Multiple myeloma (MM) is the second most common
haematological malignancy worldwide. However, MM is incurable for
the majority of patients (1).
Malignant plasma cells expand in the bone marrow, in which they
constantly develop, acquire resistance to apoptosis and eventually
cause patients to exhibit symptomatic MM (CRAB: High calcium, renal
impairment, anemia and bone lesions) (2). In most cases, MM is preceded by an
asymptomatic premalignant disease known as monoclonal gammopathy of
undetermined significance (MGUS) (3).
It has been reported that ~2% of the population over the age of 50
are affected by MGUS, with 1% of cases progressing to MM every year
(4).
MM acts as osteolytic bone metastasis, and this
behavior differs from that of other hemotologic malignancies that
infiltrate the bone marrow. The bone marrow is a semi-solid tissue
that occupies cavities within spongy or cancellous bone. Bone
marrow is composed of interstitial cells and blood vessels that are
associated with haematopoiesis, bone marrows primary function
(5). The bone marrow microenvironment
(BMME), which is also known as the bone marrow niche, consists of a
cellular and non-cellular component. The cellular component
includes stromal cells, endothelial cells, osteoclasts (OCs),
osteoblasts (OBs) and immune system cells, and the non-cellular
component includes the extracellular matrix (ECM) and liquid
milieu, which contains chemokines, cytokines and growth factors
(6). The BMME has been previously
demonstrated to be a scaffold for haematopoietic stem and
progenitor cells (HSPCs) (7–9). Further studies (10–12) have
reported that the BMME serves an important role in haematopoiesis
and in neoplastic disease. The stability of the BMME is essential
for the maintenance of normal cell proliferation, differentiation,
metabolism and mobilization. An abnormal BMME has been revealed to
lead to cell lesions or tumorigenesis. An abundance of research has
been undertaken into elucidating the biological features of MM, and
the association with the BMME (13,14). It
can be concluded from these studies that the BMME serves a key role
in the differentiation, proliferation and drug resistance of
malignant plasma cells. These studies also provide preclinical
evidence that MM cells and bone marrow stromal cells (BMSCs) can be
targeted as antitumor strategies and used in disease treatment
(15). The present article elaborates
on the complex interaction that exists between the BMME and MM.
The components of the BMME, especially the BMSCs and
the extracellular matrix (ECM) proteins, serve a key role in the
pathogenesis of MM (16) (Fig. 1). The adhesion that occurs between MM
plasma cells and both BMSCs and ECM proteins has been
well-established. These adhesions can influence the growth,
proliferation, invasion and drug resistance of MM cells and cause
angiogenesis and lytic bone lesions (13,17). The
very late antigen 4 (VLA-4), which is located on the MM plasma cell
surface, mediates the binding between tumor cells and ECM proteins,
including type I collagen and fibronectin, and can bind to BMSC
vascular cell adhesion molecule-1 (VCAM-1). The tumor cells that
adhere to BMSCs can stimulate numerous pathways, and result in the
upregulation of cytokines, which regulate the cell, and
antiapoptotic proteins (18),
including interleukin-6 (IL-6), RANKL and Activin A (19,20). The
interaction between MM and BMSC cells can be mediated via the
Notch-signaling pathway. The notch-signaling pathway in MM and
BMSCs can be stimulated by the interaction with Notch-Notch ligand,
and can lead to the secretion of IL-6, vascular endothelial growth
factor (VEGF) and insulin-like growth factor 1 (IGF-1).
Additionally, the adhesion of MM cells to BMSCs also induces
NF-κB-dependent transcription and IL-6 secretion. IL-6 promotes MM
cell production and secretion of VEGF, and regulates malignant
plasma cell apoptosis and OC differentiation (21). Conversely, the inhibition of NF-κB
activity abrogates this response (22). It has been previously demonstrated
that in patients with MM, exosomes may be released by BMSCs and
transferred to MM cells, leading to MM progression and the
induction of drug resistance (23,24).
Recently, a study has demonstrated that MM exosomes that are
derived from BMSCs can promote BMSC growth and angiogenesis by
modifying the BM microenvironment. This modification can increase
MM cell proliferation, disease progression and interfere with drug
resistance (25). Although the
specific mechanism governing this remains unclear, these findings
revealed that cell exosomes may be associated with the
intercellular transfer of genetic information in clonal plasma cell
disorders (24). Understanding MM
cell exosomes and their associated miRNA may enable the discovery
of a novel therapeutic strategy for the treatment of MM.
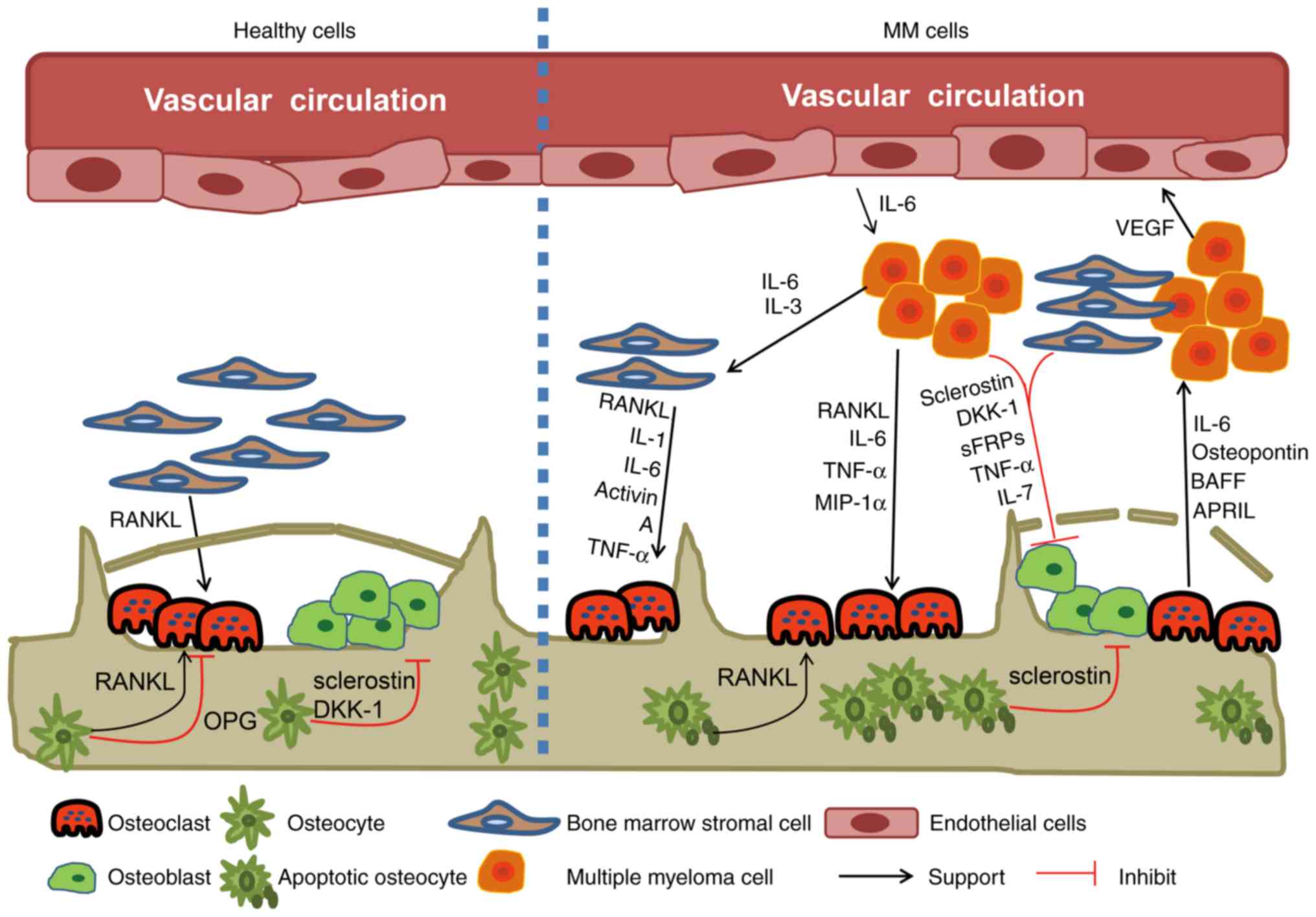
Normal bone is composed of mineralized and organic
components. In a normal BM microenvironment, osteocytes, OCs and
OBs sustain homeostasis by maintaining a balance between bone
formation and bone resorption. Osteocytes constitute the majority
of all bone cells, and OCs and OBs constitute <10% (37). The balance between the generation of
new OBs and OCs is disrupted in MM, leading to the rapid
development of bone lesions and bone resorption (38–40), which
are the major primary clinical features of this disease (41). Patients with MM exhibit a decreased
number of live osteocytes due to the presence of bone lesions,
which may be involved in MM-induced OC formation (42). Bone destruction caused by osteoclast
activation usually occurs close to MM plasma cells, and not in
normal bone marrow. A number of cytokines are associated with
osteoclast activation, including RANKL, interleukin-3 (IL-3) and
MIP-1a (6). IL-3 expression
significantly increases in patients with MM, and this can
upregulate OC activity and inhibit osteogenic protein-2 (bone
morphogenetic protein-2), which subsequently inhibits OB
differentiation (43,44). OCs produce OPN and IL-6, and the
adhesion of OCs to MM plasma cells has been demonstrated to
increase IL-6 production, a key growth factor involved in MM
progression (45). Osteoclastic
resorption induces the release of matrix-bound factors which
promote the development of MM. This process has been named ‘the
vicious cycle’ (46). Additionally,
as a member of the tumor necrosis factor (TNF) family, RANK ligand
has been associated with increased osteoclastogenesis, which is
also involved with MM. In the bone marrow, the adhesion of MM
plasma cells and neighboring BMSCs leads to an increase in RANKL
expression. RANKL binding to its associated receptor indirectly
increases osteoclast precursor cell activity, which increases OC
differentiation via the c-Jun N-terminal kinase pathway and NF-κB
(47). Osteoprotegerin (OPG) is a
soluble decoy receptor that is attracted to OBs and stromal cells.
The RANKL and OPG molecules, which are mutually competitive
receptors, serve an important role in the regulation of bone
resorption (48). In patients with
MM, higher RANKL expression and decreased OPG is exhibited in the
bone marrow microenvironment, which leads to increased osteoclast
activation and bone osteolysis. RANKL is the most important protein
associated with the prolonged survival of MM plasma cells. This is
due to RANKL inhibiting osteoclast apoptosis. It has been
previously demonstrated that obstructing RANKL, with its soluble
form, RANK, can accelerate bone loss and regulate tumor pressure in
MM animal models (49). Recently, it
has been reported that OC activity is associated with the
excitation of resting MM cells in bone marrow. These results
revealed that OCs may serve an important role in remodeling the
terminal ‘vicious cycle’, as well as inducing tumor growth and
increasing the possibility of relapse (50). Therefore, OCs are considered to be
associated with tumor growth in MM (51).
It is not yet clear what effects osteoblasts exhibit
on MM cells. In patients with MM, OC proliferation and
differentiation is inhibited due to bone cell abnormalities
(52). Numerous factors are
associated with the suppression of osteoblast activity and new bone
growth, including the secretion of DKK1, soluble Wnt inhibitor
frizzled-related protein 2 (sFRP-2) and transforming growth
factor-β (TGF-β) (53). DKK1, a
Wnt-signaling antagonist, is secreted by MM cells and is
overexpressed in patients with MM who exhibit lytic bone lesions
(54). A previous study has
demonstrated that the inhibition of DKK1 can reduce bone resorption
and tumor burden in a severe combined immunodeficient 11-rabbit
(SCID-rab) myeloma model (55).
Additionally, OB differentiation has also been revealed to be
suppressed by sFRP-2, which is produced by the majority of MM
cells, including in RPMI8226 and U266 cell lines (56). A similar effect has been indicated
subsequent to the induction of osteocyte overexpressing sclerostin
(SOST) (57). However, intriguing
findings are emerging in this field of research (58). Whether OBs are inhibitory or
supportive in MM has been revealed to depend on the source of the
MM cells (59). A previous study has
indicated that OBs contributed to MM cell survival and growth, and
also increased osteolysis development, which enhanced MM
pathogenesis (19). The capacity of
OBs to secrete IL-6 close to MM plasma cells may result in this
phenomenon and promote increased bone resorption and MM cell
proliferation (60). It has also been
reported that OBs serve a key role in activating MM cell survival,
and this may be due to the release of OPG inhibiting MM cell
apoptosis, which is mediated by TNF-related apoptosis-inducing
ligand (TRAIL) (61). Yaccoby et
al demonstrated that the majority of MM plasma cells obtained
from active MM patients, in triple-cultures with OBs+OCs, exhibited
fewer viable cells compared with cells co-cultured with OCs and MM
plasma cells alone. In myelomatous severe combined immunodeficiency
(SCID) exhibited in mice implanted with fetal human bone (SCID-hu
mouse model), OBs were revealed to inhibit tumor growth and
increase bone mineral density (59).
Overall, further research is required to determine an efficient
method to target OBs and alter tumor cell survival and growth. This
could be used as a novel therapeutic strategy in the treatment of
patients exhibiting bone disease and tumor growth (46). Future research should aim to elucidate
the mechanisms behind the interaction between OBs and MM plasma
cells. Elucidating the mechanisms behind this interaction may
support the potential use of osteoblastic mediators in the
treatment of MM.
The association between cytokines, proteolytic
enzymes, chemokines and adhesion molecules provides survival
signals that are used in MM plasma cells and are associated with
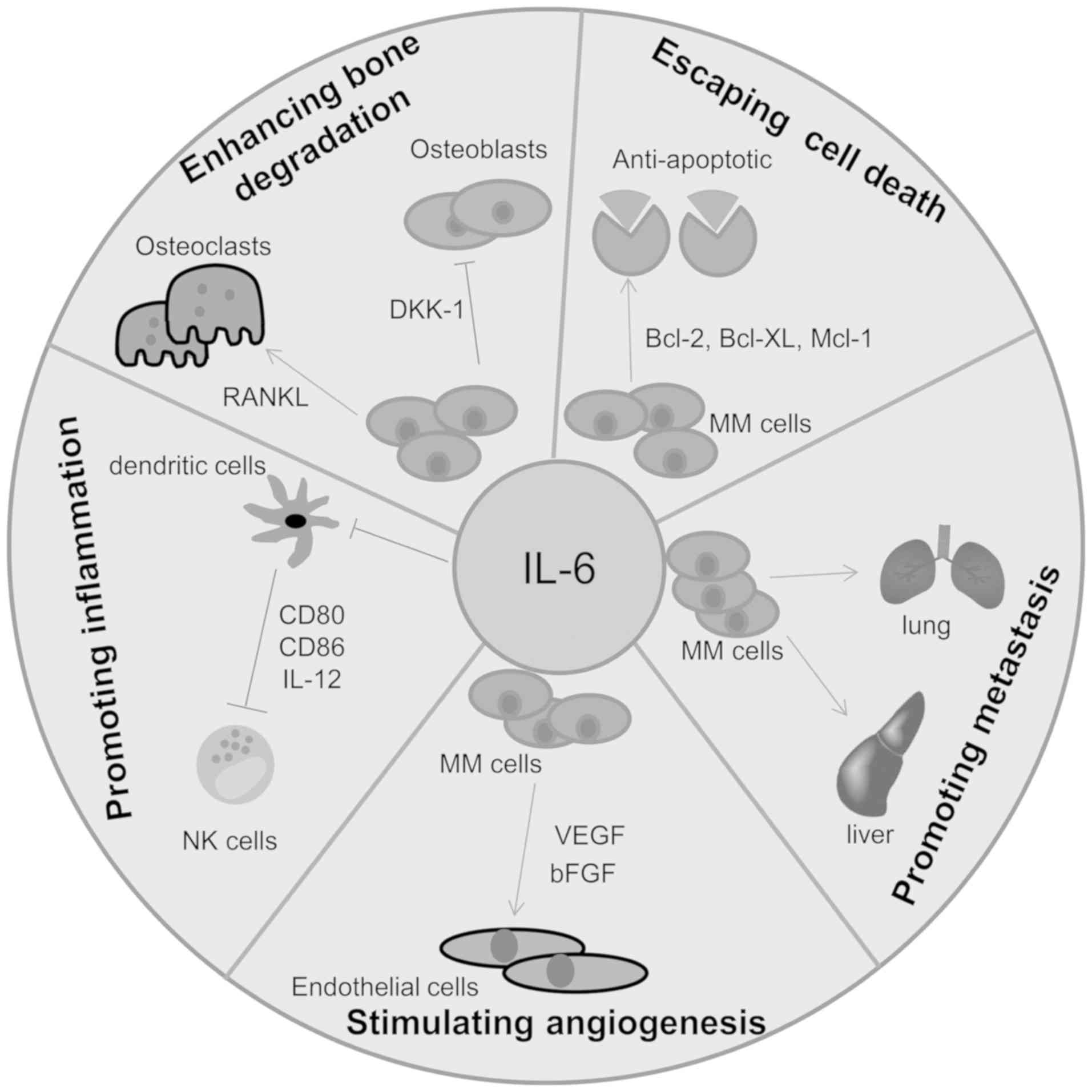
the development of MM drug resistance. Among these, IL-6 has been
reported to serve a key role in a variety of tumor activities,
including cell migration, invasion, tumor growth, apoptosis,
angiogenesis, the differentiation of tumor cells and inflammation
or infection (62,63). IL-6 was initially cloned as a B cell
stimulating factor (BSF) and designated interferon β2. IL-6 has
been demonstrated to stimulate cytotoxic T cells and promote
osteoclast precursor cells to differentiate into mature and active
osteoclasts (64). In fact, the
cellular sources of IL-6 in MM-infiltrated bone marrow are highly
controversial, with their origin usually described as paracrine or
autocrine (65). Initially, IL-6 was
regarded as an autocrine factor produced by certain MM cells.
However, the most recent consensus opinion is that the BM
environment constitutes the main source of this cytokine (Fig. 2). Several recent studies have also
confirmed the paracrine origin of IL-6 in murine models of
plasmacytomas and MM (66,67). Frank myeloma cells and myeloma
progenitors in the BMME have been suggested to use two sources of
IL-6 to promote tumor maintenance and proliferation in vivo
(68). Numerous molecules and
cytokines can upregulate the secretion of IL-6 derived from BMSCs.
It has been previously reported that IL-6 can be detected in the
supernatant of MM cells using stimulation with anti-CD40 antibody
(69,70). Additionally, IL-1β has also been
indicated to be a major cytokine that contributes to the production
of BMSC IL-6 via the paracrine loop, which promotes the growth and
survival of MM cells (71). After
binding to its receptor (IL-6R), IL-6 triggers the activation of
the PI3K/Akt/mTOR (72–74) and Ras/Raf/MEK/Erk (35) signaling pathways. IL-6 has also been
revealed to serve a key role as an MM growth factor by stimulating
IL-6R and triggering the phosphorylation of STAT3 via JAK1
(75). The Janus kinase (JAK)-signal
transducer and activator of transcription (STAT) pathway was
initially revealed to be associated with interferon-α (IFN-α),
IFN-γ and IL-6-mediated downstream signaling (76). Blocking the IL-6R/STAT3 pathway has
been indicated to induce apoptosis in MM cell line NCI-H929
(77). NF-κB has been demonstrated to
be associated with cytokine- and adhesion-mediated IL-6
upregulation. Therefore, the inhibition of NF-κB can prohibit the
secretion of IL-6 (78). In previous
studies, it was suggested that the non-specific inhibition of IL-6
could be performed due to its highly pleiotropic effects (with
drugs including lenalidomide and associated immunomodulatory
drugs), or IL-6 could be specifically targeted in MM and BMSCs
(79). Presently, it is well-known
that effective inhibition methods include the use of monoclonal
antibodies (mAbs) to IL-6 (siltuximab) (80–82) or
IL-6R (tocilizumab), or recombinant proteins, which act as IL-6R
antagonists (83). Small molecules
that inhibit the cellular IL-6 signaling pathway or cross-talked
pathways in MM can also be used (84). Additional therapeutic treatments are
performed using compounds, which either block the biochemical
pathways that promote IL-6 signaling directly, or activate
biochemical pathways which can inhibit IL-6 signaling indirectly
(85). The study of mice models is
invaluable in providing key evidence for the potential preclinical
use of IL-6-targeted therapies for MM treatment.
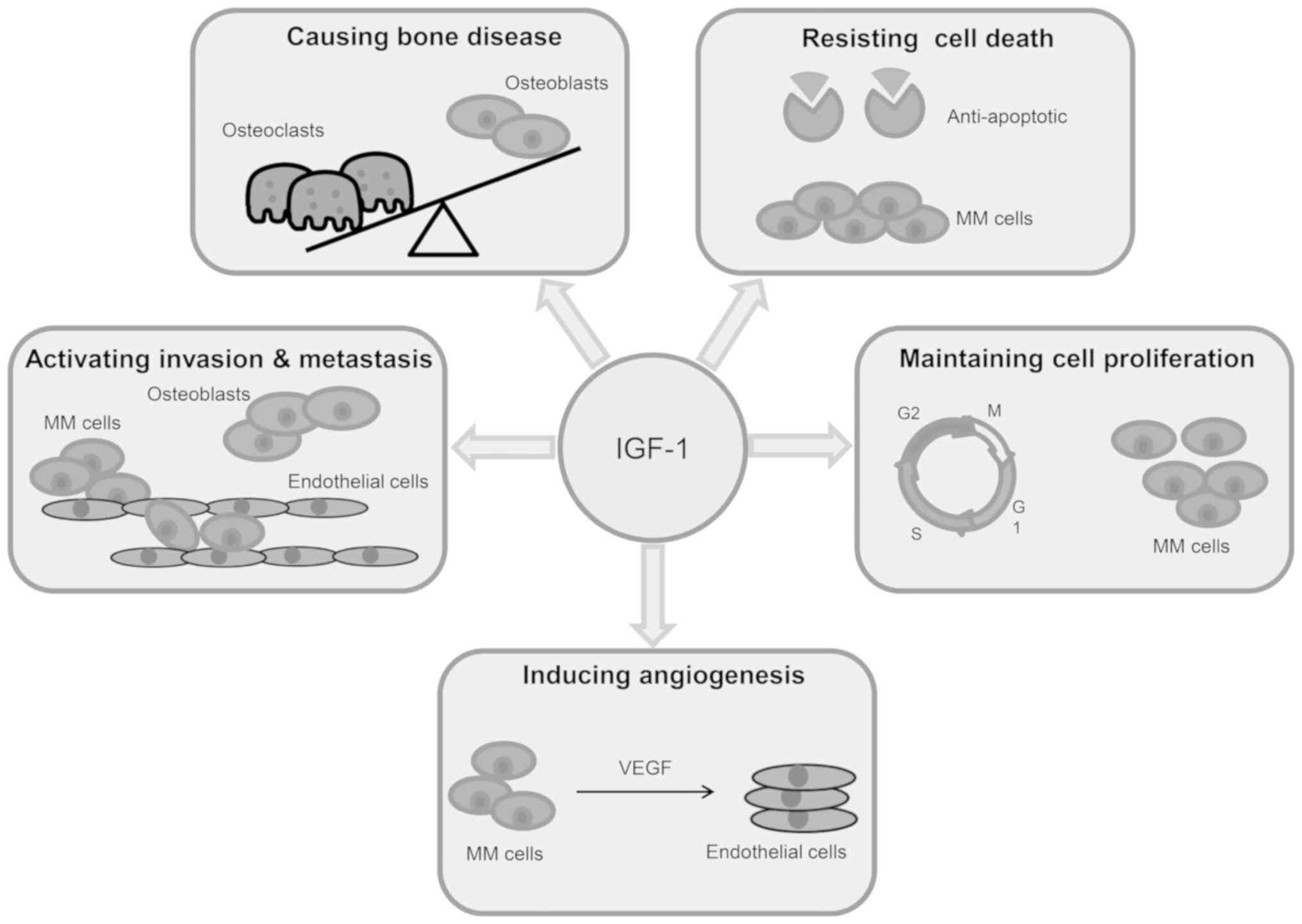
Within the past decade, the insulin-like growth
factor (IGF) system has been demonstrated to serve a key role in
the establishment and progression of MM (86,87)
(Fig. 3). It has been previously
reported that IL-6 inhibition is not sufficient to induce effects
in MM. Therefore, an increased interest has emerged into the IGF
system, which can compensate IL-6 signaling and may induce the
proliferation and survival of IL-6-independent MM cells (88). This suggests that IGF-1R inhibitors
may contribute to the drug resistance exhibited, both in
vitro and in vivo, to well-known anti-myeloma medicines
(89,90). In the extracellular environment, the
IGF system is composed of six high-affinity insulin-like growth
factor binding proteins (IGFBP1-6), IGF1, IGF2, IGF-1R and a minor
free fraction of total IGF (91). The
binding of free IGF to its receptor initiates a signaling cascade
that results in the proliferation and viability of MM cells. In MM,
IGF-1 is largely available in the BMME due to its secretion from
BMSCs and OBs. It has been previously revealed that, through the
activation of IGF-1R, IGF-1 serves an important role in MM cell
growth, increases DNA synthesis and has been demonstrated to
enhance the survival and proliferation of some MM cell lines.
However, anti-IGF-IR blocking antibody use can inhibit the
aforementioned effects (92,93). A number of studies have indicated that
MM cells exhibit high IGF-1 and IGF-1R expression and, through an
autocrine cascade, IGF-1/IGF-1R signaling contributes to the
survival, growth and drug resistance in MM cells (86,94).
Furthermore, in MM cell lines, it has been revealed that autocrine
IGF-1 is an important self-clonogenic growth factor (95). Recent research has also demonstrated
that IGF-1 can reverse the inhibition of cell proliferation induced
by the knockdown of EEN. EEN, an endocytosis-regulating molecule,
promotes the proliferation and viability of MM cells by stimulating
IGF-1 secretion (96). In MM, the
role of IGF-1/IGF-1R signaling in the inhibition of apoptosis and
the induction of tumor cell proliferation occurs via activation of
the PI3K/AKT and MAPK pathways (97).
Which of these two activities (apoptosis or cell proliferation) is
initiated depends on which pathway is induced. For example,
apoptosis can be inhibited by IGF-IR through the activation of the
PI3K/AKT pathway, which inhibits the ability of mitochondria to
release cytochrome c. Subsequent to this, pro-apoptotic
protein BAD is phosphorylated by activated AKT, separated from
anti-apoptotic protein Bcl-2 and sequestered by the adaptor protein
14-3-3, leading to the inhibition of apoptosis in PCM cells
(72,98). Additionally, IGF-1/IGF-1R is
associated with MM cell proliferation, including migration,
adhesion and invasion. These processes differentiate monoclonal
malignant plasma cells in the BM microenvironment (99). IGF-1 serves a chemo-attractant role in
murine 5T33MM cells and has been revealed to enhance cell adhesion
to the extracellular matrix glycoprotein fibronectin in
vitro by activating IGF-IR downstream targets, such as the
PI3K/AKT pathway (100). IGF-1R is
also upregulated in MM cells subsequent to interaction with the
BMME (101). It has also been
reported that enhanced IGF-1 signaling, through increased IGF-1
secretion and IGF-1R activation, is associated with bortezomib
drug-resistance. This resistance may be overcome by inhibiting
downstream targets such as PI3K and mTOR (90). IGF-1 signaling promotes the activation
of AKT. However, the AKT signaling pathway is not the sole
downstream kinase that can be activated by IGF-1. The MAPK pathway
has been revealed to be activated by IGF-1 to promote MM
proliferation in six MM cell lines (98). Conversely, one study reported that the
MAPK pathway could only be activated in IL-6-dependent MM lines.
Numerous downstream signaling pathways may also be activated by
IGF-1, including the Janus kinase pathway (102). Therefore, further research is
required to elucidate which additional signaling pathways lie
downstream of IGF-1.
An increase in BM vascularization has been
demonstrated to be positively associated with an adverse prognosis
in MM patients (103). New blood
vessel growth enhances oxygen and nutrient transport to cells and
can promote tumor growth. VEGF is a highly conservative homodimeric
glycoprotein with a relative molecular mass of 45 kDa. VEGF is the
only mitogen that is associated with endothelial cells (104). VEGF is a peptide growth factor that
belongs to the platelet-derived growth factor (PDGF) superfamily,
which exhibits high homology across species. VEGF has been
previously described as a vascular permeability factor (VPF), and
is now denoted VEGF-A (105). VEGF
serves a key pro-angiogenic role in MM and is induced by MM cells
and BMSCs (99). The secretion of
VEGF can be stimulated by numerous cytokines and cell growth
factors, including IL-6, bFGF, TGF-β and tumor necrosis TNF-α. VEGF
serves a role in the formation of new blood vessels by binding to
its receptors VEGFR-1 and VEGFR-2. This induces the survival,
proliferation, migration and differentiation of BMSCs and ECs,
through a number of signaling pathways, including Ras, GAP,
PI3K/Akt, STAT and MEK/ERK (103).
As aforementioned, angiogenesis is the formation of new vasculature
from pre-existing vasculature, and has been demonstrated to be
essential for tumor growth, invasion, and metastasis in a variety
of cancer types (106). If the pro-
and anti-angiogenic balance is broken and angiogenesis is favored,
MGUS can turn into MM (107–109). VEGF has been revealed to be
crucially implicated during osteoclastogenesis in MM (110). Research has also revealed that when
VEGF binds to the VEGFR-1 receptor, which is expressed in OCs, OC
cells are stimulated. However, OCs can also be activated
indirectly, since MM cells can secrete VEGF subsequent to IL-6
stimulation (111). VEGF directly
enhances osteoclastic bone resorption and the survival of mature
OCs (33). Furthermore, VEGF is also
correlated with high expression and activation of adhesion proteins
and has been demonstrated to activate the secretion of proteases
and ECM components, which can subsequently remodel ECM structures
and modulate tumor cell-tumor cell and tumor cell-ECM
interactions.
In recent years, rising evidence has emphasized the
role of exosomes in the cross-talk between MM cells and the BMME,
and exosomes are now considered to be associated with cell to cell
communication (24,112,113).
Exosomes, a sub-fraction of extracellular vesicles (EVs), range in
size from 30–100 nm. They transport a large variety of mRNAs,
proteins and growth factors to target cells, and are secreted
either via endocytosis, which occurs in MM cells and cells
composing the BMME, or by direct fusion with the cell membrane
(114–116). MicroRNAs (miRNAs), a class of small
non-coding RNAs, are transported by exosomes and can suppress
target gene expression at the post-transcriptional/translational
level via hybridization with mRNA transcript sequences (117). It has also been reported that
exosomes are associated with various biological processes,
including inflammation, embryonic development, hematopoiesis, the
immune response and tumorigenesis (118). In MM cells, upregulated miR-146a
increases the expression and release of cytokines, including IL-6,
CXCL1, IP-10 and CCL5, which enhance MM cell viability and
migration (119). miR-146a has also
been demonstrated to activate numerous signaling pathways,
including NF-κB, Notch and JAK/STAT, and in turn, the Notch pathway
contributes to the secretion of cytokines that are induced by
miR-146a (120). Previous studies
have indicated that MM cell-derived exosome treatment can increase
the expression of OC-specific markers, including tartrate-resistant
acid phosphatase (TRAP), cathepsin K (CTSK) and matrix
metallopeptidase 9 (MMP9) (121).
Amphiregulin (AREG), the epidermal growth factor receptor (EGFR)
ligand, is expressed by MM cells and is associated with MM-derived
exosomes, which participate in MM-induced osteoclastogenesis
(122). AREG arises from the
activation of the EGFR pathway and, subsequent to MM-derived
exosome treatment, its presence is often confirmed by a significant
increase in SNAIL mRNA, which is a downstream target of EGFR
(122). In vitro studies have
indicated that osteoblast differentiation and functionality can be
inhibited by exosomes derived from 5TGM1 cells. This study also
indicated that DKK1 expression was upregulated in MC3T3-E1 cells
containing 5TGM1 exosomes (123).
Aberrant angiogenesis has been demonstrated to be associated with
exosomes in MM-infiltrated bone marrow (124). A previous study revealed that
exosomes that are derived from hypoxia-resistant cells
significantly increased angiogenesis, and exosomal miR-135b was
considered to be a specific miRNA that is associated with chronic
hypoxia (125). Numerous animal
model studies have demonstrated that exosomes that are derived from
normal BMSCs are effective in the treatment of cardiovascular
disease (126) and kidney injury
(127). Antitumor drugs, such as
doxorubicin, could potentially be loaded into liposomes to improve
osteosarcoma chemotherapy treatment (128). Additionally, liposomal doxorubicin
could be used in conjunction with bortezomib or dexamethasone in
the treatment of patients exhibiting MM (129). A recent study also revealed that in
endothelial cells, miR-340 can inhibit angiogenesis via the
hepatocyte growth factor/c-MET (HGF/c-MET) signaling pathway
(130). Therefore, it can be
expected that additional RNA or protein components that are
associated with exosomes derived from MM cells or BMSCs exist, and
are associated with the cell-cell communication among them.
Research into exosomes provides important insights into the methods
by which MM can be promptly diagnosed. Exosomal miRNAs, which are
derived from young BMSCs, can be utilized in the formation of a
novel strategy for MM treatment.
The present review investigated the strong
interaction exhibited between MM cells and the surrounding BMME.
The present review demonstrated that OBs, OCs, BMSCs and immune
cells serve key roles in the promotion of MM cell growth and
survival, inhibit autonomous or drug-induced MM cell apoptosis and
therefore contribute to the development of osteolysis. Therefore,
these cells should be considered in research into potential new
treatment strategies for MM. The more research that investigates
the interactions between healthy cells in the BMME, the better
understanding will be gained into how MM disrupts the BMME. The
development of novel MM treatment targets is ongoing. In recent
years, the introduction of agents targeting MM cells and their
interactions with the BMME, including proteasome inhibitors
(bortezomib and carfilzomib) and immunomodulatory agents
(thalidomide, lenalidomide and pomalidomide), has reduced mortality
in patients with MM. Although bortezomib has been indicated to
inhibit VEGF, IL-6 and IGF-1 secretion in BMSC and endothelial
cells (36,131), and the OB differentiation and OC
apoptosis (132), further
elucidation into the molecular mechanisms of MM-bone interactions
and the MM niche are required to determine novel target approaches
that exhibit beneficial effects on bone disease and MM tumor
progression.
Currently, controversy exists as to whether a
constant disease control approach should be pursued, which impacts
patient quality of life and overall survival, or whether MM should
be treated with an aggressive multidrug strategy that exhibits a
complete recovery. Younger patients with poor prognosis may benefit
more from the aggressive multidrug strategies as these patients are
more likely to benefit long-term. However, for elderly, frail or
non-high-risk patients who are newly diagnosed with MGUS, the
treatment strategy should be to control the disease. This can be
done by achieving stable renal function, which exhibits stable
non-progression of bone disease, low toxic effects, high quality of
life and increased survival rate. Future study should focus on
developing combination therapies for a variety of patient subgroups
to ensure patients in the low-risk stage or those who do not
tolerate concentrated interventions will not be overtreated.
In conclusion, MM remains incurable and exhibits a
complex pathophysiology. The interaction between MM plasma cells
and BM has been demonstrated to serve a crucial role in the
pathogenesis and progression of MM. The current standard treatment,
and the majority of active anti-MM drugs, work against MM cells and
BMME. Through targeting these diseased cells and their
microenvironment, novel treatment strategies may be determined that
can improve the outcome of this incurable blood cancer.
The authors would like to thank all Lab members, and
CYG and YY for collecting literature and discussion.
The present study was supported by the National
Natural Science Foundation of China (nos. 81670200, 81770220 and
81600177; to YY and CG); the 2016 Outstanding Youth Fund of Jiangsu
Province BK20160048 (to YY); the Natural Science Foundation of
Jiangsu Province BK20161041 (to CG); the National Key Research and
Development Program-Precision Medicine sub-program 2016YFC0905900
(to YY); The Priority Academic Program Development of Jiangsu
Higher Education Institutions for Chinese Medicine; the Innovation
Team of Six Talent Peaks Project in Jiangsu Province TD-SWYY-015
(to CG).
Not applicable.
YY, CG and ZF were involved in the conception of the
review. JH and RW wrote the manuscript. JQ analyzed the data and
plotted the graphs. CG and RW edited the manuscript. All authors
have read and approved the final manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Rajkumar SV: Myeloma today: Disease
definitions and treatment advances. Am J Hematol. 91:90–100. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Landgren O, Kyle RA, Pfeiffer RM, Katzmann
JA, Caporaso NE, Hayes RB, Dispenzieri A, Kumar S, Clark RJ, Baris
D, et al: Monoclonal gammopathy of undetermined significance (MGUS)
consistently precedes multiple myeloma: A prospective study. Blood.
113:5412–5417. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kyle RA, Durie BG, Rajkumar SV, Landgren
O, Blade J, Merlini G, Kröger N, Einsele H, Vesole DH, Dimopoulos
M, et al: Monoclonal gammopathy of undetermined significance (MGUS)
and smoldering (asymptomatic) multiple myeloma: IMWG consensus
perspectives risk factors for progression and guidelines for
monitoring and management. Leukemia. 24:1121–1127. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kunisaki Y, Bruns I, Scheiermann C, Ahmed
J, Pinho S, Zhang D, Mizoguchi T, Wei Q, Lucas D, Ito K, et al:
Arteriolar niches maintain haematopoietic stem cell quiescence.
Nature. 502:637–643. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Manier S, Sacco A, Leleu X, Ghobrial IM
and Roccaro AM: Bone marrow microenvironment in multiple myeloma
progression. J Biomed Biotechnol. 2012:1574962012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Crisan M and Dzierzak E: The many faces of
hematopoietic stem cell heterogeneity. Development. 143:4571–4581.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pang WW, Schrier SL and Weissman IL:
Age-associated changes in human hematopoietic stem cells. Semin
Hematol. 54:39–42. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shiozawa Y, Pedersen EA, Havens AM, Jung
Y, Mishra A, Joseph J, Kim JK, Patel LR, Ying C, Ziegler AM, et al:
Human prostate cancer metastases target the hematopoietic stem cell
niche to establish footholds in mouse bone marrow. J Clin Invest.
121:1298–1312. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Corcoran KE, Patel N and Rameshwar P:
Stromal derived growth factor-1alpha: Another mediator in
neural-emerging immune system through Tac1 expression in bone
marrow stromal cells. J Immunol. 178:2075–2082. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shiozawa Y, Pedersen EA, Patel LR, Ziegler
AM, Havens AM, Jung Y, Wang J, Zalucha S, Loberg RD, Pienta KJ and
Taichman RS: GAS6/AXL axis regulates prostate cancer invasion,
proliferation, and survival in the bone marrow niche. Neoplasia.
12:116–127. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chinni SR, Sivalogan S, Dong Z, Filho JC,
Deng X, Bonfil RD and Cher ML: CXCL12/CXCR4 signaling activates
Akt-1 and MMP-9 expression in prostate cancer cells: The role of
bone microenvironment-associated CXCL12. Prostate. 66:32–48. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kawano Y, Moschetta M, Manier S, Glavey S,
Görgün GT, Roccaro AM, Anderson KC and Ghobrial IM: Targeting the
bone marrow microenvironment in multiple myeloma. Immunol Rev.
263:160–172. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ribatti D, Nico B and Vacca A: Importance
of the bone marrow microenvironment in inducing the angiogenic
response in multiple myeloma. Oncogene. 25:4257–4266. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ghobrial IM: Myeloma as a model for the
process of metastasis: Implications for therapy. Blood. 120:20–30.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Anderson KC and Carrasco RD: Pathogenesis
of myeloma. Annu Rev Pathol. 6:249–274. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Abdi J, Chen G and Chang H: Drug
resistance in multiple myeloma: Latest findings and new concepts on
molecular mechanisms. Oncotarget. 4:2186–2207. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bianchi G and Munshi NC: Pathogenesis
beyond the cancer clone(s) in multiple myeloma. Blood.
125:3049–3058. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ribatti D, Moschetta M and Vacca A:
Microenvironment and multiple myeloma spread. Thromb Res. 133
(Suppl 2):S102–S106. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lemaire M, Deleu S, De Bruyne E, Van
Valckenborgh E, Menu E and Vanderkerken K: The microenvironment and
molecular biology of the multiple myeloma tumor. Adv Cancer Res.
110:19–42. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wu Q, Zhou X, Huang D, Ji Y and Kang F:
IL-6 enhances osteocyte-mediated osteoclastogenesis by promoting
JAK2 and RANKL activity in vitro. Cell Physiol Biochem.
41:1360–1369. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hideshima T, Chauhan D, Schlossman R,
Richardson P and Anderson KC: The role of tumor necrosis factor
alpha in the pathophysiology of human multiple myeloma: Therapeutic
applications. Oncogene. 20:4519–4527. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang J, Hendrix A, Hernot S, Lemaire M, De
Bruyne E, Van Valckenborgh E, Lahoutte T, De Wever O, Vanderkerken
K and Menu E: Bone marrow stromal cell-derived exosomes as
communicators in drug resistance in multiple myeloma cells. Blood.
124:555–566. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Roccaro AM, Sacco A, Maiso P, Azab AK, Tai
YT, Reagan M, Azab F, Flores LM, Campigotto F, Weller E, et al: BM
mesenchymal stromal cell-derived exosomes facilitate multiple
myeloma progression. J Clin Invest. 123:1542–1555. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang J, De Veirman K, Faict S, Frassanito
MA, Ribatti D, Vacca A and Menu E: Multiple myeloma exosomes
establish a favourable bone marrow microenvironment with enhanced
angiogenesis and immunosuppression. J Pathol. 239:162–173. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kumar S, Gertz MA, Dispenzieri A, Lacy MQ,
Wellik LA, Fonseca R, Lust JA, Witzig TE, Kyle RA, Greipp PR and
Rajkumar SV: Prognostic value of bone marrow angiogenesis in
patients with multiple myeloma undergoing high-dose therapy. Bone
Marrow Transplant. 34:235–239. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Moschetta M, Mishima Y, Kawano Y, Manier
S, Paiva B, Palomera L, Aljawai Y, Calcinotto A, Unitt C, Sahin I,
et al: Targeting vasculogenesis to prevent progression in multiple
myeloma. Leukemia. 30:1103–1115. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Vacca A and Ribatti D: Angiogenesis and
vasculogenesis in multiple myeloma: Role of inflammatory cells.
Recent Results Cancer Res. 183:87–95. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Andreuzzi E, Colladel R, Pellicani R,
Tarticchio G, Cannizzaro R, Spessotto P, Bussolati B, Brossa A, De
Paoli P, Canzonieri V, et al: The angiostatic molecule Multimerin 2
is processed by MMP-9 to allow sprouting angiogenesis. Matrix Biol.
64:40–53. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ria R, Reale A, De Luisi A, Ferrucci A,
Moschetta M and Vacca A: Bone marrow angiogenesis and progression
in multiple myeloma. Am J Blood Res. 1:76–89. 2011.PubMed/NCBI
|
|
31
|
Vacca A, Ria R, Ribatti D, Semeraro F,
Djonov V, Di Raimondo F and Dammacco F: A paracrine loop in the
vascular endothelial growth factor pathway triggers tumor
angiogenesis and growth in multiple myeloma. Haematologica.
88:176–185. 2003.PubMed/NCBI
|
|
32
|
Menu E, Kooijman R, Van Valckenborgh E,
Asosingh K, Bakkus M, Van Camp B and Vanderkerken K: Specific roles
for the PI3K and the MEK-ERK pathway in IGF-1-stimulated
chemotaxis, VEGF secretion and proliferation of multiple myeloma
cells: Study in the 5T33MM model. Br J Cancer. 90:1076–1083. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tanaka Y, Abe M, Hiasa M, Oda A, Amou H,
Nakano A, Takeuchi K, Kitazoe K, Kido S, Inoue D, et al: Myeloma
cell-osteoclast interaction enhances angiogenesis together with
bone resorption: A role for vascular endothelial cell growth factor
and osteopontin. Clin Cancer Res. 13:816–823. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cackowski FC, Anderson JL, Patrene KD,
Choksi RJ, Shapiro SD, Windle JJ, Blair HC and Roodman GD:
Osteoclasts are important for bone angiogenesis. Blood.
115:140–149. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hideshima T, Mitsiades C, Tonon G,
Richardson PG and Anderson KC: Understanding multiple myeloma
pathogenesis in the bone marrow to identify new therapeutic
targets. Nat Rev Cancer. 7:585–598. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Roccaro AM, Hideshima T, Raje N, Kumar S,
Ishitsuka K, Yasui H, Shiraishi N, Ribatti D, Nico B, Vacca A, et
al: Bortezomib mediates antiangiogenesis in multiple myeloma via
direct and indirect effects on endothelial cells. Cancer Res.
66:184–191. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bonewald LF: The amazing osteocyte. J Bone
Miner Res. 26:229–238. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bataille R, Chappard D, Marcelli C,
Dessauw P, Sany J, Baldet P and Alexandre C: Mechanisms of bone
destruction in multiple myeloma: The importance of an unbalanced
process in determining the severity of lytic bone disease. J Clin
Oncol. 7:1909–1914. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Delgado-Calle J, Bellido T and Roodman GD:
Role of osteocytes in multiple myeloma bone disease. Curr Opin
Support Palliat Care. 8:407–413. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kristensen IB, Christensen JH, Lyng MB,
Møller MB, Pedersen L, Rasmussen LM, Ditzel HJ and Abildgaard N:
Expression of osteoblast and osteoclast regulatory genes in the
bone marrow microenvironment in multiple myeloma: Only
up-regulation of Wnt inhibitors SFRP3 and DKK1 is associated with
lytic bone disease. Leuk Lymphoma. 55:911–919. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Walker RE, Lawson MA, Buckle CH, Snowden
JA and Chantry AD: Myeloma bone disease: Pathogenesis, current
treatments and future targets. Br Med Bull. 111:117–138. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Giuliani N, Ferretti M, Bolzoni M, Storti
P, Lazzaretti M, Dalla Palma B, Bonomini S, Martella E, Agnelli L,
Neri A, et al: Increased osteocyte death in multiple myeloma
patients: Role in myeloma-induced osteoclast formation. Leukemia.
26:1391–1401. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ehrlich LA, Chung HY, Ghobrial I, Choi SJ,
Morandi F, Colla S, Rizzoli V, Roodman GD and Giuliani N: IL-3 is a
potential inhibitor of osteoblast differentiation in multiple
myeloma. Blood. 106:1407–1414. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lee JW, Chung HY, Ehrlich LA, Jelinek DF,
Callander NS, Roodman GD and Choi SJ: IL-3 expression by myeloma
cells increases both osteoclast formation and growth of myeloma
cells. Blood. 103:2308–2315. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Abe M, Hiura K, Wilde J, Shioyasono A,
Moriyama K, Hashimoto T, Kido S, Oshima T, Shibata H, Ozaki S, et
al: Osteoclasts enhance myeloma cell growth and survival via
cell-cell contact: A vicious cycle between bone destruction and
myeloma expansion. Blood. 104:2484–2491. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Croucher PI, McDonald MM and Martin TJ:
Bone metastasis: The importance of the neighbourhood. Nat Rev
Cancer. 16:373–386. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ehrlich LA and Roodman GD: The role of
immune cells and inflammatory cytokines in Paget's disease and
multiple myeloma. Immunol Rev. 208:252–266. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Emery JG, McDonnell P, Burke MB, Deen KC,
Lyn S, Silverman C, Dul E, Appelbaum ER, Eichman C, DiPrinzio R, et
al: Osteoprotegerin is a receptor for the cytotoxic ligand TRAIL. J
Biol Chem. 273:14363–14367. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Heath DJ, Vanderkerken K, Cheng X,
Gallagher O, Prideaux M, Murali R and Croucher PI: An
osteoprotegerin-like peptidomimetic inhibits osteoclastic bone
resorption and osteolytic bone disease in myeloma. Cancer Res.
67:202–208. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lawson MA, McDonald MM, Kovacic N, Hua
Khoo W, Terry RL, Down J, Kaplan W, Paton-Hough J, Fellows C,
Pettitt JA, et al: Osteoclasts control reactivation of dormant
myeloma cells by remodelling the endosteal niche. Nat Commun.
6:89832015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
McDonald MM, Fairfield H, Falank C and
Reagan MR: Adipose, bone, and myeloma: Contributions from the
microenvironment. Calcif Tissue Int. 100:433–448. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fu R, Liu H, Zhao S, Wang Y, Li L, Gao S,
Ruan E, Wang G, Wang H, Song J and Shao Z: Osteoblast inhibition by
chemokine cytokine ligand3 in myeloma-induced bone disease. Cancer
Cell Int. 14:1322014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Gavriatopoulou M, Dimopoulos MA,
Christoulas D, Migkou M, Iakovaki M, Gkotzamanidou M and Terpos E:
Dickkopf-1: A suitable target for the management of myeloma bone
disease. Expert Opin Ther Targets. 13:839–848. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Moester MJ, Papapoulos SE, Löwik CW and
van Bezooijen RL: Sclerostin: Current knowledge and future
perspectives. Calcif Tissue Int. 87:99–107. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhou F, Meng S, Song H and Claret FX:
Dickkopf-1 is a key regulator of myeloma bone disease:
Opportunities and challenges for therapeutic intervention. Blood
Rev. 27:261–267. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Oshima T, Abe M, Asano J, Hara T, Kitazoe
K, Sekimoto E, Tanaka Y, Shibata H, Hashimoto T, Ozaki S, et al:
Myeloma cells suppress bone formation by secreting a soluble Wnt
inhibitor, sFRP-2. Blood. 106:3160–3165. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Delgado-Calle J, Anderson J, Cregor MD,
Hiasa M, Chirgwin JM, Carlesso N, Yoneda T, Mohammad KS, Plotkin
LI, Roodman GD and Bellido T: Bidirectional notch signaling and
osteocyte-derived factors in the bone marrow microenvironment
promote tumor cell proliferation and bone destruction in multiple
myeloma. Cancer Res. 76:1089–1100. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Reagan MR, Liaw L, Rosen CJ and Ghobrial
IM: Dynamic interplay between bone and multiple myeloma: Emerging
roles of the osteoblast. Bone. 75:161–169. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yaccoby S, Wezeman MJ, Zangari M, Walker
R, Cottler-Fox M, Gaddy D, Ling W, Saha R, Barlogie B, Tricot G and
Epstein J: Inhibitory effects of osteoblasts and increased bone
formation on myeloma in novel culture systems and a myelomatous
mouse model. Haematologica. 91:192–199. 2006.PubMed/NCBI
|
|
60
|
Mitsiades CS, McMillin DW, Klippel S,
Hideshima T, Chauhan D, Richardson PG, Munshi NC and Anderson KC:
The role of the bone marrow microenvironment in the pathophysiology
of myeloma and its significance in the development of more
effective therapies. Hematol Oncol Clin North Am. 21:1007–1034,
vii-viii. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Shipman CM and Croucher PI:
Osteoprotegerin is a soluble decoy receptor for tumor necrosis
factor-related apoptosis-inducing ligand/Apo2 ligand and can
function as a paracrine survival factor for human myeloma cells.
Cancer Res. 63:912–916. 2003.PubMed/NCBI
|
|
62
|
Scheller J and Rose-John S: Interleukin-6
and its receptor: From bench to bedside. Med Microbiol Immunol.
195:173–183. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Suchi K, Fujiwara H, Okamura S, Okamura H,
Umehara S, Todo M, Furutani A, Yoneda M, Shiozaki A, Kubota T, et
al: Overexpression of Interleukin-6 suppresses cisplatin-induced
cytotoxicity in esophageal squamous cell carcinoma cells.
Anticancer Res. 31:67–75. 2011.PubMed/NCBI
|
|
64
|
Hong DS, Angelo LS and Kurzrock R:
Interleukin-6 and its receptor in cancer: Implications for
translational therapeutics. Cancer. 110:1911–1928. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Kawano M, Hirano T, Matsuda T, Taga T,
Horii Y, Iwato K, Asaoku H, Tang B, Tanabe O, Tanaka H, et al:
Autocrine generation and requirement of BSF-2/IL-6 for human
multiple myelomas. Nature. 332:83–85. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Rosean TR, Tompkins VS, Olivier AK,
Sompallae R, Norian LA, Morse HC III, Waldschmidt TJ and Janz S:
The tumor microenvironment is the main source of IL-6 for plasma
cell tumor development in mice. Leukemia. 29:233–237. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Matthes T, Manfroi B, Zeller A,
Dunand-Sauthier I, Bogen B and Huard B: Autocrine amplification of
immature myeloid cells by IL-6 in multiple myeloma-infiltrated bone
marrow. Leukemia. 29:1882–1890. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Rosean TR, Tompkins VS, Tricot G, Holman
CJ, Olivier AK, Zhan F and Janz S: Preclinical validation of
interleukin 6 as a therapeutic target in multiple myeloma. Immunol
Res. 59:188–202. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Qi C, Tian S, Wang J, Ma H, Qian K and
Zhang X: Co-expression of CD40/CD40L on XG1 multiple myeloma cells
promotes IL-6 autocrine function. Cancer Invest. 33:6–15. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Westendorf JJ, Ahmann GJ, Armitage RJ,
Spriggs MK, Lust JA, Greipp PR, Katzmann JA and Jelinek DF: CD40
expression in malignant plasma cells. Role in stimulation of
autocrine IL-6 secretion by a human myeloma cell line. J Immunol.
152:117–128. 1994.PubMed/NCBI
|
|
71
|
Dinarello CA: Interleukin-1 in the
pathogenesis and treatment of inflammatory diseases. Blood.
117:3720–3732. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Tu Y, Gardner A and Lichtenstein A: The
phosphatidylinositol 3-kinase/AKT kinase pathway in multiple
myeloma plasma cells: Roles in cytokine-dependent survival and
proliferative responses. Cancer Res. 60:6763–6770. 2000.PubMed/NCBI
|
|
73
|
Hideshima T, Nakamura N, Chauhan D and
Anderson KC: Biologic sequelae of interleukin-6 induced PI3-K/Akt
signaling in multiple myeloma. Oncogene. 20:5991–6000. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Hsu JH, Shi Y, Hu L, Fisher M, Franke TF
and Lichtenstein A: Role of the AKT kinase in expansion of multiple
myeloma clones: Effects on cytokine-dependent proliferative and
survival responses. Oncogene. 21:1391–1400. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Sansone P and Bromberg J: Targeting the
interleukin-6/Jak/stat pathway in human malignancies. J Clin Oncol.
30:1005–1014. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Matthes T, Manfroi B and Huard B:
Revisiting IL-6 antagonism in multiple myeloma. Crit Rev Oncol
Hematol. 105:1–4. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Monaghan KA, Khong T, Burns CJ and Spencer
A: The novel JAK inhibitor CYT387 suppresses multiple signalling
pathways, prevents proliferation and induces apoptosis in
phenotypically diverse myeloma cells. Leukemia. 25:1891–1899. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Chauhan D, Kharbanda S, Ogata A, Urashima
M, Teoh G, Robertson M, Kufe DW and Anderson KC: Interleukin-6
inhibits Fas-induced apoptosis and stress-activated protein kinase
activation in multiple myeloma cells. Blood. 89:227–234.
1997.PubMed/NCBI
|
|
79
|
Burger R: Impact of interleukin-6 in
hematological malignancies. Transfus Med Hemother. 40:336–343.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Orlowski RZ, Gercheva L, Williams C,
Sutherland H, Robak T, Masszi T, Goranova-Marinova V, Dimopoulos
MA, Cavenagh JD, Špička I, et al: A phase 2, randomized,
double-blind, placebo-controlled study of siltuximab (anti-IL-6
mAb) and bortezomib versus bortezomib alone in patients with
relapsed or refractory multiple myeloma. Am J Hematol. 90:42–49.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
San-Miguel J, Bladé J, Shpilberg O,
Grosicki S, Maloisel F, Min CK, Polo Zarzuela M, Robak T, Prasad
SV, Tee Goh Y, et al: Phase 2 randomized study of
bortezomib-melphalan-prednisone with or without siltuximab
(anti-IL-6) in multiple myeloma. Blood. 123:4136–4142. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Voorhees PM, Manges RF, Sonneveld P,
Jagannath S, Somlo G, Krishnan A, Lentzsch S, Frank RC, Zweegman S,
Wijermans PW, et al: A phase 2 multicentre study of siltuximab, an
anti-interleukin-6 monoclonal antibody, in patients with relapsed
or refractory multiple myeloma. Br J Haematol. 161:357–366. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Guo DJ, Han JS, Li YS, Liu ZS, Lu SY and
Ren HL: In vitro and in vivo antitumor effects of the recombinant
immunotoxin IL6(T23)-PE38KDEL in multiple myeloma. Oncol Lett.
4:311–318. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Younes A, Romaguera J, Fanale M,
McLaughlin P, Hagemeister F, Copeland A, Neelapu S, Kwak L, Shah J,
de Castro Faria S, et al: Phase I study of a novel oral Janus
kinase 2 inhibitor, SB1518, in patients with relapsed lymphoma:
Evidence of clinical and biologic activity in multiple lymphoma
subtypes. J Clin Oncol. 30:4161–4167. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Garcia-Bates TM, Bernstein SH and Phipps
RP: Peroxisome proliferator-activated receptor gamma overexpression
suppresses growth and induces apoptosis in human multiple myeloma
cells. Clin Cancer Res. 14:6414–6425. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Sprynski AC, Hose D, Caillot L, Réme T,
Shaughnessy JD Jr, Barlogie B, Seckinger A, Moreaux J, Hundemer M,
Jourdan M, et al: The role of IGF-1 as a major growth factor for
myeloma cell lines and the prognostic relevance of the expression
of its receptor. Blood. 113:4614–4626. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Barretina J, Caponigro G, Stransky N,
Venkatesan K, Margolin AA, Kim S, Wilson CJ, Lehár J, Kryukov GV,
Sonkin D, et al: The cancer cell line encyclopedia enables
predictive modelling of anticancer drug sensitivity. Nature.
483:603–607. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Mitsiades CS, Mitsiades NS, McMullan CJ,
Poulaki V, Shringarpure R, Akiyama M, Hideshima T, Chauhan D,
Joseph M, Libermann TA, et al: Inhibition of the insulin-like
growth factor receptor-1 tyrosine kinase activity as a therapeutic
strategy for multiple myeloma, other hematologic malignancies, and
solid tumors. Cancer Cell. 5:221–230. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Bieghs L, Lub S, Fostier K, Maes K, Van
Valckenborgh E, Menu E, Johnsen HE, Overgaard MT, Larsson O,
Axelson M, et al: The IGF-1 receptor inhibitor picropodophyllin
potentiates the anti-myeloma activity of a BH3-mimetic. Oncotarget.
5:11193–11208. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Kuhn DJ, Berkova Z, Jones RJ, Woessner R,
Bjorklund CC, Ma W, Davis RE, Lin P, Wang H, Madden TL, Wei C, et
al: Targeting the insulin-like growth factor-1 receptor to overcome
bortezomib resistance in preclinical models of multiple myeloma.
Blood. 120:3260–3270. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Bieghs L, Brohus M, Kristensen IB,
Abildgaard N, Bøgsted M, Johnsen HE, Conover CA, De Bruyne E,
Vanderkerken K, Overgaard MT and Nyegaard M: Abnormal IGF-binding
protein profile in the bone marrow of multiple myeloma patients.
PLoS One. 11:e01542562016. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Jelinek DF, Witzig TE and Arendt BK: A
role for insulin-like growth factor in the regulation of
IL-6-responsive human myeloma cell line growth. J Immunol.
159:487–496. 1997.PubMed/NCBI
|
|
93
|
Georgii-Hemming P, Wiklund HJ, Ljunggren O
and Nilsson K: Insulin-like growth factor I is a growth and
survival factor in human multiple myeloma cell lines. Blood.
88:2250–2258. 1996.PubMed/NCBI
|
|
94
|
Chapuis N, Tamburini J, Cornillet-Lefebvre
P, Gillot L, Bardet V, Willems L, Park S, Green AS, Ifrah N,
Dreyfus F, et al: Autocrine IGF-1/IGF-1R signaling is responsible
for constitutive PI3K/Akt activation in acute myeloid leukemia:
Therapeutic value of neutralizing anti-IGF-1R antibody.
Haematologica. 95:415–423. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Chiron D, Maiga S, Surget S, Descamps G,
Gomez-Bougie P, Traore S, Robillard N, Moreau P, Le Gouill S,
Bataille R, et al: Autocrine insulin-like growth factor 1 and stem
cell factor but not interleukin 6 support self-renewal of human
myeloma cells. Blood Cancer J. 3:e1202013. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Huang EW, Xue SJ, Li XY, Xu SW, Cheng JD,
Zheng JX, Shi H, Lv GL, Li ZG, Li Y, et al: EEN regulates the
proliferation and survival of multiple myeloma cells by
potentiating IGF-1 secretion. Biochem Biophys Res Commun.
447:271–277. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Vishwamitra D, George SK, Shi P, Kaseb AO
and Amin HM: Type I insulin-like growth factor receptor signaling
in hematological malignancies. Oncotarget. 8:1814–1844. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Ge NL and Rudikoff S: Insulin-like growth
factor I is a dual effector of multiple myeloma cell growth. Blood.
96:2856–2861. 2000.PubMed/NCBI
|
|
99
|
Bieghs L, Johnsen HE, Maes K, Menu E, Van
Valckenborgh E, Overgaard MT, Nyegaard M, Conover CA, Vanderkerken
K and De Bruyne E: The insulin-like growth factor system in
multiple myeloma: Diagnostic and therapeutic potential. Oncotarget.
7:48732–48752. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Vanderkerken K, Asosingh K, Braet F, Van
Riet I and Van Camp B: Insulin-like growth factor-1 acts as a
chemoattractant factor for 5T2 multiple myeloma cells. Blood.
93:235–241. 1999.PubMed/NCBI
|
|
101
|
Asosingh K, Günthert U, Bakkus MH, De
Raeve H, Goes E, Van Riet I, Van Camp B and Vanderkerken K: In vivo
induction of insulin-like growth factor-I receptor and CD44v6
confers homing and adhesion to murine multiple myeloma cells.
Cancer Res. 60:3096–3104. 2000.PubMed/NCBI
|
|
102
|
Ogata A, Chauhan D, Urashima M, Teoh G,
Treon SP and Anderson KC: Blockade of mitogen-activated protein
kinase cascade signaling in interleukin 6-independent multiple
myeloma cells. Clin Cancer Res. 3:1017–1022. 1997.PubMed/NCBI
|
|
103
|
Podar K and Anderson KC: The
pathophysiologic role of VEGF in hematologic malignancies:
Therapeutic implications. Blood. 105:1383–1395. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Andersen NF, Vogel U, Klausen TW, Gimsing
P, Gregersen H, Abildgaard N and Vangsted AJ: Vascular endothelial
growth factor (VEGF) gene polymorphisms may influence the efficacy
of thalidomide in multiple myeloma. Int J Cancer. 131:E636–E642.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Senger DR, Galli SJ, Dvorak AM, Perruzzi
CA, Harvey VS and Dvorak HF: Tumor cells secrete a vascular
permeability factor that promotes accumulation of ascites fluid.
Science. 219:983–985. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Weis SM and Cheresh DA: Tumor
angiogenesis: Molecular pathways and therapeutic targets. Nat Med.
17:1359–1370. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Asosingh K, De Raeve H, Menu E, Van Riet
I, Van Marck E, Van Camp B and Vanderkerken K: Angiogenic switch
during 5T2MM murine myeloma tumorigenesis: Role of CD45
heterogeneity. Blood. 103:3131–3137. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Bhutani M, Turkbey B, Tan E, Kemp TJ,
Pinto LA, Berg AR, Korde N, Minter AR, Weiss BM, Mena E, et al:
Bone marrow angiogenesis in myeloma and its precursor disease: A
prospective clinical trial. Leukemia. 28:413–416. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Hose D, Moreaux J, Meissner T, Seckinger
A, Goldschmidt H, Benner A, Mahtouk K, Hillengass J, Rème T, De Vos
J, et al: Induction of angiogenesis by normal and malignant plasma
cells. Blood. 114:128–143. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Taylor RM, Kashima TG, Knowles HJ and
Athanasou NA: VEGF, FLT3 ligand, PlGF and HGF can substitute for
M-CSF to induce human osteoclast formation: Implications for giant
cell tumour pathobiology. Lab Invest. 92:1398–1406. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Terpos E, Christoulas D, Gavriatopoulou M
and Dimopoulos MA: Mechanisms of bone destruction in multiple
myeloma. Eur J Cancer Care (Engl). 26:2017. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Neviani P and Fabbri M: Exosomic microRNAs
in the tumor microenvironment. Front Med (Lausanne).
2:472015.PubMed/NCBI
|
|
113
|
Wang X, Lu H, Li T, Yu L, Liu G, Peng X
and Zhao J: Krüppel-like factor 8 promotes tumorigenic mammary stem
cell induction by targeting miR-146a. Am J Cancer Res. 3:356–373.
2013.PubMed/NCBI
|
|
114
|
Corrado C, Raimondo S, Chiesi A, Ciccia F,
De Leo G and Alessandro R: Exosomes as intercellular signaling
organelles involved in health and disease: Basic science and
clinical applications. Int J Mol Sci. 14:5338–5366. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Raimondo S, Corrado C, Raimondi L, De Leo
G and Alessandro R: Role of extracellular vesicles in hematological
malignancies. Biomed Res Int. 2015:8216132015. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Zijlstra A and Di Vizio D: Size matters in
nanoscale communication. Nat Cell Biol. 20:228–230. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Amodio N, Di Martino MT, Neri A,
Tagliaferri P and Tassone P: Non-coding RNA: A novel opportunity
for the personalized treatment of multiple myeloma. Expert Opin
Biol Ther. 13 (Suppl 1):S125–S137. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Ohtsuka M, Ling H, Doki Y, Mori M and
Calin GA: MicroRNA processing and human cancer. J Clin Med.
4:1651–1667. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
De Veirman K, Wang J, Xu S, Leleu X, Himpe
E, Maes K, De Bruyne E, Van Valckenborgh E, Vanderkerken K, Menu E
and Van Riet I: Induction of miR-146a by multiple myeloma cells in
mesenchymal stromal cells stimulates their pro-tumoral activity.
Cancer Lett. 377:17–24. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Forloni M, Dogra SK, Dong Y, Conte D Jr,
Ou J, Zhu LJ, Deng A, Mahalingam M, Green MR and Wajapeyee N:
miR-146a promotes the initiation and progression of melanoma by
activating Notch signaling. Elife. 3:e014602014. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Raimondi L, De Luca A, Amodio N, Manno M,
Raccosta S, Taverna S, Bellavia D, Naselli F, Fontana S, Schillaci
O, et al: Involvement of multiple myeloma cell-derived exosomes in
osteoclast differentiation. Oncotarget. 6:13772–13789. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Raimondo S, Saieva L, Vicario E, Pucci M,
Toscani D, Manno M, Raccosta S, Giuliani N and Alessandro R:
Multiple myeloma-derived exosomes are enriched of amphiregulin
(AREG) and activate the epidermal growth factor pathway in the bone
microenvironment leading to osteoclastogenesis. J Hematol Oncol.
12:22019. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Faict S, Muller J, De Veirman K, De Bruyne
E, Maes K, Vrancken L, Heusschen R, De Raeve H, Schots R,
Vanderkerken K, et al: Exosomes play a role in multiple myeloma
bone disease and tumor development by targeting osteoclasts and
osteoblasts. Blood Cancer J. 8:1052018. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Kocemba KA, van Andel H, de Haan-Kramer A,
Mahtouk K, Versteeg R, Kersten MJ, Spaargaren M and Pals ST: The
hypoxia target adrenomedullin is aberrantly expressed in multiple
myeloma and promotes angiogenesis. Leukemia. 27:1729–1737. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Umezu T, Tadokoro H, Azuma K, Yoshizawa S,
Ohyashiki K and Ohyashiki JH: Exosomal miR-135b shed from hypoxic
multiple myeloma cells enhances angiogenesis by targeting
factor-inhibiting HIF-1. Blood. 124:3748–3757. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Lai RC, Chen TS and Lim SK: Mesenchymal
stem cell exosome: A novel stem cell-based therapy for
cardiovascular disease. Regen Med. 6:481–492. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Tomasoni S, Longaretti L, Rota C, Morigi
M, Conti S, Gotti E, Capelli C, Introna M, Remuzzi G and Benigni A:
Transfer of growth factor receptor mRNA via exosomes unravels the
regenerative effect of mesenchymal stem cells. Stem Cells Dev.
22:772–780. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Chi Y, Yin X, Sun K, Feng S, Liu J, Chen
D, Guo C and Wu Z: Redox-sensitive and hyaluronic acid
functionalized liposomes for cytoplasmic drug delivery to
osteosarcoma in animal models. J Control Release. 261:113–125.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Becker PS, Gooley TA, Green DJ, Burwick N,
Kim TY, Kojouri K, Inoue Y, Moore DJ, Nelli E, Dennie T and
Bensinger WI: A phase 2 study of bortezomib, cyclophosphamide,
pegylated liposomal doxorubicin and dexamethasone for newly
diagnosed multiple myeloma. Blood Cancer J. 6:e4222016. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Umezu T, Imanishi S, Azuma K, Kobayashi C,
Yoshizawa S, Ohyashiki K and Ohyashiki JH: Replenishing exosomes
from older bone marrow stromal cells with miR-340 inhibits
myeloma-related angiogenesis. Blood Adv. 1:812–823. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Hideshima T, Mitsiades C, Akiyama M,
Hayashi T, Chauhan D, Richardson P, Schlossman R, Podar K, Munshi
NC, Mitsiades N and Anderson KC: Molecular mechanisms mediating
antimyeloma activity of proteasome inhibitor PS-341. Blood.
101:1530–1534. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Chauhan D, Singh A, Brahmandam M, Podar K,
Hideshima T, Richardson P, Munshi N, Palladino MA and Anderson KC:
Combination of proteasome inhibitors bortezomib and NPI-0052
trigger in vivo synergistic cytotoxicity in multiple myeloma.
Blood. 111:1654–1664. 2008. View Article : Google Scholar : PubMed/NCBI
|