Introduction: General considerations on
protease inhibitors
Protease inhibitors (PIs) inhibit human
immunodeficiency virus (HIV)-1 and HIV-2 proteases, impeding viral
replication and liberation of viral elements from infected cells.
The mechanism of action of PIs involves competitive binding to the
enzyme. PIs were designed to halt the development of the HIV
virion, by impeding cleavage of polyproteins by the viral aspartyl
protease into their operational layout. Since 1995, by employing
the HIV protease crystal layout, various small inhibitors have been
constructed for HIV therapy. To date, numerous PIs have been
produced including ritonavir, amprenavir, lopinavir, ritonavir,
atazanavir, indinavir, darunavir, fosamprenavir and tipranavir.
Their use has caused a decrease in the mortality rate due to HIV
infection, and a reduction in the prevalence of opportunistic
infections (1). In fact, with the
use of PIs and of HIV reverse transcriptase inhibitors the era of
highly active antiretroviral treatment (HAART) started, which is
now the main therapy in HIV infection. Moreover, in HIV subjects
receiving PI-based treatment, an impressive decrease in the amount
of HIV-associated cancers, unconnected to the viral burden or CD4
amount, has been observed. This has paved the way for a series of
studies aimed at evaluating the effects of PIs on neoplastic
diseases. It has been therefore demonstrated that numerous PIs have
consequences on cancer proliferation, spread and outcome. This is
possibly due to actions on endoplasmic reticulum (ER) stress, the
proteasome, NF-κB and Akt signaling (2,3).
Employing drugs used for the treatment of infectious
diseases to cure tumors has been noted in several previous cases,
such as fludarabine and tetracycline, which have demonstrated
action on women with breast cancer (4). The possible use of anti-HIV drugs
against cancer is also not new. Research has demonstrated that PIs
display anticancer action which is independent from their capacity
to interfere with the HIV protease. Ikeoze et al
demonstrated that ritonavir, indinavir and saquinavir mediated a
decrease in the growth and differentiation of HL-60 and NB4 myeloid
leukemia cells (5). Other studies
have shown that PIs can reduce the growth of Kaposi sarcoma,
urological cancer cells and pheocromocytoma cells (5–8).
Notwithstanding their partial similarity with the
HIV protease, cellular proteases appear to be the main target of
PIs that are responsible for their anticancer action. PIs
particularly act on the proteasome and extracellular matrix
metalloproteases (1). Nevertheless,
several of the supplementary actions of PIs have been demonstrated.
PIs can reduce angiogenesis via a reduction in the phosphoinositide
3-kinase (PI3K)/Akt pathway, which regulates the production of
vascular endothelial growth factor (VEGF) (9,10),
increase apoptosis in cancer cells by inhibition of STAT3 and
c-Src. Moreover, PIs may block NF-κB activation via a reduction in
proteasomal degradation of IαB (6,11–14).
However, the actions appear to be cell type-dependent; for
instance, ritonavir may be protective against cell death in normal
T cells (15).
A fundamental work by Gills et al
demonstrated that PIs displayed an effect against all 60 cell lines
in the NCI60 panel and the authors reported several possible
mechanisms of action of PIs against tumor cells (16).
Nelfinavir
In March 1997, the Food and Drug Administration
approved nelfinavir (NFV) for HIV treatment. NFV is an orally
available drug used against HIV-1 and HIV-22 (17) (Fig.
1). NFV is a nonpeptidic PI that works by binding to the
catalytic site of the HIV protease, thus impeding the maturation of
viral polyprotein precursors into operational proteins that are
indispensable for viral proliferation. NFV is present in tablets of
250 and 625 mg or as an oral suspension powder. The suggested dose
of NFV for adult patients is 1,250 mg twice or 750 mg three times a
day (17).
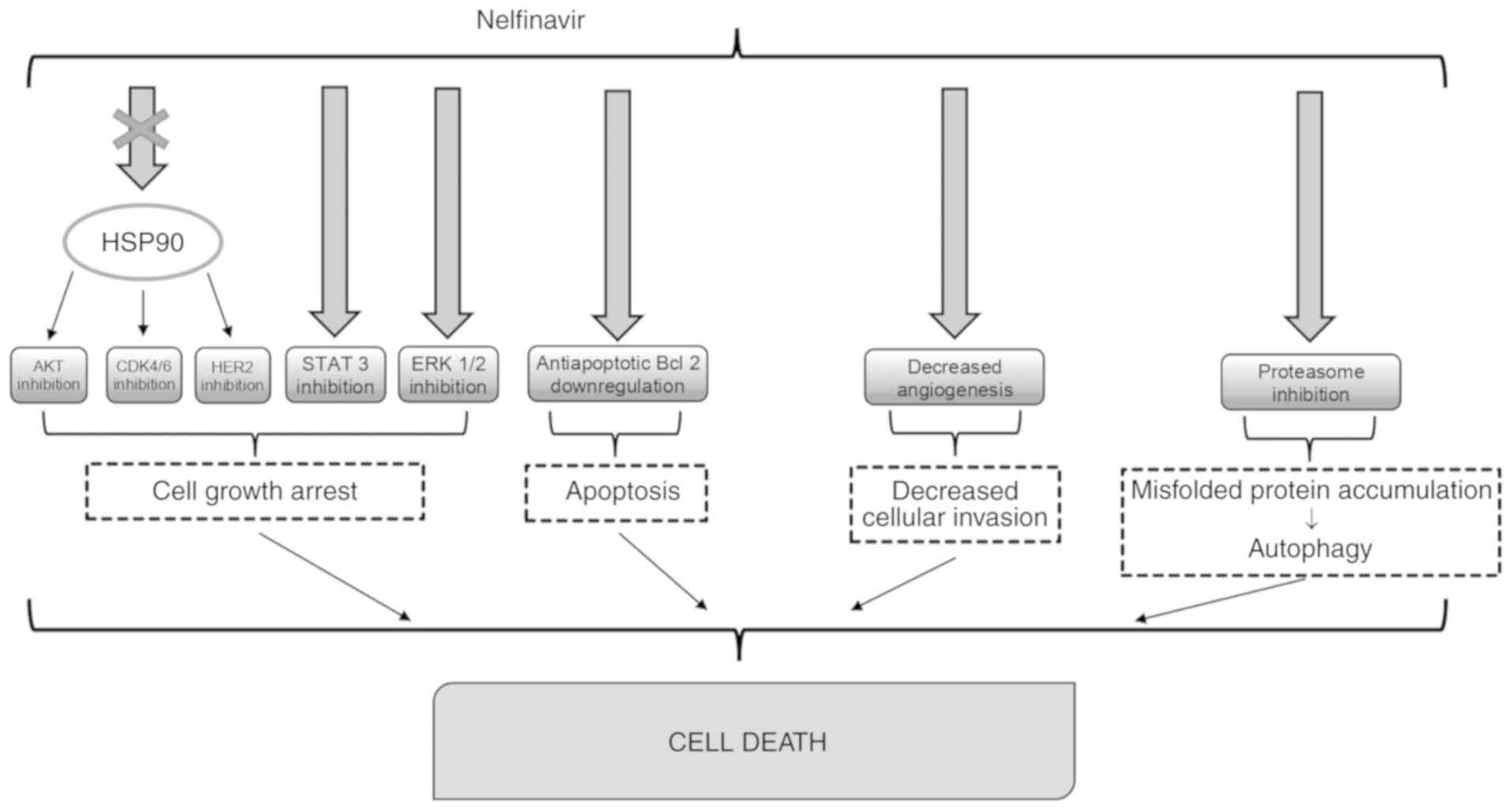
Currently, NFV is infrequently employed for
antiretroviral therapy, as it is being replaced by
second-generation HIV PIs but has demonstrated beneficial effects
in tumor patients with or without HIV infection (Fig. 2).
A previous study demonstrated that PIs have action
against cancer cells, with NFV being the most powerful among the
drugs tested (16). A complete
computational study of protein interactions has discovered 92
predicted cellular targets of NFV, among which there were 7 with
the highest binding affinities and they were aspartyl proteases
(18). The residual targets were
growth factor receptors that regulate NF-κB, Akt and other
signaling molecules (19).
Indeed, NFV appears to be the most effective
inhibitor of Akt, even though this differs by cellular type
(20). In rapamycin-resistant
diffuse large B cell lymphoma cell lines in which Akt was
upregulated, the use of NFV with Akt inhibitor MK-2206 resulted in
increased cytotoxicity (21,22).
A different mechanism through which NFV could
express its antineoplastic action could be its action on
angiogenesis. NFV can block angiogenesis via the downregulation of
PI3K/Akt, which regulates the expression of VEGF and other elements
implicated in cancer neovascularization (10). However, the concentration of Akt
does not constantly correlate with the anticancer action, and in
some experimental models NFV paradoxically stimulated Akt (23). This may result from a reduction in
growth factors or an increase in endoplasmic reticulum (ER) stress.
In fact, the most relevant anticancer action of NFV is due to ER
stress and unfolded protein response (UPR) which may be one of the
means resulting in cell death (24). NFV blocks the proteases S1P and S2P
that are implicated in SREBP-1 maturation and other proteases
essential for protein folding in ER (25).
Shim et al and Srirangam et al studied
different breast cancer cell lines and demonstrated that NFV
reduced the growth of human epidermal growth factor receptor 2
(HER2)-positive breast cancer cells when compared to HER2-negative
cells. In HER2-positive breast cancer cells, NFV provoked a
degradation of HER2 and Akt by blocking their connection with heath
shock protein 90 (HSP90) (26,27).
Nelfinavir and cancer
As stated above, NFV promotes cell death in tumor
cells by different mechanisms, including necrosis, apoptosis and
autophagy (16,28). The accumulation of misfolded
proteins could be the main mechanism of action of NFV in glioma,
ovarian cancer cells, and lung cancer (28–30).
In breast cancer cells, tamoxifen increases the
antitumor action of NFV. This synergic effect was unconnected to
the estrogen receptor status so that the combination of NFV and
tamoxifen may be useful even in subjects with no hormone responsive
tumors (31) (Table I).
 | Table I.Studies demonstrating the effects of
nelfinavir on cancer. |
Table I.
Studies demonstrating the effects of
nelfinavir on cancer.
| Cancer cells | Actions | (Refs.) |
|---|
| Breast cancer
cells | Synergic action
with tamoxifen | 31 |
| Cervical cancer
cells | Cell cycle
arrest | 32 |
| Head and neck
tumor |
Radiosensitization | 33 |
| Adenoid cystic
cancer | Reduced cell
growth | 34 |
In a different study, the proteasome inhibitor
bortezomib and NFV were used in experiments with human cervical
cancer cells. Both substances provoked cell cycle arrest in tumor
cells. An increase in the molecular chaperone BiP and in cell
stress marker ATF3 suggested the induction of UPR as the main
mechanism of cell death in tumor cells. NFV showed no actions on
proteasomal activity in the tumor cells. Even when NFV and
bortezomib were active on cisplatin-resistant cervical cancer
cells, neither of the two substances provoked a sensitization to
cisplatin therapy. Instead, both drugs augmented the effectiveness
of an apoptosis-inducing TRAIL receptor antibody (32).
In head and neck tumors, NFV promoted a reduction in
Akt and radiosensitization (33),
while in adenoid cystic cancer, NFV reduced Akt and MAPK (34), and decreased oral cell growth,
including normal keratinocytes and squamous cancer cells (35).
Finally, there is evidence that NFV is able to act
on pancreatic tumors (36), while
data have demonstrated the ability of NFV to sensitize tumor cells
to chemoradiotherapy (36).
Nelfinavir and haematological
malignancies
Several studies have demonstrated the possibility of
NFV to take action not only on solid neoplasms but also on
haematological malignancies (Table
II) such as acute myeloid leukemia (AML).
| Table II.Studies demonstrating the effects of
nelfinavir on hematologic malignancies. |
Table II.
Studies demonstrating the effects of
nelfinavir on hematologic malignancies.
| Tumor cell lines
and patients | Effect | (Refs.) |
|---|
| Leukemia cells | Increased cell
death | 37 |
| Primary acute
myeloid leukemia | Increased
apoptosis | 38 |
| Pediatric leukemia
cells | Proliferation
inhibition | 40 |
| Chronic lymphoid
leukemia | Increase of
autophagy | 41 |
| Diffuse large B
cell lymphoma cell lines | Inhibition of cell
viability | 21 |
| MM cell lines | Increased cell
death | 47 |
| Phase I trial | Overcomes
proteasome inhibition resistance in MM patients | 60 |
| Phase I/II trial in
MM patients | Overcomes
lenalidomide resistance in MM patients | 66 |
| Phase II trial in
MM patients | ORR 65% | 61 |
Brüning et al assessed the action of NFV on
leukemia cells and non-malignant bone marrow-derived cells. At a
dosage of 9 µg/ml, NFV caused 90% cell death of IM9, HL60, and
Jurkat cells. At similar levels, less than 10% of non-malignant
bone marrow-derived cells displayed NFV-provoked cell damage.
NFV-provoked death of leukemia cells was preceded by an increase in
caspases 3, 7 and 8. However, despite caspase activation, the
increase in the antiapoptotic bcl-2 family member protein mcl-1
that followed the NFV treatment stabilized the mitochondrial
membrane potential, causing mitochondrial-independent cell death.
Reduction in mcl-1 expression using sorafenib increased NFV-induced
apoptosis even at minor NFV levels but did not have supplementary
negative actions on non-malignant bone marrow cells (37).
Similarly, NFV exhibited a cytotoxic effect against
primary AML cells, stimulated PS-caused apoptosis, blocked
AKT-phosphorylation and demonstrated synergistic cytotoxicity with
carfilzomib and bortezomib at micromolar levels (38). NFV blocked intracellular proteasome
activity, including β2 proteasome activity that was not affected by
bortezomib and carfilzomib (39).
The presence of NFV-caused cytotoxicity was also
reported in pediatric leukemia cells. NFV was tested against
pediatric leukemia cells by in vitro proliferation
inhibition essays. Several substances were recognized to have a
synergistic effect with NFV in its antileukemic activity such as
AT101 (Bcl-2 family inhibitor), sunitinib (TK inhibitor), and JQ1
(BET inhibitor) (40).
NFV has also been shown to have a possible
therapeutic action in lymphoproliferative disorders. NFV provokes
moderate ER stress and autophagy in chronic lymphoid leukemia (CLL)
cells. Remarkably, NFV did not cause direct cytotoxicity against
CLL cells as a single drug. Nevertheless, co-treatment with NFV and
chloroquine markedly provoked the direct death of CLL cells in
vitro (41).
Diffuse large B cell lymphoma (DLBCL) is the most
frequent type of Non-Hodgkin's lymphoma (NHL) (42). The mTOR pathway is constitutively
stimulated in DLBCL, and blockage of mTOR is a possible treatment
for DLBCL (43), although the
response to mTOR inhibitors (mTORi) is approximately 30% in DLBCL
(44) due to the onset of
resistance to mTORi (45).
Petrich et al assessed DLBCL cell lines with
differential resistance to the mTORi. Then the authors assessed NFV
and MK-2006, chosen for their potential to synergize with rapamycin
in DLBCL. Both substances demonstrated synergistic inhibition of
cell viability in combination with rapamycin in DLBCL cell lines,
and strongly inhibited targets of activated mTOR (21).
Another research study investigated the possibility
that IPs may modify the kinetics of drugs employed in lymphoma
treatment. Comprehensive pharmacokinetics and pharmacodynamic
analysis were carried out in 19 NHL subjects during 38 cycles of
chemotherapy: 19 cycles with CHOP and 19 CHOP + HAART. Highly
active antiretroviral therapy (HAART) comprised NFV, saquinavir
(SQV), and indinavir (IDV). No substantial actions of HAART on the
pharmacokinetic values of doxorubicin (DOX) were described.
Similarly, no differential action on DOX pharmacokinetics among
NFV, SQV, and IDV was demonstrated (46).
Nelfinavir and multiple myeloma
The hematological pathology in which the effects of
NFV have been most studied is represented by multiple myeloma (MM).
Ikezoe et al demonstrated that NFV promoted suppression of
proliferation and cell death of MM RPMI8226, U266, and ARH77 cell
lines. This event occurred in association with a decrease in the
antiapoptotic protein Mcl-1. Moreover, NFV suppressed the survival
of isolated MM cells from subjects. However, NFV did not influence
survival of normal bone marrow (BM)-derived cells and colony
formation of myeloid committed stem cells (CFU-GM) from control
subjects. Furthermore, the authors observed that NFV reduced
interleukin-6 (IL-6)-stimulated phosphorylation of both signal
transducer and activator of transcription 3 (STAT3) in MM cells and
decreased basal and IL-6-stimulated STAT3/DNA binding activity
(47).
A relevant aspect of NFV action is represented by
the ability of PIs to act synergistically with the drugs used in
the treatment of MM. It was demonstrated that NFV and bortezomib
(BZ) synergistically increased proteotoxicity, reduced cell growth
and provoked cell death in MM cells. The combination of the two
drugs increased activating transcription factor (ATF)3 and
CCAAT-enhancer binding protein homologous protein (CHOP), markers
of ER stress, while their siRNA-mediated knockdown reduced cell
death. Pre-treatment with cycloheximide (a protein synthesis
inhibitor), reduced the concentrations of ubiquitinated proteins,
CHOP and ATF3, indicating that reduction in protein synthesis
augments cell survival by reducing proteotoxic stress. The use of
NFV/BZ was found to reduce the proliferation of non-small cell lung
carcinoma (NSCLC) xenografts, which was associated with the
increase in markers of ER stress and cell death (48).
Proteasome inhibitors are the mainstay of MM
treatment. Nevertheless, certain MM patients acquire proteasome
inhibitor resistance. It is well known that the proteasome
inhibitor sensitivity of MM cells is controlled by UPR (49,50),
that avoids accumulation of altered proteins in the ER by operating
on mRNA translation and protein destruction. This is controlled by
ER-associated degradation machinery, with the proteasome as its
rate-limiting terminal protease (51). Extreme activation of UPR causes
apoptosis and this is the main mechanism of action of BZ in MM
patients (52). The concentration
of UPR pre-activation regulates the proteasome
inhibitor-sensitivity of MM, so that an increase in UPR may
overcome proteasome inhibitor resistance (53). Activation of UPR is started via
inositol-requiring kinase 1 (IRE1) (54). Full plasma cell maturation requires
UPR activation via the IRE1/XBP1 axis and causes a mature,
proteasome inhibitor-sensitive MM cell type. On the contrary,
IRE1-/XBP1-MM cells are proteasome inhibitor-resistant and are
increased in proteasome inhibitor-resistant MM subjects. However,
while IRE1-targeting drugs are at an early stage of assessment
(55), it has been demonstrated
that NFV has UPR- and IRE1/XBP1-stimulating activity (24,56).
This activity may interest interference with UPR-stimulating
proteases (57), the AKT pathway
(58) and the proteasome (59). It is able to re-sensitize proteasome
inhibitor-resistant MM cells at low micromolar levels (38).
In addition to preclinical studies, numerous
clinical trials have attempted to demonstrate the efficacy of NFV
in patients with MM. In a phase I trial (SAKK 65/08), the authors
demonstrated that NFV blocked proteasome activity and increased the
amount of proteins correlated to UPR in peripheral blood
mononuclear cells. Contemporary use of NFV with BZ further
stimulated UPR and overwhelmed proteasome inhibitor resistance
(60).
In another study, the authors performed a phase 2
trial (SAKK 39/13) to study the effects and safety of NFV in MM
refractory subjects to proteasome inhibitors and previously treated
with immunomodulatory drugs. The protocol provided for
administration of NFV 2,500 mg on days 1–14 twice daily; bortezomib
1.3 mg/m2 i.v./subcutaneously on days 1, 4, 8 and 11;
and dexamethasone 20 mg orally on days 1–2, 4–5, 8–9 and 11–12 for
up to six 21-day cycles. The results of the study were remarkable.
Patients were heavily pre-treated, but ORR was 65%, a rate
comparable to the one reported in first-line BZ-naïve subjects.
Moreover, it is possible that clinical advantage was underestimated
as the treatment was administered only for 4.2 months (61).
NFV has proteasome-blocking action at high levels
(20–40 µM) in vitro (62),
but peak NFV levels at the 2×1,875 mg dose are in the 15 µM range
in treated subjects (60). Besse
et al conjectured that adjoining lenalidomide to NFV therapy
may augment intracellular NFV levels necessarily to cause the
pan-proteasome-blocking action seen with high levels of NFV. Both
NFV and lenalidomide are substrates of MDR-1 type drug efflux pumps
(63); thus, competition of the two
substances for the MDR-1 drug pump may reduce the efflux of NFV.
Similarly, NFV may augment the efficacy of the treatment by
increasing intracellular lenalidomide levels within myeloma cells
(63). Similar considerations can
also be made for the simultaneous administration of NFV with
carfilzomib, a different proteasome inhibitor (64,65).
A phase I/II trial evaluated whether adjoining NFV
to lenalidomide-dexamethasone can overcome lenalidomide resistance
in MM patients. Twenty-nine subjects were studied (lenalidomide-BZ
double-refractory 34%). They were treated with four cycles of NFV
2,500 mg/day with lenalidomide 25 mg days 1–21 and dexamethasone
(40/20 mg days 1, 8, 15 and 22). It was shown that a minor response
was attained in 55% of patients, while a partial response was
achieved in 9 patients (31%). Median overall survival was 21.6
months. Peripheral blood mononuclear cells exhibited a 45% (95% CI,
40–51%) decrease in total proteasome activity and substantial
increase of UPR and autophagy. Thus, NFV/lenalidomide/dexamethasone
appears to be an active oral treatment for lenalidomide-refractory
MM (66).
Nelfinavir and its disadvantages
Severe adverse effects of PIs are infrequent with
the exclusion of diarrhea when employed at high dosages.
Nevertheless, there are other unfavorable side events with PIs,
such as insulin resistance, hyperlipidemia and lypodystrophy. The
main factor underlying these side events is the suppression of the
breakdown of sterol regulatory element binding proteins (SREBP) in
the liver and adipose tissues causing an increased cholesterol and
fatty acid biosynthesis. SREBP storage in adipose tissue provokes
lipodystrophy. Moreover, PIs reduce proteasome-mediated breakdown
of lipoprotein (apo) B, causing an increase in the production of
triglyceride. Finally, NFV also reduces storage of the glucose
transporter GLUT-4 in adipose tissue. This may promote the onset of
insulin resistance and diabetes (67).
Moreover, an increase in serum aminotransferase may
occur in subjects receiving NFV. Considerable increase in serum
aminotransferase levels (>5 times) is present in 3–10% of
subjects. This increase is generally asymptomatic and self-limited
and does not require suspension of the drug. Hepatomegaly and
hepatic steatosis are direct effects of the metabolic changes
reported above (68). Clinical
features of hypersensitivity such as rash, fever, of eosinophilia
can arise as autoantibody formation but these events are not very
noticeable. Finally, it was reported that myelosuppression is more
common in subjects treated with chemotherapy and HAART combination
(69).
However, despite the substantially good tolerability
of treatment with NFV, some issues must be solved. Although NFV has
pro-apoptotic activity on tumor cells, an increase was reported in
the antiapoptotic mitochondrial membrane protein mcl-1 able to
increase phosphorylation of ERK1/2 (extracellular signal-regulated
kinases 1/2) (70). Upregulation of
ERK is able to reduce cell death. This condition can be solved with
the administration of sorafenib (37,70).
Moreover, in spite of the antitumor action of NFV,
this drug does not decrease the risk of tumor onset in HIV patients
and also causes a reduction in immunological functions, altering
the differentiation of monocytes into dendritic cells (71,72).
Finally, although the results from numerous studies demonstrate
that the combination of HAART with chemotherapy improves prognosis,
many uncertainties exist on the choice of the best combination of
chemotherapy and PIs. Overlapping toxicities have been reported
with combination treatment, and there is a possible risk for
pharmacokinetic associations between chemotherapy and PIs (73).
Future perspectives
Although the use of NFV has given encouraging
results in in vitro and in vivo studies, even better
results could be achieved by using drug combinations. Akt
activation plays a main role in the tumor phenotype (74). At present there are no substances
able to inhibit this protein with a convenient safety profile. The
anti-Akt action of NFV can be increased by simultaneous mTOR
blockage which provokes a synergistic cytotoxicity (21). This may be since mTOR inhibition
without Akt blockage removes a negative biofeedback loop on Akt,
causing increased phosphorylation of Akt (75,76).
The negative bio-feedback loop on Akt must be solved to achieve
appropriate results. NFV could be useful to obtain better results
when used with mTORi.
Numerous other substances have been identified as
possible synergists with NFV. NFV increases anti-malarial activity
of artemisinin in vitro on Plasmodium falciparum, but
artemisinin also has antitumor action (77). Several studies have tried to
evaluate whether the simultaneous administration of artemisinin and
NFV could enhance the antineoplastic action of the two substances.
A research study employed NFV and artemisinin, in an experimental
protocol (CUSP 9) for the therapy of relapsed glioblastoma. The
combination was reported to postpone glioblastoma spread (78).
Experimental tests are however necessary to verify
the possibility that the use of NFV with other drugs such as
celecoxib or chloroquine, may be effective for the treatment of
neoplasms. In fact, the cancer cell killing capacity of NFV can be
increased with different ER stressors such as celecoxib (79); perillyl alcohol is another stress
factor that has been employed using this rationale (80).
Hydroxicloroquine and chloroquine are autophagy
inhibitors and may also operate synergistically with NFV,
decreasing autophagy and augmenting apoptosis (41,81,82).
Conclusions
Drug repositioning leads to the identification of
new indications for current drugs and the use of the newly
recognized medicines to therapy of diseases other than the drug's
intended target. Even though the exact molecular target of NFV is
still to be identified, its efficacy and safety are well known. All
data reported in this review support the hypothesis that NFV is a
useful means of integration in cancer treatment. In the future, it
should be evaluated in combination with chemotherapy in the design
of new protocols. Indeed, it is fundamental to establish the most
appropriate associations, dosing and timing of NFV administration
in patients with MM or other hematological diseases. In fact,
concurrent therapy with PIs is not without drug interactions. As
such, clinical decisions regarding therapy should be carefully
evaluated, and dose adjustments must be made to reduce the risk for
adverse outcomes and disease progression. Moreover, new PIs are
being created with better anticancer activity and further
development of new PIs with stronger anticancer activity will be
realized in the future.
Despite the above, research for the use of
antivirals in the treatment of hematologic neoplasias and in
particular MM is ongoing. In fact, notwithstanding recent advanced
therapy opportunities such as proteasome inhibitors, histone
deacetylase inhibitors, immunomodulatory drugs and immunotherapy,
and myeloma-targeted antibodies (83–87),
MM is still judged as an incurable disease.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All information was cited by relevant references
listed in the Review.
Authors' contributions
Conceptualization of the review was carried out by
AA and CM. Curation of the data was conducted by VI, AGA, MP and
NP. Writing of the original draft was undertaken by AA. Writing of
the review and editing were carried out by AA and CM. All authors
read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interest
The authors state that they have no competing
interests.
Authors' information
Alessandro Allegra, Orcid: 0000-0001-6156-8239.
References
|
1
|
Gantt S, Casper C and Ambinder RF:
Insights into the broad cellular effects of nelfinavir and the HIV
protease inhibitors supporting their role in cancer treatment and
prevention. Curr Opin Oncol. 25:495–502. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Niehues T, Horneff G, Megahed M, Schroten
H and Wahn V: Complete regression of AIDS-related Kaposi's sarcoma
in a child treated with highly active antiretroviral therapy. AIDS.
13:1148–1149. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sgadari C, Monini P, Barillari G and
Ensoli B: Use of HIV protease inhibitors to block Kaposi's sarcoma
and tumour growth. Lancet Oncol. 4:537–547. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Xing F, Liu Y, Wu SY, Wu K, Sharma S, Mo
YY, Feng J, Sanders S, Jin G, Singh R, et al: Loss of XIST in
breast cancer activates MSN-c-Met and reprograms microglia via
Exosomal miRNA to promote brain metastasis. Cancer Res.
78:4316–4330. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ikezoe T, Daar ES, Hisatake J, Taguchi H
and Koeffler HP: HIV-1 protease inhibitors decrease proliferation
and induce differentiation of human myelocytic leukemia cells.
Blood. 96:3553–3559. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pati S, Pelser CB, Dufraine J, Bryant JL,
Reitz MS Jr and Weichold FF: Antitumorigenic effects of HIV
protease inhibitor ritonavir: Inhibition of Kaposi sarcoma. Blood.
99:3771–3779. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Okubo K, Isono M, Asano T and Sato A:
Lopinavir-ritonavir combination induces endoplasmic reticulum
stress and kills urological cancer cells. Anticancer Res.
39:5891–5901. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lu M, Dong H, Bao D, Liu B and Liu H:
Tenofovir disoproxil fumarate induces pheochromocytoma cells
apoptosis. Eur J Pharmacol. 844:139–144. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Esposito V, Verdina A, Manente L, Spugnini
EP, Viglietti R, Parrella R, Pagliano P, Parrella G, Galati R, De
Luca A, et al: Amprenavir inhibits the migration in human
hepatocarcinoma cell and the growth of xenografts. J Cell Physiol.
228:640–645. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pore N, Gupta AK, Cerniglia GJ and Maity
A: HIV protease inhibitors decrease VEGF/HIF-1alpha expression and
angiogenesis in glioblastoma cells. Neoplasia. 8:889–895. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sun L, Niu L, Zhu X, Hao J, Wang P and
Wang H: Antitumour effects of a protease inhibitor, nelfinavir, in
hepatocellular carcinoma cancer cells. J Chemother. 24:161–166.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dewan MZ, Uchihara JN, Terashima K, Honda
M, Sata T, Ito M, Fujii N, Uozumi K, Tsukasaki K, Tomonaga M, et
al: Efficient intervention of growth and infiltration of primary
adult T-cell leukemia cells by an HIV protease inhibitor,
ritonavir. Blood. 107:716–724. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Srirangam A, Milani M, Mitra R, Guo Z,
Rodriguez M, Kathuria H, Fukuda S, Rizzardi A, Schmechel S, Skalnik
DG, et al: The human immunodeficiency virus protease inhibitor
ritonavir inhibits lung cancer cells, in part, by inhibition of
survivin. J Thorac Oncol. 6:661–670. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Timeus F, Crescenzio N, Doria A, Foglia L,
Pagliano S, Ricotti E, Fagioli F, Tovo PA and Cordero di
Montezemolo L: In vitro anti-neuroblastoma activity of
saquinavir and its association with imatinib. Oncol Rep.
27:734–740. 2012.PubMed/NCBI
|
|
15
|
Weichold FF, Bryant JL, Pati S,
Barabitskaya O, Gallo RC and Reitz MS Jr: HIV-1 protease inhibitor
ritonavir modulates susceptibility to apoptosis of uninfected T
cells. J Hum Virol. 2:261–269. 1999.PubMed/NCBI
|
|
16
|
Gills JJ, Lopiccolo J, Tsurutani J,
Shoemaker RH, Best CJ, Abu-Asab MS, Borojerdi J, Warfel NA, Gardner
ER, Danish M, et al: Nelfinavir, A lead HIV protease inhibitor, is
a broad-spectrum, anticancer agent that induces endoplasmic
reticulum stress, autophagy, and apoptosis in vitro and in vivo.
Clin Cancer Res. 13:5183–5194. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
National Institute of Diabetes and
Digestive and Kidney Diseases. Protease Inhibitors (HIV). LiverTox,
. Clinical and research information on drug-induced liver injury
[Internet]. PMID: 31644200. Bethesda (MD): Sep 1–2012-2017
|
|
18
|
Xie L, Evangelidis T, Xie L and Bourne PE:
Drug discovery using chemical systems biology: Weak inhibition of
multiple kinases may contribute to the anti-cancer effect of
nelfinavir. PLoS Comput Biol. 7:e10020372011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gills JJ, Lopiccolo J and Dennis PA:
Nelfinavir, a new anti-cancer drug with pleiotropic effects and
many paths to autophagy. Autophagy. 4:107–109. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kraus M, Bader J, Overkleeft H and
Driessen C: Nelfinavir augments proteasome inhibition by bortezomib
in myeloma cells and overcomes bortezomib and carfilzomib
resistance. Blood Cancer J. 3:e1032013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Petrich AM, Leshchenko V, Kuo PY, Xia B,
Thirukonda VK, Ulahannan N, Gordon S, Fazzari MJ, Ye BH, Sparano JA
and Parekh S: Akt inhibitors MK-2206 and nelfinavir overcome mTOR
inhibitor resistance in diffuse large B-cell lymphoma. Clin Cancer
Res. 18:2534–2544. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Plastaras JP, Vapiwala N, Ahmed MS,
Gudonis D, Cerniglia GJ, Feldman MD, Frank I and Gupta AK:
Validation and toxicity of PI3K/Akt pathway inhibition by HIV
protease inhibitors in humans. Cancer Biol Ther. 7:628–635. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jiang W, Mikochik PJ, Ra JH, Lei H,
Flaherty KT, Winkler JD and Spitz FR: HIV protease inhibitor
nelfinavir inhibits growth of human melanoma cells by induction of
cell cycle arrest. Cancer Res. 67:1221–1227. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bernstein WB and Dennis PA: Repositioning
HIV protease inhibitors as cancer therapeutics. Curr Opin HIV AIDS.
3:666–675. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guan M, Fousek K and Chow WA: Nelfinavir
inhibits regulated intramembrane proteolysis of sterol regulatory
element binding protein-1 and activating transcription factor 6 in
castration-resistant prostate cancer. FEBS J. 279:2399–2411. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shim JS, Rao R, Beebe K, Neckers L, Han I,
Nahta R and Liu JO: Selective inhibition of HER2-positive breast
cancer cells by the HIV protease inhibitor nelfinavir. J Natl
Cancer Inst. 104:1576–1590. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Srirangam A, Mitra R, Wang M, Gorski JC,
Badve S, Baldridge L, Hamilton J, Kishimoto H, Hawes J, Li L, et
al: Effects of HIV protease inhibitor ritonavir on Akt-regulated
cell proliferation in breast cancer. Clin Cancer Res. 12:1883–1896.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Pyrko P, Kardosh A, Wang W, Xiong W,
Schönthal AH and Chen TC: HIV-1 protease inhibitors nelfinavir and
atazanavir induce malignant glioma death by triggering endoplasmic
reticulum stress. Cancer Res. 67:10920–10928. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Peñaranda Fajardo NM, Meijer C and Kruyt
FA: The endoplasmic reticulum stress/unfolded protein response in
gliomagenesis, tumor progression and as a therapeutic target in
glioblastoma. Biochem Pharmacol. 118:1–8. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Brüning A, Burger P, Vogel M, Rahmeh M,
Gingelmaiers A, Friese K, Lenhard M and Burges A: Nelfinavir
induces the unfolded protein response in ovarian cancer cells,
resulting in ER vacuolization, cell cycle retardation and
apoptosis. Cancer Biol Ther. 8:226–232. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Brüning A, Friese K, Burges A and Mylonas
I: Tamoxifen enhances the cytotoxic effects of nelfinavir in breast
cancer cells. Breast Cancer Res. 12:R452010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Bruning A, Vogel M, Mylonas I, Friese K
and Burges A: Bortezomib targets the caspase-like proteasome
activity in cervical cancer cells, triggering apoptosis that can be
enhanced by nelfinavir. Curr Cancer Drug Targets. 11:799–809. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gupta AK, Lee JH, Wilke WW, Quon H, Smith
G, Maity A, Buatti JM and Spitz DR: Radiation response in two
HPV-infected head-and-neck cancer cell lines in comparison to a
non-HPV-infected cell line and relationship to signaling through
AKT. Int J Radiat Oncol Biol Phys. 74:928–933. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gupta AK, Wilke WW, Taylor EN, Bodeker KL,
Hoffman HT, Milhem MM, Buatti JM and Robinson RA: Signaling
pathways in adenoid cystic cancers: Implications for treatment.
Cancer Biol Ther. 8:1947–1951. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Danaher RJ, Wang C, Roland AT, Kaetzel CS,
Greenberg RN and Miller CS: HIV protease inhibitors block oral
epithelial cell DNA synthesis. Arch Oral Biol. 55:95–100. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kimple RJ, Vaseva AV, Cox AD, Baerman KM,
Calvo BF, Tepper JE, Shields JM and Sartor CI: Radiosensitization
of epidermal growth factor receptor/HER2-positive pancreatic cancer
is mediated by inhibition of Akt independent of ras mutational
status. Clin Cancer Res. 16:912–923. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Brüning A, Rahmeh M, Gingelmaier A and
Friese K: The mitochondria-independent cytotoxic effect of
nelfinavir on leukemia cells can be enhanced by sorafenib-mediated
mcl-1 downregulation and mitochondrial membrane destabilization.
Mol Cancer. 9:192010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kraus M, Müller-Ide H, Rückrich T, Bader
J, Overkleeft H and Driessen C: Ritonavir, nelfinavir, saquinavir
and lopinavir induce proteotoxic stress in acute myeloid leukemia
cells and sensitize them for proteasome inhibitor treatment at low
micromolar drug concentrations. Leuk Res. 38:383–392. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Allegra A, Alonci A, Gerace D, Russo S,
Innao V, Calabrò L and Musolino C: New orally active proteasome
inhibitors in multiple myeloma. Leuk Res. 38:1–9. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Meier-Stephenson V, Riemer J and Narendran
A: The HIV protease inhibitor, nelfinavir, as a novel therapeutic
approach for the treatment of refractory pediatric leukemia. Onco
Targets Ther. 10:2581–2593. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Mahoney E, Maddocks K, Flynn J, Jones J,
Cole SL, Zhang X, Byrd JC and Johnson AJ: Identification of
endoplasmic reticulum stress-inducing agents by antagonizing
autophagy: A new potential strategy for identification of
anti-cancer therapeutics in B-cell malignancies. Leuk Lymphoma.
54:2685–2692. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Falduto A, Cimino F, Speciale A, Musolino
C, Gangemi S, Saija A and Allegra A: How gene polymorphisms can
influence clinical response and toxicity following R-CHOP therapy
in patients with diffuse large B cell lymphoma. Blood Rev.
31:235–249. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hess G, Smith SM, Berkenblit A and
Coiffier B: Temsirolimus in mantle cell lymphoma and other
non-Hodgkin lymphoma subtypes. Semin Oncol. 36 (Suppl 3):S37–S45.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Smith SM, van Besien K, Karrison T, Dancey
J, McLaughlin P, Younes A, Smith S, Stiff P, Lester E, Modi S, et
al: Temsirolimus has activity in non-mantle cell non-Hodgkin's
lymphoma subtypes: The University of Chicago phase II consortium. J
Clin Oncol. 28:4740–4746. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Witzig TE, Reeder CB, LaPlant BR, Gupta M,
Johnston PB, Micallef IN, Porrata LF, Ansell SM, Colgan JP,
Jacobsen ED, et al: A phase II trial of the oral mTOR inhibitor
everolimus in relapsed aggressive lymphoma. Leukemia. 25:341–347.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Toffoli G, Corona G, Cattarossi G,
Boiocchi M, Di Gennaro G, Tirelli U and Vaccher E: Effect of highly
active antiretroviral therapy (HAART) on pharmacokinetics and
pharmacodynamics of doxorubicin in patients with HIV-associated
non-Hodgkin's lymphoma. Ann Oncol. 15:1805–1809. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ikezoe T, Saito T, Bandobashi K, Yang Y,
Koeffler HP and Taguchi I: HIV-1 protease inhibitor induces growth
arrest and apoptosis of human multiple myeloma cells via
inactivation of signal transducer and activator of transcription 3
and extracellular signal-regulated kinase 1/2. Mol Cancer Ther.
3:473–479. 2004.PubMed/NCBI
|
|
48
|
Kawabata S, Gills JJ, Mercado-Matos JR,
LoPiccolo J, Wilson III W, Hollander MC and Dennis PA: Synergistic
effects of nelfinavir and bortezomib on proteotoxic death of NSCLC
and multiple myeloma cells. Cell Death and Disease. 3:e3532012.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Leung-Hagesteijn C, Erdmann N, Cheung G,
Keats JJ, Stewart AK, Reece DE, Chung KC and Tiedemann RE:
Xbp1s-negative tumor B cells and pre-plasmablasts mediate
therapeutic proteasome inhibitor resistance in multiple myeloma.
Cancer Cell. 24:289–304. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Neznanov N, Komarov AP, Neznanova L,
Stanhope-Baker P and Gudkov AV: Proteotoxic stress targeted therapy
(PSTT): Induction of protein misfolding enhances the antitumor
effect of the proteasome inhibitor bortezomib. Oncotarget.
2:209–221. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Mayor T: Navigating the ERAD interaction
network. Nat Cell Biol. 14:46–47. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Obeng EA, Carlson LM, Gutman DM,
Harrington WJ Jr, Lee KP and Boise LH: Proteasome inhibitors induce
a terminal unfolded protein response in multiple myeloma cells.
Blood. 107:4907–4916. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Ling SC, Lau EK, Al-Shabeeb A, Nikolic A,
Catalano A, Iland H, Horvath N, Ho PJ, Harrison S, Fleming S, et
al: Response of myeloma to the proteasome inhibitor bortezomib is
correlated with the unfolded protein response regulator XBP-1.
Haematologica. 97:64–72. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Reimold AM, Iwakoshi NN, Manis J,
Vallabhajosyula P, Szomolanyi-Tsuda E, Gravallese EM, Friend D,
Grusby MJ, Alt F and Glimcher LH: Plasma cell differentiation
requires the transcription factor XBP-1. Nature. 412:300–307. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Papandreou I, Denko NC, Olson M, Van
Melckebeke H, Lust S, Tam A, Solow-Cordero DE, Bouley DM, Offner F,
Niwa M and Koong AC: Identification of an Ire1alpha endonuclease
specific inhibitor with cytotoxic activity against human multiple
myeloma. Blood. 117:1311–1314. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Chow WA, Jiang C and Guan M: Anti-HIV
drugs for cancer therapeutics: Back to the future? Lancet Oncol.
10:61–71. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Guan M, Fousek K, Jiang C, Guo S, Synold
T, Xi B, Shih CC and Chow WA: Nelfinavir induces liposarcoma
apoptosis through inhibition of regulated intramembrane proteolysis
of SREBP-1 and ATF6. Clin Cancer Res. 17:1796–1806. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yang Y, Ikezoe T, Nishioka C, Bandobashi
K, Takeuchi T, Adachi Y, Kobayashi M, Takeuchi S, Koeffler HP and
Taguchi H: NFV, an HIV-1 protease inhibitor, induces growth arrest,
reduced Akt signalling, apoptosis and docetaxel sensitisation in
NSCLC cell lines. Br J Cancer. 95:1653–1662. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Bono C, Karlin L, Harel S, Mouly E,
Labaume S, Galicier L, Apcher S, Sauvageon H, Fermand JP, Bories JC
and Arnulf B: The human immunodeficiency virus-1 protease inhibitor
nelfinavir impairs proteasome activity and inhibits the
proliferation of multiple myeloma cells in vitro and in vivo.
Haematologica. 97:1101–1109. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Driessen C, Kraus M, Joerger M, Rosing H,
Bader J, Hitz F, Berset C, Xyrafas A, Hawle H, Berthod G, et al:
Treatment with the HIV protease inhibitor nelfinavir triggers the
unfolded protein response and may overcome proteasome inhibitor
resistance of multiple myeloma in combination with bortezomib: A
phase I trial (SAKK 65/08). Haematologica. 101:346–355. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Driessen C, Müller R, Novak U, Cantoni N,
Betticher D, Mach N, Rüfer A, Mey U, Samaras P, Ribi K, et al:
Promising activity of nelfinavir-bortezomib-dexamethasone in
proteasome inhibitor-refractory multiple myeloma. Blood.
132:2097–2100. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Kraus M, Malenke E, Gogel J, Müller H,
Rückrich T, Overkleeft H, Ovaa H, Koscielniak E, Hartmann JT and
Driessen C: Ritonavir induces endoplasmic reticulum stress and
sensitizes sarcoma cells toward bortezomib-induced apoptosis. Mol
Cancer Ther. 7:1940–1948. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Besse A, Stolze SC, Rasche L, Weinhold N,
Morgan GJ, Kraus M, Bader J, Overkleeft HS, Besse L and Driessen C:
Carfilzomib resistance due to ABCB1/MDR1 overexpression is overcome
by nelfinavir and lopinavir in multiple myeloma. Leukemia.
32:391–401. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Besse A, Besse L, Kraus M, Mendez-Lopez M,
Bader J, Xin BT, de Bruin G, Maurits E, Overkleeft HS and Driessen
C: Proteasome inhibition in multiple myeloma: Head-to-head
comparison of currently available proteasome inhibitors. Cell Chem
Biol. 26:340–351.e3. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Hawley TS, Riz I, Yang W, Wakabayashi Y,
Depalma L, Chang YT, Peng W, Zhu J and Hawley RG: Identification of
an ABCB1 (P-glycoprotein)-positive carfilzomib-resistant myeloma
subpopulation by the pluripotent stem cell fluorescent dye CDy1. Am
J Hematol. 88:265–272. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hitz F, Kraus M, Pabst T, Hess D, Besse L,
Silzle T, Novak U, Seipel K, Rondeau S, Stüdeli S, et al:
Nelfinavir and lenalidomide/dexamethasone in patients with
lenalidomide-refractory multiple myeloma. A phase I/II Trial (SAKK
39/10). Blood Cancer J. 9:702019. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Hui DY: Effects of HIV protease inhibitor
therapy on lipid metabolism. Prog Lipid Res. 42:81–92. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Riddle TM, Kuhel DG, Woollett LA,
Fichtenbaum CJ and Hui DY: HIV protease inhibitor induces fatty
acid and sterol biosynthesis in liver and adipose tissues due to
the accumulation of activated sterol regulatory element-binding
proteins in the nucleus. J Biol Chem. 276:37514–37519. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Orem J, Fu P, Ness A, Mwanda WO and Remick
SC: Oral combination chemotherapy in the treatment of
AIDS-associated Hodgkin's disease. East Afr Med J. 82 (Suppl.
9):S144–S149. 2005.PubMed/NCBI
|
|
70
|
Brüning A, Burger P, Vogel M, Gingelmaier
A, Friese K and Burges A: Nelfinavir induces mitochondria
protection by ERK1/2-mediated mcl-1 stabilization that can be
overcome by sorafenib. Invest New Drugs. 28:535–542. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Crum-Cianflone NF, Hullsiek KH, Marconi V,
Weintrob A, Ganesan A, Barthel RV, Fraser S, Roediger MP, Agan B
and Wegner S: The impact of nelfinavir exposure on cancer
development among a large cohort of HIV-infected patients. J Acquir
Immune Defic Syndr. 51:305–309. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Giardino Torchia ML, Ciaglia E, Masci AM,
Vitiello L, Fogli M, la Sala A, Mavilio D and Racioppi L: Dendritic
cells/natural killer cross-talk: A novel target for human
immunodeficiency virus type-1 protease inhibitors. PLoS One.
5:e110522010. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Bower M, McCall-Peat N, Ryan N, Davies L,
Young AM, Gupta S, Nelson M, Gazzard B and Stebbing J: Protease
inhibitors potentiate chemotherapy-induced neutropenia. Blood.
104:2943–2946. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Gills JJ and Dennis PA: Perifosine: Update
on a novel Akt inhibitor. Curr Oncol Rep. 11:102–110. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Wan X, Harkavy B, Shen N, Grohar P and
Helman LJ: Rapamycin induces feedback activation of Akt signaling
through an IGF-1R-dependent mechanism. Oncogene. 26:1932–1940.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Carracedo A, Bacelga J and Pandolfi PP:
Deconstructing feedback-signaling networks to improve anticancer
therapy with mTORC1 inhibitors. Cell Cycle. 7:3805–3859. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Mishra LC, Bhattacharya A, Sharma M and
Bhasin VK: HIV protease inhibitors, indinavir or nelfinavir,
augment antimalarial action of artemisinin in vitro. Am J Trop Med
Hyg. 82:148–150. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Kast RE, Boockvar JA, Brüning A, Cappello
F, Chang WW, Cvek B, Dou QP, Duenas-Gonzalez A, Efferth T, Focosi
D, et al: A conceptually new treatment approach for relapsed
glioblastoma: Coordinated undermining of survival paths with nine
repurposed drugs (CUSP9 by the International Initiative for
Accelerated Improvement of Glioblastoma Care. Oncotarget.
4:502–530. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Cho HY, Thomas S, Golden EB, Gaffney KJ,
Hofman FM, Chen TC, Louie SG, Petasis NA and Schönthal AH: Enhanced
killing of chemo-resistant breast cancer cells via controlled
aggravation of ER stress. Cancer Lett. 282:87–97. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Cho HY, Wang W, Jhaveri N, Torres S, Tseng
J, Leong MN, Lee DJ, Goldkorn A, Xu T, Petasis NA, et al: Perillyl
alcohol for the treatment of temozolomide-resistant gliomas. Mol
Cancer Ther. 11:2462–2472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Homewood CA, Warhurst DC, Peters W and
Baggaley VC: Lysosomes, pH and the anti-malarial action of
chloroquine. Nature. 235:50–52. 1972. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Shintani T and Klionsky DJ: Autophagy in
health and disease: A double-edged sword. Science. 306:990–995.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Allegra A, Penna G, Alonci A, Russo S,
Greve B, Innao V, Minardi V and Musolino C: Monoclonal antibodies:
Potential new therapeutic treatment against multiple myeloma. Eur J
Haematol. 90:441–468. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Allegra A, Sant'antonio E, Penna G, Alonci
A, D'Angelo A, Russo S, Cannavò A, Gerace D and Musolino C: Novel
therapeutic strategies in multiple myeloma: Role of the heat shock
protein inhibitors. Eur J Haematol. 86:93–110. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Allegra A, Penna G, Innao V, Greve B,
Maisano V, Russo S and Musolino C: Vaccination of multiple myeloma:
Current strategies and future prospects. Crit Rev Oncol Hematol.
96:339–354. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Allegra A, Innao V, Gerace D, Vaddinelli D
and Musolino C: Adoptive immunotherapy for hematological
malignancies: Current status and new insights in chimeric antigen
receptor T cells. Blood Cells Mol Dis. 62:49–63. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Ettari R, Zappalà M, Grasso S, Musolino C,
Innao V and Allegra A: Immunoproteasome-selective and non-selective
inhibitors: A promising approach for the treatment of multiple
myeloma. Pharmacol Ther. 182:176–192. 2018. View Article : Google Scholar : PubMed/NCBI
|