Introduction
Cervical cancer ranks as the 4th most common female
malignancy and the 2nd most common cancer in developing countries,
which affects women globally (1–3). Surgery
or a combination of chemotherapy and radiotherapy are the routine
treatments for cervical cancer patients (4–7). However,
the malignancy is largely incurable for the patient. The early
stage of cervical cancer is predisposed to develop progressively
into an advanced stage. Therefore, early detection and prognosis
are crucial. It is critical to understand the molecular mechanisms
of cervical cancer development and identify novel biomarkers for
the early detection and treatment of cervical cancer.
Long non-coding RNAs (lncRNAs) emerged as important
molecules involved in normal development and in tumorigenesis
(8). Growing evidence has revealed
that lncRNAs are involved in the aberrant pathological development
of cervical cancer (9). For example,
reduced lncRNA MEG3 (maternally expressed 3) is associated with
increased cervical cancer cell proliferation (10). LncRNA MALAT1 (metastasis-associated
lung adenocarcinoma transcript 1) markedly increased after HPV
infection in cervical cancer (11).
FTH1P3 belongs to long non-coding RNAs, and is a member of the
ferritin heavy chain (FHC) gene family located at chromosome 2p23.3
(12). FTH1P3 is widely expressed in
different human cell lines and tissues and positively modulated
during cell differentiation (12).
Zhang et al revealed that FTH1P3 is a promoting factor in
the growth and progression of OSCC by stimulating cell
proliferation, migration, and invasion (13). However, the role of FTH1P3 in cervical
cancer has not yet been elaborated.
MicroRNAs (miRNAs or miRs), ~18–25 nucleotides, are
another component of the noncoding RNA family. They participate and
regulate gene expression through a post-transcriptional pattern
(14–16). miRNAs are aberrantly expressed in
various types of malignancies and function either as oncogenes or
as tumor suppressors (17–19). Accumulating evidence has demonstrated
that miRNAs regulated various carcinogenesis processes, including
cell maturation, cell proliferation, migration, invasion,
autophagy, apoptosis, and metastasis (20–26).
Therefore, miRNAs have a large potential to serve as promising
markers in the diagnosis, prognosis, and personalized targeted
therapies (22–26). miR-145 plays a tumor-suppressive role
in several types of cancer, such as gastric (27), hepatic (28), breast (29), non-small cell lung (30), and cervical cancer (31). Zhou et al reported that
miRNA-145 inhibited tumorigenesis and invasion of cervical cancer
stem cells by inducing cancer stem cell (CSC) differentiation
through downregulation of the stem cell transcription factors that
maintain CSC pluripotency (32).
Sathyanarayanan et al stated that miRNA-145 modulated
epithelial-mesenchymal transition (EMT) and suppressed
proliferation, migration, and invasion by targeting SIP1 in human
cervical cancer cells (33).
However, the reason that miR-145 was downregulated
in cervical cancer remains obscure. In our previous bioinformatics
analysis (unpublished data), it was revealed that FTH1P3 was
possibly regulating miR-145 by anti-sense sequence matching. Since
miR-145 has a wide range of functions in a variety of tumors, the
study aimed to determine whether FTH1P3 can target miR-145 in
cervical cancer. In the present study, the functions and
associations between FTH1P3 and miR-145 in cervical cells and
tissues were demonstrated. The present findings may further provide
a therapeutic target for the treatment and diagnosis of cervical
cancer patients.
Materials and methods
Patients and materials
Fifty-two cervical cancer tissues (all females,
44–69 years, 55.7±5.5 years) were collected from patients who
underwent surgery at our hospital from March 2015 to March 2017.
The samples of patients with other major diseases were excluded.
Normal cervix healthy tissues were used as a normal control. The
dissected patient surgical specimen was immediately transferred to
the surgical laboratory. Written consent was signed from each
patient. The implemented protocol was approved by the Human Ethics
Committee of Gansu Provincial Cancer Hospital. All the tissues were
either fixed in 4% PFA (paraformaldehyde) or were snap-frozen in
liquid nitrogen for later use. Cervical cancer patients (52) were
divided into high- and low-expression groups according to the
median values of FTH1P3 (fold change=4.32) and miR-145 (fold
change=0.41) expression.
Cell culture
Human cervical cancer cell lines (SiHa, HeLa, CaSki,
and C4-1) and normal cervical epithelial cells (Ect1/E6E7) were
purchased from Cell Bank of the Chinese Academy of Sciences
(Shanghai, China). The cells were cultured in DMEM (Dulbecco's
Modified Eagle's Medium) supplemented with 10% heated inactivated
fetal bovine serum (Hi-FBS) and 100 U/ml penicillin-streptomycin
(10,000 U/ml; all from Thermo Fisher Scientific, Inc.). Cells were
maintained at 37°C, and 5% CO2 in a humidified
incubator.
Bioinformatics analysis
The binding candidates of FTH1P3 were predicted
using miRcode software (http://www.mircode.org/). All parameters were
default.
Quantitative real-time reverse
transcription-PCR (qRT-PCR)
Total RNA was extracted using TRIzol™ reagent
(Thermo Fisher Scientific, Inc.). RNA concentrations were measured
using an ND-1000 UV–Vis Spectrophotometer (Thermo Fisher
Scientific, Inc.), and the quality was monitored by an Agilent 2100
Bioanalyzer (Agilent Technologies, Inc.). qRT-PCR was carried out
using a TaqMan miRNA Assay according to the manufacturer's protocol
(Applied Biosystems; Thermo Fisher Scientific, Inc.). The
amplification conditions were: 40 cycles of 15 sec at 95°C and 1
min at 60°C. The expression levels of miR-145 and FTH1P3 were
normalized by U6.
Cell transfection
The hsa-miRNA-145 mimic/miRNA-145 scramble
(negative-control for miRNA-145 mimic) and hsa-miRNA-145
inhibitor/miRNA-145 scramble (negative-control for miRNA-145
inhibitor) were designed and synthesized from Shanghai GeneChem
Co., Ltd. The FTH1P3 siRNAs were synthesized by Invitrogen; Thermo
Fisher Scientific, Inc. FTH1P3 siRNA (targeted region) sequences
were: siRNA_1, CGCCUGUAAUCCCAGCUCUCA; siRNA_2,
AUAAGCGUAACUUCCCUCAAA; siRNA_3, CGUAACUUCCCUCAAAGCAACAACC.
pcDNA-3.1(+)-FTH1P3 (lncRNA-FTH1P3). The overexpression plasmid of
FTH1P3 was purchased from Shanghai GeneChem Co., Ltd. A
transfection assay was conducted using a Lipofectamine 3000 kit
(Thermo Fisher Scientific, Inc.). The sequence of miRNA-145 mimic,
miRNA-145 inhibitor or respective control were as follows:
miRNA-145 mimic: 5′-UCCCUAAGGACCCUUUUGACCUG-3′ (sense)
3′-AGGGAUUCCUGGGAAAACUGGAC-5′ (antisense); miRNA-145 scramble
(negative-control for miRNA-145 mimic):
5′-CUAUCCACCAGGUUGCUUUGACC-3′ (sense) 3′-GAUAGGUGGUCCAACGAAACUGG-5′
(antisense); miRNA-145 inhibitor: 5′-CAGGUCAAAAGGGUCCUUAGGGA-3′;
miRNA-145 scramble (negative-control for miRNA-145 inhibitor):
5′-GUCCAGAGGAAACUUGUCGAAGG-3′.
CCK-8 analysis
Cell Counting Kit-8 (CCK-8) was used to determine
the cell viability in cell proliferation and cytotoxicity. Briefly,
human cervical cancer cells were seeded in a 96-well plate
(5×103 cells/well) overnight. After cell confluence
reached 70–80%, the miR-145 mimic or FTH1P3 siRNA was transfected
into the cells using Lipofectamine 3000 kit (Thermo Fisher
Scientific, Inc.). After 2 days, the CCK-8 reagent (5 mg/ml) was
directly loaded into each well and incubated in the dark for 2 h at
37°C followed by measurement of the absorbance at OD 490 nm. Each
measurement was repeated in triplicate.
Flow cytometric analysis
After transfection with the desired plasmid or
negative control in 6-well plates, the cells were cultured for 48
h. Annexin V-FITC (3 µg/ml) and propidium iodide (5 µg/ml) were
incubated with cells for 30 min at RT shielded from the light. Then
the cells were collected, filtered and subjected to a BD FACSAria
Fusion Cell Sorter (BD Biosciences) with Cytomics FC 500 with CXP
Software (Beckman Coulter, Inc.). The data were quantified with
FlowJo software v10.6.1 (FlowJo LLC).
Invasion and migration assays
For the invasion assay, 2×104 HeLa cells
in FBS-free DMEM were placed in the upper chamber of an insert
(8-µm pores; Merck KGaA). Matrigel (Merck KGaA) was employed to
pre-coat the membrane of the Transwell chambers. The lower chambers
were incubated in culture medium supplemented with 10% FBS for 24
h. Then, the HeLa cells on the upper surface were scraped and
washed away. Subsequently, the cells on the lower surface were
fixed and stained with Diff-Quik staining kit (Sysmex Corp.) at RT
for 2 h. The invaded cells in the lower surface of membrane were
also fixed and stained with Diff-Quik kit (Sysmex Corp.). The
invaded cells were imaged and counted under a BX43 Upright
Microscope (Olympus Corp.) at a magnification of ×200 from 10
random fields in each well. For the migration assay,
1.5×106 cells/well were seeded in 6-well plate for
overnight culture until the cells reached an ~90% confluence. The
scratch was generated using a 200-µl sterile pipet tip in the hood.
After aspiration and washing, fresh complete medium was added. Then
the cells were cultured for another 24 h. Cell migration was
monitored under a BX43 Upright Microscope (Olympus Corporation) and
images were captured at 0 and 24 h.
Luciferase reporter assays
The fragments of the 3′-UTR of FTH1P3 were amplified
with Phusion® High-Fidelity DNA Polymerase (NEB, Inc.)
from human genomic DNA. After gel extraction, the PCR product was
cloned into the multiple cloning sites located downstream of the
firefly luciferase coding region of the pGL3 Luciferase Reporter
Vectors (Promega, USA). The recombinant vector was named
pMIR-FTH1P3-wild-type. The mutations in the miR-145 binding sites
were introduced using Site-Directed Mutagenesis (Thermo Fisher
Scientific, Inc.). The mutation sequence was confirmed by
sequencing (Shengong Bio Company). The resulting vector was
designated as pMIR-BCYRN-mutant. pMIR-FTH1P3 or pMIR-FTH1P3-mut
vector and miR-145 mimic or miR-145 inhibitor were co-transfected
into a 24-well plate according to the Lipofectamine 3000 kit
(Thermo Fisher Scientific, Inc.). After 36 h, the cells were
harvested and lysed (lysis buffer; Promega Corporation). The
luciferase reporter gene assay was detected by the
GloMax® 20/20 Luminometer (Promega Corporation)
according to the instructions.
Statistical analysis
Statistical analysis was performed using SPSS 13.0
(Pearson). All data were expressed as the mean ± standard deviation
(SD). Correlation between FTH1P3 and miR-145 expression was
determined using Spearman's correlation analysis. Differences were
assessed by two-sample t-test or chi-square test or one-way ANOVA
and Fisher's LSD post hoc tests. P<0.05 was considered to
indicate a statistically significant difference.
Results
FTH1P3 expression is upregulated in
cervical cancer tissues and cell lines
To investigate the expression of lncRNA FTH1P3 in
cervical cancer tissues, total RNA was extracted from 52 cervical
cancer tissues and paired adjacent non-cancerous normal tissues.
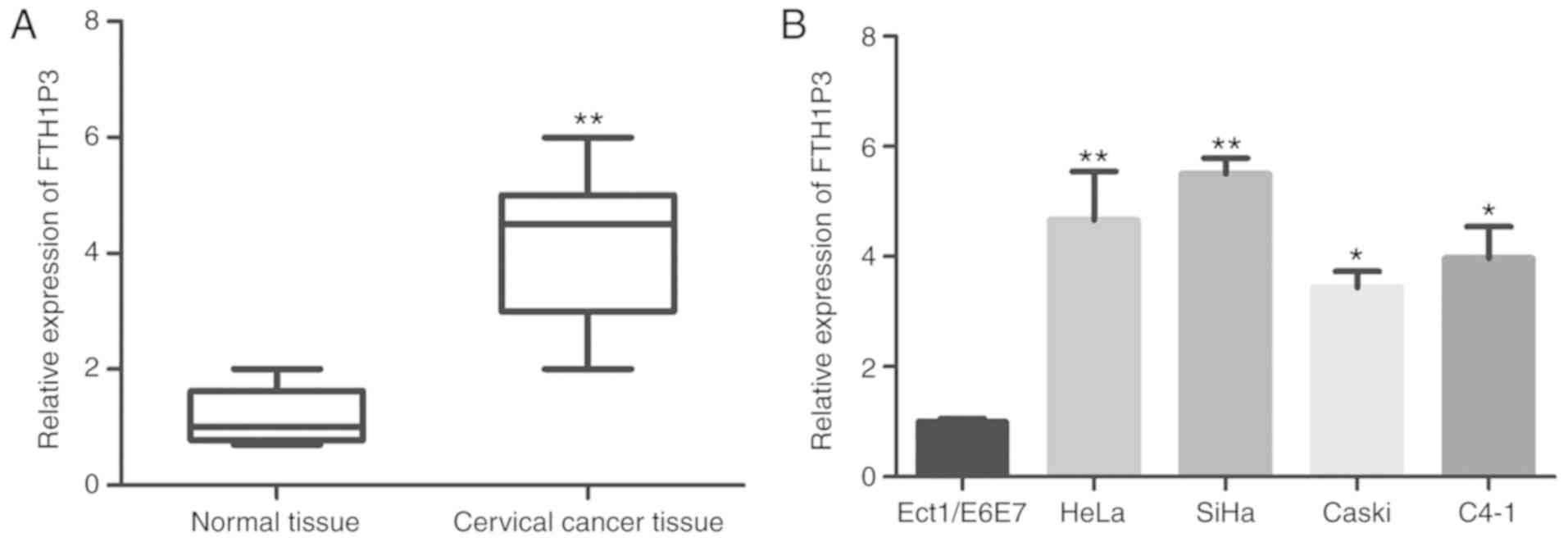
qRT-PCR was carried out to detect FTH1P3 mRNA expression level. As
revealed in Fig. 1A, FTH1P3
expression was significantly upregulated in cervical cancer tissues
compared to paired adjacent non-cancerous normal tissues
(P<0.01). Furthermore, the correlation between FTH1P3 expression
and pathological parameters of cervical cancer patients were
evaluated. The results of Table I
revealed that FTH1P3 upregulation in cervical cancer tissues was
significantly correlated with crucial clinicopathological factors,
including tumor size (P=0.014), lymph node metastasis (P=0.005) and
FIGO stage (P=0.002). However, there was no significant difference
between FTH1P3 expression and other clinicopathological factors,
such as age, menopause, depth of invasion and CEA level. Next, the
expression of FTH1P3 in cervical cancer cell lines was further
investigated. As revealed in Fig. 1B,
it was determined that FTH1P3 expression was significantly
increased in cervical cancer cell lines (HeLa, SiHa, Caski, and
C4-1) compared to cervical normal epithelial cells (ECT1/E6E7).
Notably, higher expression of FTH1P3 was observed in the HeLa and
SiHa cell lines. Therefore, HeLa and SiHa cells were selected for
further studies.
 | Table I.Correlation analysis between FTH1P3
expression and clinicopathological index in cervical cancer
patients. |
Table I.
Correlation analysis between FTH1P3
expression and clinicopathological index in cervical cancer
patients.
|
|
| FTH1P3 |
| miR-145 |
|
|---|
|
|
|
|
|
|
|
|---|
| Parameters | n | Low (n) | High (n) | P-value | Low (n) | High (n) | P-value |
|---|
| Age (years) |
|
|
|
|
|
|
|
|
<50 | 24 | 10 | 14 | 0.101 | 11 | 13 | 0.525 |
|
≥50 | 28 | 16 | 12 |
| 15 | 13 |
|
| Menopause |
|
|
|
|
|
|
|
|
Yes | 22 | 10 | 12 | 0.438 | 12 | 10 | 0.438 |
| No | 30 | 16 | 14 |
| 14 | 16 |
|
| Tumor size
(cm) |
|
|
|
|
|
|
|
|
<4 | 18 | 12 | 6 | 0.014a | 7 | 11 | 0.035a |
| ≥4 | 34 | 14 | 20 |
| 19 | 15 |
|
| Depth of
invasion |
|
|
|
|
|
|
|
|
<2/3 | 21 | 12 | 9 | 0.085 | 11 | 10 | 0.621 |
|
≥2/3 | 31 | 14 | 17 |
| 15 | 16 |
|
| LNM stage |
|
|
|
|
|
|
|
|
Negative | 35 | 21 | 14 | 0.005b | 15 | 20 | 0.011a |
|
Positive | 17 | 5 | 12 |
| 11 | 6 |
|
| FIGO stage |
|
|
|
|
|
|
|
|
I–II | 27 | 20 | 7 | 0.002b | 17 | 10 | 0.017a |
|
III–IV | 25 | 6 | 19 |
| 9 | 16 |
|
| CEA |
|
|
|
|
|
|
|
|
Negative | 33 | 18 | 15 | 0.083 | 16 | 17 | 0.662 |
|
Positive | 19 | 8 | 11 |
| 10 | 9 |
|
FTH1P3 silencing inhibits cell
viability and motility in cervical cancer cell lines
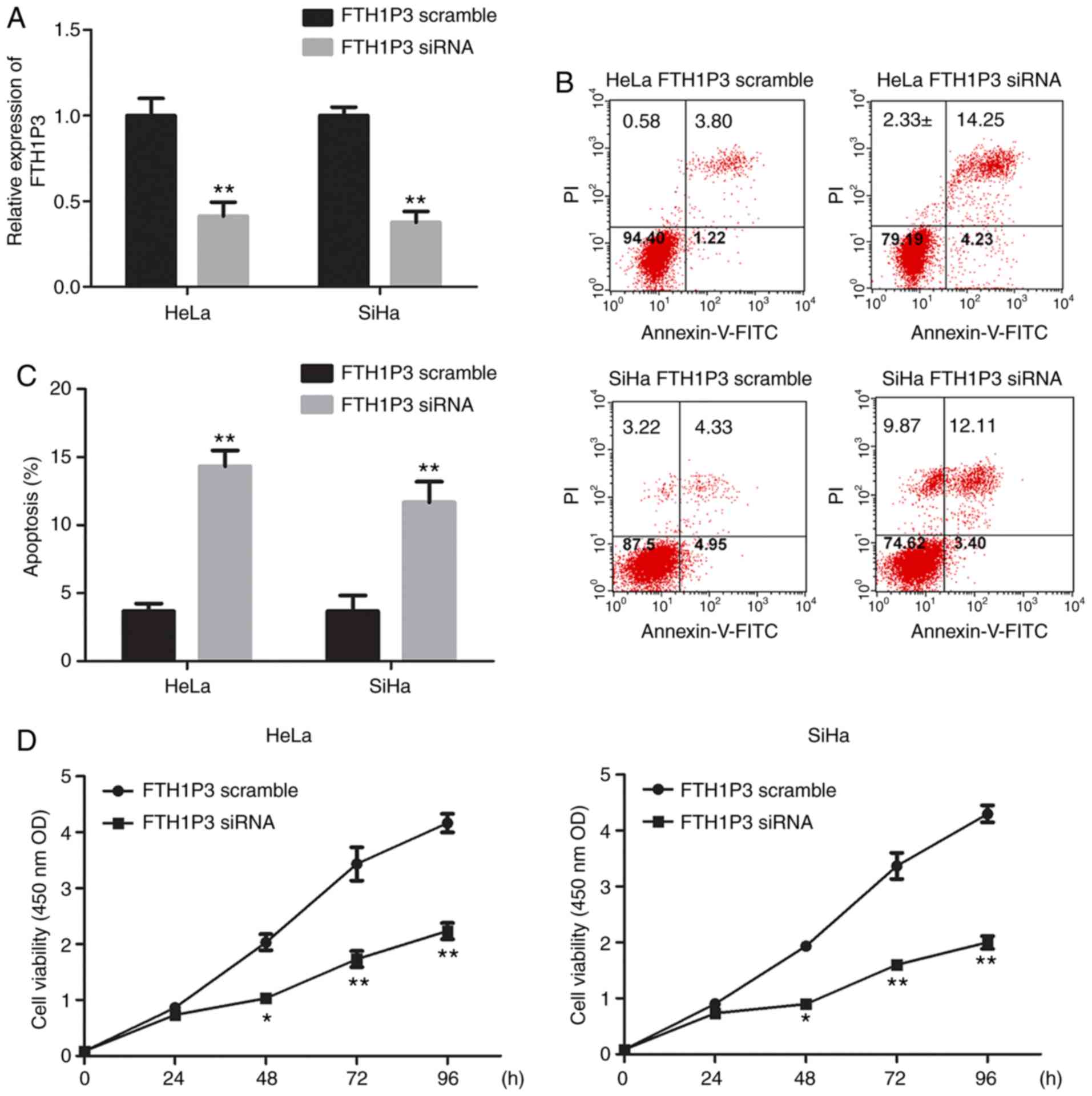
To determine the roles of FTH1P3 in cervical cancer,
siRNA of FTH1P3 was constructed and transfected into HeLa and SiHa
cells. As revealed in Fig. 2A, siRNA
of FTH1P3 significantly suppressed the expression of FTH1P3 in both
HeLa and SiHa cells (P<0.01). Furthermore, downregulation of
FTH1P3 by siRNA induced significant apoptosis in HeLa and SiHa
cells (Fig. 2B and C; P<0.01).
CCK-8 assay also revealed that FTH1P3 downregulation significantly
inhibited the proliferation of HeLa and SiHa cells (Fig. 2D; P<0.01). In addition, the effects
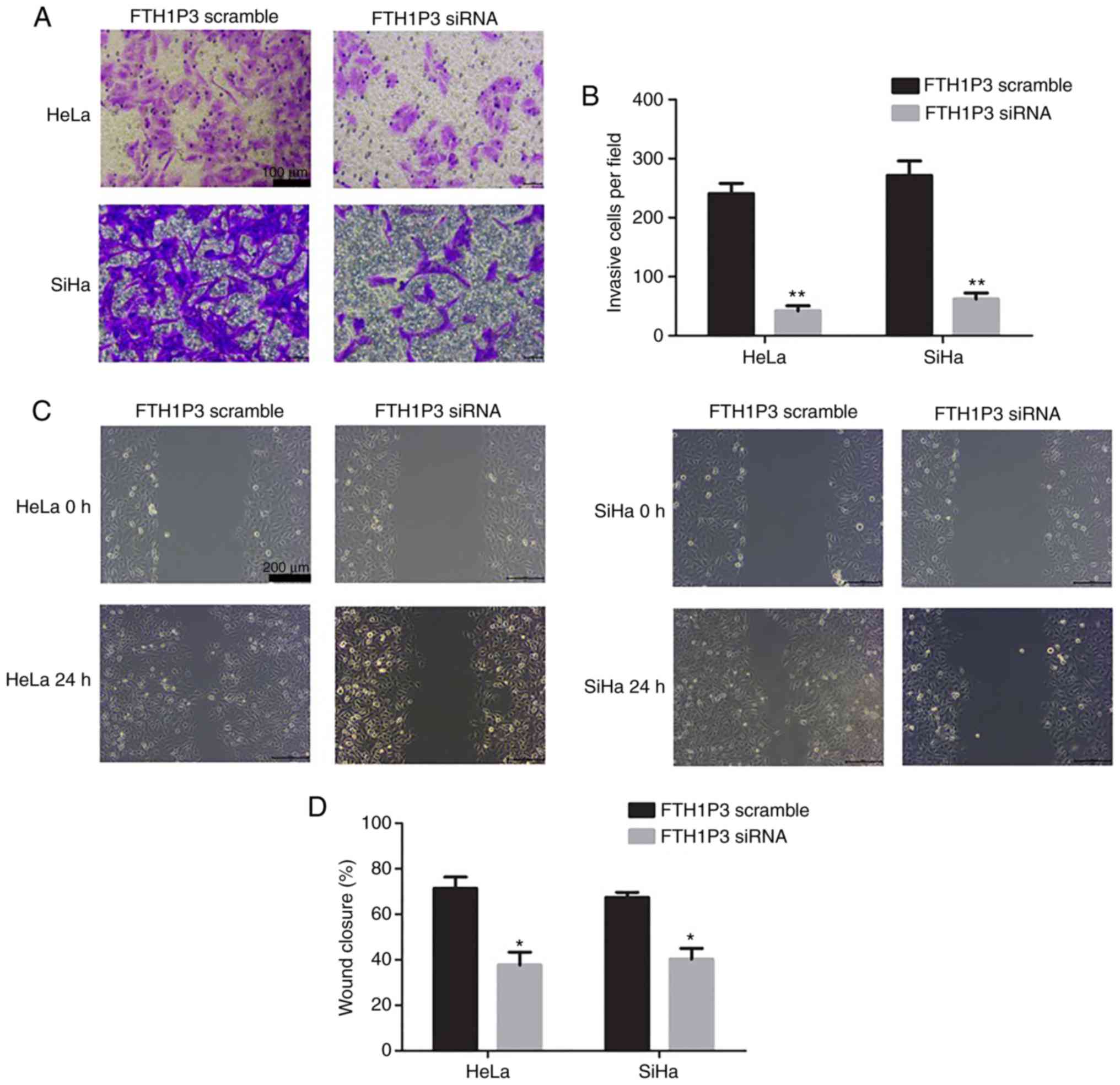
of FTH1P3 on cell motility were examined by Transwell and
wound-healing assays. The results revealed that the number of
invasive cells was significantly reduced after transfection with
FTH1P3 siRNA (P<0.01; Fig. 3A and
B). Similarly, a significantly decreased closure rate of
scratch wounds was observed in the FTH1P3 siRNA group compared with
the siRNA scramble group in HeLa and SiHa cells (P<0.05;
Fig. 3C and D).
FTH1P3 is identified as a potential
target of miR-145
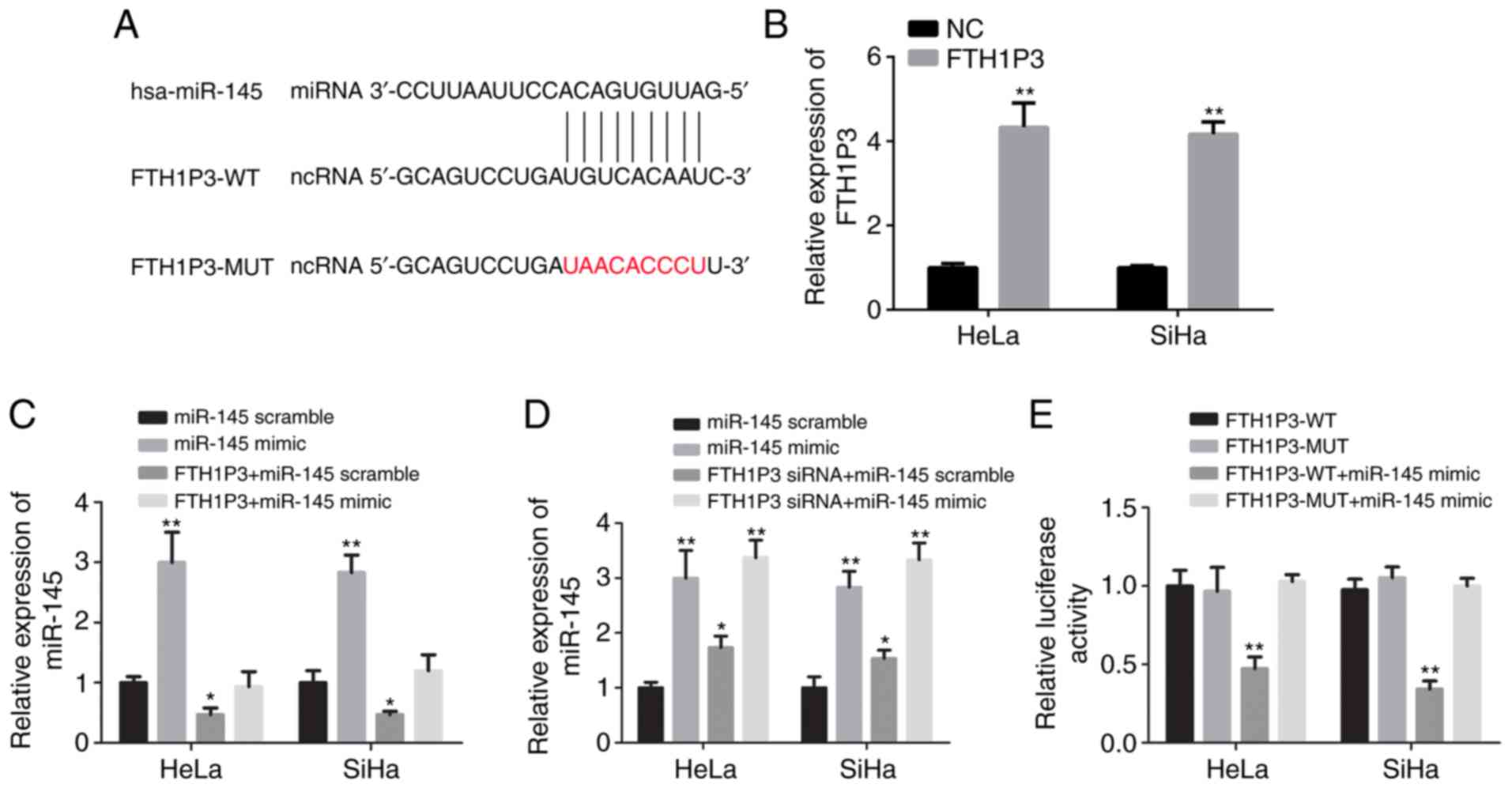
Bioinformatics analysis with miRcode software was
performed and it was revealed that human miR-145 (hsa-miR-145) was
a potential binding candidate of FTH1P3 (Fig. 4A). As revealed in Fig. 4B, FTH1P3 overexpression plasmid
significantly increased the expression of FTH1P3 in both HeLa and
SiHa cells compared with the negative control (P<0.01). After
transfection of human miR-145 mimic, a significant increase of
miR-145 expression in HeLa and SiHa cells was observed (Fig. 4C; P<0.01). The overexpression of
FTH1P3 by plasmid transfection could significantly reduce the
expression of miR-145, while miR-145 expression was inversely
increased due to the downregulation of FTH1P3 (Fig. 4C and D; P<0.05). In addition, a
luciferase reporter assay revealed that miR-145 mimic significantly
suppressed the luciferase activities of FTH1P3-WT reporter vector.
Conversely, after transfection with FTH1P3-MUT and the miRNA-145
mimic, the luciferase activities between these two cells were
nearly comparable with that in the control cells (Fig. 4E; P<0.01). These data indicated
that FTH1P3 was a potential target of miR-145.
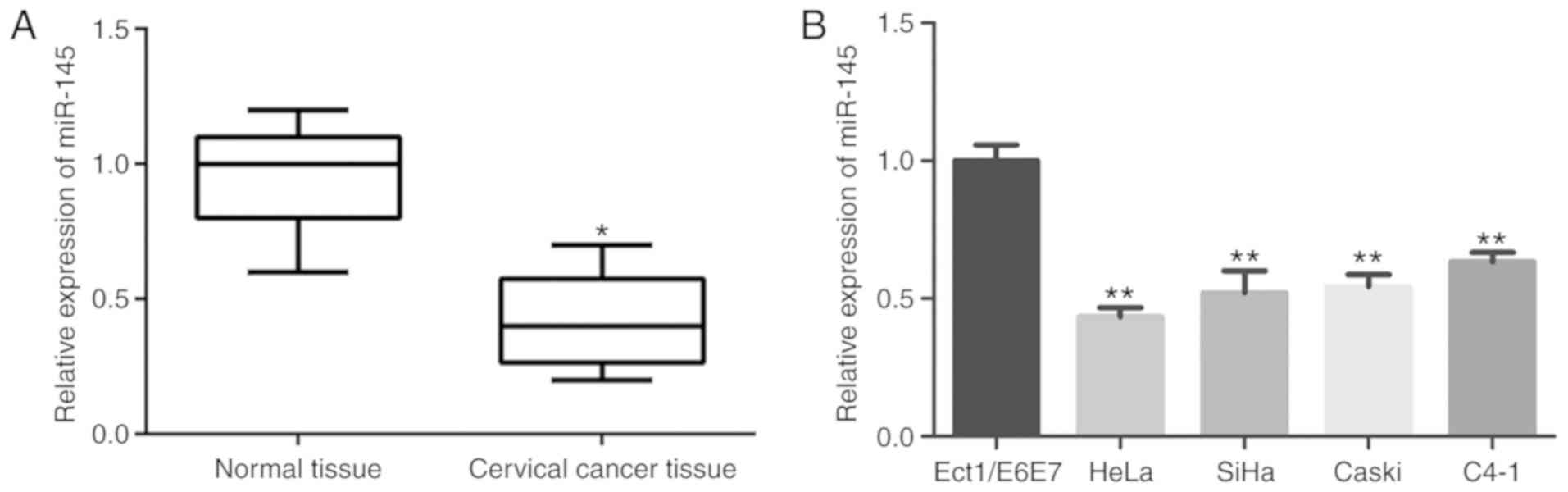
miRNA-145 is downregulated in cervical
cancer
To investigate the role of miR-145 in cervical
cancer progression, miR-145 expression in cervical cancer tissues
and non-malignant tissues was detected by qRT-PCR. As revealed in
Fig. 5A, miR-145 expression in
cervical cancer tissues was significantly downregulated compared
with the non-malignant tissues (P<0.01). Consistently, the
expression of miR-145 in five assessed cervical cancer cell lines
were all significantly lower than that in normal cervical
epithelial cells (Ect1/E6E7) (P<0.01; Fig. 5B). Moreover, the downregulated
expression of miR-145 demonstrated a significant association with
tumor size (P=0.035), lymph node metastasis (P=0.011) and FIGO
stage (P=0.017). Spearman's correlation analysis revealed that
miR-145 expression was inversely associated with FTH1P3 expression
in cervical cancer tissues (Table
II, r=−0.265, P=0.002), indicating that upregulated FTH1P3
expression in cervical cancer was correlated with downregulated
miR-145 expression.
| Table II.Correlation between FTH1P3 expression
and miR-145 expression in cervical cancer patients. |
Table II.
Correlation between FTH1P3 expression
and miR-145 expression in cervical cancer patients.
|
|
| miR-145
Expression |
|
|
|
|---|
|
|
|
|
|
|
|
|---|
| FTH1P3
expression | n | Low (n, %) | High (n, %) | rs | χ2 | P-value |
|---|
| Low (n, %) | 26 | 8 | 18 | −0.265 | 9.102 | 0.002 |
| High (n, %) | 26 | 18 | 8 |
|
|
|
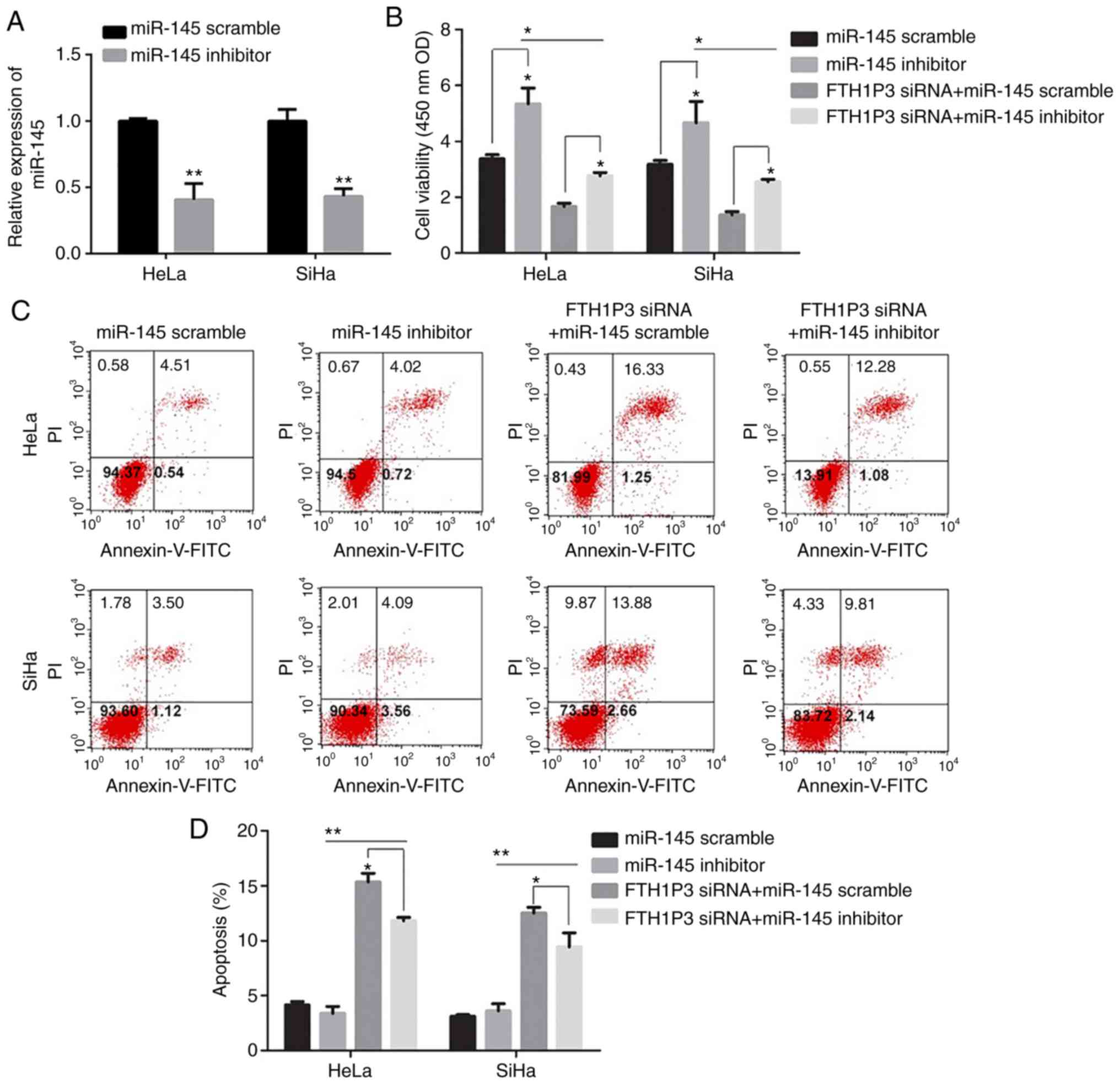
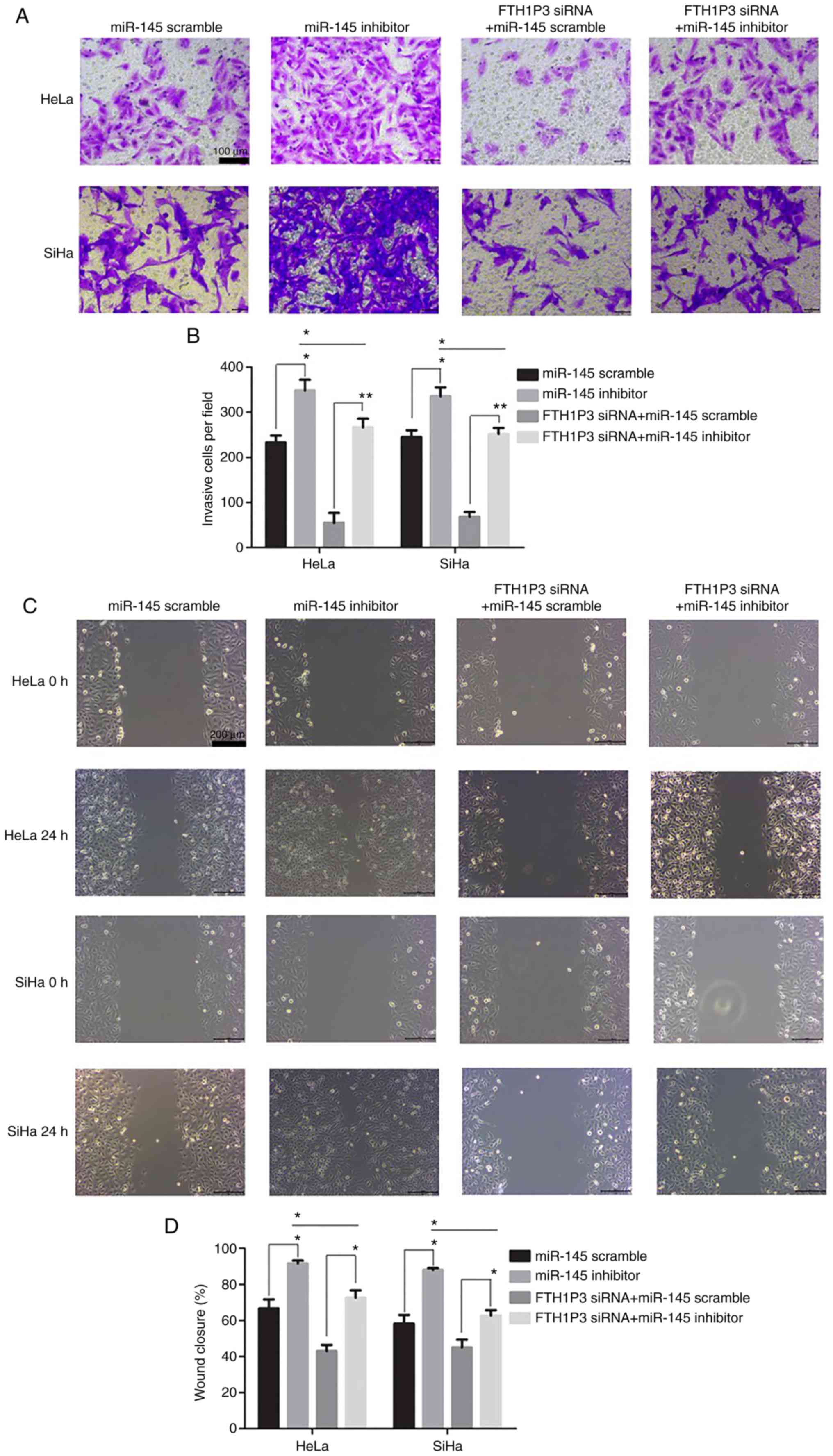
Effects of miR-145 inhibitor on the
cell proliferation, invasion, and migration in cervical cancer
cells
As illustrated in Fig.
6A, the transfection of miR-145 inhibitor caused a significant
reduction of miR-145 in cervical cancer cells (P<0.01).
Moreover, downregulation of FTH1P3 significantly inhibited the
proliferation, invasion and migration abilities and promoted
apoptosis in HeLa and SiHa cells (Figs.
6 and 7). Notably, the miRNA-145
inhibitor could enhance the cell proliferation, invasion and
migration in cervical cancer cells compared to the RNA scramble
group, while FTH1P3 siRNA revealed a significant bucking effect for
miRNA-145 inhibitor in HeLa and SiHa cells compared to cells
transfected with FTH1P3 siRNA+ RNA scramble group (Figs. 6 and 7).
Discussion
The present study revealed that FTH1P3 was highly
expressed in malignant cervical cancer tissues and cell lines.
Downregulation of FTH1P3 significantly inhibited cell
proliferation, invasion and migration, and promoted apoptosis in
cervical cancer cells. The FTH1P3 and miR-145 axis was
demonstrated, to have reciprocal repression functionally. These
results demonstrated that lncRNA FTH1P3 functioned as a promoting
factor in cervical cancer by targeting miR-145.
FTH1P3 expression was revealed to be enhanced in
uveal melanoma cell lines and tissues, and FTH1P3 upregulation
promoted cell proliferation, the cell cycle and migration in uveal
melanoma by targeting miR-224-5p (34). FTH1P3 was abundantly expressed in
paclitaxel-resistant breast cancer tissue and cells (35). FTH1P3 was revealed to play a role as a
competing endogenous RNA (ceRNA) to sponge miR-206 and increase
ABCB1 (ATP-binding cassette subfamily B member 1) protein
expression (35). FTH1P3 was
overexpressed in oral squamous cell carcinoma (OSCC) tissues
(13). Overexpression of of FTH1P3
significantly promoted OSCC cell growth, while the cell growth was
inhibited after knockout of FTH1P3 (13). In the present study, it was also
revealed that FTH1P3 was highly expressed in malignant cervical
cancer tissues and human cervical cancer cell lines (SiHa, HeLa,
CaSki and C4-1) (Fig. 1).
Collectively, these studies indicated that FTH1P3 may function as
an oncogene and be favorable to tumorigenesis and progression of
various cancers.
Emerging studies have indicated that miR-145 is
implicated in a number of cancers, such as breast and cervical
cancer, chondrosarcoma, colorectal and endometrial cancer,
esophageal squamous cell carcinoma as well as gallbladder carcinoma
(32,36,37), and
its downregulation inhibited tumor proliferation, migration,
invasion, metastasis, progression, and angiogenesis via distinctive
signaling pathways. For instance, miR-145 downregulation inhibited
invasion of bladder cancer cells by targeting PARK1 (38). Hsa-mir-145 downregulation was closely
associated with aggressive progression and poor prognosis in human
cervical cancer (31). miR-145
directly targeted p70S6K1 in cancer cells to inhibit colon cancer
tumor growth and angiogenesis (39).
miR-145 suppressed cell migration and invasion by targeting
paxillin in human colorectal cancer cells (40). In endometrioid carcinomas, miR-145 and
miR-143 were downregulated and associated with DNA
methyltransferase 3B overexpression and worse prognosis (41). In the present study, miRNA-145 was
predicted as a potential binding candidate of FTH1P33. Through
in vitro transfection with lncRNA FTH1P3 or siRNA FTH1P3
combined with miR-145 mimic and mutant (lncRNA-FTH1P3-Mut)
luciferase reporter, the association between FTH1P3 and miR-145 was
further confirmed.
Moreover, it was revealed that miRNA-145 inhibitor
could enhance cell proliferation, invasion, and migration in
cervical cancer cells, while FTH1P3 siRNA exhibited a significant
bucking effect for miRNA-145 inhibitor in HeLa and SiHa cells.
FTH1P3 siRNA partially attenuated the effects of the miR-145
inhibitor on cell viability and mobility in cervical cancer cells,
indicating that the FTH1P3-miR-145 axis functioned inversely in the
cervical tumor.
It is worth noting that this study is limited by the
small sample size. In addition, only in vitro cell
experiments were performed. Future studies with a larger sample
size and animal model experiments should be included to further
confirm the conclusions.
In conclusion, the present study demonstrated the
FTH1P3-miR-145 axis was involved in cell proliferation, invasion,
migration, apoptosis, viability, and mobility in cervical cancer
cells. These results may provide a potential therapeutic target and
strategy to impede the progression of cervical cancer.
Acknowledgments
Not applicable.
Funding
The present study was supported by Gansu Provincial
Cancer Hospital.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
RL designed and conceived this study and performed
the experiments and analysis of the data and wrote and supervised
the manuscript. QWZ performed the research and manuscript revision.
Both authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of Gansu Provincial Cancer Hospital. All patients and
healthy volunteers provided written informed consent prior to their
inclusion in the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
lncRNAs
|
long non-coding RNAs
|
|
MALAT1
|
metastasis-associated lung
adenocarcinoma transcript 1
|
|
FHC
|
ferritin heavy chain
|
|
FTH1P3
|
ferritin heavy chain 1 pseudogene
3
|
|
MEG3
|
maternally expressed 3
|
|
miRNA
|
microRNA
|
|
CSC
|
cancer stem cell
|
|
EMT
|
epithelial-mesenchymal transition
|
|
CCK-8
|
Cell Counting Kit-8
|
References
|
1
|
Zeferino LC and Derchain SF: Cervical
cancer in the developing world. Best Pract Res Clin Obstet
Gynaecol. 20:339–354. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ohazurike E, Anorlu I, Okunade K, Okunowo
A and Sajo A: Cervical cancer: An urgent need to scale up screening
in the developing world. Int J Gynecol Cancer. 27:842. 2017.
|
|
3
|
Nogueira-Rodrigues A, de Melo AC, Garces
AHI, Paulino E, Alves FV, Vilaça Mdo N, Silva LG, Gonçalves CA,
Fabrini JC, Carneiro AT and Thuler LC: Patterns of care and outcome
of elderly women diagnosed with cervical cancer in the developing
world. Int J Gynecol Cancer. 26:1246–1251. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rao GG, Rogers P, Drake RD, Nguyen P and
Coleman RL: Phase I clinical trial of weekly paclitaxel, weekly
carboplatin, and concurrent radiotherapy for primary cervical
cancer. Gynecol Oncol. 96:168–172. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Falcetta FS, Medeiros LR, Edelweiss MI,
Pohlmann PR, Stein AT and Rosa DD: Adjuvant platinum-based
chemotherapy for early stage cervical cancer. Cochrane Database
Syst Rev. 11:CD0053422016.PubMed/NCBI
|
|
6
|
Rosa DD, Medeiros LR, Edelweiss MI,
Bozzetti MC, Pohlmann PR, Stein AT and Dickinson HO: Adjuvant
platinum-based chemotherapy for early stage cervical cancer.
Cochrane Database Syst Rev. CD0053422009.PubMed/NCBI
|
|
7
|
Rosa DD, Medeiros LR, Edelweiss MI,
Pohlmann PR and Stein AT: Adjuvant platinum-based chemotherapy for
early stage cervical cancer. Cochrane Database Syst Rev.
CD0053422012.PubMed/NCBI
|
|
8
|
Kung JT, Colognori D and Lee JT: Long
noncoding RNAs: Past, present, and future. Genetics. 193:651–669.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jin XJ, Chen XJ, Hu Y, Ying F, Zou R, Lin
F, Shi Z, Zhu X, Yan X, Li S and Zhu H: LncRNA-TCONS_00026907 is
involved in the progression and prognosis of cervical cancer
through inhibiting miR-143-5p. Cancer Med. 6:1409–1423. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang J, Yao T, Wang Y, Yu J, Liu Y and
Lin Z: Long noncoding RNA MEG3 is downregulated in cervical cancer
and affects cell proliferation and apoptosis by regulating miR-21.
Cancer Biol Ther. 17:104–113. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jiang Y, Li Y, Fang S, Jiang B, Qin C, Xie
P, Zhou G and Li G: The role of MALAT1 correlates with HPV in
cervical cancer. Oncol Lett. 7:2135–2141. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Di Sanzo M, Aversa I, Santamaria G,
Gagliardi M, Panebianco M, Biamonte F, Zolea F, Faniello MC, Cuda G
and Costanzo F: FTH1P3, a Novel H-Ferritin pseudogene
transcriptionally active, is ubiquitously expressed and regulated
during cell differentiation. PLoS One. 11:e01513592016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang CZ: Long non-coding RNA FTH1P3
facilitates oral squamous cell carcinoma progression by acting as a
molecular sponge of miR-224-5p to modulate fizzled 5 expression.
Gene. 607:47–55. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zovoilis A, Mungall AJ, Moore R, Varhol R,
Chu A, Wong T, Marra M and Jones SJ: The expression level of small
non-coding RNAs derived from the first exon of protein-coding genes
is predictive of cancer status. EMBO Rep. 15:402–410. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chaudhry MA: Expression pattern of small
nucleolar RNA host genes and long non-coding RNA in X-rays-treated
lymphoblastoid cells. Int J Mol Sci. 14:9099–9110. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Aalto AP and Pasquinelli AE: Small
non-coding RNAs mount a silent revolution in gene expression. Curr
Opin Cell Biol. 24:333–340. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Braconi C, Henry JC, Kogure T, Schmittgen
T and Patel T: The role of MicroRNAs in human liver cancers. Semin
Oncol. 38:752–763. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wilmott JS, Zhang XD, Hersey P and Scolyer
RA: The emerging important role of microRNAs in the pathogenesis,
diagnosis and treatment of human cancers. Pathology. 43:657–671.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Esquela-Kerscher A: From worms to humans:
Understanding the role of microRNAs in cancer progression. Cancer
Res. 70:LB–353. 2010.
|
|
20
|
Gozuacik D, Akkoc Y, Ozturk DG and Kocak
M: Autophagy- regulating microRNAs and cancer. Front Oncol.
7:652017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chu R, Mo GQ, Duan Z, Huang M, Chang J, Li
X and Liu P: miRNAs affect the development of hepatocellular
carcinoma via dysregulation of their biogenesis and expression.
Cell Commun Signal. 12:452014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ahmad MK, Waseem M, Serajuddin M, Mahdi
AA, Sankhwar SN and Mishra DP: MicroRNA: A new potential marker for
prostate cancer. Cancer Med. 7:17. 2018.
|
|
23
|
Hironaka-Mitsuhashi A, Matsuzaki J,
Takahashi RU, Yoshida M, Nezu Y, Yamamoto Y, Shiino S, Kinoshita T,
Ushijima T, Hiraoka N, et al: A tissue microRNA signature that
predicts breast cancer recurrence in young women. PLoS One.
12:e01876382017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Baldassari F, Zerbinati C, Galasso M,
Corrà F, Minotti L, Agnoletto C, Previati M, Croce CM and Volinia
S: Screen for MicroRNA and Drug interactions in breast cancer cell
lines points to miR-126 as a modulator of CDK4/6 and PIK3CA
inhibitors. Front Genet. 9:1742018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Frixa T, Sacconi A, Cioce M, Roscilli G,
Ferrara FF, Aurisicchio L, Pulito C, Telera S, Carosi M, Muti P, et
al: MicroRNA-128-3p-mediated depletion of Drosha promotes lung
cancer cell migration. Carcinogenesis. 39:293–304. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Heydari N, Nikbakhsh N, Sadeghi F,
Farnoush N, Khafri S, Bastami M and Parsian H: Overexpression of
serum MicroRNA-140-3p in premenopausal women with newly diagnosed
breast cancer. Gene. 655:25–29. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiang SB, He XJ, Xia YJ, Hu WJ, Luo JG,
Zhang J and Tao HQ: MicroRNA-145-5p inhibits gastric cancer
invasiveness through targeting N-cadherin and ZEB2 to suppress
epithelial-mesenchymal transition. Oncotargets Ther. 9:2305–2315.
2016.
|
|
28
|
Noh JH, Chang YG, Kim MG, Jung KH, Kim JK,
Bae HJ, Eun JW, Shen Q, Kim SJ, Kwon SH, et al: MiR-145 functions
as a tumor suppressor by directly targeting histone deacetylase 2
in liver cancer. Cancer Lett. 335:455–462. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ding Y, Zhang C, Zhang J, Zhang N, Li T,
Fang J, Zhang Y, Zuo F, Tao Z, Tang S, et al: miR-145 inhibits
proliferation and migration of breast cancer cells by directly or
indirectly regulating TGF-β1 expression. Int J Oncol. 50:1701–1710.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang M, Wang J, Deng J, Li X, Long W and
Chang Y: MiR-145 acts as a metastasis suppressor by targeting
metadherin in lung cancer. Med Oncol. 32:3442015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Azizmohammadi S, Safari A, Azizmohammadi
S, Kaghazian M, Sadrkhanlo M, Yahaghi E, Farshgar R and
Seifoleslami M: Molecular identification of miR-145 and miR-9
expression level as prognostic biomarkers for early-stage cervical
cancer detection. QJM. 110:11–15. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou X, Yue Y, Wang R, Gong B and Duan Z:
MicroRNA-145 inhibits tumorigenesis and invasion of cervical cancer
stem cells. Int J Oncol. 50:853–862. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sathyanarayanan A, Chandrasekaran KS and
Karunagaran D: microRNA-145 downregulates SIP1-expression but
differentially regulates proliferation, migration, invasion and Wnt
signaling in SW480 and SW620 cells. J Cell Biochem. 119:2022–2035.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zheng X, Tang H, Zhao X, Sun Y, Jiang Y
and Liu Y: Long non-coding RNA FTH1P3 facilitates uveal melanoma
cell growth and invasion through miR-224-5p. PLoS One.
12:e01847462017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang R, Zhang T, Yang Z, Jiang C and Seng
J: Long non-coding RNA FTH1P3 activates paclitaxel resistance in
breast cancer through miR-206/ABCB1. J Cell Mol Med. 22:4068–4075.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li C, Lu L, Feng B, Zhang K, Han S, Hou D,
Chen L, Chu X and Wang R: The lincRNA-ROR/miR-145 axis promotes
invasion and metastasis in hepatocellular carcinoma via induction
of epithelial-mesenchymal transition by targeting ZEB2. Sci Rep.
7:46372017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mak IW, Singh S, Turcotte R and Ghert M:
The epigenetic regulation of SOX9 by miR-145 in human
chondrosarcoma. J Cell Biochem. 116:37–44. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kou B, Gao Y, Du C, Shi Q, Xu S, Wang CQ,
Wang X, He D and Guo P: miR-145 inhibits invasion of bladder cancer
cells by targeting PAK1. Urol Oncol. 32:846–854. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xu Q, Liu LZ, Qian X, Chen Q, Jiang Y, Li
D, Lai L and Jiang BH: MiR-145 directly targets p70S6K1 in cancer
cells to inhibit tumor growth and angiogenesis. Nucleic Acids Res.
40:761–774. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Qin J, Wang F, Jiang H, Xu J, Jiang Y and
Wang Z: MicroRNA-145 suppresses cell migration and invasion by
targeting paxillin in human colorectal cancer cells. Int J Clin Exp
Patho. 8:1328–1340. 2015.
|
|
41
|
Zhang X, Dong Y, Ti H, Zhao J, Wang Y, Li
T and Zhang B: Down-regulation of miR-145 and miR-143 might be
associated with DNA methyltransferase 3B overexpression and worse
prognosis in endometrioid carcinomas. Hum Pathol. 44:2571–2580.
2013. View Article : Google Scholar : PubMed/NCBI
|