Introduction
Despite substantial investment and years of research
worldwide, pancreatic cancer remains one of the most lethal
malignancies with very high mortality rates. For all stages
combined, the 1- and 5-year relative survival rates are 27 and 7%,
respectively, making it the only type of cancer with an overall
5-year survival rate in the single digits (1). There are currently no effective
medications for the treatment of patients with locally advanced or
metastatic pancreatic cancer who are ineligible for surgery. The
stage at which pancreatic cancer is diagnosed is strongly
correlated with overall survival, and the delayed diagnosis results
in only 20% of patients being eligible for surgery or adjuvant
therapy (2). Therefore, developing
novel and effective drugs is urgently required for this terrible
malignancy.
The discovery of new drugs has grown increasingly
difficult over the years, as itis a time-consuming, high-investment
and high-risk process in traditional drug development. Therefore,
drug repurposing has become an increasingly used strategy for
identifying novel medications or new uses for approved drugs that
have already been optimized for safety and efficacy (3). In the past decade, various
high-throughput strategies and experimental platforms have been
used for the discovery and identification of repurposable drug
candidates, with statins previously identified as potential
promising antitumor agents (4).
Statins, also known as 3-hydroxy-3-methyglutaryl coenzyme A
reductase inhibitors, are some of the most commonly prescribed
medications for the treatment of hypercholesterolemia, due to their
ability to inhibit de novo cholesterol synthesis, thereby
reducing the risk of heart attacks and other major clinical
manifestations of coronary artery diseases by up to 37% (5). In 1987, lovastatin (Mevacor, Altocor
and Altoprev) became the first FDA-approved statin. Since then,
further statins have been derived and synthesized, including
simvastatin (Zocor), atorvastatin (Lipitor and Lipex) and
fluvastatin (Lescol). In general, statins are safe and
well-tolerated by patients, and they all work in a similar way.
In addition to their powerful cholesterol-lowing
effects, statins also exert several other cholesterol-independent
effects (pleiotropic effects), including improved endothelial
function, stabilization of atherosclerotic plaque and
anti-inflammatory, antioxidant, anti-proliferative and
immunomodulatory effects (6).
Notably, there has been growing interest in the potential antitumor
effects of multiple statins based on accumulating evidence of their
anti-proliferation and anti-inflammatory activities in various
types of cancer in a number of preclinical, as well as clinical
studies (7). To date, research has
indicated that statins may inhibit tumor growth and metastasis
through a series of cellular mechanisms, including the promotion of
apoptosis, inhibition of cell cycle progression and proliferation,
reduction of angiogenesis, and interference with carcinogenesis.
For example, lovastatin treatment could cause breast cancer cell
death through the LKB1-AMPK-p38MAPK-p53-survivin signaling cascade
(8). Recently, simvastatin was
reported to attenuate macrophage-mediated gemcitabine resistance of
pancreatic cancer by regulating the TGF-β1/Gfi-1 axis (9). It has also been shown that the
suppression of cholesterol biosynthesis by lovastatin significantly
inhibited gallbladder cancer cell proliferation, possibly through
the inhibition of DNA repair. However, whether these various
statins exert their antitumor function through a consistent
mechanism remains largely unknown.
In the present study, a previously established
reporter system was used to screen potential drugs for the
treatment of pancreatic cancer from the Prestwick Chemical Library
(10,11). Three statins were identified:
Fluvastatin, lovastatin and simvastatin. Furthermore, low-(2 µM)
and high-concentration (20 µM) treatment with these three statins
was used to determine their impact on the global transcriptional
activity, as well as their concentration-dependent effect in two
human pancreatic cancer cell lines, MiaPaCa2 and PANC1. The aim of
the present study was to elucidate the unique and shared functions
of these three statins, and thereby gain some insights of
theoretical guidance for the potential use of these statins in the
future clinical treatment of pancreatic cancer.
Materials and methods
Materials
In all experiments, pure forms (≥98.0%) of
fluvastatin, lovastatin and simvastatin (Tocris Bioscience) were
used. All statins were dissolved in DMSO and tested in final
concentrations of 2 and 20 µM.
Cell culture
Human pancreatic cancer MiaPaCa2 and PANC1 cell
lines (American Type Culture Collection) were maintained in DMEM
supplemented with 2 mM glutamine, 1 mM Na-pyruvate, 100 units/ml
penicillin, 100 µg/ml streptomycin and 10% fetal bovine serum (all
from Gibco, Thermo Fisher Scientific, Inc.) at 37°C in a humidified
atmosphere containing 10% CO2.
Medium throughput screening of the
Prestwick Chemical Library on pancreatic cancer
For preliminary screening purposes, the pancreatic
cancer cell line MiaPaCa2bearing stable expressing BIRC5-GLuc was
first arrayed in 384-well plates and treated with test compounds at
20 µM for 48 h under serum-free conditions. Changes to the
luciferase expression and cell viability were quantitated using the
CellTiter-Glo® Luminescent Cell Viability Assay. For
more information, the 1,280 chemically and pharmacologically
diverse compounds (90% of which were FDA-approved drugs) from the
Prestwick Chemical Library were screened against the pancreatic
cancer MiaPaCa2 cell line. Experiments were conducted in 384-well
plates. A fully automated Hamilton Star workstation was used for
all liquid handling protocols. Compounds were loaded into black
F-bottom 384-well assay ready plates (Greiner Bio-One), followed by
200 µl of pBIRC5-eGFP, resulting in a final drug concentration of
20 µM with <0.002% DMSO in the primary screen. Assay plates were
cultured for 48 h in an incubator at 37°C in a humidified
atmosphere containing 10% CO2.
Cell viability assay
The MiaPaCa2cells were seeded in 384-well plates at
a density of 1×103 cells/well in 100 µl medium. A total
of 24 h after seeding, the cells were treated with the indicated
concentrations of statins (0–100 µM) for 48 h. Cell viability was
determined by CellTiter-Glo® Luminescent Cell Viability
Assay (Promega Corporation), according to the manufacturer's
instructions. Briefly, at the end of incubation, the culture
supernatant was replaced with CellTiter-Glo reagent (1:20 v/v
dilution in fresh culture medium) (Promega Corporation). Plates
were incubated at 37°C for 15 min and absorbance was measured at
490 nm using an Omega microplate reader (IMGEN Technologies).
Drug effect assay
The drug effect was determined by Cell Counting
Kit-8 (CCK-8) assay. Briefly, 3,000 cells/well were seeded into a
96-well plate and incubated overnight in a cell culture incubator
(37°C, 95% humidity). Cells were then treated with different
concentrations (2 and 20 µM) of statins. Following incubation, the
medium in each well was replaced with fresh culture medium
containing 10 µl CCK-8 (Dojindo Molecular Technologies, Inc.). The
plates were incubated for an additional 2 h in a cell culture
incubator (37°C, 95% humidity), and absorbance was determined at
450 nm using a microplate spectrophotometer.
Reverse transcription-quantitative
(RT-q)PCR
Total RNA was extracted from cells using TRIzol
reagent (Invitrogen; Life Technologies; Thermo Fisher Scientific,
Inc.) according to manufacturer's instructions. Reverse
transcription was performed with RevertAid First Strand cDNA
Synthesis kit (cat. no. K1622; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions. RT-qPCR was conducted
using SYBR Green (ABgene) according to the manufacturer's
instructions. The primer sequences used in qPCR were as follows:
Forward, 5′-CGCTGGCGGTACTGAAGTC-3′ and reverse,
5′-GAGGAACGGTGACATGCTCAT-3′ for CCNA2; forward,
5′-CGCTACCCCTCAGAGGACAA-3′ and reverse, 5′-ACAGACATCGGTG-3′ for
LZTS3; forward, 5′-CATGTACGTTGCTATCCAGGC-3′ and reverse,
5′-CTCCTTAATGTCACGCACGAT-3′ for β-actin (ACTB). The primers were
obtained from Funengbio Co., Ltd. PCR amplification was performed
using the following thermocycling conditions: 95°C for 30 sec,
followed by 40 cycles at 95°C for 5 sec, 60°C for 34 sec, and a
melt curve stage. All experiments were performed in triplicate. The
relative gene expression level was calculated using the
2−ΔΔCq method (12), and
ACTB was used as the internal control.
Effects of statins on cancer cell
colony formation
In each 6-well plate, 400 cells were seeded and
cultured for 24 h. The culture medium was then changed to 5, 10,
15, 20 and 25 µM statin-containing medium, and cells were incubated
for 9 days, with DMSO as the control. The cells were washed with
PBS and fixed with 100% methanol for 10 min at room temperature.
Crystal violet (1%) staining buffer (cat. no. C0121; Beyotime
Institute of Biotechnology) was then added into the plates.
Following incubation for 30 min at 37°C, the crystal violet
staining buffer was removed from the plates, which were then washed
with de-ionized water. After the plates were dried at room
temperature, the colonies in each plate were counted to measure the
colony formation rate. The colonies were visualized with a general
camera. Only colonies containing ≥50 cells were counted as
clonogenic survivors.
RNA-seq for statin-treated pancreatic
cancer cells
PANC1 and MiaPaCa2 cells were collected following
treatment or no treatment with 2 and 20 µM of the three statins for
48 h (3 replicates per sample). Total RNAs were extracted using the
RNeasy Mini kit (Qiagen AB), and the quality of the RNA was
evaluated using Agilent Bioanalyzer 2100 (Agilent Technologies,
Inc.). Sequence libraries were prepared using a TruSeq Stranded
mRNA Library Prep kit for NeoPrep (Illumina, Inc.), according to
the manufacturer's instructions, and sequenced using an Illumina
HiSeq 2000 platform. FastQC (https://github.com/s-andrews/FastQC) was used for
quality control of the sequenced data (13). RNA-seq data were trimmed using
Trimmomatic (http://www.usadellab.org/cms/?page=trimmomatic) to
remove and filter low-quality sequencing data and the adapters
(14). The human genome NCBI GRCh38
(15) and its corresponding
transcriptome gene annotation was used for read alignment. The
TopHat alignment tool was used for alignment with default parameter
settings (16).
Construction of weighted gene
co-expression network analysis (WGCNA)
WGCNA was used for scale-free network topology
analysis of RNA-seq data (17). The
WGCNA R package was used to cluster highly correlated genes and
determine clusters in which gene expression was associated with the
examined characteristics. An adjacency matrix based on expression
correlation was created using a soft threshold procedure to allow
scale-free topology. The clusters created by WGCNA were referred to
as modules, and the minimum number of genes in a module was set to
30. The functional annotation tool Database for Annotation,
Visualization and Integrated Discovery (DAVID) Bioinformatics
Resources 6.8 was used to determine Gene Ontology terms enriched by
the identified genes. DAVID analyses were performed using genes
corresponding to significant WGCNA modules.
DAVID analysis
DAVID (http://david.abcc.ncifcrf.gov) Functional Annotation
Bioinformatics Microarray Analysis was used to identify
significantly enriched Gene Ontology (GO) and Kyoto Encyclopedia of
Genes and Genomes (KEGG) terms among the genes that were
differentially expressed in control samples (18,19).
Statistically overrepresented GO and KEGG categories with a P≤0.05
were considered significant.
Survival and ROC analysis
To explore the potential prognostic value of the
hub-gene, the Gene Expression Profiling Interactive Analysis
(GEPIA) database was used to perform overall and disease-free
survival analysis, and the log-rank tests were used to measure
statistical significance (20).
Statistical analysis
All experiments in the present study were
independently performed in triplicate. Data are presented as the
mean ± SEM. All graphs were plotted and analyzed with GraphPad
Prism Software (GraphPad Prism version 7.1 for Windows; GraphPad
Software, Inc.) and one-way ANOVA followed by Dunnett's multiple
comparisons test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Statins identified through medium
throughput screening of pancreatic cancer cell lines from the
Prestwick Chemical Library
The survivin gene, also known as BIRC5, plays a
central role in cancer cell survival and proliferation, and has
therefore become a promising therapeutic target for pancreatic
cancer vaccines and therapeutics (21). In order to discover small molecular
compounds that could suppress BIRC5 gene expression and pancreatic
cancer cell proliferation, a high-content immunofluorescence
screening assay utilizing an enhanced BIRC5 super-promoter system
to drive the expression of dual Gaussia luciferase (GLuc)
and sr39 thymidine kinase (sr39TK) reporter genes was developed
(22). The screening incorporated
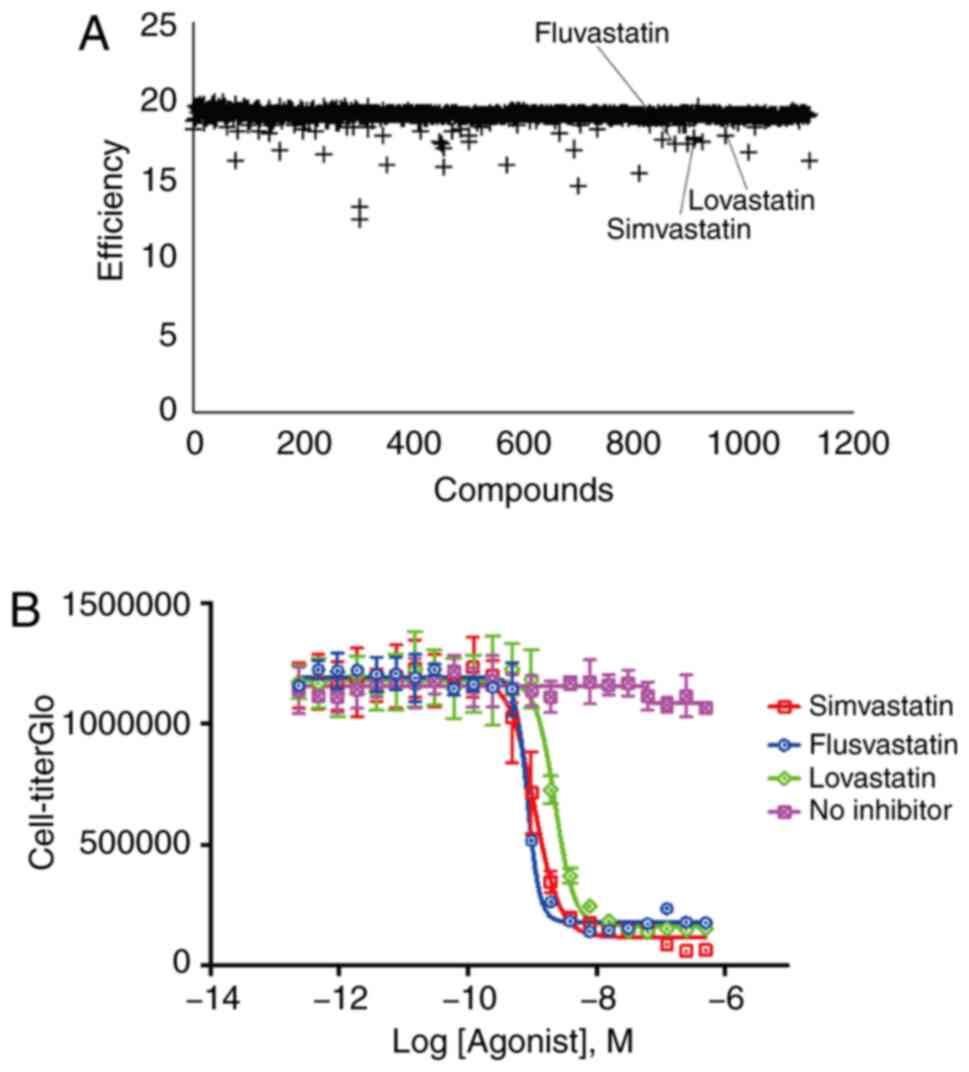
1,280 compounds from the Prestwick Chemical library. Of all tested
compounds, three statins (lovastatin, simvastatin and fluvastatin)
were successfully identified to be consistently and potently
effective at blocking BIRC5-induced Gaussia luciferase
expression (Fig. 1A and B). These
results indicated that all three statins identified through medium
throughput screening could functionally suppress the cell
proliferation of pancreatic cancer cells, which may have a
potential clinical application value.
Statins significantly inhibit
pancreatic cancer cell proliferation in a concentration-dependent
manner
A previous study suggested that, although the
MiaPaCa2 and PANC1 were commonly used pancreatic cancer cell lines,
these two cell lines had a marked difference in response to various
drugs (23). To further
characterize the antitumor effects of the three statins on
pancreatic cancer, the MiaPaCa2 and PANC1cell lines were used in
the following assays. The MiaPaCa2 and PANC1 cells were incubated
with fluvastatin, lovastatin and simvastatin at either a low (2 µM)
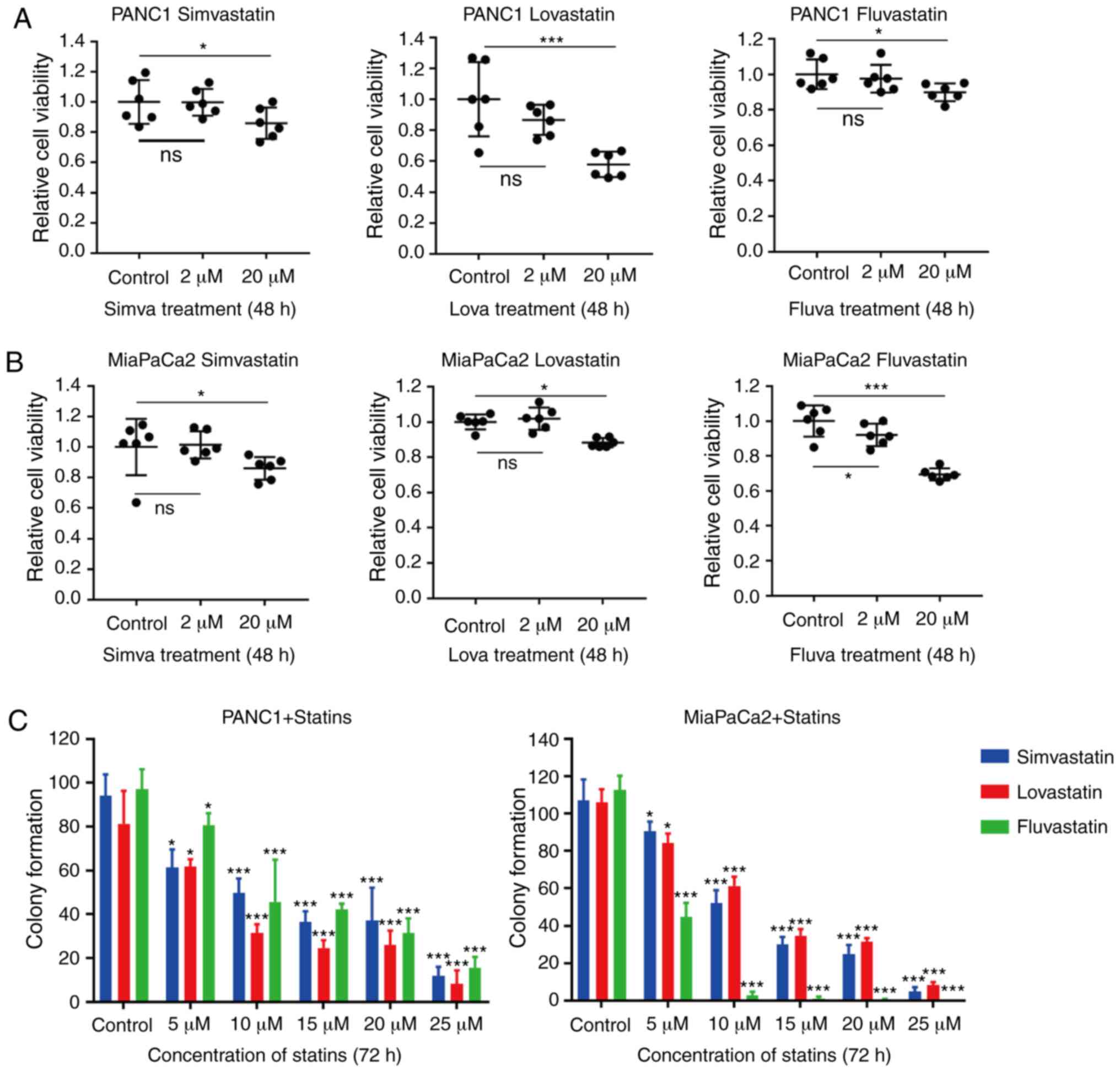
or a high (20 µM) concentration. CCK-8 assays revealed that all
three statins induced cell growth inhibition in both cell lines
following high-concentration treatment, but not with
low-concentration treatment (Fig. 2A
and B). Among the three statins, lovastatin exhibited a
relatively more efficient inhibition of PANC1 cell proliferation,
while fluvastatin exhibited a more efficient suppression of
MiaPaCa2 cell proliferation. In addition, a colony formation assay
was carried out on the MiaPaCa2 and PANC1 cells treated with
different concentrations of these three statins. The colony
formation assay revealed that 5 µM statins slightly suppressed the
proliferation of PANC1 cells (P<0.05 and P<0.01). This result
is inconsistent with Fig. 2B, which
is likely due to different concentrations of statins (5 µM vs. 2
µM) that we used. Consistent with the CCK-8 assay, the results
revealed that only at a high concentration (≥5 µM) could the three
statins significantly attenuate the proliferation of both
pancreatic cancer cell lines (Figs.
2C and S1). Notably,
lovastatin and fluvastatin had diverse inhibitory effects on
MiaPaCa2 and PANC1 cells, respectively. In conclusion, these
results revealed that the high-concentration statins significantly
inhibited the proliferation of both pancreatic cancer cell lines,
but the inhibitory efficiencies of various statins on these two
pancreatic cancer cell lines were diverse.
Identification of statin-induced
transcriptome alterations in pancreatic cancer cells through
RNA-seq
In order to investigate the transcriptional
alterations in pancreatic cancer cell lines under various treatment
conditions with the three statins, RNA-seq was performed on
MiaPaCa2 and PANC1 cells treated with either a low- or
high-concentration of the three statins. The two cancer cell lines
were treated with low-(2 µM group) and high (20 µM
group)-concentration statins for 48 h before the cells were
harvested for RNA-seq library preparation. The signature gene
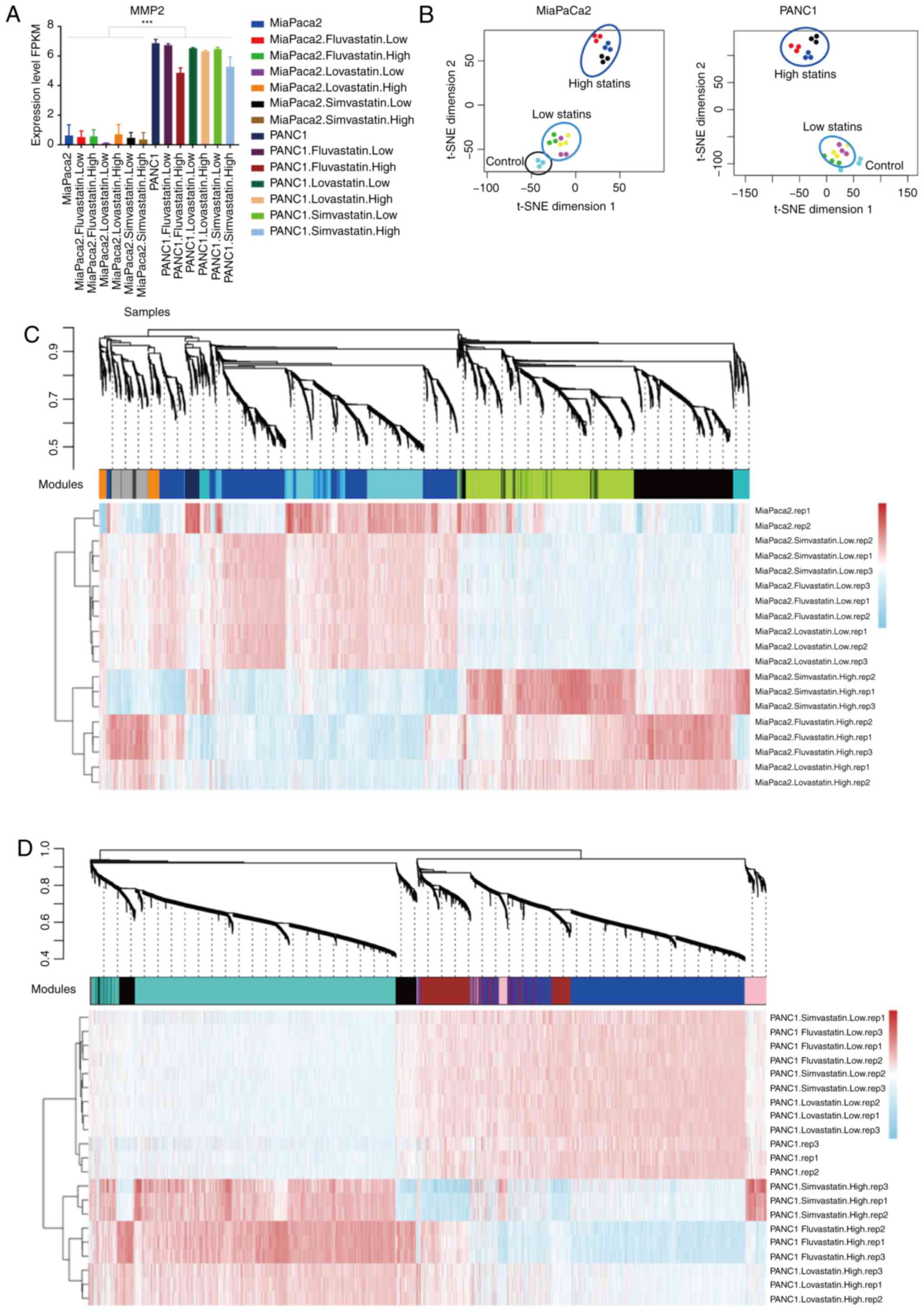
expression of MiaPaCa2 and PANC1 cells was validated by examining
the endogenous MMP2 expression of RNA-Seq data between these two
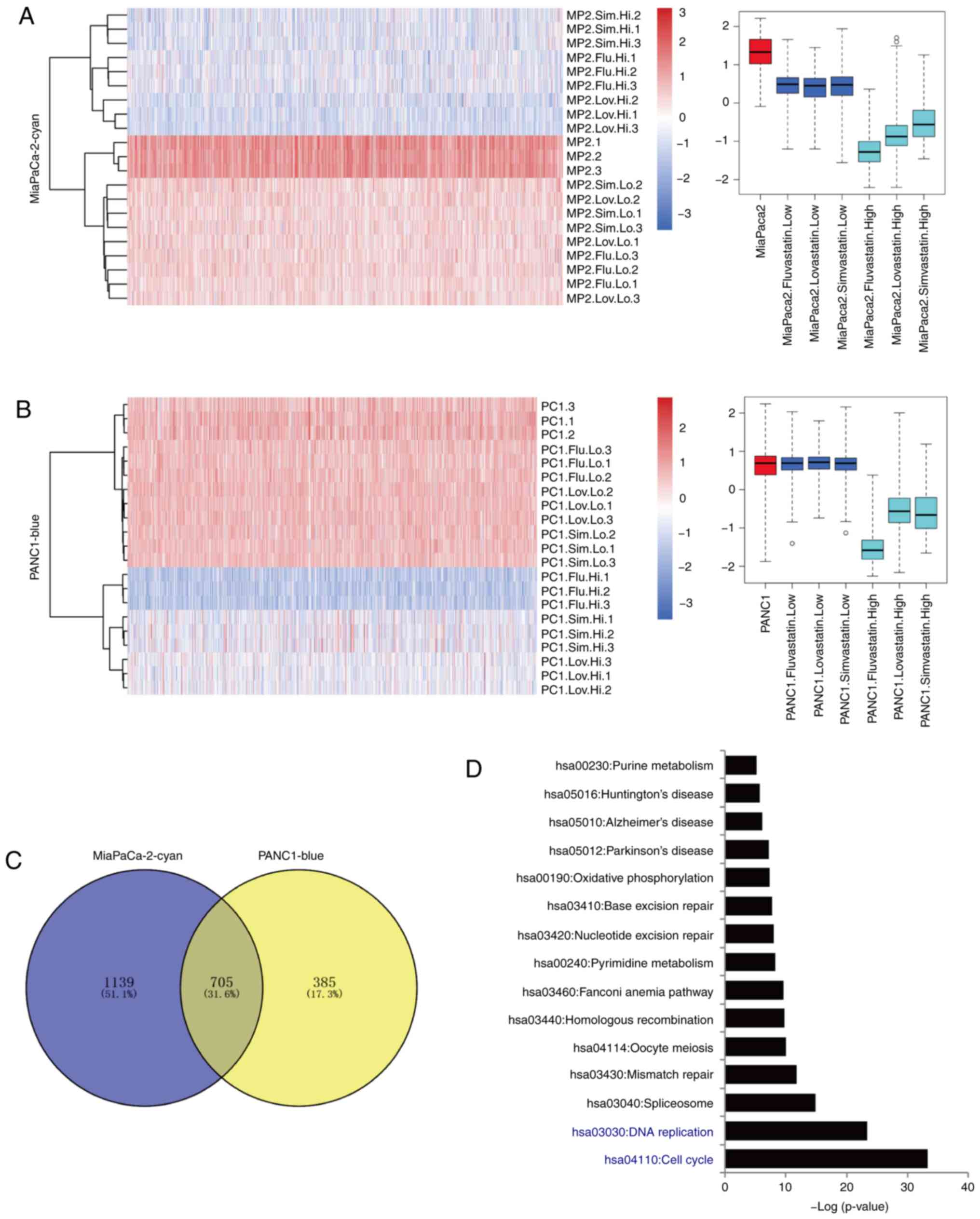
cell lines (Fig. 3A) (24). The transcriptional profiles of all
three statin-treated cancer cells were distinguished from those of
the original cancer cells. 2D t-SNE of whole genome gene expression
profiles in these treated cells revealed that the low-and
high-concentration treatment groups clustered separately (Fig. 3B). The high-concentration statins
markedly changed the expression of genes in both cell lines. On the
contrary, the low-concentration statins had relatively minor
effects. Only a subset of genes was upregulated in
low-concentration statin-treated MiaPaCa2 cells, which was not
observed in PANC1 cells (Fig. S2).
To further assess the transcriptome dynamics of statin-induced gene
alterations, the WGCNA was used, and multiple gene-network modules
associated with high- and low-concentration statins, as well as
three individual statins, were obtained (Table SI). In MiaPaCa2 cells, 8 modules
that were possibly associated with the treatment of various statins
were identified (Figs. 3C and
S3A), while 5 modules were
identified in PANC1 cells (Figs. 3D
and S3B). As compared to those
untreated cancer cells, high-concentration treatment groups
exhibited more robust gene expression alterations than that in the
low-concentration treatment groups. A high number of upregulated
and downregulated genes were identified in the high-concentration
groups, which may contribute to the cell proliferation inhibition
effects of the statins.
Low-concentration statins stimulate
considerable gene expression changes in MiaPaCa2, but not in PANC1
cells
As revealed by a previous study, the difference in
the response to various drugs between MiaPaCa2 and PANC1 was marked
(23). A significant difference
(3,028 different expression genes) was observed between the
MiaPaCa2 and PANC1 cell lines in our RNA-seq data (Fig. S4A and B). KEGG analysis of these
different expression genes revealed that the MiaPaCa2 cells
exhibited a high level of metabolism-related gene expression
(Fig. S4C), while the PANC1 cells
exhibited a high level of cancer-related gene expression (Fig. S4D). The different transcriptome
profiles of MiaPaCa2 and PANC1 suggest that they may have different
effects on statin treatment. In response to treatment with
low-concentration statins, the MiaPaCa2 cells behaved in a
considerably more sensitive way than the PANC1 cells. Similarly,
the expression of a number of genes was significantly altered in
statin-treated MiaPaCa2 cells, but not in PANC1 cells (Fig. S2). Additional WGCNA analysis also
indicated that the PANC1 cells treated with a low-concentration
statin exhibited a similar gene expression to that of original
PANC1 cells. However, 4 modules were significantly changed in
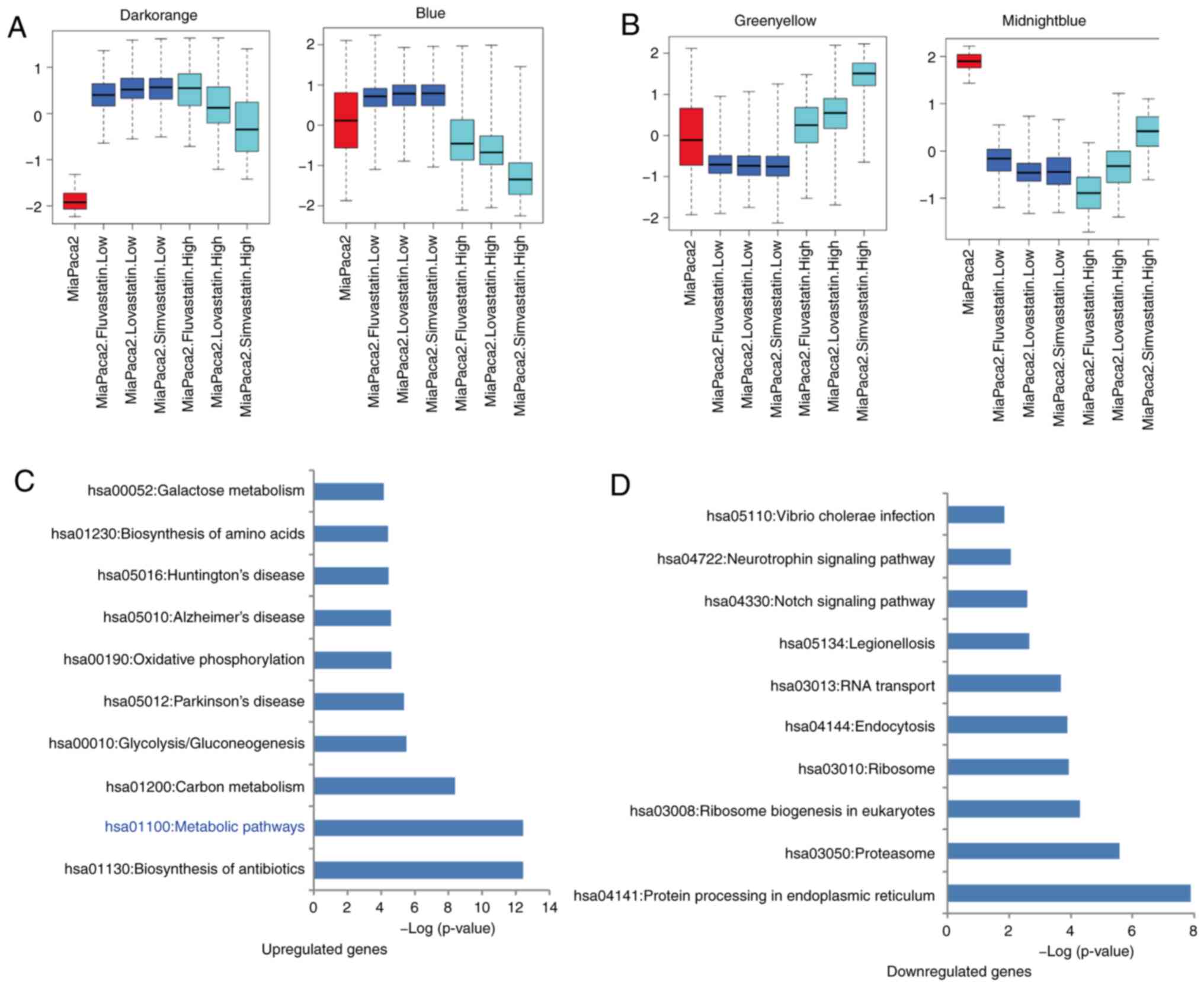
MiaPaCa2 cells. In MiaPaCa2 cells treated with low-concentration
statins, genes in the dark-orange [208 genes, module eigengene
(KME) dark-orange >0.5] and blue (1,631 genes, KME blue >0.5)
modules were significantly upregulated, while genes in
midnight-blue (158 genes, KME midnight-blue >0.5) and
green-yellow (1,575 genes, KME green-yellow >0.5) modules were
significantly downregulated (KME value=eigengene connectivity)
(Fig. 4A and B). Further DAVID GO
and KEGG analysis revealed that, upregulated genes enriched in the
signaling pathways of the biosynthesis of antibiotics (P<0.001),
metabolism (P<0.001), carbon metabolism (P<0.001) and
glycolysis/gluconeogenesis (P<0.001), while downregulated genes
were significantly enriched in the signaling pathways of the
protein processing in endoplasmic reticulum (P<0.001),
proteasome (P<0.001), ribosome biogenesis in eukaryotes
(P<0.001) and ribosome (P<0.001) (Fig. 4C and D, Table SII). In conclusion, these results
indicated that low-concentration statins did not influence the
PANC1 cells, but they affected gene expression in MiaPaCa2 cancer
cells by upregulating the metabolism-related genes and
downregulating the ribosome-related genes.
High-concentration statins
significantly suppress the proliferation of pancreatic cancer cells
by inhibiting the cyclin A2 (CCNA2)-based cell cycle pathway
In PANC1 cells treated with high-concentration
statins, the most significant variations were the gene upregulation
in the turquoise module (2,327 genes, KME turquoise >0.5) and
downregulation in the blue module (1,844 genes, KME blue >0.5;
Table SI). In MiaPaCa2 cells,
treatment with high-concentration statins caused gene upregulation
in the black (1,231 genes, KME black >0.5) and green-yellow
(1,575 genes, KME green-yellow >0.5) modules, and gene
downregulation in the cyan (1,090 genes, KME cyan >0.5) and blue
(1,631 genes, KME blue >0.5) modules (Table SI). Combined with the significant
inhibition of MiaPaCa2 cells by fluvastatin (Fig. 2B and C), the downregulated genes in
the cyan module and upregulated genes in the black module may play
a vital role on the inhibition of pancreatic cancer MiaPaCa2 cell
proliferation by statins.
In order to identify the common effects of statins
on pancreatic cancer cells, an integrated analysis of these two
cell lines was conducted. The cyan module of the MiaPaCa2 cell line
and blue module of the PANC1 cell line displayed a
concentration-dependent expression inhibition induced by all three
statins (Fig. 5A and B). The
detailed inhibition of the expression of these genes is presented.
The gene lists from these 2 modules were compared, and 705
overlapping genes that were inhibited by statins were identified
(Fig. 5C). KEGG analysis of these
genes revealed significant enrichment in the signaling pathways of
cell cycle (P<0.001), DNA replication (P<0.001) and
spliceosome (P<0.001) (Fig. 5D).
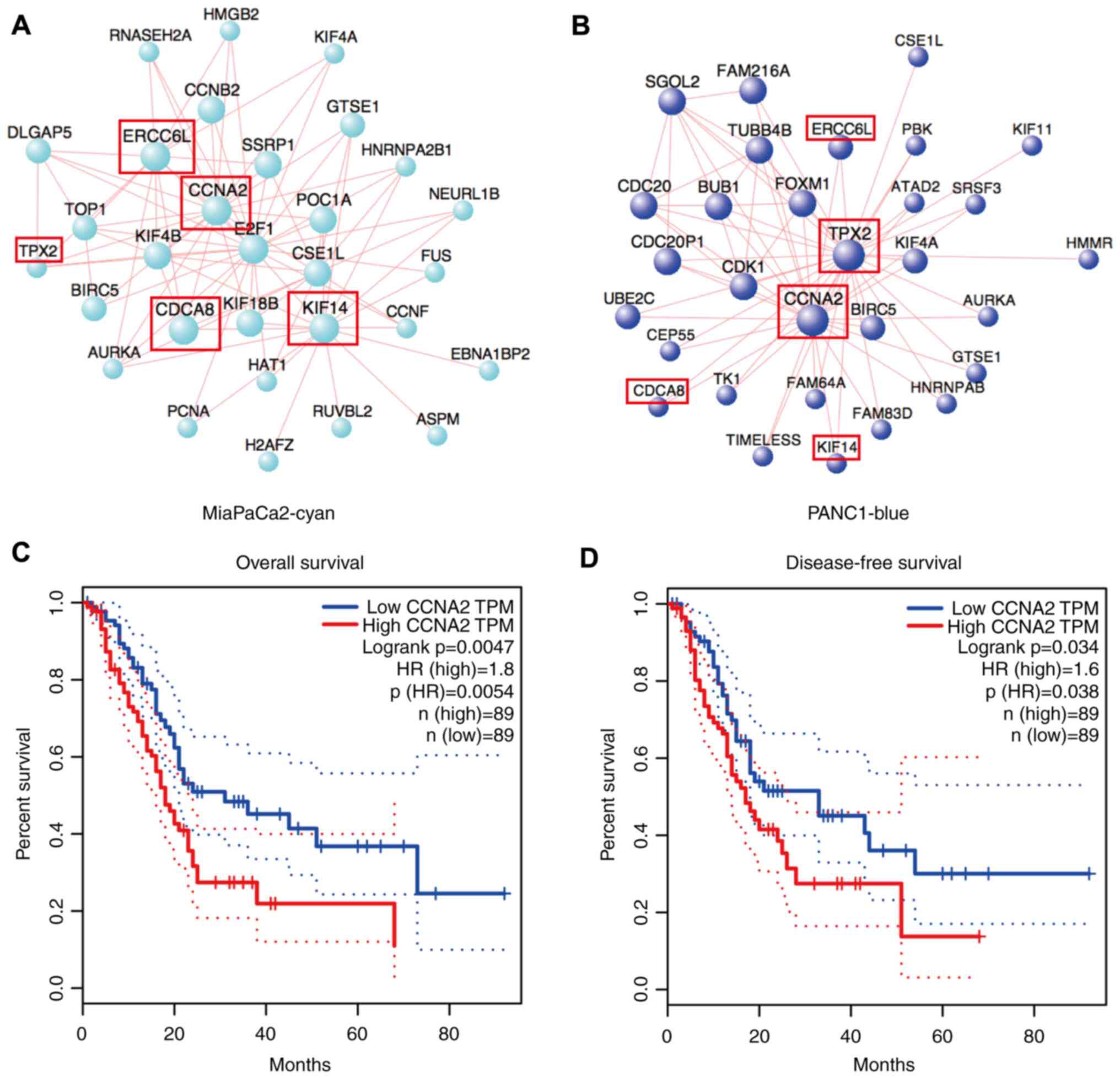
Further hub-gene analysis of these 2 modules was performed. The
result revealed that they shared some common genes that were also
involved in the cell cycle pathway, such as CCNA2, ERCC6L, TPX2,
KIF14 andCDCA8. In these two modules, CCNA2 was
the common main affecting gene (Fig. 6A
and B), while in pancreatic cancer, the high expression level
of CCNA2 was significantly associated with poor prognosis,
including overall (P=0.0047) and disease-free survival (P=0.034;
Fig. 6C and D). The suppressed
CCNA2 by high-concentration statins were also validated with
RT-qPCR (Fig. S5A-C). These data
demonstrated the inhibition of a common conserved gene by statins
in pancreatic cancer cells, and indicated that statins potentially
inhibited cancer cell proliferation through the suppression of the
cell cycle pathway, based on the core gene CCNA2.
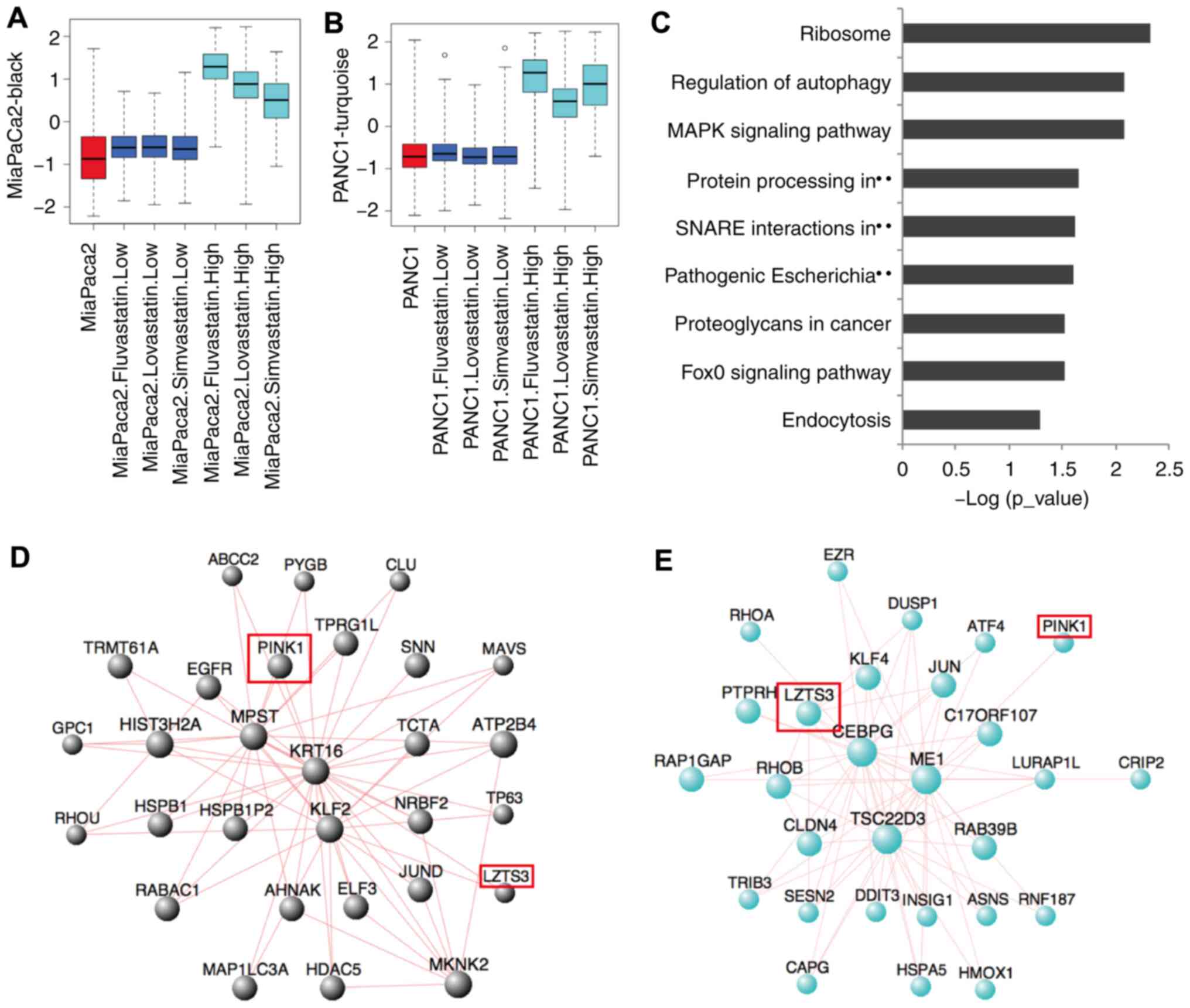
High-concentration statins
significantly upregulate the genes involved in ribosome and
autophagy pathways
These statins could also induce high gene expression
in pancreatic cancer cells. In the black module (1,297 genes;
Figs. 7A and S6A) of the MiaPaCa2 cell line and
turquoise module (2,519 genes; Figs.
7B and S6B) of the PANC1 cell
line, a markedly high gene expression of 635 genes was induced by
high-concentration treatment with all three statins (Fig. S6C). KEGG analysis of these genes
revealed significant enrichment in the ribosome (P<0.001),
regulation of autophagy (P<0.001) and MAPK signaling
(P<0.001; Fig. 7C) pathways.
Further hub-gene analysis of these 2 modules revealed that the
MiaPaCa2 and PANC1 cell lines have different core genes for
statin-induced pathway activation. In the black module of the
MiaPaCa2 cell line, the PTEN induced kinase 1 (PINK1), MPST,
KRT16, KLF2 and HIST3H2A genes form the core gene group
(Fig. 7D), while in the turquoise
module of the PANC1 cell line, the leucine zipper tumor suppressor
family member 3 (LZTS3), CEBPG, ME1 and TSC22D3 genes
form the core gene group (Fig. 7E).
Both PINK1 and LZTS3 appeared in the hub genes of
these two modules, but they have different contributions. The
induced expression of LZTS3 by high-concentration statins were
validated with RT-qPCR (Fig.
S5D-F). These results indicated that high-concentration statins
could induce massive gene expression changes in both pancreatic
cancer cells, and although the hub genes of different pancreatic
cancer cells were slightly different, their enriched signaling
pathways were identical. Both were involved in the ribosome,
autophagy and MAPK signaling pathways.
In addition to the aforementioned common effects,
these high-concentration statins could also induce the expression
of different genes in the MiaPaCa2 and PANC1 cell lines (Fig. 3C and D). In MiaPaCa2 cells, genes
from the dark-grey module were significantly upregulated in
fluvastatin-treated cells. KEGG analysis of these genes revealed a
significant enrichment in the pathways of oxidative phosphorylation
and ribosome (P<0.001; Table
SIII). Genes in the dark-turquoise module were significantly
upregulated in simvastatin-treated cells. KEGG analysis of these
genes revealed significant enrichment in the pathways of the NF-κB
signaling pathway (P=0.007; Table
SIII). In PANC1 cells, genes in the pink module were
significantly upregulated in simvastatin-treated cells. KEGG
analysis of these genes revealed that the signaling pathways of the
oxidative phosphorylation and ribosome were significantly enriched
(P<0.001), which was consistent with the dark-grey module in
MiaPaCa2 cells (Table SIII).
However, fluvastatin-treated PANC1 cells specifically upregulated
the genes in the black module, which was enriched in the signaling
pathways of the protein processing in endoplasmic reticulum and
endocytosis (P<0.001). These results indicated that, in addition
to common transcriptional influences, these three statins exhibited
different effects on the signaling pathways of the MiaPaCa2 and
PANC1 cell lines.
Discussion
Recent studies have shown that statins can be used
for cancer prevention or therapeutics (25–27),
due to their pleiotropic effects through multiple biological
pathways, particularly the mevalonate pathway (28). Among prenylated proteins, the
activated RAS proteins via farnesylation, constitute central
regulators of numerous cellular functions. It has been shown that
>90% of pancreatic cancer harbor activation mutations of the
KRAS oncogene (29). Therefore, the
beneficial effects of statins on pancreatic cancer treatment are
biologically plausible, although findings from epidemiological
studies on their therapeutic benefits are inconsistent to some
extent (30–37).
Previous studies have revealed that in addition to
the known functions on cholesterol metabolism (38), the statins have some other
pleiotropic effects, involvingthe regulation of the NF-κB signaling
pathway (39), NOTCH signaling
pathway (40,41) and endocytosis (42,43).
The present study indicated that different statins may have their
unique function on cancer cells. For example, high-concentration
fluvastatin inhibited gene expression associated with the NF-κB
signaling pathway, whereas high-concentration simvastatin
upregulated the expression of NF-κB signaling genes (dark turquoise
module of MiaPaCa2). Genes related to the NOTCH signaling pathway
were suppressed by low-concentration statins. By contrast, the
high-concentration simvastatin significantly upregulated NOTCH
signaling genes (green-yellow module of MiaPaCa2). For endocytosis,
low-concentration statins significantly inhibited genes related to
endocytosis in both MiaPaCa2 and PANC1. The inhibition of
endocytosis genes was also observed in high-concentration
simvastatin-treated PANC1 cells, but not in high-concentration
Fluvastatin-treated PANC1 cells (black module of PANC1).
Collectively, our transcriptome analyses revealed that although
these three statins exert similar function on pancreatic cancer,
the concentration of statins and different products of statins may
have different outcomes.
It is well known that the amount of drugs that could
reach cancer cells in the human body is limited (44), and therefore studying drug response
at low concentrations is important. The present study indicated
that the concentration of statins significantly affects its
potential cell death-inducing function. The low-concentration
statins (≤2 µM) did not influence the gene expression associated
with the proliferation of pancreatic cancer cells. They could
trigger transcriptomic changes in MiaPaCa2 cells, but not in PANC1
cells. Notably, as compared with PANC1 cells, the MiaPaCa2 cells
exhibited a high level of metabolism-related gene expression. In
addition, low-concentration statins could significantly inhibit
gene expression in ribosome and proteasome signaling pathways while
upregulating gene expression in metabolism signaling pathways.
These results suggested that statins may only exert limited
antitumor effects in the treatment of metabolism-related pancreatic
cancer. In addition, the low concentration of statins (2 µM) used
in the present study was markedly higher than the orally
administered therapeutic doses (1–25 nM) (45). High-concentration statins (20 µM)
can induce pancreatic cancer cell death in vitro, and this
concentration may potentially be achieved by targeted implantation.
We should therefore be cautious when statins are used clinically to
treat cancer in future, particularly when it comes to the statin
concentrations and the metabolic status of the cancer cells
(46,47).
Although low-concentration statins did not function
satisfactorily on pancreatic cancer cells, it was found that
high-concentration statins could significantly inhibit the
proliferation of cancer cells. High-concentration statins could
significantly suppress gene expression in the cell cycle and DNA
replication signaling pathways. However, they could also induce
gene expression in the ribosome, autophagy and MAPK signaling
pathways, which may eventually lead to therapeutic resistance
(48–50). In conclusion, these results
indicated that, although high-concentration statins can suppress
pancreatic cancer satisfactorily, it is still necessary to consider
combination therapy with other drugs, such as autophagy or MAPK
inhibitors, to reduce potential resistance.
Supplementary Material
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Acknowledgments
The excellent technical assistance of Dr Limei Cao
and Dr Yongjin Zhang (The NHC Key Laboratory of Drug Addiction
Medicine, The First Affiliated Hospital of Kunming Medical
University, Kunming, China) is gratefully acknowledged.
Funding
The present study was supported by grants from the
China Postdoctoral Science Foundation (grant no. 2020M673596XB),
the National Natural Science Foundation of China (grant nos.
81903046, 3171101074, 81860100, 31860306, and 81870458), the
Science and Technology Department of Yunnan Province (grant nos.
2018DH006 and 2018NS0086), the Yunling Scholar (grant no.
YLXL20170002) and Yunnan Outstanding Youth Grant. The funders had
no role in the study design, data collection, and analysis,
decision to publish, or preparation of the manuscript.
Availability of data and materials
The datasets analyzed in the present study are
available from the GEO repository, GEO GSE149566 (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE149566).
Authors' contributions
KW, JY and HW conceived the study. CC, HW and JY
performed all experiments. DK, YX, ZZ, FC, LZ, ZL, JS, HL and SHL
prepared the reagents and samples. JY, HW and FCB analyzed the
data. HW, JY, and CC wrote the manuscript, and KW and FCB reviewed
the results and participated in the discussion about the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rahib L, Smith BD, Aizenberg R, Rosenzweig
AB, Fleshman JM and Matrisian LM: Projecting cancer incidence and
deaths to 2030: The unexpected burden of thyroid, liver, and
pancreas cancers in the United States. Cancer Res. 74:2913–2921.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hamada T, Khalaf N, Yuan C, Babic A,
Morales-Oyarvide V, Qian ZR, Nowak JA, Ng K, Kraft P, Rubinson DA,
et al: Statin use and pancreatic cancer risk in two prospective
cohort studies. J Gastroenterol. 53:959–966. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Oprea TI and Mestres J: Drug repurposing:
Far beyond new targets for old drugs. AAPS J. 14:759–763. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang Y, Liang M, Sun C, Qu G, Shi T, Min
M, Wu Y and Sun Y: Statin use and risk of pancreatic cancer: An
updated meta-analysis of 26 studies. Pancreas. 48:142–150. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mitchell JD, Fergestrom N, Gage BF,
Paisley R, Moon P, Novak E, Cheezum M, Shaw LJ and Villines TC:
Impact of statins on cardiovascular outcomes following coronary
artery calcium scoring. J Am Coll Cardiol. 72:3233–3242. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mohammadkhani N, Gharbi S, Rajani HF,
Farzaneh A, Mahjoob G, Hoseinsalari A and Korsching E: Statins:
Complex outcomes but increasingly helpful treatment options for
patients. Eur J Pharmacol. 863:1727042019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tamburrino D, Crippa S, Partelli S,
Archibugi L, Arcidiacono PG, Falconi M and Capurso G: Statin use
improves survival in patients with pancreatic ductal
adenocarcinoma: A meta-analysis. Dig Liver Dis. 52:392–399. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang SW, Chyuan IT, Shiue C, Yu MC, Hsu
YF and Hsu MJ: Lovastatin-mediated MCF-7 cancer cell death involves
LKB1-AMPK-p38MAPK-p53-survivin signalling cascade. J Cell Mol Med.
24:1822–1836. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xian G, Zhao J, Qin C, Zhang Z, Lin Y and
Su Z: Simvastatin attenuates macrophage-mediated gemcitabine
resistance of pancreatic ductal adenocarcinoma by regulating the
TGF-β1/Gfi-1 axis. Cancer Lett. 385:65–74. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kanvatirth P, Jeeves RE, Bacon J, Besra GS
and Alderwick LJ: Utilisation of the Prestwick chemical library to
identify drugs that inhibit the growth of mycobacteria. PLoS One.
14:e02137132019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Moreau D, Vacca F, Vossio S, Scott C,
Colaco A, Paz Montoya J, Ferguson C, Damme M, Moniatte M, Parton
RG, et al: Drug-induced increase in lysobisphosphatidic acid
reduces the cholesterol overload in Niemann-Pick type C cells and
mice. EMBO Rep. 20:e470552019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brown J, Pirrung M and McCue LA: FQC
Dashboard: Integrates FastQC results into a web-based, interactive,
and extensible FASTQ quality control tool. Bioinformatics.
33:3137–3139. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bolger AM, Lohse M and Usadel B:
Trimmomatic: A flexible trimmer for Illumina sequence data.
Bioinformatics. 30:2114–2120. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Phan L, Hsu J, Tri LQ, Willi M, Mansour T,
Kai Y, Garner J, Lopez J and Busby B: dbVar structural variant
cluster set for data analysis and variant comparison. F1000Res.
5:6732016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Trapnell C, Pachter L and Salzberg SL:
TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics.
25:1105–1111. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu H, Yu J, Li Y, Hou Q, Zhou R, Zhang N,
Jing Z, Jiang M, Li Z, Hua Y, et al: Single-cell RNA sequencing
reveals diverse intratumoral heterogeneities and gene signatures of
two types of esophageal cancers. Cancer Lett. 438:133–143. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang da W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: Paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45((W1)):
W98–W102. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wheatley SP and Altieri DC: Survivin at a
glance. J Cell Sci. 132:jcs2238262019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu SH, Hong Y, Markowiak S, Sanchez R,
Creeden J, Nemunaitis J, Kalinoski A, Willey J, Erhardt P, Lee J,
et al: BIRC5 is a target for molecular imaging and detection of
human pancreatic cancer. Cancer Lett. 457:10–19. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Schultz RM, Merriman RL, Toth JE,
Zimmermann JE, Hertel LW, Andis SL, Dudley DE, Rutherford PG,
Tanzer LR and Grindey GB: Evaluation of new anticancer agents
against the MIA PaCa-2 and PANC-1 human pancreatic carcinoma
xenografts. Oncol Res. 5:223–228. 1993.PubMed/NCBI
|
|
24
|
Okada Y, Eibl G, Guha S, Duffy JP, Reber
HA and Hines OJ: Nerve growth factor stimulates MMP-2 expression
and activity and increases invasion by human pancreatic cancer
cells. Clin Exp Metastasis. 21:285–292. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
McGlynn KA, Hagberg K, Chen J, Graubard
BI, London WT, Jick S and Sahasrabuddhe VV: Statin use and risk of
primary liver cancer in the clinical practice research datalink. J
Natl Cancer Inst. 107:djv0092015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu O, Eberg M, Benayoun S, Aprikian A,
Batist G, Suissa S and Azoulay L: Use of statins and the risk of
death in patients with prostate cancer. J Clin Oncol. 32:5–11.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Murtola TJ, Visvanathan K, Artama M,
Vainio H and Pukkala E: Statin use and breast cancer survival: A
nationwide cohort study from Finland. PLoS One. 9:e1102312014.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Paškevičiūtė M and Petrikaitė V:
Differences of statin activity in 2D and 3D pancreatic cancer cell
cultures. Drug Des Devel Ther. 11:3273–3280. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ghaneh P, Costello E and Neoptolemos JP:
Biology and management of pancreatic cancer. Gut. 56:1134–1152.
2007.PubMed/NCBI
|
|
30
|
Archibugi L, Arcidiacono PG and Capurso G:
Statin use is associated to a reduced risk of pancreatic cancer: A
meta-analysis. Dig Liver Dis. 51:28–37. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jian-Yu E, Graber JM, Lu SE, Lin Y, Lu-Yao
G and Tan XL: Effect of metformin and statin use on survival in
pancreatic cancer patients: A systematic literature review and
meta-analysis. Curr Med Chem. 25:2595–2607. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Simon MS, Desai P, Wallace R, Wu C, Howard
BV, Martin LW, Schlecht N, Liu S, Jay A, LeBlanc ES, et al:
Prospective analysis of association between statins and pancreatic
cancer risk in the Women's Health Initiative. Cancer Causes
Control. 27:415–423. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee HS, Lee SH, Lee HJ, Chung MJ, Park JY,
Park SW, Song SY and Bang S: Statin use and its impact on survival
in pancreatic cancer patients. Medicine (Baltimore). 95:e36072016.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wu BU, Chang J, Jeon CY, Pandol SJ, Huang
B, Ngor EW, Difronzo AL and Cooper RM: Impact of statin use on
survival in patients undergoing resection for early-stage
pancreatic cancer. Am J Gastroenterol. 110:1233–1239. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Walker EJ, Ko AH, Holly EA and Bracci PM:
Statin use and risk of pancreatic cancer: Results from a large,
clinic-based case-control study. Cancer. 121:1287–1294. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jeon CY, Pandol SJ, Wu B, Cook-Wiens G,
Gottlieb RA, Merz CN and Goodman MT: The association of statin use
after cancer diagnosis with survival in pancreatic cancer patients:
A SEER-medicare analysis. PLoS One. 10:e01217832015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hong JY, Nam EM, Lee J, Park JO, Lee SC,
Song SY, Choi SH, Heo JS, Park SH, Lim HY, et al: Randomized
double-blinded, placebo-controlled phase II trial of simvastatin
and gemcitabine in advanced pancreatic cancer patients. Cancer
Chemother Pharmacol. 73:125–130. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Murai T: Cholesterol lowering: Role in
cancer prevention and treatment. Biol Chem. 396:1–11. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bahrami A, Parsamanesh N, Atkin SL, Banach
M and Sahebkar A: Effect of statins on toll-like receptors: A new
insight to pleiotropic effects. Pharmacol Res. 135:230–238. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wu F, Luo T, Mei Y, Liu H, Dong J, Fang Y,
Peng J and Guo Y: Simvastatin alters M1/M2 polarization of murine
BV2 microglia via Notch signaling. J Neuroimmunol. 316:56–64. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zacharek A, Chen J, Cui X, Yang Y and
Chopp M: Simvastatin increases notch signaling activity and
promotes arteriogenesis after stroke. Stroke. 40:254–260. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Verhulst A, D'Haese PC and De Broe ME:
Inhibitors of HMG-CoA reductase reduce receptor-mediated
endocytosis in human kidney proximal tubular cells. J Am Soc
Nephrol. 15:2249–2257. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sidaway JE, Davidson RG, McTaggart F,
Orton TC, Scott RC, Smith GJ and Brunskill NJ: Inhibitors of
3-hydroxy-3-methylglutaryl-CoA reductase reduce receptor-mediated
endocytosis in opossum kidney cells. J Am Soc Nephrol.
15:2258–2265. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Jain RK: Barriers to drug delivery in
solid tumors. Sci Am. 271:58–65. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
DeGorter MK, Tirona RG, Schwarz UI, Choi
YH, Dresser GK, Suskin N, Myers K, Zou G, Iwuchukwu O, Wei WQ, et
al: Clinical and pharmacogenetic predictors of circulating
atorvastatin and rosuvastatin concentrations in routine clinical
care. Circ Cardiovasc Genet. 6:400–408. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Qin C, Yang G, Yang J, Ren B, Wang H, Chen
G, Zhao F, You L, Wang W and Zhao Y: Metabolism of pancreatic
cancer: Paving the way to better anticancer strategies. Mol Cancer.
19:502020. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Sousa CM and Kimmelman AC: The complex
landscape of pancreatic cancer metabolism. Carcinogenesis.
35:1441–1450. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Biancur DE and Kimmelman AC: The
plasticity of pancreatic cancer metabolism in tumor progression and
therapeutic resistance. Biochim Biophys Acta Rev Cancer.
1870:67–75. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hermann PC and Sainz B Jr: Pancreatic
cancer stem cells: A state or an entity? Semin Cancer Biol.
53:223–231. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
New M and Tooze S: The role of autophagy
in pancreatic cancer-recent advances. Biology (Basel). 9:72019.
|