Introduction
Gastric cancer (GC) is the fourth most common
malignancy in humans and the second leading cause of cancer-related
death worldwide, with approximately one million new cases each year
(1). Chronic infection with
Helicobacter pylori is a major risk factor for the
initiation and progression of GC, with approximately 90% of
non-cardia GC attributed to infection with this bacterium (2,3). At
present, early diagnosis of GC patients is extremely difficult, and
the prognosis for advanced-stage patients remains extremely poor,
with a 5-year survival rate of only ~24% (2). Thus, the identification of novel
biomarkers and an improved understanding of the mechanisms
underlying GC carcinogenesis, progression, and metastasis are of
utmost importance.
MicroRNAs (miRNAs) are endogenous RNAs with a length
of ~22 nucleotides, which are key post-transcriptional regulators
of gene expression (4). In general,
these small non-coding RNAs bind to 3′-untranslated regions (UTRs)
on their target messenger RNAs (mRNAs), typically resulting in
repressed gene expression (5).
Increasing evidence indicates that miRNAs are involved in many
crucial biological processes, including cell proliferation,
invasion, migration, differentiation, and apoptosis, as well as
tumour metastasis and angiogenesis (4,5). In
addition, miRNAs are known to play significant roles in all types
of cancers, including GC, and they also function as oncogenes or
tumour suppressors by post-transcriptionally regulating
cancer-related genes (6). Recently,
microRNA-34 (miR-34) family members, including miR-34a and
miR-34b/c, have been identified as direct transcriptional targets
of the onco-suppressor p53, the expression levels of which
are significantly influenced by DNA damage and oncogenic stress
(7). In various human cancers,
including GC, the expression of miR-34 family members is
epigenetically silenced. Moreover, p53-induced miR-34
promotes cell apoptosis and senescence, induces cell cycle arrest,
represses cancer cell invasion and metastasis, and inhibits
epithelial-mesenchymal transition to inhibit carcinogenesis and
cancer development (4). Successful
cancer treatments should not only be aimed at targeting
proliferating cancer cells but also resting cancer stem cells
(CSCs) (8,9). Given its essential role in the
self-renewal and differentiation of CSCs, miR-34a may represent a
promising therapeutic target against CSCs in GC (10). Here, we review the biogenesis and
epigenetic dysregulation of miR-34 in human GC, and summarize
verified miR-34 target genes in GC. This review also discusses the
potential clinical value of miR-34 as a novel prognostic and
predictive biomarker, as well as its potential use as a therapeutic
target for the treatment of GC.
Biogenesis and structure of the miR-34
family
The miR-34 family consists of three homologous
miRNAs (miR-34a and miR-34b/c) that are encoded by two distinct
genomic loci located on chromosomes 1p36 and 11q23, respectively
(Fig. 1) (11–13).
First, miR-34 genes are transcribed as primary transcripts
(pri-miR-34) from the transcription start site in the nucleus by
RNA polymerase II/III. Next, the pri-miR-34 is polyadenylated and
capped with 7-methylguanosine. The pri-miR-34 is then cleaved into
precursor miR-34 (pre-miR-34) by DGCR8 (DiGeorge syndrome critical
region gene 8), and the pre-miR-34 is finally transported to the
cytoplasm to create a miR-34 duplex, which contains the mature
miR-34 mediating post-transcriptional gene silencing by partially
binding to the 3′-UTR of target message RNAs (14).
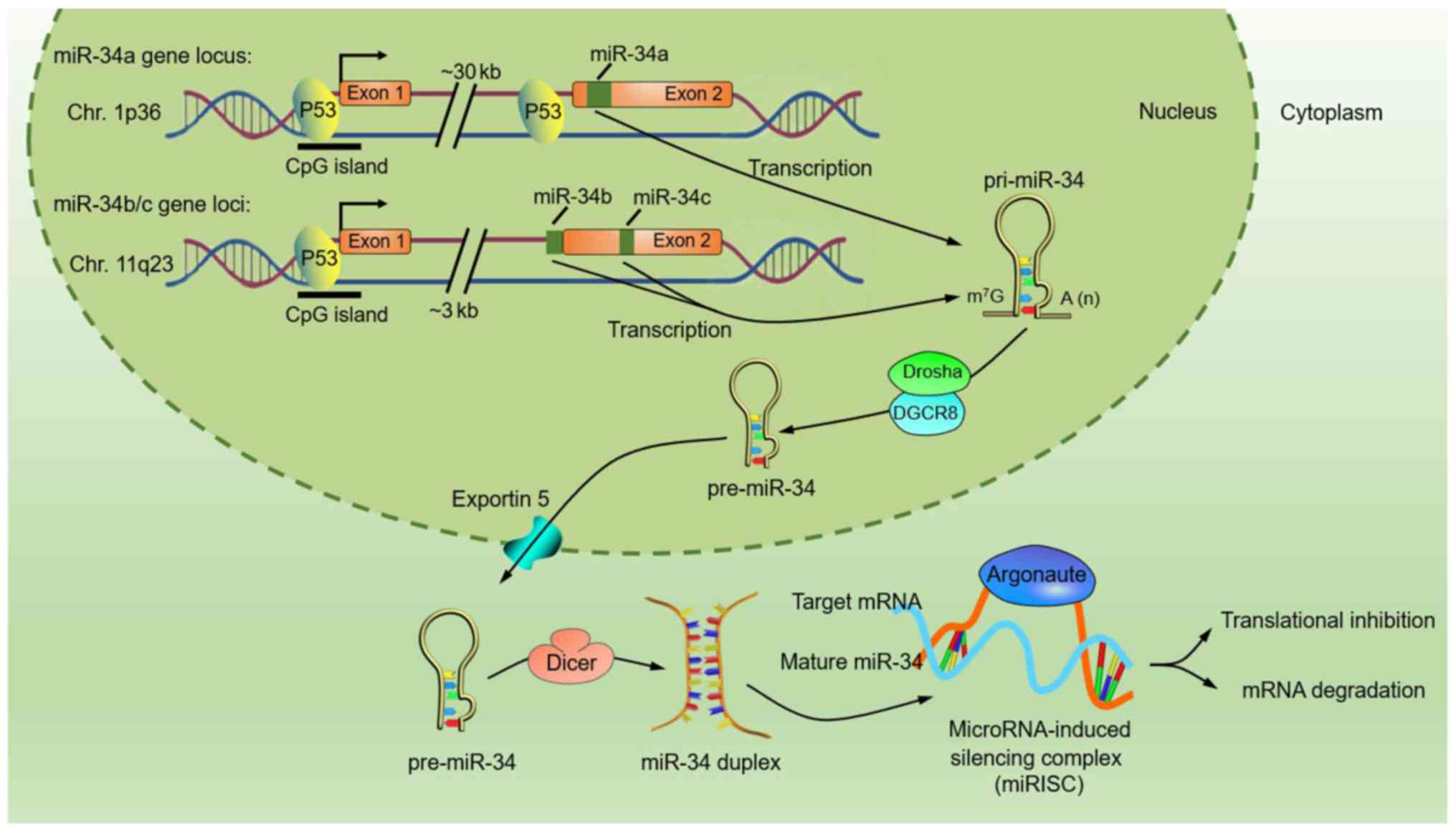 | Figure 1.Structure and biogenesis of the
miR-34 family. miR-34a is encoded by an individual transcript, and
the gene is located at the chromosome 1p36 locus (11). In contrast, miR-34b and miR-34c share
a common host gene located on chromosome 11q23 (12). The mature miR-34a sequence is located
within the second exon of its host gene, approximately 30 kb
downstream of the first exon, and the proximal region of the first
exon houses a predicted p53 binding site. miR-34b- and
miR-34c-encoding sequences are located within intron 1 and exon 2,
respectively, of the same precursor (miR-34b/c), and the proximal
regions of their transcriptional start site houses a consensus
p53-binding site (7,13). These p53-binding sites are
evolutionarily conserved and consistent with the general
characteristics of p53-regulated protein-coding targets.
First, the miR-34 gene is transcribed by RNA polymerase II/III,
thereby generating a primary miR-34 (pri-miR-34) that then
undergoes nuclear cleavage to form precursor miR-34 (pre-miR-34) by
the microprocessor complex of Drosha and DGCR8. Finally, pre-miR-34
is cleaved by RNase Dicer in the cytoplasm to create a miR-34
duplex, which contains the mature miR-34 that assembles into the
miRNA-induced silencing complex (miRISC) with Argonaute, while the
other is supposedly degraded. |
Animal studies have demonstrated that miR-34a is
ubiquitously expressed and that the highest levels are found in the
brain, whereas the highest levels of miR-34b/c expression are found
in the lungs, with lower levels in the brain and testis (7). Expression of miR-34b/c in other tissues
is scarce or undetectable (7).
miR-34c is also abundantly expressed in human sperm and testis, but
it is absent in human ovaries (15).
Thus, these findings suggest that these three miR-34 isoforms may
have tissue-specific functions. Interestingly, miR-34a and miR-34c
have identical seed sequences (nucleotides 2–7 from the 5′-UTR),
whereas the miR-34b seed sequence is similar, but not identical
(Table I) (16), indicating that miR-34a and miR-34c may
share similar mRNA targets. miR-34 isoforms use their seed
sequences to recognize the 3′-UTRs of their target mRNAs by
incomplete base pairing, inducing mRNA destabilization or
translational repression of target genes.
 | Table I.Sequences of the human miR-34 family
members. |
Table I.
Sequences of the human miR-34 family
members.
| miR-34
isoforms | Previous IDs | Sequence
(5′~3′) |
|---|
| hsa-miR-34a-5p | hsa-miR-34a | 22-UGGCAGUGUCUUAGCUGGUUGU-43 |
| hsa-miR-34a-3p | hsa-miR-34a* |
64-CAAUCAGCAAGUAUACUGCCCU-85 |
| hsa-miR-34b-5p | hsa-miR-34b;
hsa-miR-34b* | 13-UAGGCAGUGUCAUUAGCUGAUUG-35 |
| hsa-miR-34b-3p | hsa-miR-34b |
50-CAAUCACUAACUCCACUGCCAU-71 |
| hsa-miR-34c-5p | hsa-miR-34c | 13-AGGCAGUGUAGUUAGCUGAUUGC-35 |
| hsa-miR-34c-3p | – |
46-AAUCACUAACCACACGGCCAGG-67 |
miR-34 joins the p53 tumour-suppressor
network
p53, which serves as a tumour-suppressor
gene, has been implicated in many malignancies in humans (17,18). In
2007, several studies have identified miR-34a and miR-34b/c as
direct p53 target genes that regulate cell proliferation and
induce apoptosis (7,13,19,20).
Upstream regions of miR-34 family genes house one or more predicted
p53 binding sites, and p53 directly binds to those
response elements, transactivating the expression of miR-34
(7,13). Yamakuchi et al demonstrated
that restoration of miR-34a decreased the level of silent
information regulator 1 (SIRT1) that inactivates p53
by deacetylation, leading to an increase in acetylated p53
levels and expression of p21 and PUMA, which are
post-transcriptional targets of p53 that regulate the cell
cycle and apoptosis, respectively (21). Additionally, other studies have
described a positive feedback loop between p53 and miR-34a,
such as a miR-34a/MDM4/p53 loop and a
miR-34a/MYC/p53 loop, in which p53 activates
miR-34a expression and miR-34a in turn increases the activity of
p53, thereby inducing a series of cellular processes
associated with tumour suppression and apoptosis (22,23).
Notably, Christoffersen et al showed that oncogene-induced
senescence strongly upregulated miR-34a in a p53-independent
manner, which indicates that alternative cancer-related pathways
regulate miR-34a, emphasizing its significance as a tumour
suppressor (24).
Function of miR-34 and its target genes in
gastric cancer
miR-34 has been deemed to possess tumour-suppressive
properties in light of its synergistic effect with the well-known
tumour suppressor p53 (25).
In previous studies, it has been reported that restoration of
miR-34a expression induces pro-apoptotic functions in various
cancer cell lines, whereas inactivation of endogenous miR-34a
protects wild-type p53-expressing cells from DNA
damage-induced cell death (20,26,27).
Ectopic expression of miR-34 has also been shown to induce cell
cycle arrest in various cancer cell lines by suppressing cyclins D1
and E2, cyclin-dependent kinase 4 (CDK4) and CDK6
(13,28). Indeed, miR-34 has been shown to
promote the processes that inhibit carcinogenesis, such as
apoptosis and senescence, and to inhibit cancer progression via
cell cycle arrest and suppression of proliferation, invasion and
metastasis (26–29).
miR-34 is recognized as a tumour suppressor, and
generalized silencing of miR-34 results in abnormally high
expression of oncogenic targets in tumours (30,31).
Consequently, miR-34 target genes have commonly been found to be
overexpressed in GC. By microarray analysis, Zhou et al
identified IGF2BP3, a target of miR-34a, as the highest
upregulated gene in nine GC cell lines (32). Notably, some distinctions exist with
regard to their respective target genes: miR-34b targets
Notch2 and Notch4, whereas miR-34c targets
Notch1–4 (10,33).
As a vital component of the p53 suppressor
network, miR-34 has been well documented to regulate multiple
apoptosis-related genes in GC (34,35). As
shown in Table II, miR-34a promotes
GC cell apoptosis by negatively modulating survivin, Bcl-2,
and Tigf2. Among these target genes, Bcl-2, along
with its family members, is believed to be a central regulator of
apoptosis (10). miR-34 has also been
shown to be regulated by p53 during the suppression of
tumour cell proliferation. It has previously been reported that
miR-34a negatively modulates survivin in GC cells and that
survivin expression in tumours correlates with the
proliferation of cancer cells (36).
Although SIRT1 dysregulation by miR-34a in GC cells has been
demonstrated to be responsible for inhibiting proliferation and
promoting apoptosis, SIRT1 has been shown to act as a tumour
suppressor in intestinal carcinogenesis (37).
| Table II.Validated miR-34 target genes in
gastric cancer (partially listed). |
Table II.
Validated miR-34 target genes in
gastric cancer (partially listed).
| Target genes | Description | Intracellular
functions | miR-34
isoforms | Seed-matching
sequencesa | Validation
method | (Refs.) |
|---|
| Src | Tyrosine kinase
c-SRC | Proliferation and
migration | miR-34a | -CACUGCCA- | Luc., qRT-PCR,
WB | (44) |
| YY1 | Yin yang 1
transcription factor | Apoptosis | miR-34a/b/c | all, -GGCAGUG- | Luc., qRT-PCR,
WB | (33) |
| MMP7 | Matrix
metalloproteinase-7 | Metastasis | miR-34a | N.d. | qRT-PCR, WB | (45) |
|
Survivin | BIRC5 | Apoptosis | miR-34a | N.d. | qRT-PCR, WB | (46,47) |
|
PDGFRα/β | Platelet-derived
growth factor receptor | Migration, invasion
and proliferation | miR-34a | -ACUGCC- | Luc., qRT-PCR,
WB | (48) |
| FOXP1 | Forkhead box
protein 1 | B-cell
differentiation | miR-34a | N.d. | ISH and IHC | (49) |
|
MET/c-Met | A hepatocyte growth
factor receptor | Migration and
invasion | miR-34a | -CACUGCC- | Luc., qRT-PCR,
WB | (50–52) |
| Snail | Snail family zinc
finger 1 | Metastasis and
EMT | miR-34a | N.d. | IF, qRT-PCR,
WB | (52) |
| Bcl-2 | B-cell
leukaemia/lymphoma 2 | Apoptosis | miR-34 a/b/c | N.d. | Luc., qRT-PCR,
WB | (10,49) |
| Notch | Notch
homologs1/2/4 | Notch signalling
and stemness | miR-34 a/b/c | N.d. | qRT-PCR, WB | (10) |
| HMGA2 | High mobility group
AT-hook 2 | Cell cycle | miR-34a/b/c | N.d. | qRT-PCR, WB | (10) |
| Tgif2 | TGFβ-induced factor
homeobox 2 | Invasion and
metastasis | miR-34a | N.d. | qRT-PCR, WB | (53) |
| CD44 | Heparan sulphate
proteoglycan | Stemness,
proliferation and migration | miR-34a | N.d. | qRT-PCR, IHC,
WB | (54) |
|
Oct3/4 | Octamer-binding
transcription factor 3/4 | Stemness,
proliferation and migration | miR-34a | N.d. | qRT-PCR | (33,54) |
| Nanog | Nanog homeobox
transcription factor | Stemness,
proliferation and migration | miR-34a | N.d. | qRT-PCR | (10,54) |
| NOX2 | NADPH oxidase
2 | Apoptosis | miR-34a | N.d. | WB | (55) |
| IGF2BP3 | Insulin-like growth
factor-2 mRNA-binding protein 3 | Proliferation and
invasion | miR-34a | -CACUGCC- | Luc., WB | (32) |
| SIRT1 | Silent information
regulator 1 | Cell cycle,
metabolism | miR-34a | N.d. | WB | (21,37) |
The miR-34a/CD44 axis is thought to affect
the metastatic ability of GC cells by regulating the cellular
cytoskeleton-associated protein Ras homolog family member A
(RhoA), LIM domain kinase-1 (LIMK-1), and matrix
metalloproteinase (MMP)-2 (38).
Interestingly, it has been shown that simultaneous overexpression
of MET and PDGFR results in a stronger inhibitory
effect of the suppressive action of miR-34a on GC cell migration,
invasion, and proliferation than when either target is
overexpressed alone (39). Moreover,
CD44 is a significant marker of cancer stem cells (CSCs),
including gastric CSCs (40). In
recent years, the ability of tumour sphere formation of GC cells,
which generally represents the self-renewal and differentiation
potential of CSC, has been documented to be correlated with
miR-34a-mediated suppression through direct modulation of the
downstream targets CD44, Bcl-2, Notch, HMGA2, Nanog, Oct4,
SOX-2 and YY1 (16,33).
Although most information concerning the role of miRNA-34a in CSC
generation or differentiation has been obtained only from studies
in cell culture models, miR-34a may also represent a promising
therapeutic target against CSCs in GC.
Almost all the published reports have confirmed a
tumour-suppressive function of miR-34a in GC. However, there are
some reports that demonstrated an adverse function of miR-34a in
other cancers. Pu et al reported that transfection with the
miR-34a mimic made osteosarcoma G-292 cells multi-chemoresistant,
with resistance against cisplatin increasing 1.66-fold, mainly
through the downregulation of the Delta like ligand 1 (DLL1)
gene, the ligand of the Notch pathway (41). Moreover, Krause et al
demonstrated an oncogenic role for miR-34a as it promoted genomic
instability, tumourigenesis and tumour progression by regulating
the Kaposi's sarcoma-associated herpesvirus (KSHV)-encoded
chemokine receptor vGPCR (42). Additionally, in an early study, the
authors reported that the knockdown of miRNA-34a with siRNA
significantly inhibited the proliferation of renal carcinoma, HeLa
and MCF7 cells, which indicated that the overexpression of miR-34a
may be an acquired trait during carcinogenesis but warrants further
investigation (43). In a word, to
clarify the tumour-suppressive and oncogenic roles in versatile
cancers requires more mechanistic and detailed studies.
Expression of miR-34 in gastric cancer
Aberrant expression of miR-34
Since the first report of loss of expression in
glioblastoma (11), miR-34a and
miR-34b/c have been found to be aberrantly expressed in various
human cancers, including GC (56).
Indeed, miR-34 has been shown to be downregulated in most GC
tissues compared with normal mucosa (Table III). Hui et al evaluated the
expression of miR-34a in 76 GC tissues and corresponding adjacent
normal tissues by quantitative real-time polymerase chain reaction
(qPCR), and they showed that miR-34a expression was decreased in GC
tissues and that downregulation of miR-34a expression correlated
significantly with the Lauren classification of GC (57). In addition, Kim et al analysed
90 GC tissues and 34 normal samples and identified a miRNA profile
signature distinguishing GC from normal stomach epithelium,
indicating that the expression of miR-34b in GC tissue was
downregulated to 0.64 compared with normal tissue (58). Likewise, Wang et al
demonstrated that the expression of miR-34a and miR-34c, but not
miR-34b, was lower in GC than that in adjacent normal tissue
(33).
| Table III.miR-34 family expression in gastric
cancer tissues (partial list). |
Table III.
miR-34 family expression in gastric
cancer tissues (partial list).
| miRNA | Expression | Fold change (vs.
normal) | Tissue samples
(cancerous/normal) | Sample regions | Validation
methods | (Refs.) |
|---|
| miR-34b/c | Upregulated | 1.90/4.20 | 17
(11/6) | Japan | qRT-PCR | (61) |
| miR-34a | Upregulated | 1.76–3.55 | 20
(10/10) | China | qRT-PCR | (62) |
| miR-34a | Upregulated | 1.25 | 74
(37/37) | Japan | Oligo chips | (63) |
| miR-34a | Upregulated | – | 40
(20/20) | China | qRT-PCR | (64) |
| miR-34a | Upregulated | 4.03 | 27
(22/5) | Japan | Microarray
chips | (65) |
| miR-34b | Downregulated | 0.64 | 124 (90/34) | Korea | Microarray
chips | (58) |
| miR-34b | Downregulated | 0.583 | 144 (72/72) | Taiwan, China | qRT-PCR | (66) |
| miR-34a | Downregulated | <0.5 | 48
(39/9) | Hungary | qRT-PCR | (60) |
| miR-34a | Downregulated | ~0.5 | 20
(10/10) | China | qRT-PCR | (44) |
| miR-34a | Downregulated | – | 24
(12/12) | China | qRT-PCR | (45) |
| miR-34a | Downregulated | 0.299 | 6
(3/3) | China | Microarray
chips | (67) |
| miR-34a/c | Downregulated | – | 64
(32/32) | China | qRT-PCR | (33) |
| miR-34a | Downregulated | – | 137 (high 64/low
73) | China | qRT-PCR | (68) |
| miR-34a | Downregulated | 0.43–0.64 | 152 (76/76) | China | qRT-PCR | (57) |
| miR-34a | Downregulated | ~0.6 | 40 (20/20) | China | qRT-PCR | (50) |
| miR-34a | Downregulated | ~0.5 | 16 (8 /8) | China | Microarray
chips | (69) |
In human GC cell lines, expression of the miR-34
family has been extensively investigated (Table IV). For example, by qPCR, Hui et
al discovered that a panel of GC cell lines, such as NCI-N87,
AGS, MKN-45, MKN-28, BGC-823 and SGC7901, all expressed
significantly lower levels of miR-34a than the immortalized normal
human gastric epithelial cell line GES-1, indicating that decreased
miR-34a expression may be related to GC oncogenesis (57). In addition, Suzuki et al found
that miR-34b/c expression was downregulated in a larger panel of GC
cell lines (59).
| Table IV.miR-34 family expression in gastric
cancer cell lines. |
Table IV.
miR-34 family expression in gastric
cancer cell lines.
| miRNA | Expression | GC cell lines | Validation
methods | (Refs.) |
|---|
| miR-34a | Downregulated | NCI-N87, AGS,
MKN-45, MKN-28, BGC-823 and SGC-7901 | qRT-PCR | (57) |
| miR-34a | Downregulated | AGS, BGC-823,
MGC-803 and SGC-7901 | qRT-PCR | (69) |
| miR-34a | Downregulated | MGC803, SGC-7901,
HGC-27 and NCI-N87 | qRT-PCR | (46) |
| miR-34a/b/c | Downregulated | AGS, MKN45, NCI-N87
and Kato III | TaqMan RT-PCR | (10) |
| miR-34b/c | Downregulated | MKN74, SNU1,
SNU638, JRST, Kato III, AZ521, MKN28, MKN45, AGS and NCI-N87 | TaqMan RT-PCR | (59) |
Of note, miR-34 expression was also explored using
the TCGA (The Cancer Genome Atlas) database in previous studies.
For example, Wei et al investigated miR-34a expression in an
independent cohort (n=352 GC specimens) from the TCGA database
(50). The results revealed lower
levels of miR-34a expression in GC tissues and significantly lower
levels of miR-34a expression in T3 and T4 tumour stages compared
with T1, and low levels of miR-34a predicted longer survival rates
in patients with GC (50).
However, as shown in Table III, miR-34a and miR-34b/c were
upregulated in GC tissues in all three studies conducted in Japan
and some conducted in China. The lack of agreement between
different studies with respect to miR-34 expression can be
explained by a variety of factors, some of which are discussed
below. First, different studies used different reagents and
analytical approaches. Second, the inclusion of a small number of
cancer samples may affect the reliability of the miR-34 expression
level measurements. Moreover, studies reporting upregulated
expression of miR-34 generally profiled a large number of miRNAs in
human GC tissues, an approach that may lack sensitivity,
specificity and reliability. Significantly, miR-34 expression
levels in GC tissues obtained from GC patients without distant
metastases, as opposed to those obtained from patients with higher
clinical TNM stages and with poor tumour differentiation, were
demonstrated to be different. In addition, prior administration of
any type of targeted therapy, radiotherapy, chemotherapy, and/or
intervention for GC may also affect miR-34 expression levels in GC
tissues. Moreover, living conditions (rural or urban), social
status, and lifestyle behaviour, such as cigarette smoking and
alcohol consumption, are also associated with miR-34 expression
patterns in GC (60). The
inconsistency in the methods employed in the mentioned reports
requires additional analyses to draw a reliable conclusion that is
applicable for cancer diagnosis and treatment.
Mechanisms of miR-34 inactivation
In previous studies, the expression of miR-34a and
miR-34b/c was commonly lost or downregulated in many types of
cancerous tissues and cancer cell lines, including GC. However, the
correlation of downregulated miR-34 with GC and its precise role
during carcinogenesis and cancer progression are still unclear.
Therefore, identifying the underlying mechanisms involved in the
downregulated expression of miR-34 is of the utmost importance.
As miR-34 is directly transactivated by tumour
suppressor p53, missense mutations in the p53 gene
and genomic alterations of the p53 binding site may result
in the downregulation of miR-34 expression (70). In this regard, Hanazono et al
reported that a p53 gene mutation was present in
approximately 50% of GC patients and that expression of wild-type
p53 protein becomes undetectable in GC tissues as it is
replaced by overexpression of mutant forms (71). In addition, Ji et al
demonstrated the lowest level of miR-34a expression in Kato III (a
GC cell line), which is generally considered to carry a mutant form
of p53 (10). Moreover, Corney
et al demonstrated a reduction of miR-34a expression in
human epithelial ovarian cancer (EOC) tissues with both wild- or
mutant-type p53 (decreased by 93 and 100%, respectively),
and of miR-34b/c expression in 72% of EOC tissues bearing a
p53 mutation (72).
Several studies have shown that silencing of miR-34a
and miR-34b/c in GC is associated with epigenetic inactivation of
miR-34 genes (70,73). Epigenetic modifications include DNA
methylation, histone modification, and chromatin remodelling
(74). In mammalian genomes, DNA
methylation occurs almost exclusively on cytosine residues that
precede guanine (CpG), within clusters of CpG dinucleotides in
guanine-cytosine-rich regions of the genome called ‘CpG islands’.
Approximately 60% of human genes harbour CpG islands in their
promoter regions (74). The process
of CpG island methylation is catalysed by DNA methyltransferases
(DNMTs) (75). The proximal upstream
regions of miR-34a and miR-34b/c also contain CpG islands within
promoter regions, and miR-34 genes are transcribed from
transcription start sites located in the CpG island (76). Recently, several studies have
demonstrated that silenced expression of miR-34a and miR-34b/c in
GC tissues and cell lines results from aberrant CpG island
methylation in their promoter regions. Lodygin et al
reported that in a broad range of cancer tissues and cell lines,
miR-34a expression was silenced due to aberrant CpG island
methylation of its promoter (73).
Likewise, using methylation-specific PCR analysis, Suzuki et
al confirmed that the CpG islands of miR-34b and miR-34c were
completely methylated in most GC cell lines tested. In contrast, in
normal human gastric epithelial cells, the majority of CpG sites
were unmethylated (59). In their
study, these investigators also detected elevated levels of miR-34b
and miR-34c methylation in 83 of 118 (70.3%) tumour tissues from
primary GC patients by means of bisulphite-pyrosequencing. In
contrast, only limited methylation (<15.0%) was detected in
normal gastric mucosa (59). Using a
qPCR method, Tsai et al demonstrated that expression of
miR-34b was upregulated following treatment of five GC cell lines
with 5-Aza-dC (a DNA-demethylating agent) (66).
In addition to DNA methylation, histone modification
is another pivotal type of epigenetic modification that correlates
with the silencing of miR-34 expression in human GC. Liu et
al demonstrated that HOX antisense intergenic RNA (HOTAIR)
epigenetically silenced miR34a expression by binding to polycomb
repressive complex 2 (PRC2) to promote the
epithelial-to-mesenchymal transition in human GC (52). PRC2 represses miR-34a gene
transcription by trimethylating histone H3 lysine 27 (H3K27me3) in
its promoter region (52). Likewise,
Zhang et al demonstrated that SIRT7, a
NAD+-dependent acetylated lysine 18 of histone H3
(H3K18Ac) deacetylase that is overexpressed in human GC, promoted
GC growth and inhibited apoptosis of GC cells. It was found that
SIRT7 selectively bound to promoter regions of miR-34a and
deacetylated H3K18ac, resulting in decreased miR-34a expression
(77). Moreover, Lin et al
showed that depletion of HDAC1, a histone deacetylase (HDAC), in GC
cells by transfection with a specific knockdown siRNA, resulted in
a nearly 4-fold upregulated expression level of miR-34a (38). Moreover, alteration of the miR-34
biogenesis machinery may also cause aberrant miR-34 expression.
Based on the above-mentioned studies, the putative mechanisms
responsible for miR-34 gene silencing in GC, which include
p53 mutations, DNA methylation, and histone modification,
are partially described in Fig.
2.
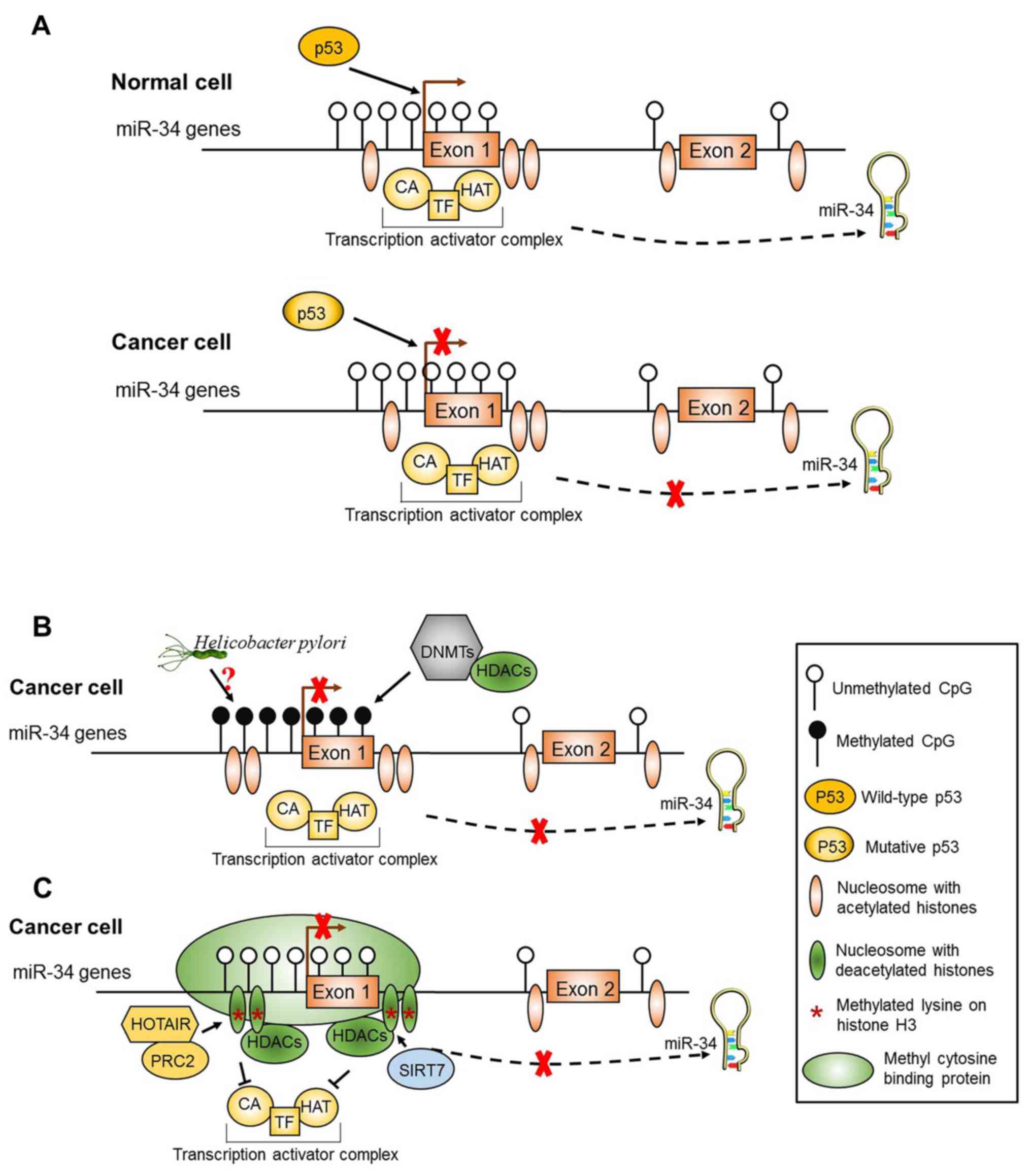 | Figure 2.Schematic diagram of the putative
regulatory mechanisms of miR-34 expression in normal cells and
gastric cancer (GC) cells. (A) The regulation of miR-34 expression
by p53. In normal cells, once activated, wild-type
p53 protein binds to p53 response elements residing
around miR-34 gene promoter regions and promotes the transcription
of miR-34. In GC cells, mutations in the p53 gene or genomic
alterations of the p53 binding site results in the
downregulation of miR-34 expression. (B) DNA methylation of the
miR-34 promoter regions. The regions surrounding the miR-34 gene
promoter regions are widely embedded with CpG islands and spaced
with nucleosomes. The DNA methyltransferase (DNMT) and histone
deacetylase (HDAC) complexes are excluded from the promoter
regions, and the miR-34 genes are normally expressed in normal
cells. But in GC cells, the promoter regions of miR-34 genes are
accessible to DNMT and HDAC complexes, causing the methylation of
CpG islands and subsequent silencing of miR-34 genes transcription.
Moreover, H. pylori infection is also associated with the
methylation of miR-34, yet the concrete mechanism is unclear. (C)
Histone modification of the miR-34 promoter regions. In GC cells,
the histones of nucleosomes and lysine on histone H3 are methylated
after methyl cytosine binding protein (MBP)- and HDAC-containing
complexes bind to the promoter regions of miR-34 genes, causing
transcriptional silencing of miR-34 genes. HOTAIR, HOX antisense
intergenic RNA; PRC2, polycomb repressive complex 2; CA,
co-activator protein; HAT, histone acetyltransferase; TF,
transcription factor. (Modified from ref. 76). |
In addition to the influence of the intrinsic
factors described above, miRNA dysregulation by Helicobacter
pylori (H. pylori) infection in gastric carcinogenesis
has also been explored. H. pylori dysregulates the
expression of various miRNAs in human gastric mucosa, which has
been linked to H. pylori-induced host inflammatory immune
responses and H. pylori-mediated gastric carcinogenesis
(78). Chang et al reported
that miR-34a was upregulated 2-fold in H. pylori-negative
versus H. pylori-positive cancers (79). Suzuki et al demonstrated that
the levels of miR-34b/c methylation were approximately three times
higher in H. pylori-positive than H. pylori-negative
mucosae (59), suggesting that
methylation of miR-34b/c is associated with H. pylori
infection. A more comprehensive understanding of the mechanisms
underlying reduced expression of miR-34 is needed to fully explain
miR-34 dysregulation by H. pylori. It is critically
important to dissect these intricate pathways and to understand how
host-pathogen interactions can disrupt these comprehensive
regulatory pathways.
Signalling pathways modulated by miR-34 in
gastric cancer
As a major downstream effector of receptor tyrosine
kinases (RTKs) and G protein-coupled receptors
(GPCRs), the phosphoinositide 3-kinase
(PI3K)/AKT pathway has been shown to play a crucial
role in carcinogenesis and tumour development (80). Recent studies have revealed that nodes
of the PI3K signal transduction system are frequently targeted by
miRNAs in a variety of human cancers, including GC. Liu et
al found that EGF-induced EGFR phosphorylation in GC
cells activated MMP-7 and consequently promoted cancer
invasion and metastasis, and this process could be inhibited by
miR-34a via modulation of PI3K/AKT signalling
cascades (45). In addition, Peng
et al found that miR-34a significantly decreased the
phosphorylation of AKT by targeting PDGFR and
MET, two upstream RTKs. They also reported that
miR-34a inhibited carcinogenesis, proliferation, invasion, and
metastasis in GC by targeting PDGFR and MET through
the PI3K/AKT pathway (48). Wang et al reported that miR-34a
mimics plus diallyl disulphide (DADS) decreased the expression of
phosphorylated PI3K (p-PI3K) and phosphorylated AKT
(p-AKT), thereby causing inactivation of the PI3K/AKT
pathway and enhancing the anti-invasion and pro-apoptotic effects
of DADS in SGC-7901 cells (81). The
PI3K/AKT pathway has been demonstrated to be
implicated in drug resistance in GC. Sun et al reported that
the upregulation of survivin expression induced by p-AKT and
hypoxia-inducible factor 1α (HIF-1α) contributed to the
cisplatin resistance of GC cells (82). Furthermore, Cao et al revealed
that restoration of miR-34a improved the sensitivity of GC cells
(SGC-7901) to cisplatin, possibly via the
PI3K/AKT/survivin signalling pathway (47).
In addition to the PI3K/AKT pathway, Tang et
al used bioinformatic analysis to discover additional pathways
that could be involved in the development and progression of GC and
be modulated by miR-34a, including the p53 signalling
pathway, the mitogen-activated protein kinase (MAPK),
Wnt/β-catenin, Smad, and Notch signalling pathways
(83). Moreover, Liu et al
reported that silencing of miR-34a induced by HOTAIR promoted
EMT in GC, possibly through activation of the
HGF/c-MET/Snail pathway (52). Kim et al demonstrated that in
Epstein-Barr virus (EBV)-infected GC cells, transcriptional
upregulation of NADPH oxidase 2 (NOX2) and the subsequent
accumulation of reactive oxygen species (ROS) was induced by EBV
nuclear antigen 1 (EBNA1), through downregulation of miR34a
expression (55). Together, these
results suggest that EBNA1 promotes cell viability by regulating
EBNA1/miR34a/NOX2/ROS signalling in GC cells
(55).
Clinical implications of miR-34
Promising biomarkers in GC
The correlation between miR-34a expression and the
prediction/prognosis for GC patients has been well established
(50,68,69). A
higher recurrence rate and an inferior survival outcome have been
observed in GC patients with low expression levels of miR-34a
compared with patients who express higher miR-34a levels (50,68). In
addition to its value as a prognostic biomarker, the methylation
level of miR-34 genes has also been explored in relation to the
predictive merits for GC. Suzuki et al observed that the
methylation level of miR-34b/c was higher in the gastric mucosa of
patients with multiple GC than in the gastric mucosa of H.
pylori-positive healthy controls or patients with single GC
(59). In addition, the methylation
level of miR-34b/c in noncancerous gastric body mucosa was an
independent predictive risk factor for metachronous GC in patients
treated with endoscopic submucosal dissection (ESD) (84). Therefore, frequent follow-up endoscopy
is needed in GC patients presenting with high miR-34b/c methylation
levels in the gastric body after ESD (84).
miR-34 polymorphism and GC risk
The most frequent genetic mutations in the human
genome are single nucleotide polymorphisms (SNPs), which are
variations of a single nucleotide. Since the regulatory function of
miRNA is dependent on base complementation, a SNP occurring either
in the miRNA seed regions or in targeted mRNA sequences may have
significant effects (85–87). By in silico searching and
database mining, Xu et al found a potentially functional
common SNP rs4938723 (T>C) in the promoter region of
pri-miR-34b/c (423-bp upstream from the transcription start site),
which was located in a typical CpG island (88). Subsequently, Yang et al
revealed that miR-34b/c rs4938723 CT/CC genotypes may be associated
with a decreased risk of GC in a Chinese population and that the C
allele may be a protective factor in GC (89). Pan et al demonstrated a lower
occurrence rate of GC among patients with CT and CT/CC genotypes in
miR-34b/c rs4938723 compared with patients with a TT genotype
(90). These results were consistent
with a meta-analysis study, which reported a reverse association
between miR-34b/c rs4938723 polymorphism and susceptibility to GC
(91).
Therapeutic applications in
cancers
As a promising tumour suppressor, miR-34a has been
considered as a therapeutic candidate in cancer. It is tempting to
hypothesize that restoration of the expression of miR-34 may
represent a novel and feasible treatment for GC patients, or that
it could become a complementary therapy to conventional surgical
resection and chemoradiation (46,50,69).
Consistent with its role as a negative regulator of CSCs in
prostatic and pancreatic cancer, restoration of miR-34a has been
documented to inhibit the self-renewal potential of gastric CSCs,
which may offer a novel target against GC (10). The sensitivity to cisplatin could be
restored to some extent by elevating cellular miR-34a, indicating
that improved treatment could be obtained in GC patients by
administering cisplatin together with miR-34a (47,51).
Moreover, ectopic expression of miR-34a can also elevate the
sensitivity of GC cells with low levels of miR-34a expression to
doxorubicin, gemcitabine, and docetaxel (10).
The development of immunotherapy targeting the
PD1 (programmed cell death protein 1) or PDL1
(programmed death ligand 1) checkpoint has led to considerable
progress in the treatment of many cancers. Recently, Wang et
al showed that the downregulation of miR-34a in acute myeloid
leukaemia was inversely correlated with high protein expression of
PDL1 and miR-34a was capable of targeting a predicted site
in the PDL1 3′-UTR, resulting in transcriptional repression
(92). Cortez et al also
demonstrated that p53 specifically modulated the tumour
immune response in non-small cell lung cancer by repressing
PDL1 via miR-34 (93). These
results suggest that miR-34a could function as a potential
immunotherapeutic target, and delivery of miR-34a combined with
standard therapies may represent a new approach in cancer
immunotherapy.
Although miR-34 overexpression has great
therapeutic potential according to the recent research findings, a
major challenge that limits the transition from basic in
vitro studies to in vivo clinical applications is the
deficiency of a miRNA delivery system that can elevate miR-34 in
target sites to therapeutically optimal levels. It is significant
that Jang et al has demonstrated that nano-vesicles
containing poly-l-lysine-graft-imidazole (PLI)/miR-34a complexes
(NVs/miR-34a) not only efficiently delivered miR-34a into GC cells
in vitro but also successfully delivered miR-34a
systemically with an accumulation of miR-34a by tumours in
vivo (54). Significant
antitumour efficacy was observed in a gastric tumour xenograft nude
mouse model that was injected with NVs/miR-34a via the tail vein of
the mouse (54). In addition, Di
Martino et al described a lentiviral miR-34a mimic delivery
system that elevated miR-34a expression significantly in multiple
myeloma cells, and the multiple myeloma xenograft formation and
size were suppressed by lentiviral miR-34a in the mouse models
(94). Lentiviral delivery system was
also used to deliver miR-34a to prostate cancer progenitor cells,
and the results showed that the lentivirus-delivered miR-34a
inhibited cancer cell metastasis (95). The stable nucleic acid lipid particle
(SNALP) is a synthesized vector that provides another successful
strategy for miR-34a mimic delivery, exhibiting high stability in
serum but low toxicity. SNALP-encapsulated miR-34a mimics were
successfully delivered in vitro in multiple myeloma (MM)
cells with a significant change of gene expression with relevant
effects on multiple signal transduction pathways (96). The same system was then experimented
in vivo and the exciting results showed a significant
inhibition of MM xenograft growth and a prolonged mice survival
(96).
The application of miR-34a is not limited to
laboratory, clinical trials have also confirmed the therapeutic
prospect of miR-34a in various cancer types. Daige et al
ever reported that the systemic delivery of MRX34, a
liposome-formulated miR-34 mimic, increased the levels of miR34a
expression by approximately 1,000-fold in vitro and caused
the inhibition of liver tumour xenograft growth (97). Recently, MRX34 was included in
clinical trials as the first miRNA-associated therapeutic agent.
And the first clinical trial (NCT01829971) of MRX34 implemented in
2013 that enrolled 155 patients with 7 cancer types, including
solid tumours and blood malignancies. In this trial a good
therapeutic effect was observed but only with some immunoreaction,
which provided bright prospect for MRX34 to clinical oncotherapy.
In 2017, Beg et al recruited 47 patients with various solid
tumours to assess the maximum tolerated dose (MTD), safety,
pharmacokinetics, and clinical activity of MRX34. The authors found
that MRX34 with dexamethasone premedication was associated with
acceptable safety. And the MTD was 93 mg/m2 for
hepatocellular carcinoma (HCC) patients and 110 mg/m2
for non-HCC patients (98).
Conclusion
The three miR-34 isoforms (i.e. miR-34a and
miR-34b/c) are encoded by two different transcriptional units.
Furthermore, the seed sequence of miR-34a and miR-34c is identical,
indicating that they hold similar mRNA targets, while miR-34b is a
little different, as described in Fig.
1 and Table I. miR-34a and
miR-34b/c are direct target genes of p53 which has been
shown to be downregulated or lost in various types of malignancies,
including gastric cancer (GC) (Table
III and Table IV). Moreover, it
has been demonstrated to play a crucial role in repressing
tumorigenesis and progression. This tumour-suppressive process is
credited to the regulation of multiple biological processes,
including cell cycle arrest, metastasis, induction of senescence
and apoptosis by targeting more than 30 different oncogenes (e.g.
SIRT1, YY1, MMP7, PDGFR-α/β, Met, Bcl-2, Snail, Tgif2, HMGA2,
CD44, NOX2 and IGF2BP3), as well as tumour immune
evasion by targeting PD-L1, death-associated protein kinase
2 (DAPK2), specificity protein 1 (Sp1) or androgen
receptor (AR) (99,100). Moreover, miR-34a and miR-34b/c have
been demonstrated to target the expression of the same oncogenes
such as YY1, Bcl-2, Notch and HMGA2 in gastric cancer
to regulate cancer metastasis, stemness and apoptosis (Table II). Several key mediators or
signalling pathway nodes (e.g. PI3K/AKT, MAPK, Wnt/β-catenin,
Smad and Notch pathways) that are abnormally activated
during the process of carcinogenesis have been shown to be
effectively modulated by miR-34 in GC. A large number of studies
concerning the use of miR-34a mimics for cancer therapy have been
carried out in vitro and in vivo, and its tumour
suppressive role has been confirmed. Many vectors such as
nanovesicles, lentiviruses, SNALPs, polymeric nanogels and
liposomes have been designed to achieve the effective systemic
delivery of miR-34a mimics. The application of miR-34a in clinical
oncotherapy has also been mentioned. MRX34, a liposome-formulated
miR-34 mimic, serving as the first miRNA-associated drug for
oncotherapy tested in clinical trials, exhibited good antitumor
activity in patients with refractory solid tumours and blood
malignancies. Moreover, the application of MRX34 for treating
numerous types of cancers is feasible, contributing to its broad
distribution in various tissues (97,98). The
combinations of miR-34a with other tumour-suppressive miRNA agents
in ‘cocktails’, such as in combination with miR-15a, may benefit
patients by means of the synergistic effects. Moreover, when
administered together, miR-34a can elevate the sensitivity of
cancer cells to chemotherapeutic agents, including cisplatin,
gemcitabine and docetaxel. Additionally, miR-34a and miR-34b/c have
also been demonstrated to play a significant role in tumour
suppression in vitro. However, few studies of miR-34b/c
in vivo have been reported, thus their application for
oncotherapy in the clinic warrants exploration.
In conclusion, miR-34 has been identified as a
novel diagnostic, prognostic, and predictive biomarker for GC, and
restoration of miR-34 levels may produce a synergistic effect with
other drugs to achieve a better clinical outcome in GC
patients.
Acknowledgements
The authors wish to thank the reviewers for the
valuable suggestions that significantly improved the quality of
this manuscript.
Funding
The present study was funded by grants from the
National Science Foundation of China (nos. 81160307 and
81560395).
Availability of data and materials
Not applicable.
Authors' contributions
SX and MH wrote the manuscript. SX constructed the
tables and diagrams. SX, MH, CL, HC and XZ checked and revised the
manuscript. SX, MH, HC and XZ were responsible for the submission
of this manuscript and final approval of the version to be
published. All the authors were also involved in the conception and
design of the study, as well as the acquisition, analysis and
interpretation of data for the work. All authors agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors have declared that there are no
conflicts of interest.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Plummer M, Franceschi S, Vignat J, Forman
D and Martel C: Global burden of gastric cancer attributable to
Helicobacter pylori. Int J Cancer. 136:487–490. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tsugane S and Sasazuki S: Diet and the
risk of gastric cancer: Review of epidemiological evidence. Gastric
Cancer. 10:75–83. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pasquinelli AE: MicroRNAs and their
targets: Recognition, regulation and an emerging reciprocal
relationship. Nat Rev Genet. 13:271–282. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hayes J, Peruzzi PP and Lawler S:
MicroRNAs in cancer: Biomarkers, functions and therapy. Trends Mol
Med. 20:460–469. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bommer GT, Gerin I, Feng Y, Kaczorowski
AJ, Kuick R, Love RE, Zhai Y, Giordano TJ, Qin ZS, Moore BB, et al:
p53-mediated activation of miRNA34 candidate tumor-suppressor
genes. Curr Biol. 17:1298–1307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dingli D and Michor F: Successful therapy
must eradicate cancer stem cells. Stem Cells. 24:2603–2610. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Dontu G, Al-Hajj M, Abdallah WM, Clarke MF
and Wicha MS: Stem cells in normal breast development and breast
cancer. Cell Prolif. 36 (Suppl 1):S59–S72. 2003. View Article : Google Scholar
|
|
10
|
Ji Q, Hao X, Meng Y, Zhang M, Desano J,
Fan D and Xu L: Restoration of tumor suppressor miR-34 inhibits
human p53-mutant gastric cancer tumorspheres. BMC Cancer.
8:2662008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Versteeg R, Caron H, Cheng NC, van der
Drift P, Slater R, Westerveld A, Voûte PA, Delattre O, Laureys G,
Van Roy N, et al: 1p36: every subband a suppressor? Eur J Cancer
31A. 538–541. 1995. View Article : Google Scholar
|
|
12
|
Rodriguez A, Griffiths-Jones S, Ashurst JL
and Bradley A: Identification of mammalian microRNA host genes and
transcription units. Genome Res. 14:1902–1910. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
He L, He X, Lim LP, de Stanchina E, Xuan
Z, Liang Y, Xue W, Zender L, Magnus J, Ridzon D, et al: A microRNA
component of the p53 tumour suppressor network. Nature.
447:1130–1134. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ha M and Kim VN: Regulation of microRNA
biogenesis. Nat Rev Mol Cell Biol. 15:509–524. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Krawetz SA, Kruger A, Lalancette C, Tagett
R, Anton E, Draghici S and Diamond MP: A survey of small RNAs in
human sperm. Hum Reprod. 26:3401–3412. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jafari N and Abediankenari S: MicroRNA-34
dysregulation in gastric cancer and gastric cancer stem cell.
Tumour Biol. 39:10104283177016522017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vogelstein B, Lane D and Levine AJ:
Surfing the p53 network. Nature. 408:307–310. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Riley T, Sontag E, Chen P and Levine A:
Transcriptional control of human p53-regulated genes. Nat Rev Mol
Cell Biol. 9:402–412. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Tarasov V, Jung P, Verdoodt B, Lodygin D,
Epanchintsev A, Menssen A, Meister G and Hermeking H: Differential
regulation of microRNAs by p53 revealed by massively parallel
sequencing: miR-34a is a p53 target that induces apoptosis and
G1-arrest. Cell Cycle. 6:1586–1593. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chang TC, Wentzel EA, Kent OA,
Ramachandran K, Mullendore M, Lee KH, Feldmann G, Yamakuchi M,
Ferlito M, Lowenstein CJ, et al: Transactivation of miR-34a by p53
broadly influences gene expression and promotes apoptosis. Mol
Cell. 26:745–752. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yamakuchi M, Ferlito M and Lowenstein CJ:
miR-34a repression of SIRT1 regulates apoptosis. Proc Natl Acad Sci
USA. 105:13421–13426. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Menssen A, Hydbring P, Kapelle K,
Vervoorts J, Diebold J, Lüscher B, Larsson LG and Hermeking H: The
c-MYC oncoprotein, the NAMPT enzyme, the SIRT1-inhibitor DBC1, and
the SIRT1 deacetylase form a positive feedback loop. Proc Natl Acad
Sci USA. 109:E187–E196. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Mandke P, Wyatt N, Fraser J, Bates B,
Berberich SJ and Markey MP: MicroRNA-34a modulates MDM4 expression
via a target site in the open reading frame. PLoS One.
7:e420342012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Christoffersen NR, Shalgi R, Frankel LB,
Leucci E, Lees M, Klausen M, Pilpel Y, Nielsen FC, Oren M and Lund
AH: p53-independent upregulation of miR-34a during oncogene-induced
senescence represses MYC. Cell Death Differ. 17:236–245. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Welch C, Chen Y and Stallings RL:
MicroRNA-34a functions as a potential tumor suppressor by inducing
apoptosis in neuroblastoma cells. Oncogene. 26:5017–5022. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Raver-Shapira N, Marciano E, Meiri E,
Spector Y, Rosenfeld N, Moskovits N, Bentwich Z and Oren M:
Transcriptional activation of miR-34a contributes to p53-mediated
apoptosis. Mol Cell. 26:731–743. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Rokavec M, Li H, Jiang L and Hermeking H:
The p53/miR-34 axis in development and disease. J Mol Cell Biol.
6:214–230. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sun F, Fu H, Liu Q, Tie Y, Zhu J, Xing R,
Sun Z and Zheng X: Downregulation of CCND1 and CDK6 by miR-34a
induces cell cycle arrest. FEBS Lett. 582:1564–1568. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mudduluru G, Ceppi P, Kumarswamy R,
Scagliotti GV, Papotti M and Allgayer H: Regulation of Axl receptor
tyrosine kinase expression by miR-34a and miR-199a/b in solid
cancer. Oncogene. 30:2888–2899. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang B, Li D, Kovalchuk I, Apel IJ,
Chinnaiyan AM, Wóycicki RK, Cantor CR and Kovalchuk O: miR-34a
directly targets tRNAiMet precursors and affects cellular
proliferation, cell cycle, and apoptosis. Proc Natl Acad Sci USA.
115:7392–7397. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Grammatikakis I, Gorospe M and Abdelmohsen
K: Modulation of cancer traits by tumor suppressor microRNAs. Int J
Mol Sci. 14:1822–1842. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou Y, Huang T, Siu HL, Wong CC, Dong Y,
Wu F, Zhang B, Wu WK, Cheng AS, Yu J, et al: IGF2BP3 functions as a
potential oncogene and is a crucial target of miR-34a in gastric
carcinogenesis. Mol Cancer. 16:772017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang AM, Huang TT, Hsu KW, Huang KH, Fang
WL, Yang MH, Lo SS, Chi CW, Lin JJ and Yeh TS: Yin Yang 1 is a
target of microRNA-34 family and contributes to gastric
carcinogenesis. Oncotarget. 5:5002–5016. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mert U, Ozgür E, Tiryakioglu D, Dalay N
and Gezer U: Induction of p53-inducible microRNA miR-34 by gamma
radiation and bleomycin are different. Front Genet. 3:2202012.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Polyak K, Xia Y, Zweier JL, Kinzler KW and
Vogelstein B: A model for p53-induced apoptosis. Nature.
389:300–305. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shen Z, Zhan G, Ye D, Ren Y, Cheng L, Wu Z
and Guo J: MicroRNA-34a affects the occurrence of laryngeal
squamous cell carcinoma by targeting the antiapoptotic gene
survivin. Med Oncol. 29:2473–2480. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Deng X, Zheng H, Li D, Xue Y, Wang Q, Yan
S, Zhu Y and Deng M: MicroRNA-34a regulates proliferation and
apoptosis of gastric cancer cells by targeting silent information
regulator 1. Exp Ther Med. 15:3705–3714. 2018.PubMed/NCBI
|
|
38
|
Lin L, Jiang H, Huang M, Hou X, Sun X,
Jiang X, Dong X, Sun X, Zhou B and Qiao H: Depletion of histone
deacetylase 1 inhibits metastatic abilities of gastric cancer cells
by regulating the miR-34a/CD44 pathway. Oncol Rep. 34:663–672.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ha SY, Lee J, Kang SY, Do IG, Ahn S, Park
JO, Kang WK, Choi MG, Sohn TS, Bae JM, et al: MET overexpression
assessed by new interpretation method predicts gene amplification
and poor survival in advanced gastric carcinomas. Mod Pathol.
26:1632–1641. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Takaishi S, Okumura T, Tu S, Wang SS,
Shibata W, Vigneshwaran R, Gordon SA, Shimada Y and Wang TC:
Identification of gastric cancer stem cells using the cell surface
marker CD44. Stem Cells. 27:1006–1020. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pu Y, Zhao F, Wang H and Cai S: miR-34a-5p
promotes multi-chemoresistance of osteosarcoma through
down-regulation of the DLL1 gene. Sci Rep. 7:442182017. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Krause CJ, Popp O, Thirunarayanan N,
Dittmar G, Lipp M and Müller G: MicroRNA-34a promotes genomic
instability by a broad suppression of genome maintenance mechanisms
downstream of the oncogene KSHV-vGPCR. Oncotarget. 7:10414–10432.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Dutta KK, Zhong Y, Liu YT, Yamada T,
Akatsuka S, Hu Q, Yoshihara M, Ohara H, Takehashi M, Shinohara T,
et al: Association of microRNA-34a overexpression with
proliferation is cell type-dependent. Cancer Sci. 98:1845–1852.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hao Q, Lu X, Liu N, Xue X, Li M, Zhang C,
Qin X, Li W, Shu Z, Song B, et al: Posttranscriptional deregulation
of Src due to aberrant miR34a and miR203 contributes to gastric
cancer development. BMB Rep. 46:316–321. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu G, Jiang C, Li D, Wang R and Wang W:
miRNA-34a inhibits EGFR-signaling-dependent MMP7 activation in
gastric cancer. Tumour Biol. 35:9801–9806. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cao W, Fan R, Wang L, Cheng S, Li H, Jiang
J, Geng M, Jin Y and Wu Y: Expression and regulatory function of
miRNA-34a in targeting survivin in gastric cancer cells. Tumor
Biol. 34:963–971. 2013. View Article : Google Scholar
|
|
47
|
Cao W, Yang W, Fan R, Li H, Jiang J, Geng
M, Jin Y and Wu Y: miR-34a regulates cisplatin-induce gastric
cancer cell death by modulating PI3K/AKT/survivin pathway. Tumour
Biol. 35:1287–1295. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Peng Y, Guo JJ, Liu YM and Wu XL:
MicroRNA-34A inhibits the growth, invasion and metastasis of
gastric cancer by targeting PDGFR and MET expression. Biosci Rep.
34(pii): e001122014.PubMed/NCBI
|
|
49
|
He M, Gao L, Zhang S, Tao L, Wang J, Yang
J and Zhu M: Prognostic significance of miR-34a and its target
proteins of FOXP1, p53, and BCL2 in gastric MALT lymphoma and
DLBCL. Gastric Cancer. 17:431–441. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wei B, Huang QY, Huang SR, Mai W and Zhong
XG: MicroRNA-34a attenuates the proliferation, invasion and
metastasis of gastric cancer cells via downregulation of MET. Mol
Med Rep. 12:5255–5261. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhang Z, Kong Y, Yang W, Ma F, Zhang Y, Ji
S, Ma EM, Liu H, Chen Y and Hua Y: Upregulation of microRNA-34a
enhances the DDP sensitivity of gastric cancer cells by modulating
proliferation and apoptosis via targeting MET. Oncol Rep.
36:2391–2397. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Liu YW, Sun M, Xia R, Zhang EB, Liu XH,
Zhang ZH, Xu TP, De W, Liu BR and Wang ZX: LincHOTAIR
epigenetically silences miR34a by binding to PRC2 to promote the
epithelial-to-mesenchymal transition in human gastric cancer. Cell
Death. 6:e18022015. View Article : Google Scholar
|
|
53
|
Hu Y, Pu Q, Cui B and Lin J: MicroRNA-34a
inhibits tumor invasion and metastasis in gastric cancer by
targeting Tgif2. Int J Clin Exp Pathol. 8:8921–8928.
2015.PubMed/NCBI
|
|
54
|
Jang E, Kim E, Son HY, Lim EK, Lee H, Choi
Y, Park K, Han S, Suh JS, Huh YM and Haam S: Nanovesicle-mediated
systemic delivery of microRNA-34a for CD44 overexpressing gastric
cancer stem cell therapy. Biomaterials. 105:12–24. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Kim S-M, Hur DY, Hong SW and Kim JH:
EBV-encoded EBNA1 regulates cell viability by modulating
miR34a-NOX2-ROS signaling in gastric cancer cells. Biochem Biophys
Res Commun. 494:550–555. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Bader AG: miR-34-a microRNA replacement
therapy is headed to the clinic. Front Genet. 3:1202012. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Hui WT, Ma XB, Zan Y, Wang XJ and Dong L:
Prognostic significance of miR-34a expression in patients with
gastric cancer after radical gastrectomy. Chin Med J (Engl).
128:2632–2637. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kim CH, Kim HK, Rettig RL, Kim J, Lee ET,
Aprelikova O, Choi IJ, Munroe DJ and Green JE: miRNA signature
associated with outcome of gastric cancer patients following
chemotherapy. BMC Med Genomics. 4:792011. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Suzuki H, Yamamoto E, Nojima M, Kai M,
Yamano HO, Yoshikawa K, Kimura T, Kudo T, Harada E, Sugai T, et al:
Methylation-associated silencing of microRNA-34b/c in gastric
cancer and its involvement in an epigenetic field defect.
Carcinogenesis. 31:2066–2073. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Stánitz E, Juhász K, Tóth C, Gombos K,
Natali PG and Ember I: Evaluation of MicroRNA expression pattern of
gastric adenocarcinoma associated with socioeconomic, environmental
and lifestyle factors in northwestern Hungary. Anticancer Res.
33:3195–3200. 2013.PubMed/NCBI
|
|
61
|
Katada T, Ishiguro H, Kuwabara Y, Kimura
M, Mitui A, Mori Y, Ogawa R, Harata K and Fujii Y: microRNA
expression profile in undifferentiated gastric cancer. Int J Oncol.
34:537–542. 2009.PubMed/NCBI
|
|
62
|
Yao Y, Suo AL, Li ZF, Liu LY, Tian T, Ni
L, Zhang WG, Nan KJ, Song TS and Huang C: MicroRNA profiling of
human gastric cancer. Mol Med Rep. 2:963–970. 2009.PubMed/NCBI
|
|
63
|
Osawa S, Shimada Y, Sekine S, Okumura T,
Nagata T, Fukuoka J and Tsukada K: MicroRNA profiling of gastric
cancer patients from formalin-fixed paraffin-embedded samples.
Oncol Lett. 2:613–619. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Su Y, Ni Z, Wang G, Cui J, Wei C, Wang J,
Yang Q, Xu Y and Li F: Aberrant expression of microRNAs in gastric
cancer and biological significance of miR-574-3p. Int
Immunopharmacol. 13:468–475. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Tsukamoto Y, Nakada C, Noguchi T, Tanigawa
M, Nguyen LT, Uchida T, Hijiya N, Matsuura K, Fujioka T, Seto M and
Moriyama M: MicroRNA-375 is downregulated in gastric carcinomas and
regulates cell survival by targeting PDK1 and 14-3-3zeta. Cancer
Res. 70:2339–2349. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Tsai KW, Wu CW, Hu LY, Li SC, Liao YL, Lai
CH, Kao HW, Fang WL, Huang KH, Chan WC and Lin WC: Epigenetic
regulation of miR-34b and miR-129 expression in gastric cancer. Int
J Cancer. 129:2600–2610. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Liu D, Hu X, Zhou H, Shi G and Wu J:
Identification of aberrantly expressed miRNAs in gastric cancer.
Gastroenterol Res Pract. 2014:4738172014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhang H, Li S, Yang J, Liu S, Gong X and
Yu X: The prognostic value of miR-34a expression in completely
resected gastric cancer: Tumor recurrence and overall survival. Int
J Clin Exp Med. 8:2635–2641. 2015.PubMed/NCBI
|
|
69
|
Yang B, Huang J, Liu H, Guo W and Li G:
miR-335 directly, while miR-34a indirectly modulate survivin
expression and regulate growth, apoptosis, and invasion of gastric
cancer cells. Tumour Biol. 37:1771–1779. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wang L, Yu J, Xu J, Zheng C, Li X and Du
J: The analysis of microRNA-34 family expression in human cancer
studies comparing cancer tissues with corresponding pericarcinous
tissues. Gene. 554:1–8. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Hanazono K, Natsugoe S, Stein HJ, Aikou T,
Hoefler H and Siewert JR: Distribution of p53 mutations in
esophageal and gastric carcinomas and the relationship with p53
expression. Oncol Rep. 15:821–824. 2006.PubMed/NCBI
|
|
72
|
Corney DC, Hwang CI, Matoso A, Vogt M,
Flesken-Nikitin A, Godwin AK, Kamat AA, Sood AK, Ellenson LH and
Hermeking H: Nikitin AYFrequent downregulation of miR-34 family in
human ovarian cancers. Clin Cancer Res. 16:1119–1128. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Lodygin D, Tarasov V, Epanchintsev A,
Berking C, Knyazeva T, Körner H, Knyazev P, Diebold J and Hermeking
H: Inactivation of miR-34a by aberrant CpG methylation in multiple
types of cancer. Cell Cycle. 7:2591–2600. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Gal-Yam EN, Saito Y, Egger G and Jones PA:
Cancer epigenetics: Modifications, screening, and therapy. Annu Rev
Med. 59:267–280. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Jones PA and Baylin SB: The fundamental
role of epigenetic events in cancer. Nat Rev Genet. 3:415–428.
2002. View
Article : Google Scholar : PubMed/NCBI
|
|
76
|
Hermeking H: The miR-34 family in cancer
and apoptosis. Cell Death Differ. 17:193–199. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Zhang S, Chen P, Huang Z, Hu X, Chen M, Hu
S, Hu Y and Cai T: Sirt7 promotes gastric cancer growth and
inhibits apoptosis by epigenetically inhibiting miR-34a. Sci Rep.
5:97872015. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Noto JM and Peek RM: The role of microRNAs
in Helicobacter pylori pathogenesis and gastric
carcinogenesis. Front Cell Infect Microbiol. 1:212012. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Chang H, Kim N, Park JH, Nam RH, Choi YJ,
Lee HS, Yoon H, Shin CM, Park YS, Kim JM and Lee DH: Different
microRNA expression levels in gastric cancer depending on
Helicobacter pylori infection. Gut Liver. 9:188–196. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Liu P, Cheng H, Roberts TM and Zhao JJ:
Targeting the phosphoinositide 3-kinase pathway in cancer. Nat Rev
Drug Discov. 8:627–644. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wang G, Liu G, Ye Y, Fu Y and Zhang X:
Upregulation of miR-34a by diallyl disulfide suppresses invasion
and induces apoptosis in SGC-7901 cells through inhibition of the
PI3K/Akt signaling pathway. Oncol Lett. 11:2661–2667. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Sun XP, Dong X, Lin L, Jiang X, Wei Z,
Zhai B, Sun B, Zhang Q, Wang X, Jiang H, et al: Up-regulation of
survivin by AKT and hypoxia-inducible factor 1α contributes to
cisplatin resistance in gastric cancer. FEBS J. 281:115–128. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Tang T, Su R, Wang B and Zhang Y: An
integrated approach of predicted miR-34a targets identifies a
signature for gastric cancer. Oncol Lett. 10:653–660. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Suzuki R, Yamamoto E, Nojima M, Maruyama
R, Yamano HO, Yoshikawa K, Kimura T, Harada T, Ashida M, Niinuma T,
et al: Aberrant methylation of microRNA-34b/c is a predictive
marker of metachronous gastric cancer risk. J Gastroenterol.
49:1135–1144. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Saunders MA, Liang H and Li WH: Human
polymorphism at microRNAs and microRNA target sites. Proc Natl Acad
Sci USA. 104:3300–3305. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Cai M, Zhang Y, Ma Y, Li W, Min P, Qiu J,
Xu W, Zhang M, Li M, Li L, et al: Association between microRNA-499
polymorphism and gastric cancer risk in Chinese population. Bull
Cancer. 102:973–978. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Jiang J, Jia ZF, Cao DH, Wu YH, Sun ZW and
Cao XY: Association of the miR-146a rs2910164 polymorphism with
gastric cancer susceptibility and prognosis. Future Oncol.
12:2215–2226. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Xu Y, Liu L, Liu J, Zhang Y, Zhu J, Chen
J, Liu S, Liu Z, Shi H, Shen H and Hu Z: A potentially functional
polymorphism in the promoter region of miR-34b/c is associated with
an increased risk for primary hepatocellular carcinoma. Int J
Cancer. 128:412–417. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Yang C, Ma X, Liu D, Wang Y, Tang R, Zhu
Y, Xu Z and Yang L: Promoter polymorphisms of miR-34b/c are
associated with risk of gastric cancer in a Chinese population.
Tumour Biol. 35:12545–12554. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Pan XM, Sun RF, Li ZH, Guo XM, Qin HJ and
Gao LB: Pri-miR-34b/c rs4938723 polymorphism is associated with a
decreased risk of gastric cancer. Genet Test Mol Biomarkers.
19:198–202. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Li H, Diao S, Li J, Ma B and Yuan S: An
updated meta-analysis of 23 case-control studies on the association
between miR-34b/c polymorphism and cancer risk. Oncotarget.
8:28888–28896. 2017.PubMed/NCBI
|
|
92
|
Wang X, Li J, Dong K, Lin F, Long M,
Ouyang Y, Wei J, Chen X, Weng Y, He T and Zhang H: Tumor suppressor
miR-34a targets PD-L1 and functions as a potential
immunotherapeutic target in acute myeloid leukemia. Cell Signal.
27:443–452. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Cortez MA, Ivan C, Valdecanas D, Wang X,
Peltier HJ, Ye Y, Araujo L, Carbone DP, Shilo K, Giri DK, et al:
PDL1 Regulation by p53 via miR-34. J Natl Cancer Inst. 108(pii):
djv3032015.PubMed/NCBI
|
|
94
|
Di Martino MT, Leone E, Amodio N, Foresta
U, Lionetti M, Pitari MR, Cantafio ME, Gullà A, Conforti F, Morelli
E, et al: Synthetic miR-34a mimics as a novel therapeutic agent for
multiple myeloma: In vitro and in vivo evidence. Clin Cancer Res.
18:6260–6270. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Liu C, Kelnar K, Liu B, Chen X,
Calhoun-Davis T, Li H, Patrawala L, Yan H, Jeter C, Honorio S, et
al: The microRNA miR-34a inhibits prostate cancer stem cells and
metastasis by directly repressing CD44. Nat Med. 17:211–215. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Di Martino MT, Campani V, Misso G, Gallo
Cantafio ME, Gullà A, Foresta U, Guzzi PH, Castellano M, Grimaldi
A, Gigantino V, et al: In vivo activity of miR-34a mimics delivered
by stable nucleic acid lipid particles (SNALPs) against multiple
myeloma. PLoS One. 9:e900052014. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Daige CL, Wiggins JF, Priddy L,
Nelligan-Davis T, Zhao J and Brown D: Systemic delivery of a miR34a
mimic as a potential therapeutic for liver cancer. Mol Cancer Ther.
13:2352–2360. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Beg MS, Brenner AJ, Sachdev J, Borad M,
Kang YK, Stoudemire J, Smith S, Bader AG, Kim S and Hong DS: Phase
I study of MRX34, a liposomal miR-34a mimic, administered twice
weekly in patients with advanced solid tumors. Invest New Drugs.
35:180–188. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Shi L, Lin H, Li G, Sun Y, Shen J, Xu J,
Lin C, Yeh S, Cai X and Chang C: Cisplatin enhances NK cells
immunotherapy efficacy to suppress HCC progression via altering the
androgen receptor (AR)-ULBP2 signals. Cancer Lett. 373:45–56. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Yan LH, Chen ZN, Li L, Chen J, Mo XW, Qin
YZ, Wei WE, Qin HQ, Lin Y and Chen JS: E2F-1 promotes DAPK2-induced
anti-tumor immunity of gastric cancer cells by targeting miR-34a.
Tumor Biol. Oct 4–2016.(Epub ahead of print). View Article : Google Scholar
|














