Introduction
In China, breast cancer is one of the leading causes
of cancer-related deaths in women. Systemic (i.e., neoadjuvant)
chemotherapy is currently an integral part of the treatment regime
for breast cancer (1). Due to the
heterogeneity of the disease and individual differences, 20–30%
patients subjected to neoadjuvant chemotherapy show primary
resistance to chemotherapeutic drugs, which could delay the timing
of local treatment and severely affect patient prognosis and
quality of life (2,3). Therefore, identification of markers
that facilitate rapid and easy prediction of sensitivity to
neoadjuvant chemotherapy is of significant value for evaluating the
individual choice of treatment for patients and clarifying the
mechanisms underlying resistance to chemotherapy.
Taxane is a traditional cytotoxic antitumor drug
that mainly promotes polymerization of tubulin and inhibits spindle
motility (4). In addition, studies
have shown that taxanes affect variouc non-mitotic functions of the
microtubules, such as molecular trafficking, signaling, and could
kill interphase cancer cells (5–7).
Taxane-based combination chemotherapy is generally the first choice
for adjuvant and neoadjuvant chemotherapy of breast cancer patients
(8,9). However, many patients show continued
resistance to taxane-based chemotherapy. Multiple molecular players
are linked to taxane resistance; these include efflux pumps
(10), DNA repair mechanisms
(11), and microtubule alterations
(12). In addition, emerging
evidence indicates that both non-coding RNAs and epigenetic
effectors may also be implicated in taxane resistance (7). Using the Gene Expression Omnibus (GEO)
online database, G-protein signaling modulator 2 (GPSM2) was
identified as a molecule significantly associated with taxane
sensitivity. GPSM2 (also known as LGN) was initially shown to
regulate proper positioning of the cell spindle, maintain symmetry
of cell division, and play an important role in the G2/M phase
involved in cell division (13–15).
He and co-researchers demonstrated that GPSM2 is overexpressed in
hepatocellular carcinoma and its expression levels are related to
clinical characteristics, supporting a potential function as a
tumor-suppressor gene (16).
Overexpression of GPSM2 in breast cancer tissues was reported by
Fukukawa et al (15).
However, the possible involvement of GPSM2 in paclitaxel
sensitivity has not been examined to date.
In the present study, we performed
bioinformatics-based screening of the paclitaxel resistance-related
gene, GPSM2, which was further confirmed in clinical specimens as
well as in vitro and in vivo experiments. Our
collective findings indicate that GPSM2 plays an important role in
paclitaxel resistance and may thus serve as a marker of paclitaxel
sensitivity in breast cancer.
Materials and methods
Bioinformatic analysis
Using the Gene Expression Omnibus database (GEO;
http://www.ncbi.nlm.nih.gov/geo/) of the
National Center for Biotechnology Information (NCBI) (17), raw gene expression profiles and
clinical data available for breast cancer were downloaded from
GSE25055 (18), GSE25065 (18) and GSE41998 (19), and data on patients receiving
paclitaxel (PTX)-based neoadjuvant chemotherapy (NAC) were selected
for further analyses. Univariate logistic regression (LR) was
performed with the gene expression level as the independent
variable and pathological complete response (pCR) status as
covariates. Multiple hypothesis testing was applied on the P-value
of LR for each gene, and genes with FDR q<0.25 were identified
as pCR-related. A Venn diagram was further constructed to identify
pCR-related genes shared by the three datasets.
Genomic and drug sensitivity data on the NCI60 cell
line was downloaded and set into the correlation analysis between
each pCR-related gene with sensitivity to PTX (20). Based on the spectrum of the
correlation coefficient calculated from two non-standard
correlations of PTX (NSC125973 and NSC758645), the first 20 genes
with higher correlation coefficients were collected into two
ranking systems. The importance of each gene was quantified based
on the sum of its ranking scores in the two-ranking system, and the
gene with the lower value was identified as more significant.
Publicly available GI50 [-log10
(IC50), molar drug concentration for 50% growth
inhibition] data on PTX (NSC125973 and NSC758645) and genomic data
on the NCI60 cell line were obtained via the rcellminer R package
(20). In total, 5 breast cancer
(MCF7, MDA-MB-231, HS578T, BT-549 and T47D) and 7 ovarian cancer
(SK-OV-3, IGROV1, OVCAR-3, OVCAR-4 and OVCAR-8) cell lines were
included in the analysis. Spearman correlation was performed to
confirm the correlation coefficients (r-value) between
GI50 and GPSM2. The ranking lists included genes with
the 20 top highest r-values for each drug (NSC125973 or NSC758645).
The summed ranking of an overlapped gene in the two ranking lists
was calculated, with lower values indicating higher importance of
that gene.
Gene set enrichment analysis (GSEA) was performed
using the JAVA program (http://www.broadinstitute.org/gsea) with GSE25055,
GSE25065 or GSE41998. The MSigDB H: hallmark gene set (50
available) and C2 CP: KEGG gene set (186 available) collections
were functional gene sets (21).
Expression of GPSM2 was set to annotate phenotypes. Gene sets with
a FDR value <0.25 were considered significantly enriched. The
overlapping significant gene sets among these three data sets were
taken as enriched gene sets.
Clinical breast cancer samples
A total of 85 invasive ductal cancer (IDC) specimens
of patients undergoing core biopsy were obtained between January
2011 and December 2014 at Shengjing Hospital of the China Medical
University (Shenyang, Liaoning, China). Demographic and clinical
characteristics, such as age, sex, and stage at diagnosis, were
collected. This study was approved by the Ethics Committee of the
China Medical University, and all patients signed informed consent,
which was consistent with the Declaration of Helsinki. All patients
underwent neoadjuvant chemotherapy and treatment with 2–3 cycles of
the PTX regimen at Shengjing Hospital, Shengjing, China. Based on
different responses to PTX, the patients were divided into four
groups: Complete remission (CR) group, where the tumor disappeared
and remained absent for at least 4 weeks; partial remission (PR),
where the longest diameter of the tumor was reduced by >30% or
the sum of tumor diameter was reduced more than 50% and maintained
more than 4 weeks; progressive disease (PD), whereby the largest
diameter of the tumor increased by >20% or the sum of the tumor
diameter increased by >25%; and stable disease (SD), a stage
between PR and PD, established following at least 2 cycles of
chemotherapy; CR + PR are proportional to efficiency. In a total of
85 patients, 2 patients were evaluated as having PD, 23 patients
were evaluated as presenting with SD, 2 patients were evaluated as
having CR, and 58 patients were evaluated with PR. PD and SD groups
were determined as exhibiting resistance to PTX; CR and PR group
were determined as being sensitive to PTX.
Cell lines and cultures
The human breast cancer cell lines MDA-MB-231, T47D
and MCF-7 were purchased from the Cell Bank of the Type Culture
Collection of the Chinese Academy (Shanghai, China). MDA-MB-231
cells were grown in L15 (Invitrogen; Thermo Fisher Scientific,
Inc.). MCF-7 and T47D cells were grown in Dulbecco's modified
Eagle's medium (DMEM; Invitrogen; Thermo Fisher Scientific, Inc.).
All human breast cancer cell lines were supplemented with 10% fetal
bovine serum (FBS) and maintained at 37°C with 5%
CO2.
Immunocytochemical staining
Paraffin sections were deparaffinized with xylene
and autoclaved with sodium citrate, pH 6.0. Sections were incubated
with the primary antibody (anti-human rabbit GPSM2; cat. no.
ab84571, Abcam) at a dilution of 1:100 overnight at 4°C and the
secondary antibodies (1:200 diluted biotinylated goat-anti-rabbit
solution) (Vector Laboratories, Inc.; cat. no. PK-4001) for 30 min
on the following day. Next, sections were stained using
3,3′-diaminobenzidine (DAB) for microscopic observation and
counterstained with hematoxylin. A semi-quantitative scoring system
was adopted. The percentage of positively stained cells was scored
as follows: 0, <10%; 1, 10–25%; 2, 26–50%; 3, 51–75%; and 4,
>75%. Intensity was scored as ‘−’ (negative), ‘+’ (weak), ‘++’
(moderate) and ‘+++’ (strong). The IHC score was the percentage
score multiplied by the staining intensity score. IHC scores ≥4
were indicative of a high expression of GPSM2.
RT-qPCR
Reverse transcription polymerase chain reaction
(RT-PCR) array was used to analyze GPSM2 expression or efficiency
of transfection. RT-qPCR was performed according to our previously
reported method (22). The primer
sequences for amplification were: Forward
(5′-AGCCAGTCGGTACTTAGCCA-3′) and reverse
(5′-TTGTGGTAGCAGGTGGTGGA-3′) for GPSM2; and forward
(5′-GGTGAAGGTCGGAGTCAACGG-3′) and reverse
(5′-GAGGTCAATGAAGGGGTCATTG-3′) for the 18S control. PCR conditions
were as follows: 95°C for 5 min, 95°C for 15 sec, 62.8°C for 15
sec, 72°C for 20 sec and 72°C for 5 min for 35 cycles. Relative
expression of mRNA was calculated via the comparative cycle
threshold method using 18S as a reference.
Western blot analysis
Cells were extracted and proteins was lysed in RIPA
lysis buffer. Equivalent amounts (40 µg) of protein were isolated
by 10% SDS-PAGE (Bio-Rad) and transferred to polyvinylidene
fluoride (PVDF) membranes (Millipore). Membranes were blocked with
5% non-fat milk for 40 min and incubated with primary antibodies:
Anti-GPSM2 (dilution 1:1,000, Abcam) and anti-β-actin (cat. no.
sc-47778; dilution 1:1,000; Santa Cruz Biotechnology) overnight at
4°C. Following further washing in TTBS, membranes were incubated
with secondary antibodies for 40 min. After extensive washing,
chemiluminescence was detected using the ECL kit (Millipore). Image
J 6.0 software (NIH, National Institutes of Health, Bethesda, MD,
USA) was used to analyze the images.
Cell proliferation assay
Cell viability was determined by
1-(4,5-demethylthiazol-2-yl)-3,5-diphenyltetrazolium bromide (MTT)
assay. Cells were cultured in 96-well plates and treated with
different concentrations of PTX for 48 h. After incubation with MTT
(0.5 mg/ml) for 4 h, the supernatant was discarded and 200 µl DMSO
was added to each well. The absorbance at 570 nm was detected
spectrophotometrically.
Cell transfection and treatments
Cell transfection was performed according to the
small interfering RNA (siRNA) sequence transfection protocol for
Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.).
The sequence of the GPSM2-specific siRNA was
5′-GGUAAUCUGGGAAACACCU-3′. Nonsense RNAi
(5′-UUCUCCGAACGUGUCACGUTT-3′) was used as a negative control (NC).
At 48 h after transfection, the transfection efficiency of GPSM2
was identified by RT-qPCR and western blot analyses.
Colony formation assay
For the colony formation assay, transfected
MDA-MB-231 cells were harvested and seeded at a density of 500
cells per well in 12-well plates at 37°C and 5% CO2 in a
humidified incubator for 2 weeks. Results were determined and
images obtained under a microscope (magnification ×100) following
Giemsa staining. The colony numbers in each well were
calculated.
Transfection of the GPSM2 WT
plasmid
GPSM2 wild-type (WT) plasmid and control plasmid
were purchased from Genechem (Shanghai, China). Cells were seeded
at 4×105 cells per well in 6-well plates overnight and
transfected by Lipofectamine 2000 reagent with the GPSM2 WT
plasmid. Forty-eight hours after transfection, GPSM2 expression was
evaluated using western blot analysis.
Cell cycle assay
Propidium iodide (PI) staining was used to detect
the cell cycle. Cells in the log phase were trypsinized,
centrifuged at 300 × g for 5 min, and washed with PBS. Next, the
sediment was suspended with 400 ml PBS with 50 mg/ml PI and 100
mg/ml RNase at 37°C for 30 min. Flow cytometry (BD AccuriTM C6 Flow
cytometer) was used to determine the percentages of cells in the
G1, S, and G2 phases, and the data were analyzed by WinMDI version
software (http://www.cyto.purdue.edu/flowcyt/software/Winmdi.htm).
Generation of stable GPSM2-knockdown
cell line
GPSM2 shRNA lentiviral particles were obtained from
Genechem Technology Corp., Ltd. (Shanghai, China). MDA-MB-231 cells
were infected with GPSM2 shRNA lentiviral particles following the
manufacturer's instructions.
Animal experiments
Male athymic nude mice (BALB/c-nu/nu) (4–6 weeks
old, 18–24 g) were purchased from Shanghai Slike Experimental
Animals Co. (Shanghai, China) and housed under specific
pathogen-free conditions. The maintenance conditions for the mice
were as follows: temperature, 23–27°C; humidity, 40–60%;
ventilation, 15 times/h; 12 h light/12 h dark cycle. The mice were
fed standard laboratory food and water. MDA-MB-231 cells
(1×107) was directly injected into the exposed no. 4
inguinal mammary pad of each mouse. Tumor volume was calculated as:
Volume = (width2 × length)/2. Paclitaxel (PTX) (10 mg/kg
once every 3 days) was administered via i.p. injection starting
from the time the tumor volume reached ~30 mm3. The
health and maintenance condition of the mice were monitored every 3
days. After 2 weeks, the mice were sacrificed by cervical
dislocation and the tumors were removed and weighed. The maximum
diameter of a single tumor found was 12 mm and no mouse developed
multiple tumors. All mice were in a good body condition throughout
the experiment. All animal experiments conformed to the Guide for
Care and Use of Laboratory Animals and were approved by the Animal
Care and Use Committee of China Medical University.
Statistical analysis
Statistical significance was examined using the
Student's t test using the SPSS 13.0 software package (SPSS, Inc.).
Growth cell data were compared using one-way analysis of variance
(ANOVA) and Tukey's multiple comparisons test. Correlations between
two variables were evaluated with Spearman's rank correlation
coefficient. For all statistical analyses, the level of
significance was set at P<0.05.
Results
Online database analysis to screen the
relevant genes closely related to paclitaxel primary drug
resistance
A total of 306, 182 and 121 patients were included
in the GSE25055 (18), GSE25065
(18) and GSE41998 (19) datasets, respectively. Logistic
regression analyses showed that 5,104, 1,823 and 991 genes were
statistically significant (P<0.05, FDR <0.25) in the three
respective datasets. In the Venn plot, 410 genes were shared by the
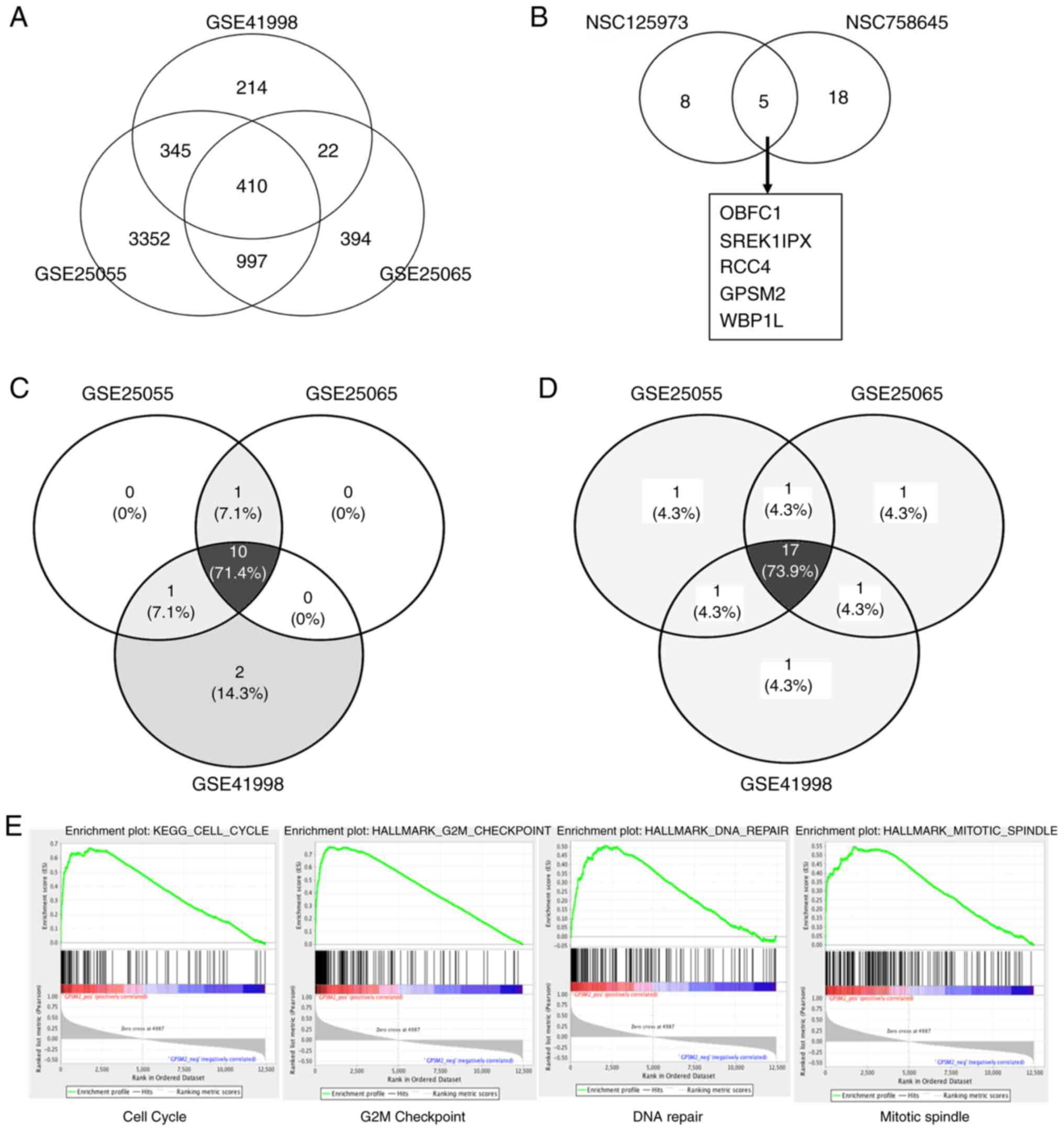
three datasets (Fig. 1A), which
were pCR-related. In NCI-60 breast and ovarian cancer datasets, the
correlation of each pCR-related gene with drug sensitivity to
NSC125973 and NSC758645 was ranked. Genes with r-values >0.4 in
NSC125973 and NSC758645 are depicted in Table I. GPSM2 was determined to be
an overlapped gene (Fig. 1B). To
further investigate the pathways regulated by GPSM2, gene set
enrichment analysis was applied. Ten gene sets showed significant
GPSM2 high-expression phenotypes when hallmarks were used as the
reference in this step (Fig. 1C).
Seventeen gene sets showed significant GPSM2 high-expression
phenotype when Kyoto Encyclopedia of Genes and Genomes (KEGG)
signaling pathways were used as the reference (Fig. 1D). The overlapping gene sets are
listed in Fig. 1E. Cell
cycle-related gene sets (‘HALLMARK_G2M_CHECKPOINT’,
‘HALLMARK_MITOTIC_SPINDLE’, ‘KEGG_CELL_CYCLE’ and
‘KEGG_DNA_REPLICATION’, among others) were significantly associated
with the GPSM2 high-expression phenotype.
 | Table I.Genes with r-values >0.4 in
NSC125973 and NSC758645. |
Table I.
Genes with r-values >0.4 in
NSC125973 and NSC758645.
|
| NSC125973 | NSC75864 |
|---|
|
|
|
|
|---|
| Gene symbol | r-value | Gene symbol | r-value |
|---|
| MEX3C | 0.661 | USP13 | 0.757 |
| WASF1 | 0.61 |
SREK1IP1 | 0.658 |
| LMNB2 | 0.556 | WBP1L | 0.618 |
| OBFC1 | 0.516 | NFIB | 0.609 |
| NUP160 | 0.516 | TCTN1 | 0.577 |
| LRPAP1 | 0.506 | ARL3 | 0.571 |
| XRCC4 | 0.459 | ACTL6A | 0.556 |
| WBP1L | 0.454 | XRCC4 | 0.538 |
| LSM6 | 0.444 | SLC39A6 | 0.521 |
|
SREK1IP1 | 0.43 | AKAP10 | 0.508 |
| IPO5 | 0.428 | TSPAN31 | 0.497 |
| GPSM2 | 0.427 | ZER1 | 0.494 |
| CDC40 | 0.414 | OBFC1 | 0.49 |
|
|
| SMC4 | 0.479 |
|
|
| HAGH | 0.475 |
|
|
| H2AFY2 | 0.435 |
|
|
| AFF3 | 0.426 |
|
|
| ABCF1 | 0.418 |
|
|
| HIF1AN | 0.417 |
|
|
| PIGQ | 0.415 |
|
|
| MLPH | 0.409 |
|
|
| GPSM2 | 0.402 |
|
|
| CCNE2 | 0.401 |
GPSM2 is significantly correlated with
paclitaxel sensitivity in breast cancer patients
To confirm the clinical significance of GPSM2 in
breast cancer, we selected 85 patients undergoing neoadjuvant
chemotherapy and performed immunohistochemical analysis of core
biopsy pathological sections. Representative examples of each level
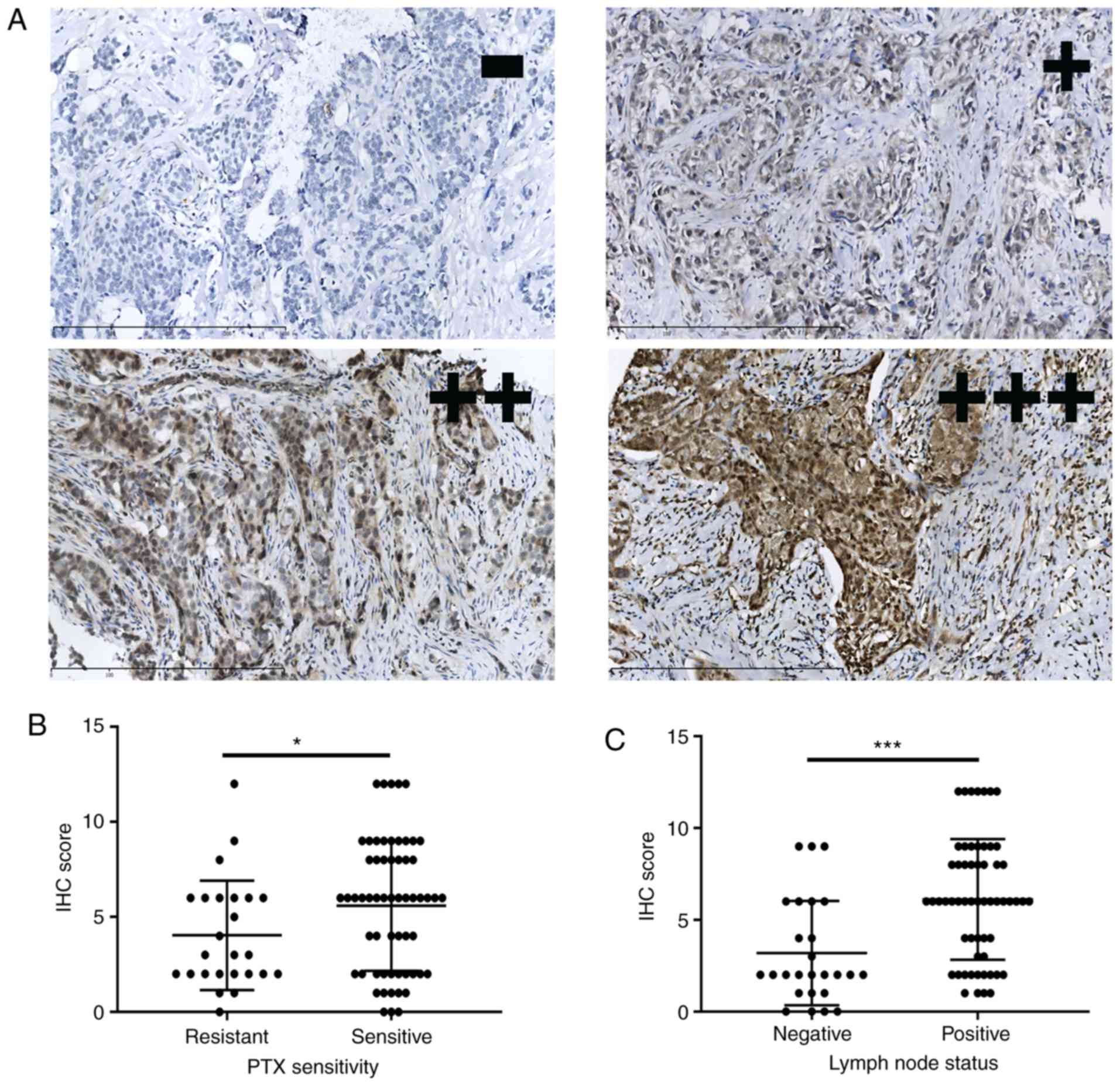
of staining are shown in Fig. 2A.
The GPSM2 staining score was significantly higher in the
PTX-sensitive patients (Fig. 2B).
Moreover, the staining score was significantly increased in
patients with a positive lymph node status (Fig. 2C). Based on the IHC results,
staining was quantified and classified into positive and negative
groups. Data from the Chi-square test showed significant
correlations between GPSM2 expression and lymph node status
(P=0.001), TNM stage (P=0.016) and PTX sensitivity (P=0.015)
(Table II). The IHC score of GPSM2
was higher in advanced-stage cancer patients (clinical stage
III–IV) than in early-stage cancer patients (clinical stage I–II)
(6.381±3.450, n=42 vs. 4.093±3.030, n=43, respectively, P=0.002).
In additional, the rate of PTX-resistance in advanced cancer was
13/42 (30.95%), while the rate of PTX-resistance in early-stage
cancer was 12/43 (27.90%). Chi-square test indicated that the
resistance of PTX had no significance associated with TNM stage
(P=0.758). Based on our results, advanced-stage cancer showed
higher GPSM2 scores, but was not associated with better clinical
benefit.
| Table II.Correlations between GPSM2 expression
and clinical characteristics of the breast cancer patients in the
present study (N=85). |
Table II.
Correlations between GPSM2 expression
and clinical characteristics of the breast cancer patients in the
present study (N=85).
|
|
| GPSM2
expression |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | Cases (n) | Positive n (%) | Negative n (%) | P-value |
|---|
| Age (years) |
|
|
| 0.377 |
|
≤45 | 27 | 19 (70.37) | 8 (26.63) |
|
|
>45 | 58 | 35 (60.34) | 23 (39.66) |
|
| Clinical stage |
|
|
| 0.016a |
|
I–II | 43 | 22 (51.16) | 21 (48.84) |
|
|
III–IV | 42 | 32 (76.19) | 10 (23.81) |
|
| Lymph node
status |
|
|
| 0.001a |
|
Negative | 26 | 10 (38.46) | 16 (61.54) |
|
|
Positive | 59 | 44 (74.58) | 15 (25.42) |
|
| Tumor size
(cm) |
|
|
| 0.474 |
|
<5 | 59 | 36 (61.01) | 23 (38.98) |
|
| ≥5 | 26 | 18 (69.23) | 8 (30.77) |
|
| ER |
|
|
| 0.349 |
|
Negative | 31 | 22 (70.97) | 9 (29.03) |
|
|
Positive | 54 | 33 (61.11) | 21 (38.89) |
|
| PR |
|
|
| 0.547 |
|
Negative | 31 | 22 (70.97) | 9 (29.03) |
|
|
Positive | 54 | 34 (62.96) | 22 (40.74) |
|
| Her2 |
|
|
| 0.138 |
|
Negative | 42 | 30 (71.43) | 12 (28.57) |
|
|
Positive | 43 | 24 (55.81) | 19 (44.19) |
|
| PTX
sensitivity |
|
|
| 0.015a |
|
Resistant | 25 | 11 (44.00) | 14 (56.00) |
|
|
Sensitive | 60 | 43 (71.67) | 17 (28.33) |
|
Knockdown of GPSM2 expression reduces
the sensitivity of breast cancer cells to paclitaxel
Next, we determined the mRNA and protein levels of
GPSM2 in three breast cancer cell lines (MDA-MB-231, T47D, MCF7).
Interestingly, MDA-MB-231 cells expressed considerably higher
levels of GPSM2 than the other cell lines (Fig. 3A and B). Cell proliferation assays
were performed after the cells were treated with different
concentrations of PTX (0, 0.1, 0.5, 1, 5 and 10 µg/ml). MDA-MB-231
cells with relatively high GPSM2 expression tended to have
significantly higher sensitivity values (Fig. 3C). Accordingly, this cell line was
selected for further investigation. The simplicity of small
interfering RNA (siRNA) manufacturing and transient nature of the
effect per dose are suited for cell assay (23). Thus, to ascertain the involvement of
GPSM2 in PTX sensitivity, knockdown of GPSM2 expression was
performed using specific siRNAs, and transfection efficiency was
assessed via RT-qPCR (Fig. 3D) and
western blot analyses (Fig. 3E).
Notably, depletion of GPSM2 led to lower PTX sensitivity in the
MDA-MB-231 cells (Fig. 3F).
Moreover, the GPSM2 siRNA-transfected groups displayed more
colonies than those transfected with NC-siRNA after PTX treatment
(Fig. 3G). MCF-7 cells were
selected for the subsequent experiments because of the low
expression level of GPSM2. Overexpression of GPSM2 was performed
using the GPSM2-WT plasmid, and transfection efficiency was
assessed via western blot analyses (Fig. 3H). Notably, overexpression of GPSM2
led to significantly higher PTX sensitivity in MCF-7 cells
(Fig. 3I). Moreover, GPSM2-WT
plasmid transfected groups displayed less colonies than those
transfected with the empty vector plasmid after PTX treatment
(Fig. 3J). PTX-treated cells are
known to undergo G2 phase arrest. Further GSEA analyses revealed
that GPSM2 is significantly correlated with cell cycle-related gene
sets. Accordingly, we assessed whether GPSM2 affects the activity
of PTX in triggering G2 phase arrest in breast cancer cells.
Knockdown of GPSM2 clearly reduced PTX-induced G2 phase arrest
(Fig. 3K). PTX induced cell cycle
arrest of 12.92±0.510% of cells in the G2 phase, whereas only
5.667±0.667 of cells were arrested in the G2 phase after knockdown
of GPSM2.
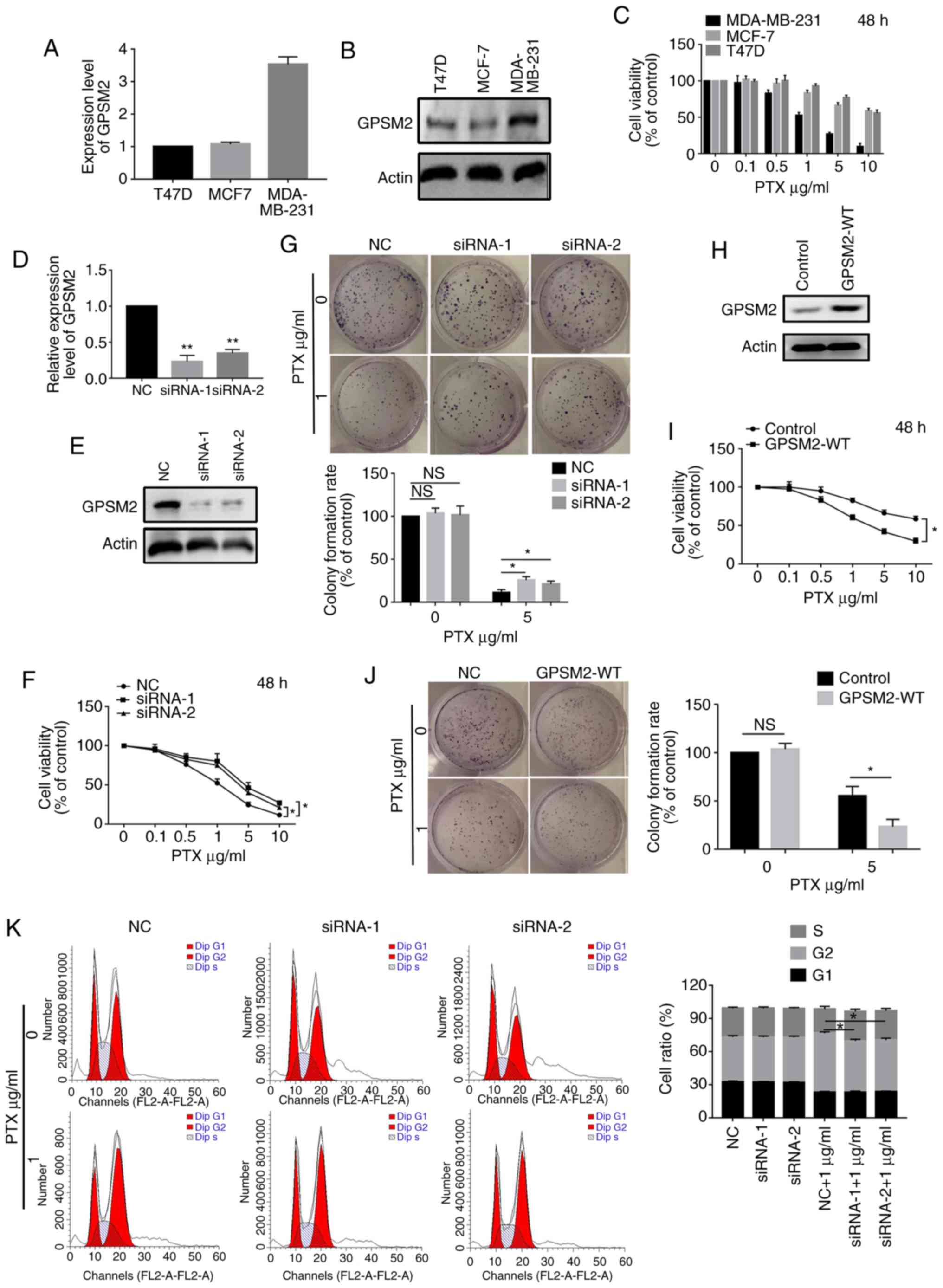 | Figure 3.Knockdown of GPSM2 increases
resistance to PTX in breast cancer MDA-MB-231 cells. (A and B)
GPSM2 mRNA and protein expression in three breast cancer cell lines
(T47D, MCF7 and MDA-MB-231). (C) Cell proliferation assays were
performed following treatment with different concentrations (0,
0.1, 0.5, 1, 5 and 10 µg/ml) of PTX in the three breast cancer cell
lines. (D and E) MDA-MB-231 cells were transfected with either
GPSM2-siRNA or NC-siRNA and GPSM2 mRNA and protein expression
levels were detected via RT-qPCR and western blot analyses,
respectively. **P<0.01 vs. the NC group. (F) Cell viability
assay showing that GPSM2 knockdown increased the resistance of
MDA-MB-231 cells to PTX. *P<0.05. (G) After treatment with PTX,
the number of colonies formed by MDA-MB-231 cells transfected with
NC siRNA was lower than those formed by cells transfected with
GPSM2 siRNA. *P<0.05; ns, not significant. (H) MDA-MB-231 cells
were transfected with either GPSM2-WT or empty vector plasmid and
GPSM2 protein expression was detected via western blot analyses.
(I) Cell viability assay showing that GPSM2 overexpression
decreased the resistance of MDA-MB-231 cells to PTX. *P<0.05.
(J) After treatment with PTX, the number of colonies formed by
MDA-MB-231 cells transfected with the empty vector plasmid was
higher than the number formed by cells transfected with the
GPSM2-WT plasmid. *P<0.05; ns, not significant. (K) GPSM2
affects the activity of PTX to trigger G2 phase arrest in breast
cancer. GPSM2 knockdown reduced paclitaxel-induced G2 phase arrest.
*P<0.05. PTX, paclitaxel; GPSM2, G protein signaling modulator
2; NC, negative control. |
GPSM2 influences paclitaxel
sensitivity in vivo
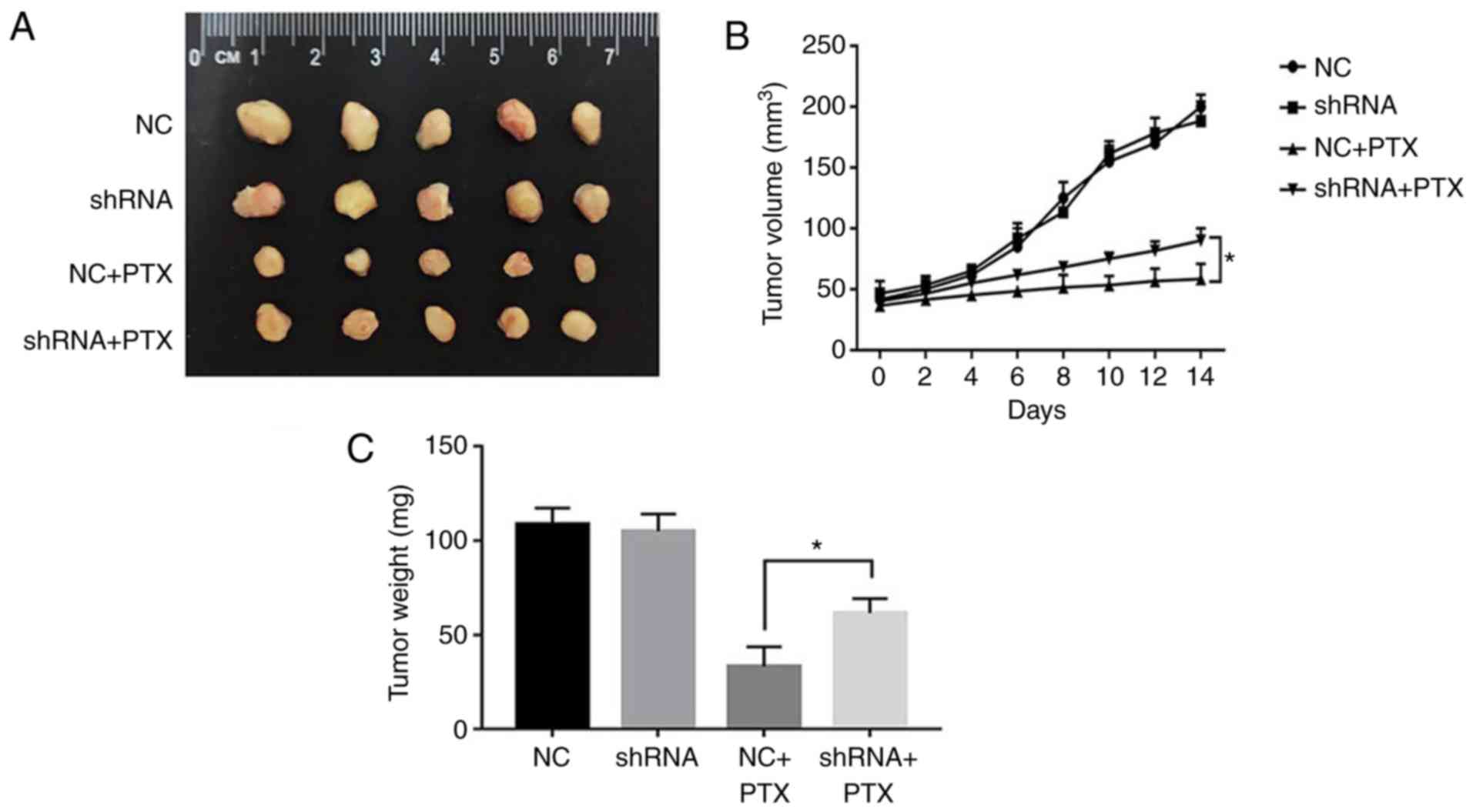
To further ascertain the potential effects of GPSM2
on paclitaxel sensitivity, we used an orthotopic implantation mouse
model of MDA-MB-231 cells. The orthotopic implantation mouse model
lasted 14 days. Sustainable characteristic of shRNA could provide
long-lasting knockdown effect (21). Thus, stable GPSM2-knockdown cells
were generated using shRNA. Compared to the mice implanted with NC
shRNA-transfected MDA-MB-231 cells, no significant differences in
tumor growth were observed in the GPSM2 shRNA-transfected group.
However, differences were evident upon treatment of both groups
with PTX. Specifically, following PTX treatment, the tumor volumes
of mice in the GPSM2 shRNA group were significantly increased,
compared with those in the NC shRNA group (P<0.05) (Fig. 4A and B). Moreover, under conditions
of knockdown of GPSM2, sensitivity of breast cancer cells to PTX
was reduced and tumor weights were higher (Fig. 4C).
Discussion
Breast cancer is the most common malignancy in women
worldwide (1). Neoadjuvant and
conventional postoperative chemotherapy strategies have been
established as the most successful treatments to date, among which
Taxol is one of the most effective drugs (2,24).
Owing to disease heterogeneity and individual differences,
approximately 20–30% patients subjected to neoadjuvant chemotherapy
are resistant to paclitaxel (25).
Thus, identification of markers that can rapidly and easily predict
sensitivity to neoadjuvant chemotherapy is currently an urgent
medical requirement.
Based on clinical information and gene expression
profiles extracted from the Gene Expression Omnibus (GEO)
(GSE25055, GSE25065 and GSE41998) database, we observed
significance differences in G-protein signaling modulator 2 (GPSM2)
expression in correlation with paclitaxel resistance. This finding
was further validated with NSC125973, NSC758645 and GSEA analyses.
GPSM2 plays an important role in controlling spindle orientation
and positioning in the context of asymmetric cell division
(26,27). Bioinformatics analyses in our study
support a novel relationship between GPSM2 and paclitaxel
resistance in breast cancer, which provides a guiding framework for
subsequent experiments.
GPSM2 has also been shown to participate in
neuroblast division and polarization of the apical complex.
Expression of GPSM2 is closely related to progression of
hepatocellular carcinoma and renal cell carcinoma (28). A previous study reported an
association of GPSM2 overexpression with occurrence of breast
cancer (15). However, its
potential involvement in paclitaxel resistance in breast cancer
patients is yet to be established. To the best of our knowledge,
this is the first study to demonstrate that GPSM2 is widely
downregulated in tissue samples of paclitaxel-resistant breast
cancer patients and is correlated with lymph node status and
clinicopathological parameters. In view of our experimental
results, we propose that GPSM2 may serve as an effective indicator
of paclitaxel drug sensitivity in breast cancer patients.
Drug resistance in tumor cells is an important
contributory factor to disease progression (29). Here, we examined the potential role
of GPSM2 expression in breast cancer by silencing its expression.
In cell viability and colony formation assays, depletion of GPSM2
expression in MDA-MB-231 cells led to increased resistance to
paclitaxel, supporting the hypothesis that GPSM2 is a potential
drug-resistance gene in breast cancer. In view of the GSEA
enrichment data showing that GPSM2 is related to the cell cycle, a
flow cytometry experiment was conducted, which revealed a decrease
in paclitaxel-induced G2 arrest of MDA-MB-231 cells following the
silencing of GPSM2. G2 phase arrest is an important mechanism for
the paclitaxel-mediated effect on cell proliferation (5). Paclitaxel, as a microtubule
stabilizing agent, induces cell cycle arrest in the G2 phase and
produces abnormal mitotic spindles (30). This suggests that G2 phase arrest
and microtubule regulation may be the potential mechanisms of
GPSM2. The association between GPSM2 and paclitaxel resistance was
further validated through in vivo experiments in nude mice.
Several studies have shown that GPSM2 acts as a guanylate
dissociation inhibitor (GDI) to inhibit the release of Gα from GDP,
resulting in a decrease in the availability of Gα-GTP (31). In addition, paclitaxel was found to
reverse the effect of GTP hydrolysis, preventing microtubules from
depolymerizing (32,33). The loss of GPSM2 may increase the
Gα-GTP level, and then the ability of paclitaxel to induce cycle
arrest by inhibiting GTP activity may decline, leading to drug
resistance in patients.
Our preliminary results clearly indicate that GPSM2
is closely related to drug resistance in breast cancer. However, it
should be noted that samples in this study were acquired
retrospectively. Further comprehensive analyses are necessary to
validate the clinical applicability of GPSM2 as a potential
biomarker.
Data from the current study support a critical role
of GPSM2 in breast cancer resistance. We propose that GPSM2 may
serve as a potential target for improving drug susceptibility in
patients as well as a marker of paclitaxel sensitivity.
Acknowledgements
Not applicable.
Funding
This study was supported by Shenyang Special Funds
of Technology Innovation Plan Projects (no. F14-231-1-46), Liaoning
Science and Technology Plan Projects (no. 2013225021) and Liaoning
Natural Science Foundation of 2018 (no. 20180530050).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
QL and YZ conceived and designed the experiments.
ZZhang, ZL, MD, BL, XX and ZZhao performed all the experiments.
ZZhang, ZL and MD analyzed the data. ZZhang and MD wrote the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All of the animal experiments conformed to the Guide
for Care and Use of Laboratory Animals and were approved by the
Animal Care and Use Committee of China Medical University (No.
2018113). All patients signed informed consent, which was
consistent with the Declaration of Helsinki.
Patient consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
References
|
1
|
Harbeck N and Gnant M: Breast cancer.
Lancet. 389:1134–1150. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pondé NF, Zardavas D and Piccart M:
Progress in adjuvant systemic therapy for breast cancer. Nat Rev
Clin Oncol. 16:27–44. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
DeMichele A, Yee D and Esserman L:
Mechanisms of resistance to neoadjuvant chemotherapy in breast
cancer. N Engl J Med. 377:2287–2289. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jordan MA and Wilson L: Microtubules as a
target for anticancer drugs. Nat Rev Cancer. 4:253–265. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Weaver BA: How Taxol/paclitaxel kills
cancer cells. Mol Biol Cell. 25:2677–2681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Herbst RS and Khuri FR: Mode of action of
docetaxel-a basis for combination with novel anticancer agents.
Cancer Treat Rev. 29:407–415. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Pucci P, Rescigno P, Sumanasuriya S, de
Bono J and Crea F: Hypoxia and noncoding rnas in taxane resistance.
Trends Pharmacol Sci. 39:695–709. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Palmieri C, Krell J, James CR,
Harper-Wynne C, Misra V, Cleator S and Miles D: Rechallenging with
anthracyclines and taxanes in metastatic breast cancer. Nat Rev
Clin Oncol. 7:561–574. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Turner N, Biganzoli L and Di Leo A:
Continued value of adjuvant anthracyclines as treatment for early
breast cancer. Lancet Oncol. 16:e362–e369. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vaidyanathan A, Sawers L, Gannon AL,
Chakravarty P, Scott AL, Bray SE, Ferguson MJ and Smith G: ABCB1
(MDR1) induction defines a common resistance mechanism in
paclitaxel- and olaparib-resistant ovarian cancer cells. Br J
Cancer. 115:431–441. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Poruchynsky MS, Komlodi-Pasztor E, Trostel
S, Wilkerson J, Regairaz M, Pommier Y, Zhang X, Kumar Maity T,
Robey R, Burotto M, et al: Microtubule-targeting agents augment the
toxicity of DNA-damaging agents by disrupting intracellular
trafficking of DNA repair proteins. Proc Natl Acad Sci USA.
112:1571–1576. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yang J, Yu Y, Liu W, Li Z, Wei Z and Jiang
R: Microtubule-associated protein tau is associated with the
resistance to docetaxel in prostate cancer cell lines. Res Rep
Urol. 9:71–77. 2017.PubMed/NCBI
|
|
13
|
Du Q, Taylor L, Compton DA and Macara IG:
LGN blocks the ability of NuMA to bind and stabilize microtubules.
A mechanism for mitotic spindle assembly regulation. Curr Biol.
12:1928–1933. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Mochizuki N, Cho G, Wen B and Insel PA:
Identification and cDNA cloning of a novel human mosaic protein,
LGN, based on interaction with G alpha i2. Gene. 181:39–43. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Fukukawa C, Ueda K, Nishidate T, Katagiri
T and Nakamura Y: Critical roles of LGN/GPSM2 phosphorylation by
PBK/TOPK in cell division of breast cancer cells. Genes Chromosomes
Cancer. 49:861–872. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
He XQ, Zhang YF, Yu JJ, Gan YY, Han NN,
Zhang MX, Ge W, Deng JJ, Zheng YF and Xu XM: High expression of
G-protein signaling modulator 2 in hepatocellular carcinoma
facilitates tumor growth and metastasis by activating the PI3K/AKT
signaling pathway. Tumour Biol. 39:1–10. 2017. View Article : Google Scholar
|
|
17
|
Barrett T, Troup DB, Wilhite SE, Ledoux P,
Rudnev D, Evangelista C, Kim IF, Soboleva A, Tomashevsky M,
Marshall KA, et al: NCBI GEO: Archive for high-throughput
functional genomic data. Nucleic Acids Res. 37:D885–D890. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hatzis C, Pusztai L, Valero V, Booser DJ,
Esserman L, Lluch A, Vidaurre T, Holmes F, Souchon E, Wang H, et
al: A genomic predictor of response and survival following
taxane-anthracycline chemotherapy for invasive breast cancer. JAMA.
305:1873–1881. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Horak CE, Pusztai L, Xing G, Trifan OC,
Saura C, Tseng LM, Chan S, Welcher R and Liu D: Biomarker analysis
of neoadjuvant doxorubicin/cyclophosphamide followed by ixabepilone
or paclitaxel in early-stage breast cancer. Clin Cancer Res.
19:1587–1595. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shoemaker RH: The NCI60 human tumour cell
line anticancer drug screen. Nat Rev Cancer. 6:813–823. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Deng M, Yu R, Wang S, Zhang Y, Li Z, Song
H, Liu B, Xu L, Wang X, Zhang Z, et al: Limb-bud and heart
attenuates growth and invasion of human lung adenocarcinoma cells
and predicts survival outcome. Cell Physiol Biochem. 47:223–234.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Rao DD, Vorhies JS, Senzer N and
Nemunaitis J: siRNA vs. shRNA: Similarities and differences. Adv
Drug Deliv Rev. 61:746–759. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jasra S and Anampa J: Anthracycline use
for early stage breast cancer in the modern era: A review. Curr
Treat Options Oncol. 19:302018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ueno NT and Mamounas EP: Neoadjuvant
nab-paclitaxel in the treatment of breast cancer. Breast Cancer Res
Treat. 156:427–440. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Morin X, Jaouen F and Durbec P: Control of
planar divisions by the G-protein regulator LGN maintains
progenitors in the chick neuroepithelium. Nat Neurosci.
10:1440–1448. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Culurgioni S, Mari S, Bonetti P, Gallini
S, Bonetto G, Brennich M, Round A, Nicassio F and Mapelli M:
Insc:LGN tetramers promote asymmetric divisions of mammary stem
cells. Nat Commun. 9:10252018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu X, Wang J and Sun G: Identification of
key genes and pathways in renal cell carcinoma through expression
profiling data. Kidney Blood Press Res. 40:288–297. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Nowakowska A and Tarasiuk J: Invasion and
metastasis of tumour cells resistant to chemotherapy. Postepy Hig
Med Dosw (Online). 71:380–397. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kaur R, Kaur G, Gill RK, Soni R and
Bariwal J: Recent developments in tubulin polymerization
inhibitors: An overview. Eur J Med Chem. 87:89–124. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wiser O, Qian X, Ehlers M, Ja WW, Roberts
RW, Reuveny E, Jan YN and Jan LY: Modulation of basal and
receptor-induced GIRK potassium channel activity and neuronal
excitability by the mammalian PINS homolog LGN. Neuron. 50:561–573.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Churchill CD, Klobukowski M and Tuszynski
JA: Elucidating the mechanism of action of the clinically approved
taxanes: A comprehensive comparison of local and allosteric
effects. Chem Biol Drug Des. 86:1253–1266. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Needleman DJ, Ojeda-Lopez MA, Raviv U,
Ewert K, Miller HP, Wilson L and Safinya CR: Radial compression of
microtubules and the mechanism of action of taxol and associated
proteins. Biophys J. 89:3410–3423. 2005. View Article : Google Scholar : PubMed/NCBI
|