Introduction
Haematological malignancies comprise a group of
heterogeneous tumours with prominent proliferative and
self-replicating abilities (1–4).
Despite advances in treatment, the quality of life of the patients
following treatment is highly affected by chemotherapy resistance,
which severely impedes the clinical treatment of B-cell lymphomas
(BCLs) (5–7). BCLs comprise a heterogenous group of
mature lymphoproliferative malignancies that are among the top 10
causes of cancer-related mortality, mainly occurring in the lymph
nodes, spleen, thymus and other lymphoid organs (4,8,9).
Almost all anti-lymphoma treatments are accompanied by chemotherapy
resistance, including resistance to genotoxic agents, monoclonal
antibodies, antibody-drug conjugates, targeted agents or multidrug
combinations (5,10–12).
Chemotherapy resistance has become the most important challenge to
the successful treatment of BCLs.
Autophagy is an evolutionarily conserved biological
process of the energy metabolism (13,14).
By degrading intracellular organelles and proteins, autophagy
provides cells with biochemical reaction substrates for the
maintenance of homeostasis under nutrient deficiencies or other
stress conditions (15,16). Autophagy has been confirmed to play
a pivotal role in chemotherapy resistance (17). The inhibition of autophagy may
enhance the cytotoxicity of chemotherapy drugs and the apoptosis of
multiple malignant tumour cells, including BCL cells (18). Various studies have also
demonstrated that multiple chemotherapeutic drugs increase
autophagic flux, thereby enhancing drug resistance and promoting
cell survival (19,20).
Autophagy-related 5 (ATG5) is a protein that, in
humans, is encoded by the ATG5 gene located on chromosome 6. ATG5
acts as a proviral factor in the autophagic vesicle formation
process. Conjugation with ATG12, through a ubiquitin-like
conjugating system involving ATG7 as an E1-like activating enzyme
and ATG10 as an E2-like conjugating enzyme, is crucial for the
function of ATG5 (21,22). The ATG5-ATG12 conjugate acts as an
E3-like enzyme, which is required for the lipidation of the ATG8
family of proteins and their association with the vesicle membranes
(23,24). This ATG5-ATG12 conjugate is
necessary for the conjugation of microtubule-associated protein
1A/1B-light chain 3 (LC3)-I and phosphatidylethanolamine to form
LC3-II (23,25).
S100A8 is a multifunctional, calcium-binding protein
that polymerizes with S100A9 to form calprotectin for the
sequestration of metals, including iron, manganese and zinc, via
chelation (26). S100A8 is part of
the 22-member calcium-binding EF hand-containing S100 superfamily
of proteins, which functions predominantly in the innate immune
system (27). S100A8 is expressed
in varying degrees in a variety of cells, and is abundant in
myeloid cells (28). According to
previous reports, S100A8 is mainly involved in cell proliferation
and inflammation, and participates in the occurrence and
development of various types of cancer (29). S100A8 has been shown to be
associated with myeloid differentiation, autophagy, apoptosis and
chemotherapy resistance (30–32).
The Bcl-2/adenovirus E1B 19-kDa protein-interacting
protein 3 (BNIP3) is a BH3-only protein primarily localized to the
mitochondria and endoplasmic reticulum (ER). BNIP3 is anchored to
the outer mitochondrial and ER membrane via its C-terminus
transmembrane domain, whereas the N-terminus faces the cytosol.
BNIP3 induces cell death by perturbing mitochondrial function
(33,34). The abnormal expression and function
of BNIP3 may interfere with Ca2+ homeostasis and promote
cell death (35,36). BNIP3 is also a potent inducer of
autophagy in multiple types of cells (32,37).
The aim of the present study was to determine the
role of S100A8 in the chemotherapy resistance of BCLs by
investigating whether the resistance of BCL cells to Adriamycin
(ADR) and vincristine (VCR) was positively correlated with
autophagy, and whether S100A8 activates autophagy in BCL cells by
promoting BNIP3, Beclin-1 (BECN1) and class III
phosphatidylinositol-3-kinase (PI3KC3) expression, hoping to
identify a novel target for the treatment of BCLs.
Materials and methods
Cell lines and cell culture
Daudi, SUDHL-4 and JeKo-1 cells were purchased from
the Cell Bank of the Chinese Academy of Sciences (Shanghai, China)
and cultured in RPMI-1640 medium supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) and 2 mM glutamine in a humidified
incubator with 5% CO2 and 95% air.
Cell viability assay
Cell viability was assessed by MTT assay. Daudi,
SUDHL-4 and JeKo-1 cells were plated in 96-well plates at a density
of 2×103 cells per well containing 100 µl culture
medium. ADR (1 mg/ml) or VCR (1 mg/ml) was added to 96-well culture
plates with 10 µl per well and incubated for 72 h, followed by
incubation with MTT. The control cells were treated only with PBS
prior to incubation with MTT. Dimethyl sulfoxide was used to
dissolve the formazan. The relative cell viability was calculated
by measuring the absorbance value/well at 570 nm and using the
following equation: Mean optical density (OD) of treated cells/mean
OD of control cells.
Lentivirus-mediated stable silencing
of ATG5 and S100A8
The lentivirus particles of shATG5 LV-ATG5-RNAi
(9513-1) were purchased from GENECHEM. The sequence for the
synthesized shRNA targeting S100A8 was 5′-CCUGAAGAAAUUGCUAGAGTT-3′.
The annealed oligonucleotide fragment was cloned into the
lentivirus plasmid PLL3.7 (Addgene, Inc.) to establish the shRNA
lentiviral vector. Recombinant lentiviruses expressing S100A8 shRNA
or negative control (Shanghai GenePharma Co., Ltd.) were obtained
by plasmid transformation. VSVG, pMDLg/pRRE, RSV-REV (Addgene,
Inc.) and Lipofectamine™ 2000 (Thermo Fisher Scientific, Inc.) were
used for lentivirus packaging in 293T cells. The supernatant of the
293T culture was collected and filtered using a 0.22-µm filter (EMD
Millipore), followed by a concentration step. Medium containing
lentivirus was stored at −80°C for future use.
Reverse transcription quantitative PCR
(RT-qPCR) analysis
Total RNA was isolated using TRIzol®
(Takara Bio, Inc.) and reverse-transcribed into cDNA using
PrimeScript RT Master Mix (Takara Bio, Inc.). according to the
manufacturer's instructions. RT-qPCR was performed using SYBR Green
Real-time PCR Master Mix kit (Takara Bio, Inc.), according to the
manufacturer's instructions. The following PCR conditions were used
on the Light Cycler: 39 cycles at 95°C for 30 sec, 95°C for 5 sec,
followed by 60°C for 30 sec in a 10-µl reaction volume. The
relative expression was normalized to that of GAPDH, which was used
as the internal control. The primers used were as follows: ATG5,
5′-CTCTGCAGTGGCTGAGTGAA-3′ forward and 5′-TCAATCTGTTGGCTGTGGGA-3′
reverse; S100A8, 5′-CCGAGCTGGAGAAAGCCTTG-3′ forward and
5′-AGGTCATCCCTGTAGACGGC-3′ reverse; GAPDH,
5′-AGAAGGCTGGGGCTCATTTG-3′ forward and 5′-AGGGGCCATCCACAGTCTTC-3′
reverse.
Western blotting (WB)
Cells were lysed in SDS lysis buffer (P0013G;
Beyotime Institute of Biotechnology) containing 1% protease
inhibitor PMSF (ST506; Beyotime Institute of Biotechnology). The
extracted protein concentration was determined using the BCA
protein assay kit (P0012S; Beyotime Institute of Biotechnology) and
stored at −80°C. A total of 20 µg of each protein sample was
separated by 12% SDS-PAGE and transferred to PVDF membranes
(IPVH00010; EMD Millipore). Following blocking with 5% non-fat milk
for 1 h at room temperature, the membranes were incubated with
appropriate primary and secondary antibodies. Proteins were
detected using an enhanced chemiluminescence kit (6305; Bio-Rad
Laboratories, Inc.) under a gel electrophoresis imager (Bio-Rad
Laboratories, Inc.). The primary antibodies for LC3 (source,
rabbit; cat. no. ab192890; dilution, 1:1,000), P62 (cat. no.
ab207305; source, rabbit; dilution, 1:2,000) and calnexin (source,
rabbit; cat. no. ab13504; dilution, 1:2,000) were purchased from
Abcam. The primary antibodies for B-cell lymphoma 2 (BCL2; cat. no.
12789-1-AP; source, rabbit; dilution, 1:500), BECN1 (cat. no.
11306-1-AP; source, rabbit; dilution, 1:500), PI3KC3 (cat. no.
20584-1-AP; source, rabbit; dilution, 1:100) and tubulin (cat. no.
10068-1-AP; source, rabbit; dilution, 1:2,000) were purchased from
ProteinTech Group, Inc. The primary antibody for S100A8 (cat. no.
HPA024372; source, rabbit; dilution, 1:500) was purchased from
Merck KGaA. The primary antibodies for manganese-dependent
superoxide dismutase 2 (cat. no. MAB3419; source, mouse; 0.5 µg/ml)
and that for BNIP3 (cat. no. MAB4147; source, mouse; 0.5 µg/ml)
were purchased from R&D. The secondary antibody for anti-rabbit
(cat. no. SA00001-2; dilution, 1:2,000) was purchased from
ProteinTech Group, Inc. The secondary antibody (anti-mouse; cat.
no. SA00001-1; dilution, 1:2,000) was purchased from ProteinTech
Group, Inc. The intensity of the western blot bands was quantified
using ImageJ software (version 1.47; National Institutes of
Health,). Only the blots with band intensity within the linear
range were included.
Autophagy flux analysis of LC3
puncta
Adenovirus expressing mCherry-GFP-LC3 fusion protein
(Ad-mCherry-GFP-LC3, C3011) was purchased from Beyotime Institute
of Biotechnology. Cells were plated in 6-well plates and allowed to
reach 50–70% confluence at the time of Ad-mCherry-GFP-LC3
transfection. Adenoviral infection was performed according to the
manufacturer's instructions. The presence of mRFP-LC3 puncta
indicated the autolysosomes in red fluorescent images.
Transmission electron microscopy (TEM)
analysis of autophagosomes
Cells were harvested and centrifuged at 1,000 × g
for 10 min at room temperature. The cell pellet was fixed with 4%
glutaraldehyde for 24 h at 4°C and 1% osmium tetroxide for 2 h at
4°C. Thereafter, the cell pellet was dehydrated in a graded series
of alcohol and acetone, followed by embedding in Epon 816 (Electron
Microscopy Sciences). Ultrathin sections were cut by a Leica
ultramicrotome (Leica Microsystems, Inc.) and stained with uranyl
acetate for 4 min and followed by lead citrate for 2 min at room
temperature. The exact thickness of the ultrathin section of the
cell samples under the transmission electron microscope was 40 nm.
TEM was conducted using a JEM-1400Plus transmission electron
microscope (JEOL, Ltd.). The mean number of autophagosomes in
tumour cells was counted and quantified.
Immunoprecipitation analysis
Cells were lysed at 4°C in ice-cold lysis buffer
(Beyotime Institute of Biotechnology) and the cell pellet was
prepared by centrifugation (12,000 × g for 10 min at room
temperature). Immunoprecipitation was performed using anti-BECN1
antibody, which was captured with protein A+G agarose beads
(Beyotime Institute of Biotechnology). The bead-bound proteins were
then eluted by boiling in SDS sample buffer, subjected to PAGE and
analyzed by WB.
Statistical analysis
Student's t-test was used when making comparisons in
datasets containing two groups. One-way ANOVA test was used when
making comparisons in datasets containing multiple groups, and
Student-Newman-Keuls test was used as the post hoc test. All data
were analyzed using GraphPad Prism software (version 8.3.1;
GraphPad Software, Inc.) and presented as the mean ± SEM.
*P<0.05, **P<0.01 and ***P<0.001 were considered to
indicate statistically significant differences.
Results
Inhibition of autophagy significantly
damages the drug resistance of BCL cells
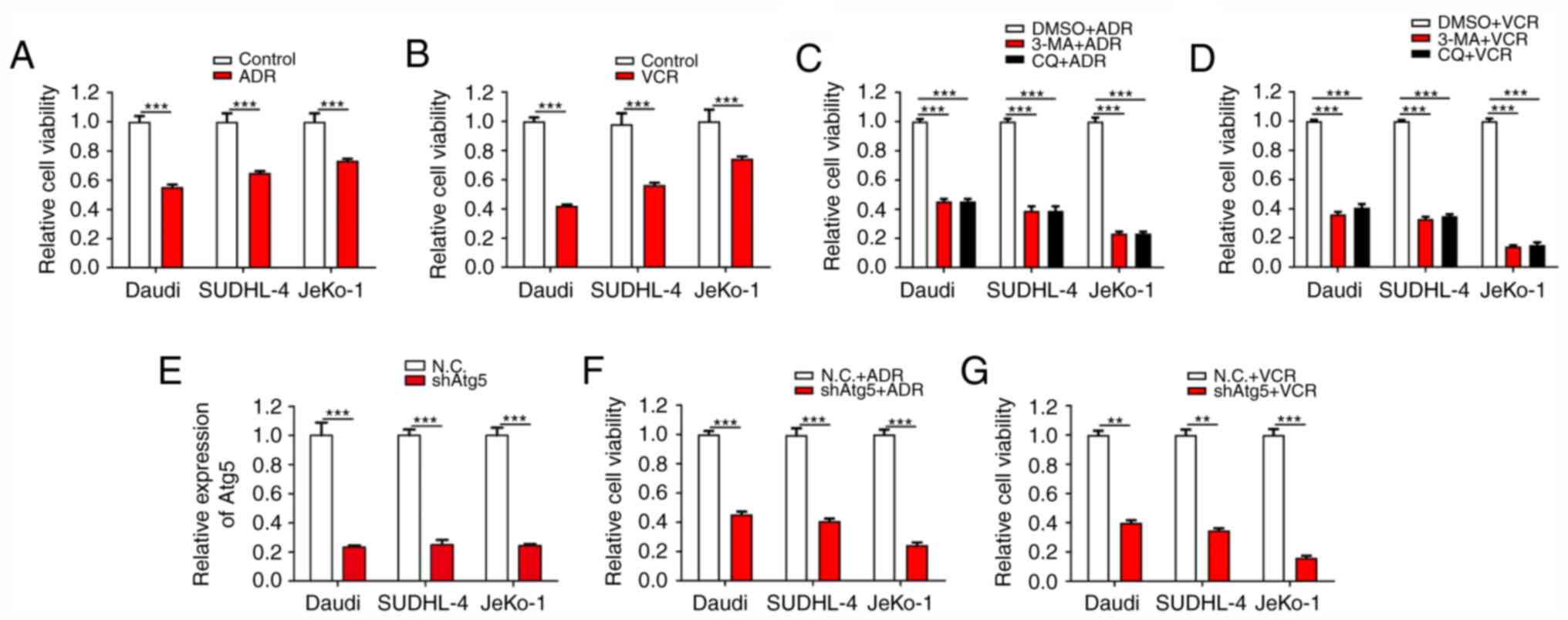
ADR and VCR are effective antitumour agents with a
broad antitumour spectrum. They exert strong inhibitory effects on
RNA synthesis and the proliferation of BCL cells. First, the
inhibitory effects of ADR and VCR on the viability of different BCL
cells, including Daudi (Burkitt lymphoma cells), SUDHL-4 (diffuse
large B-cell lymphoma cells) and JeKo-1 cells (mantle cell lymphoma
cells) were verified. As expected, both ADR and VCR notably
decreased the viability of Daudi, SUDHL-4 and JeKo-1 cells
(Fig. 1A and B). Among these three
cell lines, JeKo-1 cells exhibited the strongest drug resistance
against ADR and VCR treatments, followed by SUDHL-4 and Daudi
cells. Considering the association between autophagy and drug
resistance, it was then verified whether the inhibition of
autophagy counteracted the resistance of BCL cells to ADR and VCR.
The results demonstrated that, when the cells were treated with the
synthetic autophagy inhibitors 3-methyladenine and chloroquine
diphosphate salt combined with ADR and VCR, the viability of BCL
cells, particularly JeKo-1 cells, was significantly inhibited
(Fig. 1C and D). To further confirm
the association between autophagy and drug resistance, lentivirus
infection was conducted to inhibit autophagy via stable ATG5
interference in Daudi, SUDHL-4 and JeKo-1 cells (Fig. 1E). Cell viability assays revealed
that Daudi-shATG5, SUDHL-4-shATG5 and Jeko-1-shATG5 cells were
markedly more susceptible to ADR and VCR, as compared with their
respective control cells (Fig. 1F and
G). These results demonstrated that the resistance of BCL cells
to ADR and VCR is partially dependent on autophagy.
The resistance of BCL cells to ADR and
VCR is positively correlated with autophagy
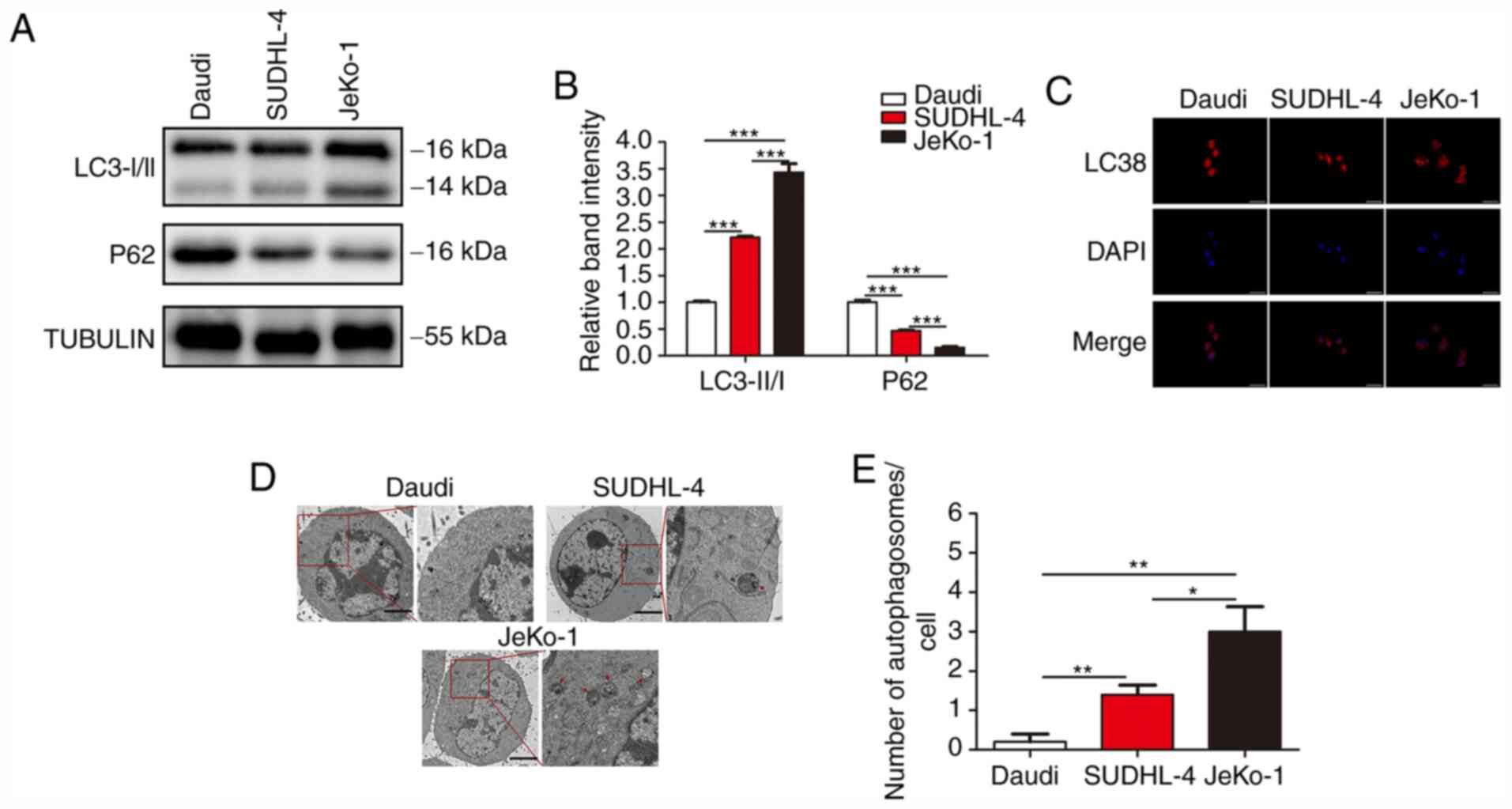
Since the drug resistance of BCL cells was found to
be closely associated with autophagy and the drug resistance of BCL
cells differed greatly, the autophagic activity in different BCL
cells was examined. Three classic assays commonly used in autophagy
research were conducted to characterize the status of autophagy: i)
The protein level of LC3 and P62 was determined by WB; ii) mRFP-LC3
punta were examined by immunofluorescence; and iii) cytoplasmic
accumulation of autophagosomes was detected by TEM. WB revealed
that, among the three B-cell lymphoma cell lines, JeKo-1 cells
exhibited the highest expression of LC3-II, followed by SUDHL-4 and
Daudi cells; furthermore, Daudi cells exhibited the highest
expression of P62, followed by SUDHL-4 and JeKo-1 cells (Fig. 2A and B). In the autophagy flux
assays, there were markedly more mRFP-LC3 punta in the JeKo-1 cells
compared with the Daudi and SUDHL-4 cells (Fig. 2C). The formation of autophagosomes
and autolysosomes as analyzed by TEM was abundant in Jeko-1 cells
and deficient in Daudi and SUDHL-4 cells (Fig. 2D and E). These results demonstrated
that autophagy was activated in BCL cells exhibiting strong drug
resistance.
S100A8 expression is closely
associated with the drug resistance of BCL cells
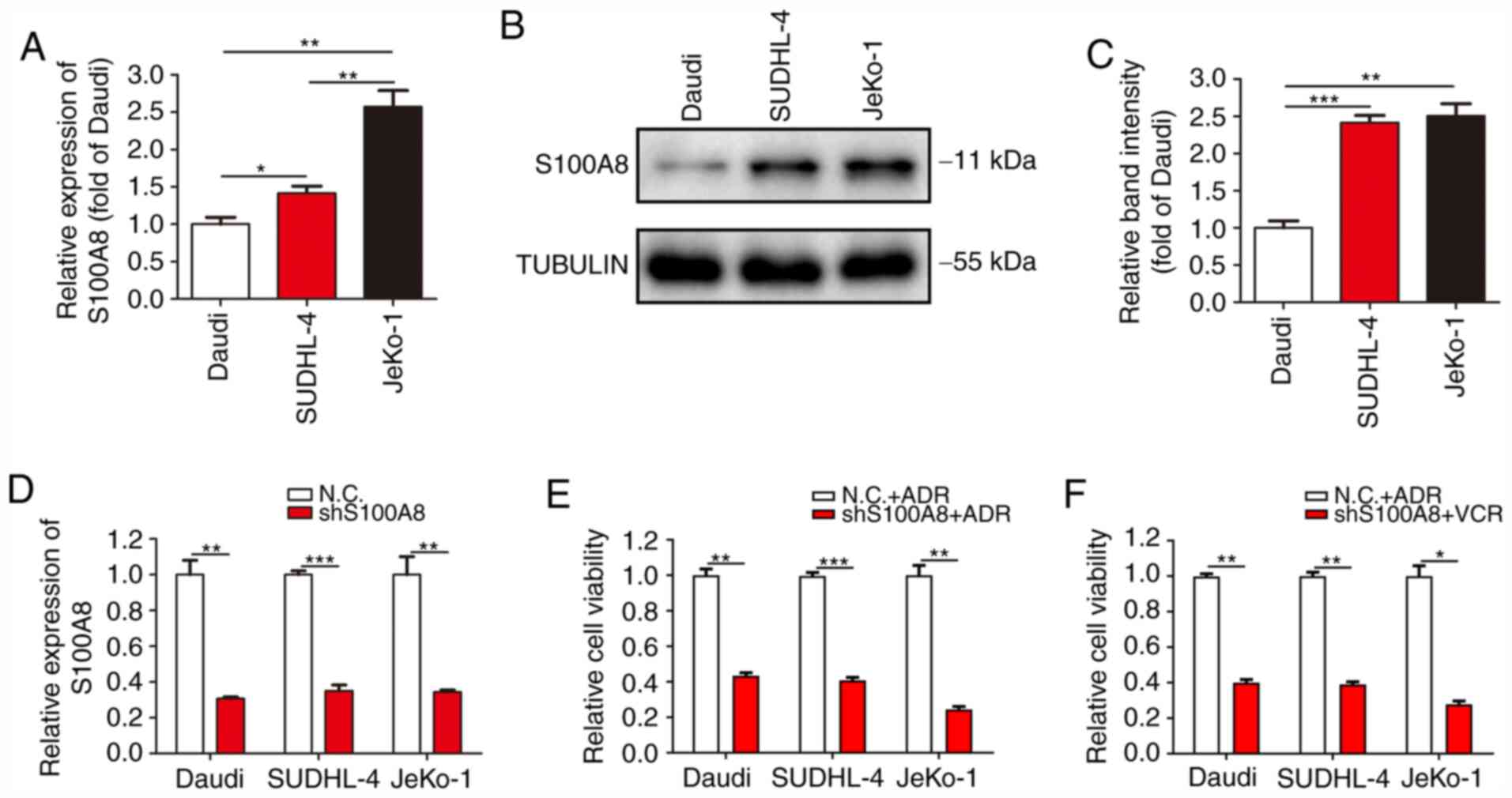
S100A8 is particularly highly expressed in myeloid
cells and plays an important regulatory role in the
differentiation, autophagy and chemotherapy resistance of myeloid
cells. To verify whether the drug resistance and autophagic
activity of Daudi, SUDHL-4 and JeKo-1 cells was associated with
S100A8, the expression of S100A8 was detected in the three B-cell
lymphoma cell lines. RT-qPCR analysis revealed that JeKo-1 cells
exhibited the highest mRNA expression of S100A8, followed by
SUDHL-4 and Daudi cells (Fig. 3A).
WB revealed a markedly higher protein expression of S100A8 in
JeKo-1 and SUDHL-4 cells, as compared with that in Daudi cells
(Fig. 3B and C). To further confirm
the association between S100A8 and drug resistance, lentivirus
infection was conducted to inhibit S100A8 expression in Daudi,
SUDHL-4 and JeKo-1 cells (Fig. 3D).
Cell viability assay revealed that Daudi-shS100A8, SUDHL-4-shS100A8
and JeKo-1-shS100A8 cells were markedly more susceptible to ADR and
VCR, as compared with their respective control cells (Fig. 3E and F). These results demonstrated
that the resistance of BCL cells to ADR and VCR was closely
associated with the expression of S100A8.
S100A8 augments autophagy in BCL
cells
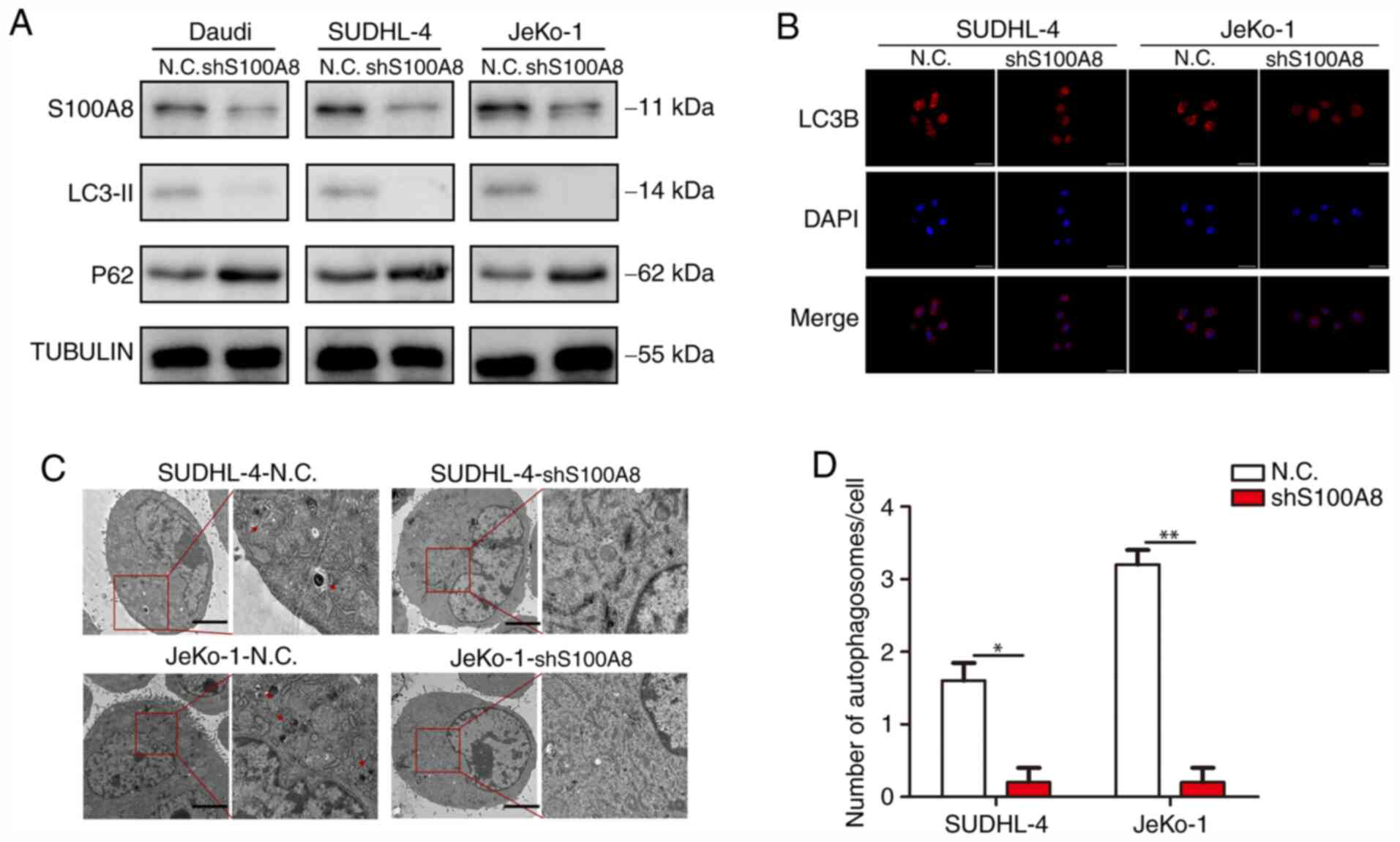
Our previous study demonstrated that the
chemotherapy resistance of BCL cells was largely dependent on
autophagy, and the stronger drug resistance of BCL cells was
associated with a higher S100A8 expression. Next, an attempt was
made to elucidate the association between S100A8 and autophagy in
BCL cells. WB revealed that Daudi-shS100A8, SUDHL-4-shS100A8 and
JeKo-1-shS100A8 cells exhibited a markedly lower LC3-II and higher
P62 protein expression, as compared with their respective control
cells (Fig. 4A). In the autophagy
flux assay, there were markedly more mRFP-LC3 punta in
SUDHL-4-shS100A8 and JeKo-1-shS100A8 cells, as compared with their
respective control cells (Fig. 4B).
Finally, the formation of autophagosomes and autolysosomes as
analyzed by TEM was markedly deficient in BCL cells with S100A8
interference (Fig. 4C and D),
indicating that S100A8 promoted autophagy in BCL cells.
S100A8 activates autophagy by
promoting BNIP3, BECN1 and PI3KC3 expression
Autophagy is an effective method used by cells to
cope with environmental stressors, such as starvation, and is
greatly involved in the normal physiological and pathological
processes of the body. BNIP3 is anchored to the outer mitochondrial
and ER membrane via its C-terminus transmembrane domain, and it can
interact with LC3. The binding of BNIP3 to LC3 on the autophagosome
is crucial for the removal of both ER (ER-phagy) and mitochondria
(mitophagy) via autophagy. To explore the molecular mechanism
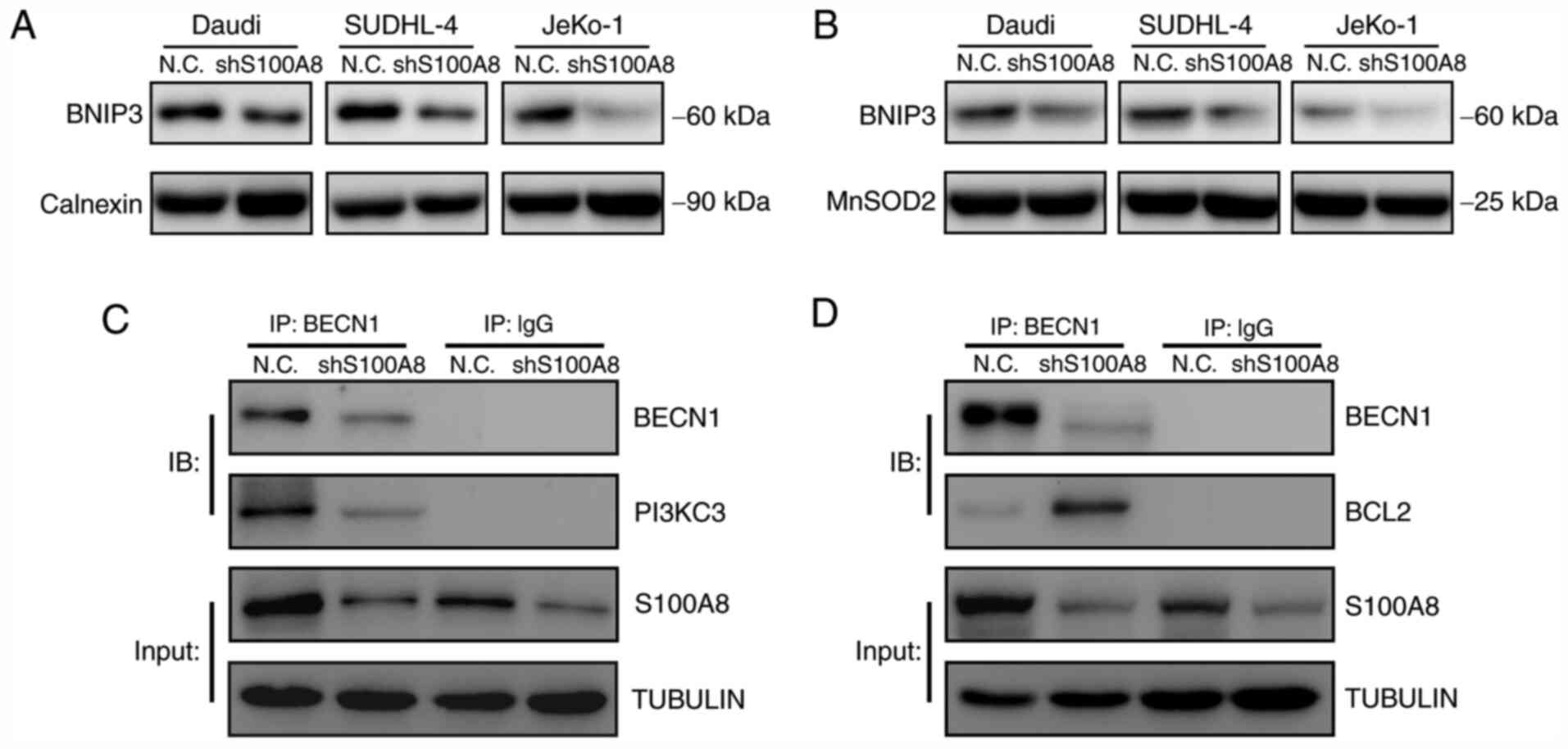
underlying S100A8-mediated autophagy, the BNIP3 expression was
tested in ER and mitochondria following S100A8 interference. The
results demonstrated that S100A8 interference significantly
decreased the BNIP3 protein expression, both in the ER and
mitochondria, in Daudi, SUDHL-4 and JeKo-1 cells (Fig. 5A and B). Next the early autophagic
signalling event of BECN1-PI3KC3 and BECN1-BCL2 complex formation
was analyzed. The co-immunoprecipitation assay revealed that S100A8
interference significantly reduced the formation of the
BECN1-PI3KC3 complex in JeKo-1 cells (Fig. 5C). Furthermore, S100A8 interference
significantly increased the formation of the BECN1-BCL2 complex in
JeKo-1 cells (Fig. 5D).
Collectively, these results indicated that S100A8 may activate
autophagy by promoting the expression of BNIP3 and the BECN1-PI3KC3
complex and inhibiting that of the BECN1-BCL2 complex.
Discussion
The understanding of the occurrence and development
of lymphomas, as well as the treatment of patients with lymphoma,
has greatly improved (38,39). Chemotherapy is the main treatment
strategy for almost all types of BCLs (40). However, drug resistance of lymphoma
cells often renders chemotherapy ineffective and greatly shortens
the long-term event-free survival of the patients. Therefore,
overcoming the drug resistance of lymphoma cells is a major
challenge for BCL treatment (5,40). In
the present study, the BCL cell lines Daudi, SUDHL-4 and JeKo-1
were examined, which represent Burkitt's lymphoma, diffuse large
B-cell lymphoma and mantle cell lymphoma, respectively, and account
for 70–85% of all BCLs. Burkitt's lymphoma is strongly associated
with infection by the oncogenic Epstein Barr virus (EBV). There is
currently no research evidence on the association between EBV
infection and chemoresistance enhancement by S100A8. It may be
hypothesized that certain EBV proteins, such as EBNA1, EBNA3C and
LMP1, may interact with S100A8, thereby affecting BECN1-PI3KC3
complex formation and eventually modifying chemoresistance by
S100A8. Diffuse large B-cell lymphoma is the most prevalent
non-Hodgkin lymphoma (NHL), accounting for 40% of all NHLs. Mantle
cell lymphoma is the most genomically unstable lymphoma that is not
sensitive to radiotherapy, chemotherapy or haematopoietic stem cell
transplantation. However, T-cell or NK-cell lymphoma cells were not
examined in the present study, which constitutes a limitation of
the present study.
Rituximab binds the CD20 antigen on the surface of B
lymphocytes, causing an enhanced immune elimination of malignant B
cells. The present study did not involve immune response
experiments in vivo but cellular chemoresistance experiments
in vitro; therefore, the broad-spectrum chemotherapy drugs
ADR and VCR were selected in the present study instead of
rituximab.
Autophagy, also referred to as type II programmed
cell death, is an evolutionarily conserved and strictly controlled
metabolic process. Autophagy plays an important role in homeostasis
and cell survival. Cancer cells have multiple responses to
chemotherapy, from activating survival pathways to triggering cell
death (41,42). Chemotherapy drugs can significantly
increase the autophagic activity of BCL cells (43). Multiple studies have demonstrated
that inhibition of autophagy enhances chemotherapy-induced BCL cell
death (44–46). However, the specific molecular
regulatory effects of S100A8 on autophagy and chemoresistance in
BCL cells remain unclear. The present study indicated that the drug
resistance of BCL cells was positively correlated with autophagic
activity, and S100A8 was closely associated with the drug
resistance of BCL cells by activating autophagy.
Prior to this study, S100A8 has been reported to
promote autophagy in cancer cells through the cross-talk between
mitochondria and lysosomes via reactive oxygen species (32), or through the activation of the
autophagy initiation complex BECN1-PI3KC3 (30). In the present study, it was
confirmed that S100A8 reduced the sensitivity of BCL cells to
chemotherapy by maintaining BCL cell autophagy, thereby increasing
the resistance of BCL cells to chemotherapy. The role of the S100A8
protein in pro-autophagic activities is mediated through the
promotion of BECN1-PI3KC3 and BECN1-BCL2 complex formation
(31). This study revealed that
S100A8 stimulated BECN1-PI3KC3 complex formation as an early
autophagic signalling event that significantly promoted autophagy.
In addition, the dissociation of BCL2 from BECN1 was found to be an
important mechanism involved in S100A8-activated autophagy.
LC3 is initially processed into its cytoplasmic
form, LC3-I, then coupled with lipid phosphatidylethanolamine,
generating its final LC3-II form (47). BNIP3 is a cell death-inducing factor
of the Bcl-2 family of proteins, more precisely a member of the
BH3-only subfamily. Proteins such as S100A8 in the BH3-only
subfamily bind through a common BH3 domain, rather than the BH1 and
BH2 domains as the other Bcl-2 family members. The binding of BNIP3
to LC3 is crucial for autophagosome formation (47,48).
The present study demonstrated that S100A8 accelerated BNIP3
expression in ER and mitochondria, resulting in the promotion of
autophagy.
In conclusion, the molecular mechanisms of
S100A8-triggered autophagy and drug resistance of BCL cells was
investigated in the present study. The involvement of the
S1008-BNIP3/BECN1-PI3KC3 complex-autophagy-chemoresistance axis in
the autophagic regulation of drug resistance in BCL cells was also
confirmed. These findings may help overcome drug resistance in the
treatment of BCLs.
Acknowledgements
Not applicable.
Funding
The present study was funded by the Key Research
Project from Science & Technology Department of Sichuan
Province (grant no. 2019YFS0301); The Doctor Research Foundation of
The Affiliated Hospital of Southwest Medical University (grant nos.
19032 and 19079); The Key Research Project from Health and Family
Planning Commission of Sichuan Province (grant no. 18ZD014); The
Applied Basic Research Project of Luzhou Science and Technology
Bureau (grant no. 2019LZXNYDJ54); and the National Natural Science
Foundation of China (grant no. 81450030).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
LZ performed the experiments and data analysis and
wrote the manuscript. SZ, TZ, KY and XL participated in conducting
this study. JT designed and oversaw this study, and contributed to
the revision of the manuscript. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
All institutional and national guidelines for the
use of laboratory materials were followed.
Patient consent for publication
Not applicable.
Competing interests
All the authors declare that they have no competing
interests.
References
|
1
|
Cruz-Rodriguez N, Combita AL and Zabaleta
J: Epigenetics in hematological malignancies. Methods Mol Biol.
1856:87–101. 2018. View Article : Google Scholar
|
|
2
|
Upadhyay VA and Fathi AT: Induction
chemotherapy in acute myeloid leukaemia: Origins and emerging
directions. Curr Opin Hematol. 25:67–74. 2018. View Article : Google Scholar
|
|
3
|
Stahl M, Kohrman N, Gore SD, Kim TK,
Zeidan AM and Prebet T: Epigenetics in cancer: A hematological
perspective. PLoS Genet. 12:e10061932016. View Article : Google Scholar
|
|
4
|
Swerdlow SH, Campo E, Pileri SA, Harris
NL, Stein H, Siebert R, Advani R, Ghielmini M, Salles GA, Zelenetz
AD and Jaffe ES: The 2016 revision of the World Health Organization
classification of lymphoid neoplasms. Blood. 127:2375–2390. 2016.
View Article : Google Scholar
|
|
5
|
Klener P and Klanova M: Drug resistance in
non-Hodgkin lymphomas. Int J Mol Sci. 21:20812020. View Article : Google Scholar
|
|
6
|
Feugier P: A review of rituximab, the
first anti-CD20 monoclonal antibody used in the treatment of B
non-Hodgkin's lymphomas. Future Oncol. 11:1327–1342. 2015.
View Article : Google Scholar
|
|
7
|
El-Galaly TC, Cheah CY, Kristensen D,
Hutchison A, Hay K, Callréus T and Villa D: Potentials, challenges
and future of chimeric antigen receptor T-cell therapy in
non-Hodgkin lymphomas. Acta Oncol. 59:766–774. 2020. View Article : Google Scholar
|
|
8
|
Swerdlow SH and Cook JR: As the world
turns, evolving lymphoma classifications-past, present and future.
Hum Pathol. 95:55–77. 2020. View Article : Google Scholar
|
|
9
|
Jones SE: Non-Hodgkin lymphomas. JAMA.
234:633–638. 1975. View Article : Google Scholar
|
|
10
|
Li Y, Wang LX, Pang P, Twitty C, Fox BA,
Aung S, Urba WJ and Hu HM: Cross-presentation of tumor associated
antigens through tumor-derived autophagosomes. Autophagy.
5:576–577. 2009. View Article : Google Scholar
|
|
11
|
Gallo S, Gatti S, Sala V, Albano R,
Costelli P, Casanova E, Comoglio PM and Crepaldi T: Agonist
antibodies activating the Met receptor protect cardiomyoblasts from
cobalt chloride-induced apoptosis and autophagy. Cell Death Dis.
5:e11852014. View Article : Google Scholar
|
|
12
|
Yu SF, Zheng B, Go M, Lau J, Spencer S,
Raab H, Soriano R, Jhunjhunwala S, Cohen R, Caruso M, et al: A
novel anti-CD22 anthracycline-based antibody-drug conjugate (ADC)
that overcomes resistance to auristatin-based ADCs. Clin Cancer
Res. 21:3298–3306. 2015. View Article : Google Scholar
|
|
13
|
Mizushima N: Autophagy: Process and
function. Genes Dev. 21:2861–2873. 2007. View Article : Google Scholar
|
|
14
|
Mizushima N and Komatsu M: Autophagy:
Renovation of cells and tissues. Cell. 147:728–741. 2011.
View Article : Google Scholar
|
|
15
|
Jardon MA, Rothe K, Bortnik S, Vezenkov L,
Jiang X, Young RN, Lum JJ and Gorski SM: Autophagy: From structure
to metabolism to therapeutic regulation. Autophagy. 9:2180–2182.
2013. View Article : Google Scholar
|
|
16
|
Mizushima N and Levine B: Autophagy in
mammalian development and differentiation. Nat Cell Biol.
12:823–830. 2010. View Article : Google Scholar
|
|
17
|
Li X, Zhou Y, Li Y, Yang L, Ma Y, Peng X,
Yang S, Liu J and Li H: Autophagy: A novel mechanism of
chemoresistance in cancers. Biomed Pharmacother. 119:1094152019.
View Article : Google Scholar
|
|
18
|
Piya S, Andreeff M and Borthakur G:
Targeting autophagy to overcome chemoresistance in acute
myleogenous leukemia. Autophagy. 13:214–215. 2017. View Article : Google Scholar
|
|
19
|
Levy JMM, Towers CG and Thorburn A:
Targeting autophagy in cancer. Nat Rev Cancer. 17:528–542. 2017.
View Article : Google Scholar
|
|
20
|
Onorati AV, Dyczynski M, Ojha R and
Amaravadi RK: Targeting autophagy in cancer. Cancer. 124:3307–3318.
2018. View Article : Google Scholar
|
|
21
|
Su LY, Luo R, Liu Q, Su JR, Yang LX, Ding
YQ, Xu L and Yao YG: Atg5- and Atg7-dependent autophagy in
dopaminergic neurons regulates cellular and behavioral responses to
morphine. Autophagy. 13:1496–1511. 2017. View Article : Google Scholar
|
|
22
|
Zheng W, Xie W, Yin D, Luo R, Liu M and
Guo F: ATG5 and ATG7 induced autophagy interplays with UPR via PERK
signaling. Cell Commun Signal. 17:422019. View Article : Google Scholar
|
|
23
|
Romanov J, Walczak M, Ibiricu I, Schüchner
S, Ogris E, Kraft C and Martens S: Mechanism and functions of
membrane binding by the Atg5-Atg12/Atg16 complex during
autophagosome formation. EMBO J. 31:4304–4317. 2012. View Article : Google Scholar
|
|
24
|
Williams RA, Woods KL, Juliano L, Mottram
JC and Coombs GH: Characterization of unusual families of ATG8-like
proteins and ATG12 in the protozoan parasite Leishmania major.
Autophagy. 5:159–172. 2009. View Article : Google Scholar
|
|
25
|
Fletcher K, Ulferts R, Jacquin E, Veith T,
Gammoh N, Arasteh JM, Mayer U, Carding SR, Wileman T, Beale R and
Florey O: The WD40 domain of ATG16L1 is required for its
non-canonical role in lipidation of LC3 at single membranes. EMBO
J. 37:e978402018. View Article : Google Scholar
|
|
26
|
Pruenster M, Vogl T, Roth J and Sperandio
M: S100A8/A9: From basic science to clinical application. Pharmacol
Ther. 167:120–131. 2016. View Article : Google Scholar
|
|
27
|
Wang S, Song R, Wang Z, Jing Z, Wang S and
Ma J: S100A8/A9 in inflammation. Front Immunol. 9:12982018.
View Article : Google Scholar
|
|
28
|
Yang J, Anholts J, Kolbe U, Stegehuis-Kamp
JA, Claas FHJ and Eikmans M: Calcium-binding proteins S100A8 and
S100A9: Investigation of their immune regulatory effect in myeloid
cells. Int J Mol Sci. 19:18332018. View Article : Google Scholar
|
|
29
|
Shabani F, Farasat A, Mahdavi M and Gheibi
N: Calprotectin (S100A8/S100A9): A key protein between inflammation
and cancer. Inflamm Res. 67:801–812. 2018. View Article : Google Scholar
|
|
30
|
Yang M, Zeng P, Kang R, Yu Y, Yang L, Tang
D and Cao L: S100A8 contributes to drug resistance by promoting
autophagy in leukemia cells. PLoS One. 9:e972422014. View Article : Google Scholar
|
|
31
|
Yang L, Yang M, Zhang H, Wang Z, Yu Y, Xie
M, Zhao M, Liu L and Cao L: S100A8-targeting siRNA enhances arsenic
trioxide-induced myeloid leukemia cell death by down-regulating
autophagy. Int J Mol Med. 29:65–72. 2012.
|
|
32
|
Ghavami S, Eshragi M, Ande SR, Chazin WJ,
Klonisch T, Halayko AJ, McNeill KD, Hashemi M, Kerkhoff C and Los
M: S100A8/A9 induces autophagy and apoptosis via ROS-mediated
cross-talk between mitochondria and lysosomes that involves BNIP3.
Cell Res. 20:314–331. 2010. View Article : Google Scholar
|
|
33
|
Chourasia AH and Macleod KF: Tumor
suppressor functions of BNIP3 and mitophagy. Autophagy.
11:1937–1938. 2015. View Article : Google Scholar
|
|
34
|
Dhingra A, Jayas R, Afshar P, Guberman M,
Maddaford G, Gerstein J, Lieberman B, Nepon H, Margulets V, Dhingra
R and Kirshenbaum LA: Ellagic acid antagonizes Bnip3-mediated
mitochondrial injury and necrotic cell death of cardiac myocytes.
Free Radic Biol Med. 112:411–422. 2017. View Article : Google Scholar
|
|
35
|
Zhang L, Li L, Leavesley HW, Zhang X,
Borowitz JL and Isom GE: Cyanide-induced apoptosis of dopaminergic
cells is promoted by BNIP3 and Bax modulation of endoplasmic
reticulum-mitochondrial Ca2+ levels. J Pharmacol Exp Ther.
332:97–105. 2010. View Article : Google Scholar
|
|
36
|
Hanna RA, Quinsay MN, Orogo AM, Giang K,
Rikka S and Gustafsson ÅB: Microtubule-associated protein 1 light
chain 3 (LC3) interacts with Bnip3 protein to selectively remove
endoplasmic reticulum and mitochondria via autophagy. J Biol Chem.
287:19094–19104. 2012. View Article : Google Scholar
|
|
37
|
Ma Z, Chen C, Tang P, Zhang H, Yue J and
Yu Z: BNIP3 induces apoptosis and protective autophagy under
hypoxia in esophageal squamous cell carcinoma cell lines: BNIP3
regulates cell death. Dis Esophagus. 30:1–8. 2017. View Article : Google Scholar
|
|
38
|
Ferreri AJM: Therapy of primary CNS
lymphoma: Role of intensity, radiation, and novel agents.
Hematology Am Soc Hematol Educ Program. 2017:565–577. 2017.
View Article : Google Scholar
|
|
39
|
Witkowska M, Smolewski P and Robak T:
Investigational therapies targeting CD37 for the treatment of
B-cell lymphoid malignancies. Expert Opin Investig Drugs.
27:171–177. 2018. View Article : Google Scholar
|
|
40
|
Reagan PM and Barr PM: Chemotherapy-free
treatment in non-Hodgkin lymphoma: A steep learning curve. Lancet
Haematol. 4:e152–e153. 2017. View Article : Google Scholar
|
|
41
|
Rivera Vargas T and Apetoh L: Danger
signals: Chemotherapy enhancers? Immunol Rev. 280:175–193. 2017.
View Article : Google Scholar
|
|
42
|
Gunjur A: Short vs. long course adjuvant
chemotherapy for colon cancer. Lancet Oncol. 19:e2362018.
View Article : Google Scholar
|
|
43
|
Gayle S, Landrette S, Beeharry N, Conrad
C, Hernandez M, Beckett P, Ferguson SM, Xu T, Rothberg J and
Lichenstein H: B-cell non-Hodgkin lymphoma: Selective vulnerability
to PIKFYVE inhibition. Autophagy. 13:1082–1083. 2017. View Article : Google Scholar
|
|
44
|
Zeng X, Li Y, Fan J, Zhao H, Xian Z, Sun
Y, Wang Z, Wang S, Zhang G and Ju D: Recombinant human arginase
induced caspase-dependent apoptosis and autophagy in non-Hodgkin's
lymphoma cells. Cell Death Dis. 4:e8402013. View Article : Google Scholar
|
|
45
|
Fan J, Zeng X, Li Y, Wang S, Wang Z, Sun
Y, Gao H, Zhang G, Feng M and Ju D: Autophagy plays a critical role
in ChLym-1-induced cytotoxicity of non-hodgkin's lymphoma cells.
PLoS One. 8:e724782013. View Article : Google Scholar
|
|
46
|
Wu K, Sun XQ, Wang CQ, Gao TX, Sun P, Wang
Y, Jiang WQ, Li ZM and Huang JJ: Metronomic combination
chemotherapy using everolimus and etoposide for the treatment of
non-Hodgkin lymphoma. Cancer Med. 8:4688–4698. 2019. View Article : Google Scholar
|
|
47
|
Schaaf MB, Keulers TG, Vooijs MA and
Rouschop KM: LC3/GABARAP family proteins: Autophagy-(un)related
functions. FASEB J. 30:3961–3978. 2016. View Article : Google Scholar
|
|
48
|
Birgisdottir AB, Lamark T and Johansen T:
The LIR motif-crucial for selective autophagy. J Cell Sci.
126:3237–3247. 2013.
|