Introduction
The most frequent malignant tumor of the head and
neck region is squamous cell carcinoma, which typically develops in
males (1,2). Squamous cell carcinoma is strongly
associated with certain environmental and lifestyle risk factors,
including tobacco smoking, alcohol consumption, UV light exposure
and infection with high-risk types of human papillomavirus
(3). In addition, betel chewing in
Southeast Asia is known to be a strong risk factor for developing
oral cavity cancers (4). Squamous
cell carcinoma can be treated through surgery, radiation therapy
and chemotherapy, as well as new investigative treatments, such as
immunotherapy and gene therapy (5).
Oral cancer has been in the top ten common cancers for decades in
Taiwan, with a continuing annual increase in its incidence rate
(6).
Arsenic is a naturally occurring element distributed
throughout the earth's crust, which exhibits both metallic and
non-metallic properties (7). In the
environment, arsenic is combined with oxygen, chlorine and sulfur
to form inorganic arsenic compounds; while in animals and plants it
combines with carbon and hydrogen to form organic arsenic
compounds. In general, inorganic arsenic is more toxic than organic
arsenic (8). Arsenic has long been
applied as a pesticide for agricultural use due to its poisonous
effects. In addition, arsenic trioxide (ATO) has been discovered to
exert antitumor effects in patients with acute promyelocytic
leukemia (APL) for clinical trial (9). Several studies have reported that ATO
induces apoptosis of malignant cells, including APL (10), multiple myeloma (11), lung cancer (12), liver cancer (13), cholangiocarcinoma (14), testicular cancer (15) and ovarian cancer (16) cells with in vitro and in
vivo studies. Another inorganic arsenic compound, arsenic
hexoxide, has also been reported to exert anticancer effects on
MCF-7 human breast cancer cells (17) and colon cancer (18). Furthermore, an organic arsenic
derivative (OAD), darinaparsin, is potentially safer and more
effective for the treatment of hematological and solid tumors
according to preclinical models (19). Dipropil-S-glycerol arsenic, a novel
OAD, also exhibits antiproliferative activity by inducing apoptosis
of human acute myeloid leukemia cell lines (20). Therefore, inorganic and organic
arsenic compounds possess the ability to suppress tumor progression
in vitro and in vivo. To the best of our knowledge,
no study has yet determined whether arsenic compounds exert any
effects on normal gingival fibroblasts or any anticancer effects on
oral tumors.
The unifying characteristics of apoptosis are
largely morphological, including cell shrinkage, blebbing of the
plasma membrane, chromatin condensation and DNA fragmentation
(21,22). Apoptosis can be distinguished into
the extrinsic and intrinsic pathways based on the initial stimuli
and regulatory caspase (aspartate-specific cysteine protease)
cascade. The extrinsic pathway, also termed the death receptor
pathway, is activated by death receptors when bound by the
appropriate ligand. Upon binding, a trimerized receptor recruits
the signal transducing molecules through interaction with death
domains, which leads to the cleavage of pro-caspase-8. Activated
caspase-8 initiates a protease cascade that cleaves targets within
a cell and results in apoptotic cell death (23). Conversely, the intrinsic pathway is
dependent on the decrease of mitochondrial membrane potential. When
cellular stress occurs, cytochrome c is released from the
mitochondrial intermembrane space into the cytosol. Subsequently,
it binds to an adapter protein, apoptotic protease activating
factor 1, which acts to recruit pro-caspase-9. Active caspase-9
subsequently cleaves pro-caspase-3, which is then released into the
cytosol to affect the proteolytic degradation of its target
substrates (23). Both apoptotic
pathways can activate downstream effector caspases and lead to
poly(ADP-ribose) polymerase (PARP) cleavage, which inhibits the
cellular function of DNA repair (24,25).
In fact, previous studies have demonstrated that ATO can induce
apoptosis with downregulation of Bcl-2 expression and activation of
a caspase cascade (24,25).
Mitogen-activated protein kinases (MAPKs), which are
crucial for the maintenance of cell proliferation, differentiation,
mitosis, cell survival, gene expression and apoptosis, consist of
three family members: c-Jun NH2-terminal kinase [(JNK)1, 2 and 3];
extracellular signal-regulated kinase (ERK1 and 2); and p38 MAPKs
(p38-MAPK α, β, γ and δ) (26,27).
MAPKs are serine/threonine/tyrosine-specific protein kinases
involved in directing cellular responses to diverse stimuli
(26,27). Depending on the cell type and
stimulus, MAPKs promote cell survival or trigger cell death. JNK
and ERK activities have been reported to sensitize ovarian
carcinoma cells to cisplatin-induced apoptosis (28). In addition, phosphorylation of JNK
and p38 is increased in response to ATO treatment, resulting in
mitochondrial apoptotic cell death in human cervical cancer cells
(29).
On account of the anticancer ability of arsenic
compounds and emerging demands for more effective therapeutic
remedies to treat oral cavity cancer, OEC-M1 cells, derived from a
surgical specimen of buccal mucosa squamous carcinoma from a
Taiwanese as a unique indigenous oral cancer cell line (30,31),
were used in the present study. Cells treated with sodium arsenite
(NaAsO2) and dimethylarsenic acid
[(CH3)2AsO2H; DMA] were
investigated to determine whether the arsenic compounds could
affect cell viability, regulate cell cycle progression, modulate
signaling pathways, and then induce apoptosis with anticancer
capabilities.
Materials and methods
Chemicals
NaAsO2 was purchased from Fluka;
Sigma-Aldrich. DMA, high glucose Dulbecco's modified Eagle's
medium, penicillin-streptomycin, staurosporine, RNase A, propidium
iodide, 30% acrylamide/Bis-acrylmide solution, methylthiazole
tetrazolium (MTT) and monoclonal antibody against β-actin were
purchased from Sigma-Aldrich; Merck KGaA. Fetal bovine serum and
trypsin-EDTA were purchased from AG Scientific, Inc. RPMI-1640
medium, Dulbecco's modified Eagle's medium/F12 and Keratinocyte-SFM
medium were purchased from Gibco; Thermo Fisher Scientific, Inc.
Sodium chloride, potassium chloride,
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) and Tris
base were purchased from JT Baker. Disodium hydrogen phosphate
(Na2HPO4), potassium dihydrogen phosphate
(KH2PO4) and sodium bicarbonate
(NaHCO3) were purchased from Riedel-de Haen.
Hydrochloric acid, sodium hydroxide, sodium dodecyl sulfate (SDS),
Tween-20 and dimethyl sulfoxide (DMSO) were purchased from Merck
KGaA. Donkey anti-rabbit IgG conjugated to horseradish peroxidase
and donkey anti-mouse IgG conjugated to horseradish peroxidase were
purchased from PerkinElmer, Inc. Annexin V-FITC apoptosis detection
kit was purchased from Strong Biotech. Micro BCA protein assay kit
was purchased from Thermo Fisher Scientific, Inc. Antibodies
against PARP, cleaved caspase-8, cleaved caspase-9, cleaved
caspase-3, phosphorylated (p)-JNK, JNK, p-ERK1/2, ERK1/2, p-p38 and
p38 were purchased from Cell Signaling Technology, Inc. Enhanced
chemiluminescence (ECL) detection kit was purchased from EMD
Millipore.
Cell culture
OEC-M1 is a cell line derived from a surgical
specimen of buccal mucosa squamous carcinoma from a Taiwanese, a
unique oral cancer indigenous in Taiwan, and was a generous gift
from Professor Kuo-Wei Chang (National Yang-Ming University,
Taipei, Taiwan) (30,31). OEC-M1 cells were maintained in
RPMI-1640 medium supplemented with 24 mM NaHCO3, 25 mM
HEPES, 10,000 U penicillin and 10,000 µg streptomycin (both from
Sigma-Aldrich; Merck KGaA) and 10% heat-inactivated fetal bovine
serum, pH 7.4, incubated in a humidified atmosphere containing 95%
air and 5% CO2 at 37°C (32).
Morphological observation
OEC-M1 cells were seeded at a concentration of
6×105 cells in a 6-cm Petri dish with 2 ml culture
medium. After reaching 70–80% confluence, cells were treated
without or with various concentrations of NaAsO2 (0.1,
1, 10, 25, 50 and 100 µM), or various concentrations of DMA (0.1,
1, 2, 5, 10, 25, 50 and 100 mM) for 24 h. Cell morphology was then
observed under an Olympus CK40 light microscope and recorded by an
Olympus DP20 digital camera (Olympus Corporation).
MTT viability test
OEC-M1 cells were seeded at a concentration of
1×104 cells/well with 100 µl culture medium. After
reaching 70–80% confluence, cells were treated without or with
various concentrations of NaAsO2 (0.1, 1, 10, 25, 50 and
100 µM) or various concentrations of DMA (0.1, 1, 10, 25, 50 and
100 mM) for 24 h. MTT was added to a final concentration of 0.5
mg/ml and incubated at 37°C for 4 h. The medium was discarded and
50 µl DMSO was added to each well to dissolve the crystals by
shaking the plate for 20 min in the dark (33,34).
The absorbance values in each treatment group were determined at a
wavelength of 570 nm by VersaMax ELISA reader (Molecular Devices,
LLC).
Cell cycle analysis
To further confirm whether NaAsO2 and DMA
could induce apoptosis among these oral cancer cell lines, the
redistribution of the cell cycle was examined by flow cytometric
analysis through propidium iodide (PI) staining. OEC-M1 cells were
seeded at a concentration of 6×105 cells. After reaching
70–80% confluence, cells were treated without or with various
concentrations of NaAsO2 (0.1, 1, 10, 25, 50 and 100
µM), or various concentrations of DMA (0.1, 1, 10, 25, 50 and 100
mM) for 24 h. Treated cells were harvested by trypsin and
centrifuged (400 × g for 12 min at 4°C), and then washed by isoton
II and fixed in 70% ethanol at −20°C for ≥2 h. After fixation,
cells were washed with isoton II again and collected by
centrifugation (400 × g for 12 min at 4°C). Cell pellets were
resuspended with isoton II and mixed with 100 µg/ml RNase and 40
µg/ml PI for 30 min (15). The
stained cells were analyzed using a flow cytometer (FACScan; BD
Biosciences) with excitation set at a wavelength of 488 nm. The DNA
contents of normal cells in the G1 phase are diploid, while that in
the G2/M phase increases after DNA synthesis progression. Cells in
the subG1 phase have less DNA contents in cell cycle distribution,
which is called hypodiploid and considered as an outcome of cell
apoptosis (34,35). The percentages of sub-G1, S and G2/M
phase cells were analyzed using FACStation v6.1× and Modfit LT v3.3
software (BD Biosciences).
Annexin V/PI double staining
assay
Treated cells were harvested by trypsin and washed
with 2 ml culture medium. Following centrifugation at 160 × g for
10 min at 4°C, the pellets were resuspended with cold isoton II and
centrifuged (400 × g for 12 min at 4°C) again. The pellets were
subsequently mixed with 100 µl staining solution for 15 min
according to the user's manual of the Annexin V-FITC apoptosis
detection kit (Strong Biotech). The stained cells were analyzed at
a wavelength of 488 nm excitation using a 515-nm band pass filter
for FITC detection and >600-nm band pass filter for PI detection
by a FACScan flow cytometer (BD Biosciences). The plots were
divided into four quadrants, which represent negative staining
(viable) cells, Annexin V-positive (early apoptosis) cells,
PI-positive (necrosis) cells, and Annexin V/PI double-positive
(late apoptosis) cells. In addition, cells were also treated with
staurosporine which was considered as a positive control. The
percentage of cells in the 4 quadrants was analyzed using
FACStation v6.1× software.
Protein extraction and immunoblotting
analysis
Cells (6×105) were seeded in a 60-mm
dish. After reaching 70–80% confluence, cells were treated without
or with various concentrations of NaAsO2 (10 and 25 µM),
or various concentrations of DMA (10 and 25 mM) for 3, 6, 12 and 24
h. Medium was then transferred to a 15-ml tube, and centrifuged at
1,500 × g for 10 min at 4°C. Attached cells were lysed using 100 µl
lysis buffer with proteinase inhibitor (cat. no. P8340;
Sigma-Aldrich; Merck KGaA). The pellets were resuspended with 10 µl
lysis buffer, blended into cell lysates, and centrifuged at 12,000
× g for 12 min at 4°C. The supernatants were collected and stored
at −80°C. The protein concentrations of cell lysates were
determined by the Lowry assay (35,36).
For immunoblotting analysis, cell lysates (30 µg)
were resolved on 12% SDS-PAGE gel with standard running buffer at
room temperature, and electrophoretically transferred to
polyvinyldifluoride membranes at 4°C. The membranes were blocked in
4% milk for 1 h at room temperature and incubated with primary
antibodies [cleaved caspase-8 (product no. 9429; 1:1,000), cleaved
caspase-9 (product no. 9509; 1:1,000), cleaved caspase-3 (product
no. 9661; 1:1,000), cleaved PARP (product no. 9542; 1:1,000),
phospho-JNK (product no. 9251; 1/4,000), JNK (product no. 9252;
1:1,000), phospho-ERK1/2 (product no. 9101; 1:4,000), ERK1/2
(product no. 9102; 1:4,000), phospho-p38 (product no. 9215;
1:1,000), p38 (product no. 9212; 1:4,000)] overnight at 4°C.
Following washing with TBS Tween-20 and incubation with horseradish
peroxidase-conjugated secondary antibodies for 1 h at room
temperature [Donkey anti-rabbit IgG; cat. no. NEF81200-1EA;
1:2,000], the membranes were visualized using the ECL detection kit
and UVP EC3 BioImaging Systems (UVP; Analytik Jena) (37). The quantification of each band was
performed using the ImageJ version 1.50 software (National
Institutes of Health).
Statistical analysis
The data are expressed as the mean ± standard error
of the mean (SEM) of at least three independent experiments. The
statistical significance of differences between the control and
treatment groups was determined by one-way ANOVA, followed by
Tukey's post hoc test comparisons. Statistical analysis was
performed with GraphPad Prism 6 software (GraphPad Software, Inc.).
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of arsenic compounds on
morphological changes in OEC-M1 cells
OEC-M1 cells were treated with medium,
NaAsO2 (trivalent arsenic) (0.1, 1, 10, 25, 50 and 100
µM) or DMA (penta-valent arsenic) (0.1, 1, 10, 25, 50 and 100 mM)
for 24 h, and the morphological changes were examined. In the
experiment with NaAsO2, OEC-M1 cells exhibited a
spindle-like shape in the control group (Fig. 1A), and became rounded up and
suspended in the medium as the concentration increased, with the
number of attached cells considerably decreasing in the 25–100 µM
treatment groups (Fig. 1E-G). In
the DMA experiment, cells started to float in the groups of OEC-M1
cells treated with 0.1–1 mM DMA (Fig.
1H and I). As the concentration of DMA increased, cells shrank
with membrane blebbing, indicating that they were undergoing
apoptosis (Fig. 1H-M). Floating
cells tended to aggregate in a high concentration of DMA treatment
compared with NaAsO2treatment. These results indicated
that both NaAsO2 and DMA could induce the morphological
changes associated with apoptosis in OEC-M1 oral cancer cells.
 | Figure 1.Effects of arsenic compounds on
morphological changes in OEC-M1 cells. Cells were treated with (A)
plain medium, (B-G) 0.1, 1, 10, 25, 50 and 100 µM sodium arsenite,
respectively, or (H-M) 0.1, 1, 10, 25, 50 and 100 mM
dimethylarsenic acid in OEC-M1 cells for 24 h, respectively.
Morphological changes were observed under light microscopy (scale
bar, 50 µm). Arrowheads indicate membrane-blebbing cells. (N)
Apoptotic cell (scale bar, 10 µm). The arrowhead indicates an
apoptotic cell with membrane blebbing). Experiments were performed
three times with similar results. |
Effects of arsenic compounds on the
viability of OEC-M1 cells
The survival rate of OEC-M1 cells treated with
arsenic compounds was further examined by MTT cell viability assay.
Cells were treated with medium, NaAsO2 (0.1, 1, 10, 25,
50 and 100 µM) or DMA (0.1, 1, 10, 25, 50 and 100 mM) for 24 h. The
OEC-M1 cell survival rate was significantly decreased by
NaAsO2 at doses of 10–100 µM (Fig. 2A) and by DMA from 1–100 mM (Fig. 2B) in dose-dependent manners
(P<0.05). The concentration of DMA required to reduce cell
viability to 50% in OEC-M1 cells was ~1,000-times higher compared
with that of NaAsO2.
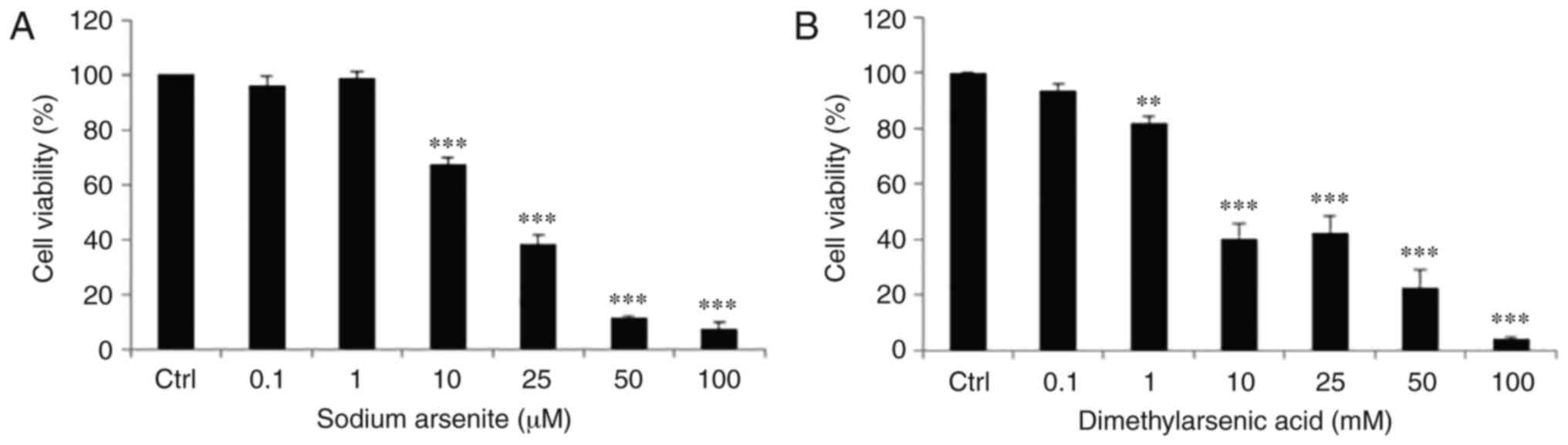 | Figure 2.Effects of arsenic compounds on cell
viability in OEC-M1 cells. Cells were treated with (A) 0, 0.1, 1,
10, 25, 50 and 100 µM NaAsO2, or (B) 0, 0.1, 1, 10, 25,
50 and 100 mM DMA for 24 h. Cell viability was quantified by MTT
assay. Results are expressed as percentages of cell growth relative
to the control groups as 100%. The data are expressed as mean ± SEM
of at least three separate experiments. **P<0.01 and
***P<0.001 represent statistical differences compared to the
Ctrl. NaAsO2, sodium arsenite; DMA, dimethylarsenic
acid; SEM, standard error of the mean; Ctrl, control. |
Effects of arsenic compounds on cell
cycle distribution of OEC-M1 cells
To assess whether NaAsO2 and DMA could
induce OEC-M1 oral cancer cell death via apoptosis, cells were
treated with arsenic compounds and then evaluated for their DNA
contents by PI staining via flow cytometric analysis. Thus, OEC-M1
cells were treated with medium, NaAsO2 (0.1, 1, 10, 25,
50 and 100 µM) or DMA (0.1, 1, 10, 25, 50 and 100 mM) for 24 h, and
the changes in the number of cells in the subG1, G1 and G2/M phases
related to cell cycle regulation were investigated.
NaAsO2 at 50 and 100 µM significantly
increased the number of cells in the subG1 phase in OEC-M1 cells
(Fig. 3A and B) (P<0.05). In
addition, the number of cells in the G1 phase was decreased by
25–100 µM NaAsO2 in OEC-M1 cells (Fig. 3A and C) (P<0.05). Furthermore,
25–100 µM NaAsO2 significantly increased the number of
OEC-M1 cells in the G2/M phase (Fig. 3A
and D) (P<0.05). DMA at 1, 10, 50 and 100 mM significantly
increased the number of cells in the subG1 phase (Fig. 4A and B); 1–100 mM significantly
reduced the number of cells in the G1 phase (Fig. 4A and C) (P<0.05); and 1–100 mM
significantly increased the number of cells in the G2/M phase
(Fig. 4A and D) (P<0.05). These
results demonstrated that both NaAsO2 and DMA could
regulate the distribution of OEC-M1 cells in the subG1, G1 and G2/M
phases of the cell cycle to induce cell apoptosis.
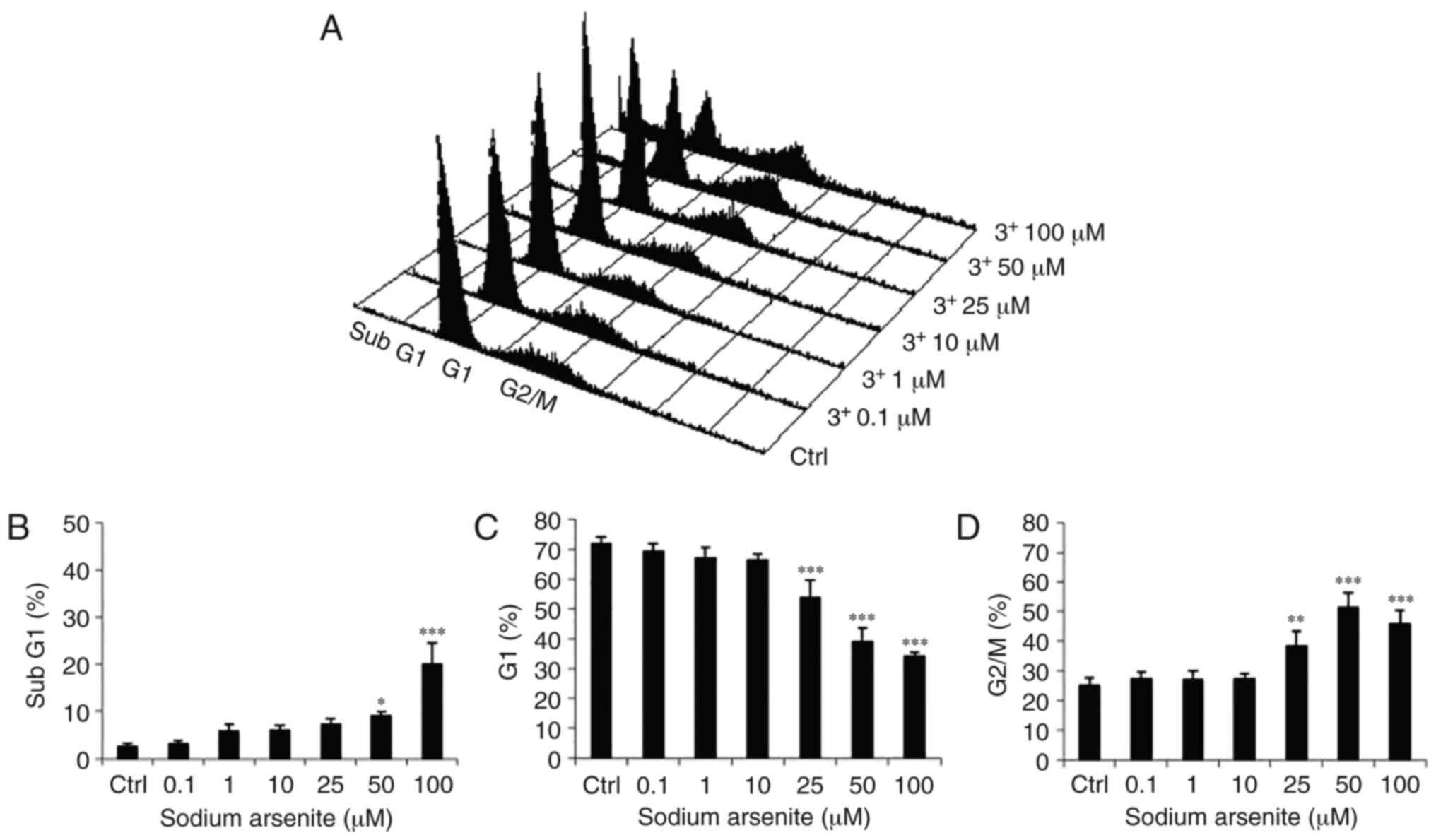 | Figure 3.Effects of NaAsO2 on cell
cycle redistribution in OEC-M1 cells. (A) Various concentrations of
NaAsO2 (0, 0.1, 1, 10, 25, 50 and 100 µM) were used to
treat OEC-M1 for 24 h. Cells were fixed, stained with PI, and
analyzed by flow cytometry. Cells in subG1 phase contain less DNA
content than normal cells, indicating apoptosis. The percentage of
(B) sub G1, (C) G1 and (D) G2/M phase cells are presented,
respectively. The data are expressed as the mean ± SEM of at least
three separate experiments. *P<0.05, **P<0.01, and
***P<0.001 represent statistical differences compared to the
Ctrl group. NaAsO2, sodium arsenite; PI, propidium
iodide; SEM, standard error of the mean; Ctrl, control. |
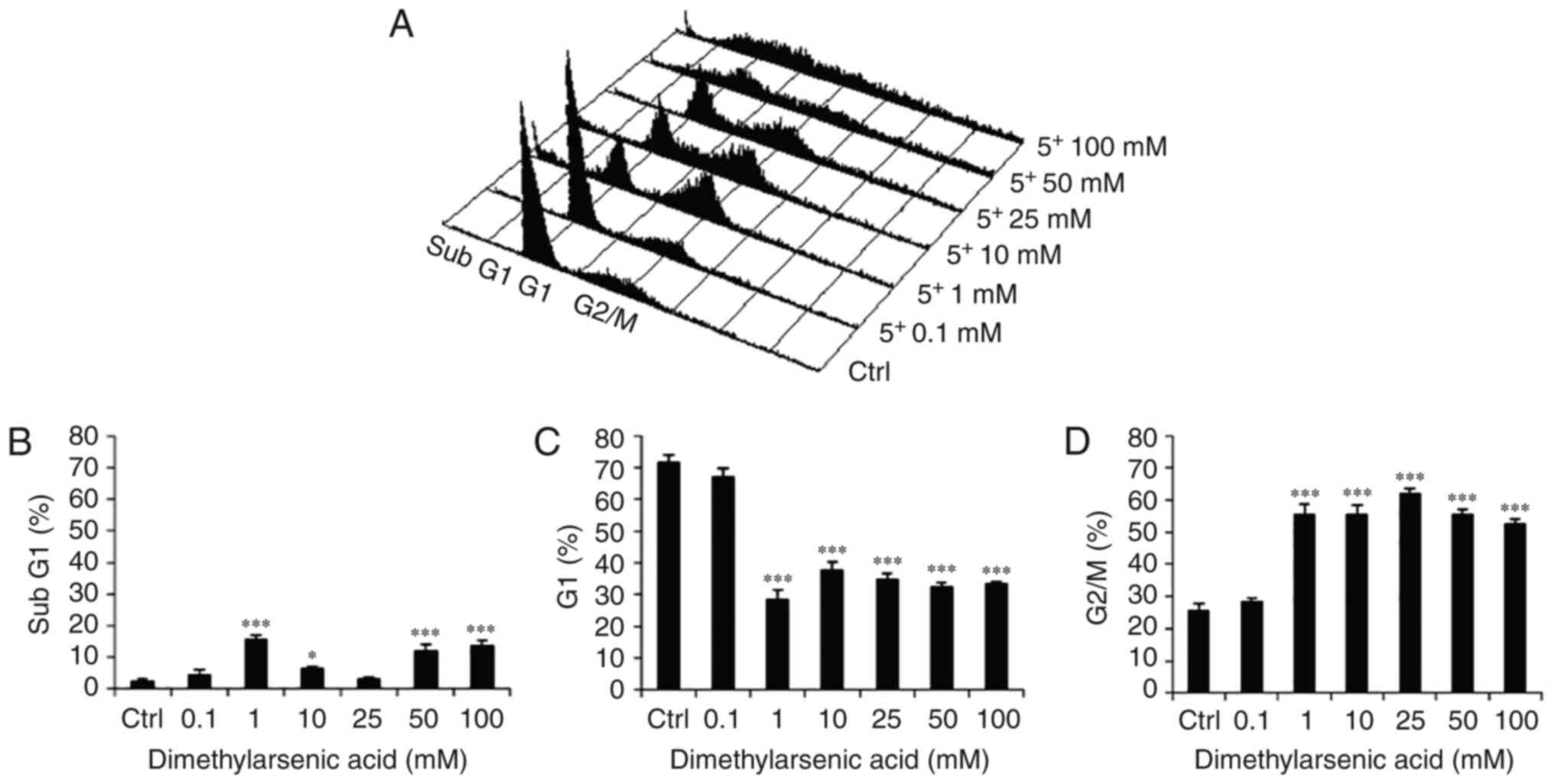 | Figure 4.Effects of DMA on cell cycle
redistribution in OEC-M1 cells. (A) Various concentrations of DMA
(0, 0.1, 1, 10, 25, 50 and 100 mM) were used to treat OEC-M1 for 24
h. Cells were fixed, stained with PI, and analyzed by flow
cytometry. Cells in subG1 phase contain less DNA content than
normal cells, indicating apoptosis. The percentage of (B) sub G1,
(C) G1 and (D) G2/M phase cells are presented, respectively. The
data are expressed as the mean ± SEM of at least three separate
experiments. *P<0.05 and ***P<0.001 represent statistical
differences compared to the Ctrl group. DMA, dimethylarsenic acid;
SEM, standard error of the mean; PI, propidium iodide; Ctrl,
control. |
Arsenic compounds induce apoptosis in
OEC-M1 cells
To further confirm whether apoptosis is induced by
arsenic compounds in OEC-M1 cells, double staining with Annexin V
and PI via flow cytometer was performed. The percentages of
double-negative (viable), PI-positive (necrotic), Annexin
V-positive (early apoptotic) and double-positive (late apoptotic)
cells can be revealed in four quadrants by the double staining
assay to assess cell apoptosis (38). Treatment with NaAsO2
(10–100 µM) (Fig. 5A) and DMA
(1–100 mM) (Fig. 6A) for 24 h
significantly induced cell apoptosis, and the number of Annexin
V-positive cells significantly increased as the concentrations of
sodium NaAsO2 (Fig. 5B and
C) and DMA increased (Fig. 6B and
C) (P<0.05). These results demonstrated that both
NaAsO2 and DMA promoted apoptosis of OEC-M1 cells.
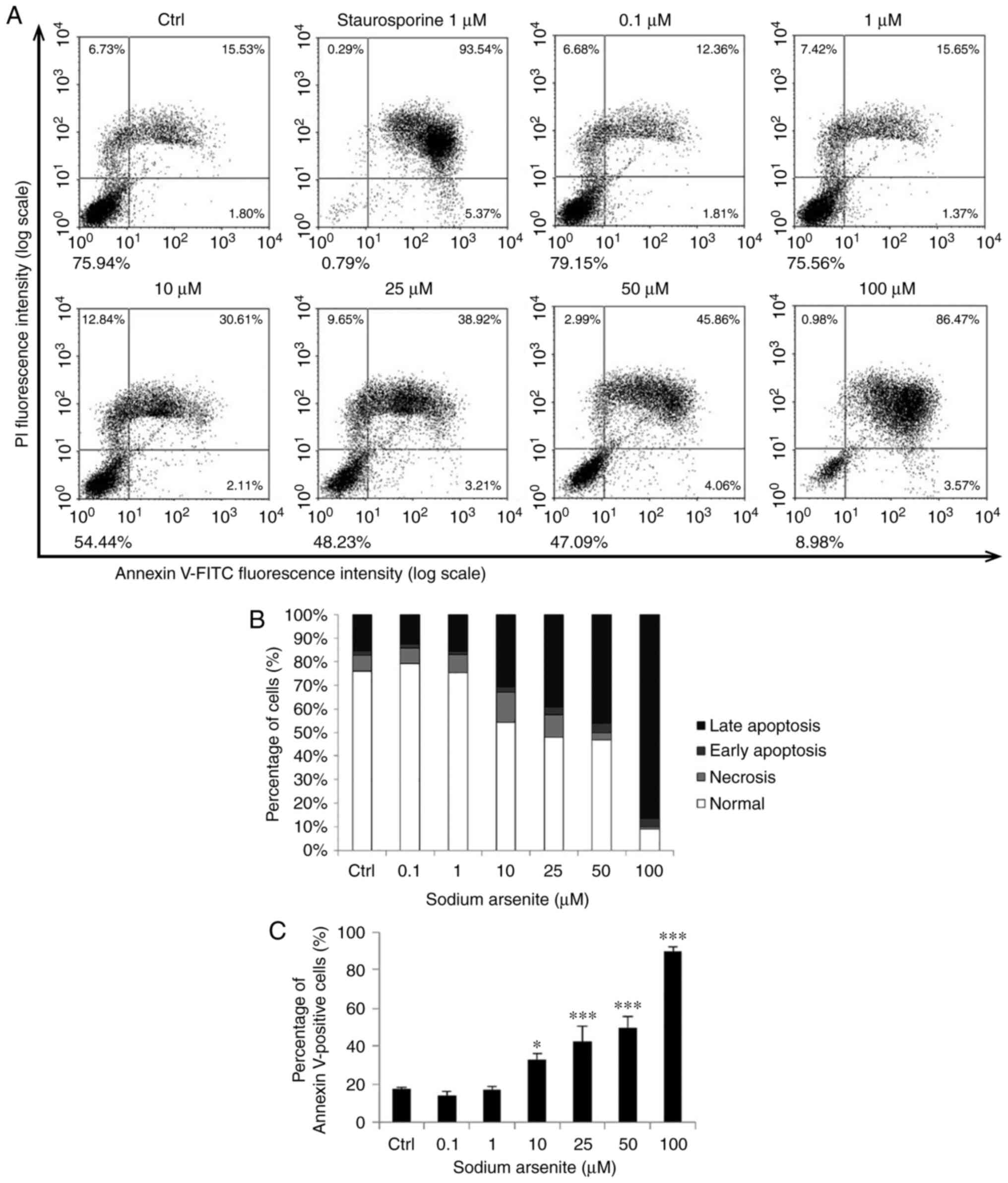 | Figure 5.NaAsO2 induces OEC-M1 cell
apoptosis. Various concentrations of NaAsO2 (0, 0.1, 1,
10, 25, 50 and 100 µM) were used to treat OEC-M1 for 24 h. (A) The
apoptotic status was determined by Annexin V/PI double staining
assay, and the staurosporine-treated cells were considered as a
positive control. (B) The percentages of double-negative (viable),
PI single-positive (necrotic), Annexin V single-positive (early
apoptotic), and double-positive (late apoptotic) cells are
presented in. (C) Annexin V-positive cells were analyzed and are
presented. The data are expressed as the mean ± SEM of at least
three separate experiments. *P<0.05 and ***P<0.001 represent
statistical differences compared to the Ctrl group.
NaAsO2 sodium arsenite; SEM, standard error of the mean;
PI, propidium iodide; Ctrl, control. |
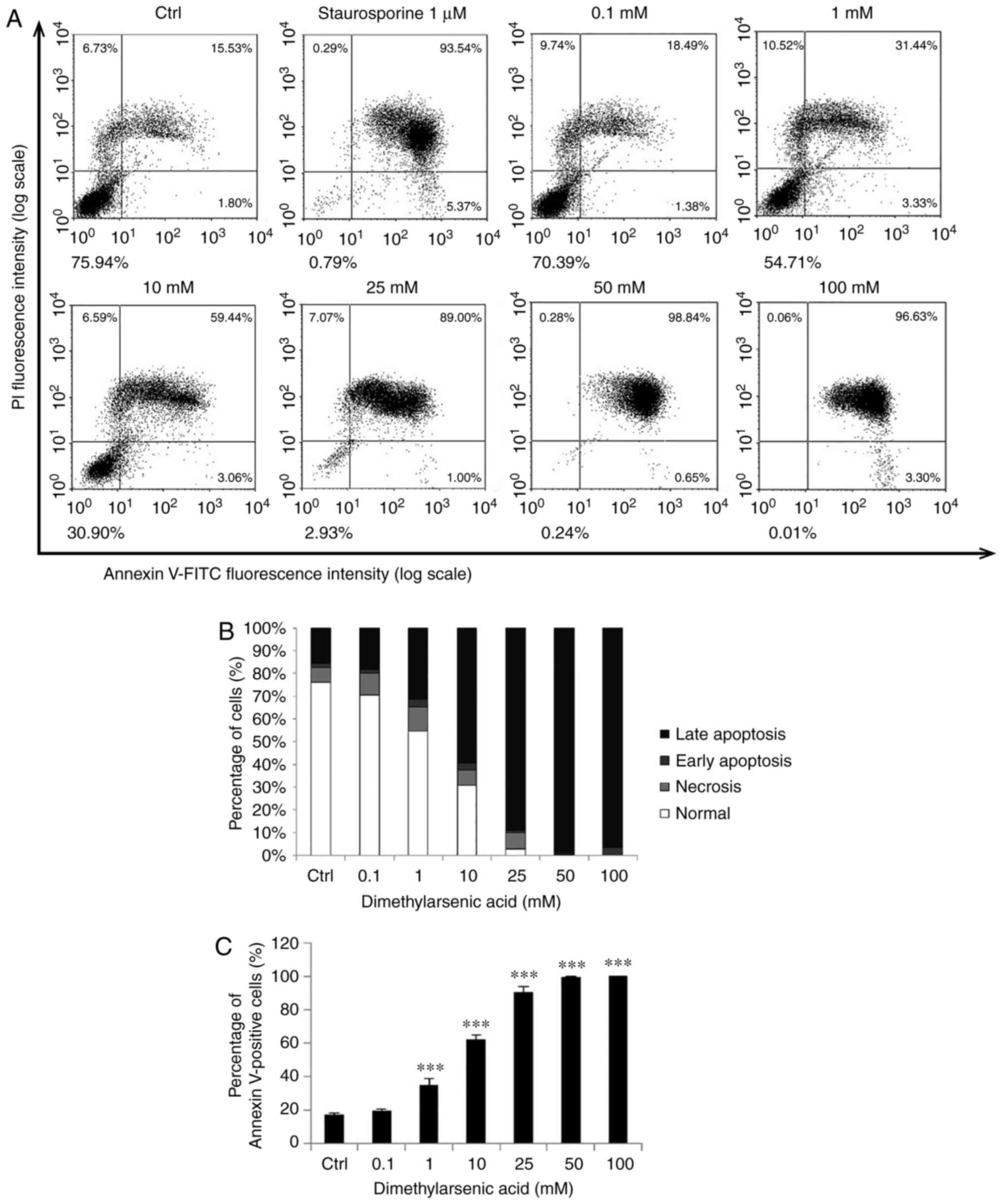 | Figure 6.DMA induces OEC-M1 cell apoptosis.
Various concentrations of DMA (0, 0.1, 1, 10, 25, 50 and 100 mM)
were used to treat OEC-M1 for 24 h. (A) The apoptotic status was
determined by Annexin V/PI double staining assay, and the
staurosporine-treated cells were considered as a positive control.
(B) The percentages of double-negative (viable), PI single-positive
(necrotic), Annexin V single-positive (early apoptotic), and
double-positive (late apoptotic) cells are presented. (C) Annexin
V-positive cells were analyzed and are presented. The data are
expressed as the mean ± SEM of at least three separate experiments.
***P<0.001 represent statistical differences compared to the
Ctrl group. DMA, dimethylarsenic acid; SEM, standard error of the
mean; PI, propidium iodide; Ctrl, control. |
Involvement of the extrinsic and
intrinsic pathways of caspases in arsenic-induced OEC-M1 cell
apoptosis
To further investigate whether arsenic compounds can
induce OEC-M1 cell apoptosis via the death receptor (extrinsic)
and/or mitochondrial (intrinsic) apoptotic pathways, the expression
levels of cleaved caspase-8 and caspase-9, and the downstream
targets of cleaved caspase-3 and PARP were examined by western
blotting (Fig. 7). The selected
concentrations of NaAsO2 (10 and 25 µM) and DMA (1 and
10 mM) used to determine the involvement of the extrinsic and
intrinsic pathways in the present experiments were based on the
doses that could reduce cell viability from ~40 to 60% covering 50%
(Fig. 2) and induce cell apoptosis
from ~30 to 45% for NaAsO2 (Fig. 5) and from ~35 to 60% for DMA,
respectively (Fig. 6).
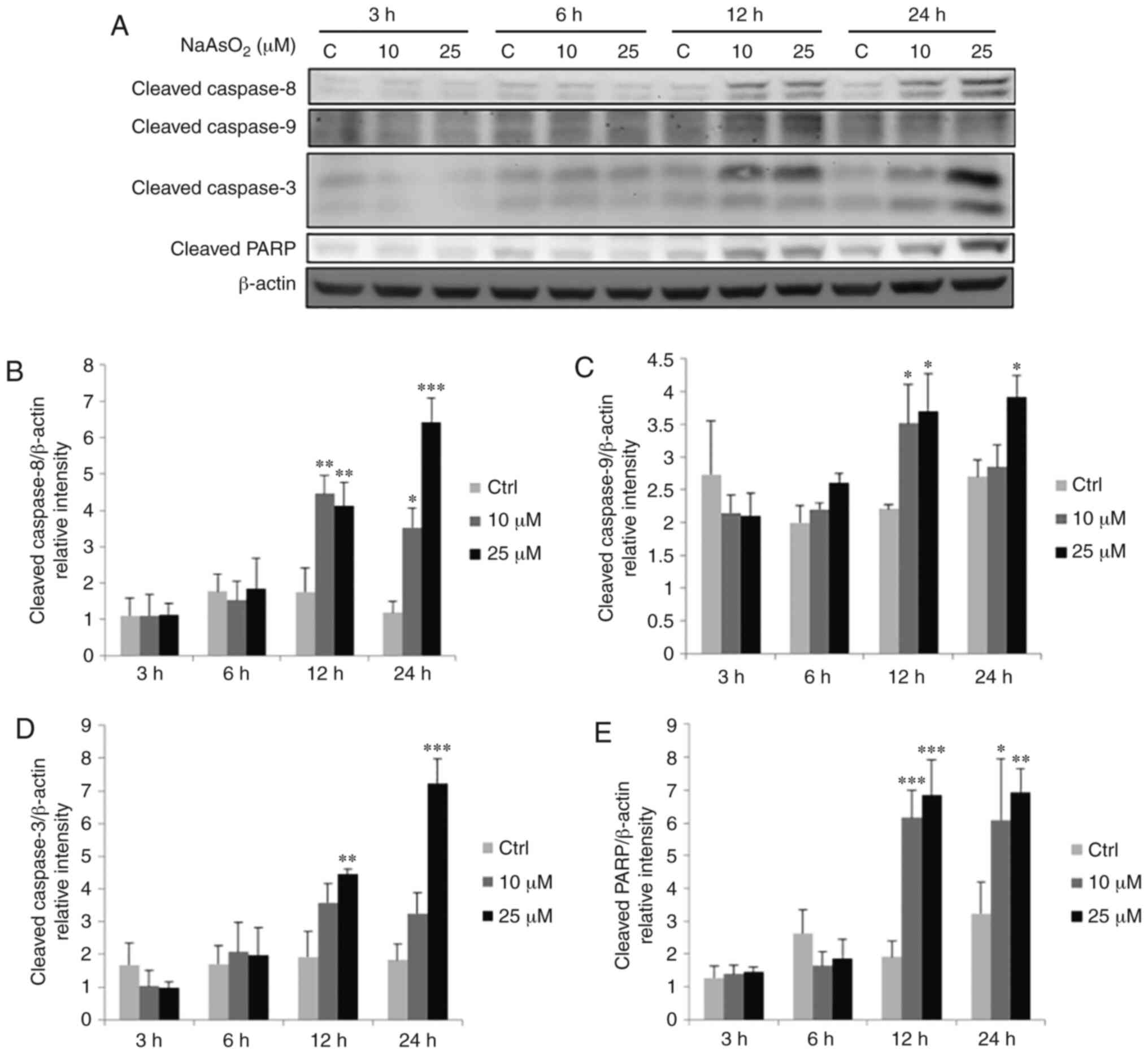 | Figure 7.Effects of NaAsO2 on the
expression levels of cleaved caspase-8, −9, −3 and PARP proteins in
OEC-M1 cells. Cells were treated without or with 10 and 25 µM
NaAsO2 for 3, 6, 12 and 24 h, respectively. (A) Cleaved
caspase-8 (43 kDa), −9 (35/37 kDa), −3 (17/19 kDa) and PARP (~85-90
kDa) were detected by western blotting. The IODs of cleaved (B)
caspase-8, (C) caspase-9, (D) caspase-3 and (E) PARP proteins were
normalized with β-actin (43 kDa) in each lane. The data are
expressed as the mean ± SEM of at least three separate experiments.
*P<0.05, **P<0.01, and ***P<0.001 represent statistical
differences compared to the Ctrl group. NaAsO2, sodium
arsenite; PARP, poly(ADP-ribose) polymerase; IODs, integrated
optical densities; SEM, standard error of the mean; C or Ctrl,
control. |
NaAsO2 at 10 and 25 µM for 12 and 24 h
significantly activated caspase-8 (Fig.
7A and B) and PARP (Fig. 7A and
E), while caspase-9 was significantly activated by 10 and 25 µM
NaAsO2 for 12 h, and 25 µM NaAsO2 for 24 h
(Fig. 7A and C) (P<0.05).
Caspase-3 was significantly activated by treatment with 25 µM
NaAsO2 for 12 and 24 h (Fig.
7A and D) (P<0.05). DMA at 1 mM for 24 h could significantly
induce the cleavage of caspase-8, −9, −3 and PARP (Fig. 8A-E) (P<0.05). The expression of
cleaved PARP was also significantly increased by 10 mM DMA for 24 h
(Fig. 8A and E) (P<0.05). In
addition, 10 mM DMA for 12 h could induce cleavage of caspase-3 and
PARP (Fig. 8A, D and E)
(P<0.05). These results indicated that long-term exposure of
both sodium compounds could activate caspase-8 and caspase-9,
followed by caspase-3 and PARP, to induce both the extrinsic and
intrinsic apoptotic pathways in OEC-M1 cells.
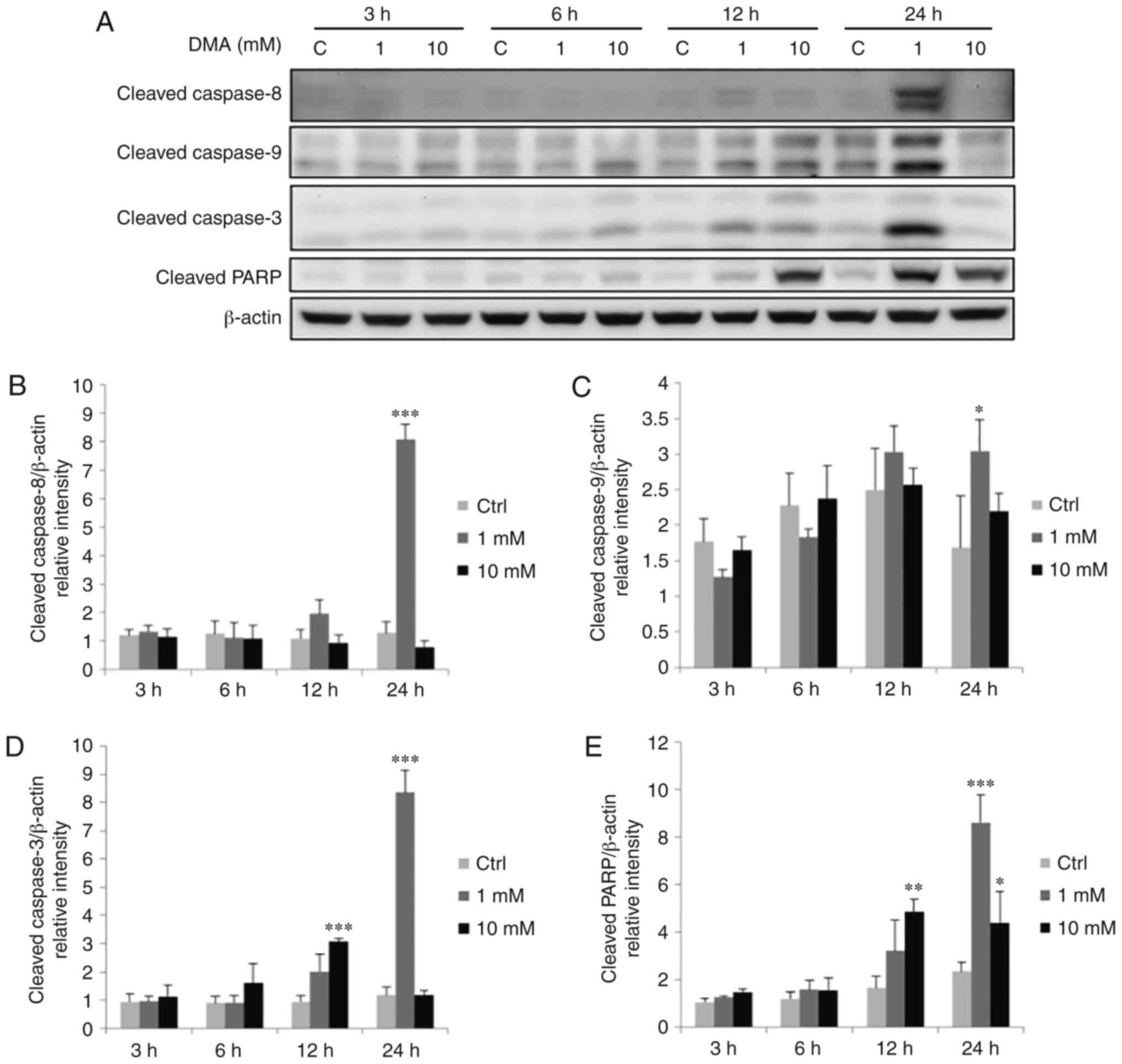 | Figure 8.Effects of DMA on the expression
levels of cleaved caspase-8, −9, −3 and PARP proteins in OEC-M1
cells. Cells were treated without or with 1 and 10 mM DMA for 3, 6,
12 and 24 h, respectively. (A) Cleaved caspase-8 (43 kDa), −9
(35/37 kDa), −3 (17/19 kDa) and PARP (~85-90 kDa) were detected by
western blotting. The IODs of cleaved (B) caspase-8, (C) caspase-9,
(D) caspase-3 and (E) PARP proteins were normalized with β-actin
(43 kDa) in each lane. The data are expressed as the mean ± SEM of
at least three separate experiments. *P<0.05, **P<0.01, and
***P<0.001 represent statistical differences compared to the
Ctrl group. DMA, dimethylarsenic acid; PARP, poly(ADP-ribose)
polymerase; IODs, integrated optical densities; SEM, standard error
of the mean; C or Ctrl, control. |
Involvement of the MAPK pathways in
arsenic-induced apoptosis of OEC-M1 cells
Previous studies have revealed that MAPK pathways
may be involved in regulating cell proliferation, differentiation,
mitosis, cell survival, gene expression and apoptosis (26,27).
To evaluate whether MAPK pathways are associated with arsenic
compound-induced OEC-M1 cell apoptosis, the phosphorylation of JNK,
ERK1/2 and p38 was investigated using western blotting.
NaAsO2 at 10 µM for 24 h and 25 µM for 3,
6 and 24 h could significantly increase the level of p-JNK
(Fig. 9A and B), while 25 µM
NaAsO2 for 3, 6, 12 and 24 h significantly increased the
expression of p-ERK1/2 (Fig. 9A and
C) (P<0.05). The p-p38 level could be significantly
increased by 25 µM NaAsO2 for 6 and 24 h (Fig. 9A and D) (P<0.05). DMA at 10 mM
for 3, 6, 12 and 24 h could significantly increase the level of
p-JNK (Fig. 10A and B), and 10 mM
DMA for 6, 12 and 24 could significantly increase the level of
p-ERK1/2 (Fig. 10A and C)
(P<0.05). The p-p38 level was significantly increased after
treatment with 10 mM DMA for 24 h (Fig. 10A and D) (P<0.05).
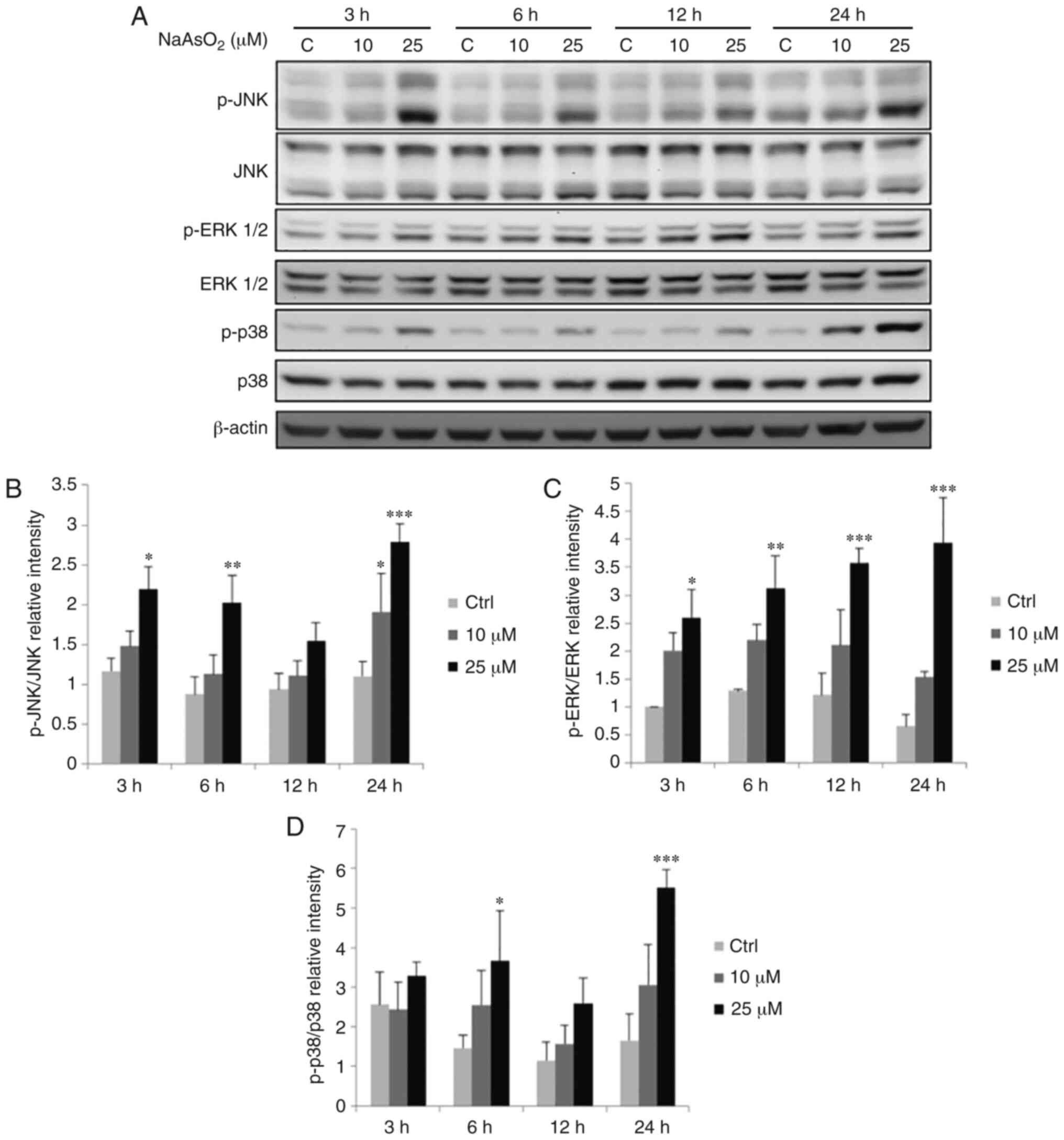 | Figure 9.Effects of NaAsO2 on the
phosphorylation of the MAPK signaling pathway in OEC-M1 cells.
Cells were treated without or with 10 and 25 µM NaAsO2
for 3, 6, 12 and 24 h, respectively. (A) p-JNK (46/54 kDa), JNK,
p-ERK1/2 (42/44 kDa), ERK1/2, p-p38 (43 kDa) and p38 were detected
by western blotting. The IODs of (B) p-JNK, (C) p-ERK and (D) p-p38
proteins were normalized with total forms of themselves in each
lane. The data are expressed as the mean ± SEM of at least three
separate experiments. *P<0.05, **P<0.01, and ***P<0.001
represent statistical differences compared to the Ctrl group.
NaAsO2, sodium arsenite; MAPK, mitogen-activated protein
kinase; p-, phosphorylated; JNK, c-Jun NH2-terminal kinase; ERK,
extracellular signal-regulated kinase; IODs, integrated optical
densities; SEM, standard error of the mean; C or Ctrl, control. |
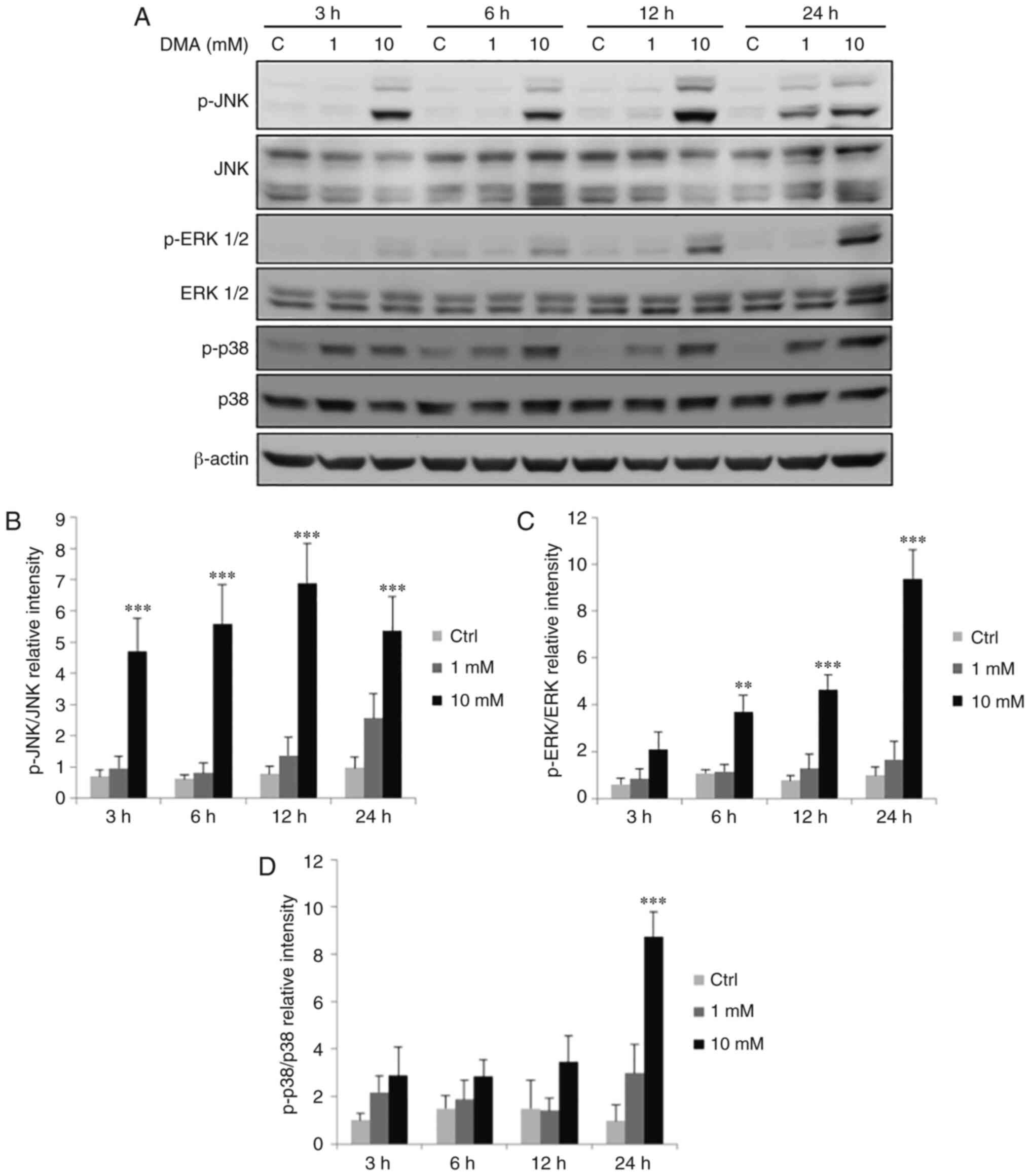 | Figure 10.Effects of DMA on the phosphorylation
of the MAPK signaling pathway in OEC-M1 cells. Cells were treated
without or with 1 and 10 mM DMA for 3, 6, 12 and 24 h,
respectively. (A) p-JNK (46/54 kDa), JNK, p-ERK1/2 (42/44 kDa),
ERK1/2, p-p38 (43 kDa) and p38 were detected by western blotting.
The IODs of (B) p-JNK, (C) p-ERK and (D) p-p38 proteins were
normalized with total forms of themselves in each lane. The data
are expressed as the mean ± SEM of at least three separate
experiments. **P<0.01 and ***P<0.001 represent statistical
differences compared to the Ctrl group. DMA, dimethylarsenic acid;
MAPK, mitogen-activated protein kinase; p-, phosphorylated; JNK,
c-Jun NH2-terminal kinase; ERK, extracellular signal-regulated
kinase; IODs, integrated optical densities; SEM, standard error of
the mean; C or Ctrl, control. |
Discussion
In the present study, NaAsO2 (inorganic
arsenic) and DMA (organic arsenic) both demonstrated the ability to
induce apoptosis of OEC-M1 oral cavity cancer cells. Studies have
revealed that the cytoskeleton may be a target of NaAsO2
and DMA (39,40), and the extent of morphological
changes upon modulating the cytoskeleton correlates with drug
concentration and incubation time (41). The present results demonstrated that
NaAsO2 resulted in a higher number of shriveled and
floating cells with increasing concentration compared with DMA in
OEC-M1 cells. In addition, a greater extent of membrane blebbing
cells could be observed, and floating cells tended to assemble at
50–100 mM DMA treatments. Compared with cells with abalone-like
shape under lower doses of NaAsO2, high doses of DMA
induced cells to attach to the ground matrix but almost rounded up,
while higher doses caused cells to float in clusters. The
regulatory mechanisms through which arsenic compounds lead to
different cell morphologies are unknown. However, the results
indicated that their effect on the cytoskeleton differs with drug
types and concentrations.
Inorganic arsenic is significantly more toxic
compared with organic arsenic compounds (42). Once entering the food chain,
inorganic compounds could be metabolized by methyltransferases in
the liver of several mammalian species (42). Inorganic arsenic could also be
methylated to less toxic compounds, which is considered to be a
detoxification reaction. For example, DMA arises from the
methylation of arsenic oxide (43).
In the present study, NaAsO2 and DMA decreased OEC-M1
cell viability in a dose-dependent manner, which was consistent
with the results of the morphological analysis. The concentration
of NaAsO2 required to perform similar effects was
1,000-fold lower compared with that of DMA, indicating that
NaAsO2 exhibited stronger cytotoxicity than DMA in
OEC-M1 oral cancer cells.
The eukaryotic cell cycle proceeds to the next phase
as the upstream events fulfill the requirements of checkpoints
(44). When undergoing cellular
damage, cells may become arrest at certain phases and may undergo
programmed cell death if the damage is not properly repaired
(44). It has been reported that
arsenite can promote disruption of the microtubule network,
resulting in spindle abnormalities and, thus, mitotic cell
apoptosis (45). In fact,
arsenic-induced mitotic arrest may be a necessary step for the
activation of apoptotic pathways in numerous human tumor cell lines
(46). A previous study has also
demonstrated a strong correlation between G2/M arrest and the
induction of apoptosis in ovarian carcinoma cell lines in response
to DNA damage (47). It is well
known that the subG1 phase is considered as a sign of DNA
fragmentation, which indicates cell apoptosis (48), and a previous study has also
revealed that G2/M-phase arrest can induce cells to undergo
apoptosis (49). In the present
study, NaAsO2 and DMA significantly induced G2/M-phase
arrest and increased the number of subG1-phase cells, which
suggests that both arsenic compounds could induce apoptosis of
OEC-M1 cells. Hence, the present data indicated that
arsenic-induced apoptosis is associated with abnormal cell cycle
redistribution. The Annexin V/PI double-staining assay further
verified that the induction of apoptosis by NaAsO2 and
DMA occurred in a dose-dependent manner in OEC-M1 cells. However,
the examination of cell cycle protein levels was not conducted in
the present study, and it is worth investigating, in a future
study, at cellular and molecular levels how the expression of cell
cycle proteins are regulated by NaAsO2 and DMA to induce
OEC-M1 cell apoptosis. Similar to the cell viability assay, once
again, NaAsO2 demonstrated 1,000-fold stronger effects
compared with DMA on cell cycle regulation in OEC-M1 oral cancer
cells.
Apoptosis is a critical process for maintaining
homeostasis (50). Dysregulation of
apoptosis is considered to be a key factor in the development of
cancer (50). It is well known that
apoptosis is initiated by either an extrinsic or an intrinsic death
signal, and executed by the activated caspase cascade (51). It is reported that arsenic trioxide
can induce the apoptosis of laryngeal cancer cells via
downregulation of survivin mRNA, which inhibits activation of
caspases (52). Arsenic trioxide
also induces apoptosis in different cancers by activating
caspase-8, −9 and −3 (11,53). In the present study, the expression
levels of cleaved caspase-8, −9, −3 and the downstream substrate
PARP were found to be increased by both NaAsO2 and DMA
in OEC-M1 cells, and NaAsO2 did exhibit a stronger
activating effect compared with DMA. Thus, these results supported
that both arsenic compounds induced oral cavity cancer cell
apoptosis through extrinsic and intrinsic apoptotic pathways. It
should be noted that, in the results, the expression of some
caspases was most prominently increased by DAM at 1 mM, but not at
10 mM. It is highly possible that DAM at 1 mM has the maximal
effect on the caspase pathway, which then starts to degrade. Thus,
DAM at 10 mM could not induce a greater effect on OEC-M1 cell
apoptosis. In addition, DAM exclusively induced some caspases at a
concentration of 1 mM, indicating that phosphorylation of JNK, ERK
and p38, significantly induced by DAM at 10 mM, could not be
associated with the activation of caspases. In fact, DAM at 10 mM
also induced the cleavage of PARP at 24 h. It is highly possible
that the expression of cleaved caspase-8, −9 and −3 had degraded at
24 h after the treatment of DAM. Thus, DAM at 10 mM could stimulate
phosphorylation of JNK, ERK and p38, and then further induce the
caspase pathway.
In fact, studies have reported that arsenic
compounds can activate different signaling pathways to induce cell
death related to apoptotic pathways among different cancer cell
types, such as myeloma cells, neutrophils and/or leukemia (54–58).
Studies have demonstrated that arsenic compounds could only
activate the caspase-8 pathway to induce human melanoma cell
apoptosis by enhancing TRAIL-induced apoptosis with the suppression
of the NF-κB and STAT3-transcriptional targets (59) and could only stimulate the caspase-9
pathway to induce liver cancer cell apoptosis by downregulating
phosphorylated-STAT3 expression with Bcl-2, XIAP, and surviving
proteins (60), respectively.
Studies have also revealed that arsenic compounds could activate
both the caspase-8 and caspase-9 pathways concurrently to induce
myeloma and promonocytic leukemia cell apoptosis by decreasing the
expression of Bcl-2 families and/or regulating p38-MAPK process
(61,62). In the present study,
NaAsO2 and DMA could concurrently induce both the
caspase-8 and −9 pathways in OEC-M1 cells, indicating that our
observations are not unprecedented. It is well known that up- and
downstream of the caspase-8 and −9 pathways, pathways including
Fas/Fas ligand, BCL-2, ER stress, and/or p53, are markedly involved
with the finer regulation of cancer cell apoptosis (59–62).
Thus, it is worth further investigating how caspase-8 and −9
pathways are regulated by up- and downstream regulators to induce
OEC-M1 cell apoptosis.
Apoptotic pathways are associated with numerous
regulating mechanisms, one of which is the MAPK signaling pathway
responding to cellular stress. MAPK signaling may either promote
survival or enhance sensitivity to apoptosis depending on the cell
types, stimuli and the latency of the activation of MAPKs (26,27).
In the present study, p-JNK, p-ERK1/2 and p-p38 were all increased
by both NaAsO2 and DMA in OEC-M1 cells. These results
indicated that both NaAsO2 and DMA could activate the
phosphorylation of JNK, ERK1/2 and/or p38 to induce apoptosis in
OEC-M1 cells. Notably, these results demonstrated that activation
of caspases and PARP was observed following treatments with arsenic
compounds for 12 and 24 h, and the phosphorylation of MAPK occurred
3–6 h earlier compared with the caspase-mediated pathway,
indicating that arsenic compounds possibly activated MAPK pathways
prior to the caspases and PARP to induce apoptosis in OEC-M1 cells.
It should be noted that 1 mM DMA significantly suppressed OEC-M1
cell viability, affected the cell cycle, increased the number of
apoptotic cells and activated caspase proteins, but not MAPK
proteins. With a closer examination of the western blot results, 1
mM DMA induced an increasing trend of MAPK protein expression
although there was no statistical difference. Thus, these
observations demonstrated that arsenic compounds activated
different MAPK pathways first and then induced apoptosis in OEC-M1
oral cavity cells, which is consistent with the findings of other
studies that have revealed that the induction of MAPK pathways and
then caspase cascade under stimuli occurs in sequence (63–65).
In the present study, both NaAsO2 and DMA
could induce cell apoptosis through extrinsic and intrinsic
apoptotic pathways, exerting potential antitumor effects on OEC-M1
oral cancer cells. Furthermore, both arsenic compounds were able to
induce phosphorylation of JNK, ERK and p38 MAPKs to modulate the
activation of the caspase cascade to stimulate apoptosis of OEC-M1
cells. However, the precise roles upon the activation of MAPK and
caspase pathways by NaAsO2 and DMA in OEC-M1 cells
remain elusive, since specific inhibitors and/or siRNA were not
used in the present study to block apoptosis. With the use of
specific inhibitors and/or siRNA to suppress OEC-M1 cell apoptosis,
the activation of MAPK and caspase pathways by NaAsO2
and DMA in OEC-M1 cells can then be confirmed, which will
conclusively reveal the underlying molecular mechanism. In
addition, studies have demonstrated that intravenous arsenic
trioxide and its derivatives both alone and/or in combination with
other agents have been used successfully for the treatment of APL
and myeloid neoplasms, and arsenic trioxide is a standard treatment
for APL patients to date (66,67).
In fact, oral liquid and pill formulations of arsenic trioxide have
recently entered clinical practice for APL patients (66,67).
Clinical trials involving arsenic compounds have never been applied
to head and neck cancers for their possible use in clinical
practice. With the observations in the present study,
NaAsO2 and DMA could possibly be applied for the therapy
of oral cancers with intravenous infusion and/or oral intake as a
tablet or pill (66,67). Thus, clinical trials should be
performed with arsenic compounds in order to possibly provide a
helpful alternative method for the treatment of oral cancers.
It should be noted that the error bars are rather
sizable in certain figures, regarding the western blot analysis of
caspase and MAPK protein expression activated by NaAsO2
and DMA in OEC-M1 cells. It is well known that in in vitro
cell culture experiments, the expression levels of proteins may
markedly vary due to the cell conditions related to the cell
passages, cell quality, and cell seeding number with the possible
changes in experimental room temperature and humidity, incubator
temperature and humidity, western blot-conducting skills, and the
integrating optical intensities of protein bands with UVP skills.
In addition, the original data from 3 independent experiments
without replicates for each treatment were used to perform
statistical analysis and to draw figures in the present study.
Thus, the error bars became rather sizable. Reliably, the
statistical analysis could still show there are differences between
treatments. Thereafter, in a future study, the same experiment
should be repeated more than 3 times with 2 and/or 3 more
replicates in each treatment, which could decrease error bar values
and increase the fidelity of results. In conclusion,
NaAsO2 and DMA stimulated the extrinsic and intrinsic
apoptotic pathways through the activation of MAPK pathways to
induce apoptosis in OEC-M1 cells, demonstrating that both arsenic
compounds exhibit promising antitumor properties and may be used
for clinical therapy with marked potential for oral cancers.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Ministry
of Science and Technology of Taiwan, R.O.C (grant no.
MOST106-2811-B-006-014, to BMH) and An-Nan Hospital, Taiwan, R.O.C
(grant no. ANHRF108-20, to NPF, CLK, ECS and BMH).
Availability of data and materials
The data that support the findings of this study are
available from the corresponding author upon reasonable
request.
Authors' contributions
NPF, CLK and CYC designed the experiments, carried
out the experiments, interpreted the results, and wrote the
manuscript. CYW, ECS and BMH participated in the design and
coordination of the study, were involved in the statistical
analysis of the obtained results and were responsible for the
revision of the manuscript for substantive and methodological
correctness. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
David MC, Randal SW and Stephen YL: Head
and neck cancer. Cancer. 113 (Suppl 7):S1911–S1932. 2008.
View Article : Google Scholar
|
|
2
|
Laraway DC, Lakshmiah R, Lowe D, Roe B and
Rogers SN: Quality of life in older people with oral cancer. Br J
Oral Maxillofac Surg. 50:715–720. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sudbø J: Human papillomavirus infection as
a risk factor for squamous-cell carcinoma of the head and neck. N
Engl J Med. 345:376–377. 2001. View Article : Google Scholar
|
|
4
|
Ko YC, Huang YL, Lee CH, Chen MJ, Lin LM
and Tsai CC: Betel quid chewing, cigarette smoking and alcohol
consumption related to oral cancer in Taiwan. J Oral Pathol Med.
24:450–453. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bernier J and Cooper JS: Chemoradition
after surgery for high-risk head and neck cancer patients: How
strong is the evidence? Oncologist. 10:215–224. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chen CJ, You SL, Lin LH, Hsu WL and Yang
YW: Cancer epidemiology and control in Taiwan: A brief review. Jpn
J Clin Oncol. 32 (Suppl):S66–S81. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mandal BK and Suzuki KT: Arsenic round the
world: A review. Talanta. 58:201–235. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Calatayud M, Devesa V and Vélez D:
Differential toxicity and gene expression in Caco-2 cells exposed
to arsenic species. Toxicol Lett. 218:70–80. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shen ZX, Chen GQ, Ni JH, Li XS, Xiong SM,
Qiu QY, Zhu J, Tang W, Sun GL, Yang KQ, et al: Use of arsenic
trioxide (As2O3) in the treatment of acute
promyelocytic leukemia (APL): II. Clinical efficacy and
pharmacokinetics in relapsed patients. Blood. 89:3354–3360. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yedjou C, Tchounwou P, Jenkins J and
McMurray R: Basic mechanisms of arsenic trioxide (ATO)-induced
apoptosis in human leukemia (HL-60) cells. J Hematol Oncol.
3:282010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu Q, Hilsenbeck S and Gazitt Y: Arsenic
trioxide-induced apoptosis in myeloma cells: p53-dependent G1 or
G2/M cell cycle arrest, activation of caspase-8 or caspase-9, and
synergy with APO2/TRAIL. Blood. 101:4078–4087. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mandegary A, Torshabi M, Seyedabadi M,
Amirheidari B, Sharif E and Ghahremani MH: Indomethacin-enhanced
anticancer effect of arsenic trioxide in A549 cell line:
Involvement of apoptosis and phospho-ERK and p38 MAPK pathways.
Biomed Res Int. 2013:2375432013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang HY, Zhang B, Zhou JN, Wang DX, Xu YC,
Zeng Q, Jia YL, Xi JF, Nan X, He LJ, et al: Arsenic trioxide
inhibits liver cancer stem cells and metastasis by targeting
SRF/MCM7 complex. Cell Death Dis. 10:4532019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim EY, Lee SS, Shin JH, Kim SH, Shin DH
and Baek SY: Anticancer effect of arsenic trioxide on
cholangiocarcinoma: In vitro experiments and in vivo xenograft
mouse model. Clin Exp Med. 14:215–224. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mu YF, Chen YH, Chang MM, Chen YC and
Huang BM: Arsenic compounds induce apoptosis through caspase
pathway activation in MA-10 Leydig tumor cells. Oncol Lett.
18:944–954. 2019.PubMed/NCBI
|
|
16
|
Byun JM, Lee DS, Landen CN, Kim DH, Kim
YN, Lee KB, Sung MS, Park SG and Jeong DH: Arsenic trioxide and
tetraarsenic oxide induce cytotoxicity and have a synergistic
effect with cisplatin in paclitaxel-resistant ovarian cancer cells.
Acta Oncol. 58:1594–1602. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim MJ, Jung JH, Lee WS, Yun JW, Lu JN, Yi
SM, Kim HJ, Chang SH, Kim GS, Hong SC and Ha WS: Arsenic hexoxide
enhances TNF-α-induced anticancer effects by inhibiting NF-κB
activity at a safe dose in MCF-7 human breast cancer cells. Oncol
Rep. 31:2305–2311. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nagappan A, Lee WS, Yun JW, Lu JN, Chang
SH, Jeong JH, Kim GS, Jung JM and Hong SC: Tetraarsenic hexoxide
induces G2/M arrest, apoptosis, and autophagy via PI3K/Akt
suppression and p38 MAPK activation in SW620 human colon cancer
cells. PLoS One. 12:e01745912017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Mann KK, Wallner B, Lossos IS and Miller
WH Jr: Darinaparsin: A novel organic arsenical with promising
anticancer activity. Expert Opin Investig Drugs. 18:1727–1734.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cheng X, Quintás-Cardama A, Golemovic M,
Zingaro R, Gao MZ, Freireich EJ, Andreeff M, Kantarjian HM and
Verstovsek S: The organic arsenic derivative GMZ27 induces
PML-RARα-independent apoptosis in myeloid leukemia cells.
Anticancer Res. 32:2871–2880. 2012.PubMed/NCBI
|
|
21
|
Kasibhatla S and Tseng B: Why target
apoptosis in cancer treatment? Mol Cancer Ther. 2:573–580.
2003.PubMed/NCBI
|
|
22
|
D'Arcy MS: Cell death: A review of the
major forms of apoptosis, necrosis and autophagy. Cell Biol Int.
43:582–592. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ashkenazi A and Dixit VM: Death receptors:
Signaling and modulation. Science. 281:1305–1308. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Akao Y, Nakagawa Y and Akiyama K: Arsenic
trioxide induces apoptosis in neuroblastoma cell lines through the
activation of caspase 3 in vitro. FEBS Lett. 455:59–62. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chen GQ, Zhu J, Shi XG, Ni JN, Zhong HJ,
Si GY, Jin XL, Tang W, Li XS, Xong SM, et al: In vitro studies on
cellular and molecular mechanisms of arsenic trioxide
(As2O3) in the treatment of acute
promyelocytic leukemia: As2O3 induces NB4
cell apoptosis with downregulation of Bcl-2 expression and
modulation of PML-RAR/PML proteins. Blood. 88:1052–1061. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
You Z, Liu SP, Du J, Wu YH and Zhang SZ:
Advancements in MAPK signaling pathways and MAPK-targeted therapies
for ameloblastoma: A review. J Oral Pathol Med. 48:201–205. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wada T and Penninger JM: Mitogen-activated
protein kinases in apoptosis regulation. Oncogene. 23:2838–2849.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hayakawa J, Ohmichi M, Kurachi H, Ikegami
H, Kimura A, Matsuoka T, Jikihara H, Mercola D and Murata Y:
Inhibition of extracellular signal-regulated protein kinase or
c-Jun N-terminal protein kinase cascade, differentially activated
by cisplatin, sensitizes human ovarian cancer cell line. J Biol
Chem. 274:31648–31654. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kang YH and Lee SJ: The role of p38 MAPK
and JNK in Arsenic trioxide-induced mitochondrial cell death in
human cervical cancer cells. J Cell Physiol. 217:23–33. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang CY and Meng CL: Regulation of PG
synthase by EGF and PDGF in human oral, breast, stomach, and
fibrosarcoma cancer cell lines. J Dent Res. 73:1407–1415. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wong DY, Chang KW, Chen CF and Chang RC:
Characterization of two new cell lines derived from oral cavity
human squamous cell carcinomas-OC1 and OC2. J Oral Maxillofac Surg.
48:385–390. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu WC, Hsiao JR, Lian YY, Lin CY and Huang
BM: The apoptotic effect of cordycepin on human OEC-M1 oral cancer
cell line. Cancer Chemother Pharmacol. 60:103–111. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kang FC, Wang SC, So EC, Chang MM, Wong
KL, Cheng KS, Chen YC and Huang BM: Propofol may increase caspase
and MAPK pathways, and suppress the Akt pathway to induce apoptosis
in MA-10 mouse Leydig tumor cells. Oncol Rep. 41:3565–3574.
2019.PubMed/NCBI
|
|
34
|
Kang FC, Chen YC, Wang SC, So EC and Huang
BM: Propofol induces apoptosis by activating caspases and the MAPK
pathways, and inhibiting the Akt pathway in TM3 mouse Leydig
stem/progenitor cells. Int J Mol Med. 46:439–448. 2020.PubMed/NCBI
|
|
35
|
Lowry OH, Rosebrough NJ, Farr AL and
Randall RJ: Protein measurement with the Folin phenol reagent. J
Biol Chem. 193:265–275. 1951.PubMed/NCBI
|
|
36
|
Chang MM, Lai MS, Hong SY, Pan BS, Huang
H, Yang SH, Wu CC, Sun HS, Chuang JI, Wang CY and Huang BM:
FGF9/FGFR2 increase cell proliferation by activating ERK1/2,
Rb/E2F1, and cell cycle pathways in mouse Leydig tumor cells.
Cancer Sci. 109:3503–3518. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chang MM, Pan BS, Wang CY and Huang BM:
Cordycepin-induced unfolded protein response-dependent cell death,
and AKT/MAPK-mediated drug resistance in mouse testicular tumor
cells. Cancer Med. 8:3949–3964. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
van Engeland M, Ramaekers FC, Schutte B
and Reutelingsperger CP: A novel assay to measure loss of plasma
membrane asymmetry during apoptosis of adherent cells in culture.
Cytometry. 24:131–139. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li W and Chou IN: Effects of sodium
arsenite on the cytoskeleton and cellular glutathione levels in
cultured cells. Toxicol Appl Pharmacol. 114:132–139. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ochi T, Nakajimar F and Fukumori N:
Different effects of inorganic and dimethylated arsenic compounds
on cell morphology, cytoskeletal organization, and DNA synthesis in
cultured Chinese hamster V79 cells. Arch Toxicol. 72:566–573. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zuk A, Targosz-Korecka M and Szymonski M:
Effect of selected drugs used in asthma treatment on morphology and
elastic properties of red blood cells. Int J Nanomedicine.
6:249–57. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Vahter M: Methylation of inorganic arsenic
in different mammalian species and population groups. Sci Prog.
82:69–88. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lin KW, Behl S, Furst A, Chien P and Toia
RF: Formation of dimethylarsinic acid from methylation of sodium
arsenite in lumbricus terrestris. Toxicol In Vitro. 12:197–199.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hartwell LH and Weinert TA: Checkpoints:
Controls that ensure the order of cell cycle events. Science.
246:629–634. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yih LH, Wu YC, Hsu NC and Kuo HH: Arsenic
trioxide induces abnormal mitotic spindles through a PIP4KIIγ/Rho
pathway. Toxicol Sci. 128:115–125. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ling YH, Jiang JD, Holland JF and
Perez-Soler R: Arsenic trioxide produces polymerization of
microtubules and mitotic arrest before apoptosis in human tumor
cell lines. Mol Pharmacol. 62:529–538. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Concin N, Stimpfl M, Zeillinger C, Wolff
U, Hefler L, Sedlak J, Leodolter S and Zeillinger R: Role of p53 in
G2/M cell cycle arrest and apoptosis in response to
gamma-irradiation in ovarian carcinoma cell lines. Int J Oncol.
22:51–57. 2003.PubMed/NCBI
|
|
48
|
Shu CH, Yang WK, Shih YL, Kuo ML and Huang
TS: Cell cycle G2/M arrest and activation of cyclin-dependent
kinases associated with low-dose paclitaxel-induced sub-G1
apoptosis. Apoptosis. 2:463–470. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tyagi AK, Singh RP, Agarwal C, Chan DC and
Agarwal R: Silibinin strongly synergizes human prostate carcinoma
DU145 cells to doxorubicin-induced growth Inhibition, G2-M arrest,
and apoptosis. Clin Cancer Res. 8:3512–3519. 2002.PubMed/NCBI
|
|
50
|
Brown JM and Attardi LD: The role of
apoptosis in cancer development and treatment response. Nat Rev
Cancer. 5:231–237. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Fan TJ, Han LH, Cong RS and Liang J:
Caspase family proteases and apoptosis. Acta Biochim Biophys Sin
(Shanghai). 37:719–727. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Cheng B, Yang X, Han Z, An L and Liu S:
Arsenic trioxide induced the apoptosis of laryngeal cancer via
down-regulation of survivin mRNA. Auris Nasus Larynx. 35:95–101.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Jiang XH, Wong BC, Yuen ST, Jiang SH, Cho
CH, Lai KC, Lin MC, Kung HF and Lam SK: Arsenic trioxide induces
apoptosis in human gastric cancer cells through up-regulation of
p53 and activation ofcaspase-3. Int J Cancer. 91:173–179. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wu X, Shi J, Wu Y, Tao Y, Hou J, Meng X,
Hu X, Han Y, Jiang W, Tang S, et al: Arsenic trioxide-mediated
growth inhibition of myeloma cells is associated with an extrinsic
or intrinsic signaling pathway through activation of TRAIL or TRAIL
receptor 2. Cancer Biol Ther. 10:1201–1214. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Binet F, Chiasson S and Girard D: Evidence
that endoplasmic reticulum (ER) stress and caspase-4 activation
occur in human neutrophils. Biochem Biophys Res Commun. 391:18–23.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lunghi P, Giuliani N, Mazzera L, Lombardi
G, Ricca M, Corradi A, Cantoni AM, Salvatore L, Riccioni R,
Costanzo A, et al: Targeting MEK/MAPK signal transduction module
potentiates ATO-induced apoptosis in multiple myeloma cells through
multiple signaling pathways. Blood. 112:2450–2462. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kircelli F, Akay C and Gazitt Y: Arsenic
trioxide induces p53-dependent apoptotic signals in myeloma cells
with SiRNA-silenced p53: MAP kinase pathway is preferentially
activated in cells expressing inactivated p53. Int J Oncol.
30:993–1001. 2007.PubMed/NCBI
|
|
58
|
Lam HK, Li K, Chik KW, Yang M, Liu VC, Li
CK, Fok TF, Ng PC, Shing MM, Chuen CK and Yuen PM: Arsenic trioxide
mediates intrinsic and extrinsic pathways of apoptosis and cell
cycle arrest in acute megakaryocytic leukemia. Int J Oncol.
27:537–545. 2005.PubMed/NCBI
|
|
59
|
Ivanov VN and Hei TK: Regulation of
apoptosis in human melanoma and neuroblastoma cells by statins,
sodium arsenite and TRAIL: A role of combined treatment versus
monotherapy. Apoptosis. 16:1268–1284. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Shen L and Zhang G, Lou Z, Xu G and Zhang
G: Cryptotanshinone enhances the effect of Arsenic trioxide in
treating liver cancer cell by inducing apoptosis through
downregulating phosphorylated- STAT3 in vitro and in vivo. BMC
Complement Altern Med. 17:1062017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Wang X and Zhang M: Synergistic effects of
valproic acid and arsenic trioxide on RPMI8226 cells in
vitro and the possible underlying mechanisms. Mol Med Rep.
12:1449–1456. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Amrán D, Sánchez Y, Fernández C, Ramos AM,
de Blas E, Bréard J, Calle C and Aller P: Arsenic trioxide
sensitizes promonocytic leukemia cells to TNFalpha-induced
apoptosis via p38-MAPK-regulated activation of both
receptor-mediated and mitochondrial pathways. Biochim Biophys Acta.
1773:1653–1663. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Alvarado-Kristensson M, Melander F,
Leandersson K, Rönnstrand L, Wernstedt C and Andersson T: p38-MAPK
signals survival by phosphorylation of caspase-8 and caspase-3 in
human neutrophils. J Exp Med. 199:449–458. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Grethe S, Ares MP, Andersson T and
Pörn-Ares MI: p38 MAPK mediates TNF-induced apoptosis in
endothelial cells via phosphorylation and downregulation of
Bcl-x(L). Exp Cell Res. 298:632–642. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Choi WS, Eom DS, Han BS, Kim WK, Han BH,
Choi EJ, Oh TH, Markelonis GJ, Cho JW and Oh YJ: Phosphorylation of
p38 MAPK induced by oxidative stress is linked to activation of
both caspase-8- and −9-mediated apoptotic pathways in dopaminergic
neurons. J Biol Chem. 279:20451–2060. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Falchi L, Verstovsek S, Ravandi-Kashani F
and Kantarjian HM: The evolution of arsenic in the treatment of
acute promyelocytic leukemia and other myeloid neoplasms: Moving
toward an effective oral, outpatient therapy. Cancer.
122:1160–1168. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Torka P, Al Ustwani O, Wetzler M, Wang ES
and Griffiths EA: Swallowing a bitter pill-oral arsenic trioxide
for acute promyelocytic leukemia. Blood Rev. 30:201–211. 2016.
View Article : Google Scholar : PubMed/NCBI
|














