Introduction
RNA binding proteins (RBPs) are an important family
of proteins related to RNA metabolism. They dynamically bind RNA to
form a variety of complexes, including ribonucleoprotein particles
(RNPs) (1). RBPs play a significant
role in various cellular processes, including RNA splicing,
polyadenylation, RNA editing, RNA transport, maintenance of RNA
stability and degradation, intracellular localization, and
translation control (2). Recent
studies have revealed that RBPs are closely associated with
muscular dystrophy, neurological disease, cancer, and Mendelian
disease (3,4).
Staufen is a member of the double-stranded
RNA-binding protein family, which is involved in mRNA transport and
localization to different subcellular compartments (5). Staufen proteins contain several
double-stranded RNA-binding domains (6). Double-stranded RNA-binding protein
Staufen homolog 1 (STAU1) plays a crucial role in mRNA output,
re-localization, translation, and STAU1-mediated mRNA decay (SMD)
(7,8). SMD is also involved in the
developmental process, such as myogenesis and adipogenesis, and
possibly in angiogenesis (9).
STAU1, by binding to the inverted repeat Alu elements (IRAlus) on
the 3′UTR of mRNA, inhibits retention of IRAlus-containing mRNA in
the nuclei, thereby enhancing its nuclear export. Moreover, STAU1
binding to the IRAlus on the 3′UTR of mRNA can promote translation
of these mRNAs by inhibiting the binding of protein kinase R (PKR)
(10). In addition, STAU1 is also
an important mRNA transport factor in neurons. After binding to the
3′UTR of mRNA, STAU1 can induce translation-dependent mRNA
degradation through direct interaction with UPF1. An upregulation
of transcripts due to depletion of STAU1 and UPF1 may play a
pivotal role in the differentiation process (11). STAU1 expression can be used to
regulate neuronal differentiation (12). Studies have revealed that STAU1
plays a substantial role in the immunity against the influenza
virus and human immunodeficiency virus type I (HIV-1) (13–15).
In order to study the possible biological functions
of STAU1, a STAU1-regulated transcriptome in HeLa cells was
obtained. To determine the gene expression profile and the
alternative splicing (AS) events in the genome, which are regulated
by STAU1, high-throughput RNA sequencing (RNA-seq) was employed. In
addition, the related results were validated in HeLa cells. A
previous research demonstrated that PTB knockdown converted highly
transformed HeLa cells to neuronal-like cells. They extended this
analysis to multiple cell types of diverse origin, including human
embryonic carcinoma stem cells (NT2), mouse neural progenitor cells
(N2A), human retinal epithelial cells (ARPE19), and primary mouse
embryonic fibroblasts (MEFs). Upon PTB knockdown, all of these
cells exhibited a neuronal-like morphology (16). Comparative transcriptome analysis
revealed that STAU1 can selectively regulate the expression of
inflammatory genes, including interferon-induced protein with
tetratricopeptide repeats 2 (IFIT2), 2′-5′-oligoadenylate
synthetase-like protein (OASL), and interferon-induced protein with
tetratricopeptide repeats 3 (IFIT3). Furthermore, it was revealed
that AS of genes widespread in the ‘nerve growth factor receptor
signaling pathway’, including PLEKHG2, ARHGEF11, NR4A1, PDGFB,
FGFR4, and RALGDS, was regulated by STAU1. Overexpression of STAU1
was closely associated with inflammation. Collectively, the present
research defined a potential regulatory pattern in which AS of
inflammatory and immune response genes was regulated by STAU1. It
indicated that STAU1 may be involved in the proliferation and
survival of neurons (17),
differentiation, cell growth and apoptosis (18), post-damage repair and regeneration,
neurite outgrowth and retraction (19), and myelination (20).
Materials and methods
Cloning and construction of
plasmids
CE Design 1.04 software (Vazyme Biotech Co., Ltd.).
was employed to design primers for Hot Fusion (21). There was a gene-specific sequence
and a pIRES-hrGFP-1a vector (part no. 240031; Agilent Technologies,
Inc.) sequence (17–30 bp) in each primer. The designed primers
contained two parts of a sequence, the sequence before ‘ATG’ in the
forward (F)-primer or ‘AGC’ in the reverse (R)-primer matches of
the pIRES-hrGFP-1a vector and the other part of the sequence
belonged to STAU1 gene sequence, presented as follows:
F-primer: AgcccgggcggatccgaattcATGAAACTTGGAAAAAAACCAATGT and
R-primer: GtcatccttgtagtcctcgagAGCACCTCCCACACACAGACA.
EcoRI and XhoI (NEB) were used to
digest pIRES-hrGFP-1a vectors for ~2–3 h at 37°C, which were then
subjected to agarose (1.0%) gel electrophoresis and purification
using Qiagen spin-column-based kit (Qiagen GmbH) according to the
manufacturer's instructions. TRIzol reagent (Ambion) was used to
extract total RNA from HeLa cells, which was used to synthesize
cDNA by oligo dT primer. The cDNA is the template to amplify insert
fragments of the STAU1 gene using the primers by polymerase
chain reaction (PCR). The vectors and insert fragments were added
to a microtube and ligated by ClonExpress II One Step Cloning kit
(Vazyme Biotech Co., Ltd.). The ligand product was transformed into
E. coli DH5α, which was then added to a Luria-Bertani (LB) agar
plate, containing 1 µl/ml ampicillin (Sigma-Aldrich; Merck KGaA),
followed by overnight incubation at 37°C. The template used for PCR
was DNA extracted from DH5α cells, using DNA polymerase 2X Green
Taq Mix (Vazyme Biotech Co., Ltd.). The sequences of the Universal
primers were as follows: Forward (F)-primer: AATTAACCCTCACTAAAGGG
and Reverse (R)-primer: GTCCTTATCATCGTCGTCTT.
PCR procedures were carried out as follows: The
samples were first denatured at 95°C for 5 min, denaturation at
95°C for 3 min, followed by 28 cycles of annealing at 55°C for 30
sec and extension at 72°C for 1 min. Universal primers were
employed to screen colonies. Sanger sequencing (22) was used to verify the insert
sequence.
Cell culture and transfections
Human cervical carcinoma cell line, HeLa (CCTCC no.
GDC0009) was obtained from the China Center for Type Culture
Collection. HeLa cells were cultured at 37°C with 5% CO2
in Dulbecco's modified Eagle's medium (DMEM; Gibco; Thermo Fisher
Scientific, Inc.) with 10% fetal bovine serum (FBS; GE Healthcare),
100 µg/ml streptomycin (Hyclone), and 100 U/ml penicillin
(Hyclone). Plasmid (400 ng) (part no. 240031; Agilent Technologies,
Inc.) was transfected into HeLa cells using Lipofectamine™ 2000
Transfection Reagent (cat. no. 11668019; Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Transfected cells were harvested after 48 h for reverse
transcription-quantitative PCR (RT-qPCR) and western blot
analysis.
Gene overexpression
STAU1 overexpression was determined using
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as the control.
cDNA was synthesized according to the standard instructions
followed by real-time quantitative PCR. RT-qPCR was conducted on
the Bestar SYBR Green RT-PCR Master Mix (DBI Bioscience). The
primers are presented in Table SI.
The concentrations of the transcripts were then normalized to GAPDH
mRNA levels using the 2−ΔΔCq method (23). The data were analyzed by Student's
t-test using GraphPad Prism software (version 8.0; GraphPad
Software, Inc.).
Cell proliferation assay
Cell proliferation was measured by the 3-(4,
5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT)
assay. Briefly, 1×103 cells were seeded in 96-well
plates in triplicate. After 48 h of transfection, 5 mg/ml MTT
solution was added into the cells. After 4 h of incubation at 37°C,
the MTT solution was removed. The insoluble MTT was dissolved in
DMSO. Absorbance at 490 nm was measured using a Benchmark Plus
microplate reader (Bio-Rad Laboratories, Inc.).
RNA isolation and high-throughput
sequencing
Four groups of samples were prepared, namely,
control cells and overexpression (OE)-STAU1 cells (two biological
replicates). Prior to RNA isolation, HeLa cells were first
harvested. TRIzol reagent (Ambion; Thermo Fisher Scientific, Inc.)
was used to isolate total RNA, which was then purified by
phenol-chloroform and treated with RQ1 DNase (Promega Corporation)
to eliminate DNA. The quantity and quality of the RNA were verified
by using SmartSpec Plus (Bio-Rad Laboratories, Inc.) to detect the
absorbance at a wavelength of 260–280 nm. Agarose (1.5%) gel
electrophoresis was employed to detect RNA integrity.
RNA-seq library was prepared using VAHTS Stranded
mRNA-seq Library Prep kit (cat. no. NR602; Vazyme Biotech Co.,
Ltd.), and 1 µg total RNA was used for one sample. After
polyadenylated mRNAs were purified, they were fragmented and
double-strand cDNAs were produced. The double-strand cDNAs were
subjected to end repair, and poly(A) tails were added. They were
then ligated to VAHTS RNA Adapters (Vazyme Biotech Co., Ltd.) and
digested by heat-labile uracil-DNA glycosylase (UDG). Before
sequencing, amplification, purification, and quantification of the
single-strand cDNA were performed, the quantity of the cDNA was
redetermined by qPCR using Agilent 2100 (Agilent Technologies,
Inc.), and was finally stored at −80°C.
The library was prepared according to the
manufacturer's protocol, which was applied to 150-nt paired-end
sequencing using Illumina HiSeq X Ten system (Illumina, Inc.).
ABLife performed the sequencing using the sequencing kit provided
from Illumina, Inc.
Data processing and alignment
Raw reads containing more than 2-N bases were first
discarded. FASTX-Toolkit (Ver. 0.0.13) was used to trim the
adaptors and bases of low quality from raw sequencing reads, and
short reads less than 16-nt were removed, filtered for quality
(fastq_quality_filter -q 25 -p 80) and against artifact sequences
(fastx_artifacts_filter) and collapsed (fastx_collapse). Base
quality Q30 was used to indicate the proportion of bases with a
sequencing error rate <0.1%. RNA-seq data presented in this
study have been deposited in the Gene Expression Omnibus of NCBI
and are accessible through GEO Series accession number GSE136890.
Using TopHat v.2.0.10 (24), clean
reads were mapped to the GRch38 genome and four mismatches were
allowed. For calculation of fragments per kilobase of transcript
per million fragments mapped (FPKM) (25) and gene read counting, we applied
specifically mapped reads, and by calculating the Pearson
correlation coefficient (PCC) for cluster analysis.
Analysis of differentially expressed
genes (DEGs)
The R Bioconductor package edgeR (26) was utilized to screen the DEGs. A
false-discovery rate (FDR)<0.05 and fold-change (FC)>2 or
<0.5 were set as the cut-off points for identifying DEGs. Next,
a volcano map was drawn to reveal the number of DEGs.
AS analysis
By applying the ABLas algorithm (27,28),
regulated differential splicing events (RDSEs) and differential
splicing events (DSEs) were identified. Briefly, 10 types of DSEs
were detected by ABLas on the basis of splice junctions directly
extracted from mapping reads, including exon skipping (ES),
alternative 5′splice site (A5SS), alternative 3′ splice site
(A3SS), intron retention (IR), mutually exclusive exons (MXE),
mutually exclusive 5′UTRs (5pMXE), mutually exclusive 3′UTRs
(3pMXE), cassette exon, A3SS and ES, and A5SS and ES. Sashimi plot
by IGV Tools was used for AS analysis.
To assess RBP-regulated DSEs, Student's t-test was
performed to evaluate the significance of AS events. The events,
which were significant at a P-value equal to 5%, were considered
RBP-regulated DSEs.
We also analyzed the overlapping genes from the DEGs
and regulated alternative splicing events (RASEs) between the
samples were defined and quantified by using the ABLas pipeline;
and a Venn diagram was drawn.
AS events and DEGs are validated by
RT-qPCR
In order to validate RNA-seq data, RT-qPCR was
performed. The primers used for RT-qPCR are presented in Table SI. RNA was reversely-transcribed
into cDNA using an M-MLV Reverse Transcriptase (Vazyme Biotech Co.,
Ltd.). RT-qPCR was performed by StepOne Real-Time PCR System using
the SYBR-Green PCR Reagent Kit (Yeasen Biotechnology Co., Ltd.).
PCR procedures were carried out as follows: The samples were first
denatured at 95°C for 10 min, followed by 40 cycles of denaturation
at 95°C for 15 sec, and then annealed and extended at 60°C for 1
min. The procedures were repeated three times for all samples. The
RNA expression levels of all the DEGs were normalized against those
of GAPDH.
In addition, an RT-qPCR assay was undertaken for DSE
validation. The primers used for detecting DSEs are presented
Table SI. To detect alternative
isoforms, a boundary-spanning primer of constitutive and
alternative exons was used, as well as an opposing primer in one
constitutive exon. The boundary-spanning primer of the alternative
exon was designed according to a ‘model exon’ to detect model
splicing, or an ‘altered exon’ to detect altered splicing.
Functional enrichment analysis
To sort out functional categories of DEGs, KOBAS 2.0
server (29) was employed to
identify Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways
and Gene Ontology (GO) terms. Enrichment was assessed using the
hypergeometric test followed by Benjamini-Hochberg FDR adjustment
for P-values. The heatmap was constructed by calculating the
Pearson correlation coefficient (PCC) of the DEGs.
Results
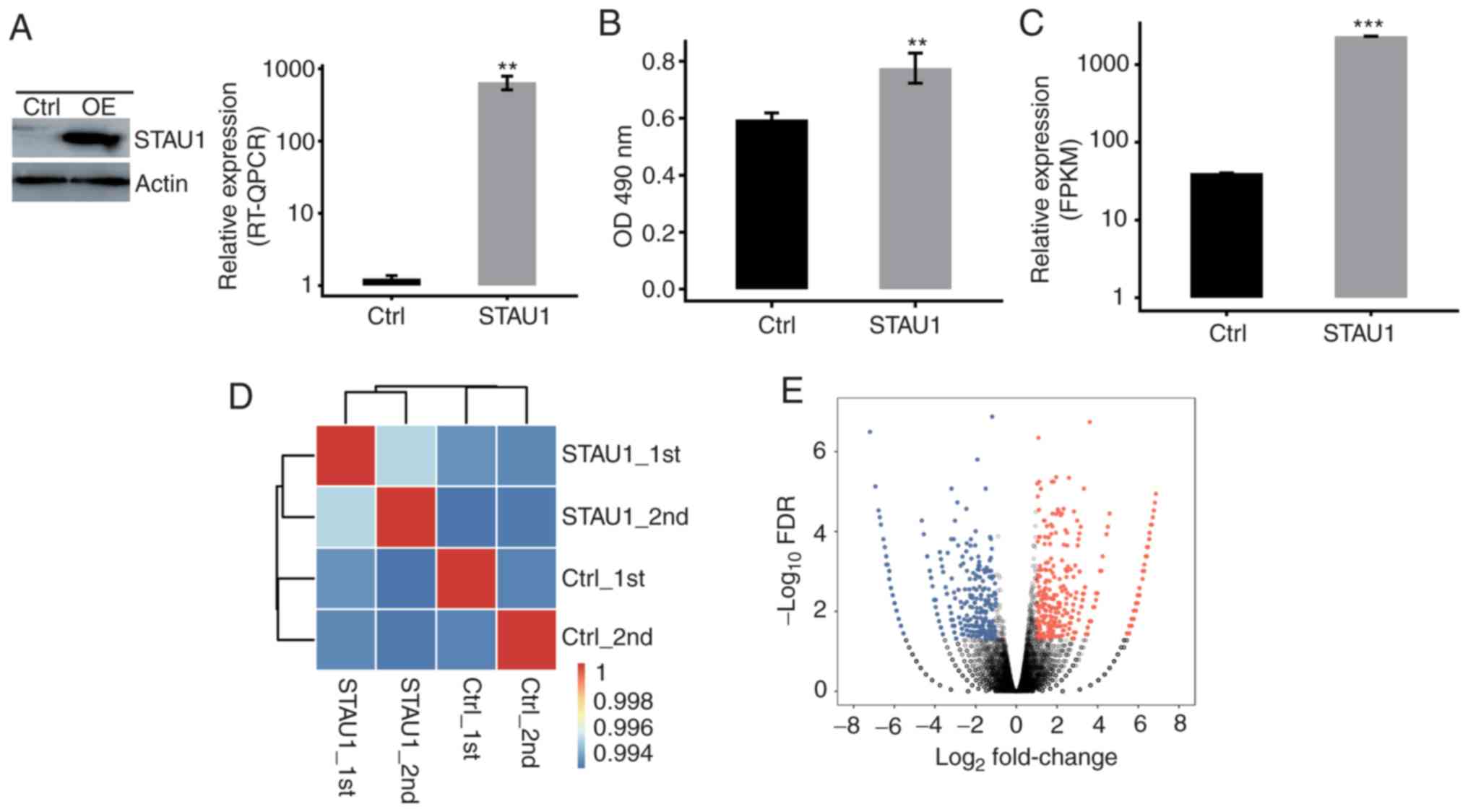
Overexpression of FLAG-tagged STAU1
promotes cell proliferation
In order to analyze the expression of
STAU1-expressing vectors and its influence on the proliferation of
human HeLa cells, the STAU1-transfected cells with GFP label and
flag label (fused with the target gene) were successfully
constructed. In addition, cells transfected with blank controls
were considered as the negative control. After 48 h of
transfection, the expression level of GFP was detected to indicate
whether the transfection was successful. The cells were harvested
and detected by RT-qPCR and western blot assays. The results
indicated a significant overexpression after cell transfection with
STAU1-expressing vectors (Fig. 1A).
Thereafter, proliferation of STAU1-transfected cells was detected
by MTT assay. Compared with the control group, overexpression of
STAU1 promoted cell proliferation (Fig.
1B).
RNA-seq profiling of the
transcriptional response to STAU1 overexpression
In order to assess STAU1-mediated transcriptional
regulation in human HeLa cells, four groups of samples were
prepared, namely, control cells and overexpression (OE)-STAU1 cells
(two biological replicates). Total RNA extraction was carried out
for the 4 groups of samples, and the cDNA library was prepared.
Then, the library was subjected to paired-end sequencing on
Illumina HiSeq X Ten to extract high-quality transcriptome data.
Quality analysis of clean reads indicated that the mean Q30 quality
score was 93.93% (30). Next,
high-quality clean reads were aligned against GRCH38 human
reference genome using TopHat2 software (Table I). RNA-Seq data were analyzed and
the expression levels of STAU1 were quantified, which further
demonstrated overexpression of STAU1 (Fig. 1C). Correlation analysis was
undertaken to determine the variability in the gene expression
level between each pair of the samples. Moreover, cluster analysis
was performed between the samples (Fig.
1D). As revealed in Fig. 1D,
there was a correlation between STAU1 OE cells and control cells;
there was also a significant correlation between the biological
replicates. With analysis of DEGs among samples, criteria for
significant difference were set to FC≥2 or ≤0.5 and FDR<0.05. A
volcano plot was drawn and the results revealed that 801
significant DEGs were related to STAU1 overexpression. Among them,
423 upregulated genes and 378 downregulated genes were identified
(Fig. 1E). This indicated that
STAU1 plays an extensive transcriptional regulatory role in HeLa
cells.
 | Table I.Mapping of the high-quality clean
reads on the reference genome. |
Table I.
Mapping of the high-quality clean
reads on the reference genome.
| Sample | STAU1_1st | STAU1_2nd | Ctrl_1st | Ctrl_2nd | Mean |
|---|
| Raw reads | 83246680 | 92553878 | 83563062 | 80069538 | 84858289.5 |
| Clean reads | 79647695 | 89540818 | 79864334 | 76548941 | 81400447 |
| Q30 (%) | 94.08 | 93.93 | 93.62 | 94.09 | 93.93 |
| Paired-end
reads | 78141536 | 87841376 | 78169980 | 75050020 | 79800728 |
| Total mapped
(%) |
71511926 (91.52) | 81141734
(92.37) | 70870280
(90.66) | 69467494
(92.56) | 73247858.5
(91.78) |
| Uniquely mapped
(%) | 69078606
(96.6) | 78068680
(96.21) | 67729842
(95.57) | 67403229
(97.03) | 70570089.3
(96.35) |
| Splice reads
(%) |
30027459 (43.47) | 33845649
(43.35) | 29491479
(43.54) | 28788551
(42.71) | 30538284.5
(43.27) |
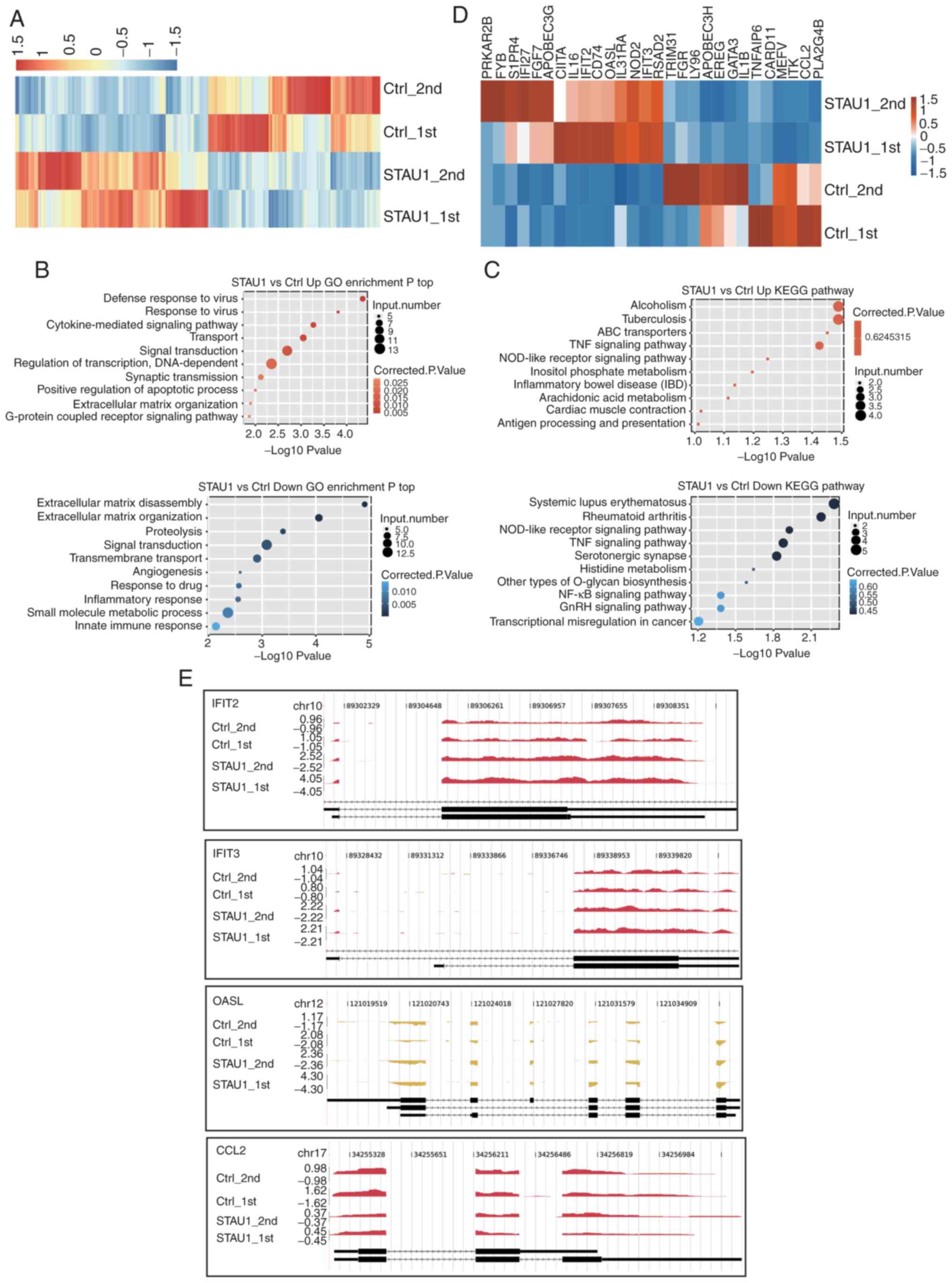
STAU1 regulates the expression of
genes enriched in inflammatory and immune response
A heatmap of the expression levels of DEGs was
plotted. The upregulated genes were represented in the red region
of the experimental group, and downregulated ones in the red region
of the control group. The results revealed a high level of
consistency in STAU1-mediated transcription between the two groups
of data (Fig. 2A). The functions
and potential biological roles of these DEGs were further analyzed.
GO enrichment and KEGG pathway analyses were conducted for the
upregulated and downregulated genes, respectively, and the top 10
GO terms were presented. It is generally believed that a P<0.05
indicates significant difference, i.e., significant enrichment.
Thus, this threshold was defined as the cut-off. GO functions were
divided into three categories, namely, molecular function,
biological process, and cellular component. As revealed in Fig. 2B, genes regulating STAU1
overexpression were primarily enriched in pathways related to
inflammation and immune response. The upregulated genes in the
OE-STAU1 were enriched in ‘defense response to virus’,
‘cytokine-mediated signaling pathways’, ‘transport’, ‘signal
transduction’ and ‘synaptic transmission’; the downregulated genes
were enriched in ‘signal transduction’, ‘transmembrane transport’,
‘inflammatory response’, and ‘innate immune response’. Studies have
confirmed that STAU1 is positively correlated with the function and
number of neuronal synapses, and can induce morphological changes
of the dendritic spine (31–33).
After overexpression of STAU1, upregulated genes associated with
inflammatory and immune responses (IFIT2, OASL, IFIT3, CCL5, and
CD74) were detected (GEO accession number GSE136890).
For KEGG pathway analysis, the cut-off was also set
to P<0.05. The results revealed that the genes were enriched in
a variety of pathways related to the immune system, inflammatory
response, and nervous system (Fig.
2C); the upregulated genes were primarily enriched in ‘ABC
transporters’ and ‘TNF signaling pathway’. The downregulated genes
were mainly enriched in ‘Systemic lupus erythematosus’, ‘Rheumatoid
arthritis’, ‘NOD-like receptor signaling pathway’, ‘TNF signaling
pathway’, ‘Serotonergic synapse’, and ‘NF-κB signaling
pathway’.
Then, among all the DEGs, 28 genes related to
cytokine-mediated signaling, inflammatory response, and immune
response were selected and presented in the DEGs-based hierarchical
clustering plot (Fig. 2D). Among
them, there were 15 upregulated genes and 13 downregulated genes.
Herein, 3 upregulated genes (IFIT2, OASL, and IFIT3) and 1
downregulated gene [chemokine (C-C motif) ligand 2 (CCL2)] were
analyzed in details in terms of coverage and distribution of reads
(Fig. 2E). Distribution of reads
reflected the relative location of genes and the relative read
abundance, which further demonstrated differential expression in
control cells and STAU1 OE cells. Therefore, it could be concluded
that STAU1 selectively regulated genes related to inflammation and
immune responses.
STAU1 positively regulates the
expression of IFIT2, OASL, IFIT3, and negatively regulates the
expression of CCL2 in HeLa cells
The expression levels of the aforementioned 4 genes
are presented in Fig. 3A. These
genes were enriched in the cytokine-mediated signaling pathway,
inflammatory response or other biological processes in the GO
database. To verify the reliability of RNA sequencing, qPCR was
conducted for HeLa cells. It was revealed that IFIT2, OASL, and
IFIT3 were significantly upregulated, while CCL2 was significantly
downregulated (Fig. 3B), which was
consistent with results of RNA-seq analysis. The results of qPCR of
the 3 genes (IFI27, S1PR4, CCL5) in Fig. S1 were also consistent with RNA-seq
analysis (data not shown), and the non-DEG CD44 was the control
gene.
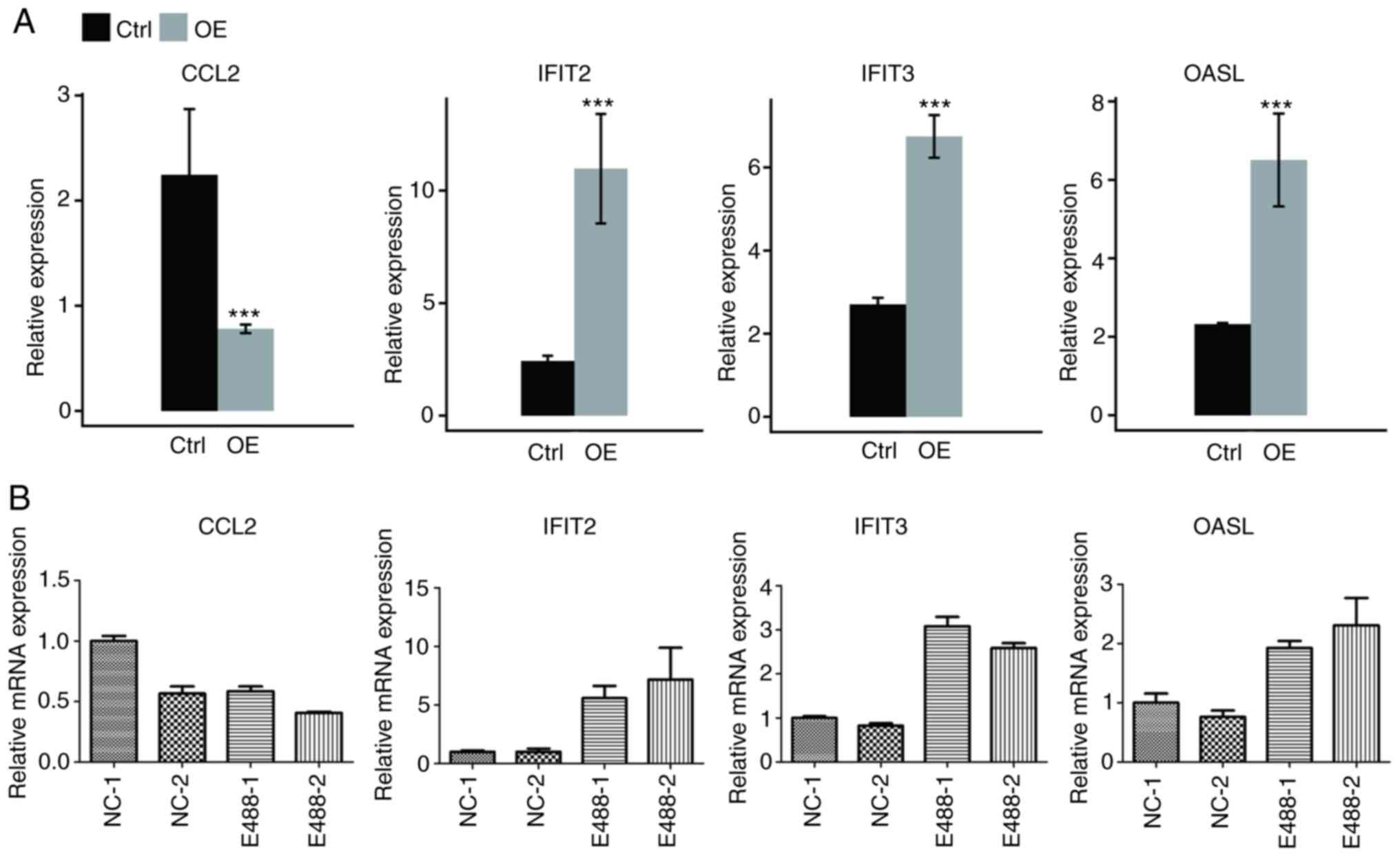 | Figure 3.Validation of the expression level of
DEGs enriched in cytokine-mediated signaling and immune response.
Gene expression quantified by (A) RNA seq data and (B) RT-qPCR.
Error bars represent mean ± SEM. ***P<0.001. FPKM values were
calculated as described in the Materials and methods. DEGs,
differentially expressed genes; RNA seq, RNA sequencing; RT-qPCR,
reverse transcription-quantitative PCR; FPKM, fragments per
kilobase of transcript per million fragments mapped; CCL2,
chemokine (C-C motif) ligand 2; IFIT2, interferon-induced protein
with tetratricopeptide repeats 2; IFIT3, interferon-induced protein
with tetratricopeptide repeats 3; OASL, 2′-5′-oligoadenylate
synthetase-like protein; Ctrl, control; OE, overexpression. |
STAU1 regulates the AS of genes
enriched in the ‘nerve growth factor receptor signaling
pathway’
Regulated AS events (RASEs) of STAU1 in human HeLa
cells were further analyzed. Every sample in the RNA-seq data was
aligned to unique mapped reads on the reference genome for RASE
analysis. The results of exon detection in 4 samples are presented
in Table II. A total of 237,791
detected exons were achieved, accounting for 64.74% of all
annotated exons in the reference genome. Splice junctions of each
sample were then analyzed by using TopHat2 software, and 160,308
known splice junctions (Known_Junction) and 163,225 novel splice
junctions (Novel_Junction) were obtained (Table III).
| Table II.Exon detection results in RNA-seq
data. |
Table II.
Exon detection results in RNA-seq
data.
| Sample | Detected exon | Annotated exon | Ratio (%) |
|---|
| Ctrl_1st | 207169 | 367321 | 56.40 |
| Ctrl_2nd | 204690 | 367321 | 55.73 |
| STAU1_1st | 213908 | 367321 | 58.23 |
| STAU1_2nd | 219650 | 367321 | 59.80 |
| Total | 237791 | 367321 | 64.74 |
| Table III.Splicing junction analysis of samples
from RNA-seq data. |
Table III.
Splicing junction analysis of samples
from RNA-seq data.
| Sample | All_Junction | Novel_Junction | Known_Junction |
|---|
| Ctrl_1st | 195462 | 55232 | 140230 |
| Ctrl_2nd | 186313 | 48162 | 138151 |
| STAU1_1st | 216284 | 71098 | 145186 |
| STAU1_2nd | 234013 | 85203 | 148810 |
| Total | 323533 | 163225 | 160308 |
Various RASEs were statistically analyzed using
ABLas, and the detection results in each sample are presented in
Table IV. There were 76,259
detected RASEs, including 19,746 annotated RASEs in the genome
(known AS) and 56,513 non-annotated novel RASEs (novel AS).
| Table IV.All AS events detected from all
samples. |
Table IV.
All AS events detected from all
samples.
| Sample | KAS (%) | NAS (%) | All AS |
|---|
| Ctrl_1st | 12728 (36.58) | 22063 (63.42) | 34791 |
| Ctrl_2nd | 11916 (36.40) | 20818 (63.60) | 32734 |
| STAU1_1st | 14251 (37.53) | 23717 (62.47) | 37968 |
| STAU1_2nd | 15375 (37.63) | 25480 (62.37) | 40855 |
| Total | 19746 (25.89) | 56513 (74.11) | 76259 |
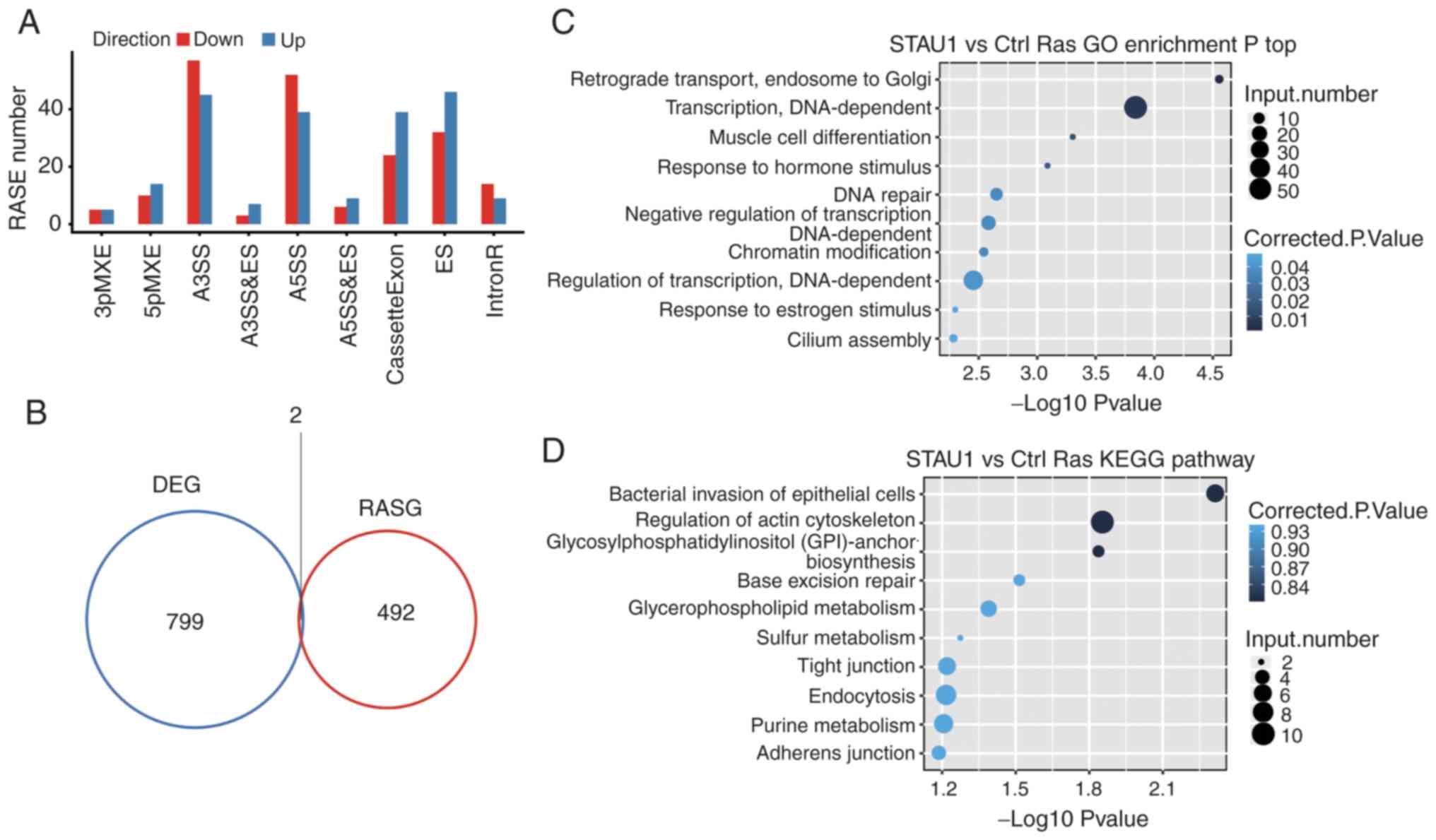
Due to the set-up of biological replicates, a
Student's t-test was used to compare the variation of AS levels of
each gene between two samples, and the criterion for significantly
different AS was set to P≤0.05. As revealed in Fig. 4A, a total of 549 RASEs were
detected, which are presented in Table
V. Among them, the major types of RASEs included 102 A3SS, 91
A5SS, 78 ES, and 63 cassette exons. This indicated that STAU1 had a
retaining and promoting effect on exons in the entire genome.
Number and types of other differential RASEs were as follows: 24
5pMXE, 10 3pMXE, 10 A3SS and ES, 16 MXE, and 15 A5SS and ES. The
aforementioned results indicated that STAU1 could regulate AS in
the genome of HeLa cells.
| Table V.Classification of all RASE events
between sample groups. |
Table V.
Classification of all RASE events
between sample groups.
| Type | STAU1_vs_Ctrl
Up | STAU1_vs_Ctrl
Down |
|---|
| 3pMXE | 5 | 5 |
| 5pMXE | 14 | 10 |
| A3SS | 45 | 57 |
| A3SS and ES | 7 | 3 |
| A5SS | 39 | 52 |
| A5SS and ES | 9 | 6 |
| ES | 46 | 32 |
| IR | 62 | 79 |
| MXE | 9 | 6 |
| Cassette Exon | 39 | 24 |
| Total | 275 | 274 |
An integrated analysis was performed for
differentially regulated alternatively spliced genes (RASGs) and
DEGs in different samples. There were 2 genes with significant
difference in terms of both the expression level and AS level
(Fig. 4B).
Similarly, GO enrichment and KEGG pathway analyses
were undertaken on the differential RASGs, and the top 10 terms are
presented in Fig. 4C and D. GO
terms and KEGG pathways, in which RASGs were enriched are presented
in Table SII and SIII, respectively.
It was revealed that the genes whose AS level was
regulated by STAU1 were mainly enriched in ‘retrograde transport,
endosome to Golgi’, ‘muscle cell differentiation’, and other
reported STAU1-related pathways. The GO term ranking the 16th was
enriched in ‘nerve growth factor receptor signaling pathway’
(~P=0.01).
STAU1-regulated AS of PLEKHG2,
ARHGEF11, NR4A1, PDGFB, FGFR4, RALGDS in HeLa cells
As revealed in Table
SII, 6 key genes were selected for the detection of RASE,
namely, ES, A5SS, 5pMXE, and IR. According to the results of
RNA-seq (Fig. 5 and Fig. S2), the number of reads sequenced
for each gene was over 10. Then, for each gene, the number of reads
for AS between STAU1 OE cells and control cells was compared, and a
significant difference in the AS levels was noted. To verify the
reliability of the results, qPCR was performed in the HeLa cells.
Primers for the qPCR verification are presented in Table SI. RASEs detected by qPCR were
consistent with those by RNA-seq, which demonstrated that STAU1 may
play a significant regulatory role in the AS of ‘nerve growth
factor receptor signaling pathway’.
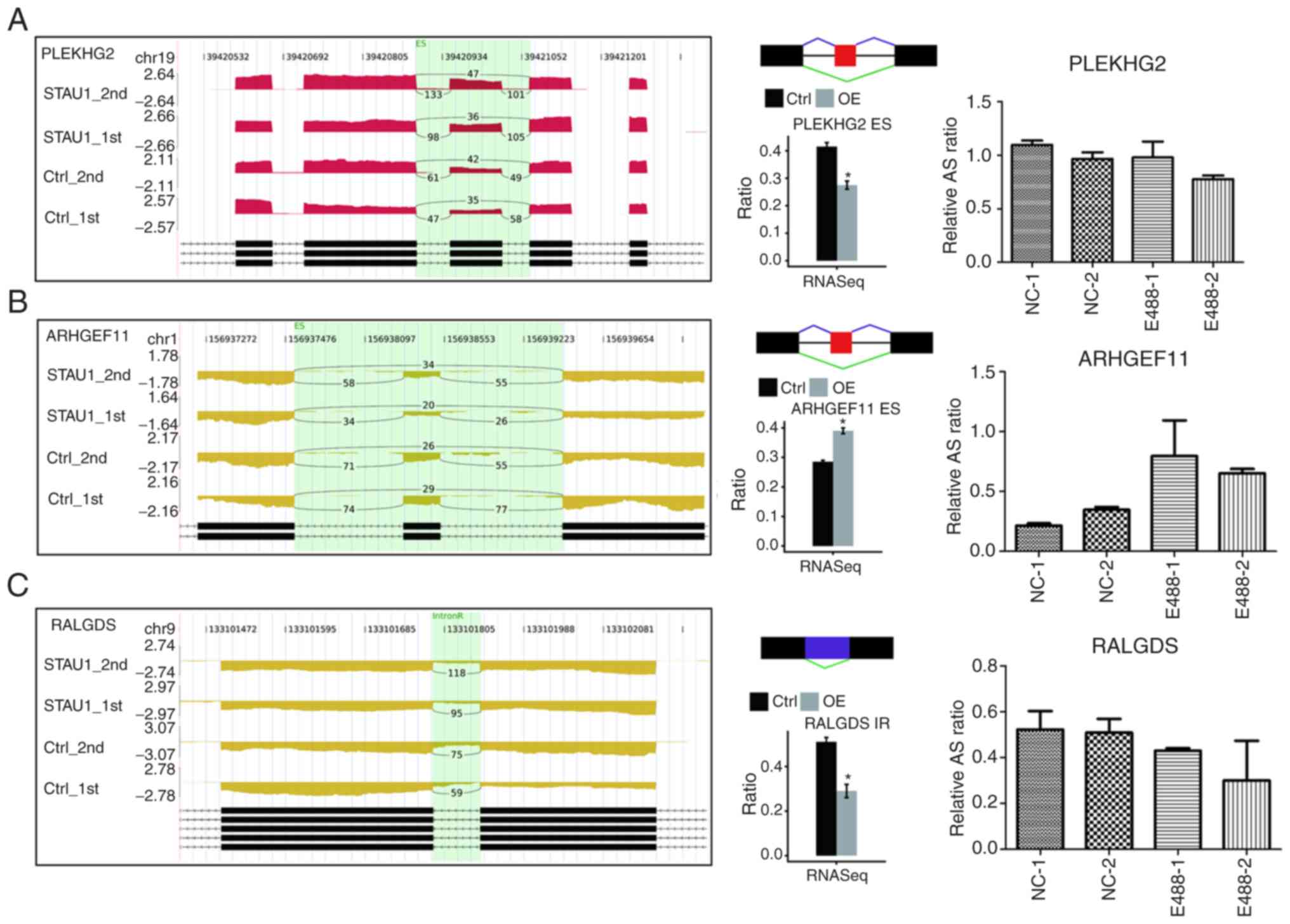 | Figure 5.Validation of STAU1-regulated AS
events. IGV-Sashimi plot revealed two (A and B) two ES and (C) one
IR AS events in three different genes. Reads distribution of each
AS event was plotted in the left panel with the transcripts of each
gene shown below. The schematic diagrams depict the structures of
AS events, AS1 (purple line) and AS2 (green line). The exon
sequences are denoted by black boxes, intron sequences by a
horizontal line (right panel, top), while the retained intron by a
purple box. RNA-seq quantification and RT-qPCR validation of ASEs
are presented in the panels on the right. STAU1, double-stranded
RNA-binding protein Staufen homolog 1; AS, alternative splicing;
ES, exon skipping; IR, intron retention; RNA-seq, RNA sequencing;
RT-qPCR, reverse-transcription quantitative PCR. Error bars
represent mean ± SEM. *P<0.05. |
Discussion
RNA-seq based on high-throughput sequencing is
currently the most widely used transcriptome sequencing technology,
which can promptly extract all the genetic information of the
samples. RNA-seq, has become one of the most representative
high-throughput sequence-based techniques due to its
high-throughput, high accuracy, and cost-effectiveness. It can be
used to study the structure and function of genes, identify changes
in gene expression, and explore AS patterns that are regulated
(34). In the field of life
sciences, this method has been used to explore the pathogenesis of
diseases, clinical diagnosis, and pharmacological research
(35,36).
In the present study it was revealed that
overexpression of STAU1 promoted the proliferation of HeLa cells,
which are useful for the study of gene regulation in the central
nervous system, while the proliferation of neurons and glial cells
in the central nervous system plays an important role in
neuropathic pain (37).
Upregulation of STAU1 caused upregulation or downregulation of
numerous genes, including IFIT2, IFIT3, OASL and CCL2. Through
functional analysis, changes in the expression levels of these
genes may affect signaling pathways, such as ‘defense response to
virus’, ‘cytokine-mediated signaling pathway’, and ‘inflammatory
response’, which are closely associated with inflammatory immune
response. In addition, the AS of multiple genes was also regulated
by STAU1, and the main enriched pathways not only include
‘retrograde transport’ and ‘muscle cell differentiation’, but also
the ‘nerve growth factor receptor signaling pathway’.
Significant upregulation of IFIT2, IFIT3, and OASL
genes was consistently indicated by RNA-seq and qPCR of HeLa cells.
The IFIT family performs multiple functions, including antitumor
effects and regulation of cell apoptosis and innate immune pathways
(38,39). It can also inhibit replication of
flavivirus and coronavirus (40).
Siegfried et al stimulated wild-type bone marrow-derived
macrophages (BMDMs), IFIT2−/−, and IFNAR−/−
BMDMs with LPS, respectively. Results of enzyme-linked
immunosorbent assay (ELISA) test indicated that the mutant BMDMs
exhibited a significant reduction in the expression levels of TNF-α
and interleukin-6 (IL-6) than the wild-type BMDMs. Furthermore,
shRNA interference targeting IFIT2 was performed in RAW264.7
macrophages. It was revealed that TNF-α and IL-6 were also
downregulated, suggesting a pro-inflammatory role of IFIT2
(41). Berchtold et al
demonstrated that overexpression of IFIT2 decreased the secretion
of TNF-α in the RAW264.7 cells (42). Niess et al compared highly
metastatic L3.6pl pancreatic tumor cells and lowly metastatic
COL0357FG pancreatic tumor cells. It was determined that
upregulation of IFIT3 promoted synthesis and secretion of IL-6
(43). Liu et al revealed
that the expression of exogenous IFIT3 enhanced the inducing effect
of NF-κB on TNF-α, without influencing TNF-α-mediated activation of
NF-κB (44). Furthermore, OASL is
an interferon-stimulated gene (ISG), playing a significant role in
the immune response to viruses (45). Activation of OASL can be induced by
interferon (IFN). The expression of OASL can further stimulate the
production of IFN, thereby forming a positive feedback (46). IFN-γ has a neuroprotective effect,
and significantly promotes secretion of IL-6 in astrocytes
(47). Inflammatory cytokines
(TNF-α and IL-6) are important molecules, mediating enhancement of
hyperalgesia via increasing glutamic acid-induced excitatory
current, thereby promoting the development of pain (48). Excitatory synaptic transmission is
mainly regulated by AMPA and NMDA receptors. Inflammatory factors
enhance their degree of excitation, promote the release of
excitatory mediators, such as glutamic acid and substance P, and
participate in the regulation of various pain signaling pathways
(49,50). Therefore, TNF-α and IL-6 play a
vital role in NP.
In the present study it was also revealed that CCL2
was markedly downregulated. CCL2, also known as monocyte
chemotactic protein-1 (MCP-1), can activate monocytes in the
inflammatory state, induce leukocyte migration reaction, regulate
T-cell function, and participate in inflammation and immune
response (51). Recently, it has
been revealed that CCL2 is highly expressed in DRG neurons and
spinal dorsal horn surface neurons during peripheral nerve injury
(52). CCL2 is released in an
activity-dependent manner from the synaptic vesicles in the central
nervous system into the spinal cord (52,53).
The CCL2 expression in the spinal cord is not limited to neurons.
After spinal nerve ligation, astrocytes can also upregulate CCL2.
Additionally, the in vitro cultured astrocytes exhibited an
upregulation of CCL2 by over 100-fold, which was rapidly released
in a JNK-dependent manner (54).
CCL2 secreted by astrocytes acts on CCR2 in the dorsal horn
neurons. CCL2 can strengthen the release of glutamic acid from the
injured neuronal presynaptic membrane and promote the function of
glutamic acid receptors in the postsynaptic membrane. This inhibits
GABA-induced inhibitory synaptic transmission, while causing rapid
phosphorylation of eukaryotic protein kinase (EPK) and activation
of NMDA receptors. As a result, central sensitization is induced in
a direct, rapid and non-transcriptional manner (55). Another study shows that MCP-1 and
its receptor CCR2 in primary sensory neurons are involved in
maintaining paclitaxel induced peripheral neuropathy (56). Therefore, overexpression or
depletion of CCL2 and CCR2 has a direct influence on NP.
The regulatory role of STAU1 overexpression on RASE
of HeLa cells was further studied. A total of 549 significantly
differential RASEs were identified, which verified our speculation
that STAU1 can globally regulate the AS events in the genome in
HeLa cells. Pathways in which the differential RASGs were enriched
have been previously aforementioned. PLEKHG2 and ARHGEF11 are both
Rho guanine nucleotide exchange factors and activators of Rho
GTPases. A variety of biological effects can be regulated by Rho
GTPases, such as transmembrane transport, cell migration, adhesion,
and proliferation (57). Moreover,
Rho GTPases can participate in the immune response by regulating
the Rho/ROCK signaling pathway (58,59).
Another study demonstrated that PLEKHG2/FLJ 00018 can regulate the
morphology of Neuro-2a cells, thereby playing a significant role in
nerve growth and cell proliferation (60). ARHGEF11 is involved in the
regulation of axonal growth by regulating the activity of RhoA
(61). RALGDS is one of the Ras
effectors and functions as a guanine nucleotide exchange factor for
the small G-protein, Ral, which regulates membrane trafficking and
cytoskeletal remodeling (62).
Notably, RALGDS has been revealed to promote neuronal
differentiation (63) and exert a
key regulatory effect on neuronal plasticity and memory formation
(64). RALGDS has also been
revealed to mediate cytoskeletal remodeling (65), promote cell proliferation (66), and facilitate oncogenic
transformation (67). Rondaij et
al revealed that RalGDS overexpression was conducive to promote
the exocytosis of endothelial Weibel-Palade bodies (WPBs) (68). The proteins encoded by the FGFR1 and
FGFR4 genes are all members of the fibroblast growth factor
receptor (FGFR) family (69). They
trigger the downstream cascade by binding to FGFRs, thereby playing
a substantial role in promoting embryonic growth and development
(69), development of the nervous
system (70), and regulating the
metabolism (71). Other biological
functions of the proteins are manifested in promoting injury repair
(72), bone formation (73), and vascular and neural regeneration
(74,75). FGFR overexpression has also been
revealed to be associated with tumor and bone diseases, as well as
arthritis (76–78). All of the aforementioned genes were
subjected to AS analysis, and the results were validated by qPCR,
which indicated consistency with RNA-seq except for the qPCR result
for the PDGFB gene that was inconsistent with that of RNA-seq.
It is already known that PDGFB plays a significant
role in the growth and proliferation of vessels and nerves
(79). Herein, we further discussed
the AS events induced by PDGFB and the resultant alterations in
gene functions as an example. Platelet-derived growth factor (PDGF)
is an important factor promoting cell growth. It consists of five
homotypic or heterotypic dimerized ligands (PDGF-AA, -AB, -BB, -CC,
-CD), which are formed by polypeptide chains (PDGF-A, PDGF-B,
PDGF-C, PDGF-D), encoded by four different genes, via the disulfide
bonds (80). Both of its receptors
PDGFR-α and PDGFR-β belong to the receptor tyrosine kinase (RTK)
family (81). It has been
demonstrated that PDGF can stimulate the division growth of
fibroblasts (82), neuroglial cells
(83) and smooth muscle cells
(84). In particular, PDGF-BB has
been revealed to promote neuronal development and differentiation
(85), and to play a neurotrophic
role as well (86,87). A number of scholars have
demonstrated that PDGF-BB can regulate neuronal proliferation and
differentiation by activating the PI3K/Akt and ERK pathways
(88), restore the proliferation
and differentiation of damaged neural precursor cells, and reverse
neuronal excitotoxicity. PDGFB has been demonstrated to play an
important role in neuropathic pain (89,90),
and in the present study it was confirmed that STAUI regulated the
alternative splicing of genes enriched in the ‘nerve growth factor
receptor signaling pathway’ including PDGFB, and thus is also
associated with neuropathic pain.
The present study confirmed that the PDGFB gene
undergoes A5SS events. The normal secretion of PDGF-B protein into
the extracellular domain to bind to the PDGFR receptor-α and -β
subunits and fulfill the biological effect is consequently
affected. The qPCR results were consistent with our theory that
STAU1 promotes the retention of the PDGFB signal peptide, which
mediates the neuroprotective mechanism and relieves neuropathic
pain. STAU1 promotes the retention of PDGFB signal peptide, which
mediates the neuroprotective mechanism and relieves the neuropathic
pain. We surmised that an even more complex regulatory mechanism is
herein involved, while further studies should be conducted to
confirm our findings.
The present study revealed that overexpression of
STAU1 had a regulatory effect on gene splicing and transcription in
HeLa cells. STAU1 could positively regulate the transcription of
genes related to inflammation and immune response. This regulatory
effect also influenced the expression levels of pro-inflammatory
factors and chemotactic factors. Moreover, AS of genes enriched in
the ‘nerve growth factor receptor signaling pathway’ as well as
‘retrograde transport, endosome to Golgi’, and ‘muscle cell
differentiation’ was regulated by STAU1. A recent study
demonstrated that several RNA binding factors involved in local
translation may play a crucial role in pain, including STAU1, a
double-stranded dsRNA binding protein, which is expressed in
peripheral sensory neurons and may play a role in axonal mRNA
transport (91). Therefore, it can
be concluded that STAU1 may be a novel potential therapeutic target
for NP.
Supplementary Material
Supporting Data
Acknowledgements
We deeply thank Dr Wen Chen for his significant
technical and scientific comments.
Funding
The present study was financially supported by the
Natural Science Foundation of Hubei Province, China (grant no.
2011CDC032). This study was partially supported by ABLife
(experimental project no. ABL-7702121).
Availability of data and materials
The data discussed in this publication are available
under GEO Series accession number GSE136890.
Authors' contributions
YZ performed the experiments and data analysis and
was the main author of the manuscript. ZH participated in the
experimental design, carried out bioinformatics analysis, and
partly contributed to the writing of the manuscript. JW and FD
conducted the cell experiments and interpretation of the data. FL
and YX designed the project, contributed to the analysis and
interpretation of the data, and revision of the manuscript. All the
authors read and approved the submitted final version of the
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Castello A, Fischer B, Hentze MW and
Preiss T: RNA-binding proteins in mendelian disease. Trends Genet.
29:318–327. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lunde BM, Moore C and Varani G:
RNA-binding proteins: Modular design for efficient function. Nat
Rev Mol Cell Biol. 8:479–490. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thandapani P, O'Connor TR, Bailey TL and
Richard S: Defining the RGG/RG motif. Mol Cell. 50:613–623. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hentze MW, Castello A, Schwarzl T and
Preiss T: A brave new world of RNA-binding proteins. Nat Rev Mol
Cell Biol. 19:327–341. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Furic L, Maher-Laporte M and
DesGroseillers L: A genome-wide approach identifies distinct but
overlapping subsets of cellular mRNAs associated with staufen1- and
staufen2-containing ribonucleoprotein complexes. RNA. 14:324–335.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bondy-Chorney E, Crawford Parks TE,
Ravel-Chapuis A, Klinck R, Rocheleau L, Pelchat M, Chabot B, Jasmin
BJ and Côté J: Staufen1 regulates multiple alternative splicing
events either positively or negatively in DM1 Indicating its role
as a disease modifier. PLoS Genet. 12:e10058272016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gong C and Maquat LE: lncRNAs
transactivate STAU1-mediated mRNA decay by duplexing with 3′UTRs
via Alu elements. Nature. 470:284–288. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kretz M, Siprashvili Z, Chu C, Webster DE,
Zehnder A, Qu K, Lee CS, Flockhart RJ, Groff AF, Chow J, et al:
Control of somatic tissue differentiation by the long non-coding
RNA TINCR. Nature. 493:231–235. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kretz M: TINCR, staufen1, and cellular
differentiation. RNA Biol. 10:1597–1601. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Elbarbary RA, Li W, Tian B and Maquat LE:
STAU1 binding 3′ UTR IRAlus complements nuclear retention to
protect cells from PKR-mediated translational shutdown. Genes Dev.
27:1495–1510. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Giorgi C, Yeo GW, Stone ME, Katz DB, Burge
C, Turrigiano G and Moore MJ: The EJC factor eIF4AIII modulates
synaptic strength and neuronal protein expression. Cell.
130:179–191. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Oh Y, Park J, Kim JI, Chang MY, Lee SH,
Cho YH and Hwang J: Lin28B and miR-142-3p regulate neuronal
differentiation by modulating staufen1 expression. Cell Death
Differ. 25:432–443. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
de Lucas S, Peredo J, Marion RM, Sanchez C
and Ortin J: Human staufen1 protein interacts with influenza virus
ribonucleoproteins and is required for efficient virus
multiplication. J Virol. 84:7603–7612. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lee JH, Oh JY, Pascua PN, Kim EG, Choi YK
and Kim HK: Impairment of the staufen1-NS1 interaction reduces
influenza viral replication. Biochem Biophys Res Commun.
414:153–158. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Rao S, Hassine S, Monette A, Amorim R,
DesGroseillers L and Mouland AJ: HIV-1 requires staufen1 to
dissociate stress granules and to produce infectious viral
particles. RNA. 25:727–736. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xue Y, Ouyang K, Huang J, Zhou Y, Ouyang
H, Li H, Wang G, Wu Q, Wei C, Bi Y, et al: Direct conversion of
fibroblasts to neurons by reprogramming PTB-regulated microRNA
circuits. Cell. 152:82–96. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gentry JJ, Casaccia-Bonnefil P and Carter
BD: Nerve growth factor activation of nuclear factor kappaB through
its p75 receptor is an anti-apoptotic signal in RN22 schwannoma
cells. J Biol Chem. 275:7558–7565. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nykjaer A, Lee R, Teng KK, Jansen P,
Madsen P, Nielsen MS, Jacobsen C, Kliemannel M, Schwarz E, Willnow
TE, et al: Sortilin is essential for proNGF-induced neuronal cell
death. Nature. 427:843–848. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rabizadeh S, Rabizadeh S, Ye X, Wang JJ
and Bredesen DE: Neurotrophin dependence mediated by p75NTR:
Contrast between rescue by BDNF and NGF. Cell Death Differ.
6:1222–1227. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Notterpek L: Neurotrophins in myelination:
A new role for a puzzling receptor. Trends Neurosci. 26:232–234.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fu C, Donovan WP, Shikapwashya-Hasser O,
Ye X and Cole RH: Hot fusion: An efficient method to clone multiple
DNA fragments as well as inverted repeats without ligase. PLoS One.
9:e1153182014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
McGinn S and Gut IG: DNA
sequencing-spanning the generations. N Biotechnol. 30:366–372.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim D, Pertea G, Trapnell C, Pimentel H,
Kelley R and Salzberg SL: TopHat2: Accurate alignment of
transcriptomes in the presence of insertions, deletions and gene
fusions. Genome Biol. 14:R362013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Trapnell C, Williams BA, Pertea G,
Mortazavi A, Kwan G, van Baren MJ, Salzberg SL, Wold BJ and Pachter
L: Transcript assembly and quantification by RNA-Seq reveals
unannotated transcripts and isoform switching during cell
differentiation. Nat Biotechnol. 28:511–515. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Robinson MD, McCarthy DJ and Smyth GK:
EdgeR: A bioconductor package for differential expression analysis
of digital gene expression data. Bioinformatics. 26:139–140. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jin L, Li G, Yu D, Huang W, Cheng C, Liao
S, Wu Q and Zhang Y: Transcriptome analysis reveals the complexity
of alternative splicing regulation in the fungus verticillium
dahliae. BMC Genomics. 18:1302017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xia H, Chen D, Wu Q, Wu G, Zhou Y, Zhang Y
and Zhang L: CELF1 preferentially binds to exon-intron boundary and
regulates alternative splicing in HeLa cells. Biochim Biophys Acta
Gene Regul Mech. 1860:911–921. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xie C, Mao X, Huang J, Ding Y, Wu J, Dong
S, Kong L, Gao G, Li CY and Wei L: KOBAS 2.0: A web server for
annotation and identification of enriched pathways and diseases.
Nucleic Acids Res. 39((Web Server issue)): W316–W322. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Erlich Y, Mitra PP, delaBastide M,
McCombie WR and Hannon GJ: Alta-cyclic: A self-optimizing base
caller for next-generation sequencing. Nat Methods. 5:679–682.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lebeau G, DesGroseillers L, Sossin W and
Lacaille JC: mRNA binding protein staufen 1-dependent regulation of
pyramidal cell spine morphology via NMDA receptor-mediated synaptic
plasticity. Mol Brain. 4:222011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lebeau G, Maher-Laporte M, Topolnik L,
Laurent CE, Sossin W, Desgroseillers L and Lacaille JC: Staufen1
regulation of protein synthesis-dependent long-term potentiation
and synaptic function in hippocampal pyramidal cells. Mol Cell
Biol. 28:2896–2907. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vessey JP, Macchi P, Stein JM, Mikl M,
Hawker KN, Vogelsang P, Wieczorek K, Vendra G, Riefler J, Tübing F,
et al: A loss of function allele for murine staufen1 leads to
impairment of dendritic staufen1-RNP delivery and dendritic spine
morphogenesis. Proc Natl Acad Sci USA. 105:16374–16379. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Z, Gerstein M and Snyder M: RNA-Seq:
A revolutionary tool for transcriptomics. Nat Rev Genet. 10:57–63.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Underwood JG, Uzilov AV, Katzman S,
Onodera CS, Mainzer JE, Mathews DH, Lowe TM, Salama SR and Haussler
D: FragSeq: Transcriptome-wide RNA structure probing using
high-throughput sequencing. Nat Methods. 7:995–1001. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Palanisamy N, Ateeq B, Kalyana-Sundaram S,
Pflueger D, Ramnarayanan K, Shankar S, Han B, Cao Q, Cao X, Suleman
K, et al: Rearrangements of the RAF kinase pathway in prostate
cancer, gastric cancer and melanoma. Nat Med. 16:793–798. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chu CT, Ji J, Dagda RK, Jiang JF, Tyurina
YY, Kapralov AA, Tyurin VA, Yanamala N, Shrivastava IH,
Mohammadyani D, et al: Cardiolipin externalization to the outer
mitochondrial membrane acts as an elimination signal for mitophagy
in neuronal cells. Nat Cell Biol. 15:1197–1205. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fensterl V, Wetzel JL, Ramachandran S,
Ogino T, Stohlman SA, Bergmann CC, Diamond MS, Virgin HW and Sen
GC: Interferon-induced Ifit2/ISG54 protects mice from lethal VSV
neuropathogenesis. PLoS Pathog. 8:e10027122012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Stawowczyk M, Van Scoy S, Kumar KP and
Reich NC: The interferon stimulated gene 54 promotes apoptosis. J
Biol Chem. 286:7257–7266. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang B, Liu X, Chen W and Chen L: IFIT5
potentiates anti-viral response through enhancing innate immune
signaling pathways. Acta Biochim Biophys Sin (Shanghai).
45:867–874. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Siegfried A, Berchtold S, Manncke B,
Deuschle E, Reber J, Ott T, Weber M, Kalinke U, Hofer MJ, Hatesuer
B, et al: IFIT2 is an effector protein of type I IFN-mediated
amplification of lipopolysaccharide (LPS)-induced TNF-α secretion
and LPS-induced endotoxin shock. J Immunol. 191:3913–3921. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Berchtold S, Manncke B, Klenk J, Geisel J,
Autenrieth IB and Bohn E: Forced IFIT-2 expression represses LPS
induced TNF-alpha expression at posttranscriptional levels. BMC
Immunol. 9:752008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Niess H, Camaj P, Mair R, Renner A, Zhao
Y, Jäckel C, Nelson PJ, Jauch KW and Bruns CJ: Overexpression of
IFN-induced protein with tetratricopeptide repeats 3 (IFIT3) in
pancreatic cancer: Cellular ‘pseudoinflammation’ contributing to an
aggressive phenotype. Oncotarget. 6:3306–3318. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu XY, Chen W, Wei B, Shan YF and Wang C:
IFN-induced TPR protein IFIT3 potentiates antiviral signaling by
bridging MAVS and TBK1. J Immunol. 187:2559–2568. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Melchjorsen J, Kristiansen H, Christiansen
R, Rintahaka J, Matikainen S, Paludan SR and Hartmann R:
Differential regulation of the OASL and OAS1 genes in response to
viral infections. J Interferon Cytokine Res. 29:199–207. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhu J, Ghosh A and Sarkar SN: OASL-a new
player in controlling antiviral innate immunity. Curr Opin Virol.
12:15–19. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sun L, Li Y, Jia X, Wang Q, Li Y, Hu M,
Tian L, Yang J, Xing W, Zhang W, et al: Neuroprotection by IFN-ү
via astrocyte-secreted IL-6 in acute neuroinflammation. Oncotarget.
8:40065–40078. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ellis A and Bennett DL: Neuroinflammation
and the generation of neuropathic pain. Br J Anaesth. 111:26–37.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kawasaki Y, Zhang L, Cheng JK and Ji RR:
Cytokine mechanisms of central sensitization: Distinct and
overlapping role of interleukin-1beta, interleukin-6, and tumor
necrosis factor-alpha in regulating synaptic and neuronal activity
in the superficial spinal cord. J Neurosci. 28:5189–5194. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Leung L and Cahill CM: TNF-alpha and
neuropathic pain-a review. J Neuroinflammation. 7:272010.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Deshmane SL, Kremlev S, Amini S and Sawaya
BE: Monocyte chemoattractant protein-1 (MCP-1): An overview. J
Interferon Cytokine Res. 29:313–326. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Dansereau MA, Gosselin RD, Pohl M, Pommier
B, Mechighel P, Mauborgne A, Rostene W, Kitabgi P, Beaudet N,
Sarret P and Melik-Parsadaniantz S: Spinal CCL2 pronociceptive
action is no longer effective in CCR2 receptor antagonist-treated
rats. J Neurochem. 106:757–769. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Thacker MA, Clark AK, Bishop T, Grist J,
Yip PK, Moon LD, Thompson SW, Marchand F and McMahon SB: CCL2 is a
key mediator of microglia activation in neuropathic pain states.
Eur J Pain. 13:263–272. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Gao YJ, Zhang L, Samad OA, Suter MR,
Yasuhiko K, Xu ZZ, Park JY, Lind AL, Ma Q and Ji RR: JNK-induced
MCP-1 production in spinal cord astrocytes contributes to central
sensitization and neuropathic pain. J Neurosci. 29:4096–4108. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Zhuang ZY, Kawasaki Y, Tan PH, Wen YR,
Huang J and Ji RR: Role of the CX3CR1/p38 MAPK pathway in spinal
microglia for the development of neuropathic pain following nerve
injury-induced cleavage of fractalkine. Brain Behav Immun.
21:642–651. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zhang H, Boyette-Davis JA, Kosturakis AK,
Li Y, Yoon SY, Walters ET and Dougherty PM: Induction of monocyte
chemoattractant protein-1 (MCP-1) and its receptor CCR2 in primary
sensory neurons contributes to paclitaxel-induced peripheral
neuropathy. J Pain. 14:1031–1044. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Rougerie P and Delon J: Rho GTPases:
Masters of T lymphocyte migration and activation. Immunol Lett.
142:1–13. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Sato K, Sugiyama T, Nagase T, Kitade Y and
Ueda H: Threonine 680 phosphorylation of FLJ00018/PLEKHG2, a Rho
family-specific guanine nucleotide exchange factor, by epidermal
growth factor receptor signaling regulates cell morphology of
Neuro-2a cells. J Biol Chem. 289:10045–10056. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Shimizu Y, Dobashi K, Iizuka K, Horie T,
Suzuki K, Tukagoshi H, Nakazawa T, Nakazato Y and Mori M:
Contribution of small GTPase Rho and its target protein rock in a
murine model of lung fibrosis. Am J Respir Crit Care Med.
163:210–217. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Sato K, Kimura M, Sugiyama K, Nishikawa M,
Okano Y, Nagaoka H, Nagase T, Kitade Y and Ueda H: Four-and-a-half
LIM domains 1 (FHL1) protein interacts with the rho guanine
nucleotide exchange factor PLEKHG2/FLJ00018 and regulates cell
morphogenesis. J Biol Chem. 291:25227–25238. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Sun X, Zhou Z, Fink DJ and Mata M: HspB1
silences translation of PDZ-RhoGEF by enhancing miR-20a and miR-128
expression to promote neurite extension. Mol Cell Neurosci.
57:111–119. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Hofer F, Fields S, Schneider C and Martin
GS: Activated ras interacts with the ral guanine nucleotide
dissociation stimulator. Proc Natl Acad Sci USA. 91:11089–11093.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Boriack-Sjodin PA, Margarit SM, Bar-Sagi D
and Kuriyan J: The structural basis of the activation of ras by
sos. Nature. 394:337–343. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
64
|
Mitra S, Cheng KW and Mills GB: Rab
GTPases implicated in inherited and acquired disorders. Semin Cell
Dev Biol. 22:57–68. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Bhattacharya M, Anborgh PH, Babwah AV,
Dale LB, Dobransky T, Benovic JL, Feldman RD, Verdi JM, Rylett RJ
and Ferguson SS: Beta-arrestins regulate a Ral-GDS Ral effector
pathway that mediates cytoskeletal reorganization. Nat Cell Biol.
4:547–555. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
66
|
Hao Y, Wong R and Feig LA: RalGDS couples
growth factor signaling to Akt activation. Mol Cell Biol.
28:2851–2859. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Yin J, Pollock C, Tracy K, Chock M, Martin
P, Oberst M and Kelly K: Activation of the RalGEF/Ral pathway
promotes prostate cancer metastasis to bone. Mol Cell Biol.
27:7538–7550. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Rondaij MG, Bierings R, van Agtmaal EL,
Gijzen KA, Sellink E, Kragt A, Ferguson SS, Mertens K, Hannah MJ,
van Mourik JA, et al: Guanine exchange factor RalGDS mediates
exocytosis of weibel-palade bodies from endothelial cells. Blood.
112:56–63. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Anteby EY, Natanson-Yaron S, Hamani Y,
Sciaki Y, Goldman-Wohl D, Greenfield C, Ariel I and Yagel S:
Fibroblast growth factor-10 and fibroblast growth factor receptors
1–4: Expression and peptide localization in human decidua and
placenta. Eur J Obstet Gynecol Reprod Biol. 119:27–35. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Kang K and Song MR: Diverse FGF receptor
signaling controls astrocyte specification and proliferation.
Biochem Biophys Res Commun. 395:324–329. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Goetz R and Mohammadi M: Exploring
mechanisms of FGF signalling through the lens of structural
biology. Nat Rev Mol Cell Biol. 14:166–180. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Jaye M, Schlessinger J and Dionne CA:
Fibroblast growth factor receptor tyrosine kinases: Molecular
analysis and signal transduction. Biochim Biophys Acta.
1135:185–199. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Barnes GL, Kostenuik PJ, Gerstenfeld LC
and Einhorn TA: Growth factor regulation of fracture repair. J Bone
Miner Res. 14:1805–1815. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Grothe C and Nikkhah G: The role of basic
fibroblast growth factor in peripheral nerve regeneration. Anat
Embryol (Berl). 204:171–177. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Pringle NP, Yu WP, Howell M, Colvin JS,
Ornitz DM and Richardson WD: Fgfr3 expression by astrocytes and
their precursors: Evidence that astrocytes and oligodendrocytes
originate in distinct neuroepithelial domains. Development.
130:93–102. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Beenken A and Mohammadi M: The FGF family:
Biology, pathophysiology and therapy. Nat Rev Drug Discov.
8:235–253. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Bono F, De Smet F, Herbert C, De Bock K,
Georgiadou M, Fons P, Tjwa M, Alcouffe C, Ny A, Bianciotto M, et
al: Inhibition of tumor angiogenesis and growth by a small-molecule
multi-FGF receptor blocker with allosteric properties. Cancer Cell.
23:477–488. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Murphy PR and Knee RS: Basic fibroblast
growth factor binding and processing by human glioma cells. Mol
Cell Endocrinol. 114:193–203. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Chang F, Steelman LS, Lee JT, Shelton JG,
Navolanic PM, Blalock WL, Franklin RA and McCubrey JA: Signal
transduction mediated by the Ras/Raf/MEK/ERK pathway from cytokine
receptors to transcription factors: Potential targeting for
therapeutic intervention. Leukemia. 17:1263–1293. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Fredriksson L, Li H and Eriksson U: The
PDGF family: Four gene products form five dimeric isoforms.
Cytokine Growth Factor Rev. 15:197–204. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Reigstad LJ, Varhaug JE and Lillehaug JR:
Structural and functional specificities of PDGF-C and PDGF-D, the
novel members of the platelet-derived growth factors family. FEBS
J. 272:5723–5741. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Kohler N and Lipton A: Platelets as a
source of fibroblast growth-promoting activity. Exp Cell Res.
87:297–301. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Westermark B and Wasteson A: A platelet
factor stimulating human normal glial cells. Exp Cell Res.
98:170–174. 1976. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Ross R, Glomset J, Kariya B and Harker L:
A platelet-dependent serum factor that stimulates the proliferation
of arterial smooth muscle cells in vitro. Proc Natl Acad Sci USA.
71:1207–1210. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Williams BP, Park JK, Alberta JA,
Muhlebach SG, Hwang GY, Roberts TM and Stiles CD: A PDGF-regulated
immediate early gene response initiates neuronal differentiation in
ventricular zone progenitor cells. Neuron. 18:553–562. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Pietz K, Odin P, Funa K and Lindvall O:
Protective effect of platelet-derived growth factor against
6-hydroxydopamine-induced lesion of rat dopaminergic neurons in
culture. Neurosci Lett. 204:101–104. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Smits A, Kato M, Westermark B, Nister M,
Heldin CH and Funa K: Neurotrophic activity of platelet-derived
growth factor (PDGF): Rat neuronal cells possess functional PDGF
beta-type receptors and respond to PDGF. Proc Natl Acad Sci USA.
88:8159–8163. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Erlandsson A, Enarsson M and
Forsberg-Nilsson K: Immature neurons from CNS stem cells
proliferate in response to platelet-derived growth factor. J
Neurosci. 21:3483–3491. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Peng F, Dhillon N, Callen S, Yao H,
Bokhari S, Zhu X, Baydoun HH and Buch S: Platelet-derived growth
factor protects neurons against gp120-mediated toxicity. J
Neurovirol. 14:62–72. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Yao H, Duan M, Yang L and Buch S:
Platelet-derived growth factor-BB restores human immunodeficiency
virus Tat-cocaine-mediated impairment of neurogenesis: Role of
TRPC1 channels. J Neurosci. 32:9835–9847. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
de la Pena JB and Campbell ZT: RNA-binding
proteins as targets for pain therapeutics. Neurobiol Pain. 4:2–7.
2018. View Article : Google Scholar : PubMed/NCBI
|