Hepatocellular carcinoma (HCC) is one of the most
common malignant tumors of the digestive system. In 2015, 466,100
patients were newly diagnosed with HCC in China, and the number of
deaths caused by HCC deaths was ~422,100 (1). In China, HCC is one of the four major
causes of cancer-related deaths. HCC is a hypermetabolic tumor that
consumes more oxygen than the surrounding normal tissues. However,
the uncontrolled proliferation of HCC cells leads to an
insufficient oxygen supply and the rapidly growing tumor not only
quickly consumes oxygen but also lacks adequate vascularization,
subsequently generating a hypoxic microenvironment.
Hypoxia-inducible factors (HIFs) are recognized as crucial
transcriptional regulators that are activated under hypoxia
(2). A number of recent studies have
documented the involvement of HIFs in HCC cell proliferation,
angiogenesis, invasion and metastasis (3,4). In
addition, progress has been made in the development of HCC
therapies involving the targeting HIFs (5). Currently, the research on HIFs is
focused on two aspects, the mechanism of transcriptional regulation
of HIFs and cancer therapy targeting HIFs. Therefore, the present
review examined the processes of regulation and activation of HIFs
in HCC, and focused on the progress of research on the function of
HIFs in HCC.
The rapid proliferation of cancer cells leads to the
rapid consumption of tissue oxygen. When the rate of oxygen
consumption exceeds the rate of oxygen supply by the circulation,
hypoxia develops (2). A hypoxic state
activates a series of adaptive responses of cells, which are
primarily mediated by HIFs. The human genome encodes three
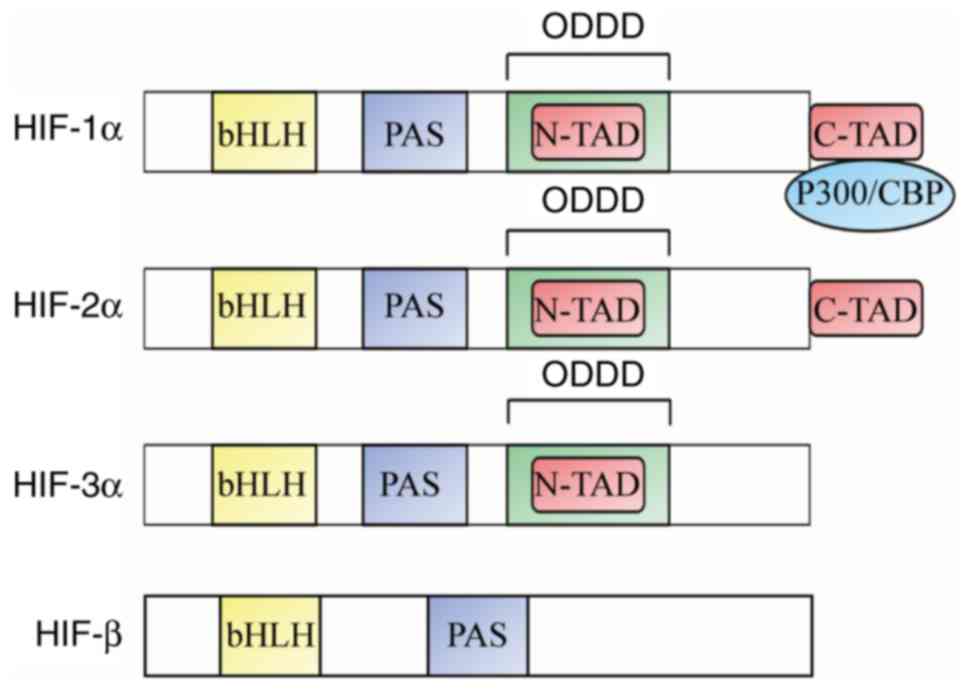
different HIF subtypes: HIF-1α, HIF-2α and HIF-3α (Fig. 1). HIFs are heterodimers composed of a
functional α subunit and a stably expressed β subunit (6). The N-terminus of HIFs has a basic
helix-loop-helix (bHLH) domain and a Per-ARNT-Sim (PAS) domain that
participate in the heterodimerization of the α and β subunits.
These domains also mediate HIF binding to the hypoxia response
element (HRE) in a target gene promoter. The C-terminus of HIF
proteins includes two transactivation domains (TAD), an N-terminal
(N)-TAD and a C-terminal (C)-TAD. The N-TAD domain serves an
essential function in activating HIF-1α or HIF-2α target genes;
N-TAD is the major transactivation domain responsible for HIF-1α or
HIF-2α target gene specificity; as a transcriptional activation
domain, N-TAD may serve as an important cofactor for interaction
sites. Transcriptional cooperation between HIF-1α and certain
factors (such as SMAD3/4 and ETS-1) can induce activation of
multiple HIF target genes under hypoxic conditions (7). The C-TAD acts to recruit
p300/CREB-binding protein (CBP) and other auxiliary transcription
factors. In addition, the structure of HIFs includes an
oxygen-dependent degradation domain (ODDD), which overlaps with
N-TAD, but its function is different from N-TAD. The ODDD serves as
the recognition site of the von Hippel-Lindau tumor suppressor
protein (pVHL) and is involved in the stabilization of proteins and
the regulation of intracellular oxygen concentration. The β subunit
is constitutively expressed, it is not regulated by intracellular
oxygen concentration, and does not have transcriptional activity
alone; only a heterodimer of HIF-α and HIF-β subunits is active.
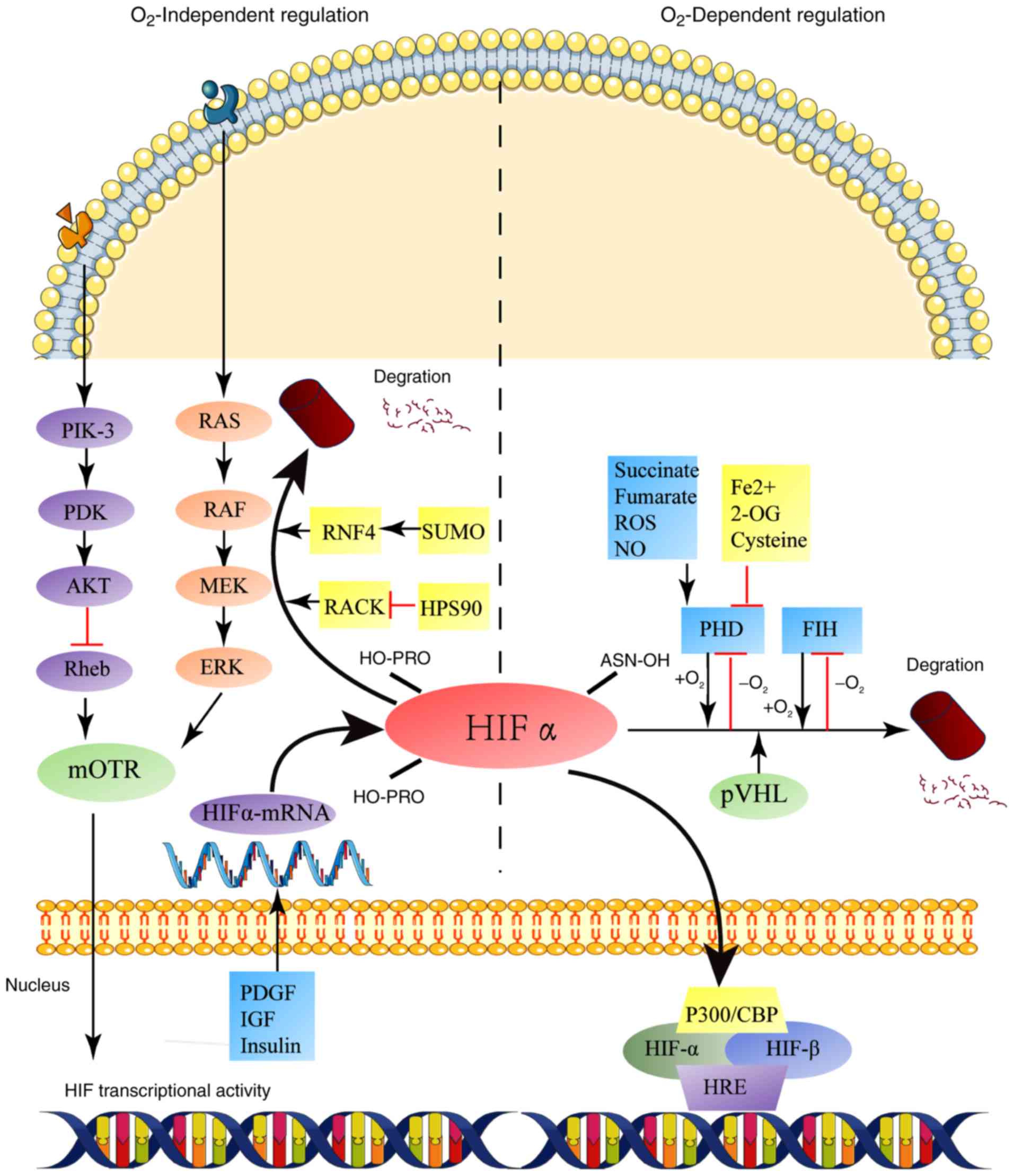
The ODDD contains two proline residues that can be hydroxylated by
the prolyl hydroxylase domain (PHD) enzymes. Hydroxylated HIF
subtypes are recognized by pVHL, which is ubiquitinated by
pVHL-related elongin BC-Cul2 ubiquitin ligase complex. Hydroxylated
HIF-1α binds to pVHL, which recruits elongin B, elongin C, cullin-2
and loop cassette 1 to form the E3 ubiquitin ligase complex. Unlike
the targeted proteasomal degradation, HIF-1α forms the E3 ubiquitin
ligase complex, which ubiquitinates HIF-1α and is ultimately
mediated by the 26S proteasome (8–10), whereas
HIF-2α is ubiquitinated by the of E2 ubiquitin-binding enzyme; but,
both HIF-1α and HIF-2α are subsequently degraded by 26S proteasome
(8).
PHDs are key enzymes of this degradation process,
which uses oxygen and 2-ketoglutarate as substrates, and
Fe2+ and ascorbate as co-factors of dioxygenase
(Fig. 2). The activity of HIFs can
also be suppressed by the HIF-1 inhibitor, such as factor
inhibiting HIF-1α (FIH-1). The catalytic effect of FIH-1 is similar
to that of PHD, which also requires oxygen and 2-ketoglutarate as
substrates. Factor inhibiting HIF-1α (FIH) is an asparaginyl
hydroxylase that catalyzes the hydroxylation of asparagine 803
(Asn803) on C-TAD, preventing HIF-1α from interacting with p300/CBP
and inhibiting its transcriptional activity. However, both PHDs and
HIFs are oxygen-dependent and, therefore, are inactive under
hypoxic conditions, forming stable aggregates of HIF subtypes in
the cytoplasm (11,12). Additionally, PHD activity can be
inhibited by numerous important metabolites, including reactive
oxygen species (ROS), nitric oxide (NO), succinate and fumarate
(13). By contrast, cysteine may
enhance PHD2 activity by inhibiting autoxidation (14).
HIF expression can also be regulated by other
factors, including growth factors such as platelet-derived growth
factor (PDGF), insulin-like growth factor 1 (IGF-1), insulin and
heregulin (Fig. 2). The
Akt/HIF-1a/PDGF-BB autocrine signaling loop is formed under hypoxic
conditions to increase the chemosensitivity of liver cancer cells
(15). Previous studies have shown
that IGF-1 affects HIF-1α and HIF-2α protein synthesis (16,17).
Insulin regulates HIF-1α by a ROS-sensitive activation of Sp1 in
3T3-L1 preadipocytes (18); this is a
novel transcriptional mechanism by which insulin is involved in
Sp1. Heregulin stimulates HIF-1α synthesis via a
rapamycin-dependent manner (19).
Acetyltransferases can acetylate the lysine residue at position 532
of HIF-1α, enhancing the binding ability of pVHL to HIF-1α and,
ultimately, promoting its degradation (20). Receptor for activated protein C kinase
1 (RACK1) and heat shock protein 90 (Hsp90) compete to bind to the
PAS region of HIF-1α; RACK1 enhances the binding of HIF-1α to E3
ligase and promotes degradation, whereas Hsp90 stabilizes HIF-1α
and prevents its degradation (Fig. 2)
(21).
The expression and activity of HIF-2α are also
regulated by certain non-oxygen-dependent pathways, such as small
ubiquitin-related modifier (SUMO) modification. SUMO modification
is the main mechanism of HIF-2α degradation under hypoxia, which
can negatively regulate the expression of HIF-2α. HIF-2α binds
covalently to SUMO-2 via Lys394, resulting in its modification by
SUMO. SUMO-modified HIF-2α is degraded by a mechanism involving
SUMO-dependent E3 ubiquitin-protein ligase RNF4 and pVHL (Fig. 2) (22).
Although numerous studies have focused on HIF-1α and
HIF-2α, our understanding of the role of HIF-3α in cancer cells is
limited (23). It has been reported
that HIF-3α can also be activated under hypoxic conditions and
regulate the transcription and protein stability of HIF-1α
(24–26). In addition, HIF-3α can activate the
transcription of a set of specific target genes, which partially
overlaps with genes upregulated by HIF-1α and HIF-2α, but their
role remains to be demonstrated in future studies (27–29).
A large number of clinical studies have demonstrated
a relationship between HIFs and metastasis, recurrence, vascular
proliferation and prognosis of patients with HCC (Table I). The data indicate that the
expression of HIF-1α in HCC tissues was higher compared with that
in corresponding adjacent tissues. Overexpression of HIF-1α is
associated with poor prognosis in patients with HCC; however, some
recent studies have not reported that expression of HIF-2α or
HIF-3α in HCC is associated with prognosis (Table I).
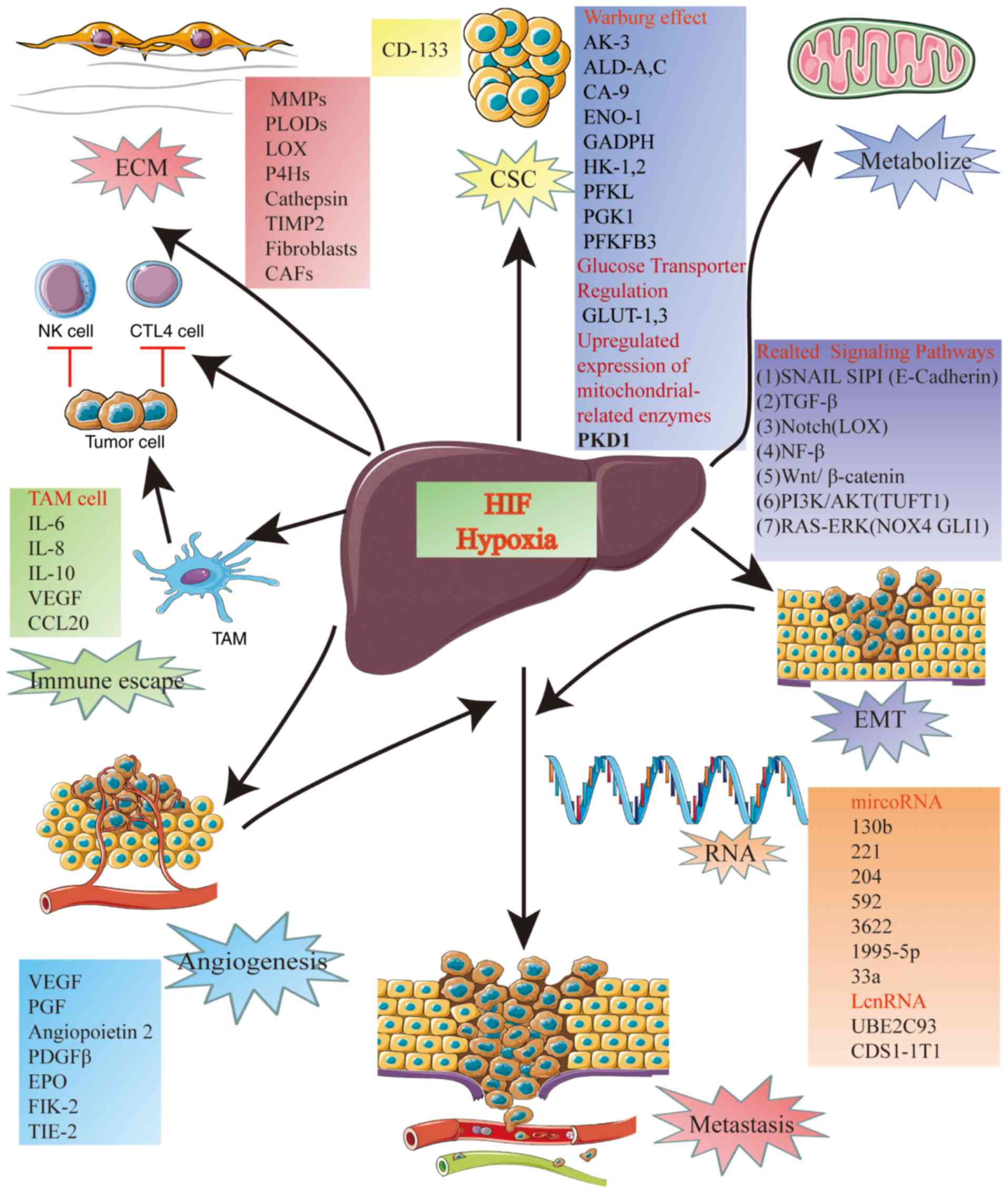
A number of previous studies have demonstrated a
complex relationship between HIF and HCC (30,31). The
relationship between HIFs and HCC include, metabolism, immune
escape, angiogenesis, metastasis, extracellular matrix (ECM)
remodeling and activity of cancer stem cells (CSCs) (Fig. 3).
The rapid proliferation of cancer cells requires a
large amount of energy, resulting in increased consumption of
oxygen, which leads to the generation of a hypoxic environment in
the tumor tissue. Under hypoxia, tumor cells undergo a transition
from aerobic to anaerobic metabolism. This difference in metabolism
between normal and cancer cells was first identified in 1920
(32). Normal cells under physiologic
oxygen concentration convert glucose into pyruvate, which is
further metabolized in the mitochondria via the tricarboxylic acid
cycle and oxidative phosphorylation. In these cells, the
availability of oxygen inhibits the rate of glycolysis (Pasteur
effect), enables mitochondrial respiration, increases ATP levels
and inhibits the activity of phosphofructokinase (PFK) responsible
for glycolysis (33). Under hypoxic
conditions, the final product of anaerobic glycolysis is pyruvic
acid, which is subsequently metabolized to lactic acid. In
comparison with non-malignant tissues, tumor cells rely more on the
use of glycolysis to support their energy needs, even when oxygen
is available, a phenomenon called the Warburg effect (34). Tumor cells are known to produce energy
by generating ATP in anaerobic glycolysis, a process mainly
regulated by HIF-1α (35,36). HIF-1α accelerates the glycolysis
pathway of cancer cells by activating related target genes and
transcription products. This activation may occur through three
distinct mechanisms.
The first mechanism, the metabolism of HCC, is often
related to the Warburg effect, involves HIF-1 activation of key
enzymes involved in glucose metabolism and glycolysis (37,38).
Overexpression of HIF-1α in cancer cells increases the activities
of several isoenzymes that are different from those in normal
tissues, including adenylate kinase 3 (AK3), aldolase-A (ALD-A) and
ALD-C, carbonic anhydrase 9 (CA9), enolase 1 (ENO1), glucose
transporter (GLUT)-1 and GLUT-3, GAPDH, hexokinase (HK)-1 and HK2,
L-lactate dehydrogenase A chain (LDHA), liver-type PKF (PFKL),
phosphoglycerate kinase 1 (PGK1) and
6-phosphofructo-2-kinase/fructose-2,6-bisphosphate 3 (PFKFB3)
(Fig. 3).
In the second mechanism, induction of glucose
transporter regulation, HIF-1α induces overexpression and increased
activity of several glycolytic protein isoforms, including GLUT1
and GLUT3. HIF1α-induced glucose transport is important for
glycolytic flux control and provides new therapeutic targets for
inhibiting HCC growth and progression (Fig. 3) (39).
Transcriptional activity of HIF-1, in the third
mechanism, increases the expression of mitochondrial-related
enzymes, such as pyruvate dehydrogenase kinase 1 (PDK1), which can
inhibit the conversion of pyruvate to acetyl coenzyme A and, as a
result, reduce the level of oxidative phosphorylation and oxygen
consumption by the mitochondria (Fig.
3) (40).
In addition to the above mechanisms, HIF-1α can
reduce intracellular pH by promoting anaerobic glycolysis and
increasing the concentration of lactic acid increase (41). Compared with normal tissues, GLUT1,
LDHA, HK1, pyruvate kinase PKM2 and voltage-dependent
anion-selective channel protein 1 (VDAC-1) expression levels were
revealed to be significantly higher in primary HCC tissues and its
metastases (42).
Progression and metastasis of tumors can take place
only if both primary and metastatic tumors have the ability to
escape immune surveillance. Numerous studies have demonstrated that
hypoxia and HIFs are associated with the evasion of immune response
by tumor cells (43,44). The function of immune cells is
regulated by HIF1-dependent signaling mechanisms. During hypoxia,
HIFs induce the resistance of tumor cells to CD8 cytotoxic T
lymphocytes (CTL) and natural killer (NK) cells. The mechanisms
involved include inhibition of apoptosis (45) and activation of autophagy (46). Additionally, hypoxia can upregulate
the expression of extracellular enzymes CD39 and CD73 that produce
adenosine, increasing its concentration in the cell environment.
Adenosine strongly inhibits the anti-tumor function of activated T
cells and NK cells by binding to its A2A receptor (47). HIFs can also suppress the immune
response against the tumor by acting on the macrophages, so-called
tumor-associated macrophages (TAMs), infiltrating the tumor
microenvironment (48). TAMs have
been repeatedly demonstrated to promote the growth, invasion and
metastasis of tumor cells by secreting cytokines such as
interleukin (IL)-10, transforming growth factor β (TGF-β), IL-6,
VEGF and IL-8, as well as matrix metalloproteinases (MMPs)
(Fig. 3). The cytokines and MMPs
stimulate the tumor cell proliferation, epithelial-mesenchymal
transition (EMT), induce neovascularization, promote remodeling of
the ECM and inhibit the anti-tumor immune function of the organism
(49). In patients with HCC, TAM
infiltration in the liver tissue around the tumor has been
associated with poor prognosis (50).
It has also been reported that in late-stage HCC, a large number of
triggering receptor expressed on myeloid cells 1 (TREM-1)-positive
TAMs indirectly affect the cytotoxic function of CD8+ T
cells and trigger their apoptosis (51). A previous study demonstrated that
specific scavenging of macrophages with chlorophosphate liposomes
resulted in significant suppression of tumor growth and
angiogenesis (52). The role of TAMs
was also documented in a study in which their inhibition delayed
the growth of HCC in nude mice (53).
The role of macrophages in HCC was also underscored by the
demonstration that expression of hypoxia-induced high mobility
group box-1 protein (HMGB1) promotes tumor invasion and metastasis
in animal models of HCC by regulating macrophage-derived IL-6
(54). A previous study demonstrated
that hypoxia promotes the immunosuppressive phenotype of HCC cell
lines through upregulation of HIF1-dependent C-C motif chemokine 20
(CCL20) expression, and CCL20 significantly induces indoleamine
2,3-dioxygenase (IDO) expression in monocyte-derived macrophages
(55). This study also showed a link
between elevated CCL20 levels and poor survival in patients with
liver cancer, suggesting a link between microenvironment of
immunosuppressive hypoxic tumors and promotion of metastasis
(55).
In contrast to HIF-1α, HIF-2α is only expressed
during normal development of blood vessels and lungs (65). It has also been detected in tumor
vascular endothelial cells, tumor cells and TAMs (66); and hypoxia-inducible expression of
HIF-2α has been reported in the brain, lung, heart, liver,
duodenum, pancreas and kidney of mice (67). HIF-2α mainly acts on
angiogenesis-related genes, including VEGF, erythropoietin (EPO),
VEGF receptor 2 (VEGFR2), angiogenin, and tyrosine-protein kinase
receptor TIE-2 (68,69); experiments using different tumor cell
lines and animal models have demonstrated that HIF-2α activates
tumor angiogenesis by upregulating VEGF. Additionally, HIF-2α forms
a complex with transcription-assisted activator ETS proto-oncogene
1 (ETS-1), and binds to HRE4 on the promoter of VEGFR2, activating
its expression (70).
Intrahepatic and extrahepatic metastasis is the
major contributor to poor prognosis in patients with HCC. Invasion
and metastasis of tumors is a complex process in which the first
step involves EMT. In the process of EMT, polar epithelial cells
transform into mobile stromal cells, gaining the ability to migrate
to distant sites. HIF-1α is a crucial regulator of EMT under
hypoxic conditions, acting through seven distinct mechanisms
detailed in the subsections below (Fig.
3).
Inactivation of epithelial (E)-cadherin, a protein
essential for cell adhesion, results in the weakening of cell-cell
contacts and increased mobility, initiating EMT. HIF-1α inhibits
the expression of E-cadherin by upregulating SNAI1 and SIP1,
transcriptional inhibitors of E-cadherin (71). HIF-1α regulates SNAI1 by binding to
two HREs on the SNAI1 promoter, affecting the expression of
E-cadherin, as well as N-cadherin and vimentin, activating EMT in
HCC cells and promoting HCC invasion and metastasis (72).
The TGF-β signaling pathway is widely involved in
embryonic development, tissue and organ formation, cell
proliferation, apoptosis, differentiation and migration. TGF-β has
a dual function in the development of tumors. TGF-β signaling
pathway induces EMT, facilitating the invasion and metastasis of
tumors (73). It has been also
demonstrated that hypoxia is an important stimulator of EMT by
activating HIFs (74). Under hypoxic
conditions, HIF expression in hepatocytes promotes TGF-β signaling;
HIF and TGF-β signaling contribute to the mechanism of
hypoxia-stimulated hepatocyte EMT (74). It has been reported that the TGF-β1
pathway serves an important role in the regulation of liver cancer
by regulating SMAD4, SMAD2/3, cleaved Notch1, and β-catenin
proteins (75).
The Notch signaling pathway regulates embryonic
development and differentiation, and proliferation and apoptosis of
mature cells. Notch signaling induces EMT primarily by two
mechanisms. The first one involves the upregulation of SNAI1
achieved by Notch-mediated recruitment of HIF-1α and the resulting
increase in lysyl oxidase (LOX), which stabilizes SNAI1, thus
promoting EMT (76). The second
mechanism relies on the interaction of Notch with the TGF-β/SMAD
pathway, which also activates EMT (77). Although the molecular mechanisms
underlying hypoxia and Notch pathway activation are not clear,
there is indeed a link between them. Hypoxia activates
Notch-responsive promoters and increases expression of Notch direct
downstream genes; the Notch intracellular domain interacts with
HIF-1α, and after activation of Notch under hypoxic conditions,
HIF-1α is recruited to the Notch reactive promoter (78).
The presence of a bi-directional correlation between
HIF and NF-κB has also been reported, in which NF-κB can induce HIF
and HIF can also regulate NF-κB (79). Cancer is characterized by the presence
of hypoxia and inflammation. Hypoxia has been demonstrated to
promote inflammation through the regulation of gene expression by
oxygen-sensitive transcriptional regulators, including HIF and
NF-κB (80). The basis for this
association includes the regulation of the components of the NF-κB
pathway and the transcriptional regulation of HIF-1 under hypoxia
(81).
Wnt regulates the growth, proliferation, invasion
and metastasis of cancer cells. Under hypoxic conditions, an
increase of Wnt3a upregulates the expression of β-catenin and
promotes EMT (82). A previous study
reported that the Wnt/β-catenin signaling pathway enhances the
transcriptional activity of HIF-1α and inhibits the apoptosis of
HCC, as well as inducing EMT and triggering HCC metastasis
(83). In addition, hypoxia promotes
HCC cell migration and angiogenesis by regulating the expression of
B-cell CLL/lymphoma 9 (BCL9), which activates Wnt/β-catenin
signaling pathway (84).
PIK3/AKT signaling is crucially involved in tumor
development. Hypoxia induces the expression of tuftelin1 (TUFT1) in
a HIF-1α-dependent manner (85). In
turn, TUFT1 activates the Ca2+/PI3K/AKT pathway,
promoting HCC cell growth, metastasis and EMT in vitro and
in vivo.
Hypoxia significantly promotes the progression of
EMT and is associated with activation of the non-canonical Hedgehog
(Hh) signaling pathway. HIF-1α knockdown attenuates hypoxia-induced
membrane-spanning protein SMO and glioma-associated oncogene 1
(GLI1) expression and inhibits EMT progression. In addition, SMO
inhibitors or GLI1 small interfering (si)RNA can also reverse
hypoxia-driven EMT under hypoxic conditions. It is suggested that
non-canonical Hh signaling serves an important role in
hypoxia-induced EMT. Hypoxia increases reactive oxygen species
(ROS) production, and ROS inhibitors (NACs) block GLI1-dependent
EMT processes under hypoxic conditions. In hypoxic HCC cells,
nicotinamide adenine dinucleotide phosphate (NADPH) oxidase 4
(NOX4) expression was found to increase at mRNA and protein levels.
siRNA-mediated knockdown of NOX4 expression abolishes
hypoxia-induced ROS production and hypoxia-induced GLI1-dependent
EMT. Hypoxia triggers ROS-mediated GLI1-dependent EMT progression
by inducing NOX4 expression. Non-canonical Hh pathway regulates
HIF-1α/NOX4/ROS signaling pathway under hypoxic conditions to
regulate EMT processes in HCC cells (86).
A relationship has also been identified between HIF
signaling and p53 family members. A previous study reported that
due to the binding of p53 protein to HIF-1α, p53 is stabilized, and
hypoxia induction of transcriptionally active wild-type p53 gene is
achieved (87). Conversely, p53 and
p73 interact with HIF-1α, suppressing its activity, thereby
inhibiting the migration and metastasis of tumor cells (88,89). A
number of studies have demonstrated that microRNAs (miRNAs) are
also closely related to the migration and metastasis of tumors, and
their effects involve the activity of HIF-1α. For example, miRNA
(miR)-130b and miR-21 can activate EMT through the PTEN/AKT/HIF-1α
pathway and enhance HCC metastasis (90,91).
Hypoxia-induced downregulation of miR-204, which acts as a
post-transcriptional regulator of vasodilator-stimulated
phosphoprotein (VASP) expression, promotes intrahepatic metastasis
of HCC (92). miR-199a-5p (93), miR-592 (94) and miR-3662 regulate the Warburg effect
and HCC progression (95) by reducing
the expression of HIF-1α. Hypoxia induction and up-regulation of
HIF can lead to downregulation of miR-33a expression in HCC cells;
miR-33a controls EMT and invasiveness of HCC by downregulating
Twist1 (96). miR-26a impacts HCC
angiogenesis through the PIK3C2/AKT/HIF-1α/VEGFA pathway (97).
In addition to miRNAs, long non-coding RNAs
(lncRNAs) can also promote HCC metastasis. The lncRNA UBE2CP3
triggers the proliferation and migration of HCC cells by activating
ERK/HIF-1α/p70S6K/VEGFA signal transduction (98). Previous in vitro experiments
demonstrated that the lncRNA CPS1-intronic transcript 1
significantly suppresses proliferation, migration and invasion of
cells by reducing the activity of Hsp90 and HIF-1α, thus inhibiting
the EMT (99).
CSCs have an important function in the initiation,
development, recurrence and metastasis of tumors. Studies on HIF
and stem cells focused on the role of HIF in hematopoietic stem
cells (105); based on data
suggesting the involvement of HIF in the function of hematopoietic
stem cells, studies have demonstrated that the HIF signaling
pathway serve an important role in the induction and maintenance of
CSC and EMT phenotypes, and regulates its function by regulating
multiple complex signaling molecules within the tumor
microenvironment (106). A recent
study reported that hypoxia significantly enhances stem
cell-related properties of HCC cells, an effect that can be
abolished by the knockdown of HIF-1α or HIF-2α (3). Additionally, HIF-1α-specific small
interfering RNA treatment markedly reduces the expression of CD133
in CSCs at the RNA and protein levels (107). Importantly, EMT activation can
induce CSC characteristics. Notch1 mediates the process of
EMT-induced CSCs by direct interaction with HIF-1α; upregulation of
the intracellular expression of Notch by HIF-1α can activate EMT
and induce HCC cells to acquire the features of CSC in vitro
(108).
Given the importance of HIF-1α in promoting the
initiation and development of tumors, the possibility of a therapy
targeting HIFs has become a focus of intense research effort. To
date, a number of drugs or compounds inhibiting HIF-1α have been
identified, but the drugs applicable for HCC treatment are still
unsatisfactory. HIF-1α inhibitors can be classified into eight
categories. i) Drugs affecting the HIF-1α signaling pathway.
Typically, these molecules inhibit mTOR and PI3K signaling.
Recombinant analgesic-antineoplastic peptide (rAGAP) is a protein
comprising small ubiquitin-related modifiers linked to
ubiquitin-histidine tags. rAGAP inhibits the AKT/PI3K pathway,
suppressing angiogenesis and tumor progression (109). Circular RNA circ-EPHB4 derived from
the gene coding for a member of the ephrin (Eph) receptor tyrosine
kinase family, EphB4, prevents tumor growth by modulating the
HIF-1α and AKT/PI3K signaling (110). The drug salidroside significantly
increases the sensitivity of HCC to platinum and inhibits
hypoxia-induced EMT by blocking the HIF-1α signaling (111). Rapamycin counteracts the process of
EMT and angiogenesis, thus inhibiting the growth and lung
metastasis in a rat model of HCC (112). Ruscogenin reduces the expression of
MMP-2, MMP-9, urokinase plasminogen activator, VEGF and HIF-1α by
interfering with the PI3K/AKT/mTOR signaling pathway, resulting in
an inhibition of tumor growth (113). The dietary phytochemical
sulforaphane prevents angiogenesis of HCC by inhibiting
STAT3/HIF-1α/VEGF signal transduction (114). N1-guanyl-1,7-diaminoheptane (GC7)
enhances the sensitivity of HCC to doxorubicin by reversing the EMT
signaling pathway induced by HIF-1α (115). Everolimus suppresses tumor growth
and angiogenesis by blocking AKT/mTOR signaling pathway in
vitro by promoting cell apoptosis and inhibiting endothelial
cell proliferation (116). Finally,
Huaier polysaccharide TP-1 is a naturally occurring bioactive
macromolecule, found in Huaier fungus, prevents tumor growth and
metastasis by downregulating HIF-1α-VEGF and AUF-1/AEG-1 signal
transduction pathways (117). ii)
Drugs inhibiting the expression of HIF-1α mRNA. Two compounds,
RO70179 and EZN-2968, have been demonstrated to markedly reduce the
expression of HIF-1α in HCC tissues (118). iii) Drugs inhibiting the synthesis
of HIF-1α protein. Topotecan, an inhibitor of topoisomerase, has
been reported to block the entry of the ribosome on HIF-1α mRNA,
preventing translation of the protein (119). Additionally, vorinostat, a histone
deacetylase inhibitor, decreases interaction between acetyl-Hsp90
and HIF-1α, inhibiting HIF-α nuclear translocation (120). iv) Drugs promoting the degradation
of HIF-1α protein. A previous study has reported that evodiamine in
combination with vorinostat accelerated the degradation of HIF-1α
in HCC cells under hypoxic conditions (121). v) Drugs inhibiting HIF-1α
stabilization. Curcumin can induce the clearance of ROS by
upregulating nuclear factor E2-related factor 2 (Nrf2) and
glutathione (GSH), which inhibit the stabilization of HIF-1α, and,
in turn, suppress the expression of connective tissue growth factor
(CTGF), providing a protective effect on HCC (122). vi) Drugs blocking the binding of
HIF-1α to target genes; for example, doxorubicin (115). vii) Drugs inhibiting HIF-1α-mediated
transcriptional activation; for example, bortezomib (123). viii) Drugs used for systemic
therapy. A previous study demonstrated that inhibition of HIF-1α by
systemic therapy with digoxin significantly delayed the development
of HCC (124). In addition,
metformin was reported to enhance the potential of regorafenib by
regulating the levels of HIV TAT-interactive protein (TIP30) and
HIF-2α, and inhibits the recurrence and metastasis of HCC after
hepatectomy (125).
The expression of HIF-1α in HCC is significantly
higher compared with expression in normal liver cells. HIF-1α is a
crucial regulator of the adaptation of HCC cells to the hypoxic
microenvironment and can affect the proliferation, growth,
invasion, metastasis, angiogenesis, apoptosis and drug resistance
of HCC cells by modulating the expression of multiple target genes.
A number of studied have demonstrated the feasibility of using
HIF-1α as a therapeutic target, which suggested that interventions
modifying the activity of HIF-1α by direct or indirect ways may
become effective for the treatment of HCC. Despite the growing
number of studies on HIF-1α and identification of many HIF-1α
inhibitors, their therapeutic application has not moved beyond the
pre-clinical stage. Clinical use of these inhibitors faces multiple
problems which have to be solved urgently. They include limitations
in the specificity of HIF-1α inhibitors and lack of definitive
cytotoxicity of HIF-1α inhibitors toward cancer cells. Therefore,
compounds need to be developed and screened for clinical
application. In the case of YC-1 and other similarly
well-investigated inhibitors, further research on their
pharmacology and toxicology is still needed. Although gene therapy
targeting HIF-1α brings new hope to the treatment of HCC, finding
the target gene is only the first step in the long road to clinical
application. How to construct a safe and efficient vector, how to
search for specific transcriptional regulatory elements in HCC, and
how to rationally apply a combined therapy targeting multiple genes
are critical questions that must be conclusively answered.
Therefore, studies on the function of HIF-1 in HCC have to be
expanded, necessitating additional time before the targeted therapy
of HIF-1α for HCC can be implemented clinically. In addition, the
understanding of the function of HIF-2 and HIF-3 in HCC has only
begun to emerge, although it is already documented that HIF-2α
affects HCC energy metabolism, angiogenesis, cell proliferation and
tumor growth. Other studies have provided information regarding the
stability, transcriptional activity and role of HIF-2α in HCC
growth and progression, but the exact role in HCC remains unclear.
It is generally believed that HIF-2α can be activated in most
hypoxic solid tumors, but whether its activation promotes or
inhibits tumor growth depends on the biological environment of the
tumor. HIF-2α can participate in modulating the progression of HCC
through different signaling pathways. However, the specific role of
HIF-2α in HCC is still controversial, and definite conclusions have
can only be provided by additional experiments. Thus, in-depth
analysis of the function of HIF-2α in HCC may help to better
understand the mechanism of development and metastasis of this
tumor type and to improve the treatment methods. In conclusion,
significant additional research effort is necessary to achieve an
in-depth understanding of the role of HIFs in HCC.
Not applicable.
This study was supported by The Key Laboratory of
Tumor Molecular Diagnosis and Individualized Medicine of Zhejiang
Province, Zhejiang Provincial People's Hospital (People's Hospital
of Hangzhou Medical College; Hangzhou, China). This study was also
supported by grants from The National Science and Technology Major
Project for New Drug (grant no. 2017ZX09302003), The National
Natural Science Foundation of China (grant nos. 81874049 and
81602179), The Co-construction of Provincial and Department Project
(grant no. WKJ-ZJ-1919), The Zhejiang Provincial Natural Science
Foundation of China (grant no. LY19H160036), and The Public
Projects of Zhejiang Province (grant no. 2018C37033).
Not applicable.
YG, ZX, DH and QX conceived and designed the review.
YG, ZX, LY, YG, QZ, LH, DH and QX were involved in the collection
and collation of references. YG and ZX collected and assembled the
data presented in Table I. YG and ZX
drew the figures. YG and ZX wrote the manuscript. All authors
approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. Ca Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mckeown SR: Defining normoxia, physoxia
and hypoxia in tumours-implications for treatment response. Br J
Radiol. 87:201306762014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cui CP, Wong CC, Kai AK, Ho DW, Lau EY,
Tsui YM, Chan LK, Cheung TT, Chok KS, Chan ACY, et al: SENP1
promotes hypoxia-induced cancer stemness by HIF-1α deSUMOylation
and SENP1/HIF-1α positive feedback loop. Gut. 66:2149–2159. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wang J, Ma Y, Jiang H, Zhu H, Liu L, Sun
B, Pan S, Krissansen GW and Sun X: Overexpression of von
Hippel-Lindau protein synergizes with doxorubicin to suppress
hepatocellular carcinoma in mice. J Hepatol. 55:359–368. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Blagosklonny MV: Hypoxia-inducible factor:
Achilles' heel of antiangiogenic cancer therapy (Review). Int J
Oncol. 19:257–262. 2001.PubMed/NCBI
|
|
6
|
Dengler VL, Galbraith MD and Espinosa JM:
Transcriptional regulation by hypoxia inducible factors. Crit Rev
Biochem Mol Biol. 49:1–15. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hu CJ, Sataur A, Wang L, Chen H and Simon
MC: The N-terminal Transactivation domain confers target gene
specificity of hypoxia-inducible factors HIF-1alpha and HIF-2alpha.
Mol Biol Cell. 18:4528–4542. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ohh M, Park CW, Ivan M, Hoffman MA, Kim
TY, Huang LE, Pavletich N, Chau V and Kaelin WG: Ubiquitination of
hypoxia-inducible factor requires direct binding to the beta-domain
of the von Hippel-Lindau protein. Nat Cell Biol. 2:423–427. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Maxwell PH, Wiesener MS, Chang GW,
Clifford SC, Vaux EC, Cockman ME, Wykoff CC, Pugh CW, Maher ER and
Ratcliffe PJ: The tumour suppressor protein VHL targets
hypoxia-inducible factors for oxygen-dependent proteolysis. Nature.
399:271–275. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ivan M, Kondo K, Yang H, Kim W, Valiando
J, Ohh M, Salic A, Asara JM, Lane WS and Kaelin WG Jr: HIFalpha
targeted for VHL-mediated destruction by proline hydroxylation:
Implications for O2 sensing. Science. 292:464–468. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Garvalov BK and Acker T: Implications of
oxygen homeostasis for tumor biology and treatment. 903:169–185.
2016.PubMed/NCBI
|
|
12
|
Benita Y, Kikuchi H, Smith AD, Zhang MQ,
Chung DC and Xavier RJ: An integrative genomics approach identifies
Hypoxia Inducible Factor-1 (HIF-1)-target genes that form the core
response to hypoxia. Nucleic Acids Res. 37:4587–4602. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Salminen A, Kauppinen A and Kaarniranta K:
2-Oxoglutarate- dependent dioxygenases are sensors of energy
metabolism, oxygen availability, and iron homeostasis: Potential
role in the regulation of aging process. Cell Mol Life Sci.
72:3897–3914. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Briggs KJ, Koivunen P, Cao S, Backus KM,
Olenchock BA, Patel H, Zhang Q, Signoretti S, Gerfen GJ, Richardson
AL, et al: Paracrine Induction of HIF by glutamate in breast
cancer: EglN1 senses cysteine. Cell. 166:126–139. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lau CK, Yang ZF, Ho DW, Ng MN, Yeoh GC,
Poon RT and Fan ST: An AKT/hypoxia-inducible
factor-1alpha/platelet-derived growth factor-BB autocrine loop
mediates hypoxia-induced chemoresistance in liver cancer cells and
tumorigenic hepatic progenitor cells. Clin Cancer Res.
15:3462–3471. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Akeno N, Robins J, Zhang M, Czyzyk-Krzeska
MF and Clemens TL: Induction of vascular endothelial growth factor
by IGF-I in osteoblast-like cells is mediated by the PI3K signaling
pathway through the hypoxia-inducible factor-2alpha. Endocrinology.
143:420–425. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fukuda R, Hirota K, Fan F, Jung YD, Ellis
LM and Semenza GL: Insulin-like growth factor 1 induces
hypoxia-inducible factor 1-mediated vascular endothelial growth
factor expression, which is dependent on MAP kinase and
phosphatidylinositol 3-kinase signaling in colon cancer cells. J
Biol Chem. 277:38205–38211. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Biswas S, Mukherjee R, Tapryal N, Singh AK
and Mukhopadhyay CK: Insulin regulates hypoxia-inducible factor-1α
transcription by reactive oxygen species sensitive activation of
Sp1 in 3T3-L1 preadipocyte. PLoS One. 8:e621282013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Laughner E, Taghavi P, Chiles K, Mahon PC
and Semenza GL: HER2 (neu) signaling increases the rate of
hypoxia-inducible factor 1alpha (HIF-1alpha) synthesis: Novel
mechanism for HIF-1-mediated vascular endothelial growth factor
expression. Mol Cell Biol. 21:3995–4004. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nayak BK, Feliers D, Sudarshan S,
Friedrichs WE, Day RT, New DD, Fitzgerald JP, Eid A, Denapoli T,
Parekh DJ, et al: Stabilization of HIF-2α through redox regulation
of mTORC2 activation and initiation of mRNA translation. Oncogene.
32:3147–3155. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu LZ, Hu XW, Xia C, He J, Zhou Q, Shi X,
Fang J and Jiang BH: Reactive oxygen species regulate epidermal
growth factor-induced vascular endothelial growth factor and
hypoxia-inducible factor-1alpha expression through activation of
AKT and P70S6K1 in human ovarian cancer cells. Free Radic Biol Med.
41:1521–1533. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Jiang BH, Jiang G, Zheng JZ, Lu Z, Hunter
T and Vogt PK: Phosphatidylinositol 3-kinase signaling controls
levels of hypoxia-inducible factor 1. Cell Growth Differ.
12:363–369. 2001.PubMed/NCBI
|
|
23
|
Lim JH, Lee YM, Chun YS, Chen J, Kim JE
and Park JW: Sirtuin 1 modulates cellular responses to hypoxia by
deacetylating hypoxia-inducible factor 1alpha. Mol Cell.
38:864–878. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu X, Chen S, Tu J, Cai W and Xu Q: HSP90
inhibits apoptosis and promotes growth by regulating HIF-1α
abundance in hepatocellular carcinoma. Int J Mol Med. 37:825–835.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
van Hagen M, Overmeer RM, Abolvardi SS and
Vertegaal AC: RNF4 and VHL regulate the proteasomal degradation of
SUMO-conjugated Hypoxia-Inducible Factor-2alpha. Nucleic Acids Res.
38:1922–1931. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Duan C: Hypoxia-inducible factor 3
biology: Complexities and emerging themes. Am J Physiol Cell
Physiol. 310:C260–C269. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Makino Y, Kanopka A, Wilson WJ, Tanaka H
and Poellinger L: Inhibitory PAS domain protein (IPAS) is a
hypoxia-inducible splicing variant of the hypoxia-inducible
factor-3alpha locus. J Biol Chem. 277:32405–32408. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Maynard MA, Qi H, Chung J, Lee EH, Kondo
Y, Hara S, Conaway RC, Conaway JW and Ohh M: Multiple splice
variants of the human HIF-3 alpha locus are targets of the von
Hippel-Lindau E3 ubiquitin ligase complex. J Biol Chem.
278:11032–11040. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Heikkilä M, Pasanen A, Kivirikko KI and
Myllyharju J: Roles of the human hypoxia-inducible factor (HIF)-3α
variants in the hypoxia response. Cell Mol Life Sci. 68:3885–3901.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chen C and Lou T: Hypoxia inducible
factors in hepatocellular carcinoma. Oncotarget. 8:46691–46703.
2017.PubMed/NCBI
|
|
31
|
Mucaj V, Shay JE and Simon MC: Effects of
hypoxia and HIFs on cancer metabolism. Int J Hematol. 95:464–470.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Warburg O, Wind F and Negelein E: The
metabolism of tumors in the body. J Gen Physiol. 8:519–530. 1927.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Alfarouk KO, Verduzco D, Rauch C,
Muddathir AK, Adil HH, Elhassan GO, Ibrahim ME, David Polo Orozco
J, Cardone RA, Reshkin SJ and Harguindey S: Glycolysis, tumor
metabolism, cancer growth and dissemination. A new pH-based
etiopathogenic perspective and therapeutic approach to an old
cancer question. Oncoscience. 1:777–802. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liberti MV and Locasale JW: The warburg
effect: How does it benefit cancer cells? Trends Biochem Sci.
41:211–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Denko NC: Hypoxia, HIF1 and glucose
metabolism in the solid tumour. Nat Rev Cancer. 8:705–713. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Semenza GL: Regulation of cancer cell
metabolism by hypoxia-inducible factor 1. Semin Cancer Biol.
19:12–16. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Semenza GL: HIF-1: Upstream and downstream
of cancer metabolism. Curr Opin Genet Dev. 20:51–56. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ke Q and Costa M: Hypoxia-Inducible
Factor-1 (HIF-1). Mol Pharmacol. 70:1469–1480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Marín-Hernández A, Gallardo-Pérez JC,
Ralph SJ, Rodríguez-Enríquez S and Moreno-Sánchez R: HIF-1alpha
modulates energy metabolism in cancer cells by inducing
over-expression of specific glycolytic isoforms. Mini Rev Med Chem.
9:1084–1091. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kim JW, Tchernyshyov I, Semenza GL and
Dang CV: HIF-1-mediated expression of pyruvate dehydrogenase
kinase: A metabolic switch required for cellular adaptation to
hypoxia. Cell Metab. 3:177–185. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Weidemann A and Johnson RS: Biology of
HIF-1 alpha. Cell Death Differ. 15:621–627. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Graziano F, Ruzzo A, Giacomini E,
Ricciardi T, Aprile G, Loupakis F, Lorenzini P, Ongaro E, Zoratto
F, Catalano V, et al: Glycolysis gene expression analysis and
selective metabolic advantage in the clinical progression of
colorectal cancer. Pharmacogenomics J. 17:258–264. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Schito L and Semenza GL: Hypoxia-inducible
factors: Master regulators of cancer progression. Trends Cancer.
2:758–770. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Samanta D, Park Y, Ni X, Li H, Zahnow CA,
Gabrielson E, Pan F and Semenza GL: Chemotherapy induces enrichment
of CD47+/CD73+/PDL1+immune evasive
triple-negative breast cancer cells. Proc Natl Acad Sci USA.
115:E1239–E1248. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Terry S, Buart S and Chouaib S: Hypoxic
stress-induced tumor and immune plasticity, suppression, and impact
on tumor heterogeneity. Front Immunol. 8:16252017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Noman MZ, Janji B, Kaminska B, Moer KV,
Pierson S, Przanowski P, Buart S, Berchem G, Romero P, Mami-Chouaib
F and Chouaib S: Blocking hypoxia-induced autophagy in tumors
restores cytotoxic t-cell activity and promotes regression.
Autophagy. 71:5976–5986. 2012.
|
|
47
|
Hatfield SM and Sitkovsky M: A2A adenosine
receptor antagonists to weaken the hypoxia-HIF-1α driven
immunosuppression and improve immunotherapies of cancer. Curr Opin
Pharmacol. 29:90–96. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Vinit K and Gabrilovich DI:
Hypoxia-inducible factors in regulation of immune responses in
tumour microenvironment. Immunology. 143:512–519. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Fukuda K, Kobayashi A and Watabe K: The
role of tumor- associated macrophage in tumor progression. Front
Biosci. 4:787–798. 2012.
|
|
50
|
Zhu XD, Zhang JB, Zhuang PY, Zhu HG, Zhang
W, Xiong YQ, Wu WZ, Wang L, Tang ZY and Sun HC: High expression of
macrophage colony-stimulating factor in peritumoral liver tissue is
associated with poor survival after curative resection of
hepatocellular carcinoma. J Clin Oncol. 26:2707–2716. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Wu Q, Zhou W, Yin S, Zhou Y, Chen T, Qian
J, Su R, Hong L, Lu H, Zhang F, et al: Blocking TREM-1
Tumor-associated macrophages induced by hypoxia reverses
immunosuppression and anti-PD-L1 resistance in liver cancer.
Hepatology. 70:198–214. 2019.PubMed/NCBI
|
|
52
|
Zeisberger SM, Odermatt B, Marty C,
Zehnder-Fjällman AH, Ballmer-Hofer K and Schwendener RA:
Clodronate-liposome- mediated depletion of tumour-associated
macrophages: A new and highly effective antiangiogenic therapy
approach. Br J Cancer. 95:272–281. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kuang DM, Peng C, Zhao Q, Wu Y, Chen MS
and Zheng L: Activated monocytes in peritumoral stroma of
hepatocellular carcinoma promote expansion of memory T helper 17
cells. Hepatology. 51:154–164. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Jiang J, Wang GZ, Wang Y, Huang HZ, Li WT
and Qu XD: Hypoxia-induced HMGB1 expression of HCC promotes tumor
invasiveness and metastasis via regulating macrophage-derived IL-6.
Exp Cell Res. 367:81–88. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Ye LY, Chen W, Bai XL, Xu XY, Zhang Q, Xia
XF, Sun X, Li GG, Hu QD, Fu QH and Liang TB: Hypoxia-induced
epithelial-to-mesenchymal transition in hepatocellular carcinoma
induces an immunosuppressive tumor microenvironment to promote
metastasis. Cancer Res. 76:818–830. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Payne SJ and Louise J: Influence of the
tumor microenvironment on angiogenesis. Future Oncol. 7:395–408.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Branco-Price C, Zhang N, Schnelle M, Evans
C, Katschinski DM, Liao D, Ellies L and Johnson RS: Endothelial
cell HIF-1α and HIF-2α differentially regulate metastatic success.
Cancer Cell. 21:52–65. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
De Francesco EM, Lappano R, Santolla MF,
Marsico S, Caruso A and Maggiolini M: HIF-1α/GPER signaling
mediates the expression of VEGF induced by hypoxia in breast cancer
associated fibroblasts (CAFs). Breast Cancer Res. 15:R642013.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Ahluwalia A and Tarnawski AS: Critical
role of hypoxia sensor-HIF-1α in VEGF gene activation. Implications
for angiogenesis and tissue injury healing. Curr Med Chem.
19:90–97. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Lee K, Zhang H, Qian DZ, Rey S, Liu JO and
Semenza GL: Acriflavine inhibits HIF-1 dimerization, tumor growth,
and vascularization. Proc Natl Acad Sci USA. 106:17910–17915. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Wang W, Xu GL, Jia WD, Wang ZH, Li JS, Ma
JL, Ge YS, Xie SX and Yu JH: Expression and correlation of
hypoxia-inducible factor-1alpha, vascular endothelial growth factor
and microvessel density in experimental rat hepatocarcinogenesis. J
Int Med Res. 37:417–425. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Liu LP, Ho RL, Chen GG and Lai PB:
Sorafenib inhibits hypoxia-inducible factor-1α synthesis:
Implications for antiangiogenic activity in hepatocellular
carcinoma. Clin Cancer Res. 18:5662–5671. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Semenza GL: Defining the role of
hypoxia-inducible factor 1 in cancer biology and therapeutics.
Oncogene. 29:625–634. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Li H, Ge C, Zhao F, Yan M, Hu C, Jia D,
Tian H, Zhu M, Chen T, Jiang G, et al: Hypoxia-inducible factor 1
alpha-activated angiopoietin-like protein 4 contributes to tumor
metastasis via vascular cell adhesion molecule-1/integrin β1
signaling in human hepatocellular carcinoma. Hepatology.
54:910–919. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Tian H, McKnight SL and Russell DW:
Endothelial PAS domain protein 1 (EPAS1), a transcription factor
selectively expressed in endothelial cells. Genes Dev. 11:72–82.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Talks KL, Turley H, Gatter KC, Maxwell PH,
Pugh CW, Ratcliffe PJ and Harris AL: The expression and
distribution of the hypoxia-inducible factors HIF-1alpha and
HIF-2alpha in normal human tissues, cancers, and tumor-associated
macrophages. Am J Pathol. 157:411–421. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Wiesener MS, Jürgensen JS, Rosenberger C,
Scholze CK, Hörstrup JH, Warnecke C, Mandriota S, Bechmann I, Frei
UA, Pugh CW, et al: Widespread hypoxia-inducible expression of
HIF-2alpha in distinct cell populations of different organs. FASEB
J. 17:271–273. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhang T, Niu X, Liao L, Cho EA and Yang H:
The Contributions of HIF-Target Genes to Tumor Growth in RCC. PLoS
One. 8:e805442013. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Feng N, Chen H, Fu S, Bian Z, Lin X, Yang
L, Gao Y, Fang J and Ge Z: HIF-1α and HIF-2α induced angiogenesis
in gastrointestinal vascular malformation and reversed by
thalidomide. Sci Rep. 6:272802016. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Aprelikova O, Wood M, Tackett S,
Chandramouli GV and Barrett JC: Role of ETS transcription factors
in the hypoxia-inducible factor-2 target gene selection. Cancer
Res. 66:5641–5647. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Evans AJ, Russell RC, Roche O, Burry TN,
Fish JE, Chow VW, Kim WY, Saravanan A, Maynard MA, Gervais ML, et
al: VHL promotes E2 box-dependent E-cadherin transcription by
HIF-mediated regulation of SIP1 and snail. Mol Cell Biol.
27:157–169. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhang L, Huang G, Li X, Zhang Y, Jiang Y,
Shen J, Liu J, Wang Q, Zhu J, Feng X, et al: Hypoxia induces
epithelial-mesenchymal transition via activation of SNAI1 by
hypoxia-inducible factor-1α in hepatocellular carcinoma. Bmc
Cancer. 13:1082013. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Willis BC, Liebler JM, Luby-Phelps K,
Nicholson AG, Crandall ED, du Bois RM and Borok Z: Induction of
epithelial-mesenchymal transition in alveolar epithelial cells by
transforming growth factor-beta1: Potential role in idiopathic
pulmonary fibrosis. Am J Pathol. 166:1321–1332. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Copple BL: Hypoxia stimulates hepatocyte
epithelial to mesenchymal transition by hypoxia-inducible factor
and transforming growth factor-beta-dependent mechanisms. Liver
Int. 30:669–682. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Wang XH, Liu MN, Sun X, Xu CH, Liu J, Chen
J, Xu RL and Li BX: TGF-β1 pathway affects the protein expression
of many signaling pathways, markers of liver cancer stem cells,
cytokeratins, and TERT in liver cancer HepG2 cells. Tumor Biol.
37:3675–3681. 2016. View Article : Google Scholar
|
|
76
|
Erler JT and Giaccia AJ: Lysyl oxidase
mediates hypoxic control of metastasis. Cancer Res. 66:10238–10241.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Sahlgren C, Gustafsson MV, Jin S,
Poellinger L and Lendahl U: Notch signaling mediates
hypoxia-induced tumor cell migration and invasion. Proc Natl Acad
Sci USA. 105:6392–6397. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Gustafsson MV, Zheng X, Pereira T, Gradin
K, Jin S, Lundkvist J, Ruas JL, Poellinger L, Lendahl U and
Bondesson M: Hypoxia requires notch signaling to maintain the
undifferentiated cell state. Dev Cell. 9:617–628. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
D'Ignazio L, Batie M and Rocha S: Hypoxia
and inflammation in cancer, focus on HIF and NF-κB. Biomedicines.
5:E212017. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Fitzpatrick SF, Tambuwala MM, Bruning U,
Schaible B, Scholz CC, Byrne A, O'Connor A, Gallagher WM, Lenihan
CR, Garvey JF, et al: An intact canonical NF-κB pathway is required
for inflammatory gene expression in response to hypoxia. J Immunol.
186:1091–1096. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Taylor CT and Cummins EP: The role of
NF-kappaB in hypoxia-induced gene expression. Ann N Y Acad Sci.
1177:178–184. 2010. View Article : Google Scholar
|
|
82
|
Zhang L, Liu H, Mu X, Cui J and Peng Z:
Dysregulation of Fra1 expression by Wnt/β-catenin signalling
promotes glioma aggressiveness through epithelial-mesenchymal
transition. Biosci Rep. 37:BSR201606432017. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Zhang Q, Bai X, Chen W, Ma T, Hu Q, Liang
C, Xie S, Chen C, Hu L, Xu S and Liang T: Wnt/β-catenin signaling
enhances hypoxia-induced epithelial-mesenchymal transition in
hepatocellular carcinoma via crosstalk with hif-1α signaling.
Carcinogenesis. 34:962–973. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Xu W, Zhou W, Cheng M, Wang J, Liu Z, He
S, Luo X, Huang W, Chen T, Yan W and Xiao J: Hypoxia activates
Wnt/β-catenin signaling by regulating the expression of BCL9 in
human hepatocellular carcinoma. Sci Rep. 7:404462017. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Dou C, Zhou Z, Xu Q, Liu Z, Zeng Y, Wang
Y, Li Q, Wang L, Yang W, Liu Q and Tu K: Hypoxia-induced TUFT1
promotes the growth and metastasis of hepatocellular carcinoma by
activating the Ca2+/PI3K/AKT pathway. Oncogene.
38:1239–1255. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Liu Z, Tu K, Wang Y, Yao B, Li Q, Wang L,
Dou C, Liu Q and Zheng X: Hypoxia accelerates aggressiveness of
hepatocellular carcinoma cells involving oxidative stress,
epithelial-mesenchymal transition and non-canonical hedgehog
signaling. Cell Physiol Biochem. 44:1856–1868. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
An WG, Kanekal M, Simon MC, Maltepe E,
Blagosklonny MV and Neckers LM: Stabilization of wild-type p53 by
hypoxia-inducible factor 1alpha. Nature. 392:405–408. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Hansson LO, Friedler A, Freund S, Rüdiger
S and Fersht AR: Two sequence motifs from HIF-1alpha bind to the
DNA-binding site of p53. Proc Natl Acad Sci USA. 99:10305–10309.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Amelio I, Inoue S, Markert EK, Levine AJ,
Knight RA, Mak TW and Melino G: TAp73 opposes tumor angiogenesis by
promoting hypoxia-inducible factor 1α degradation. Proc Natl Acad
Sci USA. 112:226–231. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Liu Z, Wang J, Guo C and Fan X:
microRNA-21 mediates epithelial-mesenchymal transition of human
hepatocytes via PTEN/AKT pathway. Biomed Pharmacother. 69:24–28.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Chang RM, Xu JF, Fang F, Yang H and Yang
LY: MicroRNA-130b promotes proliferation and EMT-induced metastasis
via PTEN/p-AKT/HIF-1α signaling. Tumor Biol. 37:10609–10619. 2016.
View Article : Google Scholar
|
|
92
|
Liu Z, Wang Y, Dou C, Xu M, Sun L, Wang L,
Yao B, Li Q, Yang W, Tu K and Liu Q: Hypoxia-induced up-regulation
of VASP promotes invasiveness and metastasis of hepatocellular
carcinoma. Theranostics. 8:4649–4663. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Li B, He L, Zuo D, He W, Wang Y, Zhang Y,
Liu W and Yuan Y: Mutual Regulation of MiR-199a-5p and HIF-1α
modulates the warburg effect in hepatocellular carcinoma. J Cancer.
8:940–949. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Jia YY, Zhao JY, Li BL, Gao K, Song Y, Liu
MY, Yang XJ, Xue Y, Wen AD and Shi L: miR-592/WSB1/HIF-1α axis
inhibits glycolytic metabolism to decrease hepatocellular carcinoma
growth. Oncotarget. 7:35257–35269. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Chen Z, Zuo X, Zhang Y, Han G, Zhang L, Wu
J and Wang X: MiR-3662 suppresses hepatocellular carcinoma growth
through inhibition of HIF-1α-mediated Warburg effect. Cell Death
Dis. 9:5492018. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Guo XF, Wang AY and Liu J:
HIFs-MiR-33a-Twsit1 axis can regulate invasiveness of
hepatocellular cancer cells. Eur Rev Med Pharmacol Sci.
20:3011–3016. 2016.PubMed/NCBI
|
|
97
|
Chai ZT, Kong J, Zhu XD, Zhang YY, Lu L,
Zhou JM, Wang LR, Zhang KZ, Zhang QB, Ao JY, et al: MicroRNA-26a
inhibits angiogenesis by down-regulating VEGFA through the
PIK3C2α/AKT/HIF-1α pathway in hepatocellular carcinoma. PLoS One.
8:e779572013. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Lin J, Cao S, Wang Y, Hu Y, Liu H, Li J,
Chen J, Li P, Liu J, Wang Q and Zheng L: Long non-coding RNA
UBE2CP3 enhances HCC cell secretion of VEGFA and promotes
angiogenesis by activating ERK1/2/HIF-1α/VEGFA signalling in
hepatocellular carcinoma. J Exp Clin Cancer Res. 37:1132018.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Wang TH, Yu CC, Lin YS, Chen TC, Yeh CT,
Liang KH, Shieh TM, Chen CY and Hsueh C: Long noncoding RNA
CPS1-IT1 suppresses the metastasis of hepatocellular carcinoma by
regulating HIF-1α activity and inhibiting epithelial-mesenchymal
transition. Oncotarget. 7:43588–43603. 2016.PubMed/NCBI
|
|
100
|
Bonnans C, Chou J and Werb Z: Remodelling
the extracellular matrix in development and disease. Nat Rev Mol
Cell Biol. 15:786–801. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Rankin EB and Giaccia AJ: Hypoxic control
of metastasis. Science. 352:175–180. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Kai AK, Chan LK, Lo RC, Lee JM, Wong CC,
Wong JC and Ng IO: Down-regulation of TIMP2 by
HIF-1α/miR-210/HIF-3α regulatory feedback circuit enhances cancer
metastasis in hepatocellular carcinoma. Hepatology. 64:473–487.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Kalluri R: The biology and function of
fibroblasts in cancer. Nat Rev Cancer. 16:582–598. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Tse AP, Sze KM, Shea QT, Chiu EY, Tsang
FH, Chiu DK, Zhang MS, Lee D, Xu IM, Chan CY, et al: Hepatitis
transactivator protein X promotes extracellular matrix modification
through HIF/LOX pathway in liver cancer. Oncogenesis. 7:442018.
View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Cheng ZC and Sadek HA: Hypoxia and
metabolic properties of hematopoietic stem cells. Antioxid Redox
Signal. 20:1891–1901. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Bao B, Azmi AS, Ali S, Ahmad A, Li Y,
Banerjee S, Kong D and Sarkar FH: The biological kinship of hypoxia
with CSC and EMT and their relationship with deregulated expression
of miRNAs and tumor aggressiveness. Biochim Biophys Acta.
1826:272–296. 2012.PubMed/NCBI
|
|
107
|
Lai FB, Liu WT, Jing YY, Yu GF, Han ZP,
Yang X, Zeng JX, Zhang HJ, Shi RY, Li XY, et al: Lipopolysaccharide
supports maintaining the stemness of CD133(+) hepatoma cells
through activation of the NF-κB/HIF-1α pathway. Cancer Lett.
378:131–141. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Jing L, Ruan Z, Sun H, Li Q, Han L, Huang
L, Yu S, Wang Y, Guo H and Jiao M: Epithelial-mesenchymal
transition induced cancer-stem-cell-like characteristics in
hepatocellular carcinoma. J Cell Physiol. 234:18448–18458.
2019.PubMed/NCBI
|
|
109
|
Cao Q, Lu W, Zhou T, Liu Y, Cai X, Zhu J
and Cao P: Analgesic-antitumor peptide inhibits angiogenesis by
suppressing AKT activation in hepatocellular carcinoma. Mol Cell
Biochem. 455:119–125. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Tan Y, Du B, Zhan Y, Wang K, Wang X, Chen
B, Wei X and Xiao J: Antitumor effects of circ-EPHB4 in
hepatocellular carcinoma via inhibition of HIF-1α. Mol Carcinog.
58:875–886. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Qin Y, Liu HJ, Li M, Zhai DH, Tang YH,
Yang L, Qiao KL, Yang JH, Zhong WL, Zhang Q, et al: Salidroside
improves the hypoxic tumor microenvironment and reverses the drug
resistance of platinum drugs via HIF-1α signaling pathway.
EBioMedicine. 38:25–36. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Lei HW, Cai J, Li CM, Yang F, Shi WQ, Shi
WQ, Wang LP and Feng YY: Rapamycin combi with TAE on the growth,
metastasis, and prognosis of hepatocellular carcinoma in rat
models. Ann Hepatol. 17:645–654. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Hua H, Zhu Y and Song YH: Ruscogenin
suppressed the hepatocellular carcinoma metastasis via
PI3K/AKT/mTOR signaling pathway. Biomed Pharmacother. 101:115–122.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Liu P, Atkinson SJ, Akbareian SE, Zhou Z,
Munsterberg A, Robinson SD and Bao Y: Sulforaphane exerts
anti-angiogenesis effects against hepatocellular carcinoma through
inhibition of STAT3/HIF-1α/VEGF signalling. Sci Rep. 7:126512017.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Zhou QY, Tu CY, Shao CX, Wang WK, Zhu JD,
Cai Y, Mao JY and Chen W: GC7 blocks epithelial-mesenchymal
transition and reverses hypoxia-induced chemotherapy resistance in
hepatocellular carcinoma cells. Am J Transl Res. 9:2608–2617.
2017.PubMed/NCBI
|
|
116
|
Chow AK, Yau TC, Ng L, Chu AC, Law WL,
Poon RT and Pang RW: A preclinical study on the combination therapy
of everolimus and transarterial chemoembolization in hepatocellular
carcinoma. Am J Cancer Res. 5:2376–2386. 2015.PubMed/NCBI
|
|
117
|
Li C, Wu X, Zhang H, Yang G, Hao M, Sheng
S, Sun Y, Long J, Hu C, Sun X, et al: A Huaier polysaccharide
restrains hepatocellular carcinoma growth and metastasis by
suppression angiogenesis. Int J Biol Macromol. 75:115–120. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Wu J, Contratto M, Shanbhogue KP, Manji
GA, O'Neil BH, Noonan A, Tudor R and Lee R: Evaluation of a locked
nucleic acid form of antisense oligo targeting HIF-1α in advanced
hepatocellular carcinoma. World J Clin Oncol. 10:149–160. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Rapisarda A, Uranchimeg B, Sordet O,
Pommier Y, Shoemaker RH and Melillo G: Topoisomerase I-mediated
inhibition of hypoxia-inducible factor 1: Mechanism and therapeutic
implications. Cancer Res. 64:1475–1482. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Zhang C, Yang C, Feldman MJ, Wang H, Pang
Y, Maggio DM, Zhu D, Nesvick CL, Dmitriev P, Bullova P, et al:
Vorinostat suppresses hypoxia signaling by modulating nuclear
translocation of hypoxia inducible factor 1 alpha. Oncotarget.
8:56110–56125. 2017.PubMed/NCBI
|
|
121
|
Li YL, Zhang NY, Hu X, Chen JL, Rao MJ, Wu
LW, Li QY, Zhang B, Yan W and Zhang C: Evodiamine induces apoptosis
and promotes hepatocellular carcinoma cell death induced by
vorinostat via downregulating HIF-1α under hypoxia. Biochem Biophys
Res Commun. 498:481–486. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Shao S, Duan W, Xu Q, Li X, Han L, Li W,
Zhang D, Wang Z and Lei J: Curcumin suppresses hepatic stellate
cell-induced hepatocarcinoma angiogenesis and invasion through
downregulating CTGF. Oxid Med Cell Longev. 2019:81485102019.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Xia Y, Choi HK and Lee K: Recent advances
in hypoxia-inducible factor (HIF)-1 inhibitors. Eur J Med Chem.
49:24–40. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Abu-Remaileh M, Khalaileh A, Pikarsky E
and Aqeilan RI: Author Correction: WWOX controls hepatic HIF1α to
suppress hepatocyte proliferation and neoplasia. Cell Death Dis.
9:11592018. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Yang Q, Guo X and Yang L: Metformin
enhances the effect of regorafenib and inhibits recurrence and
metastasis of hepatic carcinoma after liver resection via
regulating expression of hypoxia inducible factors 2α (HIF-2α) and
30 kDa HIV tat-interacting protein (TIP30). Med Sci Monit.
24:2225–2234. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Wada H, Nagano H, Yamamoto H, Yang Y,
Kondo M, Ota H, Nakamura M, Yoshioka S, Kato H, Damdinsuren B, et
al: Expression pattern of angiogenic factors and prognosis after
hepatic resection in hepatocellular carcinoma: Importance of
angiopoietin-2 and hypoxia-induced factor-1 alpha. Liver Int.
26:414–423. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Dai CX, Gao Q, Qiu SJ, Ju MJ, Cai MY, Xu
YF, Zhou J, Zhang BH and Fan J: Hypoxia-inducible factor-1 alpha,
in association with inflammation, angiogenesis and MYC, is a
critical prognostic factor in patients with HCC after surgery. BMC
Cancer. 9:4182009. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Xia L, Mo P, Huang W, Zhang L, Wang Y, Zhu
H, Tian D, Liu J, Chen Z, Zhang Y, et al: The
TNF-α/ROS/HIF-1-induced upregulation of FoxMI expression promotes
HCC proliferation and resistance to apoptosis. Carcinogenesis.
33:2250–2259. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Xiang ZL, Zeng ZC, Fan J, Tang ZY, He J,
Zeng HY and Chang JY: The expression of HIF-1α in primary
hepatocellular carcinoma and its correlation with radiotherapy
response and clinical outcome. Mol Biol Rep. 39:2021–2029. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Zheng SS, Chen XH, Yin X and Zhang BH:
Prognostic significance of HIF-1α expression in hepatocellular
carcinoma: A meta-analysis. PLoS One. 8:e657532013. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Wang B, Ding YM, Fan P, Wang B, Xu JH and
Wang WX: Expression and significance of MMP2 and HIF-1α in
hepatocellular carcinoma. Oncol Lett. 8:539–546. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Cao S, Yang S, Wu C, Wang Y, Jiang J and
Lu Z: Protein expression of hypoxia-inducible factor-1 alpha and
hepatocellular carcinoma: A systematic review with meta-analysis.
Clin Res Hepatol Gastroenterol. 38:598–603. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Liu LP, Hu BG, Ye C, Ho RL, Chen GG and
Lai PB: HBx mutants differentially affect the activation of
hypoxia-inducible factor-1α in hepatocellular carcinoma. Br J
Cancer. 110:1066–1073. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Wang D, Zhang X, Lu Y, Wang X and Zhu L:
Hypoxia inducible factor 1α in hepatocellular carcinoma with
cirrhosis: Association with prognosis. Pathol Res Pract.
214:1987–1992. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Bangoura G, Liu Z, Qian Q, Jiang C, Yang G
and Jing S: Prognostic significance of HIF-2alpha/EPAS1 expression
in hepatocellular carcinoma. World J Gastroenterol. 13:3176–3182.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Sun HX, Xu Y, Yang XR, Wang WM, Bai H, Shi
RY, Nayar SK, Devbhandari RP, He YZ, Zhu QF, et al: Hypoxia
inducible factor 2 alpha inhibits hepatocellular carcinoma growth
through the transcription factor dimerization partner 3/E2F
transcription factor 1-dependent apoptotic pathway. Hepatology.
57:1088–1097. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Yao Q, Lv Y, Pan T, Liu Y, Ma J and Xu G:
Prognostic significance and clinicopathological features of hypoxic
inducible factor-2alpha expression in hepatocellular carcinoma.
Saudi Med J. 36:170–175. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Yang SL, Liu LP, Niu L, Sun YF, Yang XR,
Fan J, Ren JW, Chen GG and Lai PB: Downregulation and pro-apoptotic
effect of hypoxia-inducible factor 2 alpha in hepatocellular
carcinoma. Oncotarget. 7:34571–34581. 2016.PubMed/NCBI
|
|
139
|
Jiang L, Liu QL, Liang QL, Zhang HJ, Ou WT
and Yuan GL: Association of PHD3 and HIF2α gene expression with
clinicopathological characteristics in human hepatocellular
carcinoma. Oncol Lett. 15:545–551. 2018.PubMed/NCBI
|
|
140
|
Liu P, Fang X, Song Y, Jiang JX, He QJ and
Liu XJ: Expression of hypoxia-inducible factor 3α in hepatocellular
carcinoma and its association with other hypoxia-inducible factors.
Exp Ther Med. 11:2470–2476. 2016. View Article : Google Scholar : PubMed/NCBI
|