Introduction
Multiple myeloma (MM) is a B-cell malignancy
characterized by the abnormal proliferation of plasma cells in the
bone marrow (1). For hematological
malignancies, the incidence of MM is second only to non-Hodgkin's
lymphoma, accounting for ~13% of hematological malignancies
(2). The disease often occurs in
the elderly over the age of 60 and with aging of the global
population, the incidence of is increasing (3). The treatment of MM mainly comprises
chemotherapy, radiotherapy, immunotherapy, autologous stem cell
transplantation and targeted therapy (4). Autologous stem cell transplantation
can improve patient survival (5);
however, as autologous stem cell transplantation is difficult to
perform in the elderly and MM is prone to relapse, chemotherapy is
administered as the main clinical treatment (4). In recent years, the pathogenesis,
development and effective therapies of MM has been extensively
studied; however, further investigation is required to identify the
mechanism underlying the pathogenesis and develop novel therapeutic
strategies for treating this disease (6). Therefore, it is urgent to identify
novel targets for the diagnosis and treatment of MM.
MicroRNAs (miRNAs/miRs) are small RNA molecules of
~22 bases. Studies have reported miRNAs to serve important roles in
cellular events, including cell proliferation, differentiation,
apoptosis and metabolism (7–10).
They also play important regulatory roles in human diseases, such
as cancer, blood diseases, cardiovascular diseases and diabetes
(11). In 2004, Chen et al
(12) demonstrated that miRNAs were
involved in the regulation of human hematopoietic system for the
first time and the miRNA expression profiles were abnormal in a
variety of hematological diseases. The miR-181 family constitutes
ubiquitous miRNAs in human cells and are abnormally expressed in a
variety of tumors (13). miR-181
has been reported to be expressed in lung cancer, gastric cancer
and hematological diseases (14–16).
miR-181a of the miR-181 family was determined to be closely related
to the regulation of the hematopoietic system (16,17). A
total of 12 out of 13 differentially expressed miRNA molecules,
including miR-181a, were upregulated in MM (18). We previously reported that miR-181a
and miR-20a were highly expressed in serum of patients with MM
(19); high expression of miR-181a
was proposed as a marker of poor prognosis of MM (20). In addition, our recent study showed
that in the serum and bone marrow of patients with MM, miR-181a was
highly expressed and positively correlated with Durie-Salmon stage,
renal function, and β2-microglobulin levels (21); however, the role of miR-181a in MM
cells and its downstream target genes remain unclear.
The present study aimed to determine the expression
profile of miR-181a in MM cells, and analyze the proliferation,
cell cycle, apoptosis and invasion of MM cells after transfection
of miR-181a in MM cells using lentiviral (LV) transduction. The
target genes of miR-181a were predicted by bioinformatics analysis.
Animal experiments using nude mice were performed to investigate
the effect of miR-181a on MM in vivo.
Materials and methods
Animals
Female severe combined immunodeficiency (SCID Berge)
mice of 3–5 weeks old (n=12) were purchased from Xi'an Jiaotong
University Animal Experimental Center. Mice were housed in standard
conditions, under a 12 h light/dark cycle, free access to rodent
chow and water, room temperature of 18–25 °C and relative humidity
of 40–60 %. All experimental procedures involving animals were
conducted according to the ethical guidelines of The First
Affiliated Hospital of Xi'an Jiaotong University. Ethical approval
was received for the use of animals, prior to the start of the
study (grant no. XJTULAC2017-782). All efforts were made to
minimize animal suffering.
Isolation of human bone marrow
lymphocytes
Bone marrow lymphocytes (22) were isolated from 5 patients with
benign blood disorders [anemia (n=3) and thrombocytopenia (n=2)].
Their average age was 55.2±14.06 years old (age range: 51–73
years). There were 3 female patients (all with anemia) and 2 male
patients (all with thrombocytopenia). Prior written and informed
consent were obtained from every patient, and the study was
approved by the ethics review board of The First Affiliated
Hospital of Xi'an Jiaotong University (approval no.
XJTU1AF2016LSK-46).
Cell culture
The human MM cell lines RPMI 8226 (a gift from
Professor Aili He, Department of Hematology, The Second Affiliated
Hospital of Xi'an Jiaotong University) and H929 (a gift from
Professor Xiequn Chen, Department of Hematology, Xijing Hospital of
Air Force Military Medical University) were cultured in RPMI 1640
medium (HyClone, GE Healthcare Life Sciences) containing 10% fetal
bovine serum (Zhejiang Tianhang Biotechnology Co., Ltd.), and
cultured in a humidified incubator containing 5% CO2 at
37°C.
Cell transduction and screening
RPMI 8226 cells were seeded into 96-well plates at a
concentration of 2×105 cells/well. Then, the cells were
transfected with LV3-HIV-miR-181a-5p mimics-GFP (LV-miR-181a mimics
group), LV3-HIV-miR-181a-5p inhibitor-GFP (LV-miR-181a inhibitor
group) and LV3-negative control-GFP (LV-NC), respectively (all from
Shanghai GenePharma Co., Ltd.). The sequences were as follows.
LV3-miR-181a mimics: 5′-AACATTCAACGCTCGGTGAGT-3′; LV3-miR-181a
inhibitor: 5′-ACTCACCGACAGCGTTGAATGTT-3′; LV3-NC:
5′-TTCTCCGAACGTGTCACGT-3′. After 48 h, the culture medium was
changed. Then, cell culture was continued for another 24 h. Cells
were screened with puromycin (4 µg/ml) (Sigma-Aldrich; Merck KGaA).
After transduction, cells were observed under a fluorescent
microscope (Olympus CKX41; Olympus, Tokyo, Japan). The experiment
was repeated three times.
Animal treatment, grouping and
sampling
SCID Berge mice were randomly divided into four
groups (n=3): miR-181a mimics, miR-181a inhibitor and two LV-NC
groups. RPMI 8226 cells (1×107/cell) transduced with
different miRNA-containing vectors were suspended in 100 µl PBS and
subcutaneously inserted into the back of corresponding SCID Berge
mice groups, respectively. The mice were monitored for tumor growth
every 3 days. The tumor growth rate was relatively slower in
miR-181a inhibitor group than that in miR-181a mimics group. Thus,
tumor growth in miR-181a mimics group and its corresponding LV-NC
group was observed for 21 days, while that in miR-181a inhibitor
group and its corresponding LV-NC group was observed for 33 days.
The tumor was measured every 3 days. Tumor volumes were calculated
as 1⁄2 ×width (mm) × length (mm). Tumor bearing mice were
sacrificed by cervical dislocation. Then, the tumors were excised
and weighed. The expression of neuro-oncological ventral antigen-1
(NOVA1) in tumor tissues was detected by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR).
RT-qPCR
Total RNAs were extracted from tumor tissues and
cells with TRIzol (Thermo Fisher Scientific, Inc.). Then, RNA was
reverse transcribed into cDNA using RNA Reverse Transcription Kit
(Tiangen Biotech Co., Ltd.) according to the manufacturer's
protocols. The miRNA Real-Time PCR Assay kit (Tiangen Biotech Co.,
Ltd.) was used to detect the expression levels of miR-181a and U6.
The PCR procedure for miRNA was 95°C for 15 min followed by 45
cycles of 95°C for 20 sec and 60°C for 34 sec. The mRNA Real-Time
PCR Assay kit (Tiangen Biotech Co., Ltd.) was used to detect the
expression levels of NOVA1 and GAPDH. U6 and GAPDH were used as
internal controls. The PCR procedure for mRNA was 95°C for 15 min
followed by 45 cycles of 95°C for 10 sec and 60°C for 30 sec.
Primers were synthesized by Beijing Dingguo Changsheng
Biotechnology Co., Ltd. and the sequences were listed in Table I. The PCR assays were performed
using the SuperReal Color Fluorescent Quantitative Premixed Reagent
(Tiangen Biotech Co., Ltd.) and an ABI Step one real-time PCR
system (Applied Biosystems; Thermo Fisher Scientific, Inc.). The
comparative 2−ΔΔCq method (23) was used for relative
quantification.
 | Table I.Primers used in the present
study. |
Table I.
Primers used in the present
study.
| Gene | Sequence
(5′-3′) |
|---|
| U6-R |
AACGCTTCACGAATTTGCGT |
| U6-F |
CTCGCTTCGGCAGCACA |
| miRNA-181a-F |
AACATTCAACGCTGTCGGTGAGT |
|
miRNA-181a-Ra | – |
| GAPDH-F |
AATGGGCAGCCGTTAGGAAA |
| GAPDH-R |
GCGCCCAATACGACCAAATC |
| NOVA1-F |
CCCTCCCCAGTGAGAACAAA |
| NOVA1-R |
CCGCCATCATGTTTGCAGTT |
Cell Counting Kit-8 (CCK-8) assay
Cell viability was measured using a Cell Counting
Kit-8 assay (Genview). Cells were seeded into 96-well plates at a
concentration of 2×103 cells/well for 24 h. After 24, 48
and 72 h, 10 µl CCK-8 was added in each well. After incubation for
2 h in 37°C CO2 incubator, the absorbance at 450 nm was
measured using an Infinite F50 microplate reader (Tecan Group,
Ltd.).
Flow cytometric analysis
Cells were cultured for 24 h, collected by
centrifugation at 800 × g for 5 min at room temperature, and washed
with pre-chilled PBS buffer (pH 7.2) three times. To investigate
the cell cycle, a Cell Cycle Detection Kit (Beyotime Institute of
Biotechnology) was used. The cells were fixed with pre-chilled 75%
alcohol at 4°C for 24 h and stained with propidium iodide at 37°C
in the dark for 30 min. A flow cytometer (BD Biosciences) was then
used to analyze cell cycle. To detect apoptosis, cells were
resuspended with 195 µl assay buffer (included in kit), stained
with 5 µl Annexin V-phycoerythrin (Beyotime Institute of
Biotechnology) in the dark for 10–20 min at room temperature, and
then 5 µl 7-amino actinomycin D (7-AAD) (Keygentec) in the dark for
5–15 min on ice. Cells were then analyzed by flow cytometry. ModFit
LT 5.0 software (Verity Software House, Inc.) was used to analyze
the cell cycle, and CellQuest Pro 5.1 (BD Biosciences) software was
used to analyze apoptosis. Cells with positive Annexin V and
negative 7-AAD were in early apoptosis; cells positive for both
Annexin V and 7-AAD were in late apoptosis.
Transwell invasion assay
Matrigel (5 mg/ml; BD Biosciences) was thawed at 4°C
overnight and diluted in pre-cooled serum-free medium to a final
concentration of 1 mg/ml. Then, 100 µl of diluted Matrigel was
added to upper chamber (EMD Mililpore) and incubated overnight at
37°C. Cells were resuspended in serum-free medium at a
concentration of 2×105/ml. A total of 750 µl of complete
medium containing 10% fetal bovine serum was added to the lower
chamber and 200 µl of cell suspension was added to the upper
chamber. Subsequently, cells were cultured for 18–24 h at 37°C.
After removing the medium and washing with PBS, cells were fixed
with 95% ethanol at room temperature for 5 min and permeabilized
with 100% methanol at room temperature for 20 min. After washing,
cells were stained with 0.1% crystal violet at room temperature for
20 min. The non-invading cells were removed using a cotton swab.
Invasive cells were counted in 10 randomly selected fields under a
fluorescence microscope under bright field (Olympus CKX41;
magnification, ×200). A hemocytometer was used to count cells under
the microscope.
Target gene prediction
Bioinformatics software miRanda (http://miranda.org.uk/), TargetScan5.2 (http://www.targetscan.org/vert_72/) miRDB 5.0
(http://www.mirdb.org/), Pictar (https://pictar.mdc-berlin.de/) and miRTarBase 6.1
(http://mirtarbase.mbc.nctu.edu.tw/php/index.php) were
used to predict the target genes of miR-181a.
Statistical analysis
The data was analyzed using SPSS 17.0 (SPSS Inc.)
and GraphPad Prism 5.0 (GraphPad Software, Inc.) for
Windows®. Data from at least three independent
experiments were expressed as the mean ± standard deviation. A
Student's t-test or one-way ANOVA followed by a
Student-Newman-Keuls test was used when appropriate. P<0.05 was
considered to indicate a statistically significant.
Results
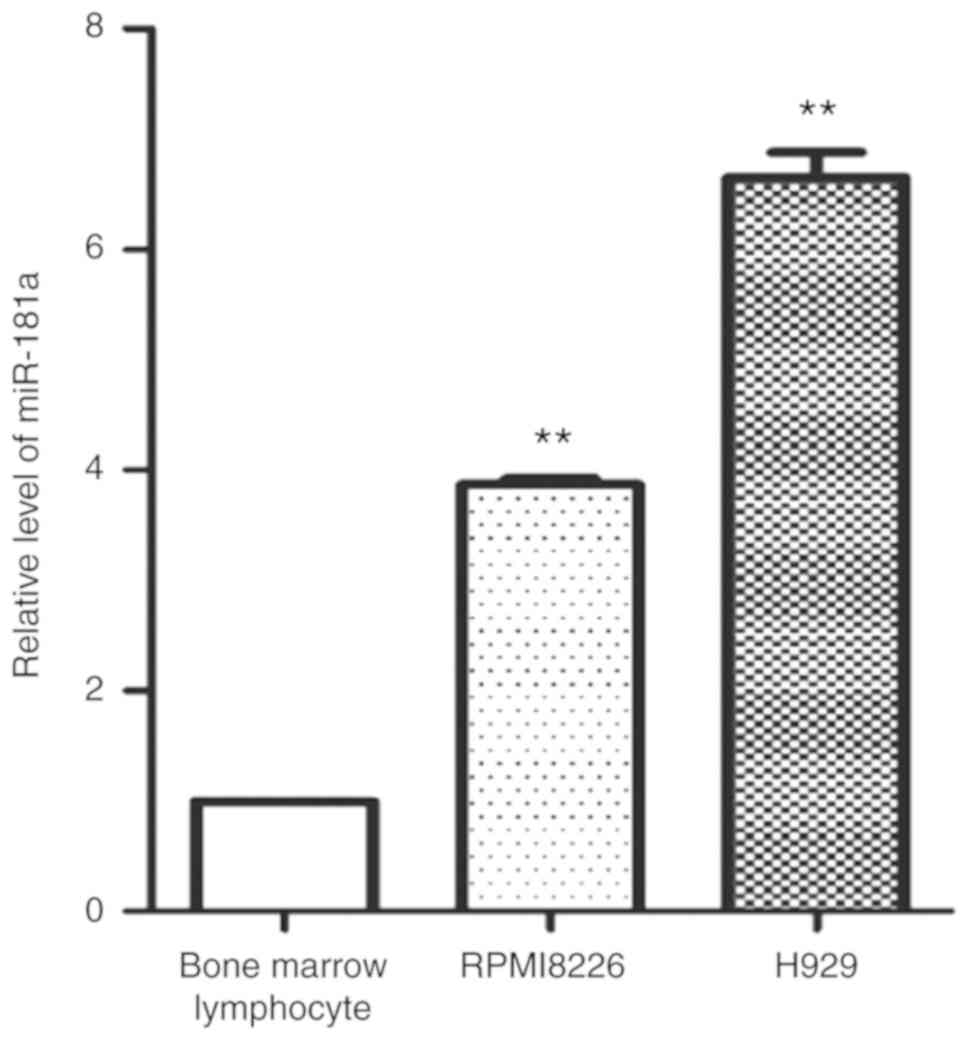
Expression of miR-181a in MM cell
lines
RT-qPCR was used to detect the expression of
miR-181a in MM cell lines (RPMI 8226 and H929 cells) and bone
marrow lymphocytes. The relative expression levels of miR-181a were
significantly higher in the MM cell lines (RPMI 8226 and H929
cells) as compared with that in bone marrow lymphocytes (Fig. 1). This result showed that miR-181a
was highly expressed in MM cell lines.
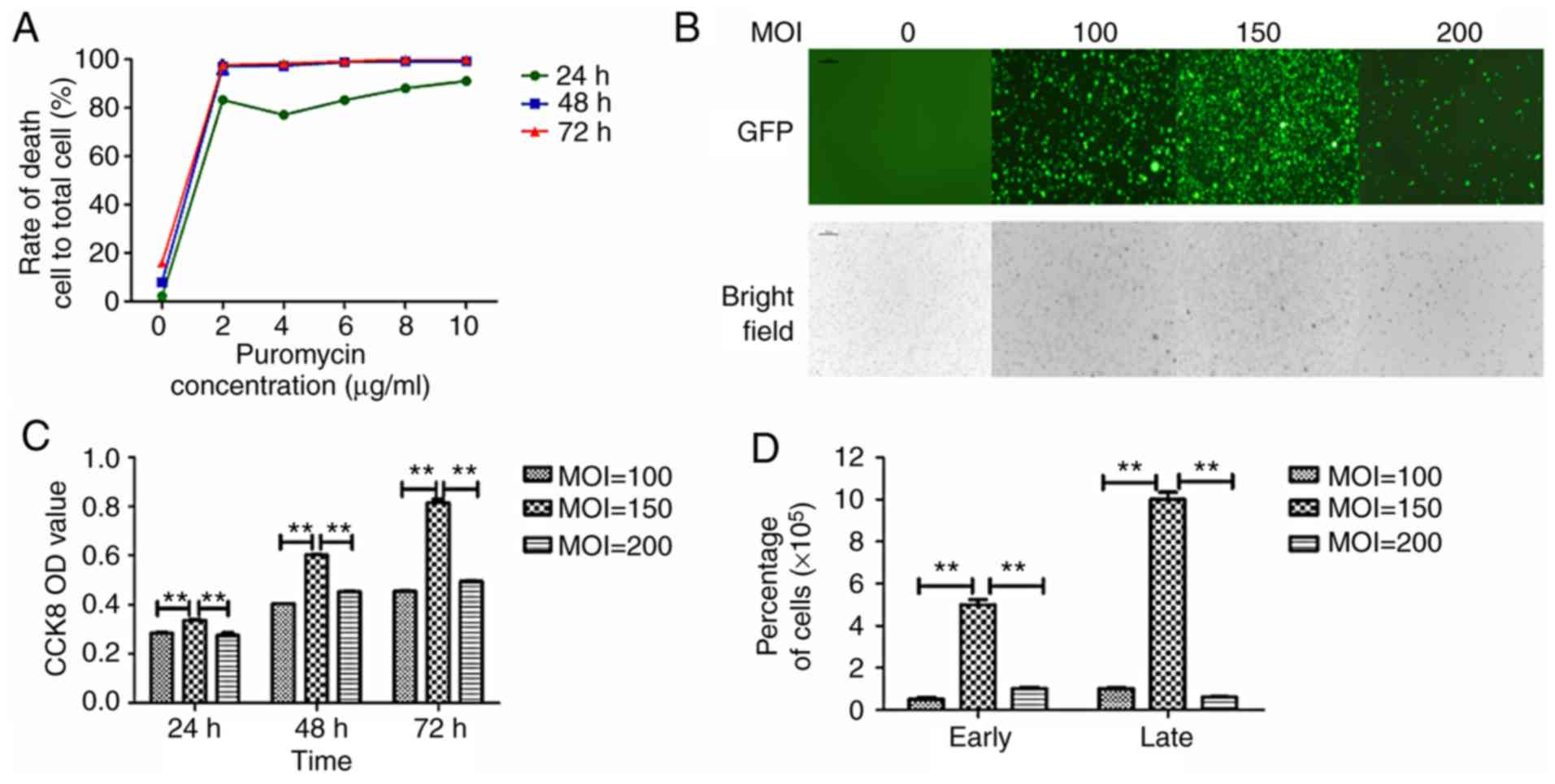
In vitro LV transfection efficiency of
MM cells
miR-181a mimics, miR-181a inhibitor and LV-NC were
transfected into RPMI 8226 cell line by lentivirus. The
transduction efficiency was detected and the effects of miR-181a on
cell proliferation, cycle, apoptosis and invasion were
investigated. A total of >99% of dead nontransfected cells were
observed on the third day when the concentration of puromycin was 4
µg/µl (Fig. 2A). Therefore, 4 µg/µl
puromycin was used to screen transfected cells and 2 µg/µl was used
to maintain cultures. Different MOI values for LV transduction
conditions were investigated in three generations. The expression
of green fluorescent protein (GFP) under fluorescence microscope
was observed (Fig. 2B). The
fluorescence intensity was strongest when the MOI value was 150.
Cell proliferation was analyzed via a CCK-8 assay (Fig. 2C) and the cell number (Fig. 2D) also revealed that an MOI of 150
resulted in significantly increased cell viability as compared with
MOIs of 100 and 200.
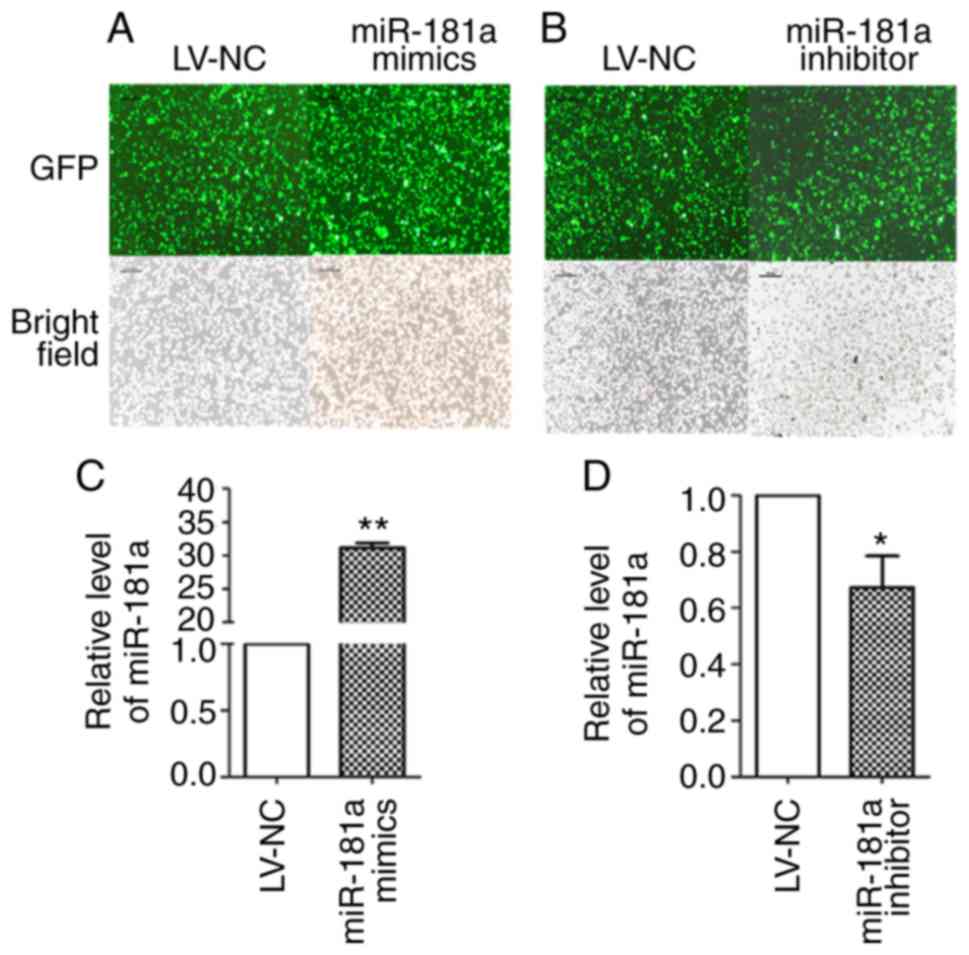
The expression of GFP in the stably transduced cell
line after transduction with LV-miR-181a mimics, LV-miR-181a
inhibitor and LV-NC was observed under an inverted fluorescence
microscope (Fig. 3A and B). The
transfection efficiency of the three groups was >90%. RT-qPCR
revealed that miR-181a expression was significantly increased in
the miR-181a mimics group compared with the LV-NC group (t=−73.968,
P<0.01; Fig. 3C), and the
expression of miR-181a in the miR-181a inhibitor group was
significantly downregulated (t=5.068, P=0.037; Fig. 3D). These results demonstrated that
LV-mediated transduction of cell lines was successfully
attained.
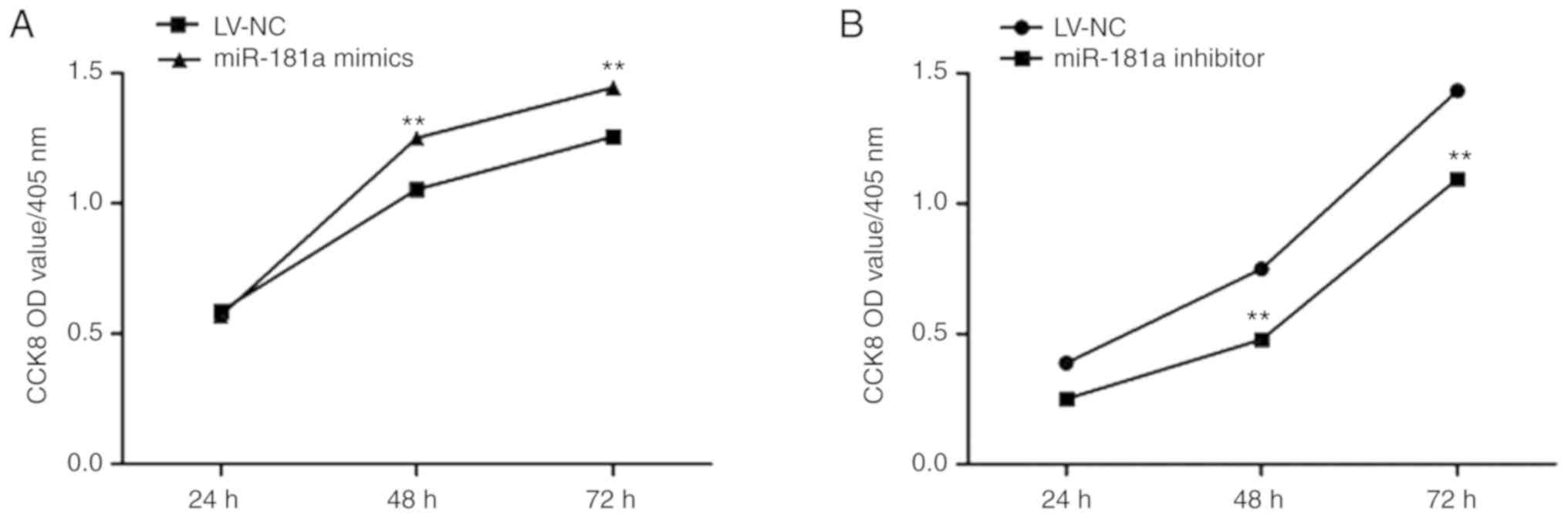
Effects of miR-181a on the
proliferation of MM cells
A CCK-8 assay was conducted to detect cell
proliferation. Compared with the LV-NC group, the proliferation of
cells of the miR-181a mimics group significantly increased at 48
and 72 h (t=−3.475, P=0.008; Fig.
4A), whereas, miR-181a inhibitor cell proliferation was
significantly inhibited (t=8.098, P<0.01; Fig. 4B). This suggests that upregulation
of miR-181a expression enhances the proliferative ability of RPMI
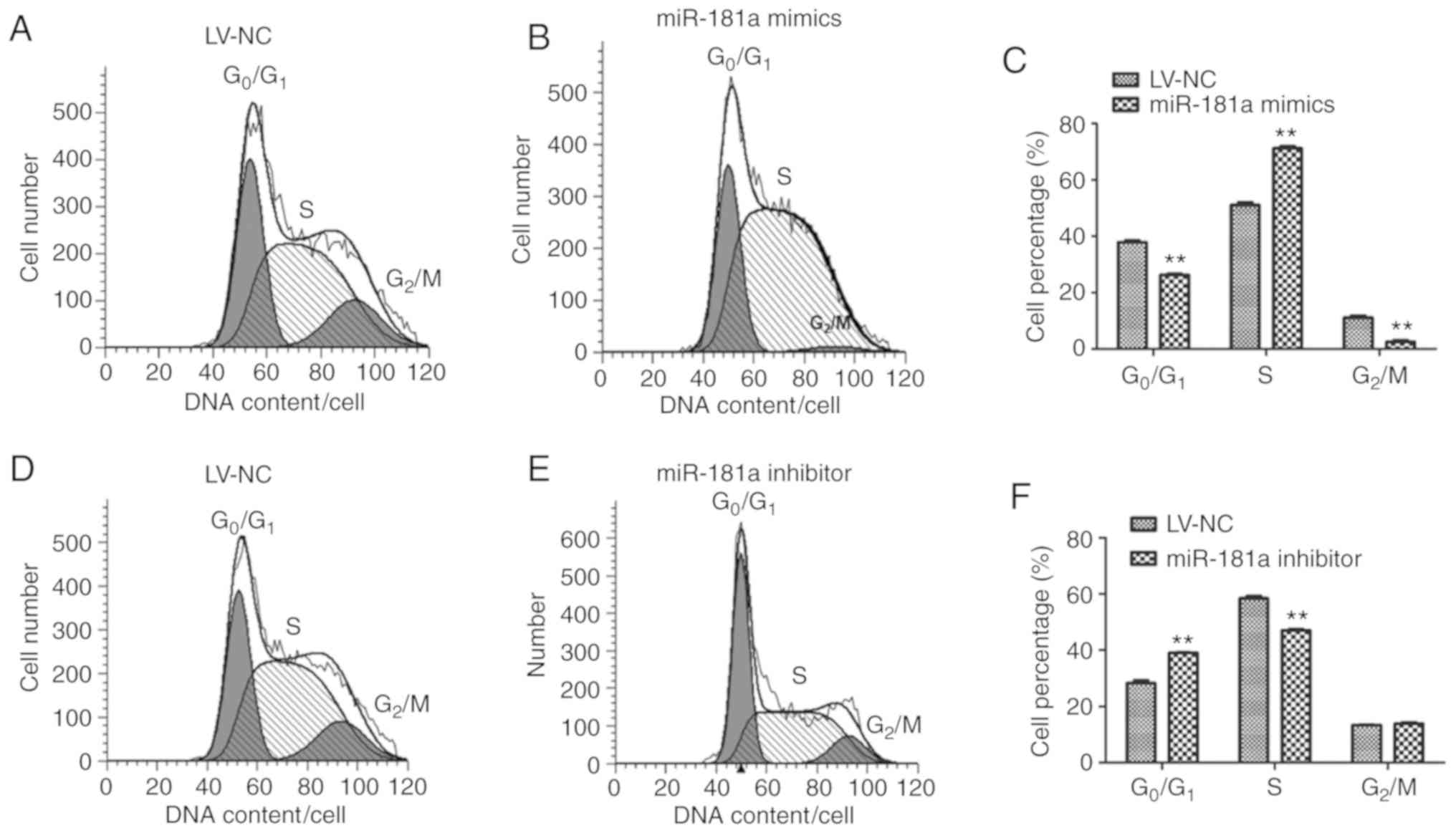
8226 cells. Flow cytometry was used to detect the cell cycle status
of RPMI 8226 cells. Compared with the LV-NC group (Fig. 5A), the number of cells in G0/G1
phase of the miR-181a mimics group (Fig. 5B) was significantly decreased
(t=11.529, P=0.006), but increased in S phase compared with the
control (t=−27.819, P<0.01) (Fig.
5C). The number of cells in G0/G1 phase of the miR-181a
inhibitor group increased significantly (t=−7.965, P=0.001), but
decreased in S phase compared with the control (t=20.610,
P<0.01) (Fig. 5D-F). These
results indicated that upregulation of miR-181a may induce the
progression of the cell cycle to S phase in RPMI 8226 cells,
promoting DNA synthesis and cell proliferation.
Effects of miR-181a transfection on
the cell cycle of MM cells
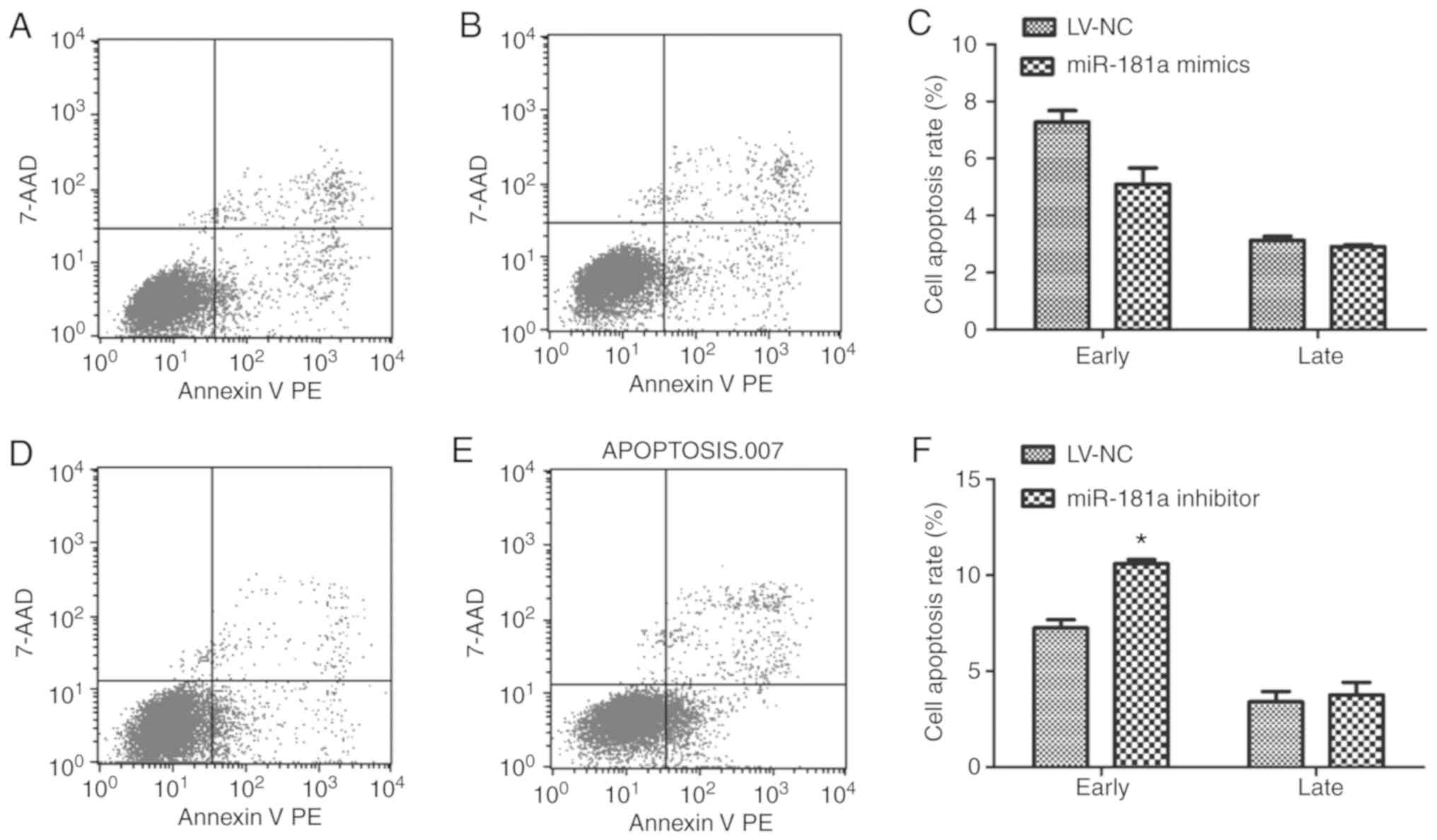
Flow cytometry was also used to detect the apoptosis
of each group. Compared with the LV-NC group, there was no
significant difference in the early apoptotic rate in the miR-181a
mimics (t=4.438, P=0.058; Fig.
6A-C). While, the early apoptotic rate in the miR-181a
inhibitor group (Fig. 6D and E) was
significantly higher than that in the LV-NC group (t=−9.812,
P=0.026; Fig. 6F). This
demonstrated that downregulation of miR-181a could enhance the
apoptotic rate of RPMI 8226 cells.
Effects of miR-181a transduction on
the invasion of MM cells
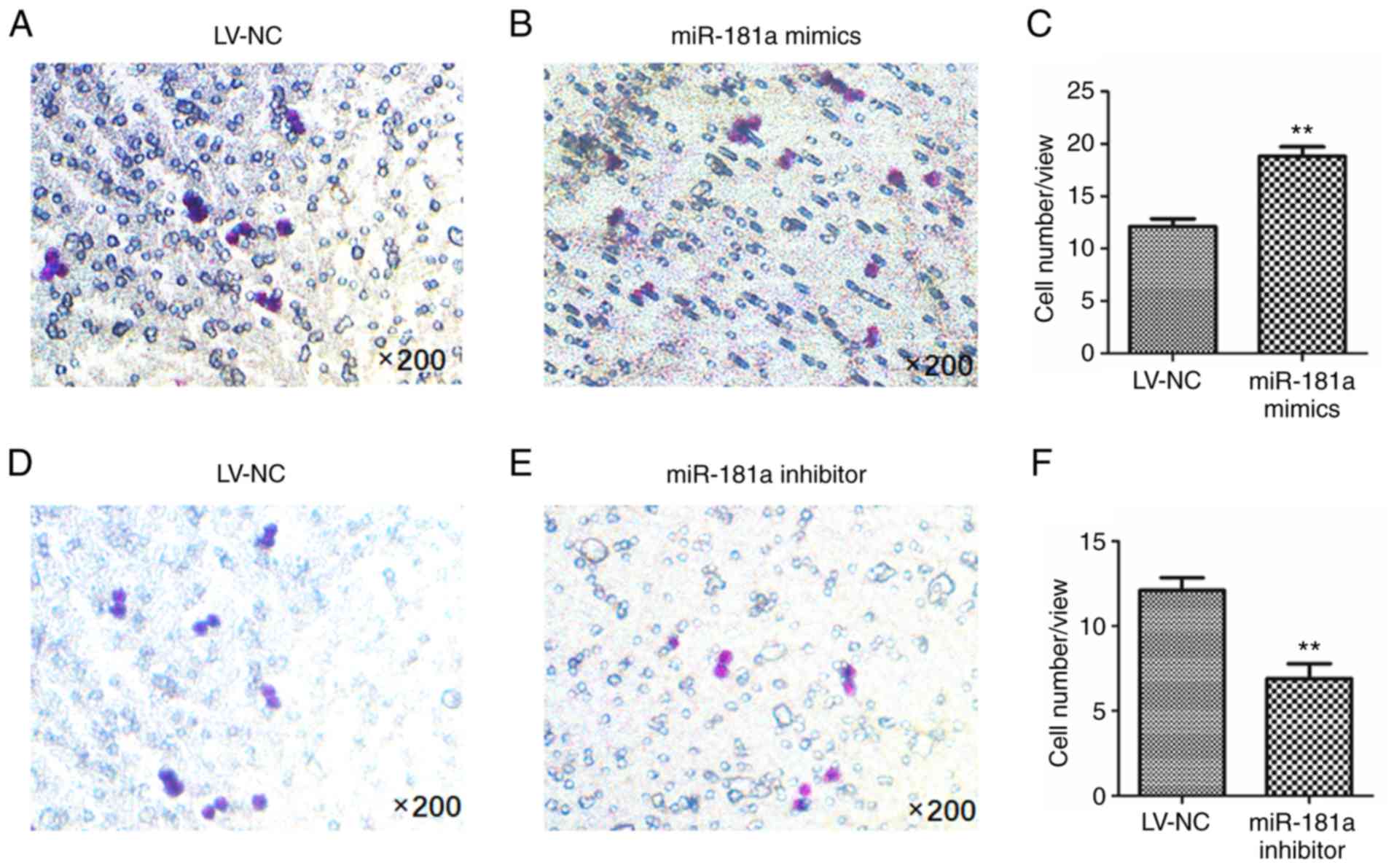
A Transwell assay was performed to detect cell
invasion. As presented in Fig. 7A and
B, LV-transduction of miR-181a mimics revealed a significant
increase in the number of invaded cells than the LV-NC group
(t=−17.978, P=0.001; Fig. 7C);
however, the number of invaded cells in the inhibitor group was
significantly less than that in the LV-NC group (t=14.361, P=0.001;
Fig. 7D-F). These results suggested
that upregulation of miR-181a may enhance the invasive ability of
RPMI 8226 cells.
Effects of miR-181a on tumor growth in
vivo
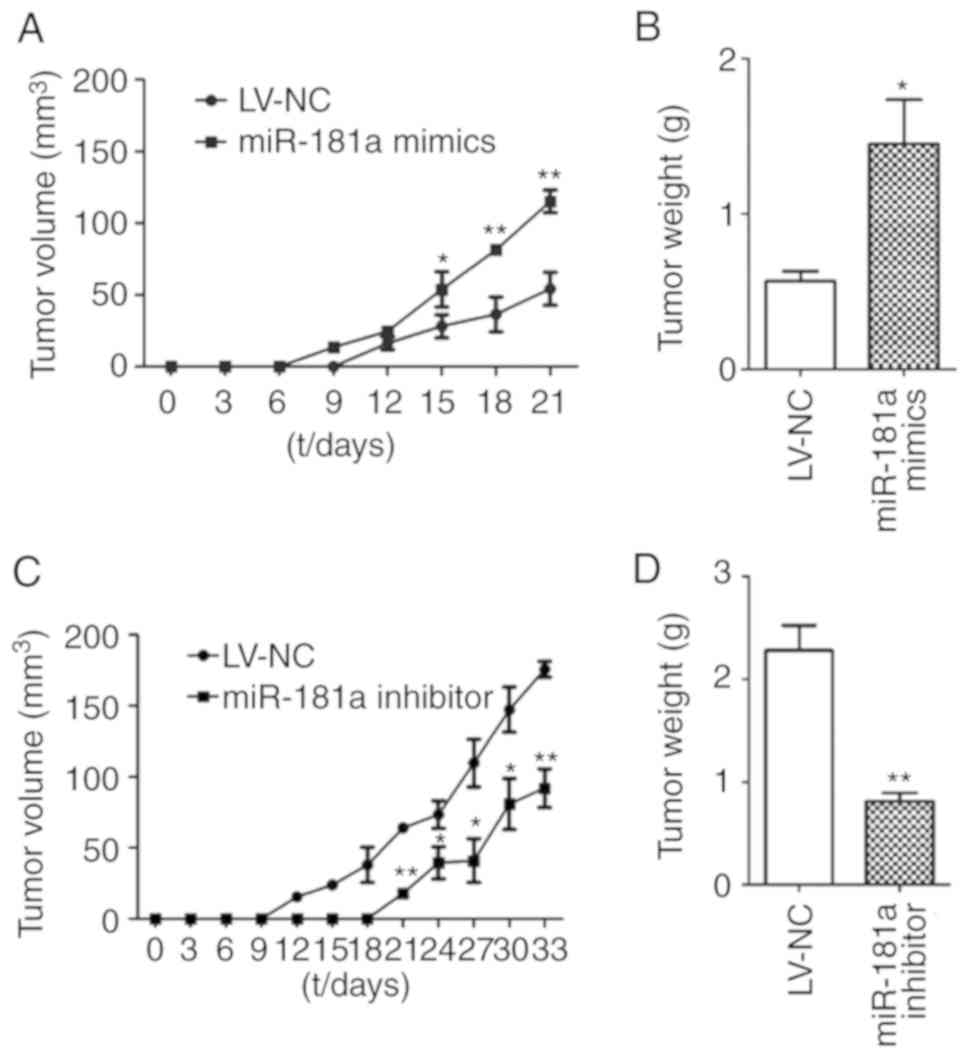
Xenografts in mice were established using stable
transduced cells. Tumor volume was measured every 3 days and the
tumors were weighed at the 4th week. The longest tumor diameter for
miR-181 mimics group and its corresponding LV-NC group was observed
at day 21, which was 13.59×18.27 mm and 10.65×12.59 mm,
respectively. The longest tumor diameter for the miR-181 inhibitor
group and the corresponding LV-NC group was observed at day 33,
which was 11.56×17.97 mm and 17.89×20.08 mm, respectively. Compared
with the LV-NC group, the tumor volume was significantly larger in
the miR-181a mimics group (t=−5.534, P<0.01; Fig. 8A); however, the tumor volume was
significantly reduced in the miR-181a inhibitor group (t=3.249,
P=0.004). The tumor weight of the miR-181a mimics group was
significantly greater than that of the LV-NC group (t=5.216,
P=0.028; Fig. 8B). On the contrary,
the tumor weight of the miR-181a inhibitor group was significantly
lower than that of the LV-NC group (t=9.852, P=0.001; Fig. 8D). These results demonstrated that
miR-181a may promote tumor growth in vivo.
Target gene prediction and expression
analysis
To study the mechanism of miR-181a on MM,
bioinformatics was used to predict the target genes of miR-181a.
The number of miR-181a target genes preselected by bioinformatics
software (miRanda, TargetScan, miRDB, Pictar, and miRTarBase) was
7,847, 1,367, 887, 457 and 559, respectively. The intersection of
the first 100 genes of each database revealed a total of 9 target
genes (Table II), including
prospero homeobox protein 1, DEAD-box helicase 3 X-linked,
diphosphoinositol pentakisphosphate kinase 2, NOVA1, interleukin-2,
zinc finger protein 780A, membrane palmitoylated protein 5,
forkhead box P1 and mitogen-activated protein kinase 1.
| Table II.Target genes of miR-181a as predicted
with bioinformatics tools. |
Table II.
Target genes of miR-181a as predicted
with bioinformatics tools.
| miR | Gene | miRanda | targetScan | miRDB | MiRTarBase | Pictar | SUM |
|---|
| Hsa-miR-181a | PROX1 |
|
| 1 | 1 |
| 2 |
| Hsa-miR-181a | DDX3X | 1 |
| 1 |
|
| 2 |
| Hsa-miR-181a | PPIP5K2 | 1 |
| 1 |
|
| 2 |
| Hsa-miR-181a | NOVA1 | 1 |
| 1 |
| 1 | 3 |
| Hsa-miR-181a | IL-2 | 1 | 1 | 1 |
|
| 3 |
| Hsa-miR-181a | ZNF780A | 1 | 1 |
|
|
| 2 |
| Hsa-miR-181a | MPP5 | 1 |
|
|
| 1 | 2 |
| Hsa-miR-181a | FOXP1 | 1 | 1 |
|
|
| 2 |
| Hsa-miR-181a | MAPK1 | 1 | 1 |
| 1 |
| 3 |
Studies have reported that NOVA1 can promote cell
proliferation in liver cancer, gastric cancer and gliomas (20,24,25).
Therefore, the mRNA expression levels of NOVA1 in
transfected MM cells was detected by RT-qPCR. Compared with the
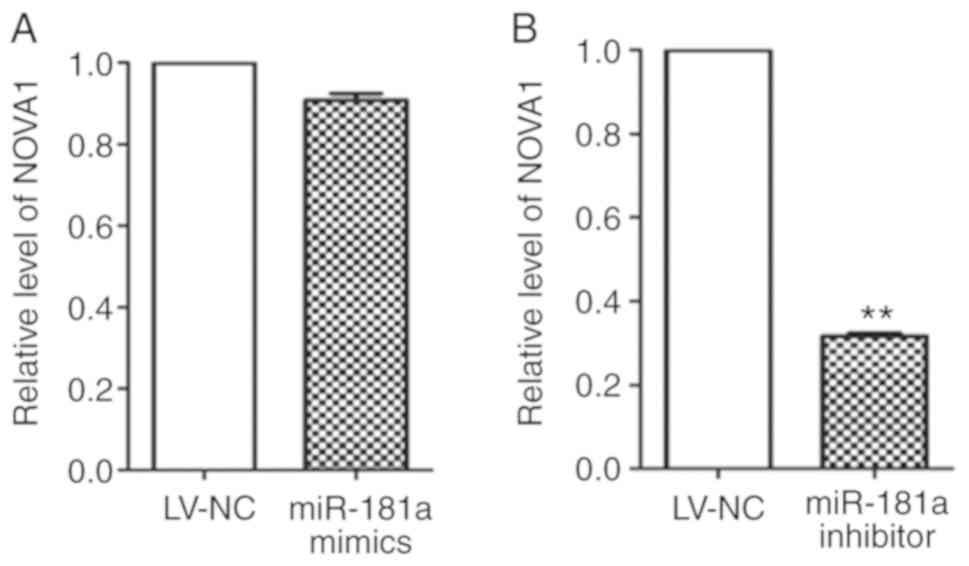
LV-NC group, the miR-181a mimics group (Fig. 9A) showed no significant change
(t=9.551, P=0.011), while the NOVA1 mRNA expression in the
miR-181a inhibitor group (Fig. 9B)
was significantly inhibited.
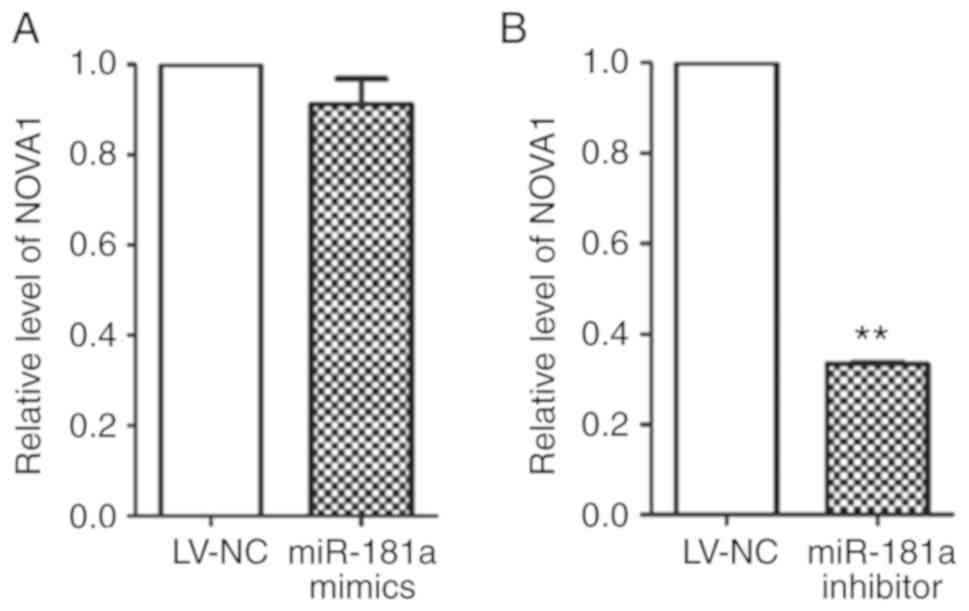
RT-qPCR was used to detect the expression of
NOVA1 at the mRNA level (Fig.
10A and B) in tumor xenograft samples. LV-mediated transduction
of miR-181a mimics exhibited no significant effects on the
expression of NOVA1, while miR-181a inhibitor significantly
downregulated the expression of NOVA1 compared with the
LV-NC group. The results were consistent with those of the in
vitro cell experiments.
Discussion
MM is a hematological malignancy that accounts for
~13% of hematological malignancies (2). In recent years, the incidence of MM
has increased annually with aging of the population worldwide
(3). The pathogenesis of MM is
complex; few symptoms are observed during the early stages of
disease and relapse frequently occurs. Clinically, this disease
cannot be cured (6). Therefore, it
is necessary to thoroughly study the pathogenesis of MM in order to
reveal its molecular mechanism, and identify novel targets for the
diagnosis and treatment of MM.
miRNAs are small RNA molecules of ~22 bases in
length that regulate gene expression at the post-transcriptional
level. The mechanism of its regulation involves the inhibition of
translation or the degradation of mRNA by binding to the mRNA
complementary sequence of the target gene, thereby inhibiting the
expression of the target gene (26). miRNA regulation serves important
regulatory roles in cell proliferation, circulation, migration and
invasion (27–29); dysregulated miRNAs have been
associated with the occurrence and development of cancer (30–32).
A study has reported that miR-181a is highly
expressed in breast and pancreatic cancers, but is downregulated in
gliomas and invasive lymphocytes (33). In non-small cell lung cancer
(33), human breast and colon
cancer (34), and cervical cancer
cells (34), miR-181a serves roles
in cell proliferation, invasion and migration, and promotes
colorectal cancer cell proliferation and drug resistance via the
Wnt/β-catenin signaling pathways (35). Furthermore, miR-181a promotes
gastric cancer cell proliferation, invasion, and inhibition of
apoptosis by downregulating myotubularin related protein 3
(30,32). These findings suggest that miR-181a
plays an important role in the development of tumors. To explore
the association between miR-181a and MM, RT-qPCR was conducted to
detect miR-181a expression in different MM cell lines. We reported
that miR-181a was upregulated in MM cell lines. We have not
identified methods for the extraction of normal plasmacytes in the
literature; MM is a B cell clone proliferative disease. Thus,
normal lymphocytes, but not normal plasmacytes, were used as
control cells in the present study.
To investigate the effects of miR-181a on MM in
vivo, tumor growth was observed every 3 days in an animal model
of xenograft tumors. The results showed that tumor growth and
weight of the miR-181a mimics group were significantly promoted
than that of the control group. The tumor growth of miR-181a
inhibitor group was inhibited, and the tumor weight was
significantly less than the control group. Thus, these results
indicated that miR-181a could promote the growth of MM tumors. This
is consistent with our findings that miR-181a promotes MM cell
proliferation in vitro. miR-181a was reported to promote
colorectal cancer cell proliferation via the Wnt/β-catenin
signaling pathway (33).
To further observe the effects of miR-181a on the
biological function of MM, the expression of miR-181a was
upregulated and downregulated via lentiviral-mediated cell
transduction. The effects of miR-181a expression on cell
proliferation, cell cycle, apoptosis and invasion were observed by
CCK-8, flow cytometry and Transwell assays. The results revealed
that cell proliferation was inhibited, the number of cells in S
phase of the cell cycle was significantly reduced, and the rate of
early apoptosis increased after downregulation of miR-181a. Cell
content in S phase can reflect the proliferative ability of the
tumor and can also be used to determine the degree of malignancy
(36). Generally, the higher the
number of cells in S phase, the greater the degree of malignancy,
which has been associated with the poor prognosis of patients
(36). These results suggest that
miR-181a may be used as a marker to predict the prognosis of
patients. Cell invasion experiments showed that miR-181a
significantly promoted the invasion of tumor cells. After
inhibiting the expression of miR-181a, the early apoptosis rate of
cells increased. This may benefit the development of therapeutic
strategies for the treatment of MM.
To further explore the molecular mechanisms
underlying the effects of miR-181a on MM cell cycle, proliferation,
apoptosis, and invasion, six target genes were predicted using
various bioinformatics software. The expression of NOVA1 was
verified by RT-qPCR. Our results revealed that NOVA1 mRNA
expression was reduced in the miR-181a inhibitor group. This
opposes the known functions of miRNAs, as the mRNA expression
should be induced providing BCL-2 and NOVA1 were
targets of miR-181a. Several studies have reported that miRNAs can
activate downstream gene expression by binding to promoters to
initiate gene transcription (37–39),
which may explain our experimental results. It is also possible
that miR-181a targets and regulates upstream of NOVA1, thus
affecting its expression; the underlying mechanism requires further
investigation.
There are some limitations to the present study. Our
previous study (19) revealed that
miR-181a and miR-20a were highly expressed in the serum of patients
with MM, whereas the present study reported that miR-181a was
upregulated in MM cell lines. These results suggest that miR-181a
may be involved in the pathogenesis of myeloma; however, we failed
to isolate primary myeloma cells from patients with MM. Further
studies with primary myeloma cells are required to verify the role
of miR-181a in myeloma.
In conclusion, this research has comprehensively
studied the biological effects of miR-181a on MM in vitro
and in vivo. miR-181a was upregulated in MM cell lines, and
promoted cell proliferation, increased the ratio of S phase cells,
inhibited apoptosis and promoted tumor growth. As a tumor-promoting
factor, miR-181a may play an important role in vitro in cell
lines and in vivo in animal xenograft models. Predicting the
target genes of miR-181a may provide a basis for further
exploration into the mechanism underlying the effects of miR-181a
in MM.
Acknowledgements
Not applicable.
Funding
This work was supported by the Ministry of Science
and Technology of Shaanxi (grant no. S2016YFKW0040).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
NL performed the experiments (including RT-qPCR,
CCK-8, flow cytometry, Transwell assays and animal experiments),
analyzed the data and wrote the manuscript. JY performed the animal
experiments and analyzed the data. RY participated in RT-qPCR and
flow cytometry analysis. JP and LL were involved in the CCK-8 and
Transwell assays. XG conceived the idea, designed the study and
helped to revise the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Prior written and informed consent were obtained
from every patient, and the study was approved by the ethics review
board of The First Affiliated Hospital of Xi'an Jiaotong University
(approval no. XJTU1AF2016LSK-46). All experimental procedures
involving animals were conducted according to the ethical
guidelines of The First Affiliated Hospital of Xi'an Jiaotong
University. Ethical approval was received for the use of animals,
prior to the start of the study (grant no. XJTULAC2017-782).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Li Y, Bai O, Liu C, Du Z, Wang X, Wang G
and Li W: Association between hepatitis B virus infection and risk
of multiple myeloma: A systematic review and meta-analysis. Intern
Med J. 46:307–314. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Palumbo A and Anderson K: Multiple
myeloma. N Engl J Med. 364:1046–1060. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dürner J, Reinecker H and Csef H:
Individual quality of life in patients with multiple myeloma.
Springerplus. 2:3972013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nooka AK, Kastritis E, Dimopoulos MA and
Lonial S: Treatment options for relapsed and refractory multiple
myeloma. Blood. 125:3085–3099. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Patriarca F, Fanin R, Silvestri F, Damiani
D and Baccarani M: Autologous stem cell transplantation in multiple
myeloma: A single center experience. Haematologica. 83:477–479.
1998.PubMed/NCBI
|
|
6
|
Zhao Z, Ma X, Sung D, Li M, Kosti A, Lin
G, Chen Y, Pertsemlidis A, Hsiao TH and Du L: microRNA-449a
functions as a tumor suppressor in neuroblastoma through inducing
cell differentiation and cell cycle arrest. RNA Biol. 12:538–554.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ambros V: The functions of animal
microRNAs. Nature. 431:350–355. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
435:834–838. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Michael MZ, O'Connor SM, van Holst
Pellekaan NG, Young GP and James RJ: Reduced accumulation of
specific microRNAs in colorectal neoplasia. Mol Cancer Res.
1:882–891. 2003.PubMed/NCBI
|
|
10
|
Tutar Y: miRNA and cancer; Computational
and experimental approaches. Curr Pharm Biotechnol. 15:4292014.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Deb B, Uddin A and Chakraborty S: miRNAs
and ovarian cancer: An overview. J Cell Physiol. 233:3846–3854.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen CZ, Li L, Lodish HF and Bartel DP:
MicroRNAs modulate hematopoietic lineage differentiation. Science.
303:83–86. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu G, Li Y and Gao XG: microRNA-181a is
upregulated in human atherosclerosis plaques and involves in the
oxidative stress-induced endothelial cell dysfunction through
direct targeting Bcl-2. Eur Rev Med Pharmacol Sci. 20:3092–3100.
2016.PubMed/NCBI
|
|
14
|
Cao Y, Zhao D, Li P, Wang L, Qiao B, Qin
X, Li L and Wang Y: MicroRNA-181a-5p impedes IL-17-induced nonsmall
cell lung cancer proliferation and migration through targeting
VCAM-1. Cell Physiol Biochem. 42:346–356. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu Z, Sun F, Hong Y, Liu Y, Fen M, Yin K,
Ge X, Wang F, Chen X and Guan W: MEG2 is regulated by miR-181a-5p
and functions as a tumour suppressor gene to suppress the
proliferation and migration of gastric cancer cells. Mol Cancer.
16:1332017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Su R, Lin HS, Zhang XH, Yin XL, Ning HM,
Liu B, Zhai PF, Gong JN, Shen C, Song L, et al: MiR-181 family:
Regulators of myeloid differentiation and acute myeloid leukemia as
well as potential therapeutic targets. Oncogene. 34:3226–3239.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lyu X, Li J, Yun X, Huang R, Deng X, Wang
Y, Chen Y and Xiao G: miR-181a-5p, an inducer of Wnt-signaling,
facilitates cell proliferation in acute lymphoblastic leukemia.
Oncol Rep. 37:1469–1476. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xiang T, Hu AX, Sun P, Liu G, Liu G and
Xiao Y: Identification of four potential predicting miRNA
biomarkers for multiple myeloma from published datasets. PeerJ.
5:e28312017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Peng J, Thakur A, Zhang S, Dong Y, Wang X,
Yuan R, Zhang K and Guo X: Expressions of miR-181a and miR-20a in
RPMI 8226 cell line and their potential as biomarkers for multiple
myeloma. Tumour Biol. 36:8545–8552. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shen B, Zhang Y, Yu S, Yuan Y, Zhong Y, Lu
J and Feng J: MicroRNA-339, an epigenetic modulating target is
involved in human gastric carcinogenesis through targeting NOVA1.
FEBS Lett. 589:3205–3211. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yuan R, Liu N, Yang J, Peng J, Liu L and
Guo X: The expression and role of miR-181a in multiple myeloma.
Medicine (Baltimore). 97:e120812018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tassone G and Fidler SJ: Separation and
cryopreservation of lymphocytes from spleen and lymph node. Methods
Mol Biol. 882:351–357. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xin Y, Li Z, Zheng H, Ho J, Chan MTV and
Wu WKK: Neuro-oncological ventral antigen 1 (NOVA1): Implications
in neurological diseases and cancers. Cell Prolif. 50:2017.
View Article : Google Scholar :
|
|
25
|
Zhang YA, Liu HN, Zhu JM, Zhang DY, Shen
XZ and Liu TT: RNA binding protein Nova1 promotes tumor growth in
vivo and its potential mechanism as an oncogene may due to its
interaction with GABAA Receptor-g2. J Biomed Sci.
23:712016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Zhou H and Wu L: The development and
function of dendritic cell populations and their regulation by
miRNAs. Protein Cell. 8:501–513. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang Y, Yang P and Wang XF:
Microenvironmental regulation of cancer metastasis by miRNAs.
Trends Cell Biol. 24:153–160. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shin VY and Chu KM: MiRNA as potential
biomarkers and therapeutic targets for gastric cancer. World J
Gastroenterol. 20:10432–10439. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Manikandan J, Aarthi JJ, Kumar SD and
Pushparaj PN: Oncomirs: The potential role of non-coding microRNAs
in understanding cancer. Bioinformation. 2:330–334. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Miska EA: How microRNAs control cell
division, differentiation and death. Curr Opin Genet Dev.
15:563–568. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yu W, Li L, Zheng F, Yang W, Zhao S, Tian
C, Yin W, Chen Y, Guo W, Zou L and Deng W: β-catenin cooperates
with CREB binding protein to promote the growth of tumor cells.
Cell Physiol Biochem. 44:467–478. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang M, Zhai X, Ge T, Yang C and Lou G:
miR-181a-5p promotes proliferation and invasion and inhibits
apoptosis of cervical cancer cells via regulating inositol
polyphosphate-5-phosphatase A (INPP5A). Oncol Res. 26:703–712.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Han P, Li JW, Zhang BM, Lv JC, Li YM, Gu
XY, Yu ZW, Jia YH, Bai XF, Li L, et al: The lncRNA CRNDE promotes
colorectal cancer cell proliferation and chemoresistance via
miR-181a-5p-mediated regulation of Wnt/β-catenin signaling. Mol
Cancer. 16:92017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Arora S and Tandon S: Achyranthes aspera
root extracts induce human colon cancer cell (COLO-205) death by
triggering the mitochondrial apoptosis pathway and S phase cell
cycle arrest. ScientificWorldJournal. 2014:1296972014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Janowski BA, Younger ST, Hardy DB, Ram R,
Huffman KE and Corey DR: Activating gene expression in mammalian
cells with promoter-targeted duplex RNAs. Nat Chem Biol. 3:166–173.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li LC, Okino ST, Zhao H, Pookot D, Place
RF, Urakami S, Enokida H and Dahiya R: Small dsRNAs induce
transcriptional activation in human cells. Proc Natl Acad Sci USA.
103:17337–17342. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Place RF, Li LC, Pookot D, Noonan EJ and
Dahiya R: MicroRNA-373 induces expression of genes with
complementary promoter sequences. Proc Natl Acad Sci USA.
105:1608–1613. 2008. View Article : Google Scholar : PubMed/NCBI
|