Introduction
Cervical cancer is the fourth most frequently
diagnosed tumor and the fourth leading cause of cancer-associated
mortality among women worldwide, with an estimated 570,000 cases
and 311,000 deaths according to the 2018 Global Cancer Statistics
(1–4). Persistent infection with high-risk
human papillomaviruses (HPVs) is a well known major cause of ~95%
of cervical squamous intraepithelial lesions and invasive cervical
cancer, and poses a major threat to the health of women (5–8). With
the advent of high-quality screening programs for cervical cancer
and precancerous lesions, such as liquid-based cytology,
Papanicolaou smear test, HPV test and the relatively recent
development and application of the HPV vaccine, the incidence rate
of cervical cancer has exhibited a marked decline, particularly in
developed countries (9). In some
developing countries or remote areas, a large proportion of women
are not screened or voluntarily vaccinated against HPV, and
therefore cervical cancer remains the leading cause of
cancer-associated mortality among women in 43 less developed
countries (10,11). Despite improvements in surgical
treatment, chemotherapy and radiotherapy in recent decades, the
prognosis of patients with advanced cervical cancer remains poor
(12). Therefore, it is urgent to
investigate the possible pathogenic mechanism underlying the
development of cervical cancer, and further research on novel
therapeutic strategies is required.
The claudin family, which includes ~18 closely
related transmembrane proteins, serves a key structural role in
tight junctions (13–17). Tight junctions are a type of
cell-to-cell adhesion molecules in epithelial or endothelial cells,
preventing the lateral diffusion of integral membrane proteins
between the lateral, basal and apical surfaces (18–21),
thereby preserving the specialized functions of each surface.
Additionally, tight junctions have been reported to be associated
with cell proliferation (22,23),
migration (24) and differentiation
in different types of cells, such as Madin-Darby Canine Kidney and
lactating mammary epithelial cells (25).
Claudin 1 (CLDN1), which has four transmembrane
domains with two extracellular loops, was the first member of the
claudin family to be identified and named (26,27).
Increased or decreased expression levels of CLDN1 have been
reported to damage the epithelial permeability barrier and disrupt
cellular polarity, which results in decreased cell adhesion
(28). For example, Ryu et
al (26) observed that
downregulation of CLDN1 may lead to atopic dermatitis through the
ERK/STAT3 signaling pathway. Furthermore, increased or decreased
expression levels of CLDN1 have been found to be associated with
several types of tumor; for example, overexpression of CLDN1 has
been observed in nasopharyngeal, ovarian and oral squamous cell
cancer (29). A number of studies
have revealed that CLDN1 upregulation may contribute to tumor
progression in esophageal squamous cell carcinoma, colon cancer and
gastric carcinoma (30–32), while in other types of cancer, such
as lung adenocarcinoma, CLDN1 acts as a tumor suppressor by
activating the expression of matrix metallopeptidase-2 and high
CLDN1 expression is associated with an improved prognosis (33). In prostate adenocarcinoma, Sheehan
et al (34) demonstrated
that patients with loss of CLDN1 protein expression had a higher
rate of disease recurrence. Therefore, CLDN1 serves distinct roles
in different types of tissue and tumor (29,35–37).
However, the role of CLDN1 in cervical cancer remains unclear.
Several studies have reported that CLDN1 protein expression may
promote the migration of cervical cancer cells and increase the
risk of lymph node metastasis (38,39).
Another study investigated the role of CLDN1 and CLDN7 in HeLa
cells and revealed that neither of them promoted tumorigenesis in
cervical cancer, but rather decreased the migratory and invasive
abilities of cells (40).
Therefore, these different results may be partially attributed to
the different cervical cancer cell subtypes.
The transcription factor p63 is a member of the p53
gene family that serves an important role in epidermal development
and diseases (41). A previous
study (42) has demonstrated that
ΔNp63α is the most abundant isoform expressed in cervical cancer
cells and exerts an antitumor effect. Further analyses, including
chromatin immunoprecipitation sequencing (Chip-seq) and RNA
sequencing (RNA-seq) results, demonstrated that ΔNp63α can directly
regulate the expression levels of the target gene CLDN1 (41,42).
The aim of the present study was to explore the
effects of CLDN1 on the biological behavior of cervical squamous
cell cancer cells, in order to determine whether CLDN1 may act as
an anti-oncogene in cervical cancer, and whether it may represent a
potential target for improving the effectiveness of diagnosis and
treatment of cervical cancer.
Materials and methods
Cell lines
293T cells and the cervical squamous cell carcinoma
SiHa and ME-180 cell lines were purchased from the American Type
Culture Collection and cultured under suitable conditions. 293T and
SiHa cells were cultured in DMEM (HyClone; Cytiva) supplemented
with 10% FBS (Biological Industries) and 1% penicillin/streptomycin
at 37°C in humidified atmosphere containing 5% CO2.
ME-180 cells were cultured in McCoy's 5A medium (HyClone; Cytiva)
supplemented with 10% FBS and 1% penicillin/streptomycin at 37°C in
a humidified atmosphere containing 5% CO2.
Overexpression of CLDN1 in SiHa
cells
The specific PCR primers of CLDN1 were as follows:
Forward, 5′-CTAGGCGCCGGAATTAGATCTGCCACCATGGCCAACGCGGGGCTGCA-3′
(restriction site, BglII) and reverse,
5′-GCGGAATTCGTTAACCTCGAGTCACACGTAGTCTTTCCCGC-3′ (restriction site,
XhoI). The full-length human CLDN1 was obtained by reverse
transcription-quantitative PCR (RT-qPCR) analysis and cloned into
the pMIGR1 plasmid (Wuhan Miaoling Biological Technology Co.,
Ltd.), which was named as the pMIGR1-CLDN1 plasmid. The plasmid was
confirmed by sequencing analysis and the US National Center for
Biotechnology Information blasting (data not shown). Subsequently,
3 µg plasmids combined with 3 µg packaging vectors pCL-10A1 (Wuhan
Miaoling Biological Technology Co., Ltd.) were transfected into
293T cells using Lipofectamine® 2000 transfection
reagent (Invitrogen; Thermo Fisher Scientific, Inc.; pMIGR1
plasmid: pCL-10A1=1:1). After 10 h at 37°C, the cell medium was
changed, and after a further 48 h of incubation at 37°C, the viral
supernatant of 293T cells was collected by centrifugation (12,000 ×
g for 10 min) at room temperature. Subsequently, 700 µl virus
supernatant mixed with 300 µl normal culture medium were used to
infect SiHa cells (multiplicity of infection ~106
PFU/ml) in a 12-well plate for 36-48 h. Subsequently, SiHa/CLDN1
stable cell lines were obtained by selection with 400 µg/ml G-418
(Beijing Solarbio Science & Technology Co., Ltd.) for 2-3 weeks
(32), after which the
untransfected and unstable SiHa cells would be dead, while the
control cells (SiHa/con) and SiHa/CLDN1 stable cells would survive.
Stably transfected cells were identified using western blotting and
RT-qPCR analysis.
Overexpression of CLDN1 in
ME-180/shp63
Firstly, ME-180/shp63 cells were constructed
successfully. The library of shRNAs used for producing
viral-packaged interfering RNAs for target genes was obtained from
Sigma-Aldrich (Merck KGaA). The p63-short hairpin (sh)RNA plasmid
contained a shRNA sequence targeting p63
(5′-ACAGACCCTTTGTAGCGTG-3′; target at position 3,648 to 3,666). The
lentiviral vectors pLKO.1 or p63-shRNA were used for plasmid
construction and transfected into 293T cells simultaneously with
helper plasmids (VSVG, 2 µg; GAG, 2 µg; Rev, 22 µg; shRNA, 1 µg)
using Lipofectamine 2000. After 10 h, the cell medium was changed
and after 48 h, the viral supernatant of 293T cells was collected
by centrifugation (12,000 × g for 10 min) at room temperature to
remove the dead cells. Subsequently, 700 µl virus supernatant mixed
with 300 µl normal culture medium were used to infect ME-180 cells
(multiplicity of infection was ~106 PFU/ml) at 37°C for
36-48 h. ME-180/shp63 stable cell lines were obtained by selection
with 2 µg/ml puromycin (Merck; KGaA) for 2 weeks. Secondly, CLDN1
overexpression was induced in ME-180/shp63 cells. To establish the
double transfected cell lines in which p63 was knocked down but
CLDN1 was overexpressed, the pLKO.1 vectors were modified by
replacing the puromycin-resistant cassette with the G418-resistant
cassette. The experimental process was performed as aforementioned
for SiHa cells, except that ME-180/shp63-CLDN1 stable cell lines
were obtained by selection with 400 µg/ml G-418 (Beijing Solarbio
Science & Technology Co., Ltd.) for 2-3 weeks.
Knockdown of CLDN1 by shRNA
First, the SiHa/p63 cells overexpressing p63 were
constructed. The specific PCR primers of p63 were as follows:
Forward, 5′-GAAGATCTGCCACCATGTTGTACCTGGA-3′ (restriction site,
BglII) and reverse,5′-CCGCTCGAGTCACTCCCCCTCCTCTTTGA-3′
(restriction site, XhoI). The full-length human p63 was
obtained by RT-qPCR and cloned into the pMIGR1 plasmid (Wuhan
Miaoling Biological Technology Co., Ltd.), which was named as the
pMIGR1-p63 plasmid. The next experimental steps were as
aforementioned for the overexpression of CLDN1 in SiHa cells.
Stably transfected cells were identified using western blotting and
RT-qPCR analysis.
The CLDN1-shRNA plasmid containing the target shRNA
sequences (5′-GCATCGTTATTAAGCCCTTAT−3′ at position 1,490 to 1,510)
and the shRNA control plasmid pLKO.1 (Shaanxi YouBio Technology
Co., Ltd.) were constructed by the School of Life Sciences,
University of Science and Technology of China (Hefei, China).
Lentiviral vectors pLKO.1 or CLDN1-shRNA were used for plasmid
construction and transduced into 293T cells simultaneously with
helper plasmids (1 µg lentiviral plasmid; 6 µg packaging vector)
using Lipofectamine 2000. After 10 h, the cell medium was changed
and after a further 48 h, the viral supernatant of 293T cells was
collected by centrifugation (12,000 × g for 10 min at room
temperature) to remove the dead cells. Subsequently, 700 µl virus
supernatant mixed with 300 µl normal culture medium was used to
infect SiHa/p63 cells (multiplicity of infection was
~106 PFU/ml) at 37°C for 36-48 h. The supernatants were
replaced with fresh medium with 2.0 µg/ml puromycin.
SiHa/p63-shCLDN1 stable cell lines were obtained by selection with
2 µg/ml puromycin for 2-3 weeks and were detected via RT-qPCR
analysis and western blotting.
Cell proliferation in vitro
A total of 6×104 cells in 100 µl of their
respective culture medium, including SiHa/con and SiHa/CLDN1,
SiHa/p63-pLKO and SiHa/p63-shCLDN1, ME-180/shp63-con and
ME-180/shp63-CLDN1, were seeded into 16-well plates (ACEA
Bioscience, Inc.) and measured using the xCELLigence Real-Time Cell
Analysis measuring instrument (Roche Diagnostics). Each type of
cell was added to >3 wells and the steps were strictly followed
according to the manufacturer's protocol. After 100 h, the cell
proliferation plot was acquired using the xCELLigence system (Roche
Diagnostics).
Colony formation assay
A total of 100 cells/well from each cell line
(SiHa/con and SiHa/CLDN1, SiHa/p63-pLKO and SiHa/p63-shCLDN1,
ME-180/shp63-con and ME-180/shp63-CLDN1) were seeded into 6-well
plates separately. SiHa/con, SiHa/CLDN1, SiHa/p63-pLKO and
SiHa/p63-shCLDN1 cells were cultured in DMEM supplemented with 10%
FBS, whereas ME-180/shp63-con and ME-180/shp63-CLDN1 cells were
cultured in McCoy's 5A supplemented with 10% FBS. Experiments in
each cell line were run in triplicate at 37°C in a humidified
atmosphere of 5% CO2. After 2 weeks, the colonies (≥10
cells) were observed under a light microscope (Olympus Corporation;
magnification, ×100). The cells were washed twice with PBS and
stained with 0.5% crystal violet for 1 min at room temperature
(Beijing Solarbio Science & Technology Co., Ltd.). The
quantitation of the colony formation assays was described using
histograms. The results represent mean values of two duplicate
experiments, and error bars indicate standard deviation.
Wound healing assays
For wound healing assays, 4×105
cells/well from each cell line (SiHa/con and SiHa/CLDN1,
SiHa/p63-pLKO and SiHa/p63-shCLDN1, ME-180/shp63-con and
ME-180/shp63-CLDN1) were seeded in 6-well plates. After the cells
were adhered, the cell monolayers were scratched using a pipette
tip and washed gently with PBS to remove cell debris. Subsequently,
all cells were cultured in DMEM or McCoy's 5A medium supplemented
with 1% FBS. In this experiment, the aim was to investigate the
migratory ability in the two cell lines (SiHa/con and SiHa/CLDN1).
Culturing cells in normal culture medium (with 10% FBS) can make it
difficult to distinguish whether cell migration or cell
proliferation are promoting wound healing (43). Therefore, DMEM or McCoy's 5A medium
supplemented with 1% FBS were used to culture cells. Cell migration
was assessed by measuring the movement of the cells into the
scratch in the well. Images were captured under a light microscope
(Olympus Corporation; magnification, ×100) at 24 and 48 h to
quantify the wound closure rate. Each experiment was performed in
triplicate.
Cell cycle analysis
SiHa/con and SiHa/CLDN1, SiHa/p63-pLKO and
SiHa/p63-shCLDN1, ME-180/shp63-con and ME-180/shp63-CLDN1 cells
were plated in 6-well plates at a density of 2×105
cells/well and cultured at 37°C overnight. Subsequently, all cells
were starved in serum-free DMEM for 48 h, after which they were
cultured with 10% FBS and DMEM for another 24 h at 37°C. The cells
in each group were fixed with 70% ethanol overnight at 4°C and
washed twice with PBS. Subsequently, the cells were re-suspended
with 400 µl binding buffer and stained with 20 µl PI
(Sigma-Aldrich; Merck KGaA) for 20 min at room temperature in the
dark. The cell cycle distribution was analyzed immediately using a
FACSCalibur flow cytometer (Becton, Dickinson and Company). The
percentage of cells at each phase of the cell cycle was calculated
using FlowJo 2.0 (FlowJo LLC). All assays were performed in
triplicate and repeated three times.
Flow cytometric cell death assay
SiHa/con and SiHa/CLDN1 cells were plated at a
density of 5×105 cells/well in 6-well plates and
cultured for 48 h at 37°C. Total cells were collected and washed
twice in binding buffer and stained with allophycocyanin-labeled
Annexin-V (BioLegend, Inc.) software in the dark for 20 min at room
temperature. Subsequently, the cells were stained with PI in the
dark for ~5 min at room temperature. The cells were analyzed using
a CytoFLEX flow cytometer (Beckman Coulter, Inc.) and then the data
was analyzed for percentage of viable, early apoptotic and late
apoptotic cells using FlowJo 2.0 software (FlowJo LLC). All assays
were performed in triplicate and repeated three times.
Tumorigenicity in mice
A total of 10 female BALB/c (nu/nu) mice (Shanghai
SLAC Laboratory Animal Co., Ltd.; age, 4 weeks; weight, 11.5g) were
kept in 12-h light/dark cycle at a temperature of 23°C under
specific pathogen-free atmosphere with 40-60% humidity and free
access to sufficient food and water. After 1 week, 100 µl
serum-free DMEM containing 6×106 SiHa/con or SiHa/CLDN1
cells (n=5 mice/group) were injected into the mouse right thigh.
The growth of solid tumors in mice was measured by Vernier calipers
every 3 days for up to 35 days. The tumor volume (V) was calculated
as follows: V=0.5 × length × width2. Subsequently, all
the mice were sacrificed by cervical dislocation at the end of the
experiment and the tumors were completely removed for analysis. The
animal experiment was approved by the Animal Care and Use Committee
of the University of Science and Technology of China.
RT-qPCR analysis
Total RNA was isolated from SiHa or ME-180 cells
using TRIzol® reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. RT was
performed using PrimeScript™ RT Reagent kit (Invitrogen; Thermo
Fisher Scientific, Inc.) following the manufacturer's protocol.
RT-qPCR was performed in a final volume of 20 µl containing 1 µl
cDNA, 1 µl primers (10 µM) and 10 µl SYBR Green PCR Master Mix
(Roche Diagnostics) in an ABI-7300 real-time PCR machine. The
following thermocycling conditions were used: 95°C for 5 min, 40
cycles at 94°C for 10 sec and 60°C for 31 sec, followed by the
dissociation stage at 95°C for 15 sec, 60°C for 60 sec and 95°C for
15 sec. The following primers (5′-3′) were used: GAPDH forward,
CTTCATTGACCTCAACTACATGG and reverse, CTCGCTCCTGGAAGATGGTGAT; and
CLDN1 forward, TCTGGGAGGTGCCCTACTTT and reverse,
CTGGAAGGTGCAGGTTTTGG. GAPDH was used as a control. Relative
expression of the mRNA was calculated using the 2−ΔΔCq
method (44) and normalized to
GAPDH.
Western blotting
Cell samples were collected and washed with PBS.
Proteins were extracted using RIPA lysis buffer (Beijing Solarbio
Science & Technology Co., Ltd.). Protein concentration was
quantified using a BCA Protein Assay kit (Thermo Fisher Scientific,
Inc.). Proteins (~30 µg/lane) were separated via 10% SDS-PAGE and
transferred onto PVDF membranes (EMD Millipore). The membranes were
blocked in 5% bovine serum albumin (Beijing Solarbio Science &
Technology Co., Ltd.; cat. no. SW3015) for 45 min at room
temperature and then probed with the indicated primary antibodies
at 4°C overnight. The following primary antibodies were used: p63
(dilution 1:1,000; Cell Signaling Technology, Inc.; cat. no.
39692), CLDN1 (dilution 1:1,000; Cell Signaling Technology, Inc.;
cat. no. 13255), PARP-1 (dilution 1:500; Santa Cruz Biotechnology
Inc.; cat. no. sc56197), cleaved PARP (dilution 1:500; Cell
Signaling Technology, Inc.; cat. no. 9548), caspase-3 (dilution
1:500; Cell Signaling Technology, Inc.; cat. no. 9662), caspase-7
(dilution 1:500; Cell Signaling Technology, Inc.; cat. no. 9492),
caspase-9 (dilution 1:1,000; Cell Signaling Technology, Inc.; cat.
no. 9502), cleaved caspase-9 (dilution 1:500; Cell Signaling
Technology, Inc.; cat. no. 20750), GAPDH (dilution 1:2,000;
ProteinTech Group, Inc.; cat. no. 10494-1-AP) and β-actin (dilution
1:2,000; ProteinTech Group, Inc.; cat. no. 20536-1-AP). The
secondary antibodies were HRP-conjugated goat anti-mouse IgG
(dilution 1:3,000; ProteinTech Group, Inc.; cat. no. SA00001-1) or
goat anti-rabbit IgG (dilution 1:3,000; ProteinTech Group, Inc.;
cat. no. SA00001-2). The appropriate secondary antibodies were
applied for 1 h at room temperature. The protein levels were
detected via enhanced chemiluminescence substrate (Advansta, Inc.;
cat. no. K-12045-D50.) and exposed to chemiluminescent film.
Statistical analysis
Data from all the experiments were presented as the
mean ± standard deviation using GraphPad Prism 6 (GraphPad
Software, Inc.), and were replicated at least three times.
Student's two-tailed unpaired t-tests were used to assess the
statistical significance of the differences between groups.
Survival curves were plotted using the Kaplan Meier-plotter and
analyzed using the log-rank test (http://kmplot.com/). P<0.05 was considered to
indicate a statistically significant difference.
Results
CLDN1 acts as an anti-oncogene in SiHa
cells
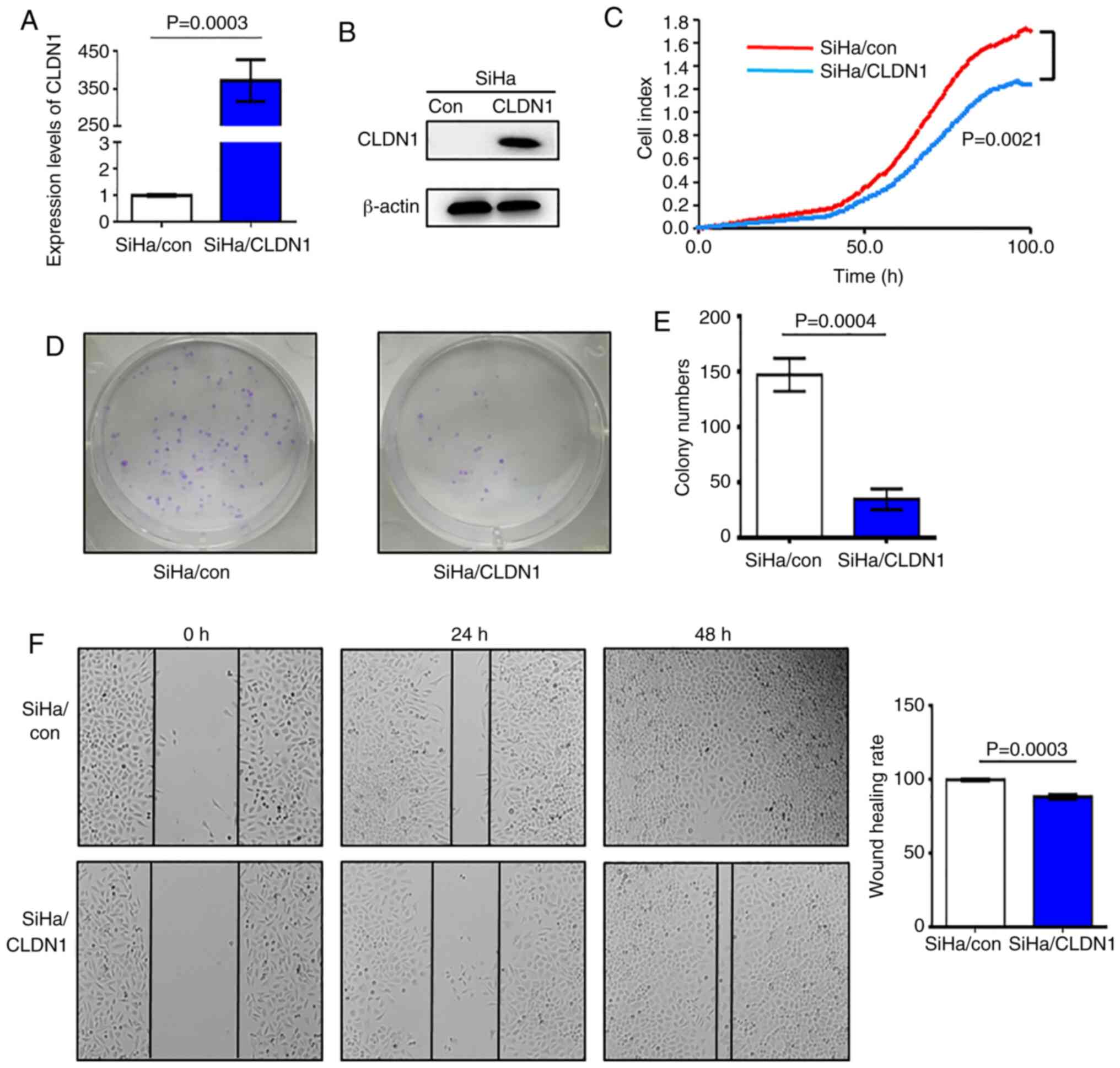
In order to study the effect of CLDN1 on cell
proliferation, SiHa/CLDN1 cell lines overexpressing CLDN1 were
constructed, and the control cell line was named SiHa/con. The
western blot and RT-qPCR analyses demonstrated that CLDN1
expression at both the mRNA and protein level in SiHa/CLDN1 cells
was higher compared with that in SiHa/con cells; therefore, stable
cell lines with CLDN1 overexpression were obtained (Fig. 1A and B). SiHa/con and SiHa/CLDN1
cells were seeded into an ACEA 16-well plate and cell proliferation
was measured. It was observed that overexpression of CLDN1
significantly suppressed cell proliferation compared with control
cells (Fig. 1C). Furthermore, the
number of cell colonies formed by SiHa/CLDN1 cells was
significantly decreased compared with that formed by SiHa/con cells
(Fig. 1D and E). In the wound
healing assays, after 48 h of culture, the wound healing rate of
SiHa/CLDN1 cells was significantly lower compared with that in the
SiHa/con cell group (Fig. 1F).
Overall, the present results indicated that CLDN1 inhibited SiHa
cervical cancer cell proliferation and migration.
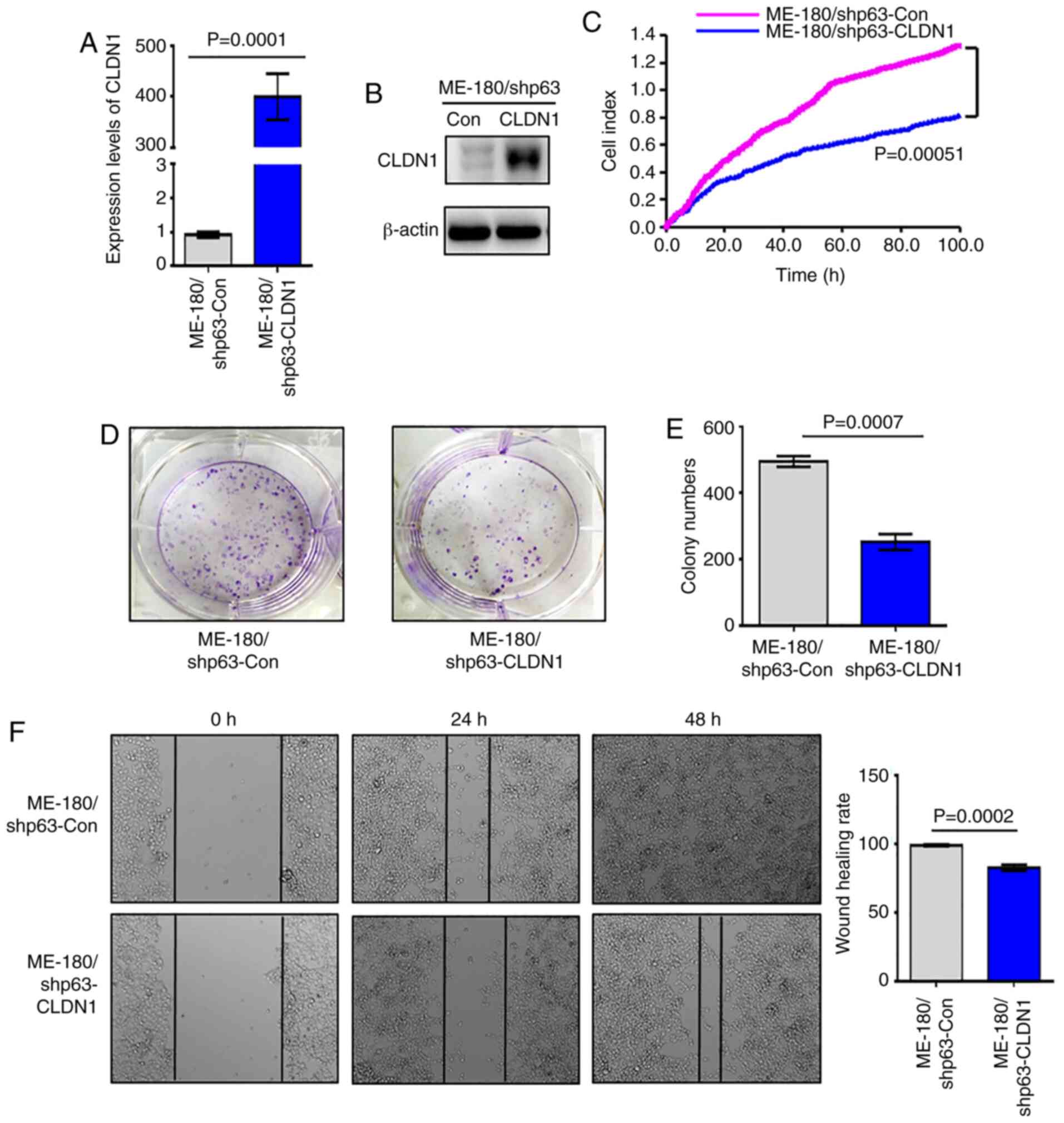
CLDN1 suppresses the proliferation of
ME-180/shp63 cells
Since the expression of CLDN1 is downregulated in
the ME-180/shp63 cell line (41),
in order to further study the function of CLDN1, a ME-180/shp63
cell line with CLDN1 overexpression was established and named
ME-180/shp63-CLDN1, whereas the control cell line was named
ME-180/shp63-con. The mRNA and protein levels of CLDN1 in the two
cell lines was then detected via western blotting and RT-qPCR
analysis. The results demonstrated that the expression levels of
CLDN1 in ME-180/shp63-CLDN1 cells were higher compared with those
in ME-180/shp63-con cells at both the mRNA and protein levels
(Fig. 2A and B). Subsequently, the
proliferation of the two cell lines was examined, and it was
observed that overexpression of CLDN1 in ME-180/shp63 cells
significantly suppressed their proliferation compared with control
cells (Fig. 2C). The number of
colonies formed by ME-180/shp63-CLDN1 cells was significantly
decreased compared with that formed by control cells (Fig. 2D and E). The wound healing assay
demonstrated that the wound healing rate of ME-180/shp63-CLDN1
cells was significantly lower compared with that of the control
cell group (Fig. 2F). Therefore,
CLDN1 was also found to act as an anti-oncogene in ME-180
cells.
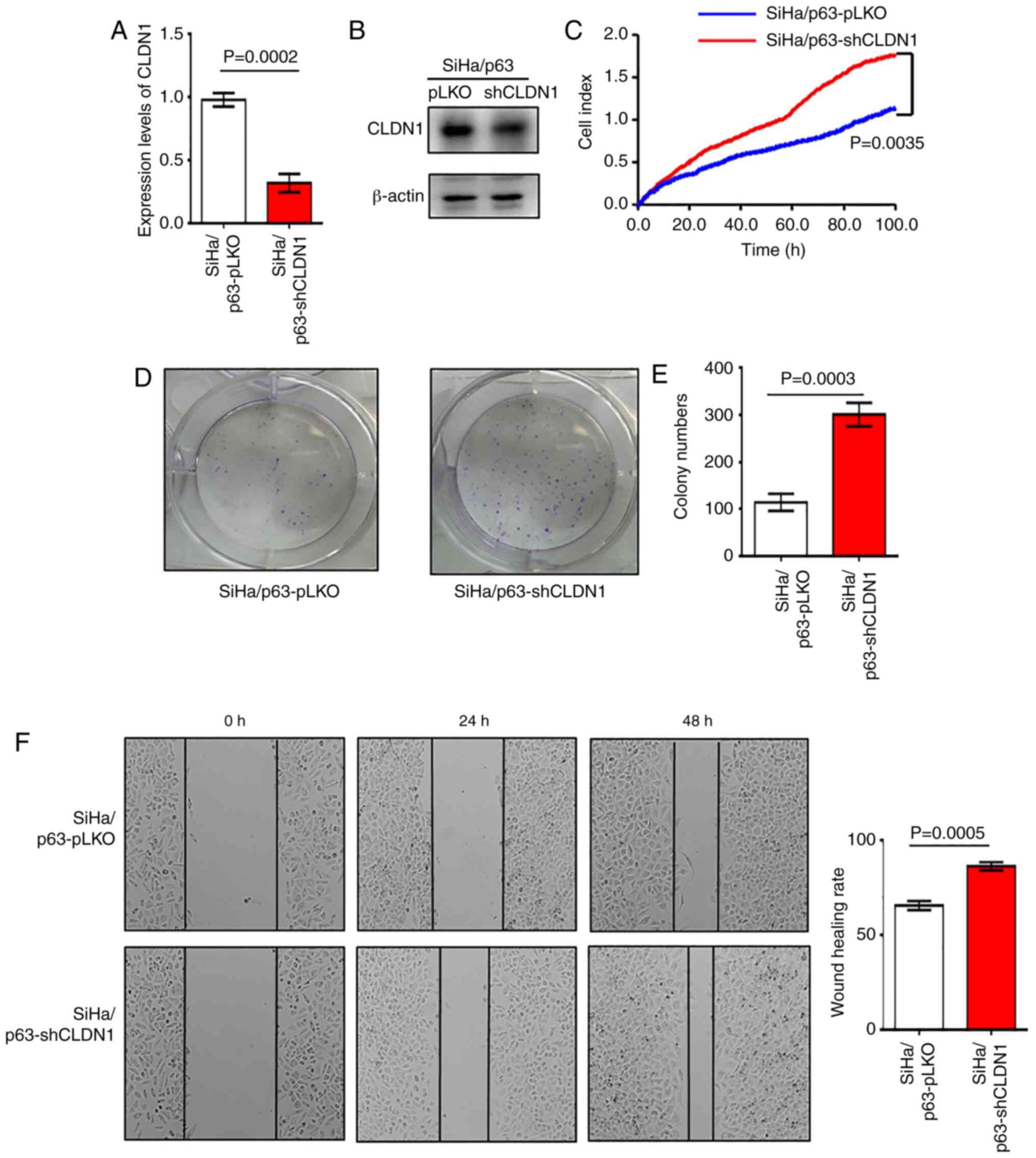
Knockdown of CLDN1 promotes the
proliferation of SiHa/p63 cells
Overexpression of CLDN1 exerted antitumor effects on
the SiHa and ME-180/shp63 cell lines. It was next determined
whether CLDN1 knockdown by shRNA could promote cervical cancer
progression. The SiHa/p63-shCLDN1 cell line was constructed and the
control cell line was named SiHa/p63-pLKO. The western blot and
RT-qPCR analyses revealed that CLDN1 expression was knocked down at
both the mRNA and protein levels in SiHa/p63-shCLDN1 cells compared
with the control cells (Fig. 3A and
B). The SiHa/p63-shCLDN1 cells exhibited increased
proliferative and colony-forming abilities compared with the
SiHa/p63-pLKO cells (Fig. 3C-E).
Additionally, knockdown of CLDN1 significantly increased cell
migration compared with that of control cells (Fig. 3F).
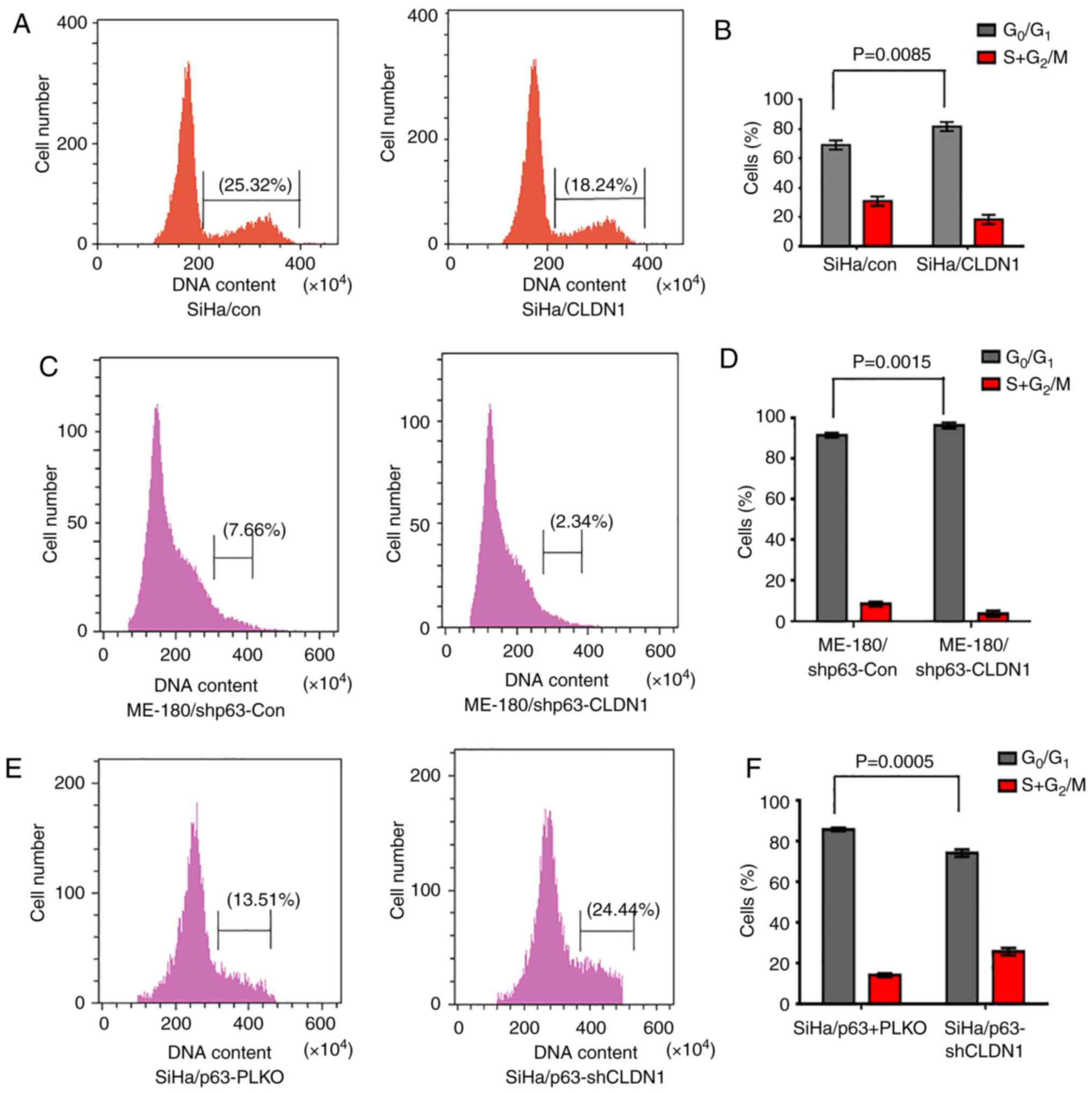
CLDN1 participates in cell cycle
regulation
In order to study the cell cycle distribution of the
3 groups of cell lines (SiHa/con and SiHa/CLDN1, ME-180/shp63-con
and ME-180/shp63-CLDN1, and SiHa/p63-pLKO and SiHa/p63-shCLDN1),
the cell cycle was examined by flow cytometry. The results
demonstrated that the proportion of SiHa/con and SiHa/CLDN1 cells
in the G1 phase was 67.42±1.26 and 81.54±1.35%,
respectively (Fig. 4A and B). The
cell cycle was primarily arrested at the G1 phase in the
SiHa/CLDN1 cell group compared with the SiHa/con cell group.
Similarly, changes in the cell cycle of ME-180 cells were detected,
and it was observed that the proportion of cells in the
G1 phase in the ME-180/shp63-con and ME-180/shp63-CLDN1
cell groups was 92.12±1.36 and 98.24±1.13%, respectively (Fig. 4C and D). Similarly, when CLDN1 was
knocked down in SiHa/p63 cells, the proportion of SiHa/p63-pLKO and
SiHa/p63-shCLDN1 cells in the G1 phase was 86.22±1.14
and 75.21±1.25%, respectively (Fig. 4E
and F). The current results demonstrated that CLDN1 affected
cell cycle distribution and arrested cells in the G1
phase.
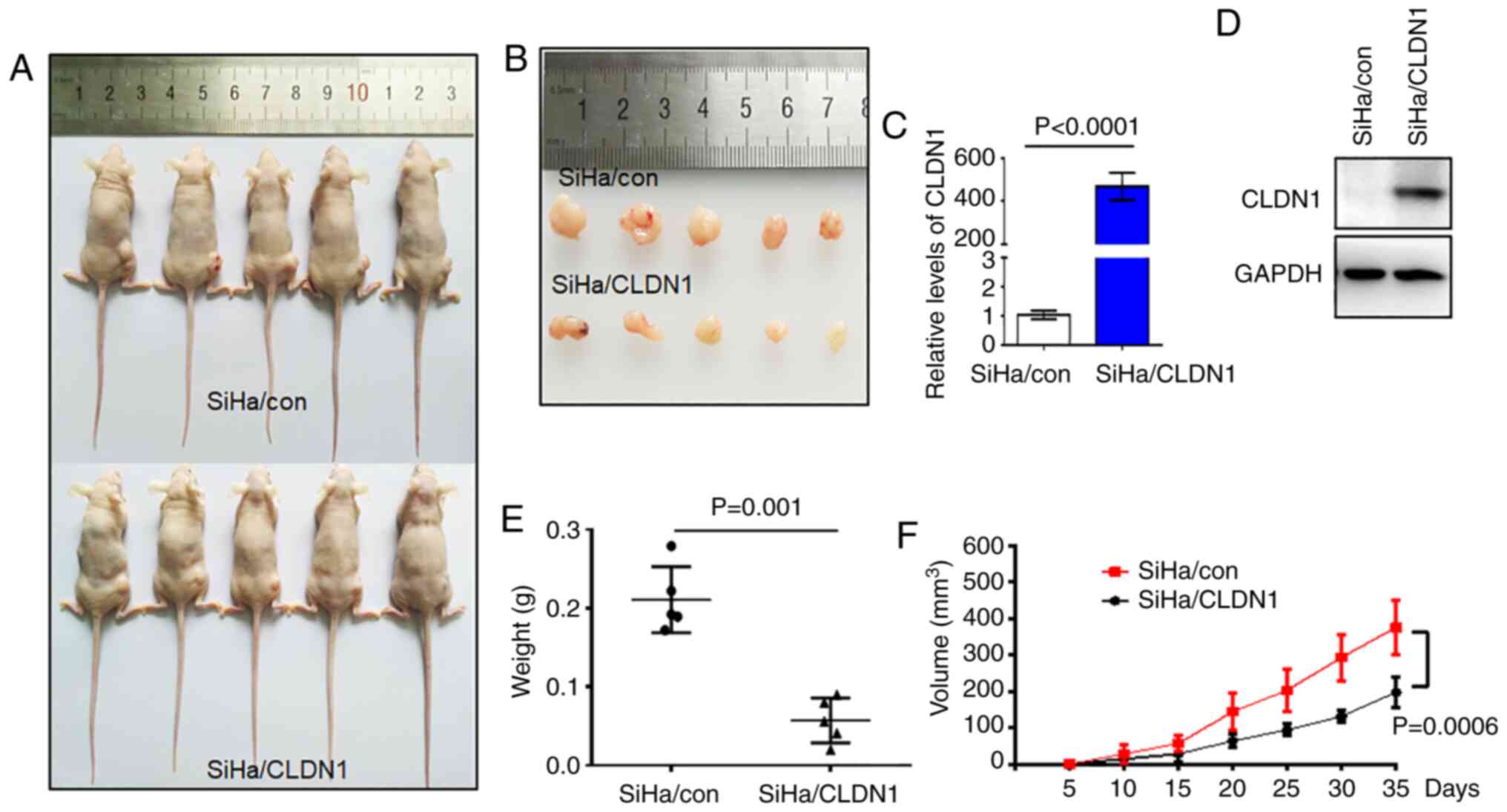
CLDN1 inhibits tumor growth in
vivo
Based on the in vitro results, the present
study next sought to verify the role of CLDN1 in cell proliferation
in vivo. A tumor-bearing mouse model was established by
transplanting SiHa/con and SiHa/CLDN1 cells. The tumor sizes are
shown in Fig. 5A and B. The
expression levels of CLDN1 in the two tumor groups were confirmed
by western blot and RT-qPCR analyses (Fig. 5C and D). Tumor size and weight in
the SiHa/con group were significantly higher compared with those in
the SiHa/CLDN1 group (Fig. 3E and
F). As expected, CLDN1 overexpression in SiHa cells inhibited
tumor growth in vivo compared with control tumors.
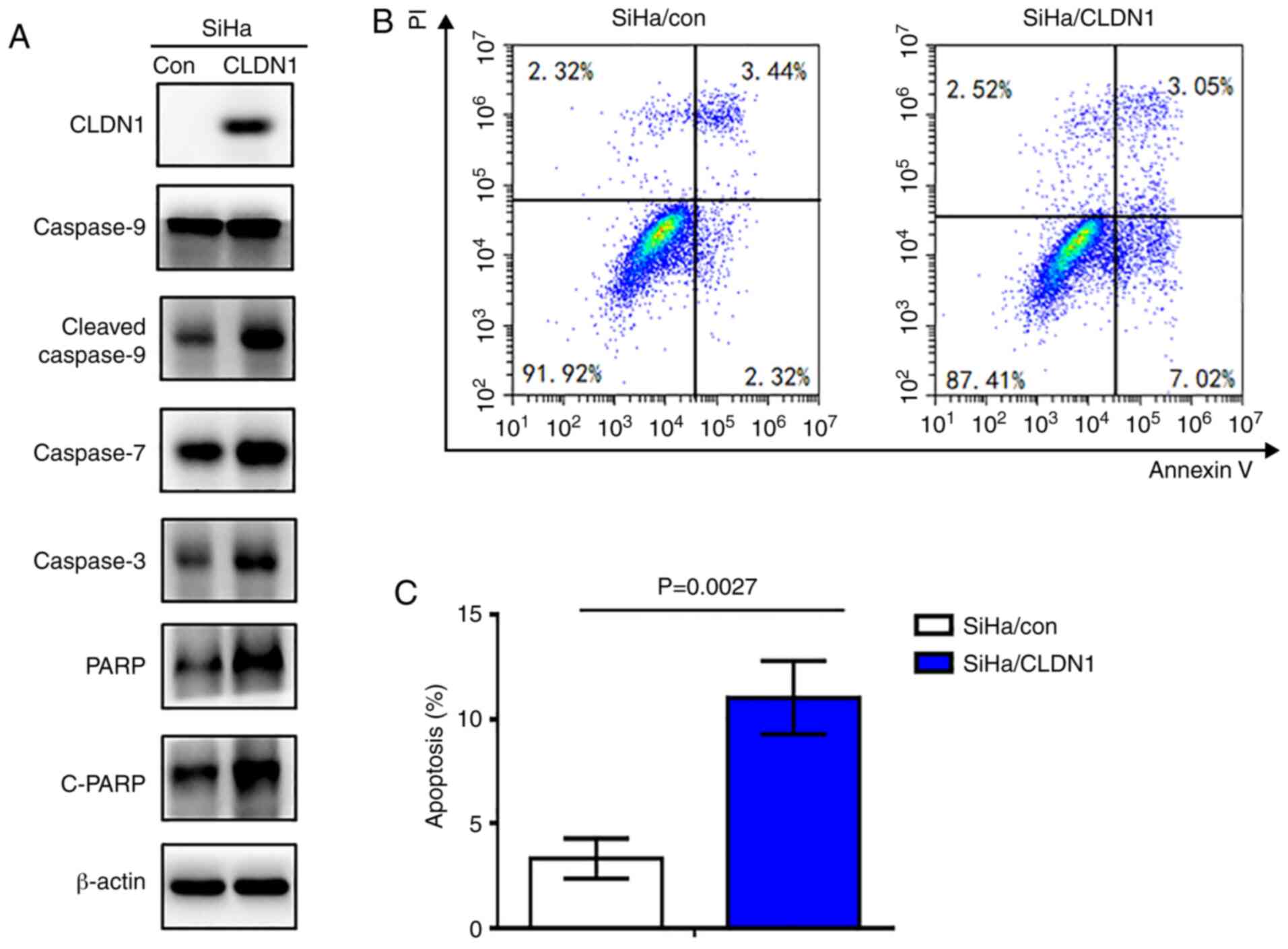
CLDN1 promotes apoptosis in cervical
cancer cells
To investigate the role of CLDN1 in cell apoptosis,
the expression levels of apoptosis-associated proteins were
detected in the SiHa/CLDN1 and SiHa/con cell groups by western
blotting. The results indicated that the expression levels of PARP,
cleaved-PARP, caspase-3, 7 and cleaved 9 in SiHa/CLDN1 cells were
markedly increased compared with those in SiHa/con cells (Fig. 6A). Additionally, flow cytometric
cell death assay was performed. As shown in Fig. 6B and C, cell apoptosis was
significantly increased in SiHa cells with CLDN1 overexpression
(P=0.0027).
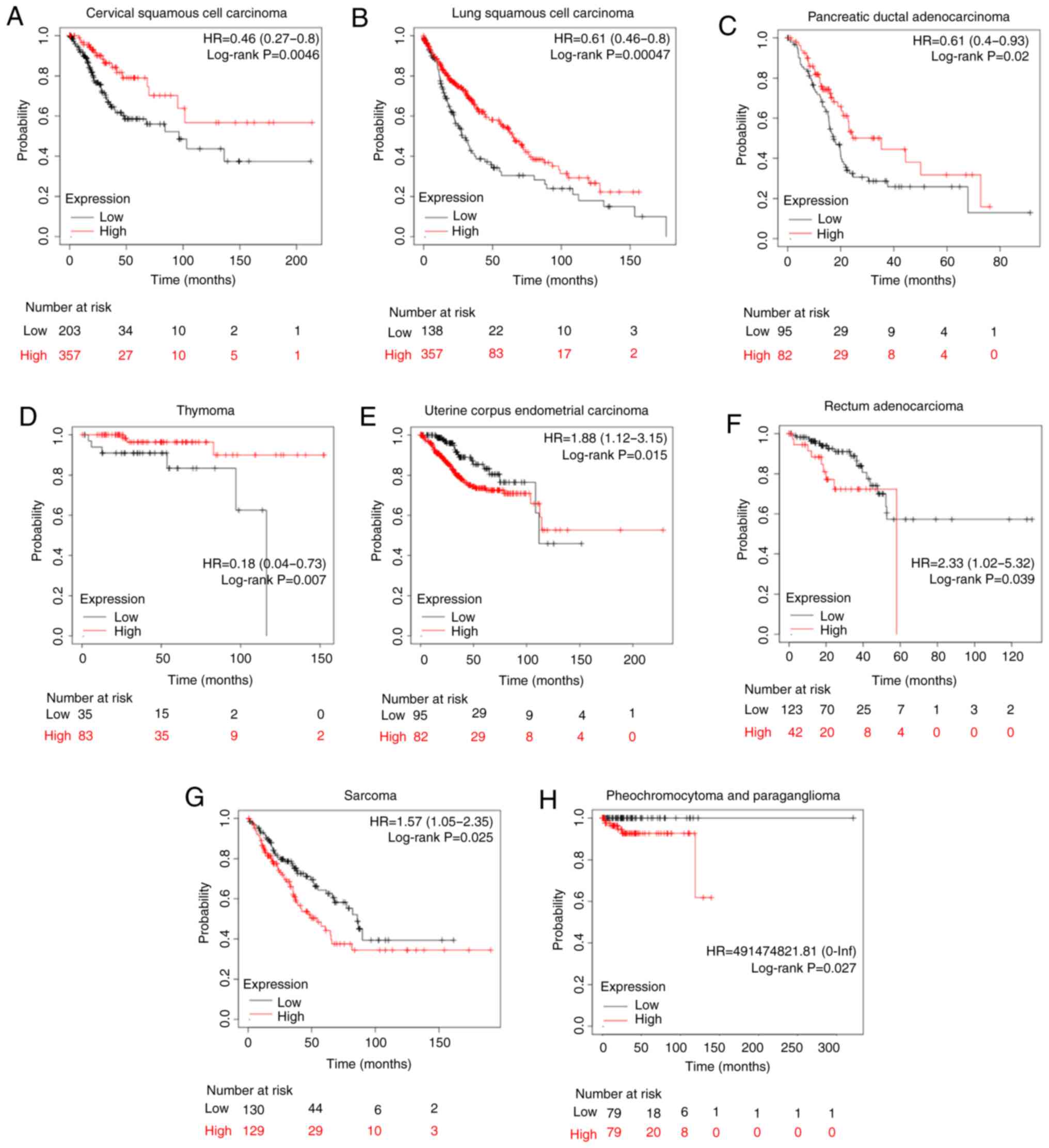
Association of CLDN1 expression with
the prognosis in other types of human tumor
To further study the effect of CLDN1 on patient
prognosis, CLDN1 expression in other types of tumor was analyzed
using the Kaplan-Meier plotter in the pan-cancer RNA-Seq mRNA
database. Sources for the databases include Gene Expression
Omnibus, the European Genome-phenome Archive and The Cancer Genome
Atlas (http://kmplot.com/analysis), and there
are a total of 21 types of cancer that can be analyzed, including
bladder carcinoma, ovarian cancer and head and neck squamous cell
carcinoma. In cervical cancer, the upper quartile survival of the
high CLDN1 expression cohort was 68.4 months, while in the low
CLDN1 expression cohort it was 27.63 months (P=0.0046; Fig. 7A), which was consistent with our
previous study (41). Additionally,
CLDN1 expression was found to be associated with prognosis in seven
other types of tumor, as shown in Fig.
7. High CLDN1 expression was directly associated with an
improved overall survival in patients with lung squamous cell
carcinoma, pancreatic ductal adenocarcinoma and thymoma (Fig. 7B-D). However, high CLDN1 expression
was directly associated with a poor prognosis in patients with
uterine corpus endometrial carcinoma, rectal adenocarcinoma,
pheochromocytoma and paraganglioma, and sarcoma (Fig. 7E-H). Therefore, the role of CLDN1
differed across different types of tumor.
Discussion
CLDN1 is a member of the claudins, which is a family
of 17-27 kDa integral membrane tight junction proteins that
determine the size of the molecules that pass through the
paracellular space in epithelial and endothelial tissues (36). There are limited data available on
the molecular interactions among the claudins. The expression
levels of CLDN1 have been investigated in several types of cancer
and different expression levels of CLDN1 have been associated with
tumorigenesis in different types of cancer (45). Downregulation of CLDN1 leads to
characteristic morphological alterations of tight junction fine
structure (46). It has been
demonstrated that loss of CLDN1 expression stimulates tumor
progression and invasion in certain types of cancer, such as
melanocytic neoplasia (47), while
in esophageal squamous cell carcinoma, increased CLDN1 expression
was associated with tumor progression (48–50).
For example, Youssefian et al (51) reported that a novel mutation in
CLDN1 was identified in neonatal ichthyosis and was associated with
sclerosing cholangitis, referred to as the NISCH syndrome, which
may be used for molecular confirmation of the diagnosis in some of
these patients. Furthermore, polymorphisms in the CLDN1 gene were
associated with the age of patients and differentiation of
triple-negative breast cancer (TNBC), but there was no association
between polymorphisms in CLDN1 and survival of patients with TNBC
(52). However, the function of
CLDN1 may vary even in the same type of cancer, such as breast
cancer (52). Loss of CLDN1
expression in patients with breast cancer was associated with tumor
invasion and metastasis by Tokés et al (53), whereas CLDN1 contributed to
migration of luminal-like MCF7 human breast cancer cells (54).
In our previous study, the results demonstrated that
the transcription factor ΔNp63α serves an antitumor role in
cervical squamous cell cancer (41). CLDN1 was found to be a direct target
of ΔNp63α by Chip-seq and RNA-Seq, which was verified by western
blot and RT-qPCR analysis. Additionally, the expression levels of
ΔNp63α and CLDN1 were significantly decreased in cervical tumor
samples compared with in adjacent normal tissues (41). Thus, it was inferred that p63 may
inhibit the progression of cervical cancer by regulating CLDN1, but
its definitive function in cervical cancer remains elusive.
Therefore, the current study further investigated the function of
CLDN1 in cervical cancer cells. In the present study, rescue
experiments were performed by knocking down CLDN1 in SiHa/p63 cells
and overexpressing CLDN1 in ME-180/shp63 cells, and the results
demonstrated that CLDN1 inhibited the proliferation and migration
of cervical cancer cells. Additionally, the in vivo
experiments in tumor-bearing mice revealed that the overexpression
of CLDN1 inhibited cell proliferation. Anticancer cytotoxic
activity was mostly mediated via arresting the cell cycle at the
G0/G1 phases, thereby inhibiting cell
proliferation, and eventually leading to apoptosis. Based on the
present findings, CLDN1 appeared to inhibit cervical cancer growth
by inducing cancer cell apoptosis. The possible underlying
mechanism may involve multiple factors or signaling pathways,
including Smad, β-catenin, the Snail family and the ERK1/2,
PI3K/AKT and AMPK signaling pathways (55,56).
In summary, the present study revealed that CLDN1
may act as a tumor suppressor gene in cervical cancer. However, the
underlying mechanisms requires further investigation in future
studies. The current findings provide a strong basis for the
further exploration of CLDN1 as a new therapeutic target for
cervical cancer.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Key
R&D Program of China (grant no. 2018YFC1003900), the National
Natural Science Foundation of China (grant nos. 81902632, 81872110
and 81272881) and the Fundamental Research Funds for the Central
Universities (grant no. WK9110000104).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
ZS and WS performed the experiments and analyzed the
data. LQ constructed the stable overexpression cell lines and wrote
part of the manuscript. YL and JZ prepared the figures. ML
performed the statistical analyses. TZ and WZ assisted with the
experiments and data interpretation. YZ performed most of the
revision experiments. XY designed the majority of the experiments
and wrote the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All experiments involving animals were performed in
accordance with the guidelines of the Animal Care and Use Committee
of the University of Science & Technology of China using a
peer-reviewed protocol (approval no. USTCACUC1801017).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Underwood SM, Ramsay-Johnson E, Browne L,
Caines N, Dean A, Duval S, Ivalis R, Lawrence ND, Lewis N, Mulkanen
M, et al: What women in the United States Virgin Islands still want
and need to know about HPV, cervical cancer, and condom use. J Natl
Black Nurses Assoc. 21:25–32. 2010.PubMed/NCBI
|
|
3
|
Malagón T, Kulasingam S, Mayrand MH,
Ogilvie G, Smith L, Bouchard C, Gotlieb W and Franco EL: Age at
last screening and remaining lifetime risk of cervical cancer in
older, unvaccinated, HPV-negative women: A modelling study. Lancet
Oncol. 19:1569–1578. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hoevel T, Macek R, Swisshelm K and Kubbies
M: Reexpression of the TJ protein CLDN1 induces apoptosis in breast
tumor spheroids. Int J Cancer. 108:374–383. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bjurberg M, Beskow C, Kannisto P and
Lindahl G: Cervical cancer is a clinical challenge. Lakartidningen.
112:2015.(In Swedish). PubMed/NCBI
|
|
6
|
Hu Z and Ma D: The precision prevention
and therapy of HPV-related cervical cancer: New concepts and
clinical implications. Cancer Med. 7:5217–5236. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fang J, Zhang H and Jin S: Epigenetics and
cervical cancer: From pathogenesis to therapy. Tumour Biol.
35:5083–5093. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Goodman A: HPV testing as a screen for
cervical cancer. BMJ. 350:h23722015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chrysostomou AC, Stylianou DC,
Constantinidou A and Kostrikis LG: Cervical cancer screening
programs in Europe: The transition towards HPV vaccination and
population-based HPV testing. Viruses. 10:7292018. View Article : Google Scholar
|
|
10
|
Vu M, Yu J, Awolude OA and Chuang L:
Cervical cancer worldwide. Curr Probl Cancer. 42:457–465. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wuerthner BA and Avila-Wallace M: Cervical
cancer: Screening, management, and prevention. Nurse Pract.
41:18–23. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Mountzios G, Soultati A, Pectasides D,
Pectasides E, Dimopoulos MA and Papadimitriou CA: Developments in
the systemic treatment of metastatic cervical cancer. Cancer Treat
Rev. 39:430–443. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zihni C, Mills C, Matter K and Balda MS:
Tight junctions: From simple barriers to multifunctional molecular
gates. Nat Rev Mol Cell Biol. 17:564–580. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tsukita S, Tanaka H and Tamura A: The
claudins: From tight junctions to biological systems. Trends
Biochem Sci. 44:141–152. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tsukita S, Furuse M and Itoh M:
Multifunctional strands in tight junctions. Nat Rev Mol Cell Biol.
2:285–293. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Suzuki T: Regulation of intestinal
epithelial permeability by tight junctions. Cell Mol Life Sci.
70:631–659. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sawada N: Tight junction-related human
diseases. Pathol Int. 63:1–12. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Campbell HK, Maiers JL and DeMali KA:
Interplay between tight junctions & adherens junctions. Exp
Cell Res. 358:39–44. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Amarnath S and Agarwala S:
Cell-cycle-dependent TGFβ-BMP antagonism regulates neural tube
closure by modulating tight junctions. J Cell Sci. 130:119–131.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang X, Adegoke EO, Ma M, Huang F, Zhang
H, Adeniran SO, Zheng P and Zhang G: Influence of Wilms' tumor
suppressor gene WT1 on bovine Sertoli cells polarity and tight
junctions via non-canonical WNT signaling pathway. Theriogenology.
138:84–93. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ruch TR and Engel JN: Targeting the
mucosal barrier: How pathogens modulate the cellular polarity
network. Cold Spring Harb Perspect Biol. 9:a0279532017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li X, Ahmad US, Huang Y, Uttagomol J,
Rehman A, Zhou K, Warnes G, McArthur S, Parkinson EK and Wan H:
Desmoglein-3 acts as a pro-survival protein by suppressing reactive
oxygen species and doming whilst augmenting the tight junctions in
MDCK cells. Mech Ageing Dev. 184:1111742019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tsugami Y, Matsunaga K, Suzuki T,
Nishimura T and Kobayashi K: Phytoestrogens weaken the blood-milk
barrier in lactating mammary epithelial cells by affecting tight
junctions and cell viability. J Agric Food Chem. 65:11118–11124.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
De Pascalis C and Etienne-Manneville S:
Single and collective cell migration: The mechanics of adhesions.
Mol Biol Cell. 28:1833–1846. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Calder MD, Edwards NA, Betts DH and Watson
AJ: Treatment with AICAR inhibits blastocyst development,
trophectoderm differentiation and tight junction formation and
function in mice. Mol Hum Reprod. 23:771–785. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ryu WI, Lee H, Bae HC, Jeon J, Ryu HJ, Kim
J, Kim JH, Son JW, Kim J, Imai Y, et al: IL-33 down-regulates CLDN1
expression through the ERK/STAT3 pathway in keratinocytes. J
Dermatol Sci. 90:313–322. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Baktash Y, Madhav A, Coller KE and Randall
G: Single particle imaging of polarized hepatoma organoids upon
hepatitis C virus infection reveals an ordered and sequential entry
process. Cell Host Microbe. 23:382–394.e5. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu H, Jiang F, Jia X, Lan J, Guo H, Li E,
Yan A and Wang Y: Cycling hypoxia affects cell invasion and
proliferation through direct regulation of claudin1/claudin7
expression, and indirect regulation of P18 through claudin7.
Oncotarget. 8:10298–10311. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bhat AA, Syed N, Therachiyil L, Nisar S,
Hashem S, Macha MA, Yadav SK, Krishnankutty R, Muralitharan S,
Al-Naemi H, et al: Claudin-1, a double-edged sword in cancer. Int J
Mol Sci. 21:5692020. View Article : Google Scholar
|
|
30
|
Dehghan Esmatabadi MJ, Farhangi B, Safari
Z, Kazerooni H, Shirzad H, Zolghadr F and Sadeghizadeh M:
Dendrosomal curcumin inhibits metastatic potential of human SW480
colon cancer cells through Down-regulation of Claudin1, Zeb1 and
Hef1-1 gene expression. Asian Pac J Cancer Prev. 16:2473–2481.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Eftang LL, Esbensen Y, Tannæs TM, Blom GP,
Bukholm IR and Bukholm G: Up-regulation of CLDN1 in gastric cancer
is correlated with reduced survival. BMC Cancer. 13:5862013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu J, Gao F, Xu T, Li J, Hu Z, Wang C,
Long Y, He X, Deng X, Ren D, et al: CLDN1 induces autophagy to
promote proliferation and metastasis of esophageal squamous
carcinoma through AMPK/STAT1/ULK1 signaling. J Cell Physiol.
235:2245–2259. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Chao YC, Pan SH, Yang SC, Yu SL, Che TF,
Lin CW, Tsai MS, Chang GC, Wu CH, Wu YY, et al: Claudin-1 is a
metastasis suppressor and correlates with clinical outcome in lung
adenocarcinoma. Am J Respir Crit Care Med. 179:123–133. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sheehan GM, Kallakury BV, Sheehan CE,
Fisher HA, Kaufman RP Jr and Ross JS: Loss of claudins-1 and −7 and
expression of claudins-3 and −4 correlate with prognostic variables
in prostatic adenocarcinomas. Hum Pathol. 38:564–569. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Emmanouilidi A, Paladin D, Greening DW and
Falasca M: Oncogenic and non-malignant pancreatic exosome cargo
reveal distinct expression of oncogenic and prognostic factors
involved in tumor invasion and metastasis. Proteomics.
19:e18001582019. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sun BS, Yao YQ, Pei BX, Zhang ZF and Wang
CL: Claudin-1 correlates with poor prognosis in lung
adenocarcinoma. Thorac Cancer. 7:556–563. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mahati S, Xiao L, Yang Y, Mao R and Bao Y:
miR-29a suppresses growth and migration of hepatocellular carcinoma
by regulating CLDN1. Biochem Biophys Res Commun. 486:732–737. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hoellen F, Waldmann A, Banz-Jansen C,
Holtrich U, Karn T, Oberländer M, Habermann JK, Hörmann M, Köster
F, Ribbat-Idel J, et al: Claudin-1 expression in cervical cancer.
Mol Clin Oncol. 7:880–884. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Zhang WN, Li W, Wang XL, Hu Z, Zhu D, Ding
WC, Liu D, Li KZ, Ma D and Wang H: CLDN1 expression in cervical
cancer cells is related to tumor invasion and metastasis.
Oncotarget. 7:87449–87461. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cunniffe C, Brankin B, Lambkin H and Ryan
F: The role of claudin-1 and claudin-7 in cervical tumorigenesis.
Anticancer Res. 34:2851–2857. 2014.PubMed/NCBI
|
|
41
|
Zhou Y, Liu H, Wang J, Wang X, Qian L, Xu
F, Song W, Wu D, Shen Z, Feng D, et al: ΔNp63α exerts antitumor
functions in cervical squamous cell carcinoma. Oncogene.
39:905–921. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Soares E and Zhou H: Master regulatory
role of p63 in epidermal development and disease. Cell Mol Life
Sci. 75:1179–1190. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hsiao YH, Hsieh MJ, Yang SF, Chen SP, Tsai
WC and Chen PN: Phloretin suppresses metastasis by targeting
protease and inhibits cancer stemness and angiogenesis in human
cervical cancer cells. Phytomedicine. 62:1529642019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kuo KT, Chen CL, Chou TY, Yeh CT, Lee WH
and Wang LS: Nm23H1 mediates tumor invasion in esophageal squamous
cell carcinoma by regulation of CLDN1 through the AKT signaling.
Oncogenesis. 5:e2392016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liebner S, Fischmann A, Rascher G, Duffner
F, Grote EH, Kalbacher H and Wolburg H: Claudin-1 and claudin-5
expression and tight junction morphology are altered in blood
vessels of human glioblastoma multiforme. Acta Neuropathol.
100:323–331. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hammer SC, Nagel S, Junginger J,
Hewicker-Trautwein M, Wagner S, Heisterkamp A, Ngezahayo A, Nolte I
and Murua Escobar H: Claudin-1, −3, −4 and −7 gene expression
analyses in canine prostate carcinoma and mammary tissue derived
cell lines. Neoplasma. 63:231–238. 2016.PubMed/NCBI
|
|
48
|
Feng S, Zhai J, Lu D, Lin J, Dong X, Liu
X, Wu H, Roden AC, Brandi G, Tavolari S, et al: TUSC3 accelerates
cancer growth and induces epithelial-mesenchymal transition by
upregulating claudin-1 in non-small-cell lung cancer cells. Exp
Cell Res. 373:44–56. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Upmanyu N, Bulldan A, Papadopoulos D,
Dietze R, Malviya VN and Scheiner-Bobis G: Impairment of the
Gnα11-controlled expression of claudin-1 and MMP-9 and collective
migration of human breast cancer MCF-7 cells by DHEAS. J Steroid
Biochem Mol Biol. 182:50–61. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Torres-Martínez AC, Gallardo-Vera JF,
Lara-Holguin AN, Montaño LF and Rendón-Huerta EP: Claudin-6
enhances cell invasiveness through claudin-1 in AGS human
adenocarcinoma gastric cancer cells. Exp Cell Res. 350:226–235.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Youssefian L, Vahidnezhad H, Saeidian AH,
Sotoudeh S, Zeinali S and Uitto J: Gene-targeted next-generation
sequencing identifies a novel CLDN1 mutation in a consanguineous
family with NISCH syndrome. Am J Gastroenterol. 112:396–398. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Hu A, Li J, Ruan S, Fan Y and Liao Y:
Polymorphisms in CLDN1 are associated with age and differentiation
of triple-negative breast cancer patients. Biosci Rep.
39:BSR201819522019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Tokés AM, Kulka J, Paku S, Szik A, Páska
C, Novák PK, Szilák L, Kiss A, Bögi K and Schaff Z: Claudin-1, −3
and −4 proteins and mRNA expression in benign and malignant breast
lesions: A research study. Breast Cancer Res. 7:R296–R305. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhou B, Blanchard A, Wang N, Ma X, Han J,
Schroedter I, Leygue E and Myal Y: Claudin 1 promotes migration and
increases sensitivity to tamoxifen and anticancer drugs in
luminal-like human breast cancer cells MCF7. Cancer Inves.
33:429–439. 2015. View Article : Google Scholar
|
|
55
|
Chen Yh, Lu Q, Schneeberger EE and
Goodenough DA: Restoration of tight junction structure and barrier
function by down-regulation of the mitogen-activated protein kinase
pathway in ras-transformed Madin-Darby canine kidney cells. Mol
Biol Cell. 11:849–862. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Lippoldt A, Liebner S, Andbjer B,
Kalbacher H, Wolburg H, Haller H and Fuxe K: Organization of
choroid plexus epithelial and endothelial cell tight junctions and
regulation of claudin-1, −2 and −5 expression by protein kinase C.
Neuroreport. 11:1427–1431. 2000. View Article : Google Scholar : PubMed/NCBI
|