Introduction
Although traditional chemotherapy drugs have
obtained high clinical efficacy in the treatment of cancer, there
are still a number of shortcomings, such as poor selectivity,
strong side effects and rapid blood clearance. In order to achieve
the characters of higher therapeutic effect, long half-life, and
low side effects, the drug conjugate consisting of target molecules
and chemotherapeutic drugs prepared by chemical coupling or genetic
recombination, which could be delivered into solid tumors and
cells, is a hot spot in anti-cancer drug research (1,2).
Human serum albumin (HSA), a single-chain
aglycosylated protein consisting of 585 amino acids, is the most
abundant protein in human plasma (3). Due to its non-immunogenicity, human
compatibility and long half-life in the serum (~19 days) (4), HSA is a widely recognized carrier for
the passive targeting to solid tumors and has been frequently
applied to construct drug conjugates for longer plasma half-life
(5–7). The albumin binding domain (ABD), a
protein domain with three helix structures discovered in the
surface proteins of gram-positive bacteria, exhibits the ability to
bind albumin (8). A mutation of ABD
(ABD035), obtained through screening and engineering and
deimmunized by substituting residues in immunogenic regions,
displays high affinity to both human serum albumin and mouse serum
albumin (MSA) with a dissociation constant of 10−13 M
(9). In addition, ABD035 exhibits
good stability, which makes it an ideal skeleton structure for
protein engineering (10).
Considering the non-covalent binding between albumin and ABD, the
construction of the recombinant protein containing ABD and a
therapeutic protein is beneficial to prolong plasma half-life,
increase treatment time and improve the efficacy of drugs (11–13).
Pancreatic carcinoma is an aggressive
gastrointestinal malignancy with high morbidity and mortality rates
worldwide; in 2018, the morbidity and mortality rates were 2.5 and
4.5%, respectively (14). Previous
studies on the molecular pathogenesis have demonstrated that the
occurrence, development and metastasis of pancreatic carcinoma are
closely associated with a variety of gene mutations and
abnormalities in cell signaling pathways, including K-ras and BRAF
mutations (15,16) and the epidermal growth factor
receptor (EGFR) and Hedgehog signaling pathways (17,18).
K-ras mutations, which are frequently present in pancreatic and
colon carcinomas, are regarded as an ideal therapeutic target owing
to their close association with oncogenesis, poor prognosis and
drug resistance (19), and they may
provide an efficient drug delivery strategy due to the intensive
macropinocytosis of extracellular nutrients observed in K-ras
mutant cells (20,21). However, no clinical drug targeting
K-ras mutations is currently available. EGFR upregulation in the
majority of human carcinomas is a validated target for cancer
therapy; therefore, a number of monoclonal antibodies and
small-molecule kinase inhibitors against EGFR, such as cetuximab
and erlotinib, have been applied for clinical treatment of
pancreatic cancer (22,23).
Lidamycin (LDM), which is a peptide antibiotic in
the process of phase II clinical trial, is considered to be an
ideal ‘warhead’ molecule for targeted drugs against tumors; LDM is
composed of an active enediyne chromophore (AE) with extremely
potent cytotoxicity and a non-covalently bound apoprotein (LDP),
which can be dissociated and reconstituted without the loss of
natural activity (24). In recent
years, a series of LDM modifications were performed to improve
tumor targeting, increase cytotoxicity on tumor cells and reduce
side effects. For example, the recombinant proteins integrating LDM
with EGFR/HER2-targeted oligopeptide or HSA enriched in solid
tumors and displayed stronger antitumor activity compared with LDM
in athymic mouse xenograft models (25,26).
In this study, a recombinant protein ABD-LDP-Ec and
its enediyne-integrated analogue ABD-LDP-Ec-AE were prepared and
their antitumor activities were studied with an aim to achieve
directional delivery of drugs against pancreatic cancer, especially
K-ras mutant pancreatic cancer, with the help of EGFR-targeting
binding, ABD-albumin combination and macropinocytosis.
Materials and methods
Preparation of the recombinant
proteins and their enediyne-integrated analogues
The DNA sequence of ABD was synthesized and inserted
into a pUC plasmid by GenScript Biotech Corporation, and the
plasmid pET30(a)-ldp containing the LDP gene was
constructed in our laboratory. The fragments of abd, ldp and
ldp-ec were amplified by PCR using the following primers:
abd forward (P1), 5′-GGAATTCCATATGCTGGCGGAAGCCAAAGTC-3′ and
reverse (P2),
5′-GAAGATCTGGCGGTGGCGGATCCGGCGGTGGCGGATCCCTGGCGGAAGCCAAAGTC-3′;
ldp and ldp-ec forward (P3),
5′-CAGAATTCGCGCCCGCCTTCTCCGTC-3′; ldp reverse (P4),
5′-CCGCTCGAGGCCGAACGTCAGTGCGAC-3′; and ldp-ec reverse (P5),
5′-CCGCTCGAGGCGCAGTTCCCACCATTTCAGATCGCGATACTGACAGCGTTCGCCAATATAGCCCACCACACAGTTTGAGCCACCTCCGCCTGAGCCACCTCCGCCGCCGAAGGTCAGAGC-3′.
The fragments were digested with NdeI, BglI and
XhoI restriction enzymes (Takara Biotechnology Co., Ltd.)
and ligated into the pET30(a) plasmid to construct
expression plasmids pET30(a)-abd-ldp and
pET30(a)-abd-ldp-ec.
The positive plasmids were confirmed by Invitrogen;
Thermo Fisher Scientific, Inc. and transformed into the expression
strain Escherichia coli BL21(DE3) Star (Novagen; Merck KGaA)
at 42°C for 45 sec. Following culture in Luria-Bertani medium (1%
NaCl, 1% peptone, 0.5% yeast extract; pH 7.4) at 37°C and induction
with 0.3 mM isopropyl β-D-thiogalactopyranoside (IPTG) at 30°C for
12 h, the bacterial cell pellets were harvested, resuspended in 20
mM Tris-HCl (pH 8.0) and sonicated on ice to collect the inclusion
bodies. The purification and refolding of the recombinant proteins
were performed as described by Sheng et al (27). The proteins were concentrated using
a centrifugal filter unit (EMD Millipore), and the concentration
was determined with a bicinchoninic acid (BCA) protein assay kit
(Thermo Fisher Scientific, Inc.).
To prepare the analogues of the recombinant proteins
(Fig. 1A), the active enediyne was
separated from lidamycin using a C4 column (GE Healthcare Life
Sciences) at 4°C overnight. The recombinant proteins reacted with
AE at room temperature for 12 h at a 1:3 molar ratio, and the free
AE was removed by ultrafiltration. The reconstituted proteins were
confirmed by reverse-phase high performance liquid chromatography
(HPLC) using a DELTA PAK C4 column (8×100 mm; Waters Corporation)
with an Alliance HPLC system (Waters Corporation) at 25°C. The
sample quantity was 40 µl, and the mobile phase consisted of 75%
solvent A (H2O with 0.05% v/v TFA) and 25% solvent B
(100% acetonitrile). The flow rate was 0.5 ml/min.
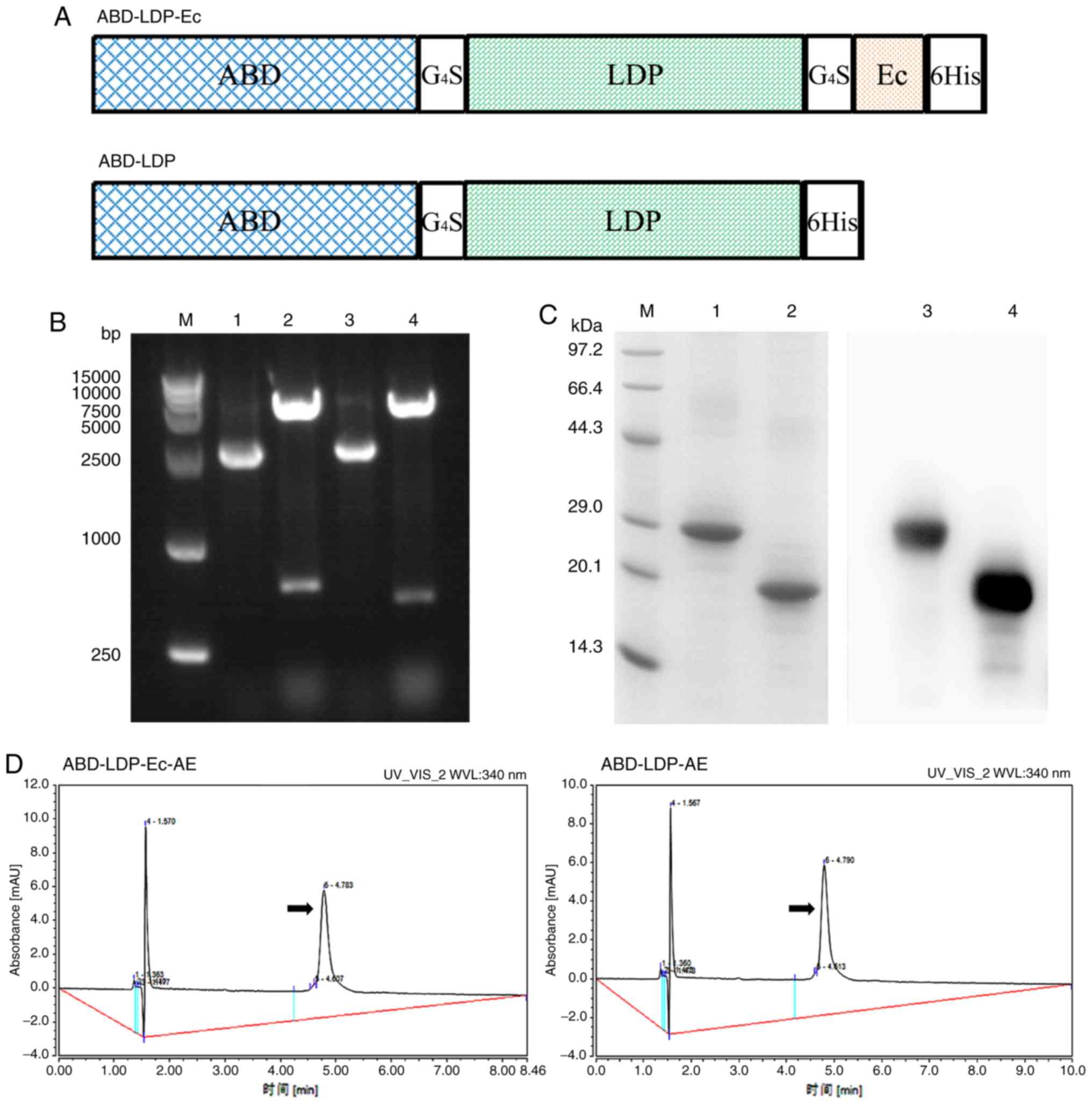 | Figure 1.Preparation of recombinant proteins
ABD-LDP-Ec, ABD-LDP and their enediyne-integrated analogues
ABD-LDP-Ec-AE, ABD-LDP-AE. (A) Schematic presentation of the
components of ABD-LDP-Ec and ABD-LDP. (B) Restriction enzyme
analysis of recombinant plasmids. Lane 1,
pET30(a)-abd-ldp-ec; lane 2, pET30(a)-abd-ldp-ec
digested with NdeI/ XhoI; lane 3,
pET30(a)-abd-ldp; lane 4, pET30(a)-abd-ldp digested
with NdeI/ XhoI. (C) SDS-PAGE and western blot
analysis of purified recombinant proteins. Lane 1 and 3, purified
ABD-LDP-Ec; lane 2 and 4, purified ABD-LDP. (D) Enediyne-integrated
analogues ABD-LDP-Ec-AE and ABD-LDP-AE determined by reverse-phase
HPLC at 340 nm. The absorption peaks of AE are indicated by arrows.
ABD, albumin-binding domain; LDP, lidamycin apoprotein; Ec,
epidermal growth factor receptor-targeting oligopeptide; AE,
enediyne chromophore; HPLC, high performance liquid
chromatography. |
Western blot analysis
The identification of proteins with His-tag was
performed by western blot analysis. Proteins (30 µl concentrated
solution per lane) were separated by 10% SDS-PAGE (5% stacking gel;
10% separating gel) and electrophoretically transferred onto PVDF
membranes (EMD Millipore). The membranes were blocked with 5% w/v
dry milk in TBS + 0.5% Tween-20 at 4°C overnight, incubated with a
horseradish peroxidase (HRP)-conjugated His-tag antibody (1:1,000;
cat. no. HRP-66005; Proteintech Group, Inc) at room temperature for
2 h, and the bands were visualized with Immobilon Western
Chemiluminescent HRP Substrate (EMD Millipore).
Cell culture
EGFR-positive human pancreatic cancer cell lines
AsPC-1, BxPC-3 and MIA PaCa-2 were obtained from American Type
Culture Collection. AsPC-1 and BxPC-3 cells were cultured in
modified RPMI medium (HyClone; GE Healthcare Life Sciences)
supplemented with 10% fetal bovine serum (FBS), 100 U/ml penicillin
and 100 µg/ml streptomycin (Gibco; Thermo Fisher Scientific, Inc.).
MIA PaCa-2 cells were cultured in high glucose DMEM (HyClone; GE
Healthcare Life Sciences) supplemented with the same additives. The
cells were cultured in a humidified incubator (Thermo Fisher
Scientific, Inc.) at 37°C with 95% air and 5% CO2.
Co-immunoprecipitation (Co-IP)
analysis
Co-IP analysis was performed using a
Co-immunoprecipitation kit (Pierce; Thermo Fisher Scientific, Inc.)
to verify the binding capacity of recombinant proteins against
albumin and EGFR, and mouse peripheral blood serum and cell lysate
of EGFR-positive AsPC-1 cells were used as the baits. The
peripheral blood was incubated for 0.5 h at room temperature and
centrifuged at 1,000 × g for 5 min at room temperature to collect
serum for further analysis. Adherent cultured AsPC-1 cells were
lysed in cell lysis buffer (Beyotime Institute of Biotechnology)
supplemented with 1 mM phenylmethylsulfonyl fluoride, and cellular
protein was obtained by high-speed centrifugation at 10,000 × g for
10 min 4°C. The protein concentration was quantified using a BCA
protein assay kit. The baits were re-cleared using the control
agarose resin to decrease non-specific binding. Each
bait/recombinant protein mixture at a 5:1 mass ratio was diluted in
IP lysis/wash buffer, added to the resin coupling albumin
(undiluted; cat. no. sc-271605) or EGFR antibody (undiluted; cat.
no. sc-373746; both from Santa Cruz Biotechnology) and incubated
with gentle agitation at 4°C overnight. Following centrifugation at
10,000 × g at 4°C for 5 min of the spin columns to remove unbound
proteins and washing the sample three times with IP lysis/wash
buffer, the flow-through of the elution buffer was collected for
western blot analysis.
ELISA assay
ELISA was performed to determine the binding
affinity of the recombinant proteins with HSA protein. The
recombinant proteins at 0.01, 0.1, 1, 10, 100, 1,000 or 10,000 nM
were added into 96-well plates coated with HSA (2 µg/well) and
incubated at 37°C for 2 h. Following incubation with an
HRP-conjugated His-tag antibody (1:1,000; cat. no. HRP-66005;
Proteintech Group, Inc) at room temperature for 2 h, 0.01%
3,3′,5,5′-tetramethylbenzidine (Tiangen Biotech Co., Ltd.) was
added as a substrate solution, and the reaction was terminated with
2 M H2SO4. The absorbance at 450 nm was
measured by a microplate reader (Thermo Fisher Scientific,
Inc.).
Transmission electron microscopy
(TEM)
TEM was used to observe the complex of ABD-LDP-Ec
and HSA. ABD-LDP-Ec was mixed with HSA at equimolar concentrations
and incubated at room temperature for 1 h. The imaging of protein
particles under TEM at ×400,000 magnification was performed by
Qingdao Sci-tech Innovation Co., Ltd.
Immunofluorescence assay
Immunofluorescence assay was performed to determine
the binding activity of ABD-LDP-Ec to pancreatic cancer cells
overexpressing EGFR. The recombinant proteins were pretreated with
DyLight 488 Antibody Labeling kit (Thermo Fisher Scientific, Inc.).
The proteins diluted in 0.05 M borate buffer were incubated with
the DyLight reagent at room temperature for 1 h protected from
light. The labeling reaction mixtures were added into the spin
columns preloaded with purification resin and mixed with the resin
by briefly vortexing, and the columns were centrifuged to collect
the labeled proteins. AsPC-1, MIA PaCa-2 and BxPC-3 cells were
seeded on coverslips, incubated at 37°C overnight and fixed with
100% methanol at −20°C for 10 min. The cells were incubated with 50
µM (~1 mg/ml) DyLight 488-labeled ABD-LDP-Ec at room temperature
for 1 h. Fluorescence was observed under a fluorescence microscope
(Nikon Corporation) at ×400 magnification and images were captured
in four random fields of view.
Flow cytometry
To compare the binding activity of the recombinant
proteins ABD-LDP-Ec, ABD-LDP and LDP to cancer cells, flow
cytometry was used. A total of 5×105 cells/tube of
AsPC-1 cells in the logarithmic phase were incubated with labeled
proteins in reaction buffer (PBS + 2% FBS) at 4°C for 1 h avoiding
internalization. Following washing with cold PBS, the cells were
resuspended and analyzed with a FACSCalibur™ cell analyzer (BD
Biosciences).
Internalization of the recombinant
proteins
Internalization of the recombinant proteins was
observed using laser scanning confocal microscopy. AsPC-1 (carrying
a K-ras mutation) and BxPC-3 (K-ras wild-type) cells were seeded in
Nunc™ Lab-Tek chambered coverglass (Thermo Fisher Scientific, Inc.)
at 2×104 cells/well and cultured overnight. The cells
were incubated with 50 µM DyLight 488-labeled ABD-LDP-Ec, ABD-LDP
or LDP and 50 µM HSA at room temperature for 1 h with or without
the specific macropinocytosis inhibitor ethyl-isopropyl amiloride
(EIPA). Fluoroshield mounting medium with DAPI (Abcam) was added to
stain the nuclei, and fluorescence images at ×400 magnification in
four random fields of view were captured by a confocal microscope
(Zeiss GmbH).
Flow cytometry was used to examine the association
between the internalization efficiency and HSA concentration or
reaction time. Following treatment with 50 µM DyLight 488-labeled
recombinant proteins and HSA, AsPC-1 cells were washed with PBS and
incubated in 0.4% trypan blue at room temperature for 10 min to
quench the fluorescence signal on the cell surface. The cells were
analyzed by flow cytometry using a FACSCalibur cell analyzer (BD
Biosciences) with FlowJo software (BD Biosciences).
In vitro cytotoxicity assay
To assess the cytotoxicity of the recombinant
proteins and their analogues to pancreatic cancer cells, clonogenic
and Cell Counting Kit-8 (CCK-8) viability assays were used,
respectively. For the recombinant proteins, AsPC-1, MIA PaCa-2 and
BxPC-3 cells were seeded in a 24-well plate at 100 cells/well and
cultured overnight for adhesion. The recombinant proteins and HSA,
at the molar ratio of 3:1, were added to treat tumor cells for 120
h. A colony was regarded as >30 cells, and the number of cell
colonies was counted under an optical microscope (Olympus
Corporation).
For the analogues of recombinant proteins, AsPC-1,
MIA PaCa-2 and BxPC-3 cells were seeded in a 96-well plate at 5,000
cells/well. Following treatment with the analogues and HSA for 48
h, CCK-8 reagent (Beyotime Institute of Biotechnology) was added
and incubated for 4 h. Absorbance at 570 nm was evaluated using a
microplate reader (Thermo Fisher Scientific, Inc.).
In vivo imaging of the recombinant
proteins
The animal studies were approved by the Ethics
Committee for Animal Experiments of The Institute of Medicinal
Biotechnology, Chinese Academy of Medical Sciences (approval no.
IMBF20060302). All animal experiments were performed in accordance
with the Good Laboratory Practice for Nonclinical Laboratory
Studies guidelines published by The Ministry of Science and
Technology of China. Female BALB/c (nu/nu) mice (4–6 weeks
old; n=41) were purchased from SPF (Beijing) Lab Animal Technology
Co., Ltd.
The recombinant proteins were labeled using a
DyLight 680 Antibody Labeling kit (Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. Each mouse (n=5) was
subcutaneously inoculated with 1×107 human pancreatic
cancer AsPC-1, MIA PaCa-2, or BxPC-3 cells suspended in 0.2 ml PBS
in the right armpit. When the tumor volume was ~400 mm3,
0.4 mg DyLight 680-labeled recombinant proteins were administered
intravenously through the tail vein. Images of fluorescence
distribution in the mice were observed and recorded by IVIS
Spectrum System (PerkinElmer, Inc.), and fluorescence signals were
measured and analyzed using Living Image software (PerkinElmer,
Inc.).
In vivo efficacy study
Due to EGFR overexpression and the K-ras mutation,
human pancreatic cancer AsPC-1 cell line was used in a xenograft
model in athymic mice to determine the therapeutic efficacy of the
recombinant proteins and their analogues. Mice were subcutaneously
inoculated with 1×107 AsPC-1 tumor cells in the right
flank and randomly divided into groups of 6 mice when the tumor
volumes were >100 mm2. The recombinant proteins and
their analogues (20 mg/kg ABD-LDP, 20 mg/kg ABD-LDP-Ec, 0.05 mg/kg
LDM, 0.1 mg/kg ABD-LDP-AE or 0.1 mg/kg ABD-LDP-Ec-AE) were
administered intravenously through the tail vein twice with a 7-day
interval. Tumor size and mouse weight were measured every three
days, and tumor volume was calculated using the following formula:
Tumor volume = 0.5 × length × (width)2. The maximum
allowed tumor volume was 1,500 mm3. On day 30, the mice
were euthanized by cervical dislocation and aseptically dissected,
and solid tumors were collected. The tumors were weighed and the
tumor growth inhibition (TGI) was calculated as follows: TGI =
(1-T/C) ×100%, where T is the mean tumor weight of the therapy
group and C is the mean tumor weight of the control group.
Statistical analysis
Statistical analysis was performed using GraphPad
Prism 7 software (GraphPad Software, Inc.). Data are presented as
the mean ± SEM. Student's t-test was used to compare two groups,
whereas Tukey's test with one-way ANOVA was used for multiple
comparisons. P<0.05 was considered to indicate a statistically
significant difference.
Results
Preparation of the recombinant
proteins and their analogues
Recombinant plasmids
pET30(a)-abd-ldp-ec and
pET30(a)-abd-ldp including gene fragments encoding
albumin binding domains, apoprotein of lidamycin, EGFR-directed
ligand peptide and glycine-serine (G4S) linkers were
constructed and identified with NdeI/XhoI digestion
(Fig. 1B). Recombinant proteins
ABD-LDP-Ec and ABD-LDP with a His-tag at C-terminus were produced
in the form of inclusion bodies by E. coli BL21(DE3) Star
following IPTG induction, purified with Ni2+ affinity
chromatography and refolded by stepwise dialysis. A total of ~20 mg
ABD-LDP-Ec and ~30 mg ABD-LDP were yielded from 1 l fermentation
broth and migrated as a band of 21.1 and 19.8 kDa in SDS-PAGE under
reducing conditions, respectively (Fig.
1C). The analogues of recombinant proteins ABD-LDP-Ec-AE and
ABD-LDP-AE were prepared by integrating the recombinant proteins
with active enediyne chromophore in vitro, and the
absorption peak of AE at 340 nm was detected by reverse-phase HPLC,
indicating successful assembly of enediyne analogues (Fig. 1D).
Affinity of the recombinant
proteins
To confirm the binding capacity of the recombinant
proteins ABD-LDP-Ec, ABD-LDP and LDP against albumin and EGFR,
co-immunoprecipitation assay and western blot analysis were used.
As demonstrated by the western blot analysis, ABD-LDP-Ec and
ABD-LDP formed complexes with albumin sourced from mouse serum
(Fig. 2A). ABD-LDP-Ec and ABD-LDP
exhibited binding capacity in AsPC-1 cell lysate. No binding was
observed between LDP and the target proteins.
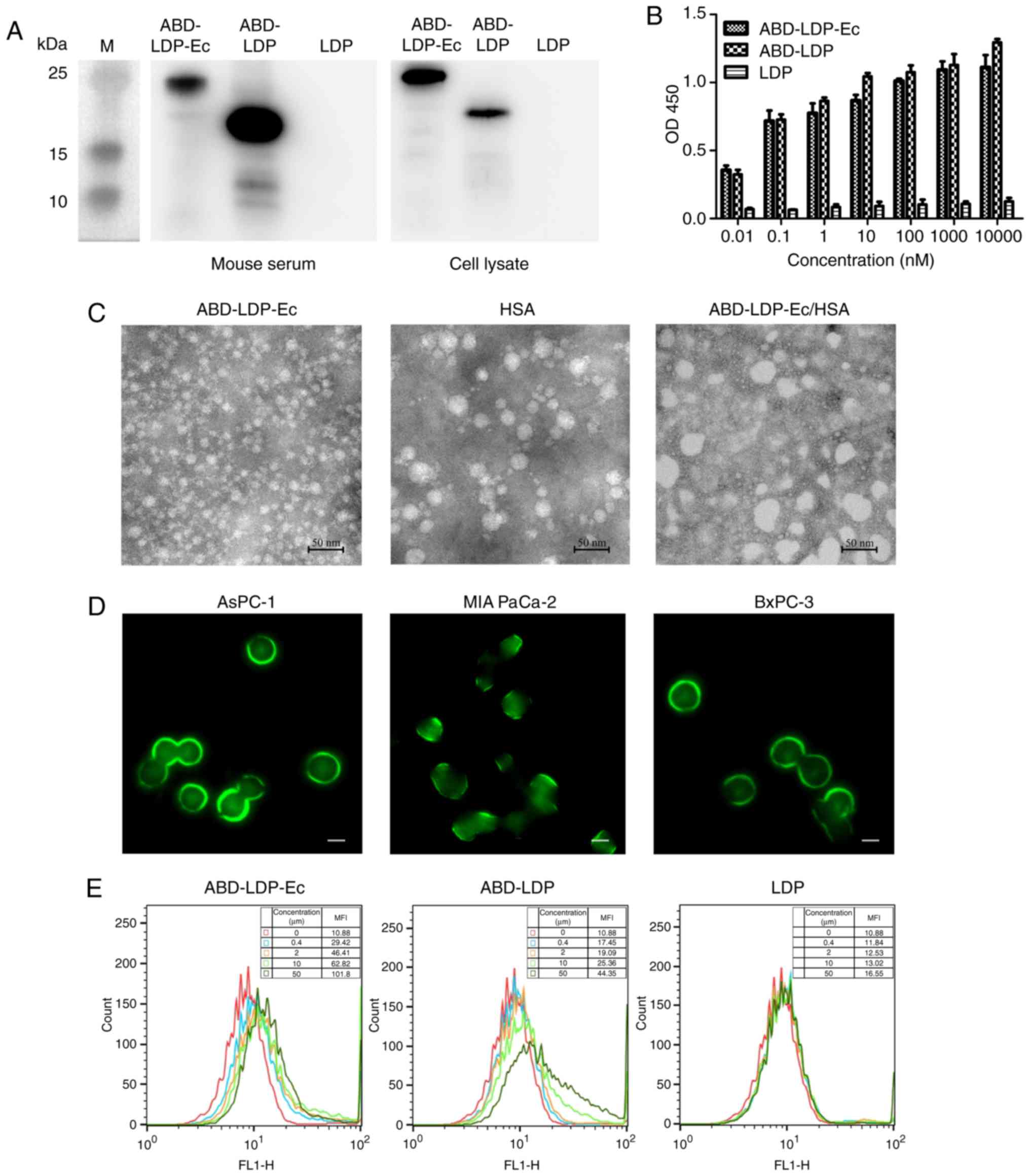 | Figure 2.Recombinant proteins bind to target
proteins and pancreatic cancer cells. (A) Co-immunoprecipitation
assay of the recombinant protein binding specificity to HSA and
EGFR. Mouse peripheral blood serum and AsPC-1 cell lysate were
incubated with ABD-LDP-Ec, ABD-LDP or LDP. The complexes were
collected using a co-immunoprecipitation kit and analyzed by
SDS-PAGE and immunoblotting with a His-tag monoclonal antibody. (B)
ELISA analysis of the recombinant protein binding with HSA protein.
(C) Transmission electron microscopy observation of ABD-LDP-Ec, HSA
and ABD-LDP-Ec/HSA complex particles. (D) Immunofluorescence assay
of ABD-LDP-Ec bound to AsPC-1, MIA PaCa-2 and BxPC-3 cells. Green
fluorescence located around the cells indicated ABD-LDP-Ec bound to
EGFR on the cell membrane. Magnification, ×400; scale bar, 5 µm.
(E) Flow cytometry analysis of the binding activity of DyLight
488-labeled ABD-LDP-Ec, ABD-LDP and LDP to AsPC-1 cells. MFI, mean
fluorescence intensity; OD, optical density; HSA, human serum
albumin; EGFR, epidermal growth factor receptor; ABD,
albumin-binding domain; LDP, lidamycin apoprotein; Ec,
EGFR-targeting oligopeptide; AE, enediyne chromophore. |
Comparison of binding to HSA protein among the three
proteins was examined by ELISA. ABD-LDP-Ec and ABD-LDP exhibited
high affinity for HSA, whereas that of LDP was weak (Fig. 2B). ABD-LDP presented slightly
stronger binding efficiency compared with ABD-LDP-Ec, which may
have been due to the steric effects of Ec. The particles of
ABD-LDP-Ec, HSA and the ABD-LDP-Ec/HSA complex were observed by
TEM; as demonstrated in Fig. 2C,
ABD-LDP-Ec/HSA particles were larger compared with the individual
proteins. The diameter of the complex was ~50 nm.
EGFR-positive AsPC-1, MIA PaCa-2 and BxPC-3 cells
were selected for the detection of the recombinant protein
cell-binding activity. Green fluorescence was observed on the
surface of cancer cells under a fluorescence microscope, indicating
that ABD-LDP-Ec possessed superior binding capacity with pancreatic
cancer cells compared with ABD-LDP and LDP (Fig. 2D). The slight fluorescence observed
in the cells may be explained by internalization of the recombinant
proteins at room temperature. In flow cytometry experiments,
ABD-LDP-Ec displayed a significantly stronger affinity to AsPC-1
cells compared with ABD-LDP and LDP (Fig. 2E) owing to the molecular recognition
of EGFR by Ec. ABD-LDP at a high concentration also exhibited a
certain affinity for AsPC-1 cells, whereas the affinity of LDP was
low.
Internalization of the recombinant
proteins in pancreatic cancer cells
AsPC-1 and BxPC-3 cells treated with recombinant
proteins labeled with DyLight 488 were observed under a laser
scanning confocal microscope. In the K-ras mutant AsPC-1 cells, HSA
substantially improved the uptake of ABD-LDP-Ec and ABD-LDP, and
the specific macropinocytosis inhibitor EIPA reversed the effects
of HSA on endocytosis (Fig. 3A). In
the K-ras wild-type BxPC-3 cells, HSA and EIPA exhibited limited
effects on protein uptake (Fig.
3B). These results suggested that ABD-LDP-Ec and ABD-LDP
benefited from HSA to internalize into K-ras mutant tumor cells by
macropinocytosis-mediated uptake. In addition, the Ec component
also promoted the internalization of ABD-LDP-Ec in EGFR-positive
tumor cells.
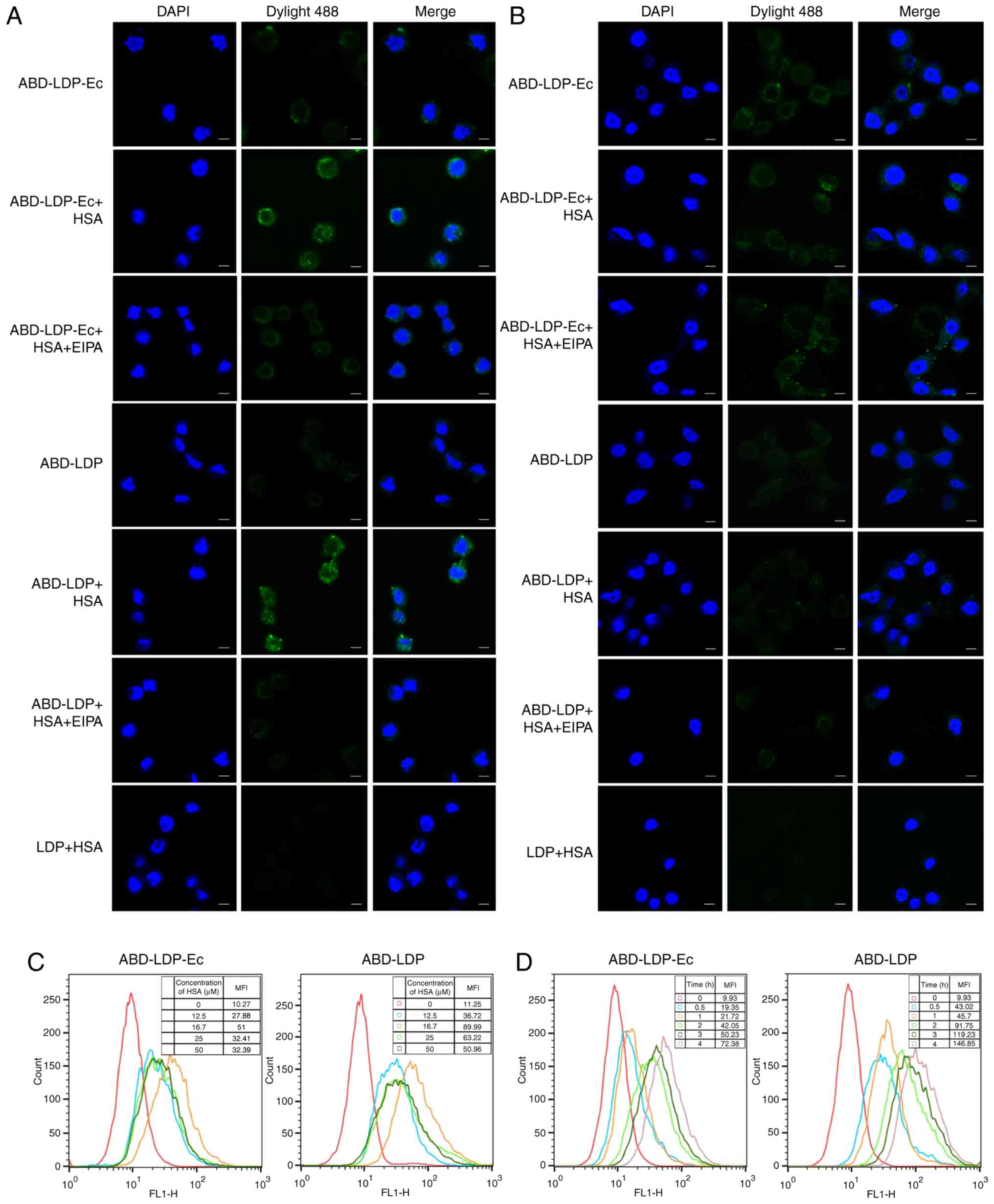 | Figure 3.Recombinant proteins are internalized
into pancreatic cancer cells. (A) Confocal microscopic observation
of K-ras mutant AsPC-1 cells treated with the recombinant proteins
with or without HSA and EIPA. (B) Confocal microscopic observation
of K-ras wild-type BxPC-3 cells treated with the recombinant
proteins with or without HSA and EIPA. Cells were observed under a
confocal microscope at ×400 magnification. Green fluorescence,
DyLight 488-labeled recombinant proteins; blue fluorescence,
DAPI-stained nuclei; scale bar, 5 µm. (C) Flow cytometry analysis
of the association between 50 µM recombinant protein uptake and HSA
concentration (0, 12.5, 16.7, 25, 50 µM) in AsPC-1 cells. (D) With
50 µM recombinant proteins and 16.7 µM HSA, flow cytometry analysis
of the association between protein uptake and incubation time (0,
0.5, 1, 2, 3, 4 h) in AsPC-1 cells. MFI, mean fluorescence
intensity. HSA, human serum albumin; EIPA, ethyl-isopropyl
amiloride; ABD, albumin-binding domain; LDP, lidamycin apoprotein;
Ec, epidermal growth factor receptor-targeting oligopeptide; AE,
enediyne chromophore. |
In K-ras mutant AsPC-1 cells, flow cytometry
detection demonstrated that the amount of protein uptake was
associated with HSA concentration and incubation time. For HSA
concentration, the dependence had two phases. As presented in
Fig. 3C, with 50 µM recombinant
proteins, the maximum amount of ABD-LDP-Ec and ABD-LDP were
internalized into AsPC-1 cells when HSA was at 16.7 µM. When the
HSA concentration was <16.7 µM, the internalization of
recombinant proteins was improved with increasing HSA
concentration, whereas when HSA concentration was >16.7 µM, the
internalization was reduced with increasing HSA concentration. The
internalization of ABD-LDP-Ec and ABD-LDP was enhanced following a
longer incubation (Fig. 3D), and
the endocytosis of ABD-LDP was higher compared with that of
ABD-LDP-Ec.
In vitro cytotoxicity of the
recombinant proteins and their analogues
The anti-proliferative effect of the recombinant
proteins and their analogues on pancreatic cancer cells was
measured by a clonogenic and CCK-8 assays, respectively. As
presented in Fig. 4A, the
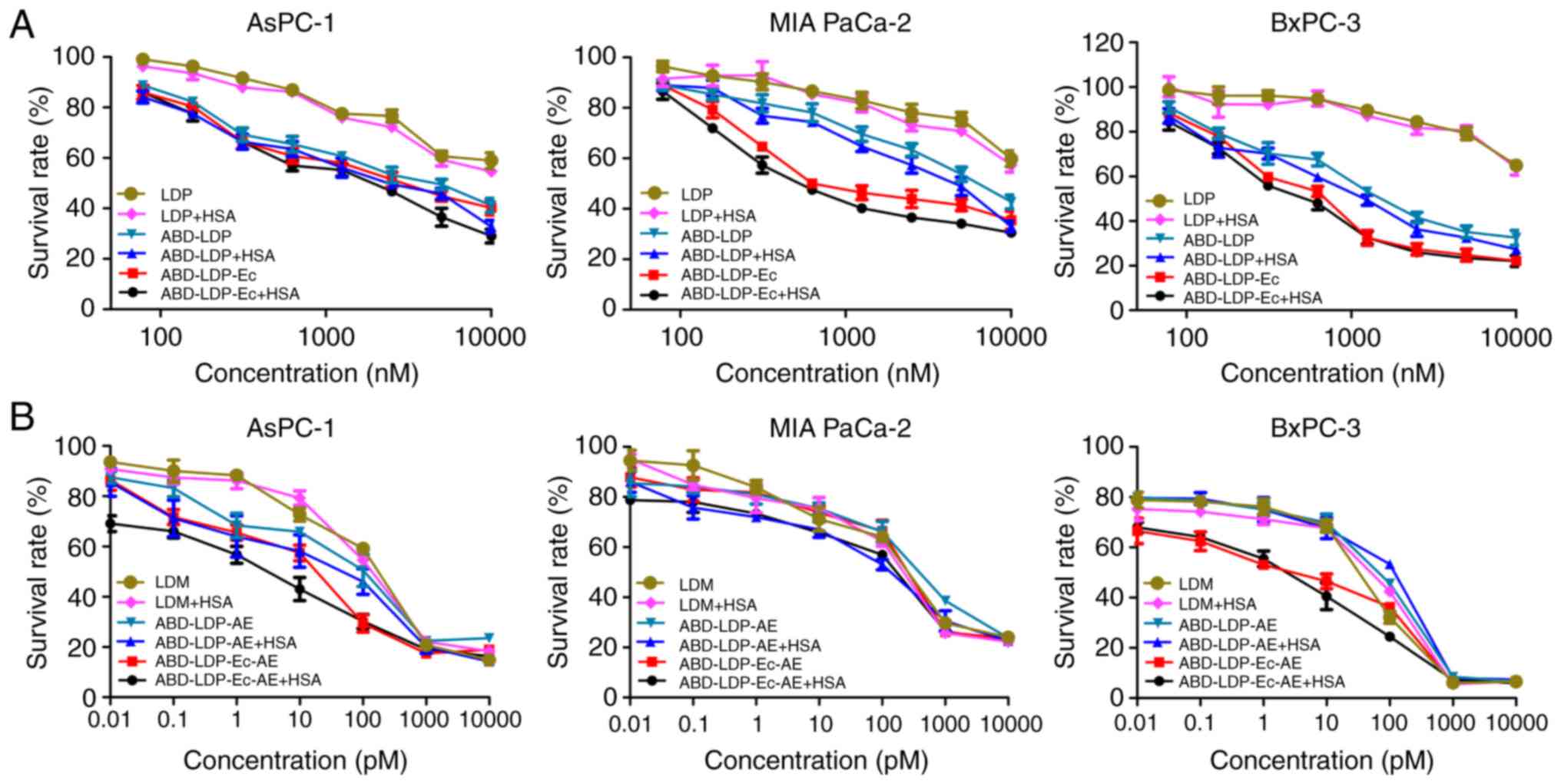
recombinant proteins at 100–10,000 nM moderately inhibited the
proliferation of cancer cells, and the efficacy of ABD-LDP-Ec and
ABD-LDP was stronger compared with LDP, especially for BxPC-3
cells. In addition, enhanced inhibitory activity of ABD-LDP-Ec and
ABD-LDP with HSA on the K-ras mutant AsPC-1 and MIA PaCa-2 cells
was observed (Table I). The
enediyne-integrated analogues ABD-LDP-Ec-AE and ABD-LDP-AE
exhibited strong proliferation inhibition on pancreatic cancer
cells with IC50 values of 1–100 pM (Fig. 4B), and HSA significantly promoted
the cytotoxicity of the analogues on K-ras mutant cells (Table II).
 | Table I.Proliferation inhibition by the
recombinant proteins in pancreatic cancer cells determined by the
clonogenic assay. |
Table I.
Proliferation inhibition by the
recombinant proteins in pancreatic cancer cells determined by the
clonogenic assay.
|
| IC50, M
(mean ± SEM) |
|---|
|
|
|
|---|
| Recombinant
protein | AsPC-1 | MIA PaCa-2 | BxPC-3 |
|---|
| ABD-LDP-Ec +
HSA |
1.44±0.03×10−6a |
1.05±0.18×10−6a |
7.10±0.71×10−7 |
| ABD-LDP-Ec |
2.16±0.11×10−6 |
1.30±0.11×10−6 |
8.12±0.65×10−7 |
| ABD-LDP + HSA |
1.80±0.08×10−6a |
3.03±0.09×10−6a |
1.43±0.18×10−6 |
| ABD-LDP |
2.44±0.14×10−6 |
4.21±0.24×10−6 |
1.75±0.09×10−6 |
| LDP + HSA |
7.23±0.31×10−6a |
8.54±0.80×10−6a |
1.36±0.17×10−5 |
| LDP |
8.10±0.48×10−6 |
1.04±0.10×10−5 |
1.50±0.11×10−5 |
| Table II.Proliferation inhibition by the
enediyne-integrated analogues and LDM on pancreatic cancer cells
determined by Cell Counting Kit-8 viability assay. |
Table II.
Proliferation inhibition by the
enediyne-integrated analogues and LDM on pancreatic cancer cells
determined by Cell Counting Kit-8 viability assay.
|
| IC50, M
(mean ± SEM) |
|---|
|
|
|
|---|
| Recombinant protein
analogue | AsPC-1 | MIA PaCa-2 | BxPC-3 |
|---|
| ABD-LDP-Ec-AE +
HSA |
1.69±0.08×10−12a |
6.04±0.48×10−11a |
9.22±0.29×10−13 |
| ABD-LDP-Ec +
AE |
7.91±0.84×10−12 |
1.61±0.10×10−10 |
1.21±0.23×10−12 |
| ABD-LDP -AE +
HSA |
9.44±0.68×10−12a |
6.88±0.28×10−11a |
2.43±0.28×10−11 |
| ABD-LDP + AE |
4.38±0.30×10−11 |
3.20±0.15×10−10 |
2.13±0.29×10−11 |
| LDM + HSA |
1.19±0.06×10−10 |
1.44±0.08×10−10 |
1.10±0.17×10−11 |
| LDM |
1.06±0.04×10−10 |
1.78±0.11×10−10 |
1.12±0.24×10−11 |
Optical imaging of the recombinant
proteins in vivo
Female BALB/c (nu/nu) mice bearing human
pancreatic cancer AsPC-1, MIA PaCa-2 and BxPC-3 ×enografts were
used to detect the biodistribution of the recombinant proteins
in vivo. 0.4 mg DyLight 680-labeled recombinant proteins
were injected intravenously, and images were captured and recorded
by an IVIS Spectrum System at 1, 2, 4, 8, 12, 24, 36 and 48 h. No
tumor site fluorescence was observed in the LDP group; by contrast,
localized fluorescence was observed in ABD-LDP-Ec and ABD-LDP
groups, which reflected the enhanced tumor-targeting ability of
recombinant proteins in vivo. ABD-LDP-Ec benefited from the
EGFR-targeting characteristic of Ec and exhibited an improved
tumor-targeting capacity compared with ABD-LDP in the AsPC-1
×enograft that presented as protein enrichment in <1 h and
maximum intensity at 8 h (Fig. 5A).
Compared with the K-ras wild-type BxPC-3 ×enograft (Fig. 5C), K-ras mutant AsPC-1 and MIA
PaCa-2 ×enografts (Fig. 5A and B)
were beneficial for the enrichment and reservation of ABD-LDP-Ec
owing to the macropinocytosis of the ABD-LDP-Ec/HSA complex and its
accumulation in the tumors.
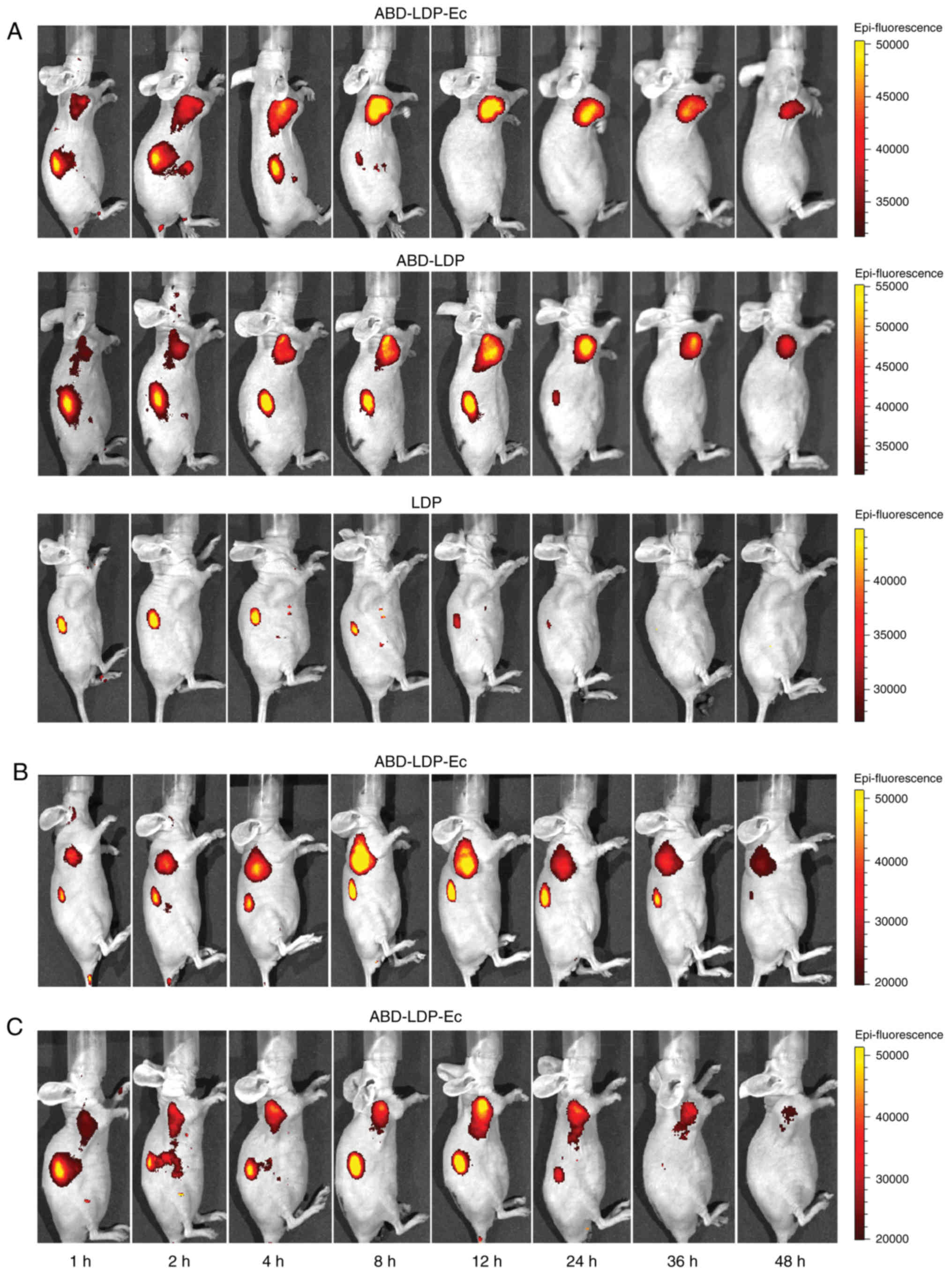 | Figure 5.Distribution of the recombinant
proteins in xenograft-bearing nude mice in vivo. (A)
Representative fluorescence images at the indicated times (1, 2, 4,
8, 12, 24, 36 and 48 h) post-intravenous injection of 0.4 mg
DyLight 680-labeled ABD-LDP-Ec, ABD-LDP or LDP in AsPC-1
×enograft-bearing nude mice. (B) Representative fluorescence images
post-intravenous injection of DyLight 680-labeled ABD-LDP-Ec in MIA
PaCa-2 ×enograft-bearing nude mice. (C) Representative fluorescence
images post-intravenous injection of DyLight 680-labeled ABD-LDP-Ec
in BxPC-3 ×enograft-bearing nude mice. ABD, albumin-binding domain;
LDP, lidamycin apoprotein; Ec, epidermal growth factor
receptor-targeting oligopeptide. |
In vivo efficacy of the recombinant
proteins and their analogues
Human pancreatic cancer AsPC-1 ×enograft in BALB/c
(nu/nu) mice was used to evaluate the antitumor activity of
the recombinant proteins and their analogues. When solid tumors
were established following subcutaneous AsPC-1 cell inoculation,
the reconstituted analogues were administrated at an equivalent
molar dose of LDM (0.05 mg/kg), which was the tolerance dose
described previously (28), through
the tail vein. The recombinant proteins were used at the same
concentration as in the optical imaging experiment (~20 mg/kg). In
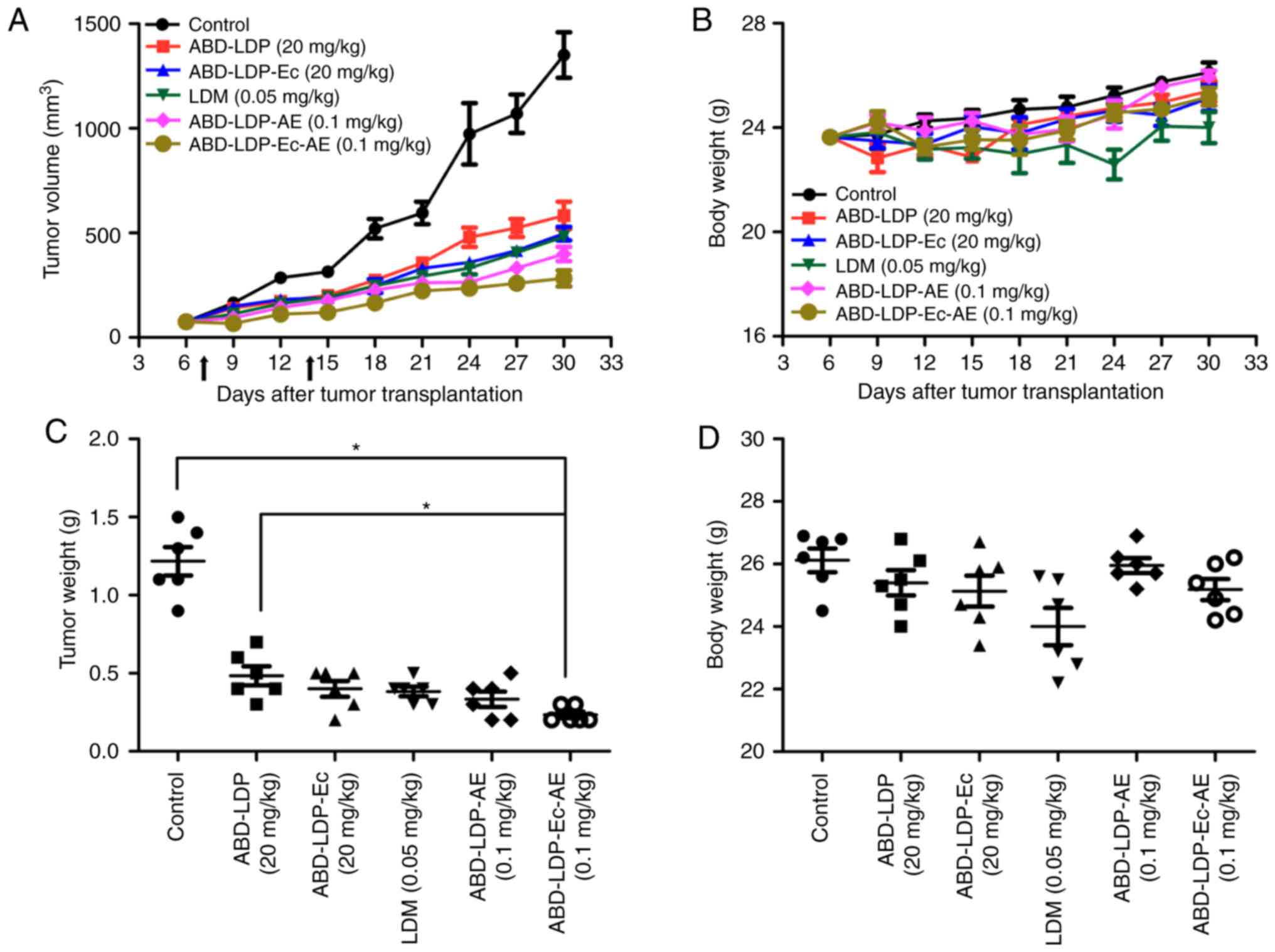
Fig. 6A, the curves of tumor volume
over time demonstrated that the energized proteins presented
stronger antitumor activity compared with the recombinant proteins
and non-targeted LDM. At the end of experiment, the inhibitory
rates based on tumor weight were 81.9% for ABD-LDP-Ec-AE and 75.2%
for ABD-LDP-AE, respectively, which were higher compared with 68.6%
for LDM (Fig. 6C). In addition,
high-dose ABD-LDP-Ec and ABD-LDP also exhibited moderate inhibition
in the AsPC-1 ×enograft. Body weights increased throughout the
experiment among all experimental groups and the control, and no
obvious differences were observed among the groups (Fig. 6B and D), which indicated that the
administered doses were well tolerated.
Discussion
Targeted drug delivery serves an important role in
improving the efficacy and reducing the side effects of drugs. A
new form of recombinant protein ABD-LDP-Ec based on an
albumin-binding domain and an EGFR-targeted oligopeptide was
produced and used to prepare an anti-cancer drug capable of active
and passive tumor targeting. Oligopeptide Ec, which comprises the
22 amino acids of the EGF COOH-terminal sufficient for
high-affinity receptor binding (29), was used as an EGFR-targeting
molecule to perform active targeting of EGFR-positive tumor cells.
In contrast to conventional drug designs such as β-defensin 2-HSA
(30), thioredoxin-albumin
(31) and HSA-eTGFBR2 (32), the ABD035 molecule instead of HSA
was fused with drug molecules in the present study and exerted the
properties of passive targeting and half-life extension, which
resulted in the enrichment and retention of ABD-LDP-Ec and ABD-LDP
in tumor tissue through capturing the functional albumin in plasma.
In addition, ABD-LDP displayed a certain binding capacity to tumor
cells, although the mechanism of action remains unclear. Compared
with the proteins combining drugs with an antibody and HSA,
Ec/ABD-fused proteins can be produced in engineered bacteria, and
the small and efficient target molecules result in less
interference with the function of fusion proteins and lower immune
response (12).
Macropinocytosis is a highly conservative endocytic
pathway observed in certain macrophages (33), dendritic (34) and tumor (35) cells that implements transmembrane
transport of extracellular substances by the macropinocytosomes
formed of plasma membrane folds. Macropinocytosis is significantly
enhanced in tumor cells carrying EGFR or proto-oncogene mutations,
such as Ras (36), Rac (37) and Sre (38). As reported by Commisso et al
(20), macropinocytosis serves an
important role in the nutrient uptake of Ras mutant tumor cells,
which enables HSA to be transported into tumor cells and further be
decomposed into essential amino acids for sustained cell growth and
proliferation, suggesting that HSA may be used as a
macropinocytosis-targeted drug delivery carrier. For example,
albumin-modified β-defensin DF-HSA displayed intensive
macropinocytosis-mediated uptake in K-ras mutant MIA PaCa-2 cells
and exerted high therapeutic efficacy against pancreatic xenograft
tumors in athymic mice (29). The
nanoparticle albumin-bound paclitaxel, which is the first-line
treatment drug for pancreatic cancer combined with gemcitabine, had
been demonstrated to be internalized into macrophages by
macropinocytosis and to induce macrophage immunostimulatory
cytokine expression (39).
ABD-LDP-Ec and ABD-LDP combined with HSA achieved higher tumor
uptake relative to LDP in AsPC-1 and MIA PaCa-2 cells, and the
tumor uptake was associated with the content of HSA. The optimum
molar ratio of ABD-LDP-Ec to HSA for the maximum uptake was 3:1,
implying that excessive HSA monomers may hinder the endocytosis of
the protein/HSA complex by occupying macropinocytosis sites.
Since LDP generally acts as the skeleton structure
of recombinant proteins and the protective group for protecting and
stabilizing the chromophore, ABD-LDP-Ec was assembled with the
active enediyne of LDM by molecular reconstitution to produce its
enediyne-integrated analogue ABD-LDP-Ec-AE, which demonstrated
1×106-fold stronger inhibition on pancreatic cancer
cells compared with ABD-LDP-Ec. Compared with LDM, ABD-LDP-Ec-AE at
a lower concentration effectively inhibited the proliferation of
AsPC-1, MIA PaCa-2 and BxPC-3 cells, whereas the TGI rate on AsPC-1
×enografts was improved by 20% at the same molar concentration.
Benefiting from the EGFR-targeting Ec and the combination of ABD
and HSA, ABD-LDP-Ec-AE was retained in tumor tissues for 48 h due
to the enhanced permeability and retention effect, and was
transported into K-ras mutant cancer cells, which significantly
prolonged the duration of LDM activity and increased the
cytotoxicity. In addition, free ABD-LDP-Ec-AE was decomposed and
eliminated through biotransformation functions of liver within 8 h,
and the damage of LDM on normal cells and tissues was weakened.
In summary, the results of the present study
demonstrated that an ABD/oligopeptide-based cancer-targeted
recombinant protein ABD-LDP-Ec and its enediyne-integrated analogue
ABD-LDP-Ec-AE exhibited more potent antitumor efficacy compared
with LDP and LDM in EGFR-positive and K-ras mutant pancreatic
cancer. With the help of the albumin-binding ABD, ABD-LDP-Ec was
enriched in solid tumors through the passive targeting of albumin,
bound to EGFR on the cell membrane and was internalized into the
cytoplasm via receptor-mediated endocytosis and albumin-induced
macropinocytosis of K-ras mutant cells. These results suggested
that the introduction of ABD-based multi-functional drug delivery
may be an effective approach to improve the efficacy of antitumor
drugs, especially for K-ras mutant cancers.
Acknowledgements
Not applicable.
Funding
This research was supported by the Chinese Academy
of Medical Sciences (CAMS) Innovation Fund for Medical Sciences
(grant nos. 2016-I2M-1-011 and 2019-I2M-1-005).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ and MJ designed this study. WS and JG conducted
main experiments and analyzed the data. LL performed molecular
reconstitution. YS performed animal experiments. WS wrote the
manuscript. All authors have read and approved the final manuscript
and agree to be accountable for all aspects of the research in
ensuring that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
The animal studies were approved by the Ethics
Committee for Animal Experiments of The Institute of Medicinal
Biotechnology, Chinese Academy of Medical Sciences (approval no.
IMBF20060302). All animal experiments were performed in accordance
with the Good Laboratory Practice for Nonclinical Laboratory
Studies guidelines published by The Ministry of Science and
Technology of China.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ABD
|
albumin-binding domain
|
|
EGFR
|
epidermal growth factor receptor
|
|
LDM
|
lidamycin
|
|
LDP
|
lidamycin apoprotein
|
|
AE
|
enediyne chromophore
|
|
Ec
|
EGFR-targeting oligopeptide
|
|
HSA
|
human serum albumin
|
|
MSA
|
mouse serum albumin
|
|
IPTG
|
isopropyl
β-D-thiogalactopyranoside
|
|
HPLC
|
high performance liquid
chromatography
|
|
HRP
|
horseradish peroxidase
|
|
TEM
|
transmission electron microscopy
|
|
EIPA
|
ethyl-isopropyl amiloride
|
|
CCK-8
|
cell counting kit-8
|
|
TGI
|
tumor growth inhibition
|
References
|
1
|
Carter PJ and Senter PD: Antibody-drug
conjugates in cancer therapy. Cancer J. 14:154–169. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Feng Q and Tong R: Anticancer
nanoparticulate polymer drug conjugate. Bioeng Trans Med.
1:277–296. 2016. View Article : Google Scholar
|
|
3
|
Ma P and Mumper RJ: Paclitaxel
nano-delivery systems: A comprehensive review. J Nanomed
Nanotechnol. 4:10001642013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nakhaei E, Kim CW, Funamoto D, Sato H,
Nakamura Y, Kishimura A, Mori T and Katayama Y: Design of a ligand
for cancer imaging with long blood circulation and an enhanced
accumulation ability in tumors. Medchemcomm. 8:1190–1195. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wei Y, Wang Y, Xia D, Guo S, Wang F, Zhang
X and Gan Y: Thermosensitive liposomal codelivery of hsa-paclitaxel
and hsa-ellagic acid complexes for enhanced drug perfusion and
efficacy against pancreatic cancer. ACS Appl Mater Interfaces.
9:25138–25151. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Byeon HJ, Min SY, Kim I, Lee ES, Oh KT,
Shin BS, Lee KC and Youn YS: Human serum albumin-TRAIL conjugate
for the treatment of rheumatoid arthritis. Bioconjug Chem.
25:2212–2221. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sheffield WP, Eltringham-Smith LJ and
Bhakta V: Fusion to human serum albumin extends the circulatory
half-life and duration of antithrombotic action of the kunitz
protease inhibitor domain of protease nexin 2. Cell Physiol
Biochem. 45:772–782. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
de Château M, Holst E and Björck L:
Protein PAB, an albumin-binding bacterial surface protein promoting
growth and virulence. J Biol Chem. 271:26609–26615. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jonsson A, Dogan J, Herne N, Abrahmsén L
and Nygren PA: Engineering of a femtomolar affinity binding protein
to human serum albumin. Protein Eng Des Sel. 21:515–527. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nilvebrant J and Hober S: The
albumin-binding domain as a scaffold for protein engineering.
Comput Struct Biotechnol J. 6:e2013030092013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cantante C, Lourenco S, Morais M, Leandro
J, Gano L, Silva N, Leandro P, Serrano M, Henriques AO, Andre A, et
al: Albumin-binding domain from Streptococcus zooepidemicus protein
Zag as a novel strategy to improve the half-life of therapeutic
proteins. J Biotechnol. 253:23–33. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Guo R, Guo W, Cao L, Liu H, Liu J, Xu H,
Huang W, Wang F and Hong Z: Fusion of an albumin-binding domain
extends the half-life of immunotoxins. Int J Pharm. 511:538–549.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Adabi E, Saebi F, Moradi Hasan-Abad A,
Teimoori-Toolabi L and Kardar GA: Evaluation of an albumin-binding
domain protein fused to recombinant human il-2 and its effects on
the bioactivity and serum half-life of the cytokine. Iran Biomed J.
21:77–83. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jonckheere N, Vasseur R and Van Seuningen
I: The cornerstone K-RAS mutation in pancreatic adenocarcinoma:
From cell signaling network, target genes, biological processes to
therapeutic targeting. Crit Rev Oncol Hematol. 111:7–19. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schönleben F, Qiu W, Bruckman KC, Ciau NT,
Li X, Lauerman MH, Frucht H, Chabot JA, Allendorf JD, Remotti HE
and Su GH: BRAF and KRAS gene mutations in intraductal papillary
mucinous neoplasm/carcinoma (IPMN/IPMC) of the pancreas. Cancer
Lett. 249:242–248. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fitzgerald TL, Lertpiriyapong K, Cocco L,
Martelli AM, Libra M, Candido S, Montalto G, Cervello M, Steelman
L, Abrams SL and McCubrey JA: Roles of EGFR and KRAS and their
downstream signaling pathways in pancreatic cancer and pancreatic
cancer stem cells. Adv Biol Regul. 59:65–81. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Thayer SP, di Magliano MP, Heiser PW,
Nielsen CM, Roberts DJ, Lauwers GY, Qi YP, Gysin S, Fernández-del
Castillo C, Yajnik V, et al: Hedgehog is an early and late mediator
of pancreatic cancer tumorigenesis. Nature. 425:851–856. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Waters AM and Der CJ: KRAS: The critical
driver and therapeutic target for pancreatic cancer. Cold Spring
Harb Perspect Med. 8(pii): a0314352018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Commisso C, Davidson SM, Soydaner-Azeloglu
RG, Soydaner-Azeloglu RG, Parker SJ, Kamphorst JJ, Hackett S,
Grabocka E, Nofal M, Drebin JA, et al: Macropinocytosis of protein
is an amino acid supply route in Ras-transformed cells. Nature.
497:633–637. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang X, Sheng W, Wang Y, Li L, Li Y, Zhang
S, Liu X, Chen S and Zhen Y: A macropinocytosis-intensifying
albumin domain-based scfv antibody and its conjugate directed
against k-ras mutant pancreatic cancer. Mol Pharm. 15:2403–2412.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tai CJ, Huang MT, Wu CH, Wang CK, Tai CJ,
Chang CC, Hsieh CI, Chang YJ, Wu CJ, Kuo LJ, et al: Combination of
two targeted medications (bevacizumab plus cetuximab) improve the
therapeutic response of pancreatic carcinoma. Medicine (Baltimore).
95:e32592016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Moon do C, Lee HS, Lee YI, Chung MJ, Park
JY, Park SW, Song SY, Chung JB and Bang S: Concomitant statin use
has a favorable effect on gemcitabine-erlotinib combination
chemotherapy for advanced pancreatic cancer. Yonsei Med J.
57:1124–1130. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shao RG and Zhen YS: Enediyne anticancer
antibiotic lidamyin: Chemistry, biology and pharmacology.
Anticancer Agents Med Chem. 30:121–131. 2008.
|
|
25
|
Guo XF, Zhu XF, Shang Y, Zhang SH and Zhen
YS: A bispecific enediyne-energized fusion protein containing
ligand-based and antibody-based oligopeptides against epidermal
growth factor receptor and human epidermal growth factor receptor 2
shows potent antitumor activity. Clin Cancer Res. 16:2085–2094.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li L, Hu L, Zhao CY, Zhang SH, Wang R, Li
Y, Shao RG and Zhen YS: The recombinant and reconstituted novel
albumin-lidamycin conjugate shows lasting tumor imaging and
intensively enhanced therapeutic efficacy. Bioconjug Chem.
29:3104–3112. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sheng W, Shang Y, Li L and Zhen Y: An
EGFR/CD13 bispecific fusion protein and its enediyne-energized
analog show potent antitumor activity. Anticancer Drugs. 25:82–91.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang YH, Shang BY and Zhen YS: Antitumor
efficacy of lidamycin on hepatoma and active moiety of its
molecule. World J Gastroenterol. 11:3980–3984. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Van de Poll ML, van Vugt MJ, Lenferink AE
and van Zoelen EJ: Indentification of the minimal requirement for
binding to the human epidermal growth factor (EGF) receptor using
chimeras of human EGF and EGF repeat of Drosophila Notch. J Bio
Chem. 273:16075–16081. 1998. View Article : Google Scholar
|
|
30
|
Du Y, Shang BY, Sheng WJ, Zhang SH, Li Y,
Miao QF and Zhen YS: A recombinantly tailored β-defensin that
displays intensive macropinocytosis-mediated uptake exerting potent
efficacy against K-Ras mutant pancreatic cancer. Oncotarget.
7:58418–58434. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tanaka KI, Shimoda M, Chuang VTG, Nishida
K, Kawahara M, Ishida T, Otagiri M, Maruyama T and Ishima Y:
Thioredoxin-albumin fusion protein prevents copper enhanced
zinc-induced neurotoxicity via its antioxidative activity. Int J
Pharm. 535:140–147. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wan A, Miao Y, Peng L, Cai Y, Chen Y, He
Y, Yang J, Jin J and Li H: Binding and biologic characterization of
recombinant human serum albumin-eTGFBR2 fusion protein expressed in
CHO cells. Bioengineered. 8:600–612. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kruth HS, Jones NL, Huang W, Zhao B, Ishii
I, Chang J, Combs CA, Malide D and Zhang WY: Macropinocytosis is
the endocytic pathway that mediates macrophage foam cell formation
with native low density lipoprotein. J Biol Chem. 280:2352–2360.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sallusto F, Cella M, Danieli C and
Lanzavecchia A: Dendritic cells use macropinocytosis and the
mannose receptor to concentrate macromolecules in the major
histocompatibility complex class II compartment: Downregulation by
cytokines and bacterial products. J Exp Med. 182:389–400. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Recouvreux MV and Commisso C:
Macropinocytosis: A metabolic adaptation to nutrient stress in
cancer. Front Endocrinol (Lausanne). 8:2612017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nakase I, Niwa M, Takeuchi T, Sonomura K,
Kawabata N, Koike Y, Takehashi M, Tanaka S, Ueda K, Simpson JC, et
al: Cellular uptake of arginine-rich peptides: Roles for
macropinocytosis and actin rearrangement. Mol Ther. 10:1011–1022.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Veltman DM, Williams TD, Bloomfield G,
Chen BC, Betzig E, Insall RH and Kay RR: A plasma membrane template
for macropinocytic cups. Elife. 5(pii): e200852016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tisdale EJ, Shisheva A and Artalejo CR:
Overexpression of atypical protein kinase C in HeLa cells
facilitates macropinocytosis via Src activation. Cell Signal.
26:1235–1242. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cullis J, Siolas D, Avanzi A, Barui S,
Maitra A and Bar-Sagi D: Macropinocytosis of Nab-paclitaxel drives
macrophage activation in pancreatic cancer. Cancer Immunol Res.
5:182–190. 2017. View Article : Google Scholar : PubMed/NCBI
|