Introduction
The multidrug resistance toward anticancer drugs
caused by efflux transporters, such as multidrug
resistance-associated protein (MRP/ABCC), P-glycoprotein
(P-gp/ABCB1), and breast cancer resistance protein (BCRP/ABCG2), is
a barrier to effective chemotherapy. The increased activities of
these transporters accompanying the malignant progression of tumor
cells results in the reduction of intracellular drug concentrations
in the tumors. The plasma membrane localization of efflux
transporters is important for transport activity, as well as the
expression levels of efflux transporters. The plasma membrane
localization of efflux transporters is governed by scaffold
proteins, such as the ezrin/radixin/moesin (ERM) proteins. For
example, the interaction of MRP2 with ezrin and/or radixin causes
the localization of MRP2 in the canalicular membrane in hepatocytes
(1–3). Ezrin and moesin are involved in the
plasma membrane localization of P-gp in brain capillary cells
(4,5). ERM proteins activated via
phosphorylation by Rho-kinase (6)
and cholecystokinin-8 (7), function
as anchoring proteins for efflux transporters and
Na+-H+ exchanger regulatory factor 1
(8). Some reports have shown that
ERM proteins are involved in the localization of transporters in
tumor cells, as well as in normal tissues (9–11).
Thus, reduction of the anchor function of ERM
proteins in tumor cells may decrease the transport activity of
efflux transporters, possibly leading to increased intracellular
accumulation and efficiency of anticancer drugs as substrates of
efflux transporters. However, it is unclear whether ERM proteins
can be used as target molecules for overcoming multidrug resistance
in tumor cells.
Therefore, the effect of the knockdown of ERM
proteins on the transport activities of MRP2, P-gp and BCRP was
examined in multiple carcinoma cell lines, and on the intracellular
accumulation and efficiency of methotrexate (MTX) as an anticancer
drug.
Materials and methods
Ethical approval
The study protocol was approved by the Committee for
the Care and Use of Laboratory Animals of the School of Pharmacy of
Kindai University (Osaka, Japan).
Chemicals and reagents
Silencer Select siRNA for human ezrin (s14796),
human radixin (s11899), human moesin (s8984), and Stealth RNAi
siRNA negative control, Lipofectamine RNAiMAX Transfection Reagent,
Opti-MEM I, BCA protein assay kit, Mem-PER Eukaryotic Membrane
Protein Extraction Kit, and D-luciferin monosodium salt were
obtained from Thermo Fisher Scientific, Inc. (Waltham, MA, USA).
Rabbit monoclonal anti-rat ezrin (EP886Y; cat. no. ab40839), rabbit
monoclonal anti-radixin antibody (EP1862Y; cat. no. ab52495) and
rabbit monoclonal anti-moesin antibody (EP1863Y. cat. no. ab52490)
(all from Abcam, Cambridge, UK), mouse monoclonal anti-β-actin
antibody (cat. no. AM00194PU-N; Acris Antibodies GmbH, Herford,
Germany), peroxidase-labeled anti-rabbit IgG antibody (cat. no.
ab214880) and peroxidase-labeled anti-mouse IgG antibody (cat. no.
ab97040; both from Abcam), and peroxidase-labeled anti-mouse IgG
antibody (cat. no. 5220-0341; KPL, Gaithersburg, MD, USA) were
commercially obtained. For assay of transport activities,
5(6)-carboxy-2′,7′-dichlorofluorescein diacetate promoiety (CDFDA,
a precursor of MRP substrate (CDF); Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany), MTX (an MRP substrate) and Rhodamine 123
(R123, a P-gp substrate; both from Wako Pure Chemical Industries,
Osaka, Japan), Hoechst 33342 hydrochloride (H33342, a BCRP
substrate; Cayman Chemical, Ann Arbor, MI, USA), MK571 sodium salt
hydrate (an MRP inhibitor; Sigma-Aldrich; Merck KGaA), verapamil
(Nacalai Tesque, Inc., Kyoto, Japan) and Ko143 (MedChemExpress,
Monmouth Junction, NJ, USA) were used. RIPA lysis buffer, including
protease inhibitors, was obtained from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA). For LC-MS/MS-based targeted proteomics, MS
grade porcine pancreatic trypsin and OASIS HLB were obtained from
Wako Pure Chemicals and Waters Corporation (Milford, MA, USA),
respectively. Sepasol RNA I Super G and MTT cell count kits were
purchased from Nacalai Tesque, Inc. In vivo jetPEI was
obtained from Polyplus-transfection® (New York, NY,
USA). ECL Prime blocking reagent, Hybond-P polyvinylidene
difluoride membranes and the ECL Prime Western Blotting Detection
system were purchased from GE Healthcare (Milwaukee, WI, USA). All
other chemicals and solvents were of the highest purity
commercially available or of mass spectrometry grade.
Cell culture and transfection of
siRNA
Human liver cancer HepG2 cells, human lung carcinoma
A549 cells, and human breast carcinoma MDA-MB-453 cells were
obtained from the RIKEN Cell Bank (Ibaraki, Japan). Cells were
maintained in Dulbecco's modified Eagle's medium (DMEM; Nacalai
Tesque, Inc.) supplemented with 10% fetal bovine serum (FBS; 100
U/ml penicillin, and 100 µg/ml streptomycin at 37°C in the presence
of 5% CO2 and 95% air.
Cells were seeded at 1×105 cells/well
onto a 24-well plate (Sumitomo Bakelite Co., Ltd., Tokyo, Japan),
and were transfected with siRNA (siEzrin, siRadixin, siMoesin, or
siNegative, 5 pmol/well) complexed with Lipofectamine RNAiMAX in
Opti-MEM. The medium was replaced with fresh DMEM 48 h after
transfection and the cells were used in the downstream
experiments.
Evaluation of siRNA knockdown by
real-time reverse transcription polymerase chain reaction (RT-PCR)
and western blotting
mRNA and protein expression levels were assessed
using RT-PCR and western blotting, as previously described
(12,13). Cells treated with siRNA for 48 h
were washed twice with cold phosphate-buffered saline (PBS). Total
RNA was extracted from each sample and reverse-transcribed to
complementary DNA. PCRs were incubated at 95°C for 10 sec, and then
amplified at 95°C for 5 sec, 55°C for 20 sec, and 72°C for 31 sec
for 40 cycles. After RIPA buffer was added to the cells, the cells
were collected by cell scrapers. Cells were sonicated for 30 sec
and were centrifuged at 14,000 × g for 15 min at 4°C. The protein
concentrations in the supernatant were determined using a BCA
protein assay kit. Sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) was performed using 7.5% e-Pagell (ATTO
Corp., Tokyo, Japan) and 20 µg of proteins/well. Resolved proteins
were transferred onto Hybond-P polyvinylidene difluoride membranes
and subjected to semi-dry blotting using a Trans-blot SD (Bio-Rad
Laboratories, Hercules, CA, USA). After a membrane blocking in a 5%
(w/v) solution of ECL Prime blocking reagent in PBS for 1 h at room
temperature, immunoreactive ezrin, radixin, moesin, and β-actin
were detected using antibodies, and an ECL Prime Western Blotting
Detection system. Primary antibodies and secondary antibodies were
used for immunoblotting at a 1:1,000 and 1:10,000 dilution for 1 h
each, respectively. The densitometric intensity of each band was
detected with Ez capture and was calculated using CS analyzer 3
(both from ATTO Corp.).
Transporter activity assays
CDFDA (10 µM), MTX (100 µM), R123 (50 µM) and H33342
(10 µM) with or without each inhibitor (50 µM MK571, 50 µM
verapamil and 10 µM Ko143) were added 30 min before the cells were
treated with siRNA for 48 h. All transporter substrates were
initially dissolved in dimethyl sulfoxide (DMSO). The final
concentration of DMSO in the medium was <0.5%. After a 1
h-incubation for CDFDA, MTX and H33342 and a 2 h-incubation for
R123, the cells were washed twice with PBS. Then, 100 µl of 1 M
NaOH was added to lyse the cells, and the fluorescence intensity in
the cells was measured using a fluorescence microplate reader
(SH-9000; Corona Electric Co., Ibaraki, Japan) at Ex/Em wavelengths
of 495/530 nm for CDF, 480/530 nm for R123, and 355/460 nm for
H33342. The determination of MTX concentrations in the cell lysates
was performed by liquid chromatography-tandem mass spectrometry
(LC-MS/MS) as previously described (14). The LC-MS/MS equipment consisted of
an LC system (UltiMate 3000 series) and a TSQ Endura Triple
Quadrupole Mass Spectrometer with electrospray ionization (both
from Thermo Fisher Scientific, Inc.). For data recording and
analysis, Finnigan XCalibur software version 3 (Thermo Fisher
Scientific, Inc.) was used.
Determination of the expression of
MRP2, P-gp and BCRP in the cytoplasm and plasma membrane by
targeted proteomics
Cytoplasm and plasma membrane proteins were
extracted from HepG2 cells transfected with siRNA using a Mem-PER
Eukaryotic Membrane Protein Extraction Kit. Proteins (10 µg)
diluted in 25 µl of phase transfer surfactant buffer were reduced
with 10 µM dithiothreitol at room temperature for 30 min. Free
cysteines were alkylated in 42 µM iodoacetamide at room temperature
for 20 min. A total of 50 µM ammonium bicarbonate and 1 µg of
trypsin was added to the cell lysates. After trypsin digestion at
37°C for 18 h, 10 pmol of isotope-labeled peptides were added. The
digested samples were desalted by OASIS HLB. The eluted samples
were dried under vacuum at 50°C and were resuspended in 100 µl of
the initial mobile phase (4.5% acetonitrile with 0.1% formic acid).
Peptides were separated on a reversed-phase column (AdvanceBio
Peptide Map C18, 2.1×150 µm, 2.7 µm; Agilent Technologies, Inc.,
Santa Clara, CA, USA). The column temperature was set at 50°C and
the autosampler was maintained at 4°C. The mobile phase (A: 0.1%
formic acid in water; B: 0.1% formic acid in 90% acetonitrile) was
pumped at a flow rate of 0.1 µl/min. The column was conditioned at
5% mobile phase B. From 2 to 26 min, mobile phase B increased
linearly from 5 to 60%. From 26 to 27 min, mobile phase B increased
linearly to 98%. From 28 to 35 min, the mobile phase was maintained
at 5% mobile phase B. The peptide sequences of surrogate peptides
determined in this study were as follows; LTIIPQDPILFSGSLR for
MRP2, IATEAIENFR for P-gp, SSLLDVLAAR for BCRP, DLTDYLMK for
β-actin, and IVEIPFNSTNK for Na+/K+ ATPase.
The protein expression levels of MRP2, P-gp and BCRP in the
cytoplasm and plasma membrane proteins were corrected by the
intensities of surrogate marker peptides of β-actin and
Na+/K+-ATPase, respectively (15–17).
Evaluation of the cytotoxicity of MTX
by MTT assay
MTX (100 µM) was added to HepG2 cells treated with
siRadixin or siNegative and incubated for 48 h. At 24 h after MTX
treatment, 10 µl of MTT cell count kit was added to the HepG2 cells
for 3 h. The absorbance was measured at 570 nm by Sunrise R (Tecan
Group, Ltd., Männedorf, Switzerland).
Inoculation of tumor cells
Male BALB/c nu/nu (4-week-old, body
weight 15–17 g) mice were purchased from Japan SLC (Shizuoka,
Japan). The animals were housed in an air-conditioned room at
22±0.5°C and relative air humidity of 55±10% with a 12-h lighting
schedule (7:00 a.m.-7:00 p.m.) and had free access to standard
laboratory food (MF; Oriental Yeast Co., Ltd., Tokyo, Japan) and
water on the light/dark schedule for a week before they were
divided into each group. The total number of mice used in the
present study was 67.
Firefly luciferase-expressing HepG2 cells
(HepG2-Luc; JCRB Cell Bank, Tokyo, Japan) were trypsinized and
suspended in Hank's buffered salt solution (HBSS). The cells
(1×105 cells in HBSS) were inoculated by subcutaneous
injection into the back of the mice.
Determination of the mRNA levels of
radixin in tumor-bearing mice by real-time reverse transcription
polymerase chain reaction (RT-PCR)
Total RNA was extracted from the mouse tumors using
Sepasol I super G, 48 h after treatment with 10 µg of siRadixin or
siNegative, complexed with in vivo jetPEI (N/P=6). The mRNA
expression was assessed using RT-PCR, as previously described
(18,19). The following primer sequences were
used: Sense primer, 5′-TTTGGCTTCGTATGCTGTCC-3′ and antisense
primer, 5′-TGTTCCAAAACACGCTGTGG-3′ for mouse radixin and sense
primer, 5′-CATTGCTGACAGGATGCAGAA-3′ and antisense primer,
5′-CCGATCCACACAGAGTACTTGC-3′ for mouse β-actin. Data were analyzed
using the StepOne Real-Time PCR System (Thermo Fisher Scientific,
Inc.), using the multiplex comparative method, as previously
reported (20). The amount of
target mRNA was normalized to β-actin as the internal control.
Accumulation of MTX in the tumors of
tumor-bearing mice
HepG2-Luc suspended in HBSS was inoculated by
subcutaneous injection into the back of the mice (1×105
cells/mice). At 1 week after inoculation of HepG2-Luc cells, the
mice received injections of MTX using a 27-gauge needle for 10 sec
at 30 µg/kg into the tumors (the long axis at ~4 µm in tumor
diameter) 48 h after treatment with 10 µg of siRadixin or
siNegative, complexed with in vivo jetPEI (N/P=6). At 1 h
after MTX treatment, the tumors were excised from mice euthanized
by cervical dislocation and the blood samples were collected in
heparinized tubes, and immediately centrifuged at 6,700 × g for 10
min at 4°C to obtain plasma. The MTX levels in the tumors and
plasma were determined by LC-MS/MS, as previously described
(14). The ratio of the MTX level
in the tumor to that in the plasma was used to indicate the
accumulation of MTX in the tumor.
IVIS imaging
The IVIS imaging was performed according to the
procedure of Puaux et al (21). Mice were injected intraperitoneally
with D-luciferin at a dose of 150 µg/kg, 1, 2 and 3 weeks after
inoculation with HepG2-Luc cells. The mice were anesthetized with
isoflurane and placed on the imaging stage of the IVIS apparatus in
the abdominal position with a continuous supply of isoflurane
through the gas anesthesia manifold. Images were collected 15 min
after D-luciferin injection using the IVIS Lumina XRMS Imaging
System and photons emitted from the tumor and its surroundings were
quantified using Living Image Software version 4 (both from
PerkinElmer, Inc., Waltham, MA, USA).
Statistical analysis
The significance of differences between mean values
was determined by Dunnett's test after analysis of variance (ANOVA)
using GraphPad Prism software version 5 (GraphPad Software, Inc.,
La Jolla, CA, USA). Values of P<0.05 were considered to indicate
statistically significant differences.
Results
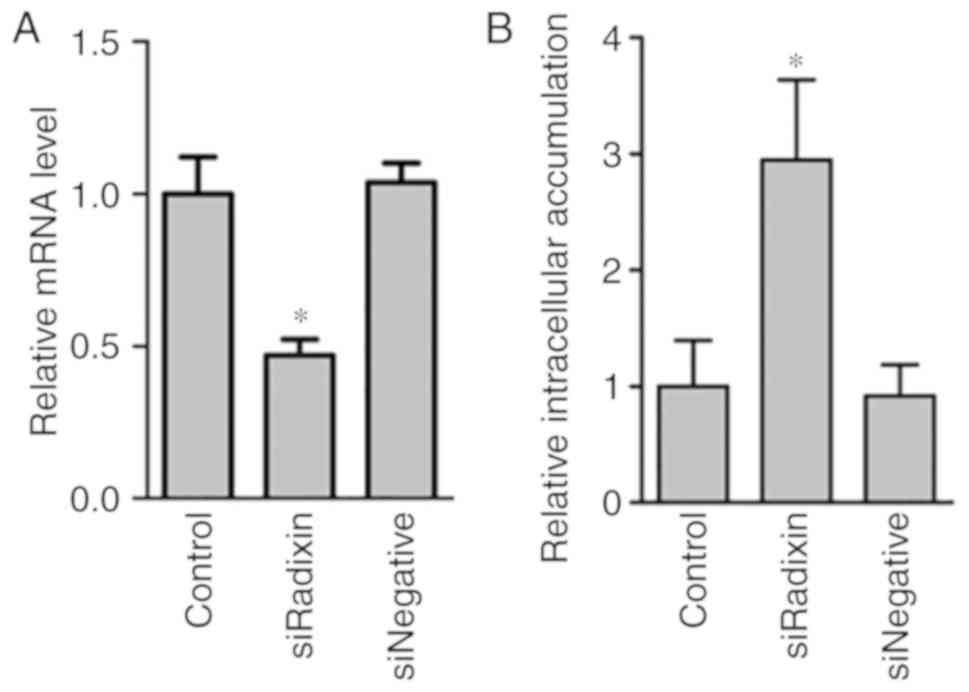
Efficiency of the knockdown of the ERM
proteins in HepG2 cells by siRNA transfection
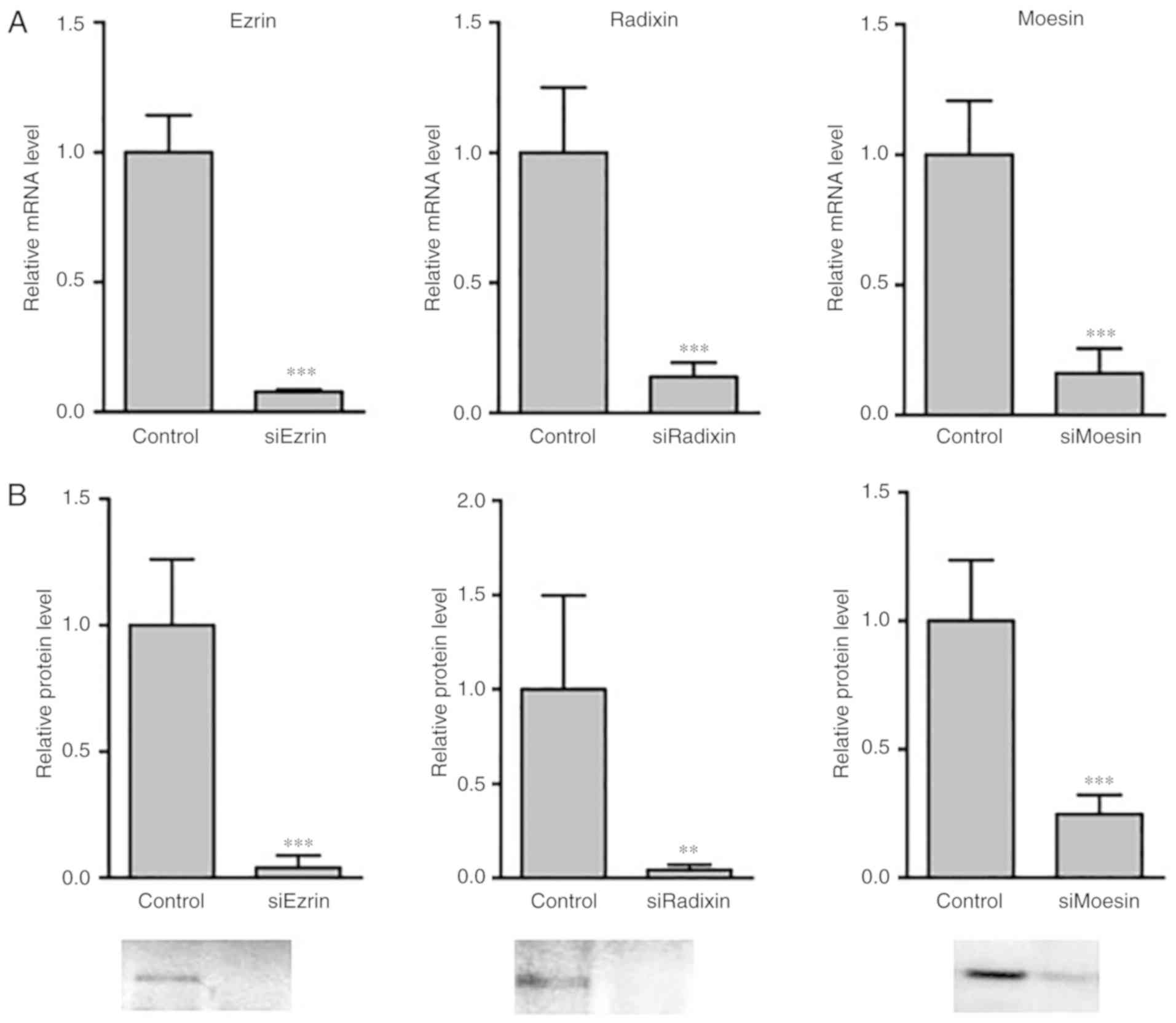
To confirm the efficiency of the knockdown of the
ERM proteins in HepG2 cells by the siRNA, the mRNA and protein
expression levels in the cell lysate were determined 48 h after
siRNA transfection (Fig. 1). The
knockdown of ERM proteins by siRNA was observed in the mRNA levels
(Fig. 1A). The protein levels of
the ERM proteins were significantly decreased by siRNA knockdown
(Fig. 1B).
Efflux activity of MRP, P-gp, and BCRP
in ERM protein-knockdown cells
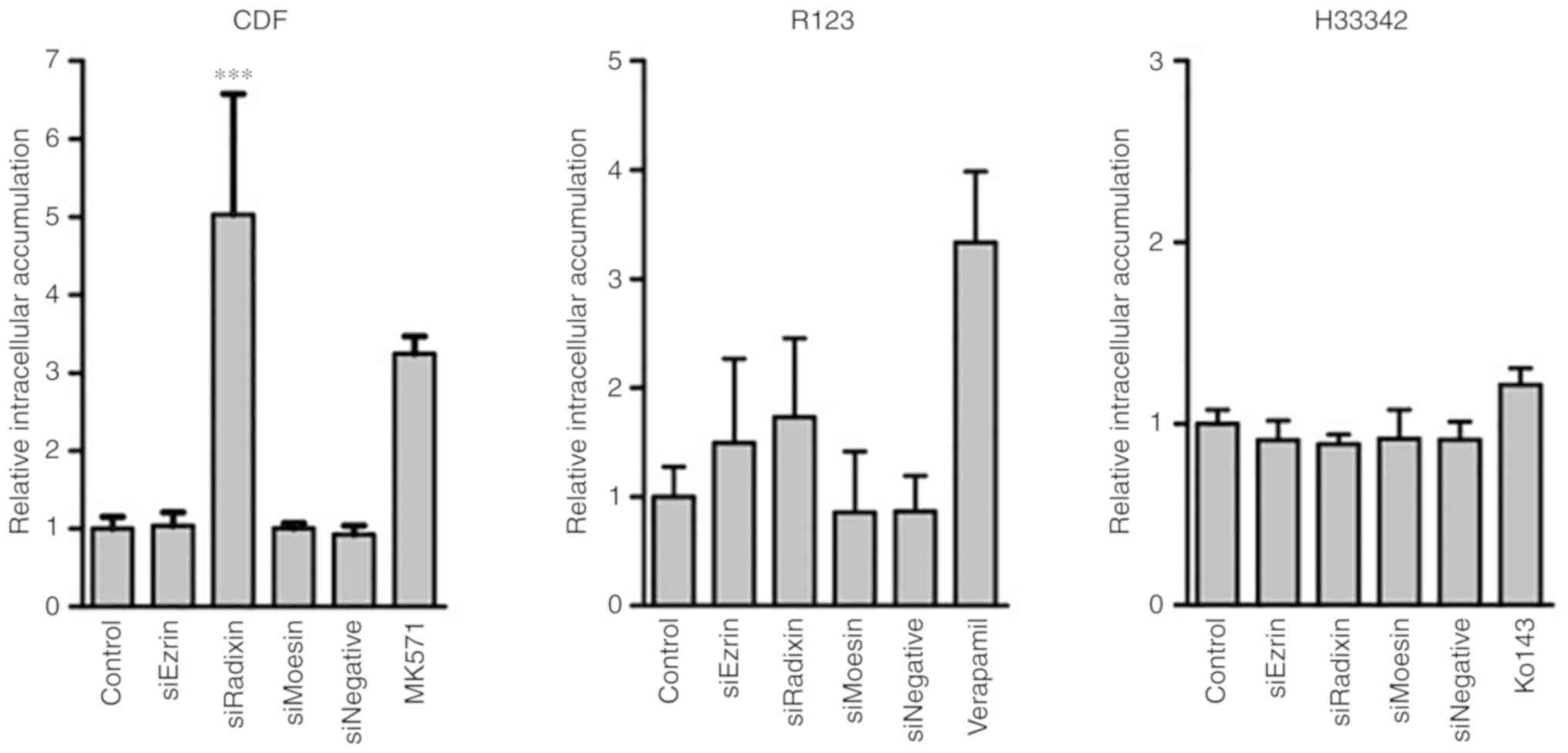
To clarify the effect of ERM protein knockdown on
the transport activity of MRP, P-gp and BCRP, the intracellular
fluorescence intensities of transporter substrates in HepG2 cells
with ERM protein knockdown were determined (Fig. 2). The knockdown of radixin, but not
ezrin or moesin, resulted in a significantly higher intracellular
accumulation of CDF compared with control HepG2 cells without ERM
protein knockdown. There was little change in the intracellular
accumulation of R123 and H33342 by ERM protein knockdown. Whether
the effects of radixin knockdown on the intracellular accumulation
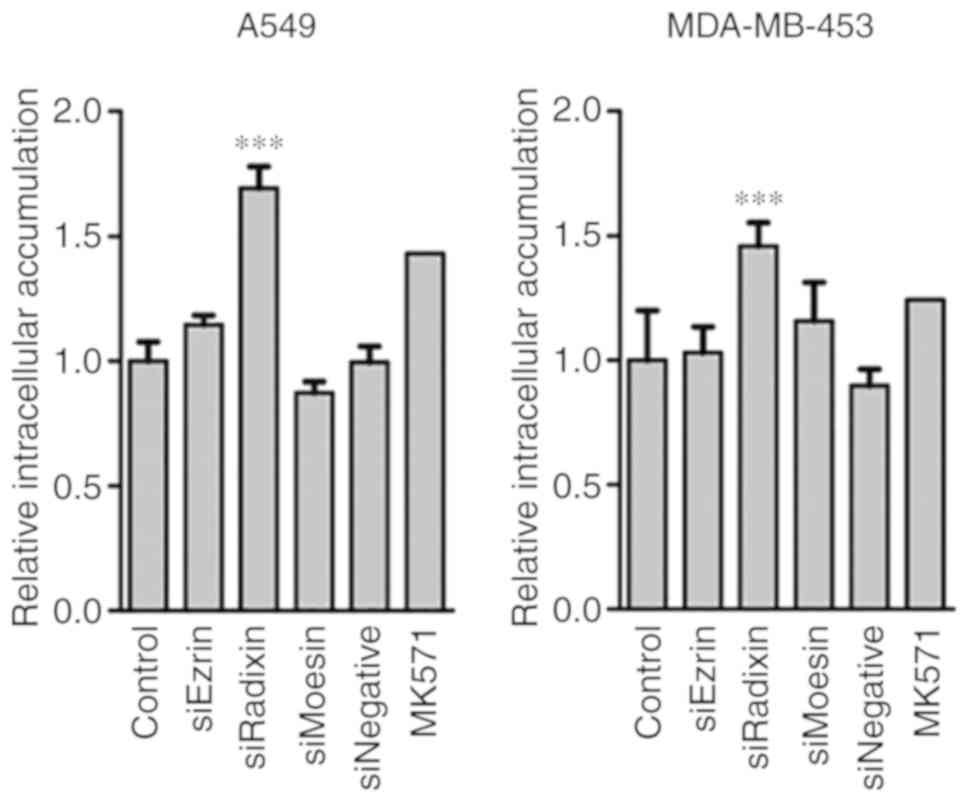
of CDF were distinct to HepG2 cells (Fig. 3) was revealed. The radixin knockdown
in A549 and MDA-MB-453 cells led to significant increases in
intracellular CDF accumulation.
Levels of MRP2, P-gp and BCRP in
radixin-knockdown cells
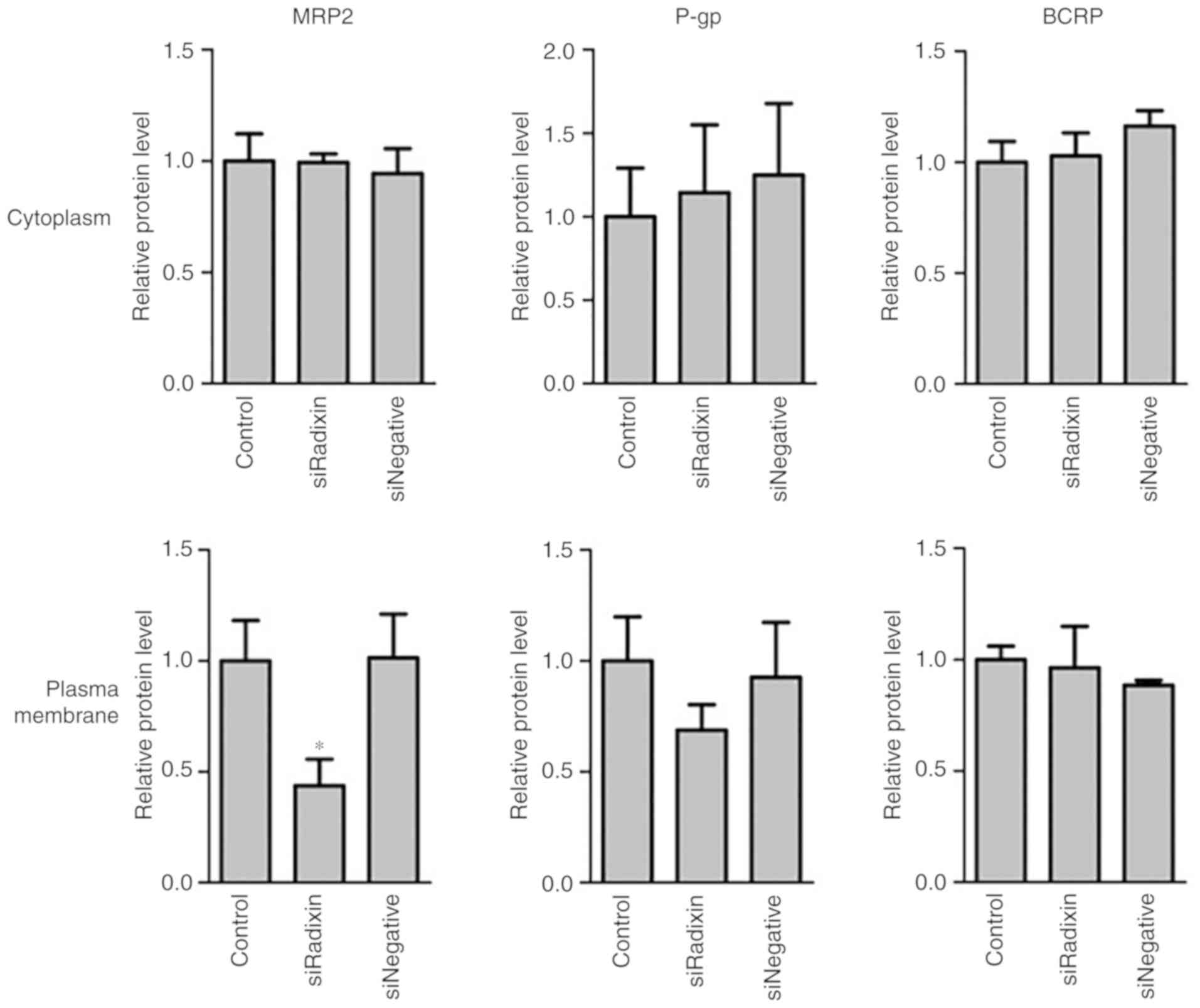
The levels of MRP2, P-gp, and BCRP in the cytoplasm
and plasma membrane of HepG2 cells with radixin knockdown were
determined (Fig. 4). The levels of
MRP2, but not P-gp or BCRP, in the plasma membrane were
significantly decreased by radixin knockdown (Fig. 4). In the cytoplasm, the levels of
MRP2, P-gp, and BCRP were unaltered by radixin knockdown.
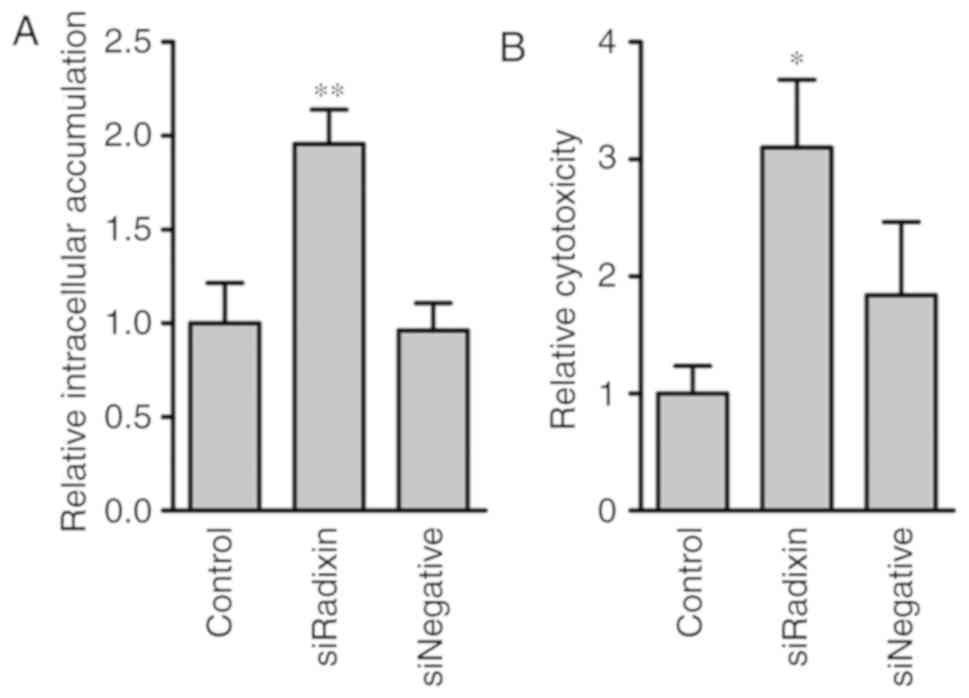
Intracellular MTX accumulation and
cytotoxicity in radixin-knockdown cells
The effect of radixin knockdown on intracellular MTX
accumulation and cytotoxicity was examined (Fig. 5). The intracellular MTX accumulation
was significantly increased by radixin knockdown compared with
control and siNegative (Fig. 5A).
The MTX cytotoxicity was also increased by radixin knockdown
(Fig. 5B).
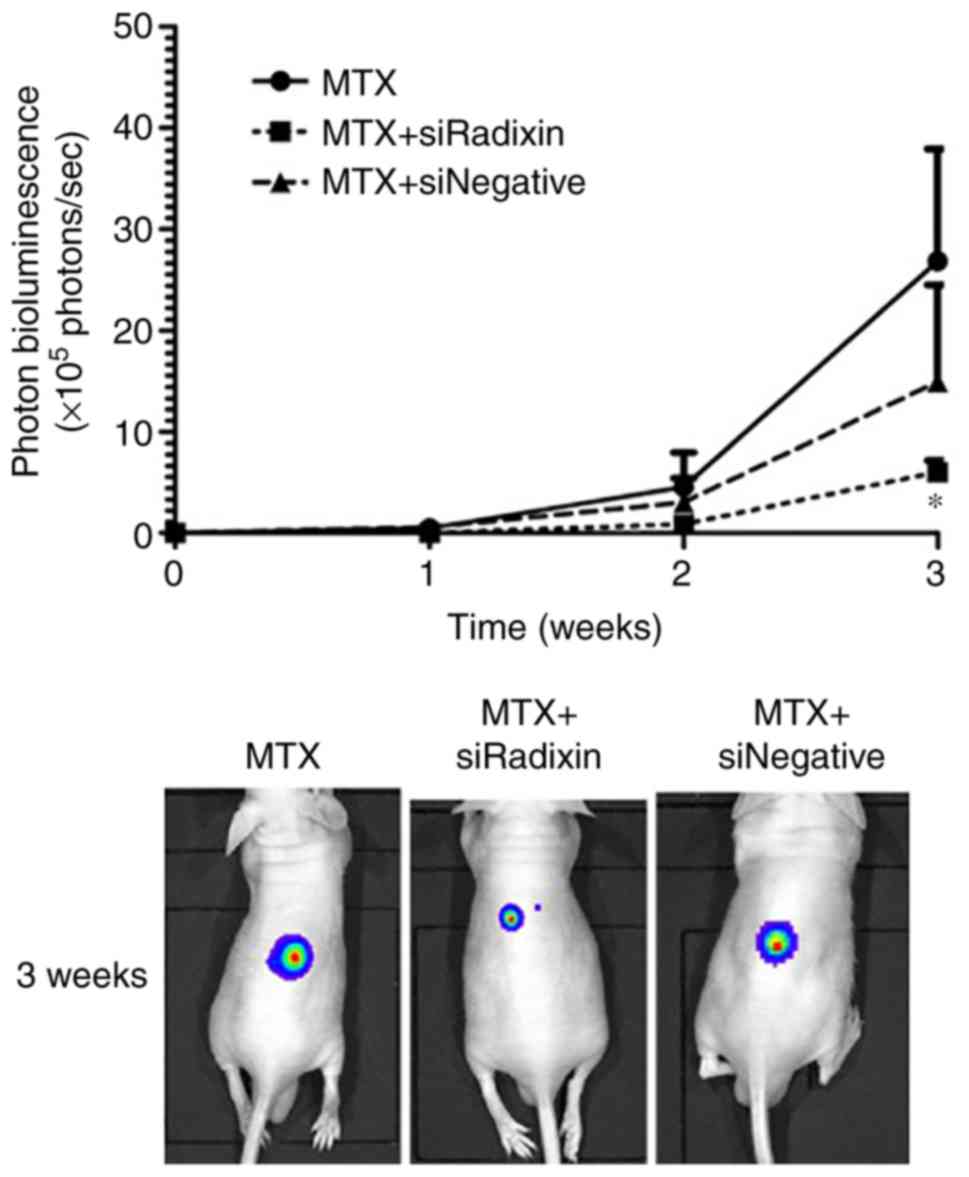
In vivo effects of radixin knockdown
on the accumulation and efficiency of MTX
The in vivo knockdown efficiency of radixin
in tumors after siRadixin treatment was determined (Fig. 6A). Significant decreases in radixin
mRNA were observed in tumors, as well as in in vitro
knockdown experiments. To clarify whether the treatments of
siRadixin caused the topical knockdown of radixin, the radixin mRNA
levels in liver were determined. The radixin mRNA levels in liver
were unaltered between the control and siRadixin 48 h after
intratumoral injection of siRNA complexed with in vivo
jetPEI (data not shown). To clarify whether the radixin knockdown
affected the accumulation of MTX in the tumors, the levels of MTX
remaining in the tumors 1 h after MTX administration were
determined (Fig. 6B). Higher
concentrations of MTX were observed in the tumors of mice with
radixin knockdown, although the plasma concentrations of MTX were
unchanged by treatment of siRadixin. The anticancer effects of MTX
after radixin knockdown were examined using in vivo imaging
(Fig. 7). Tumor proliferation in
the mice was suppressed by radixin knockdown. A significant
decrease in the photon bioluminescence derived from HepG2-Luc cells
was observed 3 weeks after inoculation of mice with HepG2-Luc
cells. MTX alone revealed a lower anticancer effect compared with
MTX with siRadixin. There was little effect with either siRadixin
alone (20.8×105 photons/sec, n=2) or in vivo
jetPEI alone (22.8×105 photons/sec, n=2) on the relative
intensity of the photon bioluminescence 3 weeks after inoculation
of mice with HepG2-Luc cells.
Discussion
It was unclear whether ERM proteins could be target
proteins for the modulation of efflux transporter activity in tumor
cells, although ERM proteins have important roles in the regulation
of transporter localization and activity in normal tissues. In the
liver, radixin deficiency causes hyperbilirubinemia with loss of
MRP2 from bile canalicular membranes (1). The plasma membrane localization and
transport activity of MRP2 were inhibited by radixin knockdown in
HepG2 cells (Figs. 2 and 4). An in vivo study revealed the
potential of radixin knockdown in tumor cells to improve the
efficiency of MTX (Fig. 7). Radixin
knockdown could promote the MRP2 internalization from the plasma
membrane surface. However, the changes of absolute amounts of MRP2
in HepG2 cells remains unclear, because the protein expression
levels were represented by relative amounts for control expression
levels.
ERM proteins are involved in the distinct regulation
of the localization and/or translation of transporters in normal
tissues. For example, radixin knockout resulted in the reduction of
P-gp expression and transport activity in the small intestine
(22). Activation of ezrin led to
apical localization of MRP2 in the small intestine (23). Radixin deficiency caused a loss of
MRP2 from bile canalicular membranes (1). In the present study, it was examined
whether ERM proteins participated in the plasma membrane
localization and transport activity of transporters in tumor cells.
The transport activity of MRP2 was significantly decreased by
radixin knockdown in HepG2, A549, and MDA-MB-453 cells (Figs. 2 and 3), indicating that radixin is involved in
the MRP2 function in tumor cells as well as in normal cells.
Pokharel et al revealed that ERM proteins are involved in
the regulation of the expression and function of P-gp in breast
cancer cells (24). Yano et
al demonstrated that the role of ERM proteins in the regulation
of P-gp was different in Caco-2 and Caki-1 cells (11), indicating that the tissue-specific
participation of ERM proteins in the regulation of P-gp in tumor
cells was maintained as in the corresponding normal tissues. In
HepG2 cells, the intracellular CDF accumulation was particularly
increased by radixin knockdown compared with A549 cells and
MDA-MB-453 cells, indicating a marked higher expression of MRP2
and/or higher interaction between radixin and MRP2 in the liver,
because the inhibition of MRP activity by MK571 was insufficient in
A549 and MDA-MB-453 cells. In addition to the regulation of the
plasma membrane localization of transporters, ezrin was revealed to
regulate the translational process of P-gp in HepG2 cells (25). However, in tumor cells, the
participation of ezrin and moesin in the transport activity of
MRP2, P-gp and BCRP may be reduced compared with radixin.
Multiple MRPs, including MRP1, MRP2 and MRP3,
exhibit resistance to MTX exposure (26,27).
Both the MRP2 activity and expression levels in the plasma membrane
of HepG2 cells were significantly decreased by radixin knockdown.
In preliminary experiments, the expression of MRP1 and MRP3 in the
cytoplasm and plasma membrane were unaltered by radixin knockdown
(data not shown). He et al demonstrated that radixin
knockdown in human gastric carcinoma SGC-7901 cells led to the
specific knockdown of MRP2, not MRP1 or MRP3 (9). These findings were consistent with the
decreased expression and activity of MRP2, compared with other MRP
family members, after radixin knockdown in HepG2 cells.
There was little effect of ERM protein knockdown on
the transport activity and expression levels of P-gp and BCRP in
HepG2 cells (Figs. 2 and 4). In HepG2 cells, it was difficult to
determine alterations in BCRP activity, because the intracellular
accumulation of H33342 was slightly increased by Ko143. An improved
evaluation system for BCRP activity is required to clarify the
effect of ERM protein knockdown on the interactions between BCRP
and ERM proteins. There was little effect of radixin knockdown on
the expression of P-gp and BCRP in the cytoplasm and plasma
membrane (Fig. 4). The changes in
transporter expression by radixin knockdown corresponded with the
alterations in the transport activity. Kano et al
demonstrated that radixin knockdown in HepG2 cells decreased the
protein expression of P-gp in plasma membrane (25). In our results, the activities of
P-gp and the protein levels of P-gp in plasma membrane were
tendency decreased, not statistically significant, although the
detailed reasons for these differences remains unclear.
Based on the results of transport activity using
typical fluorescent substrates for efflux transporters, radixin has
potential as a target protein to improve the delivery efficiency of
anticancer drugs recognized by MRP2. To clarify the effects of
radixin knockdown on the efficiency of an anticancer drug in
vitro and in vivo, the accumulation and efficiency of
MTX as a typical anticancer drug which is an MRP substrate were
examined (26–31). The intracellular accumulation of MTX
was significantly increased by radixin knockdown (Figs. 5A and 6B). Increased cytotoxicity in HepG2 cells
treated with MTX was observed with radixin knockdown (Fig. 5B), indicating that the reduction of
MRP2 activity in HepG2 cells led to an increase in the anti-cancer
effects of MTX. In an in vivo study, tumor proliferation
after inoculation of HepG2-Luc cells to mice was significantly
delayed by radixin knockdown (Fig.
7). There was little effect with MTX alone, siRNA alone, or
in vivo jetPEI alone, on the relative intensity of the
photon bioluminescence. The dose of MTX was set to evaluate the
effects of radixin knockdown on the tumor onset after ascertaining
the anticancer effects of that dose of MTX on HepG2-Luc and
referencing a previous study (32).
The anticancer effects of MTX were enhanced possibly due to the
reduction in the transport activity of MRP2 by radixin knockdown.
Alternative sustained knockdown methods, such as viral vectors or
peptides, should be used to increase the efficiency of MTX,
although radixin is a useful target for the modulation of MRP2 in
tumor cells. Further studies are required to clarify the effects of
radixin knockdown on the other doses of MTX and on the anticancer
efficiency of other drugs, such as doxorubicin, a substrate for
MRP, P-gp, and BCRP (33,34).
In conclusion, the present study provides
information on radixin as a target molecule for the modulation of
MRP activity in tumor cells. It would be useful to further
investigate whether other transporter-associated proteins could be
target proteins for the modulation of transporter functions.
Acknowledgements
Dr Victoria Muir edited a draft of this
manuscript.
Funding
The present study was supported by JSPS KAKENHI
grant no. 18K06806 from the Japanese Ministry of Education,
Culture, Sports, Science, and Technology (MEXT). This research was
also supported in part by the MEXT-Supported Program for the
Strategic Research Foundation at Private Universities, 2014–2018
(S1411037).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
AK and MI participated in the research design. AK,
YI, SN and EK conducted the experiments and performed the data
analysis AK and MI contributed to the writing of the manuscript.
All authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The study protocol was approved by the Committee for
the Care and Use of Laboratory Animals of the School of Pharmacy of
Kindai University (Osaka, Japan).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kikuchi S, Hata M, Fukumoto K, Yamane Y,
Matsui T, Tamura A, Yonemura S, Yamagishi H, Keppler D and Tsukita
S and Tsukita S: Radixin deficiency causes conjugated
hyperbilirubinemia with loss of Mrp2 from bile canalicular
membranes. Nat Genet. 31:320–325. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sekine S, Ito K, Saeki J and Horie T:
Interaction of Mrp2 with radixin causes reversible canalicular Mrp2
localization induced by intracellular redox status. Biochim Biophys
Acta. 1812:1427–1434. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chai J, Cai SY, Liu X, Lian W, Chen S,
Zhang L, Feng X, Cheng Y, He X, He Y, et al: Canalicular membrane
MRP2/ABCC2 internalization is determined by Ezrin Thr567
phosphorylation in human obstructive cholestasis. J Hepatol.
63:1440–1448. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang Y, Dong J, Zhu X, Wang W and Yang Q:
The effect of sphingomyelin synthase 2 (SMS2) deficiency on the
expression of drug transporters in mouse brain. Biochem Pharmacol.
82:287–294. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kobori T, Fujiwara S, Miyagi K, Harada S,
Nakamoto K, Nakagawa T, Takahashi H, Narita M and Tokuyama S:
Involvement of moesin in the development of morphine analgesic
tolerance through P-glycoprotein at the blood-brain barrier. Drug
Metab Pharmacokinet. 29:482–489. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Matsui T, Maeda M, Doi Y, Yonemura S,
Amano M, Kaibuchi K and Tsukita S and Tsukita S: Rho-kinase
phosphorylates COOH-terminal threonines of ezrin/radixin/moesin
(ERM) proteins and regulates their head-to-tail association. J Cell
Biol. 140:647–657. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yano K, Shimizu S, Tomono T and Ogihara T:
Gastrointestinal hormone cholecystokinin increases P-glycoprotein
membrane localization and transport activity in Caco-2 cells. J
Pharm Sci. 106:2650–2656. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vaquero J, Nguyen Ho-Bouldoires TH,
Clapéron A and Fouassier L: Role of the PDZ-scaffold protein
NHERF1/EBP50 in cancer biology: From signaling regulation to
clinical relevance. Oncogene. 36:3067–3079. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He XJ, Wang WR, Zhang Y and Yang Q: The
effect of radixin knockdown on the expression and efflux function
of MRP2 in SGC-7901 cells. Eur J Pharm Sci. 46:426–434. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang L, Xiao R, Xiong J, Leng J, Ehtisham
A, Hu Y, Ding Q, Xu H, Liu S, Wang J, et al: Activated ERM protein
plays a critical role in drug resistance of MOLT4 cells induced by
CCL25. PLoS One. 8:e523842013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yano K, Otsuka K, Kato Y, Kawabata H,
Ohmori S, Arakawa H and Ogihara T: Different regulation of
P-glycoprotein function between Caco-2 and Caki-1 cells by ezrin,
radixin and moesin proteins. J Pharm Pharmacol. 68:361–367. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kawase A, Araki Y, Ueda Y, Nakazaki S and
Iwaki M: Impact of a high-cholesterol diet on expression levels of
Niemann-Pick C1-like 1 and intestinal transporters in rats and
mice. Eur J Drug Metab Pharmacokinet. 41:457–463. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kawase A, Sakata M, Yada N, Nakasaka M,
Shimizu T, Kato Y and Iwaki M: Decreased radixin function for
ATP-binding cassette transporters in liver in adjuvant-induced
arthritis rats. J Pharm Sci. 103:4058–4065. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
den Boer E, Meesters RJ, van Zelst BD,
Luider TM, Hazes JM, Heil SG and de Jonge R: Measuring methotrexate
polyglutamates in red blood cells: A new LC-MS/MS-based method.
Anal Bioanal Chem. 405:1673–1681. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Uchida Y, Ohtsuki S, Kamiie J, Ohmine K,
Iwase R and Terasaki T: Quantitative targeted absolute proteomics
for 28 human transporters in plasma membrane of Caco-2 cell
monolayer cultured for 2, 3, and 4 weeks. Drug Metab Pharmacokinet.
30:205–208. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Prasad B and Unadkat JD: Optimized
approaches for quantification of drug transporters in tissues and
cells by MRM proteomics. AAPS J. 16:634–48. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Harwood MD, Achour B, Russell MR, Carlson
GL, Warhurst G and Rostami-Hodjegan A: Application of an LC-MS/MS
method for the simultaneous quantification of human intestinal
transporter proteins absolute abundance using a QconCAT technique.
J Pharm Biomed Anal. 110:27–33. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kawase A, Fujii A, Negoro M, Akai R,
Ishikubo M, Komura H and Iwaki M: Differences in cytochrome P450
and nuclear receptor mRNA levels in liver and small intestines
between SD and DA rats. Drug Metab Pharmacokinet. 23:196–206. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kawase A, Yamada A, Gamou Y, Tahara C,
Takeshita F, Murata K, Matsuda H, Samukawa K and Iwaki M: Increased
effects of ginsenosides on the expression of cholesterol
7α-hydroxylase but not the bile salt export pump are involved in
cholesterol metabolism. J Nat Med. 67:545–553. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–8. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Puaux AL, Ong LC, Jin Y, Teh I, Hong M,
Chow PK, Golay X and Abastado JP: A comparison of imaging
techniques to monitor tumor growth and cancer progression in living
animals. Int J Mol Imaging. 2011:3215382011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yano K, Tomono T, Sakai R, Kano T,
Morimoto K, Kato Y and Ogihara T: Contribution of radixin to
P-glycoprotein expression and transport activity in mouse small
intestine in vivo. J Pharm Sci. 102:2875–2881. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nakano T, Sekine S, Ito K and Horie T:
Correlation between apical localization of Abcc2/Mrp2 and
phosphorylation status of ezrin in rat intestine. Drug Metab
Dispos. 37:1521–1527. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pokharel D, Padula M, Lu J, Jaiswal R,
Djordjevic S and Bebawy M: The role of CD44 and ERM proteins in
expression and functionality of P-glycoprotein in breast cancer
cells. Molecules. 21:2902016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kano T, Wada S, Morimoto K, Kato Y and
Ogihara T: Effect of knockdown of ezrin, radixin, and moesin on
P-glycoprotein function in HepG2 cells. J Pharm Sci. 100:5308–5314.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kool M, van der Linden M, de Haas M,
Scheffer GL, de Vree JM, Smith AJ, Jansen G, Peters GJ, Ponne N,
Scheper RJ, et al: MRP3, an organic anion transporter able to
transport anti-cancer drugs. Proc Natl Acad Sci USA. 96:6914–6919.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hooijberg JH, Broxterman HJ, Kool M,
Assaraf YG, Peters GJ, Noordhuis P, Scheper RJ, Borst P, Pinedo HM
and Jansen G: Antifolate resistance mediated by the multidrug
resistance proteins MRP1 and MRP2. Cancer Res. 59:2532–2535.
1999.PubMed/NCBI
|
|
28
|
Zeng H, Chen ZS, Belinsky MG, Rea PA and
Kruh GD: Transport of methotrexate (MTX) and folates by multidrug
resistance protein (MRP)3 and MRP1: Effect of polyglutamylation on
MTX transport. Cancer Res. 61:7225–7232. 2001.PubMed/NCBI
|
|
29
|
Volk EL and Schneider E: Wild-type breast
cancer resistance protein (BCRP/ABCG2) is a methotrexate
polyglutamate transporter. Cancer Res. 63:5538–5543.
2003.PubMed/NCBI
|
|
30
|
Chen ZS, Robey RW, Belinsky MG,
Shchaveleva I, Ren XQ, Sugimoto Y, Ross DD, Bates SE and Kruh GD:
Transport of methotrexate, methotrexate polyglutamates, and
17beta-estradiol 17-(beta-D-glucuronide) by ABCG2: Effects of
acquired mutations at R482 on methotrexate transport. Cancer Res.
63:4048–4054. 2003.PubMed/NCBI
|
|
31
|
Vlaming ML, van Esch A, van de Steeg E,
Pala Z, Wagenaar E, van Tellingen O and Schinkel AH: Impact of
abcc2 [multidrug resistance-associated protein (Mrp) 2], abcc3
(Mrp3), and abcg2 (breast cancer resistance protein) on the oral
pharmacokinetics of methotrexate and its main metabolite
7-hydroxymethotrexate. Drug Metab Dispos. 39:1338–1344. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nakase Y, Hagiwara A, Kin S, Fukuda K, Ito
T, Takagi T, Fujiyama J, Sakakura C, Otsuji E and Yamagishi H:
Intratumoral administration of methotrexate bound to activated
carbon particles: Antitumor effectiveness against human colon
carcinoma xenografts and acute toxicity in mice. J Pharmacol Exp
Ther. 311:382–387. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Choi CH: ABC transporters as multidrug
resistance mechanisms and the development of chemosensitizers for
their reversal. Cancer Cell Int. 5:302005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chien AJ and Moasser MM: Cellular
mechanisms of resistance to anthracyclines and taxanes in cancer:
Intrinsic and acquired. Semin Oncol. 35 (Suppl 2):S1–S14; quiz S39.
2008. View Article : Google Scholar : PubMed/NCBI
|