Introduction
Pancreatic cancer is a one of the most highly
malignant tumors of the digestive tract and has an extremely poor
prognosis, with a 5-year relative survival rate of 9%. In 2019, an
estimated 56,770 individuals were newly diagnosed with pancreatic
cancer, which may result in ~45,750 cancer-related deaths in the
United States (1). In China, the
incidence rate of pancreatic cancer has also increased from 2000 to
2011. Pancreatic cancer accounts for the second leading upward
trend of age-standardized mortality rates (2). Metastasis is the leading cause of
pancreatic cancer-related mortality, as ~80% of the patients are
not deemed suitable for surgical resection due to early relapse or
advanced metastasis at diagnosis (3). Therefore, it is crucial to elucidate
the molecular mechanisms underlying the invasion and metastasis of
pancreatic cancer, and novel comprehensive and effective
therapeutic interventions are urgently needed to improve the
treatment outcome.
Pancreatic cancer is characterized by an excessive
desmoplastic reaction, which favors hypoxia, further inducing
epithelial-to-mesenchymal transition (EMT) and tumor metastasis
(4). It has become obvious that the
tumor microenvironment, which includes several types of stromal
cells, such as immune, inflammatory and endothelial cells, as well
as pancreatic stellate cells (PSCs), plays a key role in tumor
invasion and metastasis (5). PSCs
generally exist in one of two statuses: Quiescent and activated.
Under physiological conditions, non-activated PSCs contain abundant
cytoplasmic vitamin A-containing lipid droplets, which produce very
low levels of extracellular matrix (ECM). After activation, PSCs
are characterized by a decrease in lipid droplets and increased
expression of α-smooth muscle actin (α-SMA) and collagen-I, which
are crucial for tumor development, evasion of immune surveillance,
invasion and metastasis (6). In
addition, activated PSCs can produce a number of soluble factors,
such as interleukin (IL)-6, transforming growth factor (TGF)-β and
stromal cell-derived factor (SDF)-1, which promote the malignant
behavior of pancreatic cancer cells (7). IL-6 is a potent pro-inflammatory
cytokine secreted by PSCs in the pancreatic cancer
microenvironment. In addition to the inflammatory response, IL-6 is
also associated with tumor progression, including proliferation,
angiogenesis, chemoresistance, invasion, EMT and metastasis
(7,8). Our previous study demonstrated that
PSCs cultured under hypoxic conditions display higher levels of
IL-6, vascular endothelial growth factor (VEGF)-A and SDF-1
transcription and secretion (9).
Low oxygen tension (hypoxia), which is correlated
with poor survival of the patients, is most commonly present in the
microenvironment of pancreatic cancer. The adaptation of pancreatic
cancer cells to limited oxygen delivery promotes tumor invasion,
angiogenesis and distant metastasis at an early stage of tumor
development (10). Our previous
studies have demonstrated that hypoxic conditions can promote the
proliferation, migration, invasion and EMT of pancreatic cancer
cells in vitro by activating the Hedgehog (Hh) signaling
pathway (11). Using a
three-dimensional (3D) matrices model, Sada et al
demonstrated that the expression of hypoxia-induced 2-oxoglutarate
5-dioxygenase 2 in PSCs creates a permissive microenvironment for
cancer cell migration through architectural regulation of stromal
ECM in pancreatic cancer (12).
Curcumin, a natural polyphenol derived from
turmeric, is well known for its potential applications in the
treatment of multiple tumors, as well as for its anti-infectious,
anti-inflammatory, antioxidant and chemopreventive properties
(13). It has been confirmed that
curcumin can exert its antitumor effects via targeting multiple
signaling pathways (14). Our
previous study demonstrated that curcumin inhibited high
glucose-induced proliferative and invasive abilities of pancreatic
cancer cells via inhibiting the epidermal growth factor
(EGF)/extracellular signal-regulated kinase (ERK) and EGF/Akt
signaling pathways (15). Curcumin
has also been found to suppress hypoxia-induced proliferation,
migration and EMT by inhibiting the Hh signaling pathway in
pancreatic cancer cells (11).
However, whether curcumin can suppress the tumor-stromal crosstalk
in pancreatic cancer and the underlying mechanisms have yet to be
fully elucidated.
The aim of the present study was to investigate the
role of hypoxia in PSCs and pancreatic cancer cells, and examine
the potential protective effect of curcumin against hypoxia-induced
pancreatic cancer progression. It was also investigated whether
curcumin acts by suppressing tumor-stromal crosstalk and pancreatic
cancer metastasis through inhibiting the IL-6/ERK/nuclear factor
(NF)-κB axis, and whether it may be considered as a novel option
for the treatment of pancreatic cancer.
Materials and methods
Cell culture and reagents
Human PSCs were isolated from pancreatic tumor
tissue surgically resected from patients at the Department of
Hepatobiliary Surgery of the First Affiliated Hospital of Xi'an
Jiaotong University. PSCs were obtained and cultured according to
methods described in a previous study (16). The purity of the PSCs was evaluated
by morphology, Oil red O staining of intracellular fat droplets and
immunofluorescence of α-SMA, as described in our previous studies
(9,17). The experimental protocol and patient
consent forms were approved by the Ethics Committee of the First
Affiliated Hospital of Xi'an Jiaotong University (Xi'an, China).
The human pancreatic cancer cell lines, BxPC-3 and Panc-1, were
obtained from the American Type Culture Collection. The culture
medium was Gibco™ DMEM (Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.), 100 U/ml penicillin and 100 µg/ml streptomycin.
When the experiment was performed, the cells were cultured under
normoxic conditions (20% O2) or controlled hypoxic
conditions (1% O2) at 37°C. Exponentially growing cells
in complete medium were pretreated for 1 h with 20 µM curcumin,
followed by continual incubation with different O2
concentrations according to the purpose of the experiment.
Curcumin, MTT, IL-6 and PD 98059 were purchased from Sigma-Aldrich;
Merck KGaA. Curcumin was dissolved in dimethylsulfoxide (DMSO) and
then diluted to appropriate concentrations in culture medium. DMSO
was used as the control group. Millicell culture plate inserts for
the Transwell invasion assays were obtained from EMD Millipore.
Matrigel was purchased from BD Biosciences. Primary antibodies
[dilution 1:100 in phosphate-buffered saline (PBS)-Tween-20 (0.1%)]
against E-cadherin (cat. no. sc-52328), vimentin (cat. no.
sc-66002), IL-6 (cat. no. sc-130326) and matrix metallopeptidase
(MMP)-9 (cat. no. sc-12759) were obtained from Santa Cruz
Biotechnology, Inc. The anti-ERK (cat. no. 9102),
anti-phosphorylated (p)-ERK (Thr202/Tyr204, cat. no. 9106),
anti-NF-κB (cat. no. 6956), anti-p-NF-κB p65 (anti-p-NF-κB, Ser468,
cat. no. 3039) and anti-α-SMA (cat. no. 48938) antibodies (dilution
1:200 in PBS-Tween-20) were obtained from Cell Signaling
Technology, Inc. Other reagents were purchased from common
commercial sources. All drug solutions were freshly prepared on the
day of testing.
MTT assay
PSCs were seeded in 96-well plates at a density of
1×104 cells/well. The cells were then treated with
different concentrations (0–40 µM) of curcumin. After incubation
for 24, 48 and 72 h at 37°C, 15 µl of the MTT solution was added to
each well, and the cells were incubated for 4 h at 37°C.
Subsequently, 100 µl of DMSO were added to each well. The optical
density (OD) value at 490 nm was determined using a
spectrophotometer (Bio-Rad Laboratories, Inc.). The proliferation
inhibition rate was calculated as (1-ODsample/ODcontrol) ×100%.
Wound healing assay
PSCs (5×104 cells/500 µl) were seeded in
24-well plates. A 1-µl sterile pipette tip was used to produce a
linear wound in the cell monolayer after the cells had grown to
~90-100% confluence. After removing the cellular debris, PSCs were
allowed to migrate for 24 h under normoxic or hypoxic conditions,
with or without curcumin. Images were captured at 0 and 24 h
post-wounding using a Diaphot TMD inverted microscope (Nikon
Corporation) at a magnification of ×10. The relative distance
migrated by the leading edge was assessed using Photoshop software
(n=5).
Transwell Matrigel invasion assay
A chamber-based Transwell invasion assay was
performed to evaluate the invasive ability of pancreatic cancer
cells. Briefly, the 8.0-µm pore inserts were coated with 30 µl of
Matrigel. After serum starvation for 24 h, cancer cells
(5×104) were suspended in the upper chamber in DMEM
containing 1% FBS and allowed to migrate toward a serum gradient
(10%) in the lower chamber for 48 h. The non-invading cells were
then removed from the upper surface with a cotton swab. After
staining with 0.1% crystal violet solution for 10 min at room
temperature, the stained cells on the bottom surface were counted
on each membrane to test the invasion ability of cancer cells.
Three random fields were captured at a magnification of ×20
(n=3).
Reverse transcription-quantitative PCR
(RT-qPCR) assay
Total RNA was extracted from PSCs and pancreatic
cancer cells using the Fastgen200 RNA isolation system (Fastgen)
and reverse transcription was performed using the PrimeScript RT
Reagent Kit (Takara Biotechnology Co., Ltd.) according to the
manufacturer's recommendations.
The primer sequences were as follows: E-cadherin,
forward 5′-ATTCTGATTCTGCTGCTCTTG-3′ and reverse
5′-AGTCCTGGTCCTCTTCTCC-3′; vimentin, forward
5′-AATGACCGCTTCGCCAAC-3′ and reverse 5′-CCGCATCTCCTCCTCGTAG-3′;
MMP-9, forward 5′-GCAATGCTGATGGGAAACCC-3′ and reverse
5′-AGAAGCCGAAGAGCTTGTCC-3′; IL-6, forward
5′-AGTTCCTGCAGTCCAGCCTGAG-3′ and reverse
5′-TCAAACTGCATAGCCACTTTCC-3′; and β-actin, forward
5′-GACTTAGTTGCGTTACACCCTTTCT-3′ and reverse
5′-GAACGGTGAAGGTGACAGCAGT-3′.
The following PCR program was used: Denaturation at
95°C for 30 sec, followed by 40 cycles of 95°C for 5 sec, 60°C for
30 sec and 72°C for 30 sec. After each RT-qPCR experiment, a
dissociation curve analysis was conducted. The relative gene
expression was calculated using the previously described
2−ΔΔCq method (18).
ELISA
The PSCs from the indicated groups were conditioned
in serum-free medium for 72 h. The cellular culture media were then
collected and centrifuged at 1,000 × g for 5 min at 4°C to remove
particles. The production of IL-6 in the supernatants of PSCs was
detected by ELISA using the commercially available ELISA kit (cat.
no. D6050; R&D Systems, Inc.) according to the manufacturer's
instructions.
Immunofluorescence microscopy
After the designated treatment (hypoxia or
normoxia), PSCs were fixed with 4% paraformaldehyde for 10 min at
room temperature, permeabilized in 0.5% Triton X-100 for 10 min and
blocked in 1% BSA (Beyotime Institute of Biotechnology) for 1 h.
Fixed cells were incubated with mouse anti-human-α-SMA antibodies
(1:200, cat. no. 48938, Cell Signaling Technology, Inc.) at 4°C
overnight. Cells were washed and incubated with goat anti-mouse
DyLight 594 (red) IgG antibody (1:1,000, cat. no. ab96873, Abcam)
for 1 h in a dark room. Nuclei were then stained with DAPI for 5
min at room temperature. The cells were visualized by a
fluorescence microscope (Nikon Corporation) using appropriate
excitation and emission spectra at a magnification, ×400.
Western blotting
Proteins were electrophoretically resolved on a
denaturing SDS-polyacrylamide gel (10-12%) and electrotransferred
onto nitrocellulose membranes (EMD Millipore). The membranes were
initially blocked with 5% non-fat dry milk in Tris-buffered saline
for 2 h at room temperature and then probed with antibodies against
α-SMA, E-cadherin, vimentin, MMP-9, ERK, p-ERK, NF-κB, p-NF-κB or
β-actin (loading control). Following co-incubation with the primary
antibodies at 4°C overnight, the membranes were incubated with
HRP-conjugated secondary antibodies for 2 h at 37°C. The results
were visualized using the ECL western blotting substrate (Thermo
Fisher Scientific, Inc.) and photographed by GeneBox (CHEMI–XT16;
SynGene).
Statistical analysis
Statistical analysis was performed using SPSS
software (version 17.0, SPSS Inc.). Data are presented as the means
± standard error of the mean from at least three independent
experiments. Comparisons between two groups were performed using
Student's t-test. Differences among three or more groups were
evaluated by analysis of variance followed by the least significant
difference test. P<0.05 was considered to indicate statistically
significant differences. All experiments were repeated
independently at least three times.
Results
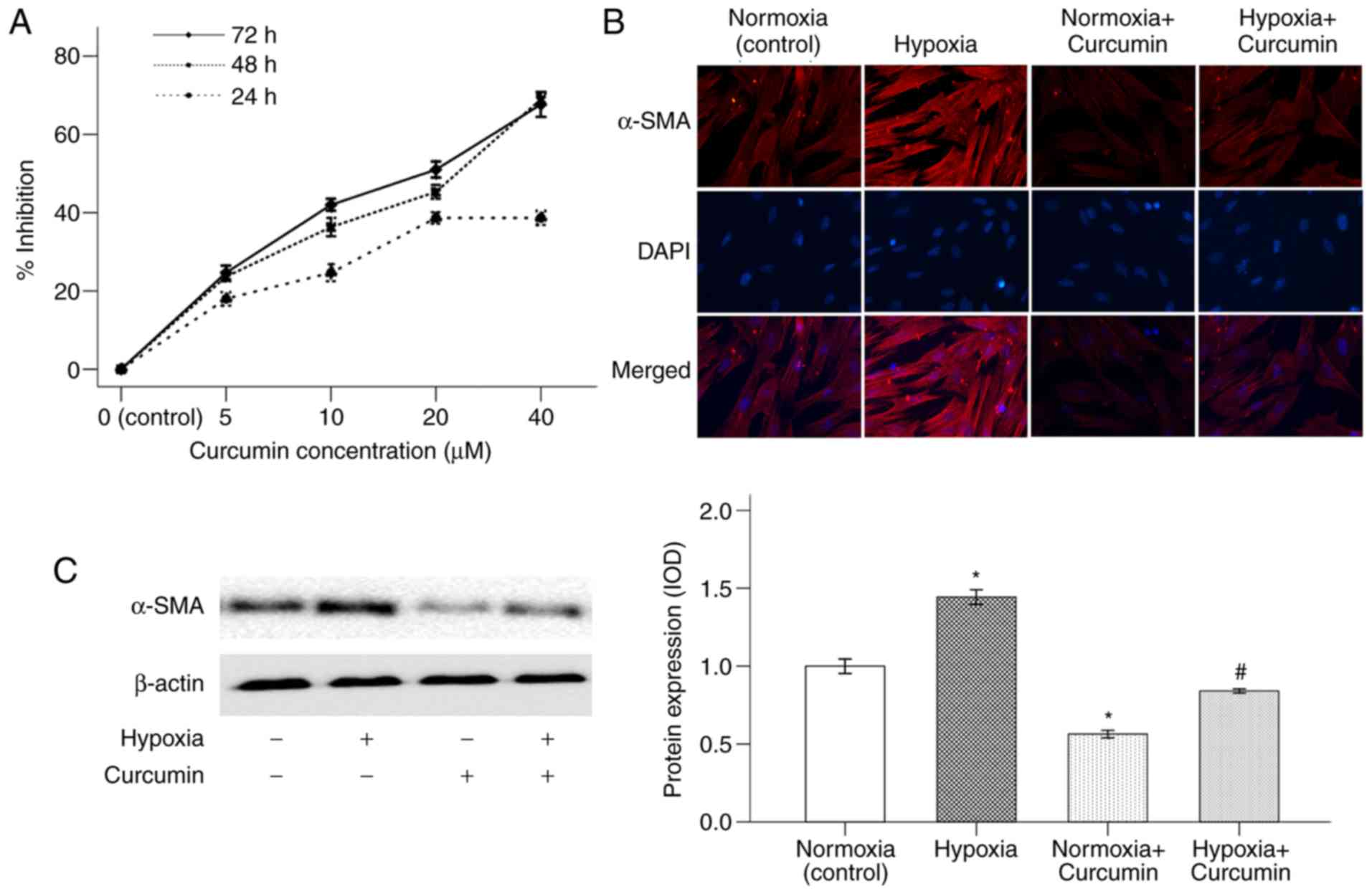
Curcumin inhibits hypoxia-induced
α-SMA expression in PSCs
A typical characteristic of activated PCSs is to
obtain a myofibroblast-like phenotype with increasing expression of
α-SMA (19). Our previous study
demonstrated that hypoxia is able to increase the activation of
PSCs (9). In the present study, the
cytotoxic effect of curcumin on PSCs was first investigated. As
shown in Fig. 1A, PSCs were treated
with curcumin at various concentrations (0–40 µM) for 24, 48 and 72
h (DMSO was used as the control group). The proliferative abilities
of PSCs decreased in response to curcumin treatment in both a time-
and dose-dependent manner. Curcumin had a 50% inhibitory
concentration (IC50) of ~20 µM, and this concentration
was used for the subsequent experiments. This result also proved
that hypoxia could enhance the expression of α-SMA in PSCs.
Treatment with 20 µM curcumin for 48 h reduced PSC activation, as
revealed by α-SMA expression (Fig. 1B
and C).
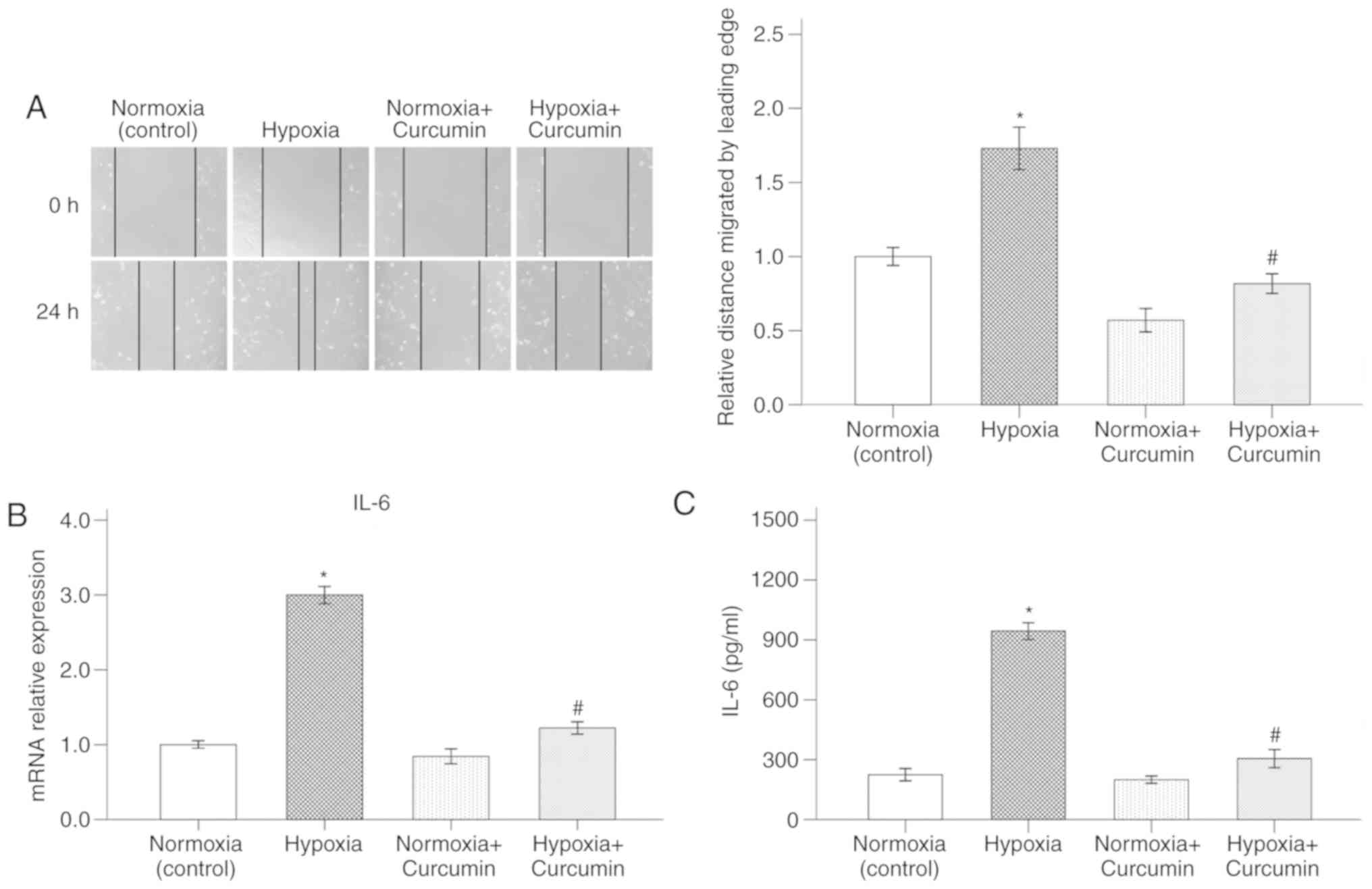
Curcumin abrogates hypoxia-activated
migratory ability and IL-6 secretion in PSCs
A hypoxic microenvironment plays a key role in
pancreatic cancer progression. It has been suggested that a hypoxic
microenvironment could affect cancer cells as well as the
surrounding PSCs (20). The present
study demonstrated that hypoxia induced migration of PSCs, as
assessed by the wound healing assay. Delayed wound closure was
observed following treatment of PSCs with curcumin (Fig. 2A).
IL-6 is a potent pro-inflammatory cytokine secreted
by PSCs in the pancreatic cancer microenvironment. Previous results
have indicated that the activated stroma is able to secrete large
amounts of IL-6, which could further promote the invasive capacity
of the surrounding tumor cells (21). To verify whether hypoxia-activated
PSCs have higher expression of IL-6, RT-qPCR and ELISA were
performed. As shown in Fig. 2B and
C, PSCs cultured under hypoxic conditions displayed higher
levels of IL-6. However, treatment with 20 µM curcumin for 72 h
counterbalanced the effect of hypoxia on the expression of IL-6 in
PSCs.
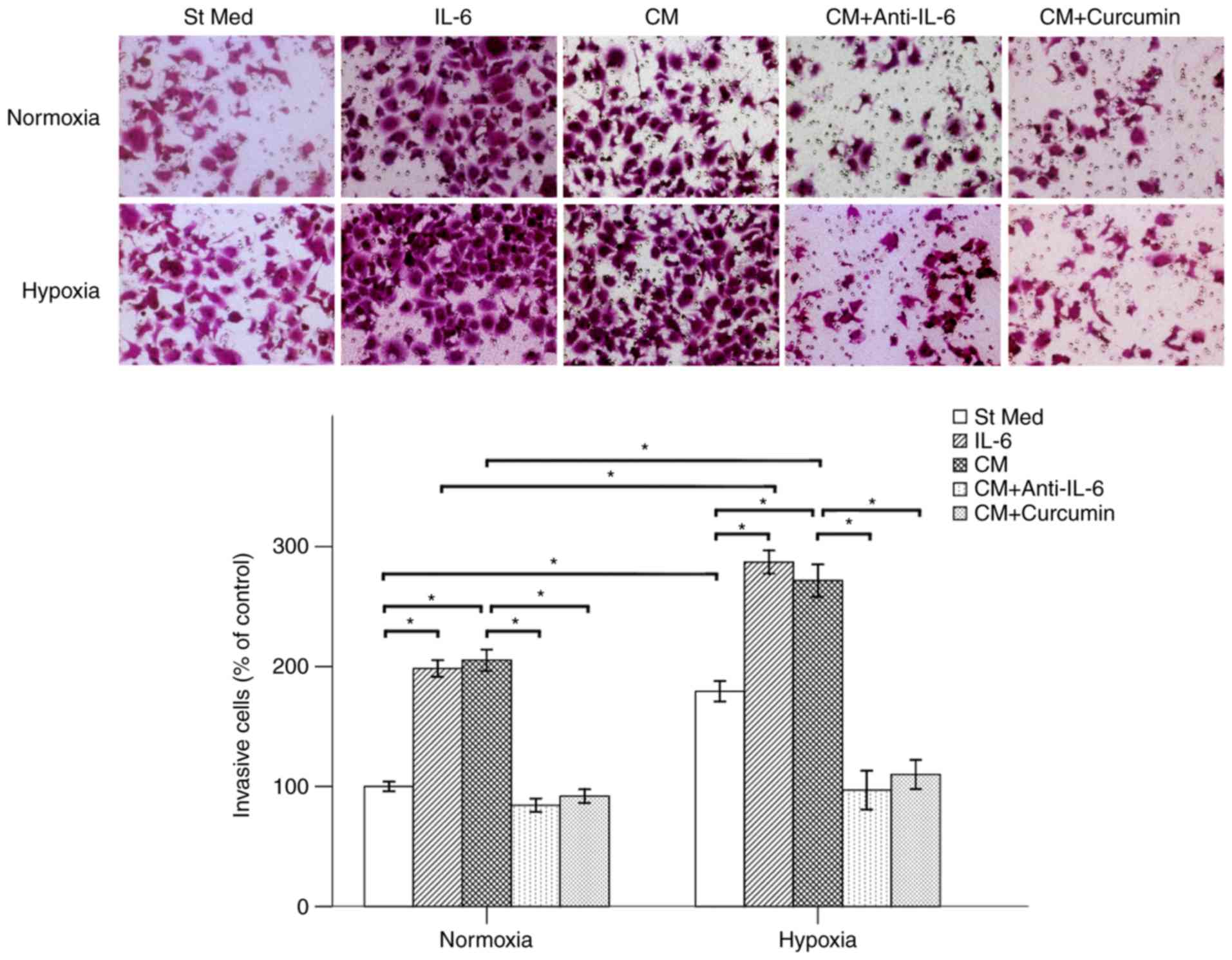
Curcumin suppresses PSC-CM-induced
invasive ability of pancreatic cancer cells
PSCs do not only form a dense fibrotic stroma and
interact with cancer cells, but may also be able to induce distant
metastasis (19). The present study
focused on whether media from the PSCs cultured under hypoxic
conditions could promote the metastatic potential of BxPC-3 cells.
The results demonstrated that both hypoxia and IL-6 increased the
invasion rates of cancer cells. PSC-CM also promoted the invasive
ability of pancreatic cancer cells. To elucidate the role of IL-6
in the PSC-CM-induced alterations in the function of pancreatic
cancer cells, a neutralizing antibody against IL-6 was employed. It
was demonstrated that treatment with anti-IL-6 antibody was able to
suppress the PSC-CM-induced invasion of BxPC-3 cells. Therefore,
PSC-CM-induced invasion may be attributed to the production of
IL-6. Our previous studies indicated that curcumin has an
IC50 of 20 µM in BxPC-3 cells (11). Thus, 20 µM of curcumin was used in
the present experiments. The mean number of cells invading into the
lower chamber, which was induced by PSC-CM, was also lower
following 48 h of co-treatment with curcumin under both hypoxic and
normoxic conditions (Fig. 3).
Therefore, curcumin may be considered as potential therapy against
IL-6-induced pathological conditions (22).
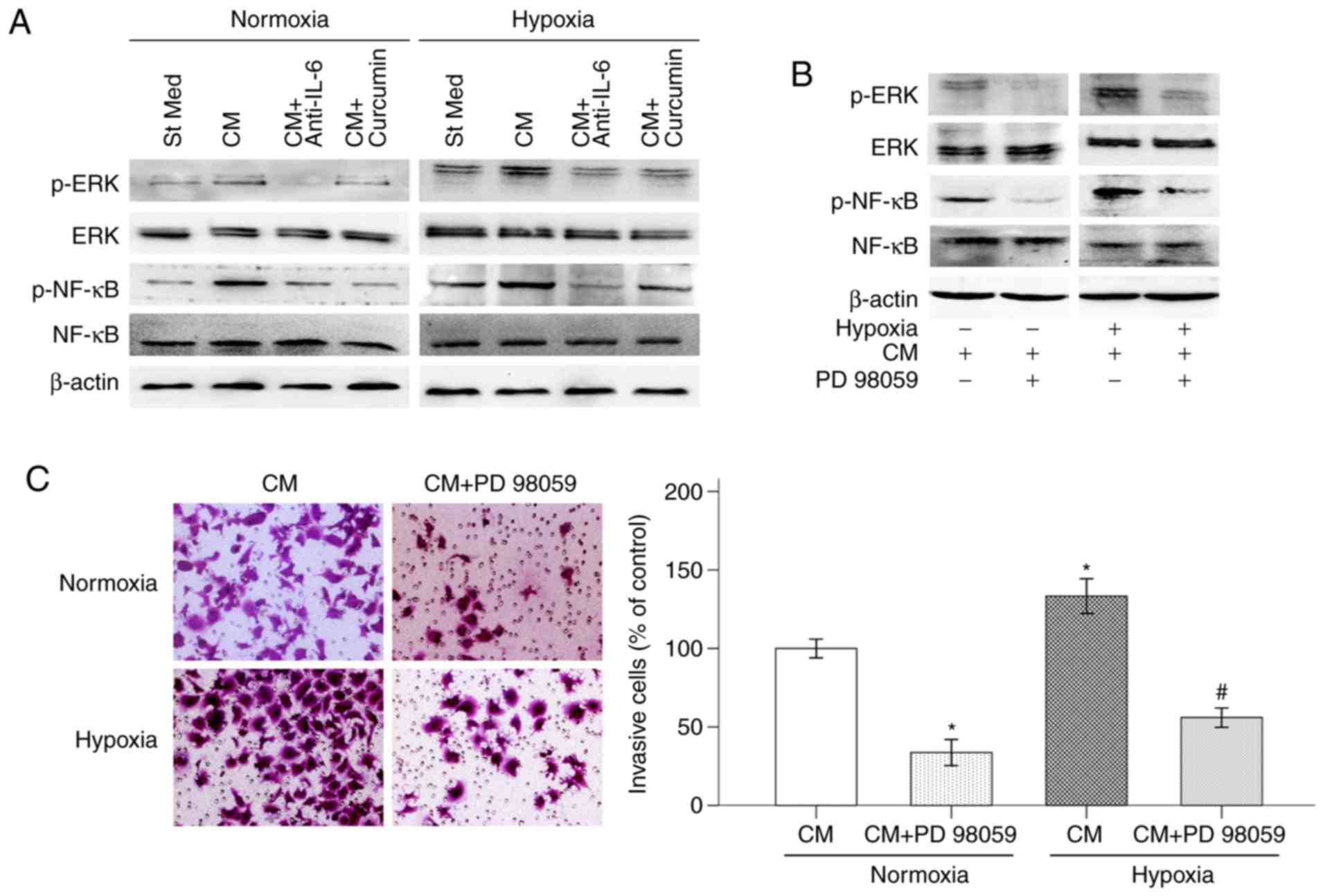
Curcumin restrains PSC-CM-induced
IL-6/ERK/NF-κB axis activation in pancreatic cancer cells under
hypoxic conditions
Previous studies have demonstrated that hypoxic
conditions (10) and PSC-CM
(23) activate multiple signaling
pathways in pancreatic cancer cells. The ERK/NF-κB pathway plays an
important role in numerous cellular processes, including tumor cell
survival, migration and invasion (24). Our previous study demonstrated that
curcumin plays an important role in inhibiting the proliferation,
migration and invasion of pancreatic cancer cells via the reactive
oxygen species/ERK/NF-κB signaling pathway (25). In the present study, BxPC-3 cells
were treated with PSC-CM activated by hypoxia in the presence of
curcumin or anti-IL-6 antibody, and the activation of the ERK/NF-κB
signaling pathway was tested via western blotting. As shown in
Fig. 4A, hypoxia significantly
increased the expression of p-ERK and p-NF-κB in BxPC-3 cells.
Treatment with PSC-CM greatly enhanced this effect. PSC-CM was also
able to activate the ERK/NF-κB signaling pathway in BxPC-3 cells,
even under normoxic conditions. The ability of the anti-IL-6
antibody to abolish the effects on hypoxia and PSC-CM-induced p-ERK
and p-NF-κB expression demonstrated that IL-6 played an important
role in ERK/NF-κB pathway activation. In addition, the
PSC-CM-induced phosphorylation of ERK and NF-κB also strongly
decreased following the addition of curcumin. Moreover, the ERK
inhibitor PD 98059 (50 µM) inhibited hypoxia-induced activation of
p-ERK and p-NF-κB, indicating that the NF-κB transcription factor
is modulated by the ERK pathway (Fig.
4B). PD 98059 also decreased pancreatic cancer cell invasion
under hypoxic conditions, indicating that the activation of the ERK
signaling pathway is involved in hypoxia-induced pancreatic cancer
cell invasion (Fig. 4C).
Curcumin inhibits PSC-CM-promoted EMT
in pancreatic cancer cells
EMT, a critical event during tumor invasion and
metastasis, includes four important steps: Loss of cellular
polarized epithelial traits, expression of mesenchymal proteins
(such as vimentin and N-cadherin), degradation of the basement
membrane, and enhancement of the cell invasive ability and entry
into the circulation (26). Our
previous study demonstrated that hypoxia is able to induce invasion
and EMT of pancreatic cancer cells (11). The focus of the present study was to
determine whether media from PSCs cultured under hypoxic conditions
could promote the EMT of pancreatic cancer cells. The results
demonstrated that cancer cell morphology changed from a typical
epithelial phenotype to a mesenchymal phenotype following PSC-CM
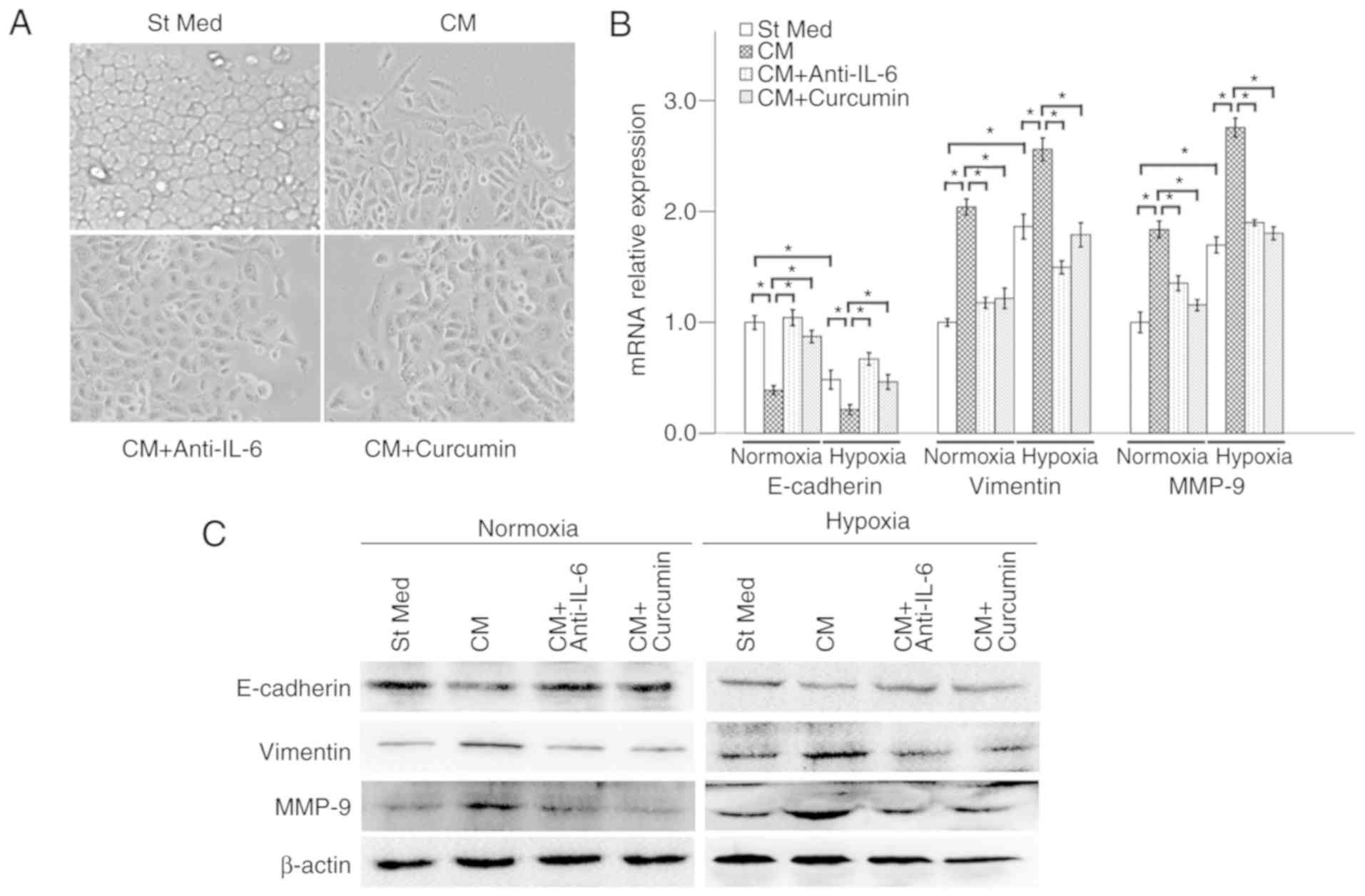
treatment for 48 h, which was counterbalanced by curcumin (Fig. 5A). In addition, PSC-CM significantly
decreased the E-cadherin level and increased the expression of
vimentin and MMP-9 in pancreatic cancer cells at both the mRNA and
protein levels, under either normoxic or hypoxic conditions, which
indicates that PSC-CM can promote EMT in pancreatic cancer cells.
These effects of hypoxia and PSC-CM were counterbalanced by either
20 µM curcumin or anti-IL-6 antibody (5 µg/ml) (Fig. 5B and C).
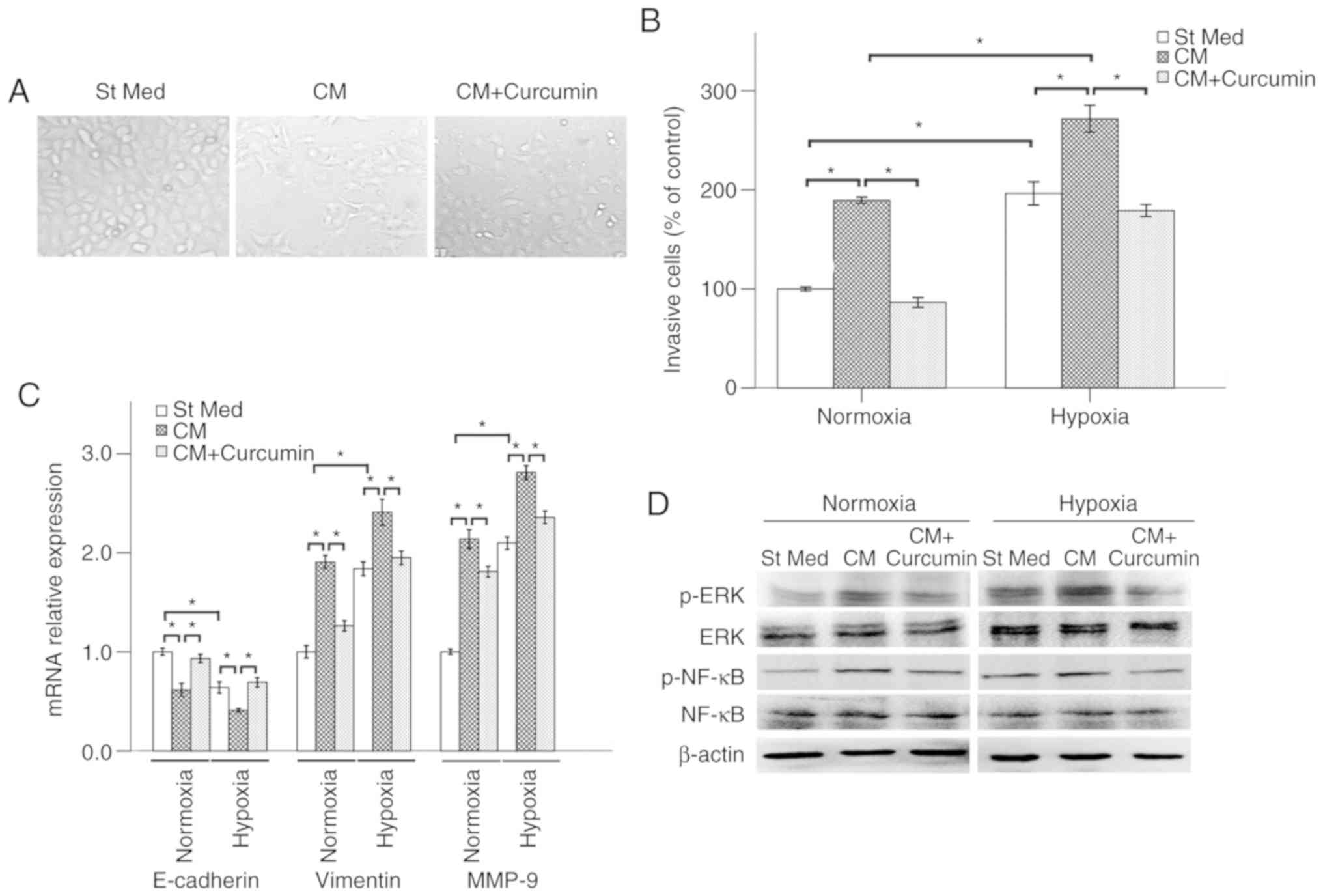
Moreover, the shape of Panc-1 cells changed into
myofibroblast-like phenotype following PSC-CM treatment for 48 h
(Fig. 6A). Hypoxic conditions and
hypoxia-induced PSC-CM promoted the invasive ability of Panc-1
cells (Fig. 6B). The expressions of
E-cadherin, vimentin, MMP-9 were also modulated by PSC-CM (Fig. 6C). As shown in Fig. 4D, hypoxia significantly increased
the expression of p-ERK and p-NF-κB in Panc-1 cells. Treatment with
PSC-CM was also able to activate the ERK/NF-κB signaling pathway
under both hypoxic and normoxic conditions, whereas these effects
of PSC-CM were suppressed by curcumin.
Taken together, these results demonstrated that
curcumin plays an important role in suppressing tumor-stromal
crosstalk and pancreatic cancer invasion and EMT by inhibiting the
IL-6/ERK/NF-κB axis.
Discussion
Pancreatic cancer is one of the most lethal types of
cancer, characterized by the lack of an effective therapeutic
strategy and poor life expectancy, mainly due to its early
metastasis and rapid progression. As pancreatic cancer is
clinically known as a hypovascular tumor, the pancreatic tumor
microenvironment is characterized by lack of oxygen and cells are
continuously exposed to hypoxia (27). A number of studies have indicated
that a hypoxic microenvironment is highly associated with poor
clinical outcome, which is attributed to the enhanced cancer cell
invasion and metastasis (28). Our
previous study also demonstrated that hypoxic conditions promote
the proliferation, migration and EMT in pancreatic cancer cells
(11). The pancreatic cancer
microenvironment contains several types of stromal cells, including
PSCs. Hypoxia activates PSCs, which further secrete a variety of
cytokines and growth factors, which in turn promote the progression
of pancreatic cancer, particularly cell migration and invasion
(9). The focus of the present study
was the underlying mechanisms through which curcumin inhibits
hypoxia-induced tumor-stromal crosstalk, as well as the invasion
and EMT of pancreatic cancer cells.
The data of the present study demonstrated that
curcumin inhibited the activation of PSCs under hypoxic conditions.
Hypoxia-induced migration and IL-6 secretion of PSCs were also
suppressed by curcumin. IL-6 and PSC-CM stimulated the invasive
ability of pancreatic cancer cells under hypoxic conditions, while
IL-6-neutralizing antibody treatment suppressed the
PSC-CM-modulated pancreatic cancer cell invasion, indicating that
hypoxia-induced IL-6 secretion of PSCs may promote the progression
of pancreatic cancer. Curcumin counterbalanced these effects of
hypoxia and PSC-CM treatment. PSC-CM-modulated cancer cell
morphology, and the changes in the expression of E-cadherin,
vimentin and MMP-9, were inhibited by curcumin and
IL-6-neutralizing antibody. In addition, hypoxia and PSC-CM-induced
activation of the ERK pathway and the transcription factor NF-κB
were also suppressed by IL-6-neutralizing antibody and curcumin.
These results indicated that curcumin inhibits hypoxia-induced
tumor-stromal crosstalk and pancreatic cancer EMT and metastasis
via the IL-6/ERK/NF-κB axis.
The pancreatic cancer microenvironment includes not
only tumor cells, but also other cell types, including PSCs, immune
cells and inflammatory cells, ECM, blood vessels, and other
signaling molecules (5). Our
previous studies have focused on the effect of hypoxia on the tumor
cells (11). The present study,
however, investigated the effect of curcumin on hypoxia-induced
tumor-stromal interaction. The normal function of PSCs includes
immune responses, phagocytosis, and stimulating the secretion of
amylase in the healthy pancreas (29). However, PSCs also act as the
sprouted seed for tumor progression, due to their important role in
inducing a fibrotic environment that facilitates tumor metastasis
(30). Several cytokines and growth
factors derived from PSCs are also involved in malignant tumor
progression. It has been proven that PSCs promote pancreatic cancer
cell invasion and migration through the hepatocyte growth factor
(HGF)/c-Met/survivin pathway, which is negatively regulated by
P53/P21 (31). PSCs may also
enhance pancreatic cancer cell invasion of the dorsal root ganglia
(DRG), promote the outgrowth of DRG and further facilitate tumor
cell perineural invasion via the HGF/c-Met pathway (32). In addition, paracrine IL-6 from PSCs
promotes the EMT of pancreatic cancer cells via the signal
transducer and activator of transcription 3 (STAT3)/nuclear factor
erythroid 2-related factor 2 (Nrf2) pathway (33). In the present study, it was observed
that hypoxia could activate PSCs and promote the paracrine
secretion of IL-6, thereby further promoting the invasion and EMT
of pancreatic cancer cells via the ERK/NF-κB signaling pathway.
As a hypovascular tumor, pancreatic cancer thrives
under hypoxic conditions. Hypoxia-inducible factor (HIF)-1, a key
mediator of the cellular response to hypoxia, is overexpressed in a
variety of solid tumors, including pancreatic cancer (34). It has been proven that HIF-1α can
recruit macrophages by promoting secretion of C-C motif chemokine
ligand 2, which further accelerates the activation of PSCs
(35). Hirakawa et al
(27) identified a
motility-stimulating effect of pancreatic cancer-associated
fibroblasts on pancreatic cancer cells through the paracrine
insulin-like growth factor-1 (IGF1)/IGF1R signaling axis,
particularly under hypoxic conditions. Hypoxia is also able to
induce PSCs to secrete connective tissue growth factor (CTGF),
which in turn promotes the invasion of pancreatic cancer cells
(36). Masamune et al
(20) demonstrated that PSCs
expressed a variety of angiogenesis-regulating molecules, including
VEGF receptors, angiopoietin-1, and the angiopoietin receptor
Tie-2. Hypoxia stimulates PSCs to induce fibrosis and angiogenesis
in pancreatic cancer. PD 98059, a MEK inhibitor, has been shown to
suppress the ERK pathway and invasion of pancreatic cancer cells in
our previous study (37). PD 98059
was used to inhibit the ERK phosphorylation, in order to verify
that this process occurs upstream of ERK signaling. The result
demonstrated that PD 98059 not only inhibited hypoxia-induced
expression of p-ERK and p-NF-κB, but also decreased pancreatic
cancer cell invasion under hypoxic conditions. These results
indicated that hypoxia can promote the invasion and EMT of
pancreatic cancer cells by interfering with tumor-stromal crosstalk
via the IL-6/ERK/NF-κB axis.
As a pro-inflammatory cytokine, IL-6 is involved in
multiple biological processes, including tumor progression. In the
cancer microenvironment, IL-6 is produced by a number of cell
types, including fibroblast stromal cells, immune cells, and even
tumor cells (38). It has been
reported that IL-6 is closely associated with pancreatic cancer
development, progression, invasion, EMT and metastasis by
activating Janus kinase 2 (JAK2)/STAT3, mitogen-activated protein
kinase (MAPK) and other signaling pathways that contribute to
oncogenesis (39). High circulating
levels of IL-6 have been associated with short overall survival in
gastrointestinal cancer patients (38). In a study using an orthotopic
xenograft model with pancreatic cancer cells in SCID/bg mice,
Goumas et al (40)
demonstrated that treatment with the anti-IL-6-receptor antibody
tocilizumab resulted in a marked decrease in tumor weight and new
distant metastases compared with the control group. The results of
the present study also indicated that hypoxia can increase the
production of IL-6 by PSCs. IL-6 treatment or hypoxia-induced
PSC-CM promoted the invasion of pancreatic cancer cells. PSC-CM
also promoted the EMT of BxPC-3 and Panc-1 cells, which was
counterbalanced by the anti-IL-6 antibody or curcumin.
Curcumin (diferuloylmethane) has been proven to
suppress the proliferation, invasion, EMT and metastasis of a
variety of tumors, including pancreatic cancer (11). Curcumin exerts its anticancer
effects via targeting multiple signaling pathways, such as the MAPK
and NF-κB signaling pathways. Our recent study demonstrated that
curcumin attenuated hyperglycemia-driven epidermal growth factor
(EGF)-induced invasive and migratory abilities of pancreatic cancer
cells by inhibiting the EGF/EGFR signaling pathway and its
downstream signaling molecules, including ERK and Akt (15). Curcumin may also play an important
role in suppressing superoxide dismutase-driven hydrogen peroxide
-induced pancreatic cancer EMT and metastasis by inhibiting the
phosphoinositide 3 kinase/Akt/NF-κB pathway (41). In recent years, an increasing number
of studies have been focusing on the role of curcumin in the tumor
microenvironment. Shao et al (42) recently revealed that curcumin exerts
a promising therapeutic effect on hepatic stellate cell-induced
hepatocellular carcinoma invasion and angiogenesis via
downregulating CTGF. Wang et al (43) also provided evidence that curcumin
was able to suppress the migration, EMT and metastasis of
pancreatic cancer cells by reducing the mesenchymal characteristics
of cancer-associated fibroblasts. The data presented herein
indicate that curcumin inhibited hypoxia-activated migratory
ability and IL-6 secretion in PSCs. Curcumin may also abrogate
hypoxia-induced PSC-CM-mediated invasive ability and EMT of
pancreatic cancer cells. The focus of our future study will be to
investigate the components of the pathway directly activated by
IL-6, such as JAK2, STAT3 or Nrf2, which was a limitation of the
present study. Interestingly, in the present study it was also
observed that curcumin was able to inhibit α-SMA expression and
cell migration under normoxic conditions. The effect of curcumin on
these processes appeared to be independent of IL-6, since this
molecule was not induced under normoxia in pancreatic cancer cells.
As curcumin can inhibit several important factors secreted by
stromal cells, including SDF-1 and CTGF (42), future work will also focus on these
factors. Curcumin has also been applied in the clinical setting,
either alone or in combination with other drugs. In a phase I/II
study, gemcitabine-resistant patients with pancreatic cancer
received oral curcumin daily (8 g) in combination with
gemcitabine-based chemotherapy. The result demonstrated that
curcumin was safe and able to increase the median survival time
(161 days) when compared with the gemcitabine alone group, the mean
survival rate of which was ~10 weeks (44). Our future research may also include
more in vivo experiments as well as preclinical trials to
further validate the present findings.
In conclusion, the present study demonstrated that
curcumin plays an important role in suppressing tumor-stromal
crosstalk and pancreatic cancer cell invasion and EMT by inhibiting
the IL-6/ERK/NF-κB axis under hypoxic conditions. Therefore,
curcumin may hold promise as a potential anticancer agent for the
treatment of patients with pancreatic cancer.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
Fundamental Research Funds for the Central Universities (grant no.
xjj2018117) and the Natural Science Basic Research Project of
Shaanxi Province (grant no. 2020JM-367).
Availability of data and materials
The datasets generated and/or analyzed during the
present study are available from the corresponding author upon
reasonable request.
Authors' contributions
WL, QM and ZWa conceived and designed the study; WL,
LS and JL conducted the experiments; ZWu and JL performed the data
analysis; WL wrote the paper; ZWa and QM reviewed and edited the
manuscript. All the authors have read and approved the final
version of the manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The experimental protocol and patient consent forms
were approved by the Ethics Committee of the First Affiliated
Hospital of Xi'an Jiaotong University (Xi'an, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen WQ, Zheng RS, Baade PD, Zhang SW,
Zeng HM, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in
China, 2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vincent A, Herman J, Schulick R, Hruban RH
and Goggins M: Pancreatic cancer. Lancet. 378:607–620. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li H, Wang X, Wen C, Huo Z, Wang W, Zhan
Q, Cheng D, Chen H, Deng X, Peng C and Shen B: Long noncoding RNA
NORAD, a novel competing endogenous RNA, enhances the
hypoxia-induced epithelial-mesenchymal transition to promote
metastasis in pancreatic cancer. Mol Cancer. 16:1692017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
von Ahrens D, Bhagat TD, Nagrath D, Maitra
A and Verma A: The role of stromal cancer-associated fibroblasts in
pancreatic cancer. J Hematol Oncol. 10:762017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tang D, Wang D, Yuan Z, Xue X, Zhang Y, An
Y, Chen J, Tu M, Lu Z, Wei J, et al: Persistent activation of
pancreatic stellate cells creates a microenvironment favorable for
the malignant behavior of pancreatic ductal adenocarcinoma. Int J
Cancer. 132:993–1003. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu Q, Tian Y, Zhang J, Zhang H, Gu F, Lu
Y, Zou S, Chen Y, Sun P, Xu M, et al: Functions of pancreatic
stellate cell-derived soluble factors in the microenvironment of
pancreatic ductal carcinoma. Oncotarget. 8:102721–102738. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Razidlo GL, Burton KM and McNiven MA:
Interleukin-6 promotes pancreatic cancer cell migration by rapidly
activating the small GTPase CDC42. J Biol Chem. 293:11143–11153.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lei J, Huo X, Duan W, Xu Q, Li R, Ma J, Li
X, Han L, Li W, Sun H, et al: α-Mangostin inhibits hypoxia-driven
ROS-induced PSC activation and pancreatic cancer cell invasion.
Cancer Lett. 347:129–138. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Erkan M, Kurtoglu M and Kleeff J: The role
of hypoxia in pancreatic cancer: A potential therapeutic target?
Expert Rev Gastroenterol Hepatol. 10:301–316. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cao L, Xiao X, Lei J, Duan W, Ma Q and Li
W: Curcumin inhibits hypoxia-induced epithelial-mesenchymal
transition in pancreatic cancer cells via suppression of the
hedgehog signaling pathway. Oncol Rep. 35:3728–3734. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sada M, Ohuchida K, Horioka K, Okumura T,
Moriyama T, Miyasaka Y, Ohtsuka T, Mizumoto K, Oda Y and Nakamura
M: Hypoxic stellate cells of pancreatic cancer stroma regulate
extracellular matrix fiber organization and cancer cell motility.
Cancer Lett. 372:210–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shanmugam MK, Rane G, Kanchi MM, Arfuso F,
Chinnathambi A, Zayed ME, Alharbi SA, Tan BK, Kumar AP and Sethi G:
The multifaceted role of curcumin in cancer prevention and
treatment. Molecules. 20:2728–2769. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Allegra A, Innao V, Russo S, Gerace D,
Alonci A and Musolino C: Anticancer activity of curcumin and its
analogues: Preclinical and clinical studies. Cancer Invest.
35:1–22. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li W, Wang Z, Xiao X, Han L, Wu Z, Ma Q
and Cao L: Curcumin attenuates hyperglycemia-driven EGF-induced
invasive and migratory abilities of pancreatic cancer via
suppression of the ERK and AKT pathways. Oncol Rep. 41:650–658.
2019.PubMed/NCBI
|
|
16
|
Bachem MG, Schneider E, Gross H,
Weidenbach H, Schmid RM, Menke A, Siech M, Beger H, Grunert A and
Adler G: Identification, culture, and characterization of
pancreatic stellate cells in rats and humans. Gastroenterology.
115:421–432. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun L, Chen K, Jiang Z, Chen X, Ma J, Ma Q
and Duan W: Indometacin inhibits the proliferation and activation
of human pancreatic stellate cells through the downregulation of
COX-2. Oncol Rep. 39:2243–2251. 2018.PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ferdek PE and Jakubowska MA: Biology of
pancreatic stellate cells-more than just pancreatic cancer.
Pflugers Arch. 469:1039–1050. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Masamune A, Kikuta K, Watanabe T, Satoh K,
Hirota M and Shimosegawa T: Hypoxia stimulates pancreatic stellate
cells to induce fibrosis and angiogenesis in pancreatic cancer. Am
J Physiol Gastrointest Liver Physiol. 295:G709–G717. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nagathihalli NS, Castellanos JA, VanSaun
MN, Dai X, Ambrose M, Guo Q, Xiong Y and Merchant NB: Pancreatic
stellate cell secreted IL-6 stimulates STAT3 dependent invasiveness
of pancreatic intraepithelial neoplasia and cancer cells.
Oncotarget. 7:65982–65992. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ghandadi M and Sahebkar A: Curcumin: An
effective inhibitor of interleukin-6. Curr Pharm Des. 23:921–931.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hamada S, Masamune A, Yoshida N, Takikawa
T and Shimosegawa T: IL-6/STAT3 plays a regulatory role in the
interaction between pancreatic stellate cells and cancer cells. Dig
Dis Sci. 61:1561–1571. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Neuzillet C, Hammel P, Tijeras-Raballand
A, Couvelard A and Raymond E: Targeting the Ras-ERK pathway in
pancreatic adenocarcinoma. Cancer Metastasis Rev. 32:147–162. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cao L, Liu J, Zhang L, Xiao X and Li W:
Curcumin inhibits H2O2-induced invasion and migration of human
pancreatic cancer via suppression of the ERK/NF-κB pathway. Oncol
Rep. 36:2245–2251. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li W, Ma Q, Liu J, Han L, Ma G, Liu H,
Shan T, Xie K and Wu E: Hyperglycemia as a mechanism of pancreatic
cancer metastasis. Front Biosci (Landmark Ed). 17:1761–1774. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hirakawa T, Yashiro M, Doi Y, Kinoshita H,
Morisaki T, Fukuoka T, Hasegawa T, Kimura K, Amano R and Hirakawa
K: Pancreatic fibroblasts stimulate the motility of pancreatic
cancer cells through IGF1/IGF1R signaling under hypoxia. PLoS One.
11:e01599122016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu H, Ma Q, Xu Q, Lei J, Li X, Wang Z and
Wu E: Therapeutic potential of perineural invasion, hypoxia and
desmoplasia in pancreatic cancer. Curr Pharm Des. 18:2395–2403.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mato E, Lucas M, Petriz J, Gomis R and
Novials A: Identification of a pancreatic stellate cell population
with properties of progenitor cells: New role for stellate cells in
the pancreas. Biochem J. 421:181–191. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Thomas D and Radhakrishnan P:
Tumor-stromal crosstalk in pancreatic cancer and tissue fibrosis.
Mol Cancer. 18:142019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang XP, Liu SL, Xu JF, Cao SG, Li Y and
Zhou YB: Pancreatic stellate cells increase pancreatic cancer cells
invasion through the hepatocyte growth factor /c-Met/survivin
regulated by P53/P21. Exp Cell Res. 357:79–87. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Nan L, Qin T, Xiao Y, Qian W, Li J, Wang
Z, Ma J, Ma Q and Wu Z: Pancreatic stellate cells facilitate
perineural invasion of pancreatic cancer via HGF/c-Met pathway.
Cell Transplant. 28:1289–1298. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu YS, Chung I, Wong WF, Masamune A, Sim
MS and Looi CY: Paracrine IL-6 signaling mediates the effects of
pancreatic stellate cells on epithelial-mesenchymal transition via
Stat3/Nrf2 pathway in pancreatic cancer cells. Biochim Biophys Acta
Gen Subj. 1861:296–306. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lei J, Ma J, Ma Q, Li X, Liu H, Xu Q, Duan
W, Sun Q, Xu J, Wu Z and Wu E: Hedgehog signaling regulates hypoxia
induced epithelial to mesenchymal transition and invasion in
pancreatic cancer cells via a ligand-independent manner Mol Cancer
12. 66:2013.
|
|
35
|
Li N, Li Y, Li Z, Huang C, Yang Y, Lang M,
Cao J, Jiang W, Xu Y, Dong J and Ren H: Hypoxia inducible factor 1
(HIF-1) recruits macrophage to activate pancreatic stellate cells
in pancreatic ductal adenocarcinoma. Int J Mol Sci. 17(pii):
E7992016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Eguchi D, Ikenaga N, Ohuchida K, Kozono S,
Cui L, Fujiwara K, Fujino M, Ohtsuka T, Mizumoto K and Tanaka M:
Hypoxia enhances the interaction between pancreatic stellate cells
and cancer cells via increased secretion of connective tissue
growth factor. J Surg Res. 181:225–233. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li W, Cao L, Han L, Xu Q and Ma Q:
Superoxide dismutase promotes the epithelial-mesenchymal transition
of pancreatic cancer cells via activation of the H2O2/ERK/NF-κB
axis. Int J Oncol. 46:2613–2620. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vainer N, Dehlendorff C and Johansen JS:
Systematic literature review of IL-6 as a biomarker or treatment
target in patients with gastric, bile duct, pancreatic and
colorectal cancer. Oncotarget. 9:29820–29841. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pop VV, Seicean A, Lupan I, Samasca G and
Burz CC: IL-6 roles-molecular pathway and clinical implication in
pancreatic cancer-A systemic review. Immunol Lett. 181:45–50. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Goumas FA, Holmer R, Egberts JH,
Gontarewicz A, Heneweer C, Geisen U, Hauser C, Mende MM, Legler K,
Röcken C, et al: Inhibition of IL-6 signaling significantly reduces
primary tumor growth and recurrencies in orthotopic xenograft
models of pancreatic cancer. Int J Cancer. 137:1035–1046. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li W, Jiang Z, Xiao X, Wang Z, Wu Z, Ma Q
and Cao L: Curcumin inhibits superoxide dismutase-induced
epithelial-to-mesenchymal transition via the PI3K/Akt/NF-κB pathway
in pancreatic cancer cells. Int J Oncol. 52:1593–1602.
2018.PubMed/NCBI
|
|
42
|
Shao S, Duan W, Xu Q, Li X, Han L, Li W,
Zhang D, Wang Z and Lei J: Curcumin suppresses hepatic stellate
cell-induced hepatocarcinoma angiogenesis and invasion through
downregulating CTGF. Oxid Med Cell Longev. 2019:81485102019.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang Q, Qu C, Xie F, Chen L, Liu L, Liang
X, Wu X, Wang P and Meng Z: Curcumin suppresses
epithelial-to-mesenchymal transition and metastasis of pancreatic
cancer cells by inhibiting cancer-associated fibroblasts. Am J
Cancer Res. 7:125–133. 2017.PubMed/NCBI
|
|
44
|
Kanai M, Yoshimura K, Asada M, Imaizumi A,
Suzuki C, Matsumoto S, Nishimura T, Mori Y, Masui T, Kawaguchi Y,
et al: A phase I/II study of gemcitabine-based chemotherapy plus
curcumin for patients with gemcitabine-resistant pancreatic cancer.
Cancer Chemother Pharmacol. 68:157–164. 2011. View Article : Google Scholar : PubMed/NCBI
|