Introduction
Chemotherapy has been demonstrated to reduce the
risk of metastasis by killing the cancer cells circulating in the
bloodstream, thereby increasing the disease-free survival rates
(1). The common types of chemotherapy
for cancer treatment are DNA intercalating agents, such as
epirubicin and doxorubicin, causing lethal DNA damage in the cells
(2). Although chemotherapy is highly
potent in killing cancer cells, some patients do not respond to
chemotherapy, and are referred to as chemoresistant. There is clear
evidence that the Forkhead box M1 (FOXM1) transcription factor is a
protein crucial for DNA damage repair and plays a key role in
cancer progression and chemotherapy resistance (3–6). It was
previously demonstrated (5,7,8) that
doxorubicin-resistant breast cancer cells exhibit a significant
increase in FOXM1 protein expression levels, with no FOXM1
degradation after doxorubicin treatment (3). The active forms of FOXM1, particularly
the FOXM1b and FOXM1c isoforms, can increase DNA double-strand
repair via homologous recombination (3,5).
Conversely, inhibition of FOXM1 expression and activity has been
revealed to result in a marked decrease in cancer metastatic rates
and an increase in the sensitivity of cancer cells to chemotherapy
(4,6,9). Targeting
FOXM1 in cancer cells may prove to be a novel approach to cancer
therapy and may also enhance the efficacy of combination therapies
by increasing chemosensitivity in cancer patients.
Recent research has demonstrated that thiostrepton
can specifically inhibit FOXM1 activity, leading to proliferation
arrest and apoptosis (10,11). Thiostrepton is a natural antibiotic
product, containing a thiazole ring, that is produced by the
Steptomyces species and is recognized to have a broad
spectrum of antibacterial and antiparasitic properties (10). Notably, Kwok et al (11) demonstrated that thiostrepton can
inhibit cell proliferation by causing G1/S and G2/M cell cycle
arrest. Additionally, thiostrepton has been shown to decrease the
expression of X-linked inhibitor of apoptosis protein and matrix
metallopeptidase 9 in FOXM1-overexpressing cells (12).
Cellular senescence is an anticancer event caused by
irreversible cell cycle arrest (13,14). Our
previous research has demonstrated that knockdown of FOXM1 by small
interfering RNA (siRNA) may cause DNA damage-induced senescence
(4), namely irreversible cell cycle
arrest (13,14). However, the mechanism through which
thiostrepton inhibits FOXM1 activity is unclear, since the
interaction between thiostrepton and FOXM1 at the molecular level
remains elusive (15,16). Gartel (15,17,18)
indicated that thiostrepton does not directly bind to and form
complexes with FOXM1; instead, it stabilizes the negative
regulators (p21Cip1 and p53) of FOXM1 by inhibiting the
proteasome degradation pathway, similar to other proteasome
inhibitors, such as siomycin A and MG132 (15,19,20). By
contrast, our previous study demonstrated that FOXM1 protein levels
were increased when MCF-7 breast cancer cells were treated with
MG132 (8). In addition, other
experimental and computational results demonstrated that
thiostrepton can interact directly with the DNA-binding domain
(DBD) of FOXM1 (16,21). Isothermal titration calorimetry (ITC)
measurement of the thiostrepton-FOXM1 interaction also revealed
that thiostrepton binds directly to FOXM1 with 1:1 stoichiometry
(16). Moreover, the binding of
thiostrepton and FOXM1 was also confirmed by affinity pull-down
assays (16). However, crucial
evidence remains elusive due to limitations in time and length
scale of the binding process (15,17,18,22).
The advances in computational simulations offer an alternative
complementary approach, and high-throughput screenings for small
molecules binding to the dimer FOXM1-DNA complex have been
successfully carried out (21).
The aim of the present study was to investigate the
role of thiostrepton in inducing senescence in cancer cells, in
order to gain a better understanding of its antiproliferative
properties and its functional doses at low concentrations. The
association between the effects of thiostrepton on FOXM1 inhibition
and cellular senescence was also investigated. In addition, in
order to gain further insight into the interactions between
thiostrepton and FOXM1, computational simulations were performed.
Collectively, these data may reveal the mechanism through which
thiostrepton inhibits the transactivation activity of FOXM1 and
help design a novel, effective molecular inhibitor of FOXM1 in
breast cancer treatment. Furthermore, a deeper understanding of the
binding mechanism between thiostrepton and FOXM1 may also aid in
the rational structure-based design of drug candidates.
Materials and methods
Cell culture and thiostrepton
treatment
The MCF-7 breast cancer cell line was used in the
present study, originating from the American Type Culture
Collection and acquired through the Cancer Research UK Cell Bank.
The triple-negative breast cancer MDA-MB-436 cell line was obtained
from the National Nanotechnology Centre (NANOTEC) in Thailand. Both
cell types were cultured in Dulbecco's modified Eagle's medium
supplemented with 10% foetal bovine serum, and 100 U/ml
penicillin/streptomycin (all from Gibco; Thermo Fisher Scientific,
Inc.). The maintenance conditions were at 37°C in a humidified
incubator with 5% CO2, as previously described (4). Thiostrepton (Sigma-Aldrich; Merck KGaA)
was used for cell treatment at various final concentrations,
ranging between 0 and 100 µg/ml.
MTT assay
The cell viability MTT assay was used to measure the
cytotoxic effect of thiostrepton on MCF-7 or MDA-MB-436 cells after
treatment for 24, 48 and 72 h. In brief, cells were seeded in a
96-well plate at a density of 2,000 cells/well. After 24 h, the
culture medium was replaced with various concentrations of
thiostrepton. At each time-point, the viability of the tested cells
was determined by adding MTT (Sigma-Aldrich; Merck KGaA) and
incubating at 37°C, allowing formazan crystals to form. The
crystals were dissolved in dimethyl sulfoxide (Merck KGaA) and the
absorbance at 490 nm was measured with a microplate reader (BioTek
PowerWave XS; BioTek Instruments, Inc.). Five replicate wells were
used for each analysis, and three independent experiments were
performed.
Reverse transcription-polymerase chain
reaction (RT-PCR) analysis
MCF-7 and MDA-MB-436 cells were cultured and treated
with various concentrations of thiostrepton, similar to the MTT
assay. Total RNA was extracted from the cells using Ribospin™ kit
(GeneAll) and the concentration was measured using NanoDrop
(DeNovix). Gene expression analyses were conducted using 2X
HyperScript™ One-step RT-PCR Master mix (GeneAll). The forward and
reverse primers (Macrogen, Inc.) were as follows (23,24):
FOXM1 (all isomers a, b and c), forward
5′-CACCCCAGTGCCAACCGCTACTTG-3′ and reverse
5′-AAAGAGGAGCTATCCCCTCCTCAG-3′; cyclin B1 (CCNB1), a
downstream target gene, forward 5′-AAGAGCTTTAAACTTTGGTCTGGG-3′ and
reverse 5′-CTTTGTAAGTCCTTGATTTACCATG-3′; and b-actin (ACTB),
serving as a housekeeping control, forward
5′-ATCTGGCACCACACCTTCTACAATGAGCTGCG-3′ and reverse
5′-CGTCATACTCCTGCTTGCTGATCCACATCTGC-3′. The PCR conditions for all
genes were 30 cycles at 96°C for 30 sec, annealing at 56°C for 30
sec and extension at 72°C for 30 sec.
Senescence-associated β-galactosidase
(SA-β-gal) assay
Cells were seeded in 6-well plates at a density of
10,000 cells/well, then treated with thiostrepton at concentrations
of 0–4 µM for 24 h before changing to a drug-free medium, and
incubated for another 24 h. To harvest, the cells were fixed and
stained using a Senescence-associated-β-galactosidase Staining Kit
(cat. no. 9860, Cell Signaling Technology, Inc.), as previously
described (4). The plate was
incubated in a dry incubator overnight at 37°C without
CO2. The cells that were SA-β-gal-positive were
indicated by blue staining. The images were captured under a
converted bright-field microscope (Nikon Eclipse TS100; Nikon
Corporation) with a Nikon digital sight DS-L2 monitor (Nikon
Corporation) for at least 3–5 fields. The percentages of
SA-β-gal-positive cells were quantified by the amount of positive
stained cells divided by the total number of cells counted.
Molecular modelling and
simulations
To investigate the effects of thiostrepton on the
stability of the FOXM1-DNA-binding domain, atomistic molecular
dynamics (MD) simulations of the FOXM1-DNA-binding domain and
thiostrepton were performed. The structures of FOXM1-DNA (25) and thiostrepton (26) were obtained from the Protein Data Bank
with accession nos. 3G73 (https://www.rcsb.org/structure/3G73) and 2L2W
(https://www.rcsb.org/structure/2L2W),
respectively. The isolated FOXM1-DNA-binding domain was expressed
in E. coli and the 3D structure was determined by X-ray
diffraction with a resolution of 2.21 Ångström (25). The DNA double-strand target consisted
of a DNA-A and a DNA-B chain, with a base-paired DNA sequence of
AAATTGTTTATAAACAGCCCG and TTCGGGCTGTTTATAAACAAT for DNA-A and
DNA-B, respectively. For thiostrepton, the molecular structure was
determined by nuclear magnetic resonance (26). The topology of thiostrepton was
created by a web-accessible Automated force field Topology Builder
(ATB; http://compbio.biosci.uq.edu.au/atb/) (27–29).
Initially, the protein structures of FOXM1-DNA and thiostrepton
were submitted to the AutoDockTools-1.5.6 (Autodock-4.2) (30) to search the binding region. To
identify the thiostrepton-binding complex structure, three
different models were performed, including i) a FOXM1 monomer, ii)
a FOXM1 dimer and iii) a FOXM1 dimer complex with DNA. To study the
structure stability of the binding complex, the structure with
thiostrepton in the binding domain region of the FOXM1 dimer model
was selected for further MD simulations. The details of the
selected structures are discussed in Results and Discussion.
Two MD simulations of FOXM1-DNA with and without
thiostrepton were considered in this study and carried out using
GROMACS version 5.1.3 (31). The
structure of thiostrepton at the lowest binding energy in the FOXM1
dimer model was used as the initial structure. The parameters of
all molecules were derived from GROMOS53a6 force field (32) and the simple point-charge (SPC) model
(33) was used for water. Each system
was solvated by 37,337 water molecules, and 17 Mg2+ ions
were added for neutralizing the system charges. The system energy
was minimized by the steepest descent algorithm to remove steric
conflicts between atoms. In order to avoid rearrangement of the
FOXM1 protein structures, position restraints were applied to the
FOXM1 backbone with a force constant of 1,000 kJ (mol
nm)−1 in all directions. Subsequently, MD simulations
were performed under NPT (constant particle number, pressure and
temperature) ensemble for 200 nsec with an integration time step of
1 fsec. The last 50 nsec of the trajectories were used to determine
the binding interactions among FOXM1, DNA and thiostrepton. The
temperature was maintained constant using the velocity-rescale
(V-rescale) algorithm (34,35) at 298 K with a time constant of 0.1
psec. Semi-isotropic pressure was applied under a constant pressure
of 1 bar with a time constant of 3.0 psec and a compressibility of
4.5×10−5 bar−1 using the Parrinello-Rahman
algorithm (36). Periodic boundary
conditions in all directions were used. A cut-off with a distance
of 1.0 nm was applied for the real-space part of the electrostatic
and Lennard-Jones interaction. In the reciprocal-space interactions
of long-range electrostatics, the Particle-mesh Ewald (37,38) method
was employed with a 0.12-nm grid and the cubic interpolation of
order four. The simulation protocol had been tested to avoid
physical artefacts (39). All bond
lengths were constrained using the P-LINCS algorithm (40). The Visual Molecular Dynamics (VMD)
software was used for system visualization (41).
Statistical analysis
All statistical analyses were performed using SPSS
16.0 (Imperial College London) and Microsoft Excel 2013 (Kasetsart
University). Values are presented as the mean ± standard deviation
(SD). One-way ANOVA was performed followed by post hoc analysis
with Dunett's test. P-values <0.05 were considered to indicate
statistically significant differences.
Results and Discussion
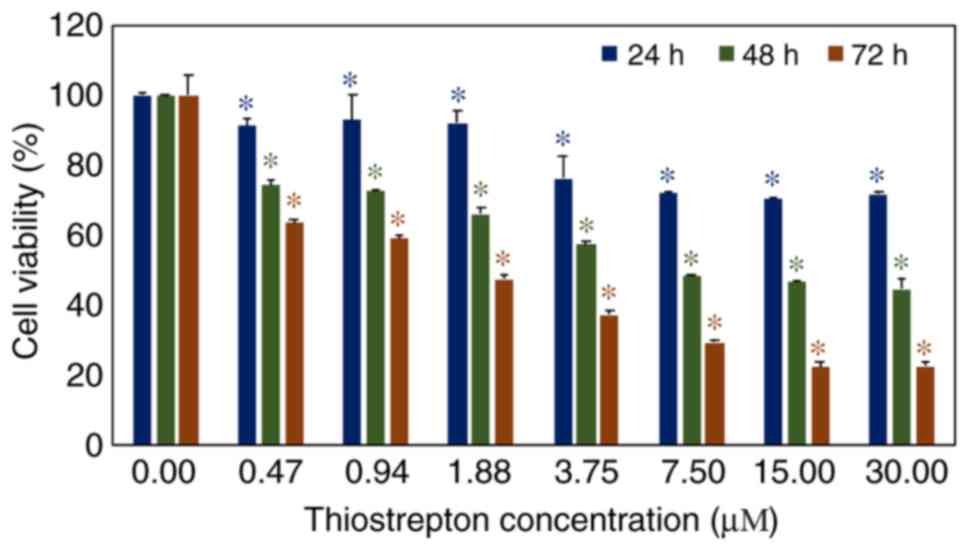
In vitro investigation of thiostrepton
suppresses breast cancer cell growth
The effect of thiostrepton on the suppression of
MCF-7 breast cancer cell growth was studied using an MTT assay. The
results revealed that thiostrepton significantly restricted cancer
cell proliferation in a dose-dependent manner. Following
thiostrepton exposure for 24 h, cell viability was significantly
decreased (P<0.05) after being treated with a dose of 0.47
µM (Fig. 1). After 48 and 72 h
of treatment, the viability of MCF-7 cells was significantly
reduced by lower doses of thiostrepton (0.47 µM).
Additionally, the effect of thiostrepton on the triple-negative
breast cancer cell line MDA-MB-436 was similar to that on MCF-7
cells (Fig. S1). Both breast cancer
cell types were affected in a dose-dependent manner. These results
confirmed that thiostrepton is an effective candidate compound for
developing a novel anticancer treatment (11).
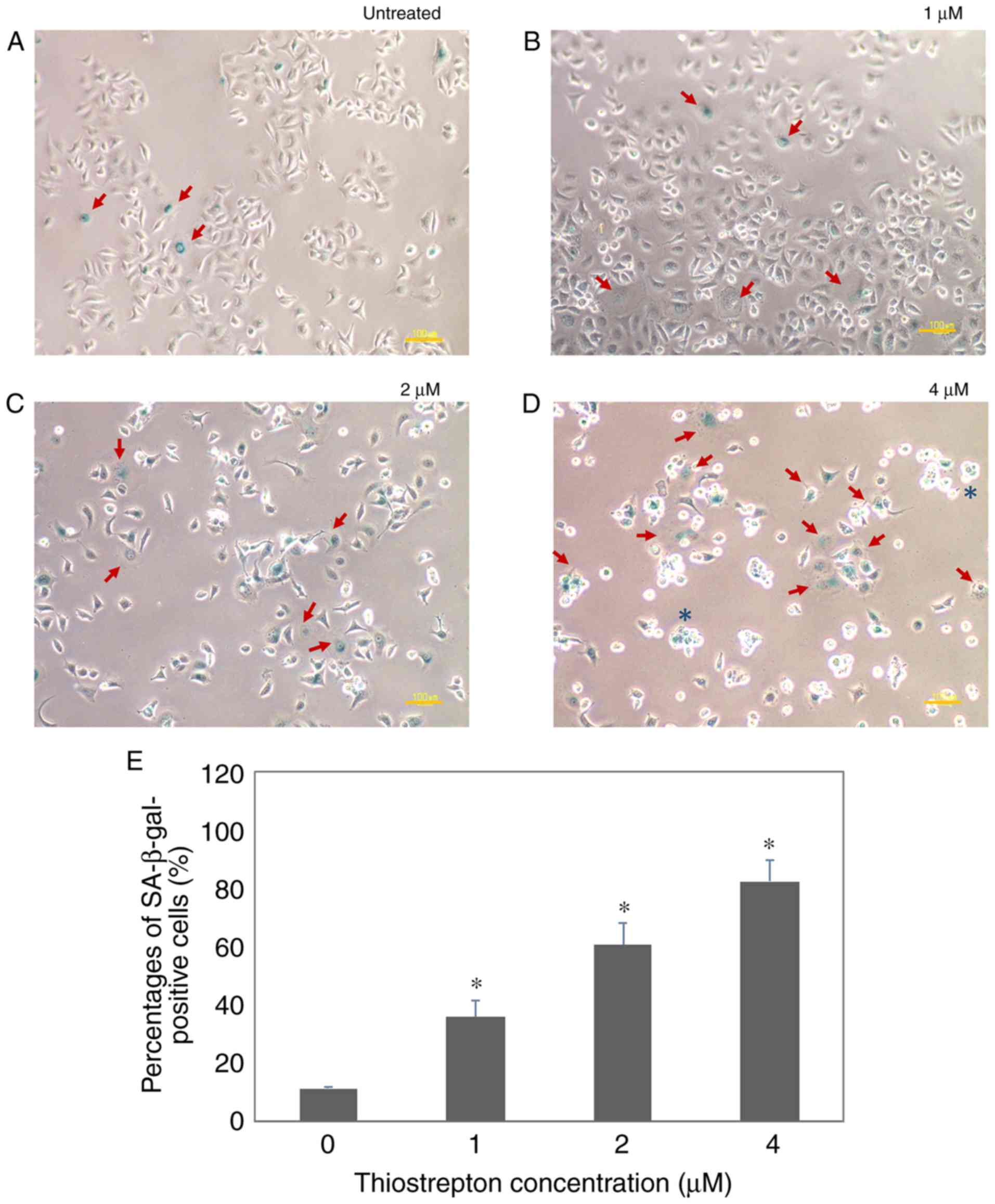
Thiostrepton potently induces
senescence in MCF-7 breast cancer cells
Inhibition of FOXM1 has been shown to induce cell
cycle arrest due to its crucial role in G1 and G2/M cell cycle
phase progression. Depletion of FOXM1 results in DNA damage and
induces cellular senescence (4).
Therefore, we investigated whether the effects of FOXM1 inhibition
by thiostrepton treatment are mediated by causing irreversible cell
cycle arrest (also referred to as cellular senescence). MCF-7 cells
were assayed for SA-β-gal positivity after the cells were subjected
to a sequence of thiostrepton concentrations (0, 1, 2 and 4
µM), as revealed in Fig. 2.
Senescent cells displayed SA-β-gal activity and were stained green
in the presence of chromogenic substrate
5-bromo-4-chloro-3-indolyl-β-d-galactopyranoside (X-gal).
Consistently, MCF-7 cells treated with thiostrepton exhibited
SA-β-gal activity and their morphology changed (Fig. 2B-D, red arrows), with the cells
becoming enlarged and flattened. This was consistent with our
previous findings on cell cycle analysis, demonstrating that
thiostrepton can cause cell cycle arrest at the G1 phase in MCF-7
cells, but not in untransformed breast epithelial MCF-10 cells
(11). The SA-β-gal activity was
determined to be increased in a dose-dependent manner, with the
highest activity observed at 4 µM. At this concentration,
some cells also became rounded, which is consistent with the
morphology of dying cells (Fig. 2D,
blue asterisk). In addition, MDA-MB-436 cells also exhibited
senescence morphology, with positive SA-β-gal activity, after being
exposed to thiostrepton (Fig. S2).
These thiostrepton-treated cells displayed typical senescence
characteristics, such as enlargement, flattening and reduction in
numbers (13,14). Under the bright-field microscope,
additional cell death morphological characteristics, such as
vacuolization, were also observed. Moreover, the cells started to
detach from the culture plate and assume a rounder shape. Of note,
the cells incubated with 2 and 4 µM of thiostrepton
exhibited lower viability, as some of the cells died after
treatment (Fig. 2). These findings
highlight the ability of thiostrepton to induce senescence in
breast cancer cells. The results strongly indicated that
thiostrepton can inhibit cancer progression by inducing both cell
death and senescence via a FOXM1-dependent pathway.
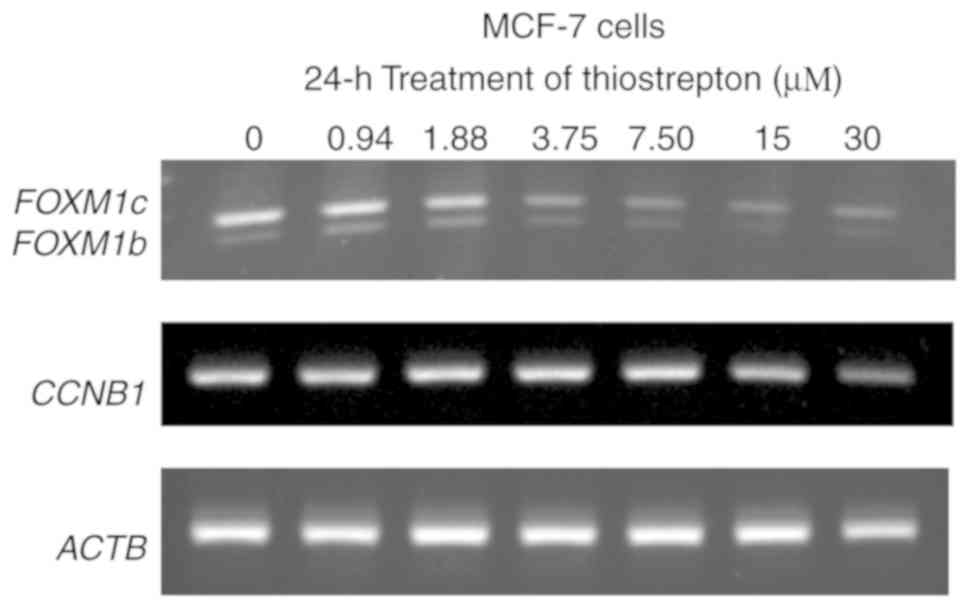
Thiostrepton reduces FOXM1 and CCNB1
expression at the transcriptional level
Next, the effect of thiostrepton on mRNA expression
of FOXM1 (auto-regulation) (42) and the well-known FOXM1 downstream
target CCNB1 (43) were
investigated. After 24 h of thiostrepton exposure, MCF-7 cells
exhibited downregulation of FOXM1 (Fig. 3). The decrease in FOXM1 levels
was evident at a thiostrepton concentration of 3.75 µM. This
downregulation of FOXM1 mRNA may be attributed to the
auto-regulation ability of FOXM1, as it can bind to its own
promoter and regulate its transcription (18,42). For
CCNB1, the mRNA expression was decreased after thiostrepton
treatment at 15 µM. Of note, the FOXM1 and
CCNB1 mRNA levels decreased with increasing drug
concentrations (Fig. 3) and time of
exposure (Fig. S3), which were also
correlated with significant decreases in the viability of MCF-7
cells (Fig. 3). Notably, the great
changes on the CCNB1 mRNA level at 30 µM indicated
the effect of transactivation activity of thiostrepton on FOXM1. A
similar phenomenon was also observed in MDA-MB-436 cells. The mRNA
expression of FOXM1 and CCNB1 was also decreased
after thiostrepton exposure in dose- and time-dependent manners
(Figs. S3 and S4). Collectively, these data indicated that
thiostrepton inhibited FOXM1 mRNA expression for both
isoforms b and c. These two isoforms of FOXM1 play a crucial role
in carcinogenesis, cancer progression and metastasis (44,45), and
the downregulation of FOXM1 mRNA expression appears to be an
interesting target for anticancer therapy (18,46).
Therefore, our results suggest that thiostrepton may be a good
candidate for inhibition of cancer cell growth and metastasis in
patients with FOXM1 overexpression (12,18,46).
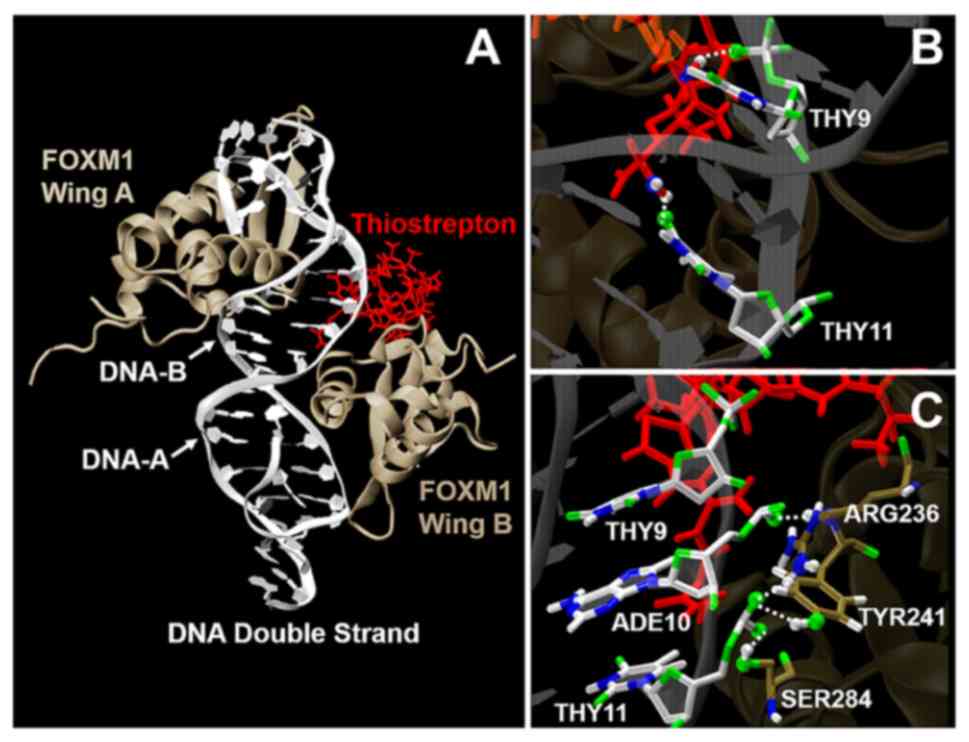
Interactions of thiostrepton and
binding domain of FOXM1 forkhead/winged-helix domain (FKH) with
targeted DNA promoter motif
In a previous study (25), the FOXM1 forkhead/winged helix domain
(FKH) sequence at residues 222–360 was identified as the DBD with a
tandem recognition sequence of the downstream target (TAAACA)
(47). Specific mutations in the DBD
have been revealed to lead to the decrease of FOXM1 transcriptional
activity (48). The binding energy
(affinity) between FOXM1 and DNA promoter motif were calculated by
the molecular docking technique (30). FOXM1 was selected as the protein
macromolecule and the DNA promoter motif served as the ligand. The
nine complex structures with different binding modes were used to
calculate the binding energy (∆G), with mode 1 having the lowest
binding energy. The ∆G between FOXM1 and promoter DNA motif was
determined to be between −6.0 and −5.3 kcal/mol, in agreement with
the preferential binding of the FOXM1 to the targeted promoter
motif DNA sequence. Next, the thiostrepton binding affinity to
FOXM1 was determined by performing three different models, as
revealed in Fig. 4: i) FOXM1 monomer
model, ii) FOXM1 dimer model and iii) FOXM1 dimer complex with DNA.
The first model (FOXM1 monomer) revealed that thiostrepton could
bind to the DBD at three possible positions and the binding energy
of this model was in the range of −6.4 to −5.5 kcal/mol (Fig. 4A). For the second model (FOXM1 dimer;
wing A and wing B), five possible positions between thiostrepton
and the FOXM1 DBD proteins were found, with the binding energy
ranging from −7.8 to −6.9 kcal/mol (Fig.
4B). The last model (FOXM1 dimer complex with DNA binding
motif) revealed that the binding affinity was decreased (−8.0 to
−7.6 kcal/mol), with two possible positions (Fig. 4C); however, neither position was
suitable to accommodate thiostrepton inside the binding domain of
FOXM1. Conversely, the first and second models enabled thiostrepton
to prevent the binding of FOXM1 to the DNA motif. Chen et al
(21) previously investigated the
notable behavior of thiostrepton in contact with the FOXM1 dimer
and partial binding to the DNA motif. Therefore, the thiostrepton
structure in the FOXM1 dimer from the second model (Mode 1 in
Fig. 4B) in a complex with DNA
binding motif was selected to perform molecular dynamic (MD)
simulations in order to compare to the structure without
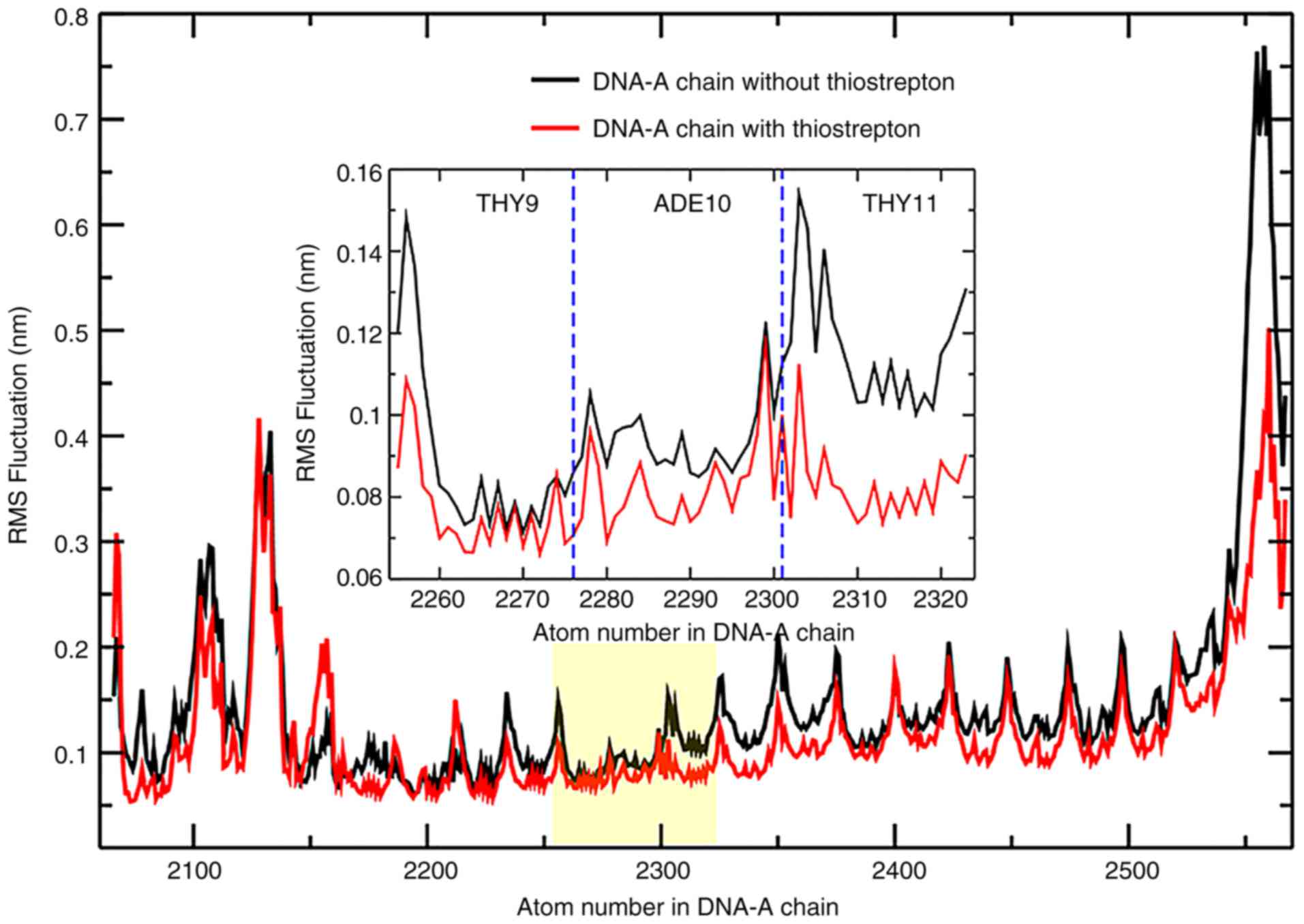
thiostrepton. The root-mean-square fluctuation (RMSF) analysis
indicated a stabilized structure of DNA, particularly at residues
in the binding domain regions of FOXM1 (Fig. 5). This result demonstrated that
thiostrepton can increase the stability of the binding between DNA
and FOXM1 inside the DBD.
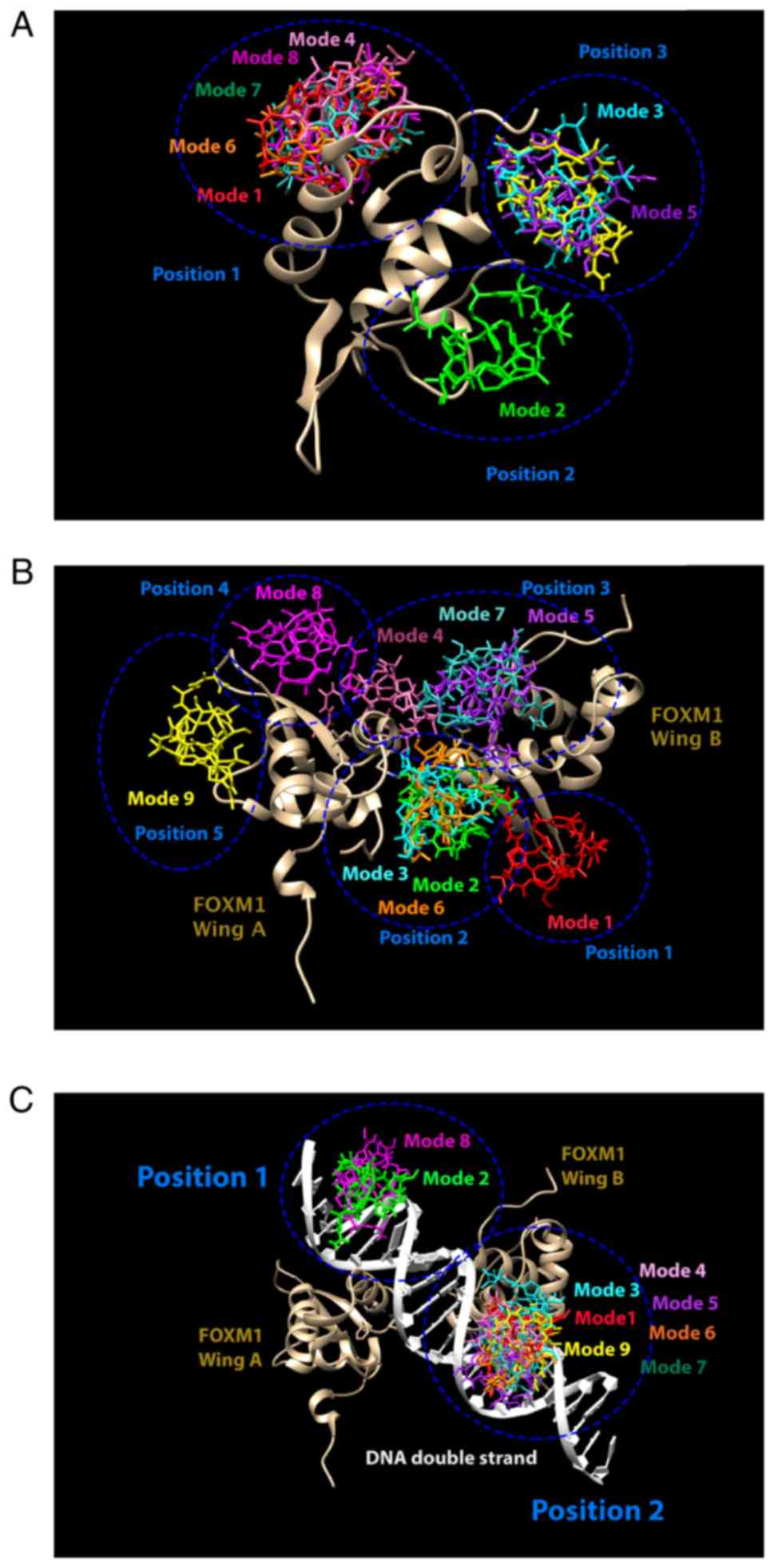 | Figure 4.Different modes of thiostrepton
binding to FOXM1 (golden bronze) with different models: (A) Monomer
FOXM1, (B) dimer FOXM1 and (C) FOXM1-DNA complex. The lowest
binding energy of thiostrepton was found in mode 1 (red), followed
by mode 2 (green), 3 (cyan), 4 (pink), 5 (purple), 6 (orange), 7
(sea blue-green), 8 (magenta) and 9 (yellow). Blue circles,
different possible positions of binding area. FOXM1, Forkhead box
M1. |
The mean number and lifetime of hydrogen bonds among
FOXM1, DNA and thiostrepton were analyzed, as it was previously
suggested that FOXM1-DNA complex interaction involves van der Waals
forces and hydrogen bonding (49). A
hydrogen bond was formed when the distance between donor and
acceptor (rHB) was <0.35 nm and the
hydrogen-donor-acceptor angle (αHB) was <30°. The
value of 0.35 nm corresponds to the first minimum of the radial
distribution function of water. The overall hydrogen bond lifetime
(τHB) is calculated from the mean over all
autocorrelation functions C(τ) of the lifetime distribution P(τ) of
all hydrogen bonds from time 0 to t: (50):
C(τ)=1-∫0tP(τ)dτ
τHB=∫0∞C(τ)dτ
The number of hydrogen bonds in the binding promoter
region among FOXM1 wing A, DNA-A and thiostrepton were analyzed and
are presented in Table I and Fig. 6. The hydrogen bond binding domains
were observed in residues Tyr241(tyrosine), Ser284(serine) and
Arg236 (arginine) of the FOXM molecule, and residues Ade10
(adenine) and Thy11 (thymine) of the DNA-A chain. These binding
residues were consistent with those reported previously (25). The number of hydrogen bonds between
FOXM1 wing A and DNA-A in the system was 4.34±0.24 and decreased
slightly to 4.17±0.05 when the thiostrepton molecule formed a
complex with the FOXM1-DNA binding domain. Moreover, the increase
in binding stability may be explained by the presence of 0.95±0.66
hydrogen bonds between thiostrepton and DNA-A in the promoter
binding region. In addition, thiostrepton forming a complex with
FOXM1-DNA was associated with an increase in hydrogen bond lifetime
in the binding domain. The mean number of hydrogen bonds and their
lifetimes are presented in Table I.
Notably, our results indicated that the increase in the stability
of hydrogen bonds in the promoter binding region may result in
transcription inhibition. Moreover, these results also indicated a
mechanism through which thiostrepton inhibits the transcriptional
activity of a transcription factor. However, protein-DNA
interaction experiments such as chromatin immunoprecipitation
sequencing (ChIP) assay, electrophretic mobility shift assay, and
reporter assay may be performed in a future study to confirm the
experimentally assessable potential of FOXM1-DNA interaction in
vitro.
 | Table I.Components of number of hydrogen
bonds and hydrogen bond lifetimes in each residue among FOXM1 wing
A, DNA-A, and thiostrepton inside the DNA binding domain. |
Table I.
Components of number of hydrogen
bonds and hydrogen bond lifetimes in each residue among FOXM1 wing
A, DNA-A, and thiostrepton inside the DNA binding domain.
| Residues | FOXM1-DNA | FOXM1-DNA with
thiostrepton |
|---|
|
|
|
|---|
| FOXM1 Wing A | DNA-A | Thiostrepton | Number of
H-bonds | H-bond lifetimes
(psec) | Number of
H-bonds | H-bond lifetimes
(psec) |
|---|
| Tyr241 | Ade10 | – | 1.09±0.02 | 32.20 | 1.02±0.03 | 100.44 |
| Tyr241 | Thy11 | – | 1.00±0.01 | 72.36 | 1.04±0.00 | 75.96 |
| Ser284 | Thy11 | – | 0.64±0.00 | 97.97 | 0.99±0.00 | 193.73 |
| Arg236 | Thy11 | – | 0.17±0.03 | 6.40 | 0.04±0.02 | 8.19 |
| – | Thy9 | Drg | – | – | 0.62±0.00 | 14.90 |
| – | Ade10 | Drg | – | – | 0.22±0.00 | 25.32 |
| – | Thy11 | Drg | – | – | 0.11±0.00 | 7.58 |
Clinically, FOXM1 expression has been confirmed to
be significantly associated with resistance to chemotherapy and
suggested to be a reliable biomarker for adverse prognosis in
cancer patients (51). Conversely,
depletion of FOXM1 was revealed to lead to the decrease of
homologous recombination repair in breast cancer cells and the
increase in sensitivity to genotoxic chemotherapy drugs (5). Three isoforms of the FOXM1 protein
exist, and they are the result of alternative splicing (6). FOXM1a is entirely inactive, as both
alternative exons insert in the C-terminal transactivation domain
(6,9,52). By
contrast, the other two isoforms, FOXM1b and FOXM1c, are
transcriptionally active and responsible for cancer cell growth and
resistance to chemotherapy by promoting the expression of genes
involved in cell proliferation, DNA repair and other
characteristics important for cancer development and progression
(6).
In the present study, the effect of thiostrepton on
cell viability was examined by using an MTT cell viability assay
and RT-PCR. The results confirmed that thiostrepton alone was able
to reduce the percentage of viable breast cancer cells by
suppressing the ability of FOXM1 to regulate the expression of
target genes, including FOXM1 and CCNB1, at the
transcriptional level. Moreover, the present study also
demonstrated that thiostrepton can inhibit the transcriptional
activity of both FOXM1b and FOXM1c, the dominant isoforms in
various types of cancer (6,53). These results indicated that
thiostrepton has the potential to be used in both FOXM1b- and
FOXM1c-overexpressing cancer patients. In addition, thiostrepton
has been reported to promote cancer cell death by apoptosis and by
inducing cell cycle arrest at the G1/S phase (11). The status of the cell cycle was
further investigated and it was revealed that the cycle arrest of
breast cancer cells induced by thiostrepton is an irreversible
event via cellular senescence (4,13,14). The present results demonstrated that
senescent cancer cells characteristically change their shape and
size (13,14). Although thiostrepton was clearly
revealed to decrease cell viability and induce cellular senescence,
the molecular mechanism underlying inhibition of FOXM1 expression
remains controversial (15,16) Previous studies demonstrated that a
small-molecule inhibitor, such as FDI-6, may interfere with the
binding of DBD and the targeted DNA, resulting in the inhibition of
FOXM1 function and transcriptional programing (49,54–56)
Notably, our molecular docking results also demonstrated the
ability of thiostrepton to bind to the FOXM1-DNA complex, thereby
interfering with the transcription of the downstream targets. This
evidence further confirms that thiostrepton is a suitable compound
for developing FOXM1-targeted therapy, as it can directly and
specifically bind to FOXM1 (11,16,21) The MD
simulation demonstrated that the lifetime of hydrogen bonds between
wing-helix DBD FOXM1 and the targeted DNA motif can be increased by
forming complexes with thiostrepton. Changes in interaction between
protein and DNA, such as a tight binding, will cause unfavorable
conditions for binding and sliding between protein and DNA
(57). Since FOXM1 is an
autoregulated protein, the increasing stability of the hydrogen
bonds may play a key role in suppressing the activity of FOXM1 and
leading to downregulation of FOXM1.
Collectively, our findings may enable a better
understanding of the DNA-FOXM1 binding with thiostrepton and may
aid in developing future anticancer strategies. In silico
and in vitro studies help to verify the efficiency of the
anticancer effects of thiostrepton at an atomic level (25). In agreement with previous studies
(11,17,22), we
herein demonstrated that thiostrepton may be an optimal compound
for overcoming cancer resistance to chemotherapy (particularly to
cisplatin, carboplatin, epirubicin and doxorubicin) via targeting
FOXM1 (11,58).
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Associate Professor
Kiattawee Choowongkomon, Department of Biochemistry, Faculty of
Science, Kasetsart University, for the valuable advice and ideas
during the research process. Computing facilities were provided by
the Department of Physics, Faculty of Science, Kasetsart
University.
Funding
The present was supported by a grant from Kasetsart
University Research and Development Institute (KURDI), Kasetsart
University, Bangkok, Thailand. SW received a grant by the National
Research Council of Thailand (NRCT: Graduate Scholarship, 2019) and
the Thailand Graduate Institute of Science and Technology (TGIST:
grant no. SCA-CO-2561-6950-TH) affiliated to the National Science
and Technology Development Agency (NSTDA). The support of the
Thailand Research Fund through the Royal Golden Jubilee Ph.D.
Program (grant no. PHD/0204/2559) and the TRF Research Scholar
(grant no. RSA6180021) to PB and JW is acknowledged. The research
of EW-FL was supported by the Medical Research Council (MRC)
(MR/N012097/1), Cancer Research UK (CRUK) (C37/A12011;C37/A18784),
Breast Cancer Now (2012MayPR070; 2012NovPhD016), the Cancer
Research UK Imperial Centre, the Imperial Experimental Cancer
Medicine Centre (ECMC) and the National Institute for Health
Research (NIHR) Imperial Biomedical Research Centre (BRC).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
MKo designed and performed the experiments,
analysed and the interpreted data. SW performed the in vivo
experiments and the statistical analysed the data. MKh designed and
interpreted the data. EWFL designed the experiments, analysed and
provided some critical idea during analyses. WV analysed and
interpreted the data. PB and JWE designed, performed the MD
simulations and analysed the data. All authors read and approved
the final manuscript and agree to be accountable for all aspects of
the research in ensuring that the accuracy or integrity of any part
of the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ATB
|
Automated force field Topology
Builder
|
|
ChIP-seq
|
chromatin immunoprecipitation
sequencing
|
|
DBD
|
DNA binding domain
|
|
DMEM
|
Dulbecco's modified Eagle's
medium
|
|
DMSO
|
dimethyl sulfoxide
|
|
FBS
|
foetal bovine serum
|
|
FKH
|
Forkhead box domain
|
|
FOXM1
|
Forkhead box M1
|
|
ITC
|
isothermal titration calorimetry
|
|
MD
|
molecular dynamics
|
|
MMP-9
|
matrix metallopeptidase 9
|
|
MTT
|
3-(4, 5-dimethylthiazolyl-2)-2,
5-diphenyltetrazoliumbromide
|
|
NMR
|
nuclear magnetic resonance
|
|
PME
|
Particle-mesh Ewald
|
|
RMSF
|
root-mean-square fluctuation
|
|
RT-PCR
|
reverse transcription polymerase
chain reaction
|
|
SA-β-gal
|
senescence-associated
β-galactosidase
|
|
siRNA
|
small interfering RNA
|
|
SPC
|
simple point-charge model
|
|
TAD
|
transactivation domain
|
|
V-rescale
|
velocity-rescale
|
|
VMD
|
visual molecular dynamics
|
|
X-gal
|
5-bromo-4-chloro-3-indolyl-β-d-galactopyranoside
|
|
XIAP
|
X-linked inhibitor of apoptosis
protein
|
References
|
1
|
Krebs MG, Hou JM, Ward TH, Blackhall FH
and Dive C: Circulating tumour cells: Their utility in cancer
management and predicting outcomes. Ther Adv Med Oncol. 2:351–365.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thorn CF, Oshiro C, Marsh S,
Hernandez-Boussard T, McLeod H, Klein TE and Altman RB: Doxorubicin
pathways: Pharmacodynamics and adverse effects. Pharmacogenet
Genomics. 21:440–446. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Myatt SS, Kongsema M, Man CW, Kelly DJ,
Gomes AR, Khongkow P, Karunarathna U, Zona S, Langer JK, Dunsby CW,
et al: SUMOylation inhibits FOXM1 activity and delays mitotic
transition. Oncogene. 33:4316–4329. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Khongkow P, Karunarathna U, Khongkow M,
Gong C, Gomes AR, Yagüe E, Monteiro LJ, Kongsema M, Zona S, Man EP,
Tsang JW, et al: FOXM1 targets NBS1 to regulate DNA damage-induced
senescence and epirubicin resistance. Oncogene. 33:4144–4155. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Monteiro LJ, Khongkow P, Kongsema M,
Morris JR, Man C, Weekes D, Koo CY, Gomes AR, Pinto PH, Varghese V,
et al: The Forkhead Box M1 protein regulates BRIP1 expression and
DNA damage repair in epirubicin treatment. Oncogene. 32:4634–4645.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Koo CY, Muir KW and Lam EW: FOXM1: From
cancer initiation to progression and treatment. Biochim Biophys
Acta. 1819:28–37. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Millour J, de Olano N, Horimoto Y,
Monteiro LJ, Langer JK, Aligue R, Hajji N and Lam EW: ATM and p53
regulate FOXM1 expression via E2F in breast cancer epirubicin
treatment and resistance. Mol Cancer Ther. 10:1046–1058. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Karunarathna U, Kongsema M, Zona S, Gong
C, Cabrera E, Gomes AR, Man EP, Khongkow P, Tsang JW, Khoo US, et
al: OTUB1 inhibits the ubiquitination and degradation of FOXM1 in
breast cancer and epirubicin resistance. Oncogene. 35:1433–1444.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Laoukili J, Stahl M and Medema RH: FoxM1:
At the crossroads of ageing and cancer. Biochim Biophys Acta.
1775:92–102. 2007.PubMed/NCBI
|
|
10
|
Nicolaou KC, Zak M, Rahimipour S, Estrada
AA, Lee SH, O'Brate A, Giannakakou P and Ghadiri MR: Discovery of a
biologically active thiostrepton fragment. J Am Chem Soc.
127:15042–15044. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kwok JM, Myatt SS, Marson CM, Coombes RC,
Constantinidou D and Lam EW: Thiostrepton selectively targets
breast cancer cells through inhibition of forkhead box M1
expression. Mol Cancer Ther. 7:2022–2032. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ahmed M, Hussain A, Begum R, Thangavel S,
Ajarim DS, Beg S, Uddin S and Al-Kuraya KS: Abstract 55:
Over-expression of FoxM1 in breast cancer can be therapeutically
targeted using thiostrepton. Cancer Res. 75:55. 2015.
|
|
13
|
Collado M and Serrano M: Senescence in
tumours: Evidence from mice and humans. Nat Rev Cancer. 10:51–57.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rodier F and Campisi J: Four faces of
cellular senescence. J Cell Biol. 192:547–556. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gartel AL: Thiostrepton, proteasome
inhibitors and FOXM1. Cell Cycle. 10:4341–4342. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hegde NS, Sanders DA, Rodriguez R and
Balasubramanian S: The transcription factor FOXM1 is a cellular
target of the natural product thiostrepton. Nat Chem. 3:725–731.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gartel AL: Suppression of the oncogenic
transcription factor FOXM1 by proteasome inhibitors. Scientifica
(Cairo). 2014:5965282014.PubMed/NCBI
|
|
18
|
Gartel AL: Targeting FOXM1 auto-regulation
in cancer. Cancer Biol Ther. 16:185–186. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bhat UG, Halasi M and Gartel AL: Thiazole
antibiotics target FoxM1 and induce apoptosis in human cancer
cells. PLoS One. 4:e55922009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gartel AL: A new target for proteasome
inhibitors: FoxM1. Expert Opin Investig Drugs. 19:235–242. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen Y, Ruben EA, Rajadas J and Teng NN:
In silico investigation of FOXM1 binding and novel inhibitors in
epithelial ovarian cancer. Bioorg Med Chem. 23:4576–4582. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Gartel AL: Thiazole antibiotics siomycin a
and thiostrepton inhibit the transcriptional activity of FOXM1.
Front Oncol. 3:1502013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nakamura S, Hirano I, Okinaka K, Takemura
T, Yokota D, Ono T, Shigeno K, Shibata K, Fujisawa S and Ohnishi K:
The FOXM1 transcriptional factor promotes the proliferation of
leukemia cells through modulation of cell cycle progression in
acute myeloid leukemia. Carcinogenesis. 31:2012–2021. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Villa-Diaz LG, Garcia-Perez JL and
Krebsbach PH: Enhanced transfection efficiency of human embryonic
stem cells by the incorporation of DNA liposomes in extracellular
matrix. Stem Cells Dev. 19:1949–1957. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Littler DR, Alvarez-Fernández M, Stein A,
Hibbert RG, Heidebrecht T, Aloy P, Medema RH and Perrakis A:
Structure of the FoxM1 DNA-recognition domain bound to a promoter
sequence. Nucleic Acids Res. 38:4527–4538. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jonker HR, Baumann S, Wolf A, Schoof S,
Hiller F, Schulte KW, Kirschner KN, Schwalbe H and Arndt HD: NMR
structures of thiostrepton derivatives for characterization of the
ribosomal binding site. Angew Chem Int Ed Engl. 50:3308–3312. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Malde AK, Zuo L, Breeze M, Stroet M, Poger
D, Nair PC, Oostenbrink C and Mark AE: An automated force field
topology builder (ATB) and repository: Version 1.0. J Chem Theory
Comput. 7:4026–4037. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Canzar S, El-Kebir M, Pool R, Elbassioni
K, Mark AE, Geerke DP, Stougie L and Klau GW: Charge Group
Partitioning in Biomolecular Simulation. J Comput Biol. 20:188–198.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Koziara KB, Stroet M, Malde AK and Mark
AE: Testing and validation of the Automated Topology Builder (ATB)
version 2.0: Prediction of hydration free enthalpies. J Comput Aid
Mol Des. 28:221–233. 2014. View Article : Google Scholar
|
|
30
|
Trott O and Olson AJ: Software news and
update AutoDock Vina: Improving the speed and accuracy of docking
with a new scoring function, efficient optimization, and
multithreading. J Comput Chem. 31:455–461. 2010.PubMed/NCBI
|
|
31
|
Abraham MJ, Murtola T, Schulz R, Páll S,
Smith JC, Hess B and Lindahl E: GROMACS: High performance molecular
simulations through multi-level parallelism from laptops to
supercomputers. SoftwareX 1–2. 19–25. 2015. View Article : Google Scholar
|
|
32
|
Oostenbrink C, Villa A, Mark AE and Van
Gunsteren WF: A biomolecular force field based on the free enthalpy
of hydration and solvation: The GROMOS force-field parameter sets
53A5 and 53A6. J Comput Chem. 25:1656–1676. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Berendsen HJC, Postma JPM, Gunsteren WF
and Hermans J: Interaction models for water in relation to protein
hydration. Intermolecular forces. Pullman B: 14. Springer; Berlin:
pp. 331–334. 1981, View Article : Google Scholar
|
|
34
|
Bussi G, Donadio D and Parrinello M:
Canonical sampling through velocity rescaling. J Chem Phys.
126:2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bussi G, Zykova-Timan T and Parrinello M:
Isothermal-isobaric molecular dynamics using stochastic velocity
rescaling. J Chem Phys. 130:0741012009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Parrinello M and Rahman A: Polymorphic
transitions in single crystals: A new molecular dynamics method. J
Appl Phys. 52:1981. View Article : Google Scholar
|
|
37
|
Darden T, York D and Pedersen L: Particle
mesh Ewald: An N I log(N) method for Ewald sums in large systems. J
Chem Phys. 98:100891993. View Article : Google Scholar
|
|
38
|
Essmann U, Perera L and Berkowitz ML: A
smooth particle mesh Ewald method. J Chem Phys. 103:85771995.
View Article : Google Scholar
|
|
39
|
Wong-Ekkabut J and Karttunen M: Assessment
of common simulation protocols for simulations of nanopores,
membrane proteins, and channels. J Chem Theory Comput. 8:2905–2911.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hess B: P-LINCS: A parallel linear
constraint solver for molecular simulation. J Chem Theory Comput.
4:116–122. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Humphrey W, Dalke A and Schulten K: VMD:
Visual molecular dynamics. J Mol Graph. 14:33–38, 27-38. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Halasi M and Gartel AL: A novel mode of
FoxM1 regulation: Positive auto-regulatory loop. Cell Cycle.
8:1966–1967. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Leung TW, Lin SS, Tsang AC, Tong CS, Ching
JC, Leung WY, Gimlich R, Wong GG and Yao KM: Over-expression of
FoxM1 stimulates cyclin B1 expression. FEBS Lett. 507:59–66. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liao GB, Li XZ, Zeng S, Liu C, Yang SM,
Yang L, Hu CJ and Bai JY: Regulation of the master regulator FOXM1
in cancer. Cell Commun Signal. 16:572018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kong X, Li L, Li Z, Le X, Huang C, Jia Z,
Cui J, Huang S, Wang L and Xie K: Dysregulated expression of FOXM1
isoforms drives progression of pancreatic cancer. Cancer Res.
73:3987–3996. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cheng XH, Black M, Ustiyan V, Le T,
Fulford L, Sridharan A, Medvedovic M, Kalinichenko VV, Whitsett JA
and Kalin TV: SPDEF inhibits prostate carcinogenesis by disrupting
a positive feedback loop in regulation of the Foxm1 oncogene. PLoS
Genet. 10:e10046562014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Korver W, Roose J and Clevers H: The
winged-helix transcription factor Trident is expressed in cycling
cells. Nucleic Acids Res. 25:1715–1719. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sanders DA, Gormally MV, Marsico G,
Beraldi D, Tannahill D and Balasubramanian S: FOXM1 binds directly
to non-consensus sequences in the human genome. Genome Biol.
16:1302015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tabatabaei-Dakhili SA, Aguayo-Ortiz R,
Domínguez L and Velázquez-Martínez CA: Untying the knot of
transcription factor druggability: Molecular modeling study of
FOXM1 inhibitors. J Mol Graph Model. 80:197–210. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Van Der Spoel D, Lindahl E, Hess B,
Groenhof G, Mark AE and Berendsen HJ: GROMACS: Fast, flexible, and
free. J Comput Chem. 26:1701–1718. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Tassi RA, Todeschini P, Siegel ER, Calza
S, Cappella P, Ardighieri L, Cadei M, Bugatti M, Romani C, Bandiera
E, et al: FOXM1 expression is significantly associated with
chemotherapy resistance and adverse prognosis in non-serous
epithelial ovarian cancer patients. J Exp Clin Cancer Res.
36:632017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Ye H, Kelly TF, Samadani U, Lim L, Rubio
S, Overdier DG, Roebuck KA and Costa RH: Hepatocyte nuclear factor
3/fork head homolog 11 is expressed in proliferating epithelial and
mesenchymal cells of embryonic and adult tissues. Mol Cell Biol.
17:1626–1641. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yao KM, Sha M, Lu Z and Wong GG: Molecular
analysis of a novel winged helix protein, WIN. Expression pattern,
DNA binding property, and alternative splicing within the DNA
binding domain. J Biol Chem. 272:19827–19836. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gormally M, Marsico G, Rai G, Lowe C,
Thomas C, Maloney D, Michael S, Matak-Vincovic D, Jadhav A,
Simeonov A and Balasubramanian S: Abstract 3088: Transcription
factor as target: Novel small molecule inhibits FOXM1 DNA binding
and oncogenic gene products. Cancer Res. 76:3088. 2016.PubMed/NCBI
|
|
55
|
Gormally MV, Dexheimer TS, Marsico G,
Sanders DA, Lowe C, Matak-Vinković D, Michael S, Jadhav A, Rai G,
Maloney DJ, et al: Suppression of the FOXM1 transcriptional
programme via novel small molecule inhibition. Nat Commun.
5:51652014. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Marsico G and Gormally MV: Small molecule
inhibition of FOXM1: How to bring a novel compound into genomic
context. Genom Data. 3:19–23. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Marklund EG, Mahmutovic A, Berg OG, Hammar
P, van der Spoel D, Fange D and Elf J: Transcription-factor binding
and sliding on DNA studied using micro- and macroscopic models.
Proc Natl Acad Sci USA. 110:19796–19801. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang X, Cheng L, Minn K, Madan R, Godwin
AK, Shridhar V and Chien J: Targeting of mutant p53-induced FoxM1
with thiostrepton induces cytotoxicity and enhances carboplatin
sensitivity in cancer cells. Oncotarget. 5:11365–11380.
2014.PubMed/NCBI
|