Introduction
Chronic inflammation promotes the development,
growth and metastasis of cancer. Acute inflammation, if not
controlled develops into chronic inflammation, which may increase
the risk of cancer. Epidemiological studies have confirmed that 25%
of tumors are developed from inflammation (1). Patients with the inflammatory
autoimmune disease ulcerative colitis are 10 times more likely to
develop colon cancer than healthy individuals (2).
The accumulation of inflammatory cells and
inflammatory cytokines in the tumor microenvironment promotes
malignant cell proliferation, metastasis and epithelial-mesenchymal
transition (EMT), and can lead to the loss of the acquired immune
response (3). The majority of
cytokines are low molecular weight soluble proteins, which have
many functions, such as regulating cell growth, hematopoiesis and
the immune response and repairing damaged tissues (4). Cytokines can be divided into
lymphokines, produced by lymphocytes, and mononuclear factors,
produced by mononuclear macrophages. Interleukins (ILs),
interferons, colony stimulating factors, tumor necrosis factors and
transforming growth factors are all cytokines produced by immune
cells, which play important regulatory roles in the immune system
and can lead to pathological reactions when expressed at altered
levels.
IL-1β is a member of the IL-1 family of cytokines.
IL-1β is produced by activated macrophages as a proprotein, which
is proteolytically processed to its active form by caspase-1
(CASP1). CASP1 is an important mediator of the inflammatory
response, and is involved in a variety of cellular activities,
including cell proliferation, differentiation and apoptosis
(5). A number of studies have
determined that IL-1β also plays an important role in the
occurrence and development of tumors. In breast cancer, IL-1β
induces EMT and promotes disease relapse (6). In gastric cancer, IL-1β-induced p38
pathway activation promoted cell invasion and migration via
increased matrix metalloproteinase (MMP)2 and MMP9 expression
(7). In addition, IL-1 receptor
antagonist (IL-1RA) is an agent that binds to the cell surface IL-1
receptor (IL-1R), the same receptor that binds to IL-1 family
members, preventing IL-1 from sending a signal to cells. A previous
study suggested that serum concentrations of IL-1RA in colorectal
cancer (CRC) patients were significantly higher than those in
healthy patients, however, the specific role of IL-1RA in CRC has
not been clarified (8).
In the present study, recombinant human (rh)IL-1β
and rhIL-1RA were used to study the role of IL-1β and IL-1RA in
CRC.
Materials and methods
Patients and specimens
CRC tumor specimens and paired non-tumor mucosa were
collected between July 2012 and July 2018. Patients with the
following criteria were excluded from participation in the study:
i) The patient had received adjuvant chemotherapy or radiotherapy
prior to surgery; ii) the patient had additional cancer diagnoses.
All patients were classified according to the 7th edition of the
TNM staging system 23. Postoperative adjuvant therapies were
performed, according to standard schedules and doses. All
participating patients provided their written informed consent.
This study was approved by the Ethical Committee of Shanghai Pudong
Hospital. Patient details are summarized in Table I.
 | Table I.Clinical characteristics of
patients. |
Table I.
Clinical characteristics of
patients.
| Characteristics | No. of patients |
|---|
| UICC (TNM) stage |
|
| I | 7 |
| II | 27 |
| III | 25 |
| IV | 8 |
| Tumor (T) stage |
|
|
pTis-1 | 2 |
| pT2 | 17 |
| pT3 | 43 |
| pT4 | 5 |
| N stage |
|
| N0 | 33 |
| N1 | 30 |
| N2 | 4 |
| M stage |
|
| M0 | 60 |
| M1 | 7 |
| Age |
|
|
<65 | 30 |
|
65–75 | 20 |
|
75–85 | 16 |
|
>85 | 1 |
| Sex |
|
| Male | 40 |
|
Female | 27 |
| Tumor location |
|
| Right
colon | 15 |
| Left
colon | 3 |
|
Transverse colon | 5 |
| Sigmoid
colon | 15 |
|
Rectum | 29 |
| Histological
grade |
|
| Well
differentiated | 66 |
| Poorly
differentiated | 1 |
| Mucinous Colloid
Type |
|
| No | 53 |
| Yes | 14 |
TCGA database and analysis
TCGA-Colon Adenocarcinoma (https://cancergenome.nih.gov/) contains 480 colon
cancer cases and 41 normal control cases, accompanied with clinical
characteristics. All mRNA expression data accompanied by clinical
data was downloaded by R-software for subsequent analysis. The
expression of IL-1β and IL-1RA in CRC were analyzed by UALCAN web
tools based on the TCGA database (9) (http://ualcan.path.uab.edu/). Co-expression, overall
survival (OS) and recurrence-free survival (RFS) of IL-1β and
IL-1RA genes were identified via GEPIA web tools based on TCGA
database (10) (http://gepia.cancer-pku.cn/).
Immunohistochemical (IHC)
staining
IHC staining was carried out according to the
antibody manufacturers' instructions. Briefly, formalin-fixed and
paraffin-embedded tissue sections were deparaffinized in xylene and
hydrated with decreasing concentrations of ethanol (100, 95, 80 and
75%). The slices were then soaked in 10% BSA to inhibit endogenous
peroxidase activity and incubated with IL-1β and IL-1RA rabbit
polyclonal antibody (dilution 1:100; cat. nos. AF-201 and AF-280;
R&D Systems, Inc.) at 4°C overnight. A horseradish
peroxidase-conjugated rabbit secondary antibody (cat. no. ab6721;
Abcam) was added for 60 min at room temperature; then,
3,3′-diaminobenzidine development (DAB Substrate Chromogen System;
Dako; Agilent Technologies, Inc.) and hematoxylin staining were
performed according to standard protocols. Slides were fixed via
neutral gum for ~1 min at room temperature and images were obtained
using an Olympus IX71 inverted microscope with a DP2-BSW Olympus
image acquisition software system (Olympus Corp.).
Cell line and reagents
The human CRC cell line HCT116 was purchased from
the University of Colorado Cancer Center Cell Bank and cultured in
RPMI-1640 medium supplemented with 10% FBS (Invitrogen; Thermo
Fisher Scientific, Inc.) at 37°C in a 5% CO2 atmosphere.
Cells were digested and passaged when cell confluence reached 80%.
Recombinant human (rh) IL-1β protein and IL-1RA protein were
purchased from R&D Systems, Inc. The working concentration was
100 nM.
Protein extraction and western blot
analysis
Total protein was extracted from HCT116 cells using
RIPA lysis buffer (Beyotime Institute of Biotechnology) with 1%
phenylmethanesulfonyl fluoride (PMSF). Then, equal amounts (20 µg)
of protein determined by BCA protein assay kit (Thermo Fisher
Scientific, Inc.) were separated using 10% SDS-PAGE gels. The
proteins were then transferred to PVDF membranes. The membranes
were blocked with 5% non-fat milk for 1 h at room temperature and
then incubated with primary antibodies at 4°C for 12 h. The
following antibodies were tested: Epithelial (E)-cadherin (cat. no.
20874), neural (N)-cadherin (cat. no. 22018), vimentin (cat. no.
10366) and zinc finger protein Snail1 (snail1; cat. no. 13099)
rabbit polyclonal antibodies (1:1,000; ProteinTech Group, Inc.);
p62 (cat. no. ab109012) and LC3B (cat. no. ab192890) rabbit
polyclonal antibodies (1:2,000; Abcam); IL-1RA rabbit polyclonal
antibody (1:1,000; cat. no. AF-201; R&D Systems, Inc.). β-actin
rabbit polyclonal antibodies (1:4,000; cat. no. 60008; ProteinTech
Group, Inc.) were used as loading controls for normalization. The
secondary antibodies were anti-rabbit antibodies and conjugated to
horseradish peroxidase (1:4,000; cat. no. SA00001; ProteinTech,
Inc.). The membranes were incubated with the secondary antibodies
for approximately 1 h at room temperature. The bands were
visualized with ECL reagents (Thermo Fisher Scientific, Inc.) and
developed using the Omega Lum™ G (Aplegen/Gel Company). ImageJ
(version 2017; National Institutes of Health) was used for
densitometry.
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
Total RNA was extracted from HCT116 cells using
TRIzol™ reagent (Invitrogen; Thermo Fisher Scientific, Inc.). cDNA
was obtained from total RNA using a PrimeScript™ RT reagent kit
(Takara Bio, Inc.). The expression of mRNA was assessed by RT-qPCR,
which was carried out in triplicate using a SYBR Premix Ex Taq™ kit
(Takara Bio, Inc.) and an ABI 7900HT Real-Time PCR system (95°C for
30 sec; 95°C for 3 sec, 60°C for 30 sec, 40 cycles; Applied
Biosystems; Thermo Fisher Scientific, Inc.). The primers used are
presented in Table II. The
comparative cycle threshold values method (2−ΔΔCq) was
used to analyze the final results (11).
| Table II.The primers of RT-qPCR. |
Table II.
The primers of RT-qPCR.
| Gene | Forward primer | Reverse primer |
|---|
| Beclin1 |
CAAGATCCTGGACCGTGTCA |
TGGCACTTTCTGTGGACATCA |
| p62 |
GACTACGACTTGTGTAGCGTC |
AGTGTCCGTGTTTCACCTTCC |
| VSP34 |
GGACCTTCTGACCACGAT |
GCAACAGCATAACGCCTC |
| ATG7 |
TGTATAACACCAACACACTCGA |
GGCAGGATAGCAAAACCAATAG |
| ATG4b |
AGAGCCCGTTTGGATACT |
GTCGATGAATGCGTTGAG |
| Actin |
GGGACCTGACTGACTACCTC |
TCATACTCCTGCTTGCTGAT |
| E-cadherin |
AGTCACTGACACCAACGATAAT |
ATCGTTGTTCACTGGATTTGTG |
| Vimentin |
AGTCCACTGAGTACCGGAGAC |
CATTTCACGCATCTGGCGTTC |
| Snail1 |
AAGGATCTCCAGGCTCGAAAG |
GCTTCGGATGTGCATCTTGA |
| N-cadherin |
TGTATGTGGGCAAGATCCACT |
CTCGTCGATCAGGAAGATGGT |
Cell proliferation assay
For this assay, 5×103 HCT 116 cells were
seeded into 96-well plates and incubated for the following
time-points: 0, 24, 48 and 72 h. Before determination, 10 µl of
Cell-Counting Kit-8 (CCK-8; Dojindo Molecular Technologies, Inc.)
solution was added to each well of the plate, and the incubation
was continued for 2 h. Finally, the absorbance of each well was
measured at a 450 nm wavelength.
Clone formation test
For this assay, 500 HCT 116 cells were seeded into
each well of 6-well plates and incubated at 37°C. Clone size was
observed under an ZEISS Axiovert 40 inverted microscope (Carl Zeiss
AG; magnification, ×200) every day until the number of cells in
most wells was more than 50. The medium was then removed and cells
were stained with 0.2% crystal violet for 30 min. The cells were
washed three times with PBS, then images were acquired and clones
were counted. The ratio of clone formation was calculated following
the equation: Ratio of clone formation (%) = clone number/500
×100%.
Flow cytometry
A total of 2×105 HCT 116 cells were
harvested and washed with PBS 3 times. The samples were then
resuspended in 100 µl binding buffer and stained with 5 µl of
Annexin V and propidium iodide (PI) for 20 min at room temperature
in the dark. Subsequent to staining, an additional 400 µl binding
buffer was added to the sample and resuspended. Analyses were
performed with flow cytometry (BD Biosciences).
Cell migration and invasion
assays
Cell migration and invasion were analyzed using
Transwell plates (24-well insert; 8 µm pore size; BD Biosciences).
The filters (Corning, Inc.) were coated with Matrigel for invasion
assays and uncoated for migration assays. A total of 55 µl Matrigel
suspended in PBS was used to coat the plates (1:8 dilution; BD
Biosciences). For the migration assays, 5×104 HCT116
cells were suspended in 200 µl of serum-free medium and seeded into
the upper chambers of the Transwells. A 600-µl volume of medium
containing 10% FBS was then added to the lower chamber as a
chemoattractant. After incubation at 37°C for 24 h, the membranes
were fixed with 4% formaldehyde for 30 min and stained with 0.1%
crystal violet at room temperature for 30 min. For invasion assays,
1×105 HCT116 cells suspended in 200 µl serum-free medium
were seeded into the Matrigel-coated upper chambers of the
Transwells and the protocol followed was as aforementioned. The
cells were counted and images were captured under an inverted
microscope (magnification, ×400) in 5 different fields of view per
filter. Each condition was studied in triplicate.
Wound healing assay
For this assay, 5×105 HCT 116 cells were
seeded into 6-well plates and cultured at 37°C for 24 h. A 200-µl
sterile micro-pipette tip was used to scratch the confluent
monolayers in a straight line when cells were 80–90% confluent.
Floating cells were then removed by washing with PBS three times
and the cells were cultured in serum-free medium. Images of the
same wound position were captured after at 0 and 48 h under a
microscope. The migration results were analyzed using by ImageJ
software (version 2017; National Institutes of Health).
Subcutaneous xenografts of nude
mice
Balb/c-nude mice (age, 5 weeks) were provided by the
Beijing Vital River Laboratory Animal Technology Co., Ltd. All
detailed experimental procedures were approved by the Institutional
Animal Care and Utilization Committee of Fudan University Pudong
Animal Experimental Center. All the mice (n=16) were randomly
divided between the rhIL-1β group (n=8) and the normal control (NC)
group (n=8). HCT-116 cells (5×106) suspended in 100 µl
PBS were injected subcutaneously from the axilla of each nude
mouse. After 2 weeks, the rhIL-1β group were treated with 1 µg
rhIL-1β dissolved in 100 µl normal saline via intraperitoneal
injection (once every two days, for 14 days); while, the NC group
was treated with 100 µl normal saline as a placebo. The long (L)
and short (S) diameter of the tumors were measured with a Vernier
caliper every 3 days (tumor volume = LxS2/2). The growth
curve of subcutaneous tumors was plotted based on the measured
tumor volume. All mice were euthanized 2 weeks after treatment by
injection of excessive 2% sodium pentobarbital, followed with rapid
cervical vertebra dislocation.
Statistical analysis
SPSS software (version 19.0, IBM Corp.) was used for
statistical analysis of all experimental data. GraphPad Prism
(version 7; GraphPad Software, Inc.) was used to determine the
statistical results. All data are expressed as the mean ± standard
deviation. Statistical analysis of data from 2 groups was performed
using a t-test. The comparison between multiple groups was
performed using one-way ANOVA and then an LSD test. P<0.05 was
considered to indicate a statistically significant difference.
Results
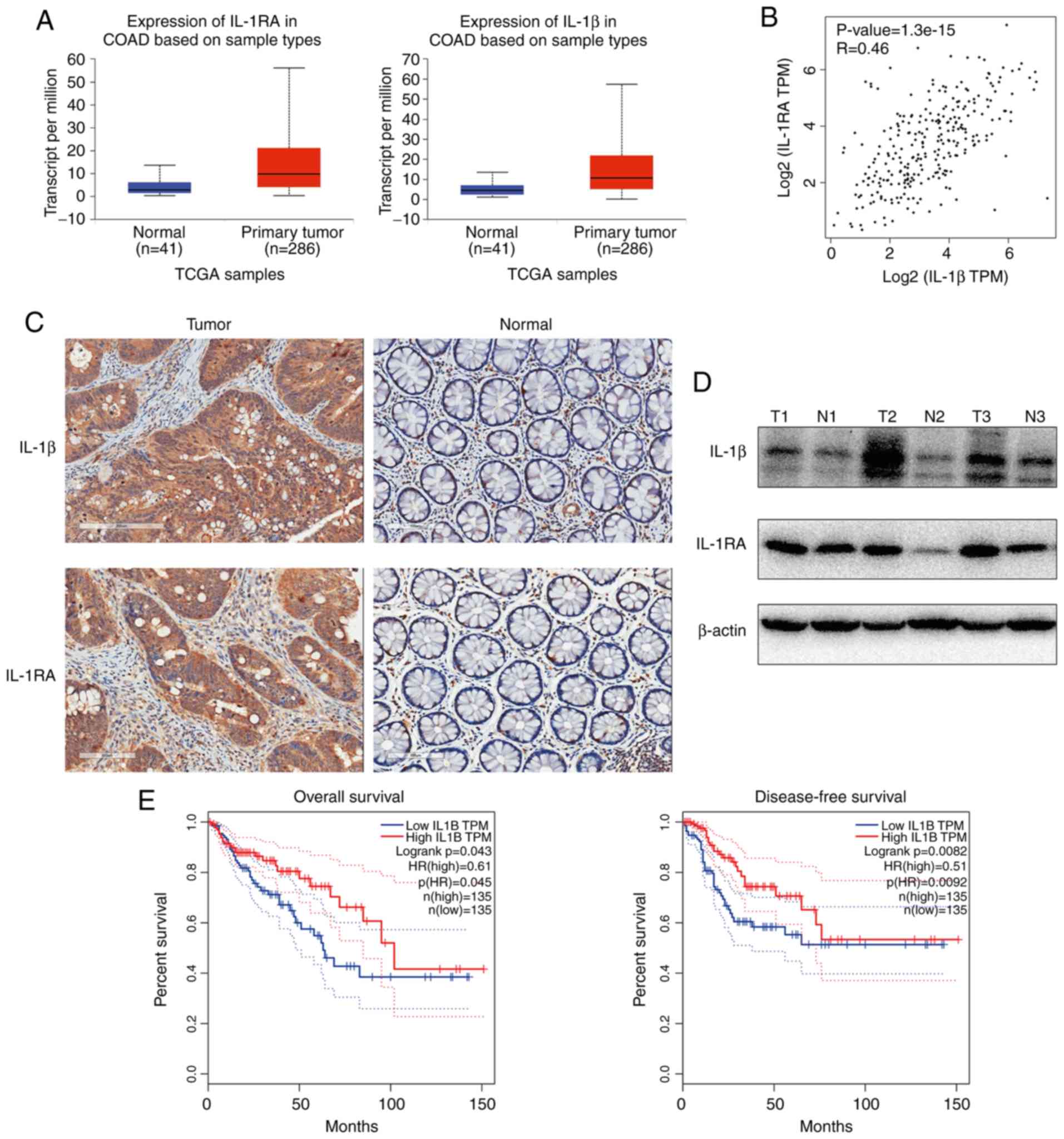
IL-1β and IL-1RA are upregulated in
CRC and are associated with an increased rate of overall survival
(OS)
The expression of IL-1β and IL-1RA in CRC were
analyzed by UALCAN web tools based on the TCGA database, and
significantly higher levels of both IL-1β and IL-1RA were observed
in CRC tumor tissue compared with normal mucosa (9) (http://ualcan.path.uab.edu/; Fig. 1A). Significant co-expression between
IL-1β and IL-1RA genes was identified via GEPIA web tools based on
TCGA database (10) (http://gepia.cancer-pku.cn/; Fig. 1B). Additionally, western blotting
(Fig. 1C) and IHC (Fig. 1D) were performed to determine the
protein expression levels, and the results indicated that both
IL-1β and IL-1RA were increased in CRC compared with paired
non-tumor mucosa. Collectively, these data indicated increased
expression of IL-1β and IL-1RA in tumor tissues at both the mRNA
and protein levels. To explore the clinical significance of
increased IL-1β expression in CRC, the GEPIA tool was used to
analyze OS and recurrence-free survival (RFS) rates in CRC patients
based on the TCGA database. Increased expression of IL-1β was
associated with an increased rate of OS and RFS (Fig. 1E)
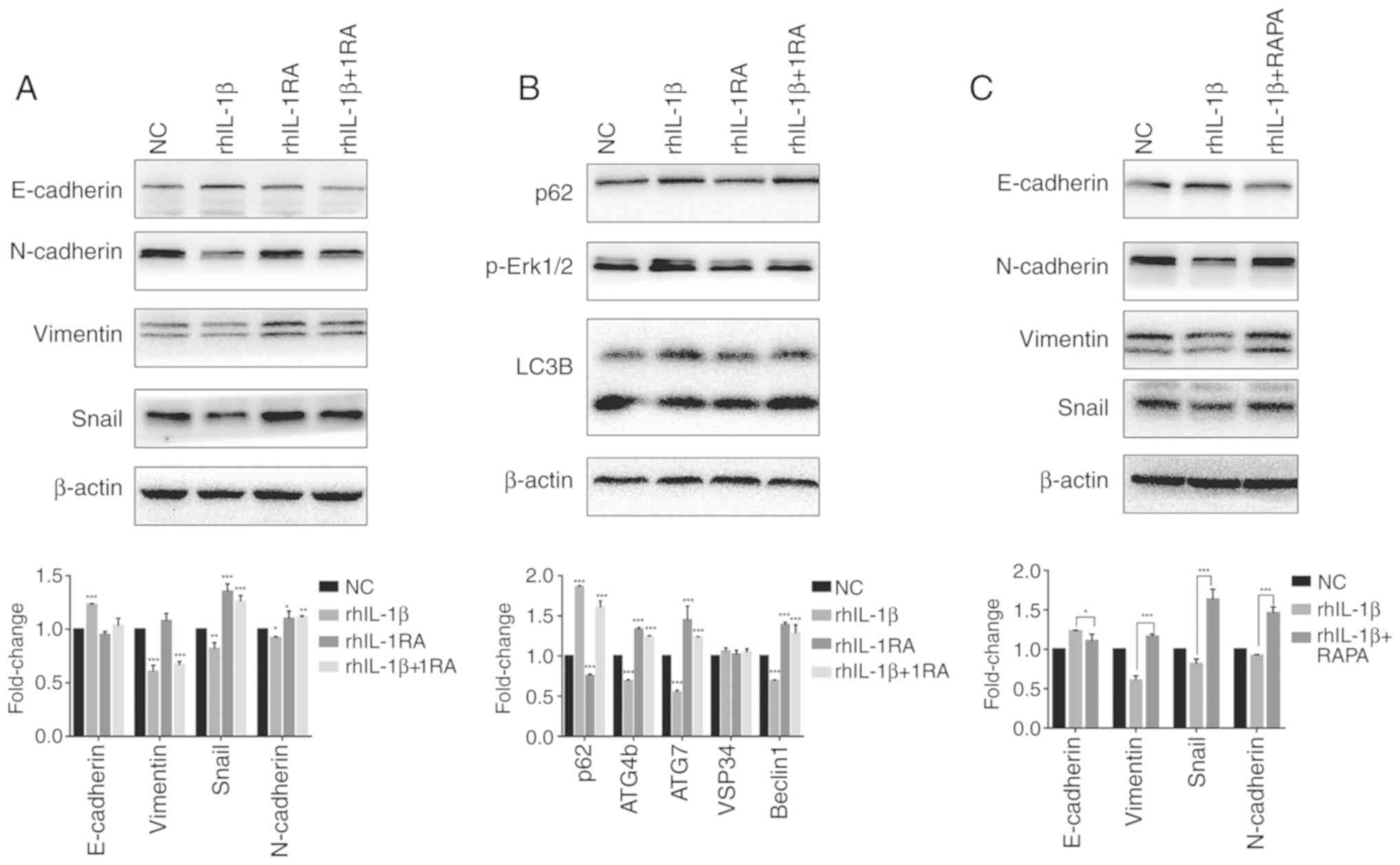
The IL-1β/IL1RA axis regulates EMT via
autophagy in vitro
Western blotting and RT-qPCR were performed to
determine the expression levels of several EMT-associated markers
(Fig. 2A). rhIL-1β increased the
expression of E-cadherin, whereas it reduced the expression of
N-cadherin, vimentin and Snail. rhIL-1RA revealed the opposite
effect in EMT markers compared with rhIL-1β; while, rhIL-1RA
rescued the increased expression of E-cadherin and reduced
expression of N-cadherin, vimentin and Snail in the rhIL-1β-treated
group. These results indicated that rhIL-1β inhibited EMT whereas
rhIL-1RA induced EMT in HCT-116 cells. The levels of several key
autophagy-associated markers in each group were also determined
(Fig. 2B). rhIL-1β increased the
expression of p62 and decreased the ratio of LC3 II/I, expression
of cysteine protease ATG4b (ATG4b), ubiquitin-like
modifier-activating enzyme ATG7 (ATG7) and Beclin1. rhIL-1RA
revealed the opposite effect on autophagy markers when compared
with rhIL-1β; while, rhIL-1RA rescued the increased expression of
p62 and reduced ratio of LC3 II/I, expression of ATG4b, ATG7, VSP34
and beclin1. These results indicated that rhIL-1β inhibited
autophagy whereas rhIL-1RA induced autophagy in HCT-116 cells. To
study the association between rhIL-1β/1RA-regulated autophagy and
EMT, autophagy activator RAPA (5 mM for 48 h) was used to treat the
rhIL-1β group. It was revealed that RAPA significantly rescued
increased expression of E-cadherin and reduced expression of
N-cadherin, vimentin and Snail in the rhIL-1β group compared with
levels in mice untreated with RAPA. These results indicated that
IL-1β/IL-1RA regulated EMT via autophagy.
IL-1β inhibits cell migration and
invasion
Transwell assays were performed to assess the impact
of IL-1β and IL-1RA on cell invasion and migration. In the invasion
assay, rhIL-1β treatment decreased the invasion ability of HCT-116
cells, whereas rhIL-1RA increased the invasion ability compared
with the NC group. Both rhIL-1RA and RAPA could rescue this
invasion ability in the rhIL-1β group (Fig. 3A). Migration assays exhibited
similar results to the invasion assay (Fig. 3B). Furthermore, the migratory
ability of each group was also confirmed by wound healing assay.
rhIL-1β increased the wound area in HCT-116 cells compared with the
control cells, whereas rhIL-1RA decreased it. Both rhIL-1RA and
RAPA treatment decreased the wound area in the rhIL-1β group
compared with control treatments (Fig.
3C).
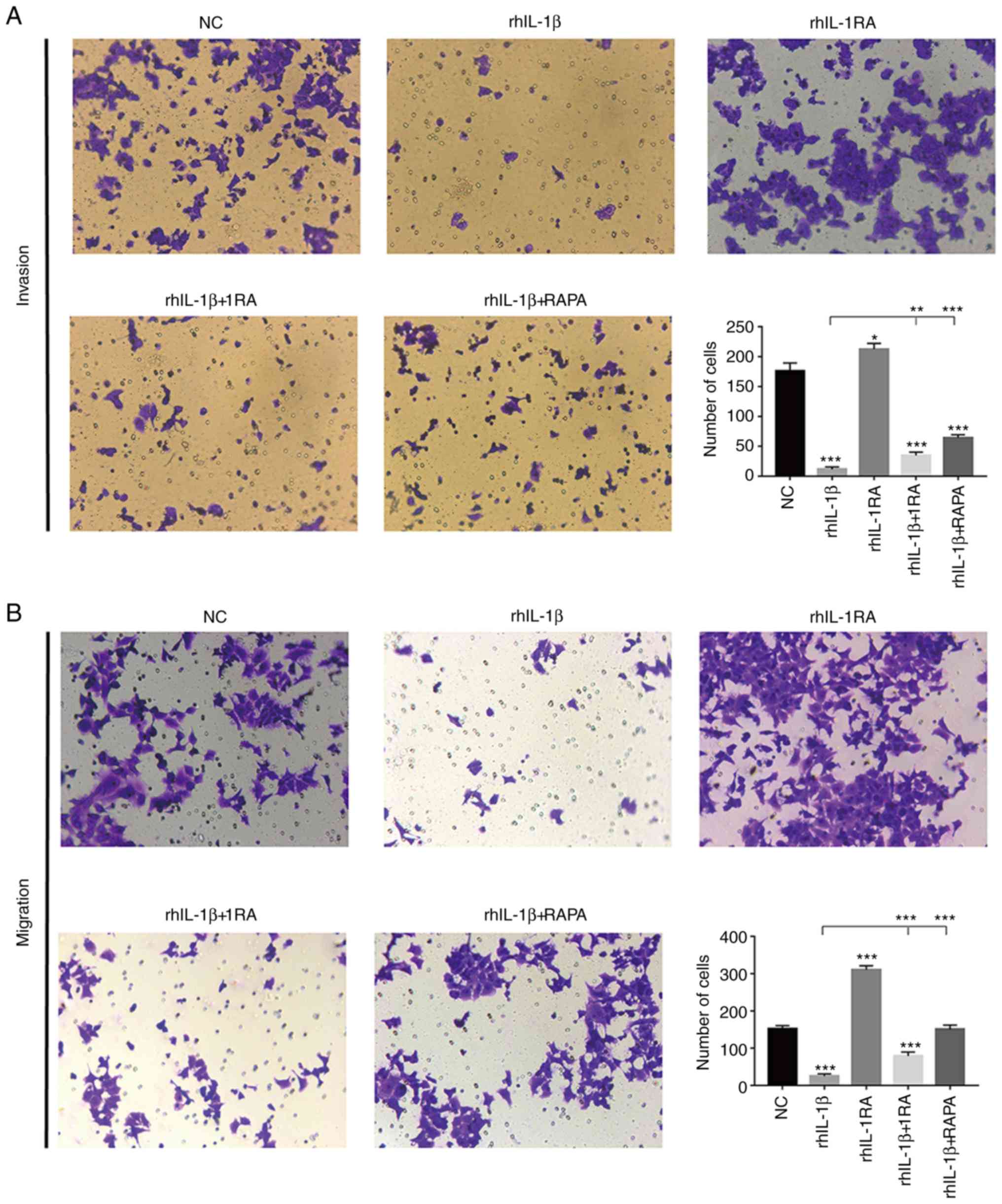 | Figure 3.(A) Invasion ability of each group
analyzed by Transwell assay. (B) Migration ability of each group
analyzed by Transwell assay. *P<0.05, **P<0.01 and
***P<0.001. rh, recombinant human, IL, interleukin; RAPA,
rapamycin. (C) Migration ability of each group analyzed by wound
healing assay. *P<0.05, **P<0.01 and ***P<0.001. rh,
recombinant human, IL, interleukin; RAPA, rapamycin. |
IL-1β reduces cell proliferation and
clone formation ability and promotes cell apoptosis
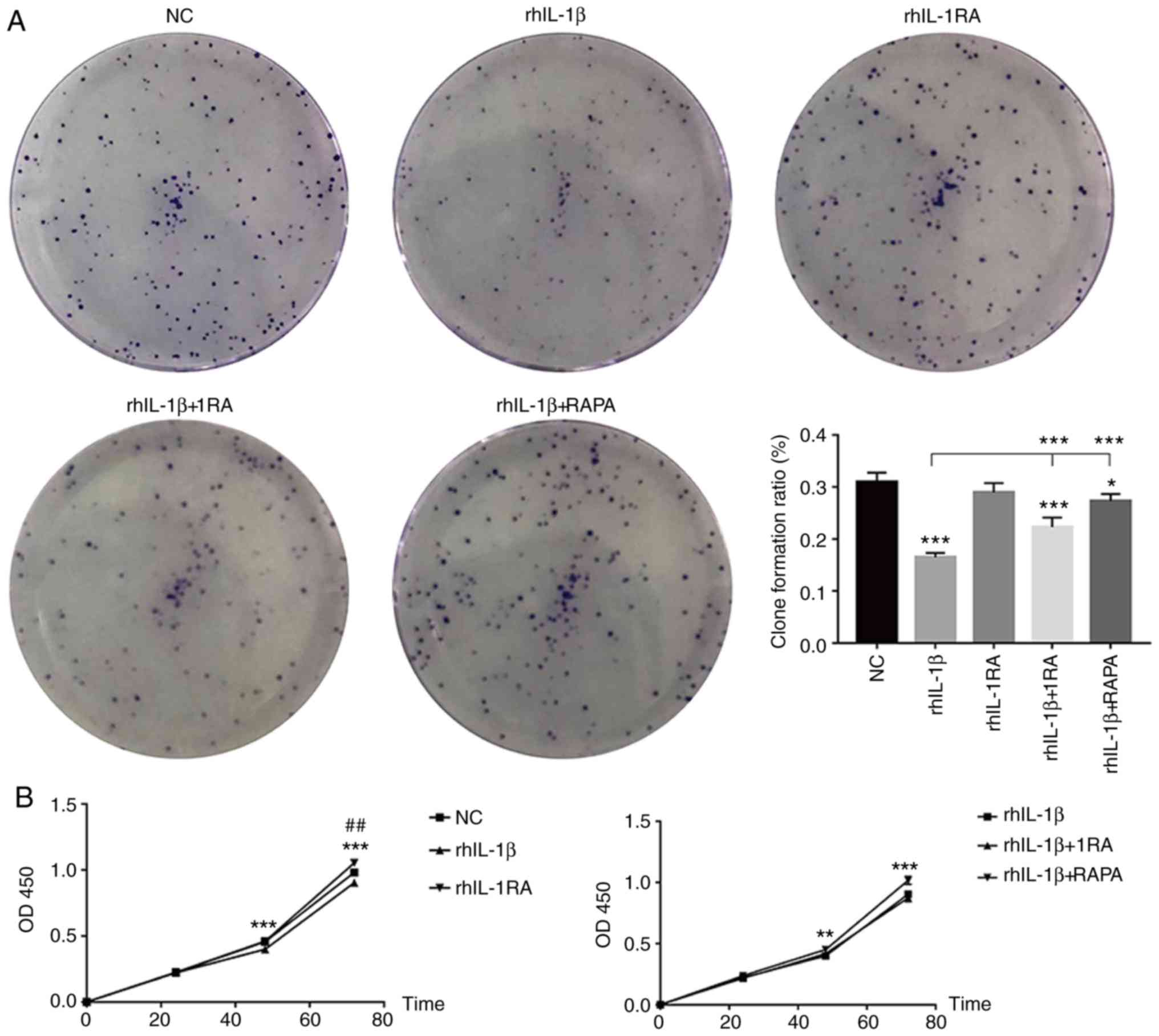
Clone formation assays and a CCK-8 assay were
performed to assess clone formation ability and proliferation in
each group. rhIL-1β reduced clone formation in HCT-116 cells
compared to untreated cells, whereas rhIL-1RA treatment exhibited
no effect. In addition, both rhIL-1RA and RAPA could rescue
inhibited clone formation in the rhIL-1β-treated group (Fig. 4A). Furthermore, rhIL-1β inhibited
proliferation of HCT-116 at 48 h and 72 h compared with the control
treatment; whereas rhIL-1RA promoted proliferation of HCT-116 cells
at 72 h compared with the control treatment. RAPA could rescue
inhibited proliferation in the rhIL-1β group, whereas rhIL-1RA
exhibited no effect (Fig. 4B). Flow
cytometry was performed to assess cell apoptosis in each group.
rhIL-1β treatment (51.97±1.73%) promoted apoptosis of HCT-116 cells
compared with the control treatment (27.19±0.33%), whereas rhIL-1RA
exhibited no effect (25.56±0.09%). In addition, both rhIL-1RA
(28.3±0.24%) and RAPA (27.09±0.24%) could rescue the increase in
apoptosis observed in the rhIL-1β group (Fig. 4C).
 | Figure 4.(A) Clone formation ability of each
group. *P<0.05 and ***P<0.001. (B) Proliferation of each
group detected by CCK-8 assay. rh1L-β vs. NC, **P<0.01 and
***P<0.001; rhIL-1RA vs. NC, ##P<0.01. (C) Cell
apoptosis of each group detected by flow cytometry (***P<0.001).
rh, recombinant human, IL, interleukin; RAPA, rapamycin. (C) Cell
apoptosis of each group detected by flow cytometry (***P<0.001).
rh, recombinant human, IL, interleukin; RAPA, rapamycin. |
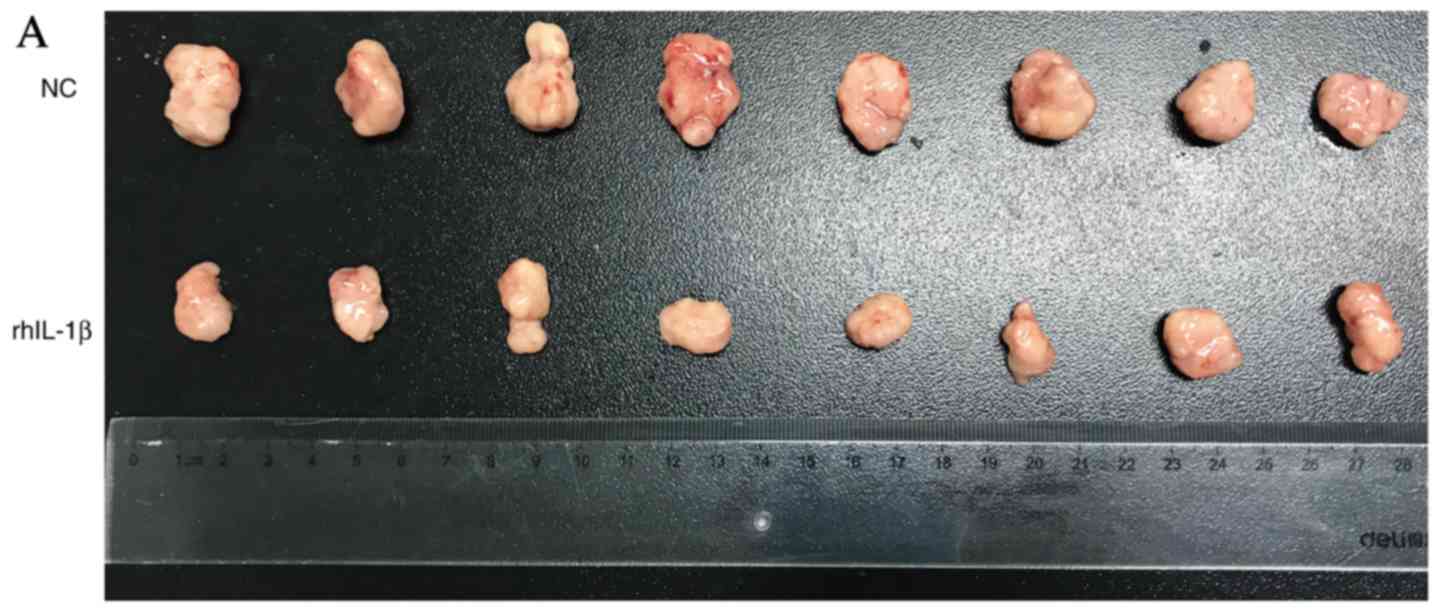
IL-1β inhibits growth of subcutaneous
xenografts in nude mice
To demonstrate the effect of rhIL-1β in vivo,
subcutaneous xenografts were injected into nude mice. A total of 1
µg rhIL-1β was administered intraperitoneally (once per two days)
and the results revealed that it significantly inhibited the growth
of xenografts between day 5 and 15 (Fig. 5A and B). In addition, rhIL-1β
exhibited no effect on mouse weight, health, food intake, sleep and
activity compared with control treatment (Fig. 5C).
Discussion
The tumor microenvironment is the area between tumor
cells and adjacent normal tissues. Its components include
extracellular matrix, soluble molecules and tumor stromal cells.
Once the tumor microenvironment is formed, numerous immune cells,
such as T cells, macrophages are chemotactically drawn to this
point (12). In addition to
cellular factors, the molecules in the tumor microenvironment
include extracellular matrix molecules, cytokines and chemokines.
Cells and molecules in the tumor microenvironment are dynamic,
reflecting the essence of tumor microenvironment evolution, and the
final outcome is that a large number of immunosuppressive cells
such as myeloid-derived suppressor cells aggregate in the tumor
microenvironment to regulate immune escape, growth and metastasis
of the tumor (13).
In the present study expression of IL-1β was
assessed in CRC patients and a significant increase was observed
when compared with normal tissues. In previous research of other
tumors, IL-1β was associated with metastasis and induction of EMT
(7). It was hypothesized that IL-1β
would play a tumorigenic role in CRC. However, analysis found
higher expression of IL-1β was associated with better OS and RFS,
indicating a beneficial role in CRC. Significant co-expression
between IL-1β and IL-1RA was also observed, indicating a possible
interaction between them.
The regulation of EMT by rhIL-1β was studied as was
demonstrated to have an activation effect in breast cancer
(6). Notably, 100 nM rhIL-1β for 48
h significantly inhibited EMT, in contrast with the results of a
previous study in breast cancer. In addition, rhIL-1RA induced EMT
and rescued inhibition of EMT by rhIL-1β. Autophagy is one of the
main regulatory mechanisms of EMT. Long non-coding RNA CPS1-IT may
suppress metastasis and EMT by inhibiting hypoxia-induced autophagy
through inactivation of hypoxia inducible factor-1α in CRC
(14). In the present study levels
of several key markers of autophagy were determined, and the
results indicated that rhIL-1β inhibited autophagy in CRC, whereas
rhIL-1RA activated autophagy. Furthermore, autophagy activator RAPA
significantly rescued inhibited autophagy in the rhIL-1β group.
Collectively, these results indicated that IL-1β inhibited EMT
in vitro via inhibition of autophagy; while, IL-1RA induced
EMT in vitro via activation of autophagy. EMT is an
important process for tumors to acquire invasiveness, and autophagy
had also been demonstrated to promote invasion and metastasis
(15,16). Autophagy is a process of
phagocytosis of cytoplasmic proteins or organelles into vesicles
and fusion with lysosomes to form autophagic lysosomes, which
degrade the contents of the lysosomes, thereby realizing the
metabolic needs of the cells themselves and leading to the renewal
of some organelles. Many malignant tumors are positively or
negatively correlated with autophagy in many stages of occurrence,
development and metastasis (17,18).
Transwell assays and wound healing assay were
performed to assess cell invasion and migration. As anticipated,
rhIL-1β significantly decreased cell invasion and migration and
rhIL-1RA promoted invasion and migration. In addition, both
rhIL-1RA and RAPA exhibited a similar rescue effect of inhibited
invasion and migration abilities in the rhIL-1β treated group.
These data indicated that the IL-1β/1RA axis regulated EMT via
autophagy. In addition, rhIL-1β decreased clone formation ability
in comparison with the control treatment, whereas rhIL-1RA
exhibited no effect. In addition, both rhIL-1RA and RAPA could
rescue inhibited clone formation ability in the rhIL-1β group.
Furthermore, rhIL-1β inhibited proliferation of HCT-116 cells after
48 h, whereas rhIL-1RA promoted proliferation of HCT-116 cells
after 72 h. RAPA could rescue inhibited proliferation in the
rhIL-1β group, whereas rhIL-1RA exhibited no effect. In addition,
rhIL-1β promoted apoptosis of HCT-116 cells, whereas rhIL-1RA
exhibited no effect. Both rhIL-1RA and RAPA could rescue the
increase of apoptosis in rhIL-1β group. These results indicated
IL-1β-1RA autophagy-regulated clone formation, cell proliferation
and apoptosis may be a complex process.
Finally, the therapeutic effect of rhIL-1β was
assessed in vivo. rhIL-1β treatment significantly inhibited
the growth of xenografts between 5 and 15 days compared with the
control treatment, with no effect on general health status.
A limitation of the present study was that our
research was performed only in HCT-116 cells, and more CRC cell
lines remain to be investigated. We will further investigate the
specific mechanism of the IL-1β-1RA-autophagy axis in more CRC cell
lines as well as other tumors.
In conclusion, IL-1β and IL-1RA were highly
expressed in CRC patients. The IL-1β/1RA axis was revealed to
regulate EMT, cell invasion, migration, clone formation,
proliferation and apoptosis in vitro via autophagy. In
addition, IL-1β also inhibited the growth of xenografts in
vivo, and may be suitable as a new therapeutic drug for CRC
patients.
Acknowledgements
Not applicable.
Funding
The present study was funded by Puxiu Medical
Talents Training Program of Pudong Hospital (grant no.
PX201702).
Availability of data and materials
The datasets used and analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YC and ZY contributed equally to the cell
experiments and mice model. BD and DW contributed to the
statistical analysis of the data and collection of specimens. YQ
and ZM contributed to the design of the study and supervision. All
authors read and approved the final version of the manuscript. All
authors read and approved the manuscript and agree to be
accountable for all aspects of the research in ensuring that the
accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
All procedures involving human participants were
performed in accordance with Shanghai Pudong Hospital Ethics
Committee and with the 1964 Declaration of Helsinki and its later
amendments or comparable ethical standards. All patients provided
their written informed consent. The study protocol was approved by
the Pudong Hospital Committee on human research. All detailed
experimental animal procedures were approved by the Institutional
Animal Care and Utilization Committee of Fudan University Pudong
Animal Experimental Center.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Taniguchi K and Karin M: IL-6 and related
cytokines as the critical lynchpins between inflammation and
cancer. Semin Immunol. 26:54–74. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Long AG, Lundsmith ET and Hamilton KE:
Inflammation and colorectal cancer. Curr Colorectal Cancer Rep.
13:341–351. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Huang C, Yang G, Jiang T, Zhu G, Li H and
Qiu Z: The effects and mechanisms of blockage of STAT3 signaling
pathway on IL-6 inducing EMT in human pancreatic cancer cells in
vitro. Neoplasma. 58:396–405. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lee S and Margolin K: Cytokines in Cancer
immunotherapy. Cancers (Basel). 3:3856–3893. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Masters SL, Dunne A, Subramanian SL, Hull
RL, Tannahill GM, Sharp FA, Becker C, Franchi L, Yoshihara E, Chen
Z, et al: Activation of the NLRP3 inflammasome by islet amyloid
polypeptide provides a mechanism for enhanced IL-1β in type 2
diabetes. Nat Immunol. 11:897–904. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Soria G, Ofrishahak M, Haas I,
Yaal-Hahoshen N, Leider-Trejo L, Leibovich-Rivkin T, Weitzenfeld P,
Meshel T, Shabtai E, Gutman M and Ben-Baruch A: Inflammatory
mediators in breast cancer: Coordinated expression of TNFα and
IL-1β with CCL2 and CCL5 and effects on epithelial-to-mesenchymal
transition. BMC Cancer. 11:1302011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang Q, Lan F, Wang X, Yu Y, Ouyang X,
Zheng F, Han J, Lin Y, Xie Y, Xie F, et al: IL-1β-induced
activation of p38 promotes metastasis in gastric adenocarcinoma via
upregulation of AP-1/c-fos, MMP2 and MMP9. Mol Cancer. 13:182014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ito H and Miki C: Profile of circulating
levels of interleukin-1 receptor antagonist and interleukin-6 in
colorectal cancer patients. Scand J Gastroenterol. 34:1139–1143.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression Data using real-time quantitative PCR and
the 2-(Delta Delta C (T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lewis CE and Pollard JW: Distinct role of
macrophages in different tumor microenvironments. Cancer Res.
66:605–612. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kessenbrock K, Plaks V and Werb Z: Matrix
metalloproteinases: Regulators of the tumor microenvironment. Cell.
141:52–67. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang W, Yuan W, Song J, Wang S and Gu X:
LncRNA CPS1-IT1 suppresses EMT and metastasis of colorectal cancer
by inhibiting hypoxia-induced autophagy through inactivation of
HIF-1α. Biochimie. 144:21–27. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li J, Yang B, Zhou Q, Wu Y, Shang D, Guo
Y, Song Z, Zheng Q and Xiong J: Autophagy promotes hepatocellular
carcinoma cell invasion through activation of
epithelial-mesenchymal transition. Carcinogenesis. 34:1343–1351.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Galavotti S, Bartesaghi S, Faccenda D,
Shaked-Rabi M, Sanzone S, McEvoy A, Dinsdale D, Condorelli F,
Brandner S, Campanella M, et al: The autophagy-associated factors
DRAM1 and p62 regulate cell migration and invasion in glioblastoma
stem cells. Oncogene. 32:699–712. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fan D, Liu SY, van Hasselt CA, Vlantis AC,
Ng EK, Zhang H, Dong Y, Ng SK, Chu R, Chan AB, et al: Estrogen
receptor α induces prosurvival autophagy in papillary thyroid
cancer via stimulating reactive oxygen species and extracellular
signal regulated kinases. J Clin Endocrinol Metab. 100:E561–E571.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang W, Kang H, Zhao Y, Min I, Wyrwas B,
Moore M, Teng L, Zarnegar R, Jiang X and Fahey TJ III: Targeting
autophagy sensitizes BRAF-Mutant thyroid cancer to vemurafenib. J
Clin Endocrinol Metab. 102:634–643. 2017. View Article : Google Scholar : PubMed/NCBI
|