Introduction
Hepatocellular carcinoma (HCC) accounts for 75–85%
of all primary liver cancer (PLC) cases and is the fourth leading
cause of cancer-related deaths worldwide (1). In Asia, liver cirrhosis caused by the
hepatitis virus infection contributes to the total number of
HCC-associated mortalities. Despite increased awareness and
improved clinical diagnosis of HCC, the patient prognosis remains
poor (1). A recent American study
based on the Surveillance, Epidemiology, and End Results (SEER) 18
Registry Database has shown that the incidence of HCC will continue
to rise until 2030 (2). Hence,
there is a need for developing effective therapeutic strategies for
improved HCC treatment.
Aspirin, one of the classical nonsteroidal
anti-inflammatory drugs (NSAIDs), is widely used as an
anti-inflammatory and anti-coagulation agent, and for reducing the
risk of cardiovascular disorders. Recently, many in vitro
and in vivo studies, epidemiological investigations, and
randomized clinical trials have generated evidence of the antitumor
effects of aspirin in various cancers such as colon (3), breast (4), pancreas (5), and lung (6) cancers. A meta-analysis showed that
aspirin is linked to a lower risk of HCC development and a
prolonged survival rate of HCC patients (7). According to the latest clinical
statistics, regular [≥2 standard-dose (325 mg) tablets per week]
and long-term use of aspirin are associated with a dose-dependent
reduction in HCC risk (8). The
functional effects of aspirin partly rely on the inhibition of the
cyclooxygenase (COX) enzyme; unlike other NSAIDs, the effect of
aspirin by this mechanism is irreversible. Furthermore, aspirin is
reported to activate key molecular targets in AMPK, mTOR, STAT3 and
NF-κB pathways in various carcinomas (4). It is also suggested to suppress cell
proliferation by inducing cell cycle arrest and apoptosis (9).
Regarding HCC cells, aspirin may decrease the levels
of reactive oxygen species (ROS) and glucose consumption by
downregulating the glucose transporter (10); inducing autophagy via
JNK/p-Bcl2/beclin-1, AMPK/mTOR, and GSK-3 signaling pathways
(11); inducing apoptosis and
mitochondrial dysfunction by increasing oxidative stress (12); and altering the tumor
microenvironment due to an effect on platelets (13,14).
Therefore, the antitumor effects of aspirin require in-depth
investigation in order to completely elucidate its underlying
molecular mechanisms.
The aim of the present study was to determine the
antitumor effects of aspirin on HCC-derived cell lines and a liver
cancer cell line and on an in vivo xenograft tumor model,
and to identify the key molecular targets and microRNAs (miRNAs)
associated with the functional effects exerted by aspirin.
Materials and methods
Chemicals
Aspirin was purchased from Wako Pure Chemical
Industries, Ltd. (Osaka, Japan). The prepared solution was diluted
with the cell culture medium as per cell requirement and used fresh
(pH 7.2 to 7.5, within the range suitable for cell growth).
Cell lines and culture
The HCC cell lines (HLE, HLF, Huh-7, PLC/PRF/5,
Hep-3B, Li-7) and a liver cancer cell line (Hep-G2) were obtained
from the Japanese Research Resources Bank (Tokyo, Japan). HCC Huh-7
cells were maintained in low glucose Dulbecco's modified Eagle's
media (DMEM) (Gibco-Invitrogen; Thermo Fisher Scientific, Inc.)
supplemented with 10% fetal bovine serum (FBS) (533-69545; FUJIFILM
Wako) and penicillin/streptomycin (100 mg/l; Invitrogen; Thermo
Fisher Scientific, Inc.) Liver cancer Hep-G2 cells and HCC Hep-3B
cells were cultured in Modified Eagle's Media (MEM)
(Gibco-Invitrogen; Thermo Fisher Scientific, Inc.) supplemented
with 10% FBS and penicillin/streptomycin. HCC HLE and PLC/PRF/5
cells were maintained in DMEM supplemented with 10% FBS and
penicillin/streptomycin. HCC HLF cells were maintained in DMEM
supplemented with 5% FBS and penicillin/streptomycin. HCC Li-7
cells were grown in RPMI-1640 (FUJIFILM Wako) supplemented with 10%
FBS and penicillin/streptomycin. Hepatocytes were grown in
endothelial cell medium (ECM) (Upcyte Technologies) with 5% FBS,
penicillin/streptomycin, 1% supplement A, and 1% L-glutamine. All
cell lines were grown in a humidified incubator at 5%
CO2 and 37°C.
Cell proliferation assay
The cell proliferation assay was performed using the
Cell Counting Kit-8 (Dojindo Laboratories) according to the
manufacturer's instructions. HLE, HLF, Huh-7, PLC/PRF/5, Hep-3B,
Li-7 and Hep-G2 cells (5,000 cells/100 µl/well) were seeded in
96-well plates and allowed to adhere, followed by treatment with
different concentrations of aspirin (0, 2.5, 5, or 10 mmol/l) for
48 h at 37°C. Subsequently, cells received 100 µl of fresh medium
containing the CCK-8 reagent and were incubated for an additional 3
h at 37°C. The absorbance was measured at 450 nm using an automated
microplate reader. The experiments were repeated thrice.
Flow cytometric analysis of the cell
cycle
To analyze the underlying mechanism of the
aspirin-mediated inhibition of tumor cell growth, flow cytometric
analyses were performed using the Cycle Phase Determination kit
(Cayman Chemical Co.). HCC Huh-7 cells (1.0×106
cells/100-mm diameter dish) were treated with 2.5 mmol/l aspirin
for 24 to 48 h. Cells were scraped and centrifuged to obtain the
cell pellet, which was resuspended in phosphate-buffered saline
(PBS) (106 cells/ml). An equal volume of cell suspension
was added to the cycle phase determination fixative and stored at
−20°C until further analysis. For cell cycle analysis, the cells
were suspended in 100 µl of PBS with 10 µl RNase A (250 µg/ml) and
10 µl propidium iodide (PI) stain (100 µg/ml), followed by
incubation at room temperature in the dark for 30 min. Flow
cytometry was performed to compare the proportion of
aspirin-treated and control cells in each phase of the cell cycle.
Flow cytometry was performed using a Cytomics FC 500 flow cytometer
(Beckman Coulter) with an argon laser (488 nm), and the percentages
of cells were analyzed using Kaluza software version v2.1 (Beckman
Coulter). The experiments were repeated thrice.
Apoptosis analysis
Aspirin-mediated apoptosis was analyzed using flow
cytometry and the Annexin V-FITC Early Apoptosis Detection kit
(Cell Signaling Technology, Inc.). Briefly, HCC Huh-7 cells
(1.0×106 cells/100-mm dish) were treated with 2.5 mmol/l
aspirin for 48 h at 37°C. Cells undergoing apoptosis and necrosis
were analyzed by double staining with FITC-conjugated Annexin V and
PI as per the manufacturer's protocol. This staining method is
based on the binding of Annexin V to apoptotic cells with exposed
phosphatidylserines, and the PI-labeling of the damaged membrane in
late apoptotic/necrotic cells. Flow cytometry was conducted using a
Cytomics FC 500 flow cytometer (Beckman Coulter) with an argon
laser (488 nm) and data were analyzed using Kaluza software version
v2.1 (Beckman Coulter). The experiments were repeated thrice to
compare the proportion of apoptotic cells in the aspirin-treated
and control groups.
Apoptosis analysis by ELISA
Caspase-cleaved cytokeratin 18 (cCK-18) levels were
measured using the M30 Apoptosense ELISA kit (Peviva Ab). Briefly,
HCC Huh-7 cells (5,000 cells/well) were seeded in 96-well plates
and treated with 2.5 mmol/l aspirin for 48 h at 37°C. Subsequently,
the cells were lysed in polyoxyethylene octyl phenyl ether (NP-40)
(Wako) and further analyzed according to the manufacturer's
instructions.
Western blot analysis
HCC Huh-7 cells were seeded (1.0×106
cells/100-mm dish) and treated with 2.5 mmol/l aspirin for 24 or 48
h. The untreated cells were used as control. The cells were lysed
with PRO-PREP complete protease inhibitor mixture (iNtRON
Biotechnology). Supernatants were collected and stored at −80°C
until further analysis. Protein concentrations were measured using
a Nanodrop 2000 spectrofluorometer (Thermo Fisher Scientific,
Inc.), and aliquots (1–10 µg) were resolved by a 12% Tris-glycine
gradient gel and sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE). The resolved proteins were transferred
to a nitrocellulose membrane and blocked with a blocking buffer.
Subsequently, the membranes were incubated with primary antibodies
followed by horseradish peroxidase (HRP)-conjugated secondary
antibodies. The following antibodies were used for developing the
blot: primary antibodies against cyclin D1 (SP4) (#MA5-14512)
(dilution 1:500), retinoblastoma protein (Rb) (#MA1-34070)
(dilution 1:1,000), epidermal growth factor receptor (EGFR)
(#PA1-1110) (dilution 1:1,000), and cyclin E (HE-12) (MS-870-P1)
(dilution 1:1,000) were obtained from Thermo Fisher Scientific,
Inc.; primary antibody against phosphorylated Rb (pS780) (dilution
1:1,000) was obtained from BD Pharmingen; against cyclin-dependent
kinase 6 (Cdk6) (sc-177) (dilution 1:500) and Cdk2 (sc-163)
(dilution 1:5,000) specific antibodies were obtained from Santa
Cruz Biotechnology, Inc.; and anti-β-actin (A5441) (dilution
1:5,000) was purchased from Sigma-Aldrich (a brand of Merck KGaA).
HRP-conjugated anti-mouse (#7076) (dilution 1:2,000) and
anti-rabbit (#7074) (dilution 1:2,000) IgG secondary antibodies
were obtained from Cell Signaling Technology.
Antibody arrays of apoptosis-related
and phosphorylated receptor tyrosine kinase (p-RTK) proteins
HCC Huh-7 cells (1.0×106 cells/100-mm
dish) were treated with aspirin for 48 h at 37°C and lysed in
PRO-PREP complete protease inhibitor mixture (iNtRON
Biotechnology). The human Apoptosis Antibody Array kit (R&D
Systems) and Human p-RTK Array kit (R&D Systems) were used to
analyze the apoptosis-related and p-RTK proteins in the
aspirin-treated and control cells as per the manufacturer's
protocol. Each array was repeated thrice to validate the
results.
miRNA assay
HCC Huh-7 cells (1.0×106 cells/100-mm
dish) were treated with 2.5 mmol/l aspirin for 48 h and total RNA
was extracted using the miRNeasy Mini kit (Qiagen) according to the
manufacturer's instructions. After confirming the purity and
quantity of each RNA using an Agilent 2100 Bioanalyzer (Agilent
Technologies) and RNA 6000 Nano kit (Agilent Technologies)
respectively, the samples were labeled using a miRCURY Hy3 Power
Labeling kit (Exiqon A/S) and hybridized to a human miRNA Oligo
Chip (v.21; Toray Industries, Inc.). Scanning was conducted using
the 3D-Gene Scanner 3000 (Toray Industries). The 3D-Gene extraction
software version 1.2 (Toray Industries) was used to calculate the
raw signal intensity of the images. The raw data were analyzed
using GeneSpring GX 10.0 software (Agilent Technologies) to assess
the differences in miRNA expression between the aspirin-treated and
control samples. Global normalization was performed on raw data
obtained above the background level. Differentially expressed
miRNAs were determined using the Welch's t-test. The false
discovery rate (FDR) was computed using the Benjamini-Hochberg
method. Hierarchical clustering was performed using the farthest
neighbor method with the absolute uncentered Pearson's correlation
coefficient as a metric. The heatmap was produced with the relative
expression intensity of each miRNA, in which the base-2 logarithm
of intensity was median-centered for each row.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis of miRNAs
We compared the miRNA expression levels obtained in
the miRNA arrays with real-time qPCR measurements to validate the
data for miR-137 and miR-7-5p, which were significantly modulated
according to the microarray analysis. Total RNA was extracted and a
diluted sample (2.0 ng/µl) was used in TaqMan microRNA assays
(Applied Biosystems) to determine the expression levels of miRNAs;
U6 small nuclear RNA (RNU6B) was used as the internal control.
miRNAs were reverse transcribed using the TaqMan microRNA Reverse
Transcription kit (Applied Biosystems). The reverse transcription
reaction mixture was prepared in 15-µl reaction volumes with the
following constituents: 5 µl of RNA, 3 µl of 5X RT primer and 7 µl
of reverse transcription Master Mix. PCRs were performed in the
MicroAmp Fast Optical 96-Well Reaction Plate (Applied Biosystems).
For real-time qPCR, reaction mixtures were prepared in 20-µl
volumes with 2 µl of cDNA, 1 µl of 20X qPCR assay, 7 µl of
nuclease-free water and 10 µl of TaqMan Fast Advanced Master Mix
(Applied Biosystems). The PCRs were carried out using the ViiA7
real-time PCR system (Applied Biosystems) with the following
reaction steps: Hold at 50°C for 2 min, denaturation at 95°C for 20
sec followed by 40 cycles of 1 sec at 95°C and 20 sec at 60°C. The
relative expression levels of miR-137 and miR-7-5p were calculated
using the comparative Ct method according to the following formula:
2−ΔΔCq (ΔCq=miRCq-U6Cq) (15).
Cell transfection
HCC Huh-7 cells were transfected with the miR-137
mimic (3′-GAUGCGCAUAAGAAUUCGUUAUU-5′) (50 nM) or scrambled miRNA
(3′-TTAAGAGGCCUGUGCAAGCCUCUU-5′) as a control (Life Technologies;
Thermo Fisher Scientific, Inc.) using Lipofectamine RNAiMAX Reagent
(Invitrogen; Thermo Fisher Scientific, Inc.). Briefly,
5×105 cells were seeded in 6-well plates with
antibiotic-free medium 1 day before transfection to reach a
confluence of 90% at the time of transfection. The transfection
complex was prepared according to the manufacturer's instructions
and added to the cells, and the plates were incubated in a
humidified atmosphere with 5% CO2 at 37°C. The medium
was replaced 6 h post-transfection. Cell samples were collected 0
or 72 h after transfection for further analysis.
Colony forming assay
Cells prepared for assay were trypsinized for 5 min
and resuspended; 500 µl of cells (500/ml) were seeded in 6-well
plates and to each well we added 1.5 ml DMEM (containing 10% FBS).
The plates were incubated at 37°C in 5% CO2, and the
medium was changed every 3 days until conspicuous colonies were
observed. Finally, the colonies were stained with 0.1% crystal
violet for 5 min at room temperature and counted using a
microscope.
Xenograft model analysis
The animal study was approved by and conducted in
accordance with guidelines set by the Committee on Experimental
Animals of the Kagawa University. Female athymic mice
(BALB/c-nu/nu; 6 weeks old; 19–21 g) were purchased from Japan SLC
(Shizuoka, Japan). The mice were maintained under specified
pathogen-free conditions using a laminar airflow rack and had
continuous free access to sterilized (γ-irradiated) food (CL-2;
Clea Japan, Tokyo) and autoclaved water. Mice were subcutaneously
inoculated with HCC Huh7 cells (1×106 cells/mice) in the
right flank region. Once the xenografts were identifiable as masses
with a maximal diameter >3 mm, the animals were randomly
assigned to 2 groups of 7 animals each. These groups were treated
with 60 mg/kg of aspirin or vehicle (PBS, 10% ethanol) by
intraperitoneal injection every day. The tumor growth was monitored
daily and the tumor volume (V) (mm3) was calculated as
V=tumor length (mm) × tumor width (mm)2/2. All animals
were sacrificed on day 12 after the beginning of the treatment.
Notably, all animals remained alive during this period.
Statistical analysis
GraphPad Prism software version 6.0 (GraphPad
Software, USA) was used for all the analyses. A Student's t-test
was used to determine statistical significance between different
groups. Two-way analysis of variance (ANOVA) or mixed ANOVA was
performed to test the comparisons and corrected by the Tukey's post
hoc test. Results with P-value <0.05 were considered
statistically significant.
Results
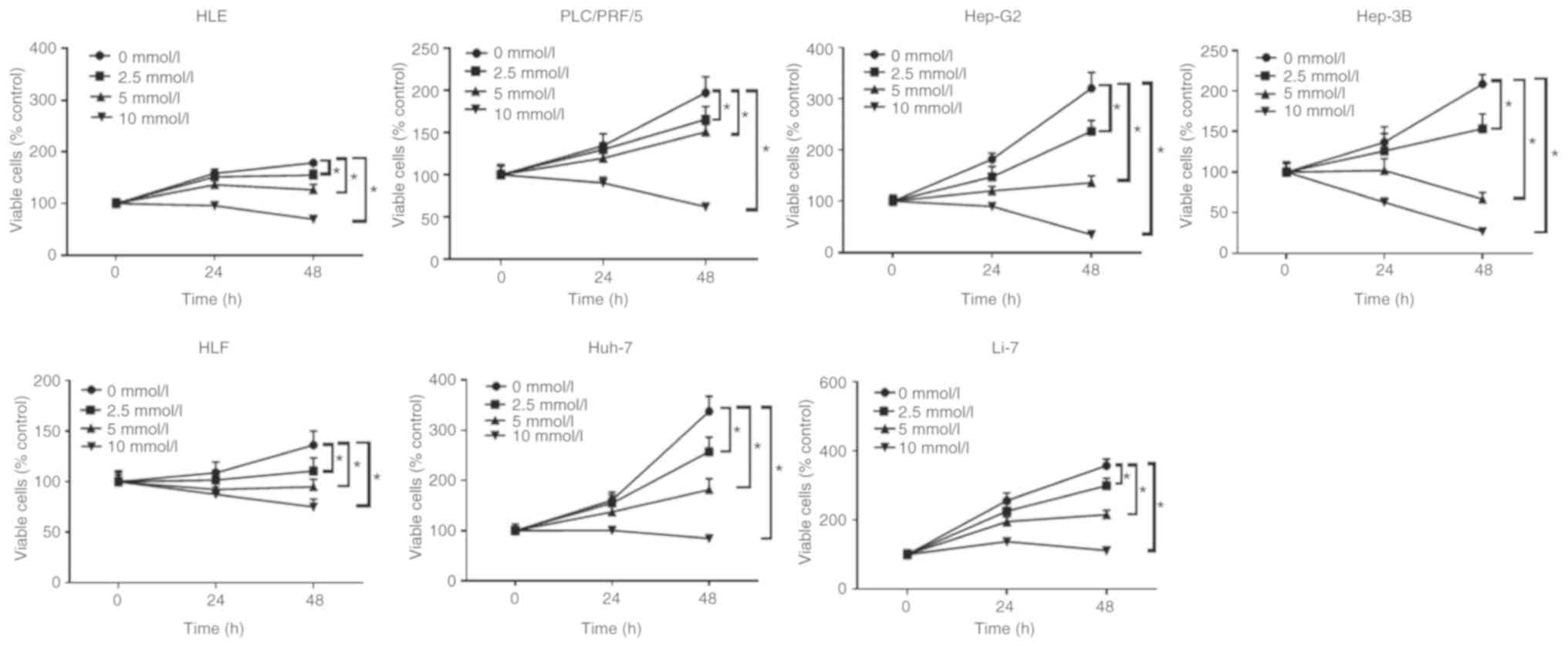
Aspirin inhibits the proliferation of
6 human HCC and a liver cancer cell line
The anti-proliferative effects of aspirin on human
liver cancer cells were determined using the following cell lines:
Huh-7, Hep-G2, Hep-3B, Li-7, HLE, HLF and PLC/PRF/5. Each cell line
was treated with 2.5, 5, or 10 mmol/l aspirin for 48 h, followed by
the estimation of the anti-proliferative activity of aspirin using
a cell proliferation assay. Untreated cells were used as control.
Our results revealed that aspirin significantly inhibited cell
proliferation in all liver cancer cell lines in a dose- and
time-dependent manner (Fig. 1).
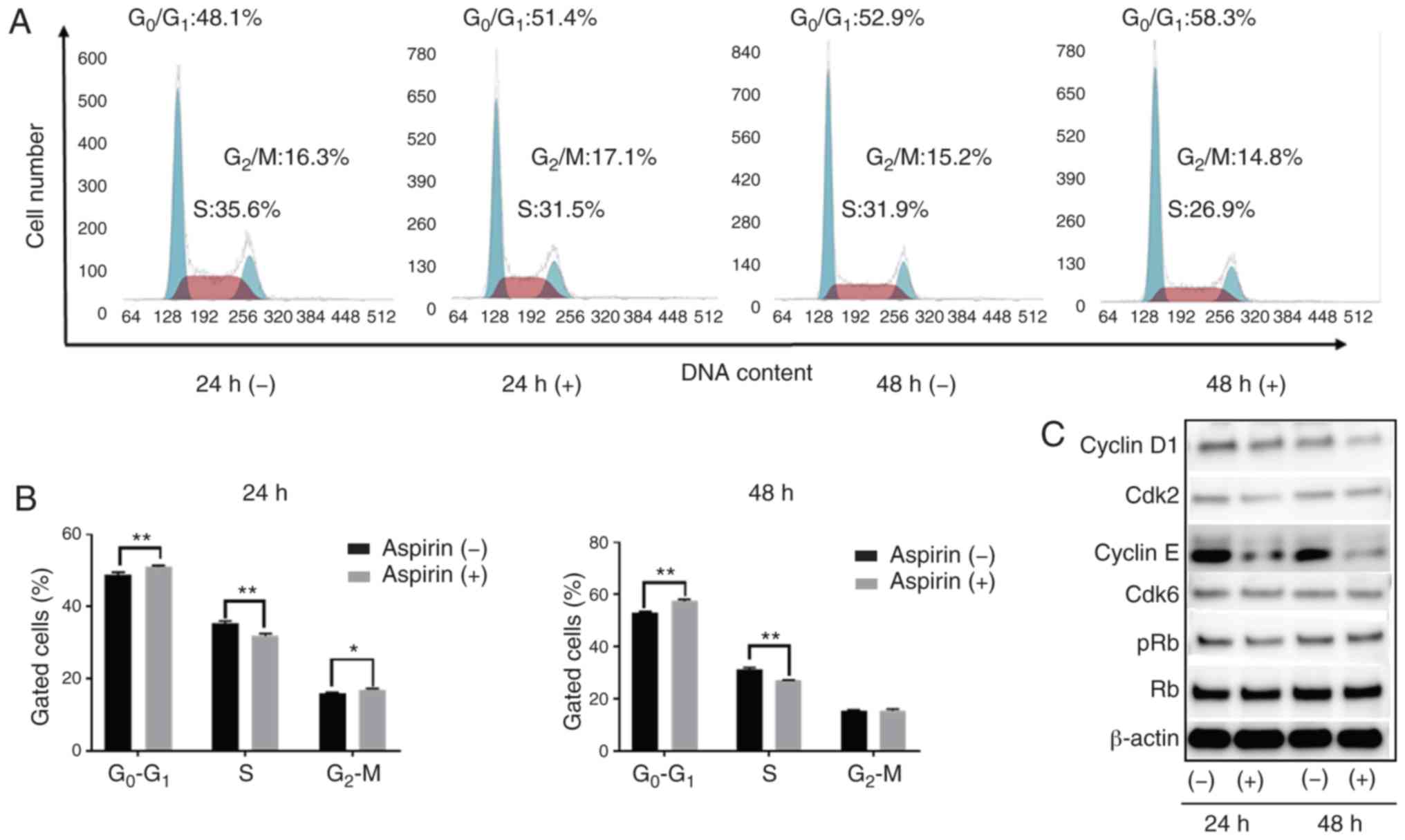
Aspirin induces cell cycle arrest in
the G0/G1 phase and regulates cell
cycle-related proteins in HCC Huh-7 cells
Based on the anti-proliferation assay, HCC Huh-7
cells are sensitive to aspirin. In addition, considering the
xenograft model, Huh-7 cells can be easily transplanted and present
black xenograft tumors, providing clear observation and
measurement. To study whether aspirin affects the cell cycle in
Huh-7 cells, flow cytometry was performed to examine cell cycle
progression and western blotting was carried out to evaluate the
expression of cell cycle-related proteins. Cells were treated with
2.5 mmol/l aspirin for 24 or 48 h, and untreated cells were used as
control. We observed that following aspirin treatment for 24 and 48
h, the cell population in the G0/G1 phase was
significantly increased, while cells in the S phase were
significantly decreased (Fig. 2A and
B). Western blot results showed that aspirin treatment
significantly modulated cyclin E, a key protein implicated in the
transition from G0/G1 phase to the S phase.
Additionally, Cdk2, the catalytic subunit of cyclin E, was
decreased after 24 and 48 h of aspirin treatment. The expression of
cyclin D1 decreased slightly after 48 h of aspirin treatment,
whereas the levels of phosphorylated Rb decreased progressively
after 24 h, suggesting that the treated cells were in G1
phase arrest (Fig. 2C).
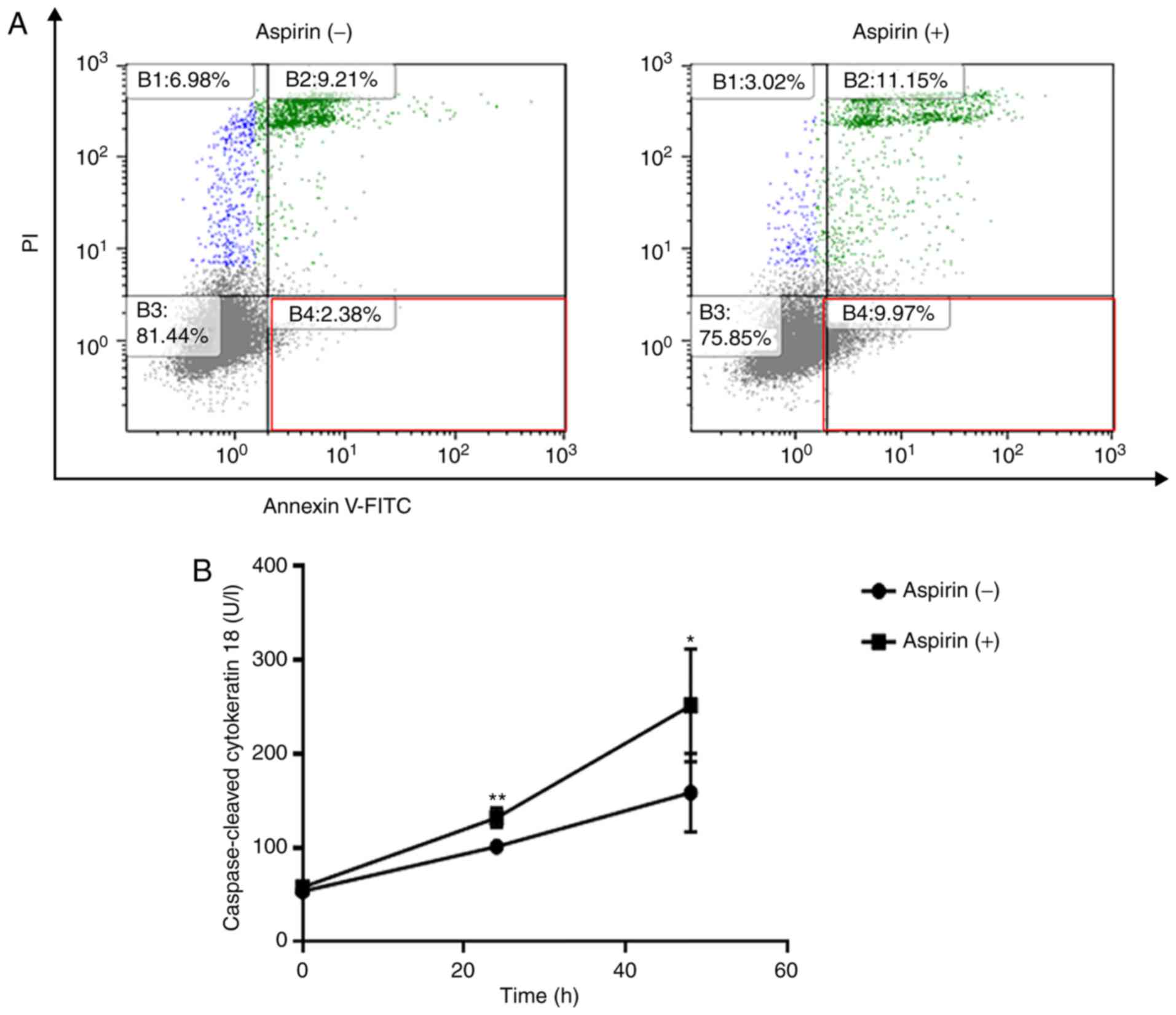
Aspirin partially induces cell
apoptosis in the Huh-7 cells
Flow cytometry was used to detect apoptotic cells
after aspirin treatment. The different quadrants represent living
cells (lower left square), early apoptotic cells (lower right
square), and late apoptotic cells (upper right square). After 48 h
of treatment, a higher percentage of cells (9.97%) were early
apoptotic in the 2.5 mmol/l aspirin-treated group in comparison to
the control group (2.38%, Fig. 3A).
In related results, aspirin significantly increased the levels of
cCK-18 after 24 and 48 h of treatment (Fig. 3B).
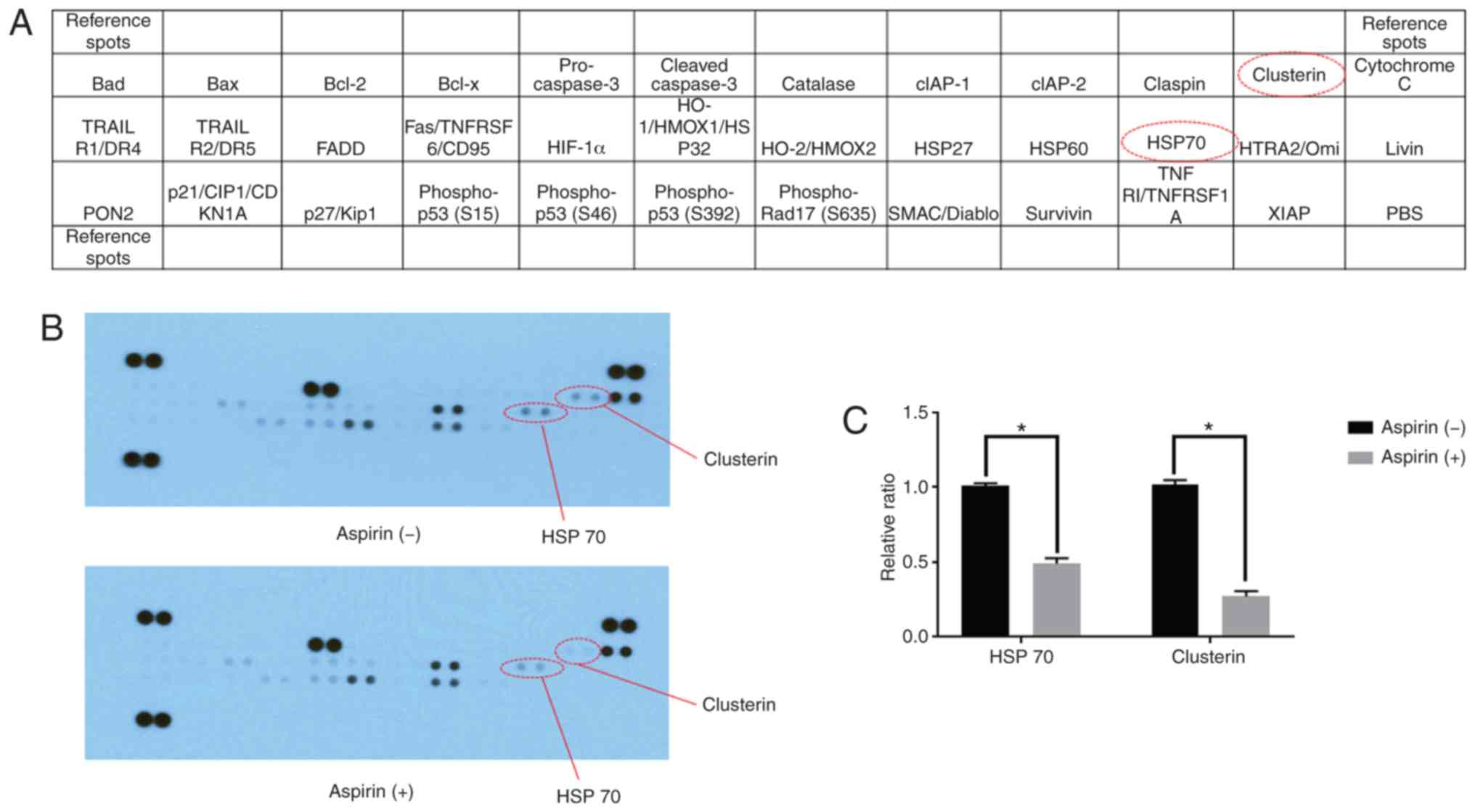
Using an apoptosis array system, we identified
apoptosis-associated proteins involved in the antitumor effects of
aspirin. We observed that aspirin decreased the levels of stress
proteins, HSP 70, and clusterin in the HCC Huh-7 cells (Fig. 4A and B). Densitometric analyses
showed that the intensities of HSP 70 and clusterin spots from
aspirin-treated Huh-7 cells were 48 and 63% weaker than those from
the control cells, respectively (Fig.
4C).
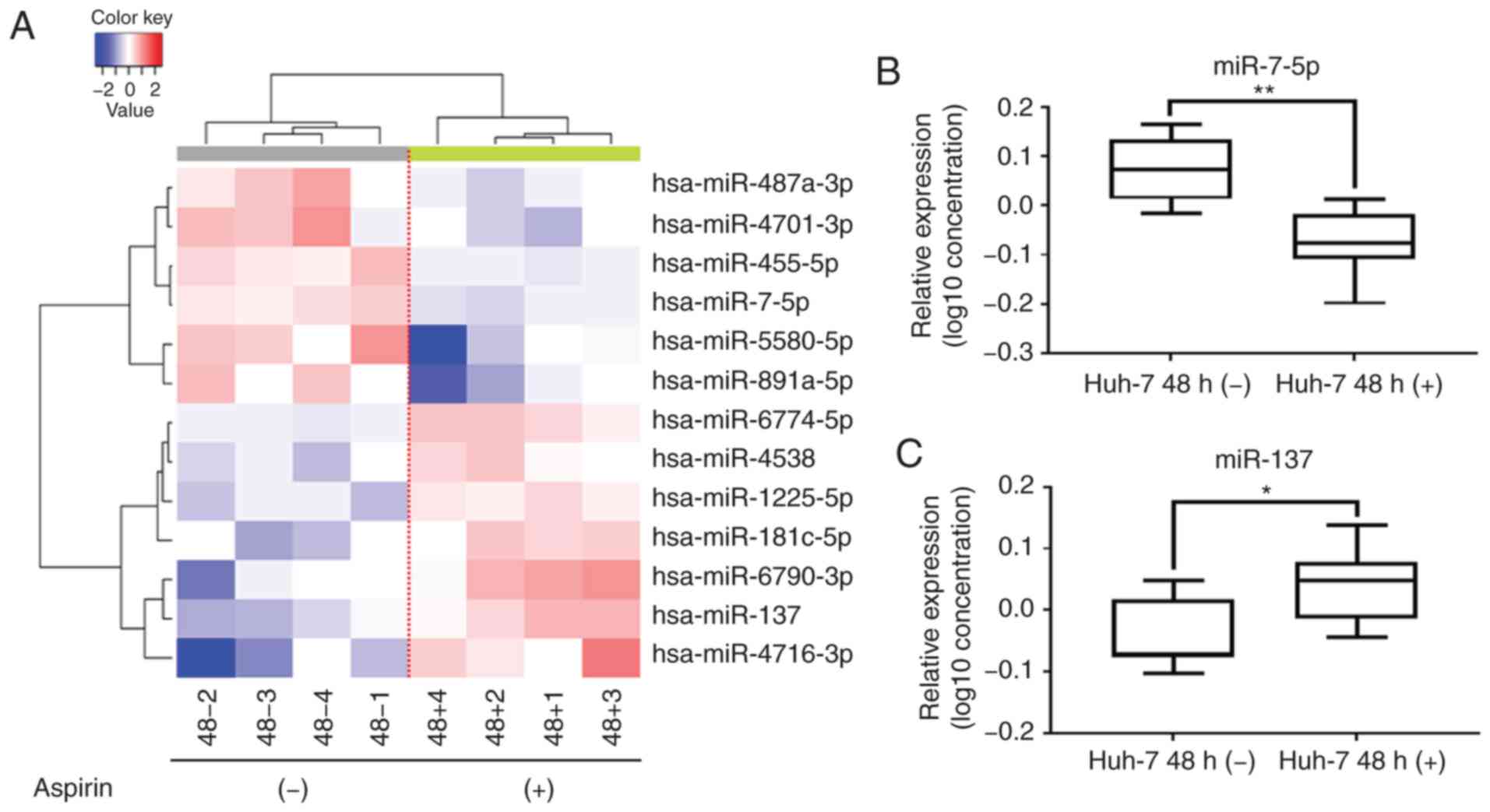
Aspirin affects miRNA expression in
Huh-7 cells
A customized microarray platform was used to analyze
the expression of 2,555 miRNAs in the aspirin-treated or control
Huh-7 cells. Treatment with 2.5 mmol/l aspirin for 48 h induced the
upregulated expression of 7 miRNAs, among which the expression of
miR-137 was found to be significantly upregulated; meanwhile, the
expression of 6 miRNAs was suppressed (Table I), and the expression of miR-7-5p
was found to be significantly downregulated.
 | Table I.Statistical results and chromosomal
locations of miRNAs that exhibited a fold change (FC)>1.5,
FC<0.67, or P<0.05 in HCC Huh-7 cells treated with aspirin
when compared with untreated cells. |
Table I.
Statistical results and chromosomal
locations of miRNAs that exhibited a fold change (FC)>1.5,
FC<0.67, or P<0.05 in HCC Huh-7 cells treated with aspirin
when compared with untreated cells.
| miRNA | Fold change
(treated/untreated) | P-value | Chromosomal
location |
|---|
| Upregulated |
|
|
|
|
hsa-miR-4716-3p | 2.597567946 | 0.0460088 | 15 |
|
hsa-miR-137 | 2.049649799 | 0.00398366 | 1p21.3 |
|
hsa-miR-6790-3p | 1.989701339 | 0.0321397 | 19 |
|
hsa-miR-181c-5p | 1.700223375 | 0.0150924 | 19 |
|
hsa-miR-1225-5p | 1.607423491 | 0.00100997 | 16 |
|
hsa-miR-6774-5p | 1.589034149 | 0.00181634 | 16 |
|
hsa-miR-4538 | 1.504758828 | 0.0281657 | 14q32.33 |
| Downregulated |
|
|
|
|
hsa-miR-455-5p | 0.657530227 | 0.00349551 | 9 |
|
hsa-miR-7-5p | 0.641358785 | 0.000504317 | 9 |
|
hsa-miR-487a-3p | 0.620582971 | 0.0388511 | 14 |
|
hsa-miR-4701-3p | 0.536963485 | 0.040636 | 12 |
|
hsa-miR-891a-5p | 0.520306733 | 0.0329557 | Xq27.3 |
|
hsa-miR-5580-5p | 0.459580271 | 0.0261789 | 14 |
Unsupervised hierarchical clustering analysis was
conducted by calculating Pearson's centered correlation
coefficient, and the results indicated that aspirin-treated Huh-7
cells clustered together (Fig. 5A).
The results of the qPCR assay confirmed that miR-7-5p levels were
significantly decreased (Fig. 5B)
and miR-137 levels were significantly upregulated (Fig. 5C) in the aspirin-treated cells
compared to the untreated cells.
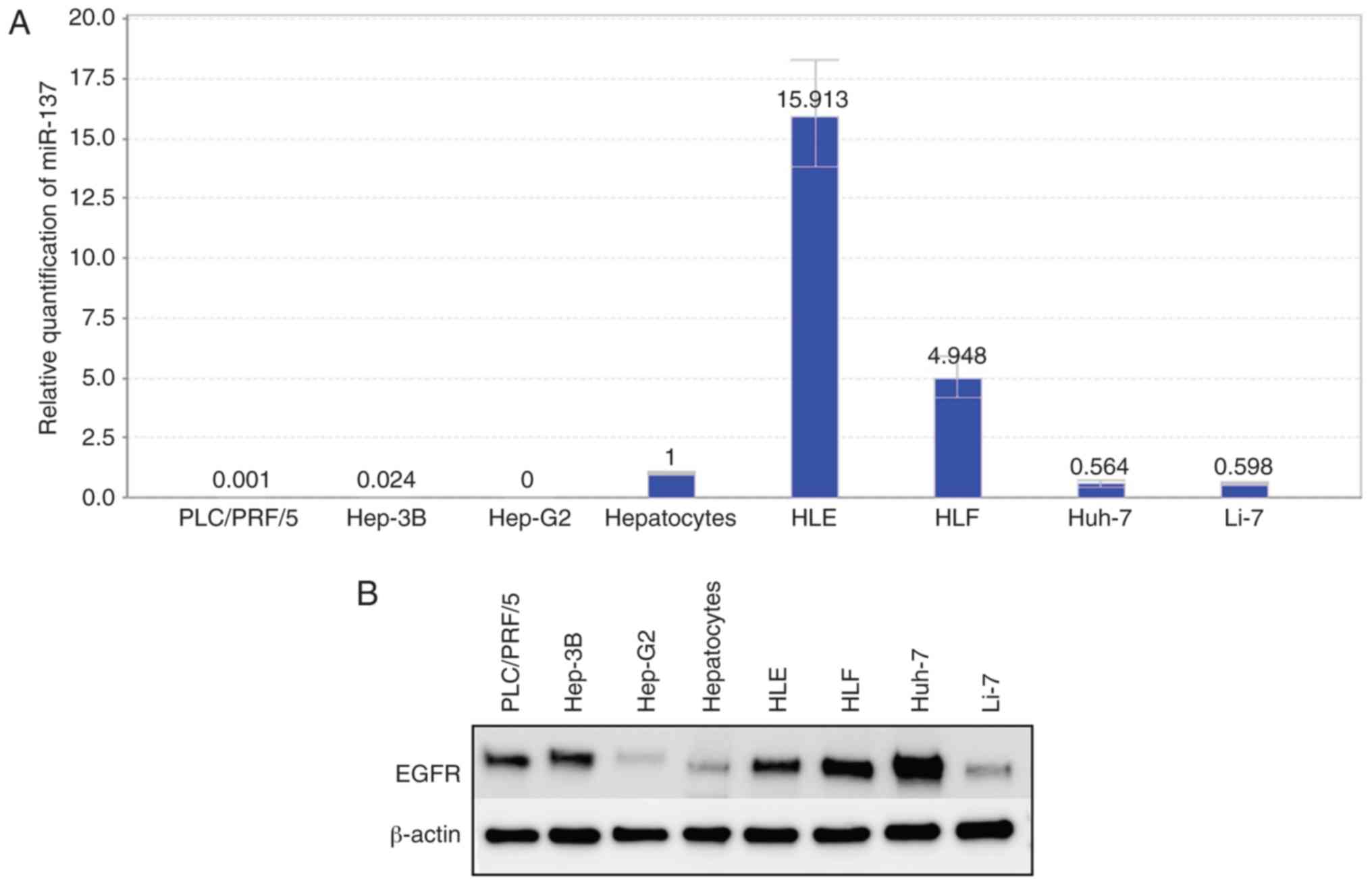
miRNA-137 inhibits the proliferation
of Huh-7 cells and decreases the expression levels of EGFR
The prediction of the miRNA target gene (www.targetscan.org) showed that the mature human
miR-137 target was consistent with the partial sequence of EGFR
(Fig. 6). Relative quantification
of miR-137 was detected in all cell lines. In the differentiated
cell lines, the miR-137 levels were downregulated, and in the
undifferentiated cell lines (HLE, HLF), miR-137 levels were
upregulated (Fig. 7A). Fig. 1 shows that cell lines with low
expression of miR-137 were more sensitive to aspirin treatment. In
addition, the expression levels of EGFR in hepatoma cell lines were
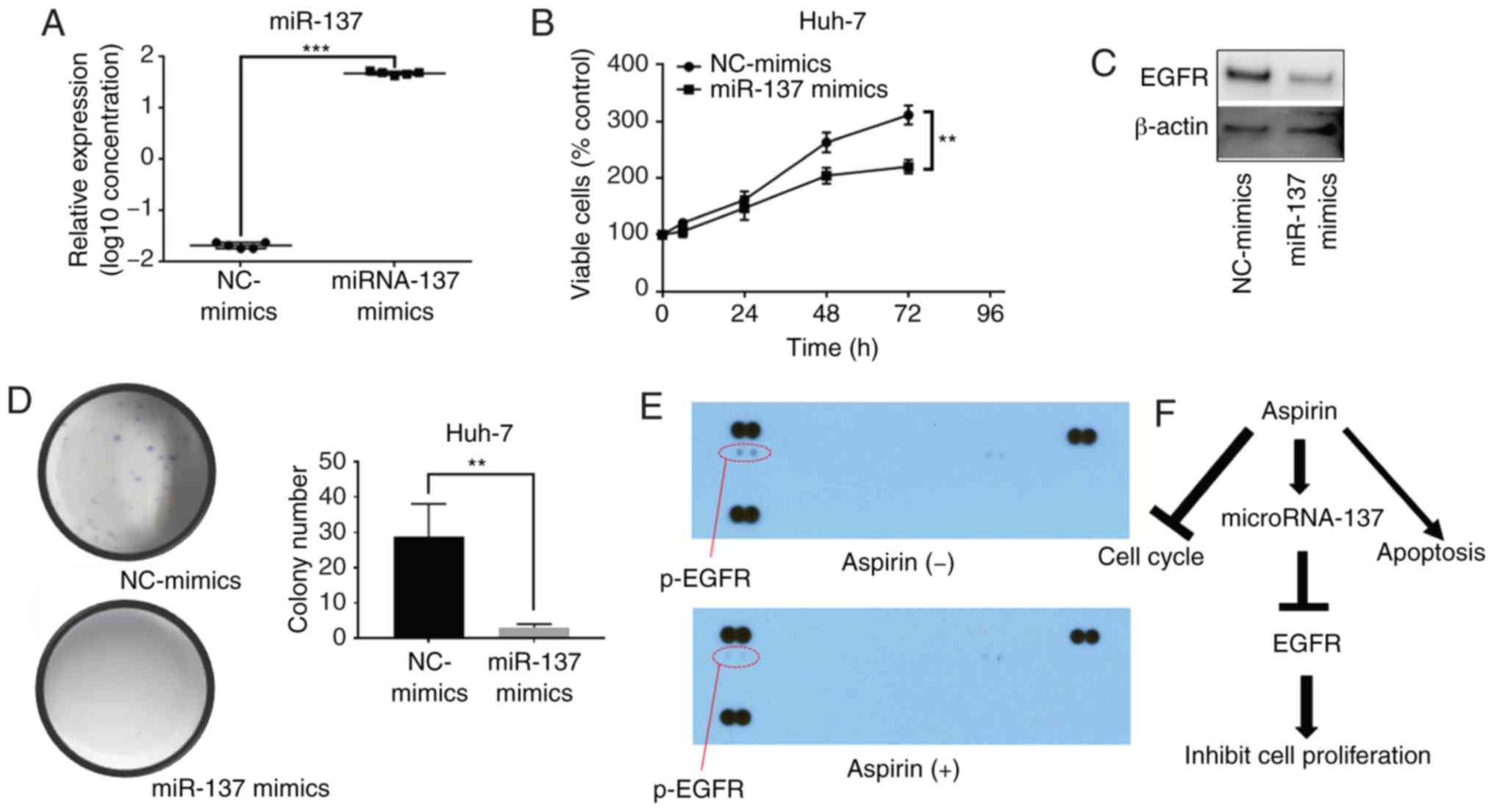
almost as upregulated as those in normal hepatocytes (Fig. 7B). After transfection of miR-137
mimics, miR-137 expression was significantly increased in the Huh-7
cells (Fig. 8A). Our results
indicated that the overexpression of miR-137 inhibited Huh-7 cell
proliferation (Fig. 8B). After
transfection with miR-137, the levels of EGFR were downregulated
(Fig. 8C), and the colony formation
assay indicated that overexpression of miR-137 decreased the cell
proliferation ability of Huh-7 cells (Fig. 8D). Meanwhile, we used a p-RTK array
system to identify the levels of p-EGFR after aspirin treatment,
also showing that aspirin inhibited the expression of p-EGFR in the
treated Huh-7 cells (Fig. 8E).
Aspirin therefore upregulated miR-137 levels, inhibiting EGFR
expression and decreasing cell proliferation ability (Fig. 8F).
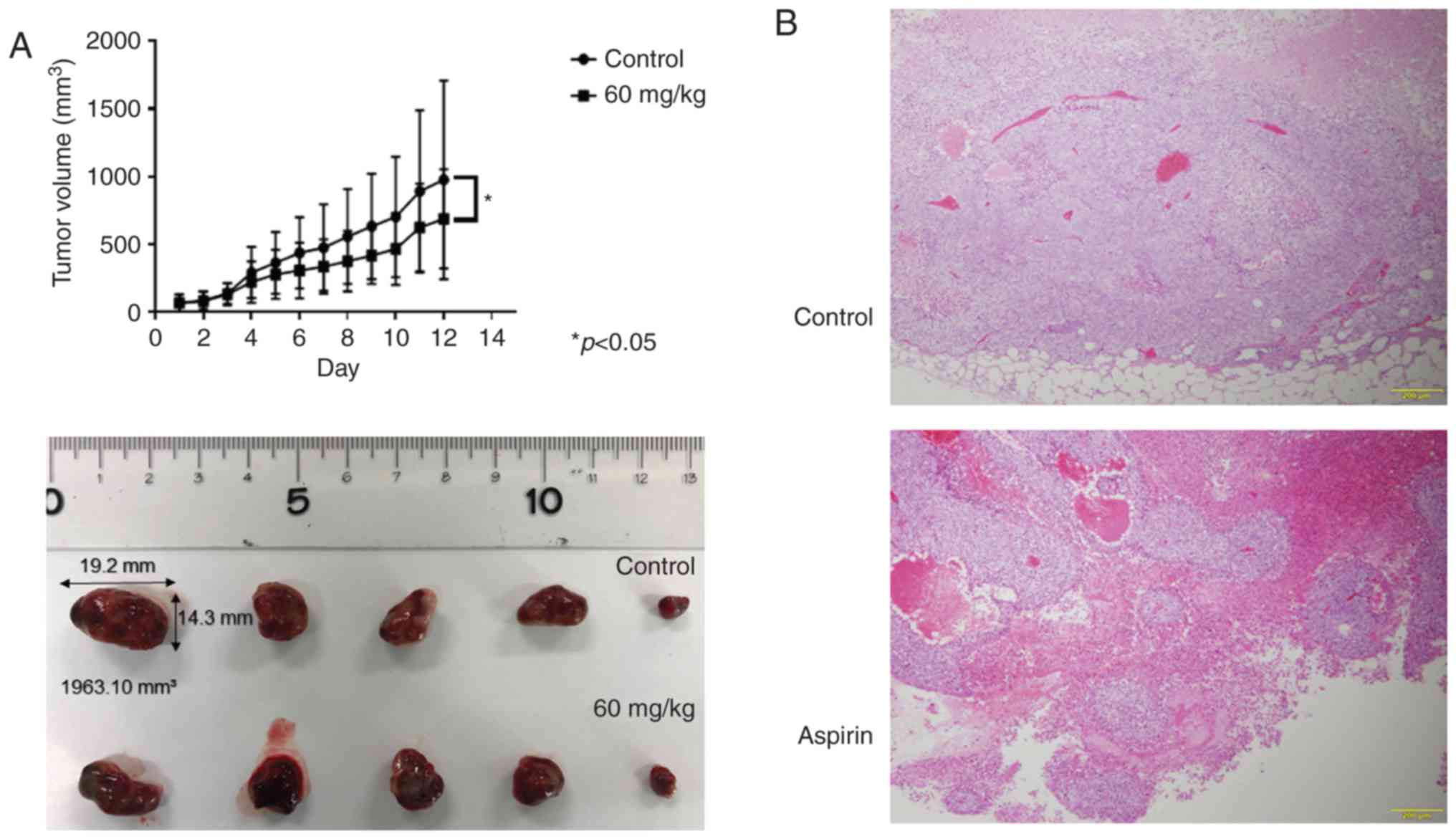
Aspirin inhibits tumor proliferation
in vivo
Based on the results obtained from in vitro
studies, nude mice were injected subcutaneously with Huh-7 cells
followed by intraperitoneal injection of aspirin. Our results
showed that tumor growth was significantly inhibited in the mice
treated with aspirin compared to the untreated mice (P<0.05;
Fig. 9A). Histopathological
sections of xenografted tumors revealed an invariable encapsulation
by connective tissue and no significant histopathological
differences between the aspirin-treated and control mice, except
for the size of the tumor (Fig.
9B). Additionally, the body weight of aspirin-treated mice was
slightly lower than that of the untreated mice (data not shown).
All mice survived for the entire observation period.
Discussion
Recent studies have reported that aspirin may have a
role in the prevention and improvement of HCC outcomes (7,8), but
conflicting results from sparse studies warrant further
investigation of the anti-proliferative effects of aspirin on
hepatocellular carcinoma (HCC) cells. The present study showed that
aspirin was indeed capable of inhibiting the proliferation of HCC
cells.
HCC is associated with high morbidity and a poor
patient survival rate, thus effective therapeutic strategies are
urgently needed for improving the disease outcome (1). Aspirin, a vastly used drug, has been
evaluated for its anti-proliferative activities, and evidence from
multiple studies has indicated that it may function effectively in
the prevention of various tumors, including HCC (10,11).
In the present study, we demonstrated that aspirin exerted
antitumor effects on HCC by inducing cell cycle arrest and
apoptosis and suppressing cell proliferation partially through the
microRNA-137/epidermal growth factor receptor (EGFR) pathway.
Aspirin is used for its antiplatelet, analgesic, and
anti-inflammatory effects over a wide dosage ranging from 75 to
1,200 mg (16). Recent analyses
indicate a significant inverse association between aspirin dose and
the risk of liver cancer (17).
Some studies have shown that similarly to aspirin, salicylic acid
exhibits anti-proliferative and antitumor activity in vitro
and in vivo (18,19), although in this study we
specifically researched only the aspirin concentration.
In the present study, aspirin treatment induced cell
cycle arrest at the G0/G1 phases and
significantly decreased the levels of cell cycle-related proteins
such as cyclin E, cyclin D1, and cyclin-dependent kinase 2 (Cdk2)
in the HCC Huh-7 cells. Cyclin, cyclin-dependent kinase and
cyclin-dependent kinase inhibitor work together in regulating the
cell cycle phase. Cyclin D1 is expressed in the early G1 phase, as
a molecular start of the cell cycle. Cyclin E is mainly involved in
a later phase of the cell cycle and combines with Cdk2 to induce
phosphorylation of the retinoblastoma protein (Rb) protein. We thus
believe that this process, especially in the early G1 phase, is
important. Our results confirm earlier findings of aspirin-induced
cell cycle arrest in many types of tumor cells (20–22).
Our flow cytometry results showed that aspirin treatment
significantly increased the proportion of early apoptotic cells.
Meanwhile, the increased levels of caspase-cleaved cytokeratin 18
(cCK-18) and the decreased levels of clusterin and HSP 70 indicated
apoptosis (23,24). Clusterin is a stress-activated
chaperone protein that plays an important role in various human
malignancies, including HCC, by promoting cell survival (25). It protects HCC cells from
endoplasmic reticulum (ER) stress-induced apoptosis, partially by
interacting with glucose-regulated protein 78 (GRP78) (26,27).
Many reports have indicated that clusterin could inhibit
mitochondrial apoptosis by interacting with BAX (28) and activating the Akt and NF-κB
pathways to promote cancer cell survival (29,30).
In a recent study, clusterin expression in nontumor liver tissue
was found to be an independent adverse prognostic factor for
post-operative HCC patients (31).
HSP 70, an evolutionarily conserved protein, is known to increase
the survivability of cells under stress. Cells with HSP 70
knockdown are sensitive to apoptosis, while HSP 70 overexpression
is shown to inhibit apoptosis (32). Previous research has indicated that
aspirin induces apoptosis by altering the Bax/Bcl-2 ratio and
activating caspase activity (33).
Here, the aspirin-mediated decrease in the levels of clusterin and
HSP 70 may partially induce apoptosis and therefore regulate cell
proliferation in HCC cells.
miRNAs are a class of small (17–25 nucleotides),
noncoding, endogenous, single-stranded RNA molecules that regulate
target gene expression by interfering with their transcription or
by inhibiting translation (34,35).
They are known to regulate the development and progression of
various types of cancer (36,37).
In order to identify the miRNAs associated with the antitumor
effects of aspirin, we used miRNA expression arrays following
aspirin treatment of Huh-7 cells. Our results showed that in
response to aspirin treatment, miR-7-5p levels were significantly
decreased and miR-137 levels were significantly upregulated.
Studies have reported a role of miR-137 in the inhibition of tumor
growth and metastasis by targeting AKT serine/threonine kinase 2. A
low miR-137 expression was significantly associated with lymph node
metastasis, vein invasion, advanced clinical stage and poor
prognosis in HCC (38–40). A related study showed that the
overexpression of miR-137 downregulated the expression of
ZBTB, a proto-oncogene reported to enhance HCC proliferation
and multi-drug resistance (41).
Based on these previous studies on miR-137, it was the first miRNA
to be selected. Here, the results obtained following miR-137 mimic
transfection indicated that miR-137 overexpression inhibited HCC
cell proliferation, in accordance with previous research. In
addition, EGFR is a predicted target gene of miR-137, and in
7 HCC cell lines, EGFR was overexpressed when compared to normal
hepatocytes. We also detected that after transfection with miR-137
in Huh-7 cells, the levels of EGFR were significantly decreased, as
well as the cell proliferation ability. Meanwhile, the
undifferentiated cell lines (HLE, HLF), presented higher miR-137
levels when compared to hepatocytes and were less sensitive to the
aspirin treatment. Furthermore, using p-RTK arrays, we demonstrated
that aspirin reduced EGFR phosphorylation in Huh-7 cells.
Overexpression of activated EGFR, an important signaling molecule
associated with tumorigenesis, has been reported in 40–70% of human
HCCs (42). EGFR can induce cyclin
D1, an important protein in the cell cycle (43,44),
and may be pivotal in activating the sEGFR/Akt/NF-κB/cyclin D1
(43) and JAK2/STAT3 signaling
pathways (45,46). Therefore, EGFR is an important link
in regulating multiple signaling pathways and its inhibition by
aspirin may affect regulatory proteins from multiple signaling
pathways and induce apoptosis. Published research indicates that a
close interaction between COX2 and EGFR signaling pathways, as well
as COX2-derived prostanoids may be important in EGFR activation
during the early stages of liver inflammation and carcinogenesis
(47). Therefore, the miR-137/EGFR
pathway could contribute to the aspirin-mediated antitumor effects
noted in HCC cells. Additionally, miRNA-1225-5p has been reported
as a tumor suppressor (48–50). These other upregulated miRNAs
identified in this study have not yet been reported in HCC.
However, these other upregulated miRNAs may be related to the
antitumor effect of aspirin. In the near future, we aim to study
the effects of these miRNAs on liver cancer. miR-7 was shown to
modulate several signaling pathways and play a significant role in
tumors such as HCC; miR-7 was found to inhibit p21-activated kinase
1 (Pak1) expression (51) and the
PI3K/Akt pathway (52). MiR-7 was
reported as being related to sorafenib resistance (SR) in HCC by
modulating the expression of the TYRO3/phosphoinositide
3-kinase/protein kinase B signal transduction pathway (53). However, other research indicates the
opposite role of miR-7 in the regulation of the complex networks of
oncogenes (54).
A previous in vivo study used aspirin at a 60
mg/kg dose, which is equivalent to the daily human dosage of
240–330 mg (55). Our results
revealed that aspirin markedly suppressed the growth of
subcutaneous HCC tumors in athymic nude mice, suggesting that it
may be effective in the development of a combination therapy with
other anticancer drugs (56) for
the treatment of HCC and possibly of other cancers.
Collectively, our results suggest that aspirin can
inhibit human HCC cell proliferation by its multiple effects on the
suppression of cell cycle-related molecules, induction of cell
apoptosis and regulation of miRNA expression. In addition, we
observed that aspirin may suppress cell proliferation partially
through the miRNA-137/EGFR pathway. Although we observed a
significant modulation in the expression of many miRNAs, the
downstream targets of some of these molecules remain unknown and
the roles of miRNAs in HCC remain to be elucidated.
Acknowledgements
We thank Ms. Kayo Hirose, Ms. Keiko Fujikawa, Ms.
Miwako Watanabe, Ms. Megumi Okamura, and Ms. Fuyuko Kokado for
their assistance.
Funding
No funding was received.
Availability of data and materials
All data supporting the conclusions of the present
study have been documented in this article.
Authors' contributions
TM, HK, AM and TTS conceived and designed the study.
TTS, KF, JG, MN and HI performed the experiments. TTS wrote the
paper. TTS, HI, SL, HY, TN, JT, KT, TT, TH, KO, KT and TM reviewed
and edited the manuscript. All authors read and approved the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Institutional Review Board of the Department of Laboratory Animal
Science of Kagawa University (Kida, Japan).
Patient consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Glossary
Abbreviations
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
NSAID
|
non-steroidal anti-inflammatory
drug
|
|
EGFR
|
epidermal growth factor receptor
|
|
HSP 70
|
heat shock protein 70
|
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Petrick JL, Kelly SP, Altekruse SF,
McGlynn KA and Rosenberg PS: Future of hepatocellular carcinoma
incidence in the United States forecast through 2030. J Clin Oncol.
34:1787–1794. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Michel P, Boige V, Andre T, Aparicio T,
Bachet JB, Dahan L, Guimbaud R, Lepage C, Manfredi S, Tougeron D,
et al: Aspirin versus placebo in stage III or high-risk stage II
colon cancer with PIK3CA mutation: A French randomised double-blind
phase III trial (PRODIGE 50-ASPIK). Dig Liver Dis. 50:305–307.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Henry WS, Laszewski T, Tsang T, Beca F,
Beck AH, McAllister SS and Toker A: Aspirin suppresses growth in
PI3K-mutant breast cancer by activating AMPK and inhibiting mTORC1
signaling. Cancer Res. 77:790–801. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yue W, Zheng X, Lin Y, Yang CS, Xu Q,
Carpizo D, Huang H, DiPaola RS and Tan XL: Metformin combined with
aspirin significantly inhibit pancreatic cancer cell growth in
vitro and in vivo by suppressing anti-apoptotic proteins Mcl-1 and
Bcl-2. Oncotarget. 6:21208–21224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gan H, Lin L, Hu N, Yang Y, Gao Y, Pei Y,
Chen K and Sun B: Aspirin ameliorates lung cancer by targeting the
miR-98/WNT1 axis. Thorac Cancer. 10:744–750. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tao Y, Li Y, Liu X, Deng Q, Yu Y and Yang
Z: Nonsteroidal anti-inflammatory drugs, especially aspirin, are
linked to lower risk and better survival of hepatocellular
carcinoma: A meta-analysis. Cancer Manag Res. 10:2695–2709. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Simon TG, Ma Y, Ludvigsson JF, Chong DQ,
Giovannucci EL, Fuchs CS, Meyerhardt JA, Corey KE, Chung RT, Zhang
X and Chan AT: Association between aspirin use and risk of
hepatocellular carcinoma. JAMA Oncol. 4:1683–1690. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang X, Feng H, Li Z, Guo J and Li M:
Aspirin is involved in the cell cycle arrest, apoptosis, cell
migration, and invasion of oral squamous cell carcinoma. Int J Mol
Sci. 19:E20292018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu YX, Feng JY, Sun MM, Liu BW, Yang G,
Bu YN, Zhao M, Wang TJ, Zhang WY, Yuan HF and Zhang XD: Aspirin
inhibits the proliferation of hepatoma cells through controlling
GLUT1-mediated glucose metabolism. Acta Pharmacol Sin. 40:122–132.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang Z, Fang W, Liu W, Wang L, Liu B and
Liu S and Liu S: Aspirin induces Beclin-1-dependent autophagy of
human hepatocellular carcinoma cells. Eur J Pharmacol. 823:58–64.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Raza H, John A and Benedict S:
Acetylsalicylic acid-induced oxidative stress, cell cycle arrest,
apoptosis and mitochondrial dysfunction in human hepatoma HepG2
cells. Eur J Pharmacol. 668:15–24. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Pavlovic N, Rani B, Gerwins P and
Heindryckx F: Platelets as key factors in hepatocellular carcinoma.
Cancers (Basel). 11:e10222019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Malehmir M, Pfister D, Gallage S,
Szydlowska M, Inverso D, Kotsiliti E, Leone V, Peiseler M,
Surewaard BGJ, Rath D, et al: Platelet GPIbα is a mediator and
potential interventional target for NASH and subsequent liver
cancer. Nat Med. 25:641–655. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dovizio M, Bruno A, Tacconelli S and
Patrignani P: Mode of action of aspirin as a chemopreventive agent.
Recent Results Cancer Res. 191:39–65. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang S, Yu Y, Ryan PM, Dang M, Clark C,
Kontogiannis V, Rahmani J, Varkaneh HK, Salehisahlabadi A, Day AS
and Zhang Y: Association of aspirin therapy with risk of
hepatocellular carcinoma: A systematic review and dose-response
analysis of cohort studies with 2.5 million participants. Pharmacol
Res. 151:1045852020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pathi S, Jutooru I, Chadalapaka G, Nair V,
Lee SO and Safe S: Aspirin inhibits colon cancer cell and tumor
growth and downregulates specificity protein (Sp) transcription
factors. PLoS One. 7:e482082012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Law BK, Waltner-Law ME, Entingh AJ, Chytil
A, Aakre ME, Nørgaard P and Moses HL: Salicylate-induced growth
arrest is associated with inhibition of p70s6k and down-regulation
of c-myc, cyclin D1, cyclin A, and proliferating cell nuclear
antigen. J Biol Chem. 275:38261–38267. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao L and Williams JL: Nitric
oxide-donating aspirin induces G2/M phase cell cycle arrest in
human cancer cells by regulating phase transition proteins. Int J
Oncol. 41:325–330. 2012.PubMed/NCBI
|
|
21
|
Dachineni R, Ai G, Kumar DR, Sadhu SS,
Tummala H and Bhat GJ: Cyclin A2 and CDK2 as novel targets of
aspirin and salicylic acid: A potential role in cancer prevention.
Mol Cancer Res. 14:241–252. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pozzoli G, Marei HE, Althani A, Boninsegna
A, Casalbore P, Marlier LNJL, Lanzilli G, Zonfrillo M, Petrucci G,
Rocca B, et al: Aspirin inhibits cancer stem cells properties and
growth of glioblastoma multiforme through Rb1 pathway modulation. J
Cell Physiol. 234:15459–15474. 2019. View Article : Google Scholar
|
|
23
|
Calderwood SK, Khaleque MA, Sawyer DB and
Ciocca DR: Heat shock proteins in cancer: Chaperones of
tumorigenesis. Trends Biochem Sci. 31:164–172. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mustafi S, Sant DW, Liu ZJ and Wang G:
Ascorbate induces apoptosis in melanoma cells by suppressing
Clusterin expression. Sci Rep. 7:36712017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang F, Kumano M, Beraldi E, Fazli L, Du
C, Moore S, Sorensen P, Zoubeidi A and Gleave ME: Clusterin
facilitates stress-induced lipidation of LC3 and autophagosome
biogenesis to enhance cancer cell survival. Nat Commun. 5:57752014.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kang YK, Hong SW, Lee H and Kim WH:
Overexpression of clusterin in human hepatocellular carcinoma. Hum
Pathol. 35:1340–1346. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang C, Jiang K, Gao D, Kang X, Sun C,
Zhang Q, Li Y, Sun L, Zhang S, Guo K and Liu Y: Clusterin protects
hepatocellular carcinoma cells from endoplasmic reticulum stress
induced apoptosis through GRP78. PLoS One. 8:e559812013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xie D, Sham JS, Zeng WF, Che LH, Zhang M,
Wu HX, Lin HL, Wen JM, Lau SH, Hu L and Guan XY: Oncogenic role of
clusterin overexpression in multistage colorectal tumorigenesis and
progression. World J Gastroenterol. 11:3285–3289. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ammar H and Closset JL: Clusterin
activates survival through the phosphatidylinositol 3-kinase/Akt
pathway. J Biol Chem. 283:12851–12861. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zoubeidi A, Ettinger S, Beraldi E,
Hadaschik B, Zardan A, Klomp LW, Nelson CC, Rennie PS and Gleave
ME: Clusterin facilitates COMMD1 and I-kappaB degradation to
enhance NF-kappaB activity in prostate cancer cells. Mol Cancer
Res. 8:119–130. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kuo PC, Chau IY, Li AF, Chau YP, Hsia CY
and Chau GY: Clusterin expression in nontumor tissue in patients
with resectable hepatocellular carcinoma related with
postresectional survival. J Chin Med Assoc. 82:929–934. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kumar S, Stokes J III, Singh UP, Scissum
Gunn K, Acharya A, Manne U and Mishra M: Targeting Hsp70: A
possible therapy for cancer. Cancer Lett. 374:156–166. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hossain MA, Kim DH, Jang JY, Kang YJ, Yoon
JH, Moon JO, Chung HY, Kim GY, Choi YH, Copple BL and Kim ND:
Aspirin induces apoptosis in vitro and inhibits tumor growth of
human hepatocellular carcinoma cells in a nude mouse xenograft
model. Int J Oncol. 40:1298–1304. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Geng L, Chaudhuri A, Talmon G, Wisecarver
JL, Are C, Brattain M and Wang J: MicroRNA-192 suppresses liver
metastasis of colon cancer. Oncogene. 33:5332–5340. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yuan Q, Cao G, Li J, Zhang Y and Yang W:
MicroRNA-136 inhibits colon cancer cell proliferation and invasion
through targeting liver receptor homolog-1/Wnt signaling. Gene.
628:48–55. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yuan K, Xie K, Fox J, Zeng H, Gao H, Huang
C and Wu M: Decreased levels of miR-224 and the passenger strand of
miR-221 increase MBD2, suppressing maspin and promoting colorectal
tumor growth and metastasis in mice. Gastroenterology.
145:853–864.e9. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Morishita A and Masaki T: miRNA in
hepatocellular carcinoma. Hepatol Res. 45:128–141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Liu LL, Lu SX, Li M, Li LZ, Fu J, Hu W,
Yang YZ, Luo RZ, Zhang CZ and Yun JP: FoxD3-regulated microRNA-137
suppresses tumour growth and metastasis in human hepatocellular
carcinoma by targeting AKT2. Oncotarget. 5:5113–5124. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Cui S, Sun Y, Liu Y, Liu C, Wang J, Hao G
and Sun Q: MicroRNA-137 has a suppressive role in liver cancer via
targeting EZH2. Mol Med Rep. 16:9494–9502. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liang L, Li X, Zhang X, Lv Z, He G, Zhao
W, Ren X, Li Y, Bian X, Liao W, et al: MicroRNA-137, an HMGA1
target, suppresses colorectal cancer cell invasion and metastasis
in mice by directly targeting FMNL2. Gastroenterology.
144:624–635.e4. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhu M, Li M, Wang T, Linghu E and Wu B:
MicroRNA-137 represses FBI-1 to inhibit proliferation and in vitro
invasion and migration of hepatocellular carcinoma cells. Tumour
Biol. 37:13995–14008. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Buckley AF, Burgart LJ, Sahai V and Kakar
S: Epidermal growth factor receptor expression and gene copy number
in conventional hepatocellular carcinoma. Am J Clin Pathol.
129:245–251. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kato K, Gong J, Iwama H, Kitanaka A, Tani
J, Miyoshi H, Nomura K, Mimura S, Kobayashi M, Aritomo Y, et al:
The antidiabetic drug metformin inhibits gastric cancer cell
proliferation in vitro and in vivo. Mol Cancer Ther. 11:549–560.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu W, Yin T, Ren J, Li L, Xiao Z, Chen X
and Xie D: Activation of the EGFR/Akt/NF-κB/cyclinD1 survival
signaling pathway in human cholesteatoma epithelium. Eur Arch
Otorhinolaryngol. 271:265–273. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Li B, Ding CM, Li YX, Peng JC, Geng N and
Qin WW: Over-regulation of microRNA-133b inhibits cell
proliferation of cisplatin-induced non-small cell lung cancer cells
through PI3K/Akt and JAK2/STAT3 signaling pathway by targeting
EGFR. Oncol Rep. 39:1227–1234. 2018.PubMed/NCBI
|
|
46
|
Han W and Lo HW: Landscape of EGFR
signaling network in human cancers: Biology and therapeutic
response in relation to receptor subcellular locations. Cancer
Lett. 318:124–134. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Dajani OF, Meisdalen K, Guren TK, Aasrum
M, Tveteraas IH, Lilleby P, Thoresen GH, Sandnes D and
Christoffersen T: Prostaglandin E2 upregulates EGF-stimulated
signaling in mitogenic pathways involving Akt and ERK in
hepatocytes. J Cell Physiol. 214:371–380. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li D, Chi G, Chen Z and Jin X:
MicroRNA-1225-5p behaves as a tumor suppressor in human
glioblastoma via targeting of IRS1. OncoTargets Ther. 11:6339–6350.
2018. View Article : Google Scholar
|
|
49
|
Sun P, Zhang D, Huang H, Yu Y, Yang Z, Niu
Y and Liu J: MicroRNA-1225-5p acts as a tumor-suppressor in
laryngeal cancer via targeting CDC14B. Biol Chem. 400:237–246.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang S, Chen X, Zhang Z and Wu Z:
MicroRNA-1225-5p inhibits the development and progression of
thyroid cancer via targeting sirtuin 3. Pharmazie. 74:423–427.
2019.PubMed/NCBI
|
|
51
|
Reddy SD, Ohshiro K, Rayala SK and Kumar
R: MicroRNA-7, a homeobox D10 target, inhibits p21-activated kinase
1 and regulates its functions. Cancer Res. 68:8195–8200. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Fang Y, Xue JL, Shen Q, Chen J and Tian L:
MicroRNA-7 inhibits tumor growth and metastasis by targeting the
phosphoinositide 3-kinase/Akt pathway in hepatocellular carcinoma.
Hepatology. 55:1852–1862. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kabir TD, Ganda C, Brown RM, Beveridge DJ,
Richardson KL, Chaturvedi V, Candy P, Epis M, Wintle L, Kalinowski
F, et al: A microRNA-7/growth arrest specific 6/TYRO3 axis
regulates the growth and invasiveness of sorafenib-resistant cells
in human hepatocellular carcinoma. Hepatology. 67:216–231. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Cheng AM, Byrom MW, Shelton J and Ford LP:
Antisense inhibition of human miRNAs and indications for an
involvement of miRNA in cell growth and apoptosis. Nucleic Acids
Res. 33:1290–1297. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Mikami J, Kurokawa Y, Takahashi T,
Miyazaki Y, Yamasaki M, Miyata H, Nakajima K, Takiguchi S, Mori M
and Doki Y: Antitumor effect of antiplatelet agents in gastric
cancer cells: An in vivo and in vitro study. Gastric Cancer.
19:817–826. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Li S, Dai W, Mo W, Li J, Feng J, Wu L, Liu
T, Yu Q, Xu S, Wang W, et al: By inhibiting PFKFB3, aspirin
overcomes sorafenib resistance in hepatocellular carcinoma. Int J
Cancer. 141:2571–2584. 2017. View Article : Google Scholar : PubMed/NCBI
|