Introduction
Triple-negative breast cancer (TNBC) is a subtype of
breast cancer that lacks the expression of the human epidermal
growth factor receptor 2 (HER2), estrogen and progesterone
receptors (1,2). Metastasis is a major clinical feature
of late stage TNBC, and is the main cause of death in patients
(3,4). It has been widely recognized that the
migration and invasion of TNBC cells are considered to be key steps
in the metastatic process (5). One
study revealed that TNBC cells with high migratory and high
invasive properties are closely associated with a poor prognosis in
TNBC (6). In a previous study,
thymoquinone, an active ingredient of Nigella sativa,
markedly inhibited cell growth, migration and invasion in both TNBC
cells and orthotopic TNBC model mice by regulating the elongation
factor 2 kinase (eEF-2K) signaling axis (7). Furthermore, in another study,
luteolin, a natural flavonoid compound, reportedly suppressed the
epithelial-mesenchymal transition (EMT) and migration of TNBC cells
by inhibiting YAP/TAZ activity (8).
These findings suggest that preventing the migration and invasion
of TNBC cells may inhibit TNBC metastasis and represent a target
for TNBC treatment.
EMT is a biological process in which epithelial
cells lose key proteins (such as E-cadherin) involved in cell
junctions and obtain a mesenchymal phenotype, which is evidenced by
upregulated protein expression levels of mesenchymal markers (such
as vimentin) (9–11). The activation of EMT has been found
to serve an important role in the initial stages of the metastatic
cascade by enhancing the migratory and invasive abilities of cancer
cells (12). It has been
demonstrated that irisin (a novel myokine) could inhibit the EMT
process by suppressing the migration and invasion of MIA PaCa-2 and
Panc03.27 cells (13). In addition,
isorhamnetin could inhibit migration and invasion by suppressing
AKT/ERK-mediated EMT in A549 human non-small cell lung cancer cells
(14). These aforementioned
findings suggest that blocking the process of EMT may represent a
potential approach to inhibit the migration and invasion of TNBC
cells.
AMP-activated protein kinase (AMPK), a vital
metabolic energy sensor that regulates protein and lipid metabolism
responses, has been demonstrated to serve an important role in the
EMT process associated with tumor metastasis (15–19).
Increasing evidence has revealed that AMPK is inactivated in TNBC
cells, and that the absence of AMPK in patients predicted a poor
prognosis (20,21). Conversely, the activation of AMPK
could inhibit the EMT process of tumor cells by regulating the
expression levels of EMT-related markers (22). For example, metformin could suppress
the EMT process in TNBC cells by activating the AMPK signaling
pathway (23). Similarly,
rosmarinic acid was found to suppress the metastatic
characteristics of colorectal cancer (CRC) cells by activating AMPK
(24). These previous studies
suggest that the activation of AMPK may be a target for the
inhibition of metastasis and the EMT process in TNBC.
Peroxisome proliferator-activated receptor γ
(PPARγ), a ligand-dependent transcription factor that regulates
lipid metabolism, has been proven to participate in tumor
metastasis (25). Accumulating
evidence indicates that PPARγ is inactivated in TNBC cells, and its
activation may inhibit the metastasis of breast cancer cells
(26,27). VSP-17 has been identified as a novel
PPARγ agonist, and it has been shown to suppress the metastasis of
TNBC by upregulating the expression levels of E-cadherin (28). However, the mechanism through which
VSP-17 may inhibit TNBC metastasis remains unclear. The present
study aimed to determine the mechanism of action of the
VSP-17-induced upregulation of E-cadherin levels in inhibiting TNBC
metastasis, with a focus on the EMT and PPARγ/AMPK signaling
pathways.
Materials and methods
Reagents
VSP-17
(C23H18F3N3O, MW:
409.1402, purity ≥98%) was designed and synthesized by our team
(Guangxi Colleges and Universities Key Laboratory of Pharmacology),
the details of which have been described in our previous study
(28). siAMPK (cat. no. 203834) and
siPPARγ (cat. no. 202738) were obtained from Sangon Biotech;
Lipofectamine 3000 reagents (cat. no. 2149659) were purchased from
Thermo Fisher Scientific, Inc., and compound C (cat. no.
866405-64-3) and GW9662 (cat. no. 22978-75-2) were purchased from
MedChemExpress. Primary antibodies against the following targets:
AMPK (cat. no. BS6271), p-AMPK (cat. no. BS4010), E-cadherin (cat.
no. BS1098) and vimentin (cat. no. BS1491) were supplied by
Bioworld Technology, Inc.; glyceraldehyde-3-phosphate dehydrogenase
(GAPDH, cat. no. 60004-1-Ig) was obtained from ProteinTech Group,
Inc. HiScript II One-Step RT-PCR Kit (cat. no. R323-01) and ChamQ
SYBR qPCR Master Mix (cat. no. Q311-02) were purchased from Vazyme;
4′,6-diamidino-2-phenylindole (DAPI; cat. no. C0060) was obtained
from Beijing Solarbio Science & Technology Co., Ltd. Other
analytical reagent grade chemicals were purchased from Chemical
Reagent Co. Ltd.
Cell culture
MDA-MB-231 and MDA-MB-453 cells were obtained from
the American Type Culture Collection (ATCC) and supplemented with
Leibovitz's (L-15) medium (cat. no. 11415064; Thermo Fisher
Scientific, Inc.) with 10% fetal bovine serum (FBS) and 1%
penicillin-streptomycin in 5% CO2, 95% air cell culture
at 37°C. The cells were starved for 2 h before the experiments.
Cell migration assay
According to the manufacturer's protocol, the cell
migration assay was carried out using Transwell plates.
Post-treated cells or compound C co-cultured cells
(1×105) were detached and suspended in the culture
medium. Then, the cells were added to the upper chamber of the
Transwell plates, while the lower chamber was filled with 600 µl of
culture medium containing 10% FBS as a chemical attractant. After
being incubated for 6 h, the non-migratory cells on the upper
surface of the membrane were removed with a soaked cotton swab. In
addition, the cells that migrated to the bottom face of the
membranes were counted after being stained with 0.5% crystal violet
solutions at 37°C for 30 min. Then, 5 microscopic fields per filter
were captured randomly with the help of an inverted microscope
(Olympus, Tokyo, Japan; ×200 magnification).
Cell invasion assay
The invasion potential of the MDA-MB-231 and
MDA-MB-453 cells was evaluated using a Matrigel-coated Transwell
chamber with 8.0-µm pore size. Post-transfected cells or compound C
co-cultured cells (1×105) were resuspended with L-15
medium and plated into the upper chamber, and L-15 medium
containing 20% FBS was added to the lower chambers as an
attractant. The suspension was discarded after 24 h, counting the
cells which invaded the lower chamber under an inverted microscope
(Olympus, Tokyo, Japan; ×200 magnification).
Cell transfection
Under the guidance of the manufacturer's
instructions, Lipofectamine 2000 and small interfering RNA (siRNA)
duplex specific for AMPK and PPARγ were used to transiently
transfect MDA-MB-231 and MDA-MB-453 cells for 6 h at 37°C, and then
the cells were incubated with L-15 medium. At 24 h post-infection,
the cells were used for subsequent experiments. The sequences of
AMPK- and PPARγ-specific siRNAs were as follows: SiAMPK: sense,
5′-GUUGCCUACCAUCUCAUAAUATT-3′ and antisense,
5′-UAUUAUGAGAUGGUAGGCAACTT-3′; and siPPARγ: Sense,
5′-GACAGUGACUUGGCUAUAUTT-3′ and antisense,
5′-GCGAUCUUGACAGGAAAGATT-3′.
Quantitative real-time polymerase
chain reaction (qPCR)
For verifying the results of RNA sequencing, qPCR
assay was performed. Total RNA was extracted from the cells with
TRIzol reagent. The mRNA was reverse transcribed to cDNA with the
help of HiScript II One-Step RT-PCR Kit. ChamQ SYBR qPCR Master Mix
and quantitative fluorescence analysis were used to analyze the
expression of EMT-related factors, and the analysis system was
CFX96 Touch optical system (Bio-Rad Laboratories, Inc.). The primer
sequences of E-cadherin are as follows: sense,
5′-GCCATCGCTTACACCATCCTCAG-3′ and antisense,
5′-CTCTCTCGGTCCAGCCCAGTG-3′. PCR conditions were as follows: 95°C
for 5 min (1×), followed by 95°C for 10 sec, 63°C for 30 sec (40×),
60°C for 10 sec (1×), and then 95°C for 10 sec (1×). Relative
quantification was calculated with the 2−ΔΔCq method as
described by Livak and Schmittgen (29).
Western blot (WB) analysis
Total proteins were extracted from the MDA-MB-231
and MDA-MB-453 cells using RIPA lysis buffer with phosphatase
inhibitor and PMSF for 30 min on ice. The proteins were quantified
using the BCA protein concentration determination kit, separated by
8% SDS-PAGE gel electrophoresis and transferred onto polyvinylidene
fluoride (PVDF) membranes. The membranes were blocked with 5%
non-fat milk and incubated overnight at 4°C with specific primary
antibodies (vimentin, E-cadherin, p-AMPK and AMPK) at a dilution
rate of 1:1,000 (v/v). After washing 3 times with TBST for 5 min,
the membranes were incubated with HRP-conjugated secondary
antibodies and then the membrane was visualized on a gel imager
after adding Immobilon Western Chemiluminescent HRP substrate (cat.
no. SQ201; Shanghai EpiZyme Biotechnology, Inc.). GAPDH was used as
a normalized control. Images were captured by Bio-Rad Gel-Doc XR+
system with Image Lab (version 4.1) software and processed with
Quantity One software (Bio-Rad Laboratories, Inc.).
Immunofluorescence (IF)
The suspended cells were seeded in a 96-well plate
at a density of 5×103 per well. At 80–90% confluence,
the cells were treated with compound C or siAMPK for 24 h, fixed
with 4% buffered paraformaldehyde, and permeabilized with 1% Triton
X-100. After being blocked using 5% non-fat milk solution for 30
min, the cells were incubated with primary anti-E-cadherin or
anti-vimentin monoclonal antibody at 37°C for 2 h and subsequently
incubated with secondary antibody (cat. no. AMJ-AB2005, 200 µl;
Wuhan AmyJet Scientific, Inc.) at 37°C for 1 h. Then the cells were
incubated with DAPI at 37°C for 5 min. Finally, the cells were
visualized with an Olympus fluorescence microscope (Olympus).
Statistical analysis
Data are expressed as the means ± SEM. Statistical
analyses were performed by a one-way analysis of variance (ANOVA),
followed by Tukey's test. A value of P<0.05 was accepted as a
statistically significant difference.
Results
VSP-17 treatment suppresses the
migration and invasion of TNBC cells in an AMPK-dependent
manner
To determine whether AMPK serves a key role in
mediating the anti-migratory and anti-invasive effects of VSP-17,
the expression levels of AMPK and p-AMPK in MDA-MB-231 and
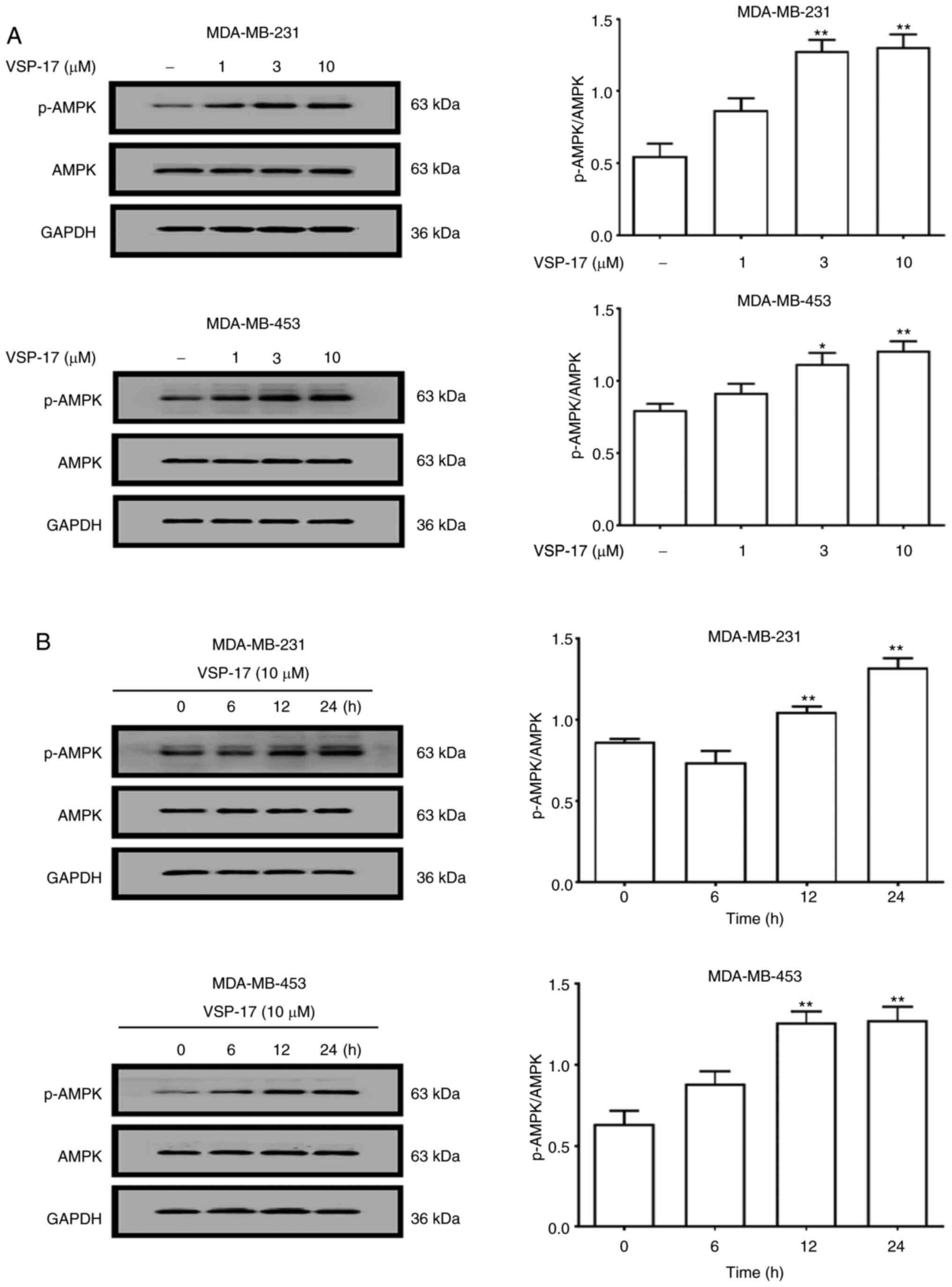
MDA-MB-453 cells were determined using western blotting. As shown
in Fig. 1A, the treatment with 1, 3
or 10 µM VSP-17 significantly upregulated the expression levels of
p-AMPK in a dose-dependent manner, while VSP-17 had no significant
effect on the expression levels of AMPK in the MDA-MB-231 and
MDA-MB-453 cells. Moreover, the expression levels of p-AMPK in both
cell lines were upregulated by 10 µM VSP-17 treatment in a
time-dependent manner (Fig.
1B).
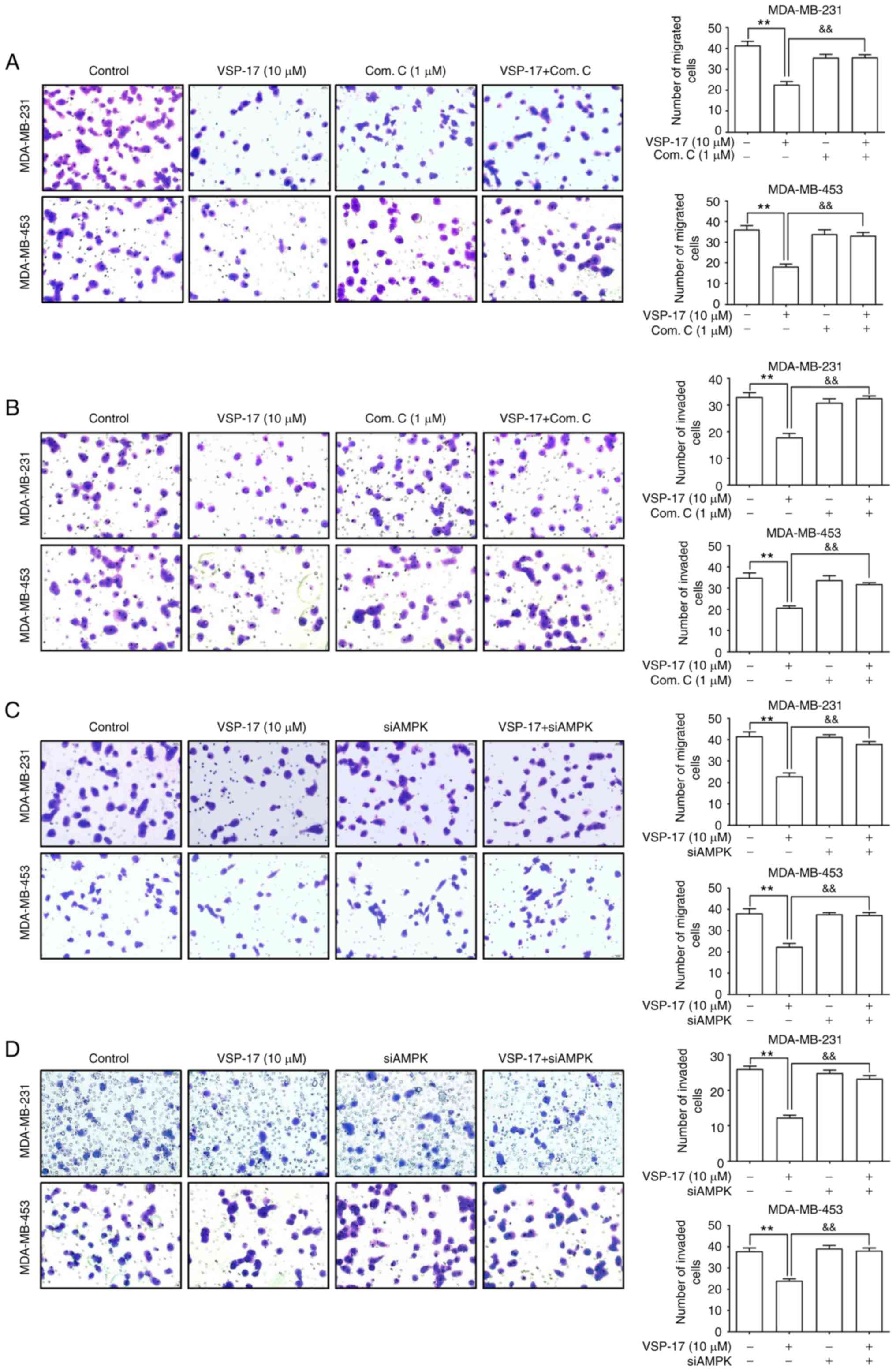
Subsequently, to further investigate the role of
AMPK in the inhibitory effect of VSP-17 on cell migration and
invasion, MDA-MB-231 and MDA-MB-453 cells were treated with
compound C or transfected with siAMPK. The data revealed that both
compound C (Fig. 2A and B) and
siAMPK (Fig. 2C and D)
significantly reversed the inhibitory effect of VSP-17 on cell
migration and invasion.
VSP-17 treatment inhibits the EMT
process in TNBC cells in an AMPK-dependent manner
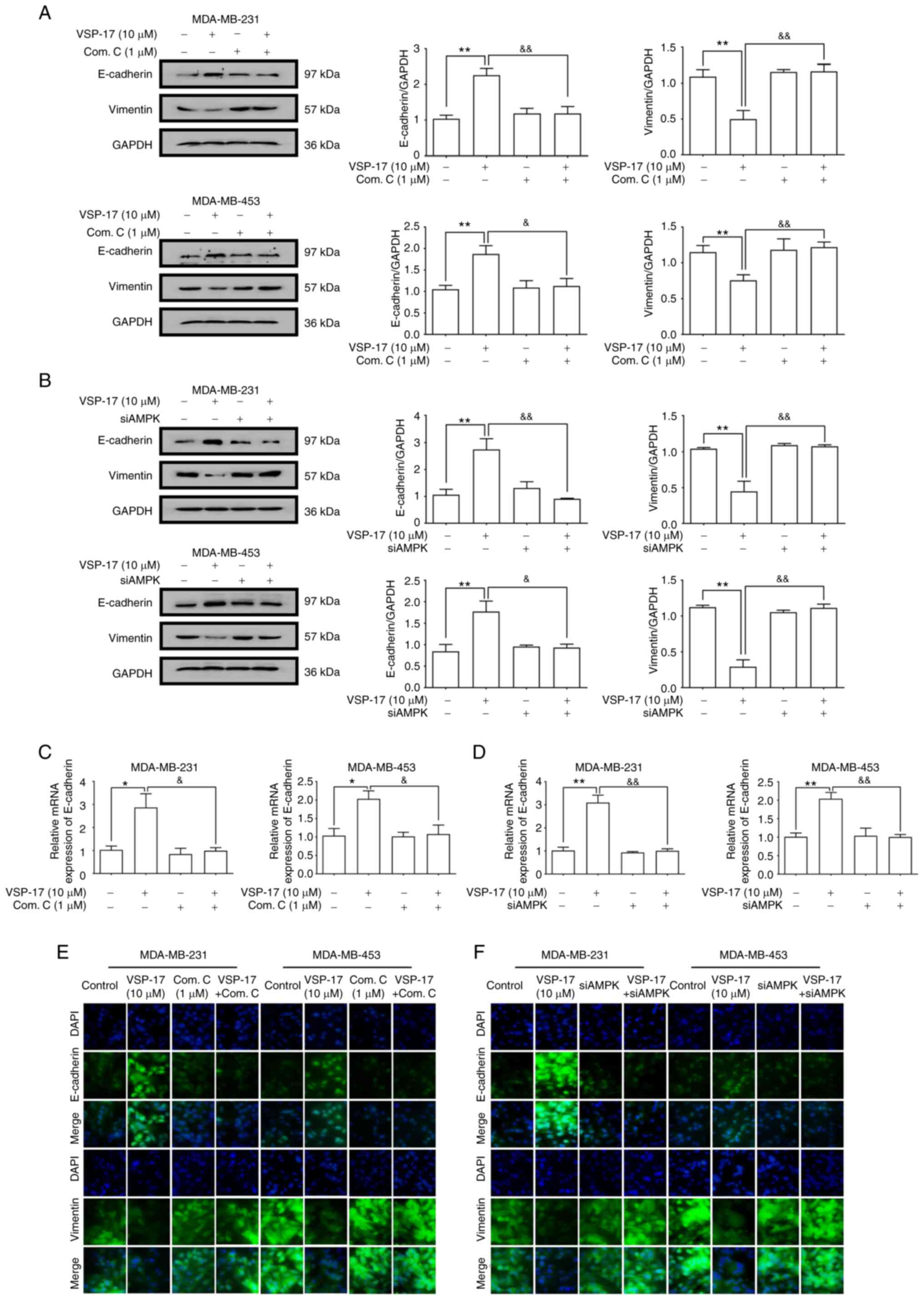
E-cadherin is a key marker of EMT (30). A previous study reported that VSP-17
treatment could upregulate the mRNA expression levels of E-cadherin
in TNBC cells (28). To further
determine the relationship between the activation of AMPK and the
inhibition of the EMT process by VSP-17, MDA-MB-231 and MDA-MB-453
cells were treated with VSP-17 (10 µM) in combination with compound
C (1 µM) or siAMPK transfection, respectively. As determined by
western blot analysis (Fig. 3A and
B), both compound C and siAMPK markedly abolished the
VSP-17-induced downregulation of vimentin expression levels and
upregulation of E-cadherin expression levels, and reverse
transcription-quantitative PCR analysis (Fig. 3C and D) and immunofluorescent
analysis (Fig. 3E and F) further
confirmed this result, suggesting that AMPK may play an important
role in the VSP-17-induced inhibition of EMT in MDA-MB-231 and
MDA-MB-453 cells.
VSP-17 treatment suppresses the
migration and invasion of TNBC cells in a PPARγ-dependent
manner
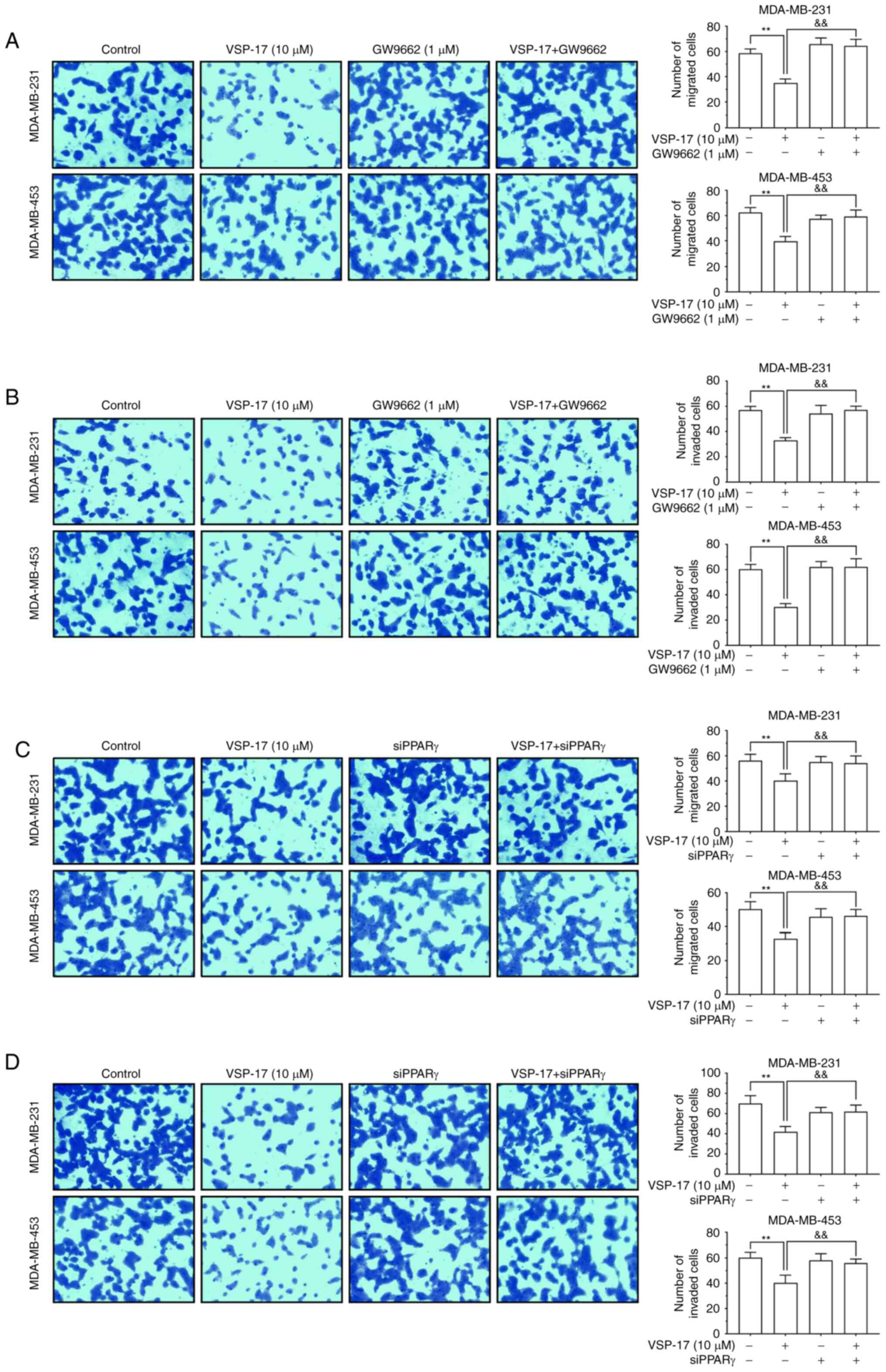
To identify the role of PPARγ in the inhibitory
effect of VSP-17 on migration and invasion, the PPARγ antagonist,
GW9662 and siPPARγ were used. The findings demonstrated that both
GW9662 and siPPARγ reversed the inhibitory effect of VSP-17 on cell
migration and invasion (Fig. 4A-D),
indicating that PPARγ may also serve an important role in the
VSP-17-induced inhibition of migration and invasion in TNBC
cells.
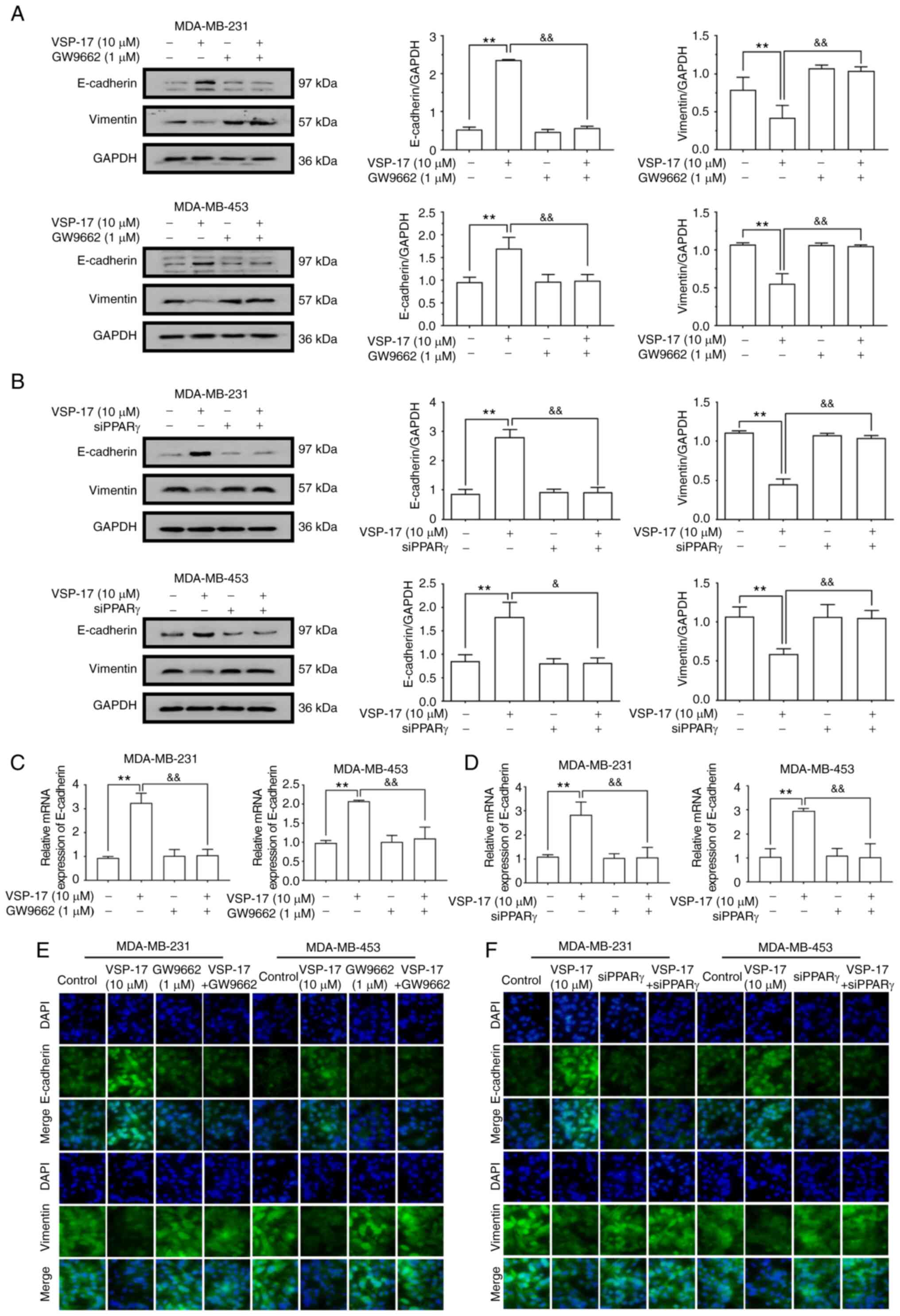
VSP-17 treatment inhibits the EMT
process in TNBC cells in a PPARγ-dependent manner
To further determine the relationship between the
activation of PPARγ and the inhibition of the EMT process by
VSP-17, MDA-MB-231 and MDA-MB-453 cells were co-treated with VSP-17
(10 µM) and GW9662 (1 µM) or treated with VSP-17 (10 µM) and
transfected with siPPARγ. As determined by western blot analysis
(Fig. 5A and B), both GW9662 and
siPPARγ significantly reversed the VSP-17-induced downregulation of
vimentin expression levels and upregulation of E-cadherin
expression levels, and reverse transcription-quantitative PCR
analysis (Fig. 5C and D) and
immunofluorescent analysis (Fig. 5E and
F) further confirmed this result, suggesting that PPARγ may
serve an important role in the VSP-17-induced inhibition of EMT in
TNBC cells.
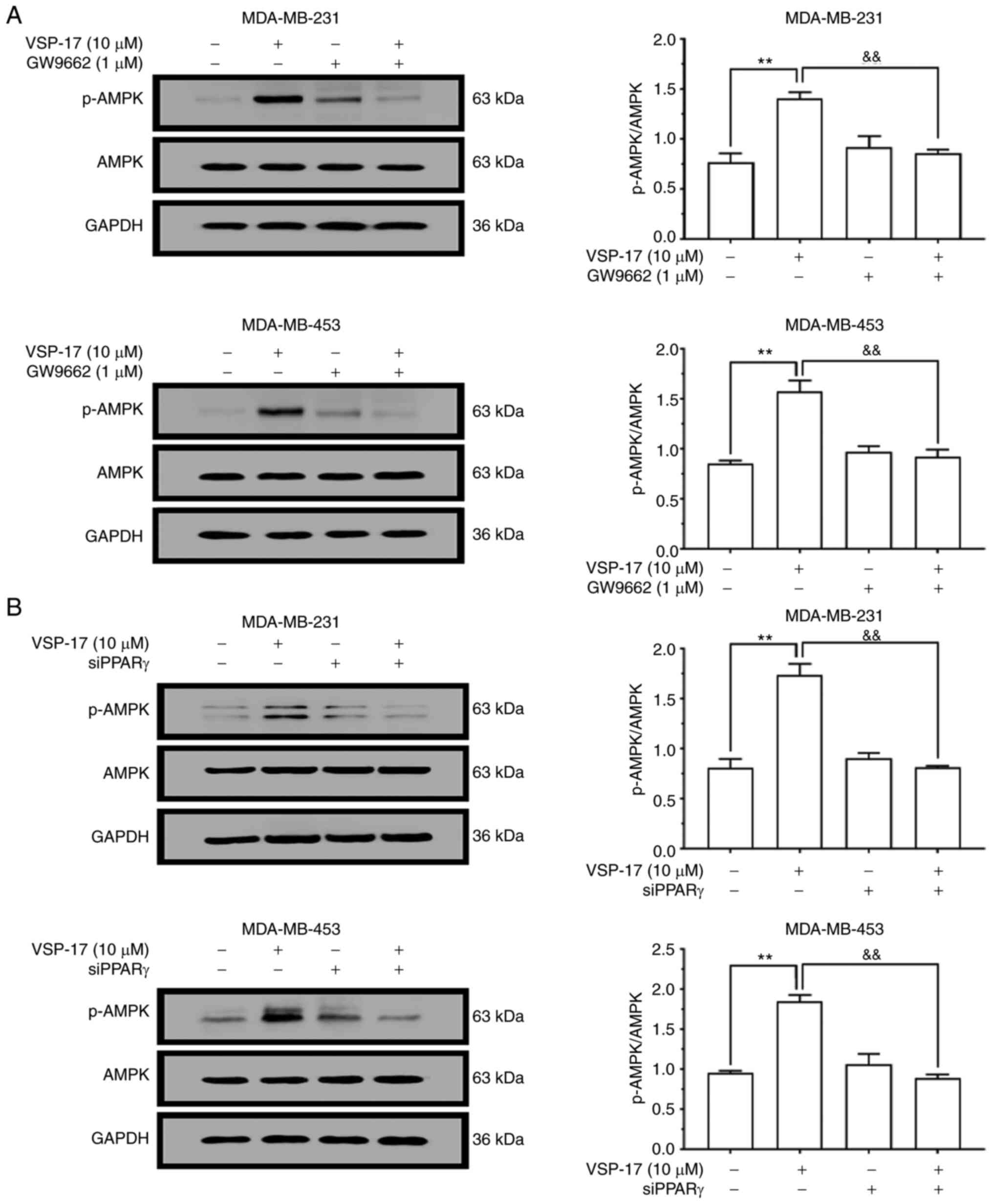
VSP-17 treatment activates AMPK in a
PPARγ-dependent manner
To determine whether the AMPK signaling pathway is
involved in the effects of VSP-17, the effect of VSP-17 treatment
on the activation of AMPK in MDA-MB-231 and MDA-MB-453 cells was
investigated. As shown in Fig. 6A and
B, the expression levels of p-AMPK in MDA-MB-231 and MDA-MB-453
cells were markedly upregulated following the treatment with 10 µM
VSP-17. In addition, both GW9662 treatment (Fig. 6A) and siPPARγ transfection (Fig. 6B) reversed the VSP-17-induced
upregulation in p-AMPK expression levels, indicating that the
VSP-17-induced activation of the AMPK signaling pathway may be
mediated by PPARγ.
Discussion
Emerging evidence has revealed that triple-negative
breast cancer (TNBC) metastasis involves the migration and invasion
of TNBC cells (28,31), and TNBC cells with increased
migratory and invasive abilities are closely associated with tumor
metastasis (32). Therefore, the
inhibition of metastasis may represent a potential strategy for
treating TNBC (33). Previous
studies have demonstrated that PPARγ agonists, such as
rosiglitazone, could inhibit the migration and invasion of breast
cancer cells (34–37). However, it is worth noting that the
long-term use of these drugs has been found to promote numerous
adverse reactions, such as hepatotoxicity, weight gain and heart
disease (38–40). Therefore, the development of a novel
PPARγ agonist with low toxicity and high selectivity may represent
a potential strategy to inhibit the metastasis of TNBC. In previous
studies, the new PPARγ agonist, VSP-17, synthesized by our team,
was found to suppress the metastasis of TNBC (28). However, the mechanism of action
remains unclear. Therefore, the present study sought to further
clarify the mechanism of action of VSP-17 with the aim of
progressing VSP-17 into clinical trials.
Previous studies have reported that
epithelial-mesenchymal transition (EMT) is a crucial initial step
for cancer invasion (41,42) and it has been proven to promote the
metastasis of tumors (43,44). The expression levels of EMT markers
are significantly altered following the occurrence of EMT in tumor
cells. In particular, the downregulation of E-cadherin expression
levels and upregulation of vimentin expression levels are the main
characteristics of EMT (45,46). A
previous study revealed that puerarin could inhibit hepatocellular
carcinoma metastasis by upregulating the expression levels of
epithelial markers and downregulating the expression levels of
mesenchymal markers (47).
Similarly, rosmarinic acid, an abundant phenolic ester found in
Prunella vulgaris, could inhibit the EMT process by
upregulating the expression levels of E-cadherin and downregulating
the expression levels of vimentin, and then inhibited the
development of colorectal cancer (24). These findings indicate that the
inhibition of the EMT process may be a potential strategy for
suppressing TNBC metastasis. In the present study, VSP-17 treatment
significantly upregulated the mRNA and protein expression levels of
E-cadherin, and downregulated the protein expression levels of
vimentin, which suggest that VSP-17 treatment may inhibit the EMT
process in TNBC cells.
Peroxisome proliferator-activated receptor γ (PPARγ)
is a ligand-activated nuclear receptor, which belongs to the
nuclear receptor gene superfamily (25). After ligand binding, PPARγ forms a
heterodimer with retinoid X receptors, which then binds to the
promoter of target genes, including NF-κB, nuclear factor erythroid
2-related factor 2 (Nrf2) and AMPK, to regulate the expression
levels (48–50). In a previous study, the PPARγ
agonist pioglitazone was found to prevent hypoxia/reoxygenation
(H/R) damage through enhancing autophagy via the AMPK/mTOR
signaling pathway (51). In
addition, ciglitazone-mediated PPARγ activation led to the
downregulation of oxidized low-density lipoprotein receptor 1
(LOX-1) expression levels and the activation of the AMPK signaling
pathway (52). These previous
studies suggest that PPARγ agonists may exert their biological
roles by activating AMPK. An increasing number of studies have also
reported that the activation of PPARγ could reverse the EMT process
by upregulating the expression levels of E-cadherin and
downregulating the expression levels of vimentin (53,54),
which suggests that the activation of PPARγ may also be considered
as a potential strategy for inhibiting the EMT process. In the
present study, GW9662 treatment or the transfection with siPPARγ
reversed the effect of VSP-17 treatment on the inhibition of
metastasis, and reversed the upregulation of E-cadherin expression
levels and downregulation of vimentin expression levels in TNBC
cells. In addition, the VSP-17-induced upregulation of p-AMPK
expression levels was blocked by GW9662 treatment or siPPARγ
transfection. These findings suggest that VSP-17 treatment may
suppress the EMT process and induce the AMPK signaling pathway in a
PPARγ-dependent manner.
Adenosine 5′-monophosphate (AMP)-activated protein
kinase (AMPK), a major mediator in cellular energy and metabolic
control, has been proven to be closely associated with the EMT
process (55,56). Experimental studies have found that
AMPK activators could inhibit the EMT transition in tumors through
suppressing the TGF-β-induced phosphorylation of SMAD2/3 (57). Increasing evidence has also revealed
that the AMPK activator, metformin, could reverse the EMT process
in breast cancer cells by activating the AMPK signaling pathway
(58). It was also previously
reported that the activation of AMPK inhibited the metastasis of
melanoma cells (59). The results
of the present study demonstrated that VSP-17 treatment could
inhibit the EMT process in an AMPK-dependent manner. It was worth
noting that multiple targets, such as mTOR, cyclooxygenase-2
(COX-2) and acetyl-CoA carboxylase (ACC), are also known to be
regulated by AMPK activation (60).
Based on these findings, it was hypothesized that VSP-17 may
regulate proteins in the downstream signaling pathway of AMPK, such
as mTOR, COX-2 and ACC, thereby leading to the regulation of the
expression levels of EMT-related markers. However, further studies
are required to identify the exact mechanism of action.
In conclusion, the findings of the present study
suggest that VSP-17 may inhibit the migration and invasion of TNBC
cells by suppressing the EMT process via the PPARγ/AMPK signaling
pathway.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Fund of China (no. 81660604), the Natural Science
Fund of Guangxi (nos. 2017GXNSFBA198104 and 2018GXNSFAA138098), the
Special Fund for Talents of Guangxi (no. AD18281016), the special
funding for 2017 Guangxi BaGui Scholars, and the special funding
for 2018 Guangxi Basic Skills of Young Teachers in the University
(nos. 2018KY0408 and 2018KY0421).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
XX and XD conceived and designed the experiments. ML
and YY performed the experiments. CW and XZ analyzed the data. HS
was involved in project management and supervised the study. YW
contributed to revising the manuscript for intellectual content and
language editing. All authors read and approved the manuscript and
agree to be accountable for all aspects of the research in ensuring
that the accuracy or integrity of any part of the work are
appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors have no financial competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ACC
|
acetyl-CoA carboxylase
|
|
AKT
|
protein kinase B
|
|
AMPK
|
adenosine 5′-monophosphate
(AMP)-activated protein kinase
|
|
COX-2
|
cyclooxygenase-2
|
|
CRC
|
colorectal cancer
|
|
DAPI
|
4,6-diamidino-2-phenylindole
|
|
EMT
|
epithelial-mesenchymal
transformation
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
FBS
|
fetal bovine serum
|
|
GAPDH
|
glycer-aldehyde-3-phosphate
dehydrogenase
|
|
HCC
|
hepatocellular carcinoma
|
|
H/R
|
hypoxia/reoxygenation
|
|
HRP
|
horseradish peroxidase
|
|
LOX-1
|
low-density lipoprotein receptor-1
|
|
mTOR
|
mechanistic target of rapamycin
|
|
NF-κB
|
nuclear factor-κB
|
|
Nrf2
|
nuclear factor erythroid 2-related
factor 2
|
|
PMSF
|
phenylmethanesulfonyl fluoride
|
|
PPARγ
|
peroxisome proliferator-activated
receptor γ
|
|
PVDF
|
polyvinylidene fluoride
|
|
RA
|
rosmarinic acid
|
|
TNBC
|
triple-negative breast cancer
|
|
TGF-β
|
transforming growth factor-β
|
References
|
1
|
da Silva JL, Cardoso Nunes NC, Izetti P,
de Mesquita GG and de Melo AC: Triple negative breast cancer: A
thorough review of biomarkers. Crit Rev Oncol Hematol.
145:1028552020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bianchini G, Balko JM, Mayer IA, Sanders
ME and Gianni L: Triple-negative breast cancer: Challenges and
opportunities of a heterogeneous disease. Nat Rev Clin Oncol.
13:674–690. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
He Z, Xu Q, Wang X, Wang J, Mu X, Cai Y,
Qian Y, Shao W and Shao Z: RPLP1 promotes tumor metastasis and is
associated with a poor prognosis in triple-negative breast cancer
patients. Cancer Cell Int. 18:1702018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Braicu C, Chiorean R, Irimie A, Chira S,
Tomuleasa C, Neagoe E, Paradiso A, Achimas-Cadariu P, Lazar V and
Berindan-Neagoe I: Novel insight into triple-negative breast
cancers, the emerging role of angiogenesis, and antiangiogenic
therapy. Expert Rev Mol Med. 18:e182016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Koedoot E, Fokkelman M, Rogkoti VM, Smid
M, van de Sandt I, de Bont H, Pont C, Klip JE, Wink S, Timmermans
MA, et al: Uncovering the signaling landscape controlling breast
cancer cell migration identifies novel metastasis driver genes. Nat
Commun. 10:29832019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang W, Xia D, Li Z, Zhou T, Chen T, Wu
Z, Zhou W, Li Z, Li L and Xu J: Aurora-A/ERK1/2/mTOR axis promotes
tumor progression in triple-negative breast cancer and
dual-targeting Aurora-A/mTOR shows synthetic lethality. Cell Death
Dis. 10:6062019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kabil N, Bayraktar R, Kahraman N, Mokhlis
HA, Calin GA, Lopez-Berestein G and Ozpolat B: Thymoquinone
inhibits cell proliferation, migration, and invasion by regulating
the elongation factor 2 kinase (eEF-2K) signaling axis in
triple-negative breast cancer. Breast Cancer Res Treat.
171:593–605. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Cao D, Zhu GY, Lu Y, Yang A, Chen D, Huang
HJ, Peng SX, Chen LW and Li YW: Luteolin suppresses
epithelial-mesenchymal transition and migration of triple-negative
breast cancer cells by inhibiting YAP/TAZ activity. Biomed
Pharmacother. 129:1104622020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yeung KT and Yang J:
Epithelial-mesenchymal transition in tumor metastasis. Mol Oncol.
11:28–39. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Savagner P: Leaving the neighborhood:
Molecular mechanisms involved during epithelial-mesenchymal
transition. Bioessays. 23:912–923. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Heimes AS and Schmidt M: Atezolizumab for
the treatment of triple-negative breast cancer. Expert Opin
Investig Drugs. 28:1–5. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu J, Song N, Huang Y and Chen Y: Irisin
inhibits pancreatic cancer cell growth via the AMPK-mTOR pathway.
Sci Rep. 8:152472018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Luo W, Liu Q, Jiang N, Li M and Shi L:
Isorhamnetin inhibited migration and invasion via suppression of
Akt/ERK-mediated epithelial-to-mesenchymal transition (EMT) in A549
human non-small-cell lung cancer cells. Biosci Rep.
39:BSR201901592019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Eroles P, Bosch A, Pérez-Fidalgo JA and
Lluch A: Molecular biology in breast cancer: Intrinsic subtypes and
signaling pathways. Cancer Treat Rev. 38:698–707. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Green AS, Chapuis N, Lacombe C, Mayeux P,
Bouscary D and Tamburini J: LKB1/AMPK/mTOR signaling pathway in
hematological malignancies: From metabolism to cancer cell biology.
Cell Cycle. 10:2115–2120. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
He K, Guo X, Liu Y, Li J, Hu Y, Wang D and
Song J: TUFM downregulation induces epithelial-mesenchymal
transition and invasion in lung cancer cells via a mechanism
involving AMPK-GSK3β signaling. Cell Mol Life Sci. 73:2105–2121.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shaw RJ, Kosmatka M, Bardeesy N, Hurley
RL, Witters LA, DePinho RA and Cantley LC: The tumor suppressor
LKB1 kinase directly activates AMP-activated kinase and regulates
apoptosis in response to energy stress. Proc Natl Acad Sci USA.
101:3329–3335. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Khan S, Shukla S, Sinha S and Meeran SM:
Role of adipokines and cytokines in obesity-associated breast
cancer: Therapeutic targets. Cytokine Growth Factor Rev.
24:503–513. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Si Y, Wang J, Liu X, Zhou T, Xiang Y,
Zhang T, Wang X, Feng T, Xu L, Yu Q, et al: Ethoxysanguinarine, a
novel direct activator of AMP-activated protein kinase, induces
autophagy and exhibits therapeutic potential in breast cancer
cells. Front Pharmacol. 10:15032020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Baba Y, Nosho K, Shima K, Meyerhardt JA,
Chan AT, Engelman JA, Cantley LC, Loda M, Giovannucci E, Fuchs CS
and Ogino S: Prognostic significance of AMP-activated protein
kinase expression and modifying effect of MAPK3/1 in colorectal
cancer. Br J Cancer. 103:1025–1033. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cazarin JM, Coelho RG, Hecht F, Andrade BM
and Carvalho DP: 5′-AMP-activated protein kinase regulates
papillary (TPC-1 and BCPAP) thyroid cancer cell survival,
migration, invasion, and epithelial-to-mesenchymal transition.
Thyroid. 26:933–942. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qu C, Zhang W, Zheng G, Zhang Z, Yin J and
He Z: Metformin reverses multidrug resistance and
epithelial-mesenchymal transition (EMT) via activating
AMP-activated protein kinase (AMPK) in human breast cancer cells.
Mol Cell Biochem. 386:63–71. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Han YH, Kee JY and Hong SH: Rosmarinic
acid activates AMPK to inhibit metastasis of colorectal cancer.
Front Pharmacol. 9:682018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lamers C, Schubert-Zsilavecz M and Merk D:
Therapeutic modulators of peroxisome proliferator-activated
receptors (PPAR): A patent review (2008-present). Expert Opin Ther
Pat. 22:803–841. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Krishnan A, Nair SA and Pillai MR: Biology
of PPAR gamma in cancer: A critical review on existing lacunae.
Curr Mol Med. 7:532–540. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Nunez-Anita RE, Cajero-Juarez M and Aceves
C: Peroxisome proliferator-activated receptors: Role of isoform
gamma in the antineoplastic effect of iodine in mammary cancer.
Curr Cancer Drug Targets. 11:775–786. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang Y, Zhu M, Yuan B, Zhang K, Zhong M,
Yi W, Xu X and Duan X: VSP-17, a New PPARγ agonist, suppresses the
metastasis of triple-negative breast cancer via upregulating the
expression of E-Cadherin. Molecules. 23:1212018. View Article : Google Scholar
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Jolly MK, Ware KE, Xu S, Gilja S, Shetler
S, Yang Y, Wang X, Austin RG, Runyambo D, Hish AJ, et al:
E-Cadherin represses anchorage-independent growth in sarcomas
through both signaling and mechanical mechanisms. Mol Cancer Res.
17:1391–1402. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kang DY, Sp N, Kim DH, Joung YH, Lee HG,
Park YM and Yang YM: Salidroside inhibits migration, invasion and
angiogenesis of MDA-MB 231 TNBC cells by regulating EGFR/Jak2/STAT3
signaling via MMP2. Int J Oncol. 53:877–885. 2018.PubMed/NCBI
|
|
32
|
Chien YC, Liu LC, Ye HY, Wu JY and Yu YL:
EZH2 promotes migration and invasion of triple-negative breast
cancer cells via regulating TIMP2-MMP-2/-9 pathway. Am J Cancer
Res. 8:422–434. 2018.PubMed/NCBI
|
|
33
|
Zardavas D, Baselga J and Piccart M:
Emerging targeted agents in metastatic breast cancer. Nat Rev Clin
Oncol. 10:191–210. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bojková B, Garajová M, Kajo K, Péc M,
Kubatka P, Kassayová M, Kisková T, Orendás P, Ahlersová E and
Ahlers I: Pioglitazone in chemically induced mammary carcinogenesis
in rats. Eur J Cancer Prev. 19:379–384. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Colmers IN, Bowker SL and Johnson JA:
Thiazolidinedione use and cancer incidence in type 2 diabetes: A
systematic review and meta-analysis. Diabetes Metab. 38:475–484.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fogo AB: Potential for peroxisome
proliferator-activated receptor-gamma agonists in progression:
Beyond metabolism. Curr Opin Nephrol Hypertens. 17:282–285. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu YY, Liu H, Su L, Xu N, Xu DH, Liu HY,
Spaner D, Bed-David Y and Li YJ: PPARγ inhibits breast cancer
progression by upregulating PTPRF expression. Eur Rev Med Pharmacol
Sci. 23:9965–9977. 2019.PubMed/NCBI
|
|
38
|
Burstein HJ, Demetri GD, Mueller E, Sarraf
P, Spiegelman BM and Winer EP: Use of the peroxisome
proliferator-activated receptor (PPAR) gamma ligand troglitazone as
treatment for refractory breast cancer: A phase II study. Breast
Cancer Res Treat. 79:391–397. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Spence JD, Viscoli CM, Inzucchi SE,
Dearborn-Tomazos J, Ford GA, Gorman M, Furie KL, Lovejoy AM, Young
LH and Kernan WN; IRIS Investigators, : Pioglitazone therapy in
patients with stroke and prediabetes: A post Hoc analysis of the
IRIS randomized clinical trial. JAMA Neurol. 76:526–535. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Scheen AJ: Thiazolidinediones and liver
toxicity. Diabetes Metab. 27:305–313. 2001.PubMed/NCBI
|
|
41
|
Barriga EH and Mayor R: Adjustable
viscoelasticity allows for efficient collective cell migration.
Semin Cell Dev Biol. 93:55–68. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Huang X, Zeng Y, Xing X, Zeng J, Gao Y,
Cai Z, Xu B, Liu X, Huang A and Liu J: Quantitative proteomics
analysis of early recurrence/metastasis of huge hepatocellular
carcinoma following radical resection. Proteome Sci. 12:222014.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Martin FT, Dwyer RM, Kelly J, Khan S,
Murphy JM, Curran C, Miller N, Hennessy E, Dockery P, Barry FP, et
al: Potential role of mesenchymal stem cells (MSCs) in the breast
tumour microenvironment: Stimulation of epithelial to mesenchymal
transition (EMT). Breast Cancer Res Treat. 124:317–326. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Dongre A and Weinberg RA: New insights
into the mechanisms of epithelial-mesenchymal transition and
implications for cancer. Nat Rev Mol Cell Biol. 20:69–84. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Peinado H, Olmeda D and Cano A: Snail, Zeb
and bHLH factors in tumour progression: An alliance against the
epithelial phenotype? Nat Rev Cancer. 7:415–428. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhou Y, Xue R, Wang J and Ren H: Puerarin
inhibits hepatocellular carcinoma invasion and metastasis through
miR-21-mediated PTEN/AKT signaling to suppress the
epithelial-mesenchymal transition. Braz J Med Biol Res.
53:e88822020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Juge-Aubry CE, Gorla-Bajszczak A, Pernin
A, Lemberger T, Wahli W, Burger AG and Meier CA: Peroxisome
proliferator-activated receptor mediates cross-talk with thyroid
hormone receptor by competition for retinoid X receptor. Possible
role of a leucine zipper-like heptad repeat. J Biol Chem.
270:18117–18122. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Xia H, Ge Y, Wang F, Ming Y, Wu Z, Wang J,
Sun S, Huang S, Chen M, Xiao W and Yao S: Protectin DX ameliorates
inflammation in sepsis-induced acute lung injury through mediating
PPARγ/NF-κB pathway. Immunol Res. 68:280–288. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Rani N and Arya DS: Chrysin rescues rat
myocardium from ischemia-reperfusion injury via PPAR-γ/Nrf2
activation. Eur J Pharmacol. 883:1733892020. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Xi X, Zou C, Ye Z, Huang Y, Chen T and Hu
H: Pioglitazone protects tubular cells against
hypoxia/reoxygenation injury through enhancing autophagy via
AMPK-mTOR signaling pathway. Eur J Pharmacol. 863:1726952019.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Xu L, Wang S, Li B, Sun A, Zou Y and Ge J:
A protective role of ciglitazone in ox-LDL-induced rat
microvascular endothelial cells via modulating PPARγ-dependent
AMPK/eNOS pathway. J Cell Mol Med. 19:92–102. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kao HF, Chang-Chien PW, Chang WT, Yeh TM
and Wang JY: Propolis inhibits TGF-β1-induced
epithelial-mesenchymal transition in human alveolar epithelial
cells via PPARγ activation. Int Immunopharmacol. 15:565–574. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Reka AK, Kurapati H, Narala VR, Bommer G,
Chen J, Standiford TJ and Keshamouni VG: Peroxisome
proliferator-activated receptor-gamma activation inhibits tumor
metastasis by antagonizing Smad3-mediated epithelial-mesenchymal
transition. Mol Cancer Ther. 9:3221–3232. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen YC, Li H and Wang J: Mechanisms of
metformin inhibiting cancer invasion and migration. Am J Transl
Res. 12:4885–4901. 2020.PubMed/NCBI
|
|
56
|
Zhang ZG, Zhang HS, Sun HL, Liu HY, Liu MY
and Zhou Z: KDM5B promotes breast cancer cell proliferation and
migration via AMPK-mediated lipid metabolism reprogramming. Exp
Cell Res. 379:182–190. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lin H, Li N, He H, Ying Y, Sunkara S, Luo
L, Lv N, Huang D and Luo Z: AMPK inhibits the stimulatory effects
of TGF-β on Smad2/3 activity, cell migration, and
epithelial-to-mesenchymal transition. Mol Pharmacol. 88:1062–1071.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Banerjee P, Surendran H, Chowdhury DR,
Prabhakar K and Pal R: Metformin mediated reversal of epithelial to
mesenchymal transition is triggered by epigenetic changes in
E-cadherin promoter. J Mol Med (Berl). 94:1397–1409. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Cerezo M, Tichet M, Abbe P, Ohanna M,
Lehraiki A, Rouaud F, Allegra M, Giacchero D, Bahadoran P,
Bertolotto C, et al: Metformin blocks melanoma invasion and
metastasis development in AMPK/p53-dependent manner. Mol Cancer
Ther. 12:1605–1615. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Li W, Saud SM, Young MR, Chen G and Hua B:
Targeting AMPK for cancer prevention and treatment. Oncotarget.
6:7365–7378. 2015. View Article : Google Scholar : PubMed/NCBI
|