
Furthermore, the present study revealed that inhibiting IL-6 could
not suppress VEGF secretion completely in CAFs, contrary to the
case in NFs. One possible reason for this is that there are many
cytokines other than IL-6 released from CAFs without any
stimulation (1); these may also
influence VEGF secretion. In the present study, the authors
succeeded in demonstrating the anti-angiogenic effect of EPA is
associated with CAFs; however, there are a number of EPA mechanisms
that may explain the suppression of IL-6 secretion from CAFs that
remain confounding.
Despite these previous reports that demonstrated the
anticancer effects of EPA (11,14–21),
it has not been used as an anticancer drug, but just as an
anti-coagulant (for example ethyl icosapentate). However, there
have not been any studies that investigated cancer stroma, CSI and
the utility of EPA in cancer therapy. Cancer tissues consist of
cancer cells and stromal cells, and as much as 60–90 % of the mass
of colon cancers is composed of stromal cells, including
fibroblasts, vascular endothelial cells and immune cells (1,39).
Actually, stromal fibroblasts are a major component of tumors
(40); they have been termed CAFs
and they have specific effects on tumor growth (22,41,42).
Thus, inhibiting CSI will serve a more important role in cancer
therapy compared with solely inhibiting cancer cell activity. The
present study demonstrated the effects of EPA on stromal cells in
terms of IL-6 and VEGF secretion. From these results, VEGF
suppression seems to cause many anticancer effects other than
anti-angiogenic effects.
CAFs have not been developed as a cell line for
widespread use. Thus, a great number of studies have used cells
cultured in the laboratory (43,44).
While this may not appear to give reproducible results, the authors
of the present study confirmed that the same results were obtained
using cells derived from other cancer patients. Thus, this method
is universal.
In conclusion, the present study determined that EPA
inhibited angiogenesis caused by CAFs and CSI. Thus, EPA could be
important for obtaining a variety of anticancer effects in cancer
therapies.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
NA, NN and KT performed most of the experiments. TY,
YuM and TH collected the cultured fibroblasts. MH, KS and ST
designed the study. NA, MH, HI, YoM and HT analyzed the obtained
data. ST conducted the entire study. TY, YuM, TH, MH and KS
contributed to the conception and design of the study and were also
responsible for the acquisition, analysis and interpretation of the
data. All authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by Nagoya City
University Graduate School of Medical Sciences and Nagoya City
University Hospital Institutional Review Board (18–127). Written
informed consent was obtained from the patient.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
EPA
|
eicosapentaenoic acid
|
|
VEGF
|
vascular endothelial growth factor
|
|
CAF
|
cancer-associated fibroblast
|
|
HUVECs
|
human umbilical vein endothelial
cells
|
|
IL-6
|
interleukin-6
|
|
LPS
|
lipopolysaccharide
|
|
PUFA
|
polyunsaturated fatty acid
|
|
CSI
|
cancer-stromal interaction
|
References
|
1
|
Shiga K, Hara M, Nagasaki T, Sato T,
Takahashi H and Takeyama H: Cancer-associated fibroblasts: Their
characteristics and their roles in tumor growth. Cancers (Basel).
7:2443–2458. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wu MH, Hong HC, Hong TM, Chiang WF, Jin YT
and Chen YL: Targeting galectin-1 in carcinoma-associated
fibroblasts inhibits oral squamous cell carcinoma metastasis by
downregulating MCP-1/CCL2 expression. Clin Cancer Res.
17:1306–1316. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang Y, Tang H, Cai J, Zhang T, Guo J,
Feng D and Wang Z: Ovarian cancer-associated fibroblasts contribute
to epithelial ovarian carcinoma metastasis by promoting
angiogenesis, lymphangiogenesis and tumor cell invasion. Cancer
Lett. 303:47–55. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rosen LS: Clinical experience with
angiogenesis signaling inhibitors: Focus on vascular endothelial
growth factor (VEGF) blockers. Cancer Control. 9 (Suppl 2):S36–S44.
2002. View Article : Google Scholar
|
|
5
|
Procaccio L, Damuzzo V, Di Sarra F, Russi
A, Todino F, Dadduzio V, Bergamo F, Prete AA, Lonardi S, Prenen H,
et al: Safety and tolerability of anti-angiogenic protein kinase
inhibitors and vascular-disrupting agents in cancer: Focus on
gastrointestinal malignancies. Drug Saf. 42:159–179. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cavaco A, Rezaei M, Niland S and Eble JA:
Collateral damage intended-cancer-associated fibroblasts and
vasculature are potential targets in cancer therapy. Int J Mol Sci.
18:E23552017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Jung JG and Le A: Targeting metabolic
cross talk between cancer cells and cancer-associated fibroblasts.
Adv Exp Med Biol. 1063:167–178. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Melzer C, von der Ohe J, Lehnert H,
Ungefroren H and Hass R: Cancer stem cell niche models and
contribution by mesenchymal stroma/stem cells. Mol Cancer.
16:282017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ramamonjisoa N and Ackerstaff E:
Characterization of the tumor microenvironment and tumor-stroma
interaction by non-invasive preclinical imaging. Front Oncol.
7:32017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nagasaki T, Hara M, Nakanishi H, Takahashi
H, Sato M and Takeyama H: Interleukin-6 released by colon
cancer-associated fibroblasts is critical for tumour angiogenesis:
Anti-interleukin-6 receptor antibody suppressed angiogenesis and
inhibited tumour-stroma interaction. Br J Cancer. 110:469–478.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kubota H, Matsumoto H, Higashida M,
Murakami H, Nakashima H, Oka Y, Okumura H, Yamamura M, Nakamura M
and Hirai T: Eicosapentaenoic acid modifies cytokine activity and
inhibits cell proliferation in an oesophageal cancer cell line.
Anticancer Res. 33:4319–4324. 2013.PubMed/NCBI
|
|
12
|
Adam O: Dietary fatty acids and immune
reactions in synovial tissue. Eur J Med Res. 8:381–387.
2003.PubMed/NCBI
|
|
13
|
Pappalardo G, Almeida A and Ravasco P:
Eicosapentaenoic acid in cancer improves body composition and
modulates metabolism. Nutrition. 31:549–555. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Serini S and Calviello G: Modulation of
Ras/ERK and phosphoinositide signaling by long-chain n-3 PUFA in
breast cancer and their potential complementary role in combination
with targeted drugs. Nutrients. 9:E1852017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu Z, Hopkins MM, Zhang Z, Quisenberry
CB, Fix LC, Galvan BM and Meier KE: Omega-3 fatty acids and other
FFA4 agonists inhibit growth factor signaling in human prostate
cancer cells. J Pharmacol Exp Ther. 352:380–394. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liang P, Henning SM, Schokrpur S, Wu L,
Doan N, Said J, Grogan T, Elashoff D, Cohen P and Aronson WJ:
Effect of dietary Omega-3 fatty acids on tumor-associated
macrophages and prostate cancer progression. Prostate.
76:1293–1302. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cao W, Ma Z, Rasenick MM, Yeh S and Yu J:
N-3 poly-unsaturated fatty acids shift estrogen signaling to
inhibit human breast cancer cell growth. PLoS One. 7:e528382012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Han L, Zhang Y, Meng M, Cheng D and Wang
C: Eicosapentaenoic acid induced SKOV-3 cell apoptosis through
ERK1/2-mTOR- NF-kappaB pathways. Anticancer Drugs. 27:635–642.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Eltweri AM, Howells LM, Thomas AL,
Dennison AR and Bowrey DJ: Effects of Omegaven®, EPA,
DHA and oxaliplatin on oesophageal adenocarcinoma cell lines
growth, cytokine and cell signal biomarkers expression. Lipids
Health Dis. 17:192018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Calviello G, Di Nicuolo F, Gragnoli S,
Piccioni E, Serini S, Maggiano N, Tringali G, Navarra P, Ranelletti
FO and Palozza P: n-3 PUFAs reduce VEGF expression in human colon
cancer cells modulating the COX-2/PGE2 induced ERK-1 and −2 and
HIF-1alpha induction pathway. Carcinogenesis. 25:2303–2310. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Morin C, Rodriguez E, Blier PU and Fortin
S: Potential application of eicosapentaenoic acid monoacylglyceride
in the management of colorectal cancer. Mar Drugs. 15:E2832017.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mueller L, Goumas FA, Himpel S, Brilloff
S, Rogiers X and Broering DC: Imatinib mesylate inhibits
proliferation and modulates cytokine expression of human
cancer-associated stromal fibroblasts from colorectal metastases.
Cancer Lett. 250:329–338. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Maeda Y, Takahashi H, Nakai N, Yanagita T,
Ando N, Okubo T, Saito K, Shiga K, Hirokawa T, Hara M, et al:
Apigenin induces apoptosis by suppressing Bcl-xl and Mcl-1
simultaneously via signal transducer and activator of transcription
3 signaling in colon cancer. Int J Oncol. Mar 7–2018.(Epub ahead of
print). doi: 10.3892/ijo.2018.4308. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Matsuo Y, Campbell PM, Brekken RA, Sung B,
Ouellette MM, Fleming JB, Aggarwal BB, Der CJ and Guha S: K-Ras
promotes angiogenesis mediated by immortalized human pancreatic
epithelial cells through mitogen-activated protein kinase signaling
pathways. Mol Cancer Res. 7:799–808. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nakagawa H, Liyanarachchi S, Davuluri RV,
Auer H, Martin EW Jr, de la Chapelle A and Frankel WL: Role of
cancer-associated stromal fibroblasts in metastatic colon cancer to
the liver and their expression profiles. Oncogene. 23:7366–7377.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Leung DW, Cachianes G, Kuang WJ, Goeddel
DV and Ferrara N: Vascular endothelial growth factor is a secreted
angiogenic mitogen. Science. 246:1306–1309. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Li CC, Hou YC, Yeh CL and Yeh SL: Effects
of eicosapentaenoic acid and docosahexaenoic acid on prostate
cancer cell migration and invasion induced by tumor-associated
macrophages. PLoS One. 9:e996302014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hopkins MM, Zhang Z, Liu Z and Meier KE:
Eicosopentaneoic acid and other free fatty acid receptor agonists
inhibit lysophosphatidic acid- and epidermal growth factor-induced
proliferation of human breast cancer cells. J Clin Med. 5:E162016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Denys A, Hichami A and Khan NA:
Eicosapentaenoic acid and docosahexaenoic acid modulate MAP kinase
(ERK1/ERK2) signaling in human T cells. J Lipid Res. 42:2015–2020.
2001.PubMed/NCBI
|
|
30
|
Szymczak M, Murray M and Petrovic N:
Modulation of angiogenesis by omega-3 polyunsaturated fatty acids
is mediated by cyclooxygenases. Blood. 111:3514–3521. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Greten FR, Eckmann L, Greten TF, Park JM,
Li ZW, Egan LJ, Kagnoff MF and Karin M: IKKbeta links inflammation
and tumorigenesis in a mouse model of colitis-associated cancer.
Cell. 118:285–296. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Grivennikov S, Karin E, Terzic J, Mucida
D, Yu GY, Vallabhapurapu S, Scheller J, Rose-John S, Cheroutre H,
Eckmann L and Karin M: IL-6 and Stat3 are required for survival of
intestinal epithelial cells and development of colitis-associated
cancer. Cancer Cell. 15:103–113. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mocellin MC, Camargo CQ, Nunes EA, Fiates
GMR and Trindade EBSM: A systematic review and meta-analysis of the
n-3 polyunsaturated fatty acids effects on inflammatory markers in
colorectal cancer. Clin Nutr. 35:359–369. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jung YD, Nakano K, Liu W, Gallick GE and
Ellis LM: Extracellular signal-regulated kinase activation is
required for up-regulation of vascular endothelial growth factor by
serum starvation in human colon carcinoma cells. Cancer Res.
59:4804–4807. 1999.PubMed/NCBI
|
|
35
|
Kojima Y, Acar A, Eaton EN, Mellody KT,
Scheel C, Ben-Porath I, Onder TT, Wang ZC, Richardson AL, Weinberg
RA and Orimo A: Autocrine TGF-beta and stromal cell-derived
factor-1 (SDF-1) signaling drives the evolution of tumor-promoting
mammary stromal myofibroblasts. Proc Natl Acad Sci USA.
107:20009–20014. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tomiyama K, Murase N, Stolz DB, Toyokawa
H, O'Donnell DR, Smith DM, Dudas JR, Rubin JP and Marra KG:
Characterization of transplanted green fluorescent protein+ bone
marrow cells into adipose tissue. Stem Cells. 26:330–338. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ishii G, Sangai T, Oda T, Aoyagi Y, Hasebe
T, Kanomata N, Endoh Y, Okumura C, Okuhara Y, Magae J, et al:
Bone-marrow-derived myofibroblasts contribute to the cancer-induced
stromal reaction. Biochem Biophys Res Commun. 309:232–240. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Iwano M, Plieth D, Danoff TM, Xue C, Okada
H and Neilson EG: Evidence that fibroblasts derive from epithelium
during tissue fibrosis. J Clin Invest. 110:341–350. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Powell DW, Adegboyega PA, Di Mari JF and
Mifflin RC: Epithelial cells and their neighbors I. Role of
intestinal myofibroblasts in development, repair, and cancer. Am J
Physiol Gastrointest Liver Physiol. 289:G2–G7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Worthley DL, Giraud AS and Wang TC:
Stromal fibroblasts in digestive cancer. Cancer Microenviron.
3:117–125. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Paland N, Kamer I, Kogan-Sakin I, Madar S,
Goldfinger N and Rotter V: Differential influence of normal and
cancer-associated fibroblasts on the growth of human epithelial
cells in an in vitro cocultivation model of prostate cancer. Mol
Cancer Res. 7:1212–1223. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Fuyuhiro Y, Yashiro M, Noda S, Kashiwagi
S, Matsuoka J, Doi Y, Kato Y, Hasegawa T, Sawada T and Hirakawa K:
Upregulation of cancer-associated myofibroblasts by TGF-beta from
scirrhous gastric carcinoma cells. Br J Cancer. 105:996–1001. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Higashino N, Koma YI, Hosono M, Takase N,
Okamoto M, Kodaira H, Nishio M, Shigeoka M, Kakeji Y and Yokozaki
H: Fibroblast activation protein-positive fibroblasts promote tumor
progression through secretion of CCL2 and interleukin-6 in
esophageal squamous cell carcinoma. Lab Invest. Jan 25–2019.(Epub
ahead of print). doi: 10.1038/s41374-018-0185-6. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ebbing EA, van der Zalm AP, Steins A,
Creemers A, Hermsen S, Rentenaar R, Klein M, Waasdorp C, Hooijer
GKJ, Meijer SL, et al: Stromal-derived interleukin 6 drives
epithelial-to-mesenchymal transition and therapy resistance in
esophageal adenocarcinoma. Proc Natl Acad Sci USA. 116:2237–2242.
2019. View Article : Google Scholar : PubMed/NCBI
|