Introduction
Salivary adenoid cystic carcinoma (SACC) is one of
the most common salivary gland malignancies and is associated with
a high local recurrence rate, neurotropic invasion and hematogenous
metastasis (1). The standard
treatment for SACC involves surgery with or without radiotherapy
and chemotherapy, although the recurrence rate is still very high
and the long-term survival rate is poor, with 5-year survival rates
of 75–80% and 15-year survival rates of 10–30% (2). Thus, there is an urgent need to
clarify the molecular mechanisms underlying SACC invasion and
metastasis, which may help guide the development of effective
therapies.
Collective cell migration is an important process in
both normal epithelial development and cancer invasion (3). Previous studies have indicated that
the classic mode of single-cell invasion accompanies the
epithelial-mesenchymal transition (EMT) in epithelial carcinomas
(4–6). Collective invasion is another
important process that involves tumour cells maintaining their
cell-cell contacts and moving as inter-connected multicellular
clumps with or without partial EMT (3,7,8).
Therefore, it is important to clarify which features of tumour
progression regulate the transition from single-cell invasion to a
collective invasive phenotype, and how these features might be
related.
Collective invasion is generally speculated to be
driven by leader cells, which maintain different cell polarities,
extend plate-like and filamentous pseudopods, adhere to the
extracellular matrix, promote actin-mediated cell contraction, and
secrete a variety of proteolytic enzymes to degrade the
extracellular matrix and guide follower cells (9). Researchers have also demonstrated that
breast cancer leader cells express basal epithelial genes [keratin
14 (K14) and p63] and that silencing these genes could inhibit
collective invasion (10). Han
et al (11) also reported
that netrin-1 expression was increased in leader cells at the
invasion front, which promoted collective migration of hepatoma
cells through increased N-cadherin expression in a
three-dimensional (3D) culture model. Our previous research also
revealed that SACC cells formed cellular masses to invade the
surrounding tissues, such as nerves, blood vessels and glands, and
that this form of collective invasion was the primary method of
SACC invasion (12). However, it is
difficult to clarify the relative contributions of specific genetic
and microenvironmental changes to carcinoma cell collective
invasion, especially as the molecular characteristics of SACC
leader and follower cells are not completely understood.
δ-like ligand 4 (Dll4) is the ligand of Notch1 and
is mainly expressed in peritumoral vascular endothelial cells
(13). Previous studies have
indicated that Dll4 upregulation in various cancers can promote
invasion and is closely related to a malignant phenotype and poor
prognosis in cases of gastric cancer and cervical cancer (14–18).
Xu et al (19) also reported
that blocking Dll4 could inhibit tumour cell proliferation and
mammosphere formation in breast cancer. However, there are no data
regarding the potential role of the Dll4/Notch1 signalling pathway
in the collective invasion of SACC. Therefore, the present study
used immunohistochemical staining to evaluate Dll4 and Notch1
expression at the invasion front of SACC. Furthermore, a 3D culture
model was used to examine the expression of Dll4 and Notch1 in the
leader and follower cells, and it was evaluated whether the
expression of these genes changed in differently simulated tumour
microenvironments. The results may help guide further research
regarding the molecular markers of collective invasion and how the
tumour microenvironment influences the collective invasion of
carcinoma cells.
Materials and methods
Histological analysis
This retrospective study evaluated 84 SACC specimens
and 5 normal salivary gland specimens (collected between January
2009 and December 2014) from the Department of Oral Pathology
database (West China Hospital of Stomatology, Sichuan University,
Chengdu, China). The patients had provided informed consent for the
use of their surgical specimens and clinical data in research. The
study protocol was approved by the Institutional Ethics Committee
of the West China Medical Center, Sichuan University (approval no.
WCHSIRB-D-2017-208). All specimens had been fixed with 10% buffered
formalin at 4°C for 24 h, and then embedded in paraffin.
Cell culture and transfection
Commonly used cell lines (SACC-83, SACC-LM and
HUVECs) were obtained from the State Key Laboratory of Oral
Diseases (Sichuan University, Chengdu, China). All cells were
cultured in high-glucose Dulbecco's modified Eagle's medium (DMEM;
Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% foetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin (HyClone; Cytiva) and 100 µg/ml streptomycin (HyClone;
Cytiva) and maintained in an incubator at 37°C with 5%
CO2. The experimental conditions for hypoxia were
conducted by incubating cells at 1% O2 in a hypoxia
workstation (Ruskinn Invivo 400 Hypoxic Workstation). Three small
interfering (si)RNAs against Dll4 and a negative control siRNA were
designed by Shanghai GeneChem Co., Ltd. SACC-83 and SACC-LM cells
were seeded at a density of 2×105 cell/ml and were
transfected with the siRNA (20 pM) using EndoFectin™ reagent
(GeneCopoeia, Inc.), according to the manufacturer's instructions.
The sequences of the siRNA were as follows: Duplex-1,
GCGUCUGCCUUAAGCACUUTTAAGUGCUUAAGGCAGACGCTT; duplex-2,
CCAGAAGGACAACCUGAUUTTAAUCAGGUUGUCCUUCUGGTT; duplex-3,
GCAACUGCCCUUAUGGCUUTTAAGCCAUAAGGGCAGUUGCTT; and negative control,
UUCUCCGAACGUGUCACGUTTACGUGACACGUUCGGAGAATT. The transfected cells
were then used for reverse transcription-quantitative polymerase
chain reaction (RT-qPCR), migration and invasion assays, 3D
cultures, and immunofluorescence (IFC) staining 24 h after
transfection.
Haematoxylin and eosin (H&E) and
immunohistochemical (IHC) staining
The sections of SACC specimens and normal glands
were deparaffinized using xylene, and then rehydrated using a
series of ethanol and water solutions. For H&E staining, slices
were stained with hematoxylin solution for 5 min, dipped in acid
ethanol, and then washed with distilled water in turn, all at room
temperature. The slices were subsequently stained with eosin
solution for 3 min and re-immersed in alcohol and xylene at room
temperature. For IHC staining, the sections were autoclaved for
antigen retrieval, blocked with 3% hydrogen peroxide for 2 min and
10% goat serum albumin (OriGene Technologies, Inc.) for 30 min at
room temperature, and then subjected to routine IHC staining
procedures as previously described (16,17).
The primary antibodies were mouse anti-Dll4 (1:500; cat. no.
220726; ZenBio, Inc.) and rabbit anti-Notch1 (1:300; cat. no.
516673; ZenBio, Inc.). Then, sections were incubated with a
commercial goat anti-mouse/rabbit IgG secondary antibody (cat. no.
SA1020; Wuhan Boster Biological Technology, Ltd.) for 15 min, and
detected with a DAB Staining Kit (Wuhan Boster Biological
Technology, Ltd.) at room temperature. The IHC staining was
independently scored by two researchers using the average
percentage of stained cells: i) 0 points, <5% of cells (no
staining); ii) 1 point, 5–25% of cells (mild staining); iii) 2
points, 25–50% of cells (moderate staining); and iv) or 3 points,
>50% of cells (strong staining).
RT-qPCR
Total RNA was extracted using a RNeasy Mini kit
(Qiagen China Co., Ltd.) and reverse transcribed into cDNA using a
High-Capacity cDNA Reverse Transcription kit (Applied Biosystems;
Thermo Fisher Scientific, Inc.), according to the manufacturer's
instructions. The primer sequences used were as follows: Dll4
forward, 5′-AGCTGTAAGGACCAGGAG-3′ and reverse,
5′-ACATTCACAAGCATAGTTGG-3′; Notch1 forward,
5′-GAGGCGTGGCAGACTATGC-3′ and reverse, 5′-CTTGTACTCCGTCAGCGTGA-3′;
transcription factor HES-1 (Hes1) forward,
5′-ACGTGCGAGGGCGTTAATAC-3′ and reverse, 5′-GGGGTAGGTCATGGCATTGA-3′;
hairy/enhancer-of-split related with YRPW motif protein 1 (Hey1)
forward, 5′-GAAGTTGCGCGTTATCTGAGC-3′ and reverse,
5′-ATGCGAAACCAGTCGAACTCG-3′; MMP9 forward,
5′-GGGACGCAGACATCGTCATC-3′ and reverse, 5′-TCGTCATCGTCGAAATGGGC-3′;
C-X-C chemokine receptor type 4 (CXCR4) forward,
5′-GGGCAATGGATTGGTCATCCT-3′ and reverse, 5′-TGCAGCCTGTACTTGTCCG-3′;
Twist-related protein 1 (Twist1) forward,
5′-GCCTAGAGTTGCCGACTTATG-3′ and reverse,
5′-TGCGTTTCCTGTTAAGGTAGC-3′; vascular endothelial growth factor A
(VEGFA) forward, 5′-GGACCCTGGCTTTACTGCTGTACC-3′ and reverse,
5′-TCACCGCCTTGGCTTGTCACA-3′; hypoxia-inducible factor 1-α (HIF-1α)
forward, 5′-TGCTCATCAGTTGCCACTTCC-3′ and reverse,
5′-CGCTGTGTGTTTTGTTCTTTACCC-3′; GADPH forward,
5′-ACAACTTTGGTATCGTGGAAGG-3′ and reverse,
5′-GCCATCACGCCACAGTTTC-3′. The RT-qPCR reactions were performed
using SYBR-Green (Roche Diagnostics) at the following thermocycling
conditions: Preincubation at 95°C for 30 sec, followed by 45 cycles
of 5 sec at 95°C, 10 sec at 55°C, and 25 sec at 72°C. The sample
values were normalized to the housekeeping gene (GAPDH) and
calculated using the 2−ΔΔCq method (20).
Migration and invasion assays
The cells were seeded in 6-well plates at a density
of 1.5×105 cell/well (at 50% confluence), and then
transfected with Dll4 siRNA or control siRNA. After 24 h, the cells
were cultured in serum-free medium, and wounds were created by
scraping each plate with a 200-µl pipette tip. Images of identical
scratch regions were captured at various time points using a phase
contrast microscope (Olympus IX 71; Olympus Corporation). The wound
healing area of five random scopes was quantified using ImageJ
software (version 1.52a; National Institutes of Health). Invasion
assays were performed using Transwell chambers (8-µm pore size;
Costar; Corning Inc.) that were coated with 100 µl Matrigel for 30
min at 37°C. A total of 5–6×104 cells were seeded into
the upper compartment with 200 ml serum-free medium and the
chambers were then placed in 24-well dishes containing 700 µl DMEM
with 10% FBS in the lower chamber. After 24 h or 48 h of co-culture
in an incubator at 37°C, the membranes were fixed with 4%
formaldehyde for 20 min at room temperature, and stained with 10%
Giemsa staining solution for 30 min at room temperature. After
removing the cells from the membrane's upper surface, the cells on
the lower membrane surface of five random scopes (magnification,
×10) were counted under a microscope (Olympus IX 71; Olympus
Corporation).
Tube formation assay
Conditioned medium (CM) was harvested 48 h after
SACC cells were transfected with Dll4 siRNA. Then, the 48-well
plates were coated with Matrigel (150 µl per well) and HUVECs were
added (4×104 cells/well). After incubation at 37°C for 6
h, tube formation was observed under a microscope (Olympus IX 71;
Olympus Corporation), and the number of nodes as well as meshes of
five random scopes were quantified (magnification, ×10) by ImageJ
software (version 1.52a; National Institutes of Health).
Cell co-culture
The SACC-83 or SACC-LM cells were seeded in 6-well
plates at a density of 1.5×105 cells/well and
transfected with Dll4 siRNA or Control siRNA. HUVECs were seeded
into the upper chamber of the Transwell (0.4 µm; EMD Millipore)
co-culture system at the same density of SACC cells. Then, the
upper and lower compartment were combined for 24–48 h, followed by
subsequent assays.
3D culture
Multicellular spheroids were generated using the
hanging drop technique or by plating the cells on 96-well ultra-low
attachment plates (Corning, Inc.) or soft agar-coated 96-well
plates. For the hanging drop method, the cells were re-suspended in
DMEM supplemented with 10% methylcellulose at a density of
5×104 cells/ml, and incubated overnight in 25 µl
droplets as previously described (21). For the other methods, 1% agar was
used to coat 96-well plates, the cells were re-suspended in 200 µl
DMEM at a density of 5×104 cells/ml, and then the cells
were seeded into 96-well ultra-low attachment plates or soft
agar-coated plates. The cells were cultured for 48–72 h to ensure
multicellular aggregation, and then the spheroids were incorporated
into a rat tail collagen solution (cat. no. C8062; Beijing Solarbio
Science & Technology Co., Ltd.) before collagen polymerization
occurred. The DMEM was then added and the plates were incubated at
37°C for 24–48 h. Leader and follower cells were defined based on
their positions in the cell cluster (at the front of the cluster
and at the back of the cluster) (3). Leader cells appeared as protrusive
cells at the front of invasive strands. After 4 days, the plates
were scanned and the relative diameters of the well and the gel
were measured using ImageJ software (version 1.52a; National
Institutes of Health).
Statistical analysis
All statistical analyses were performed using SPSS
software (version 22.0; IBM Corp.) and GraphPad Prism software
(version 5.03; GraphPad Software, Inc.). Continuous data were
reported as the mean ± standard deviation and analysed using
unpaired Student's t-test (comparisons between 2 groups) or using
one-way analysis of variance with Dunnett's post hoc multiple
comparisons (comparisons between ≥3 groups). The correlation
between the expression of Dll4 and Notch1 was assessed with a
Spearman's correlation coefficient. A Chi-squared test was used to
evaluate whether expression of Dll4 or Notch1 was associated with
the patients' clinicopathological characteristics. P<0.05 was
considered to indicate a statistically significant difference.
Results
Increased expression of Dll4 at the
invasion front is associated with metastasis and recurrence of
SACC
The significance of Dll4 and Notch1 expression in
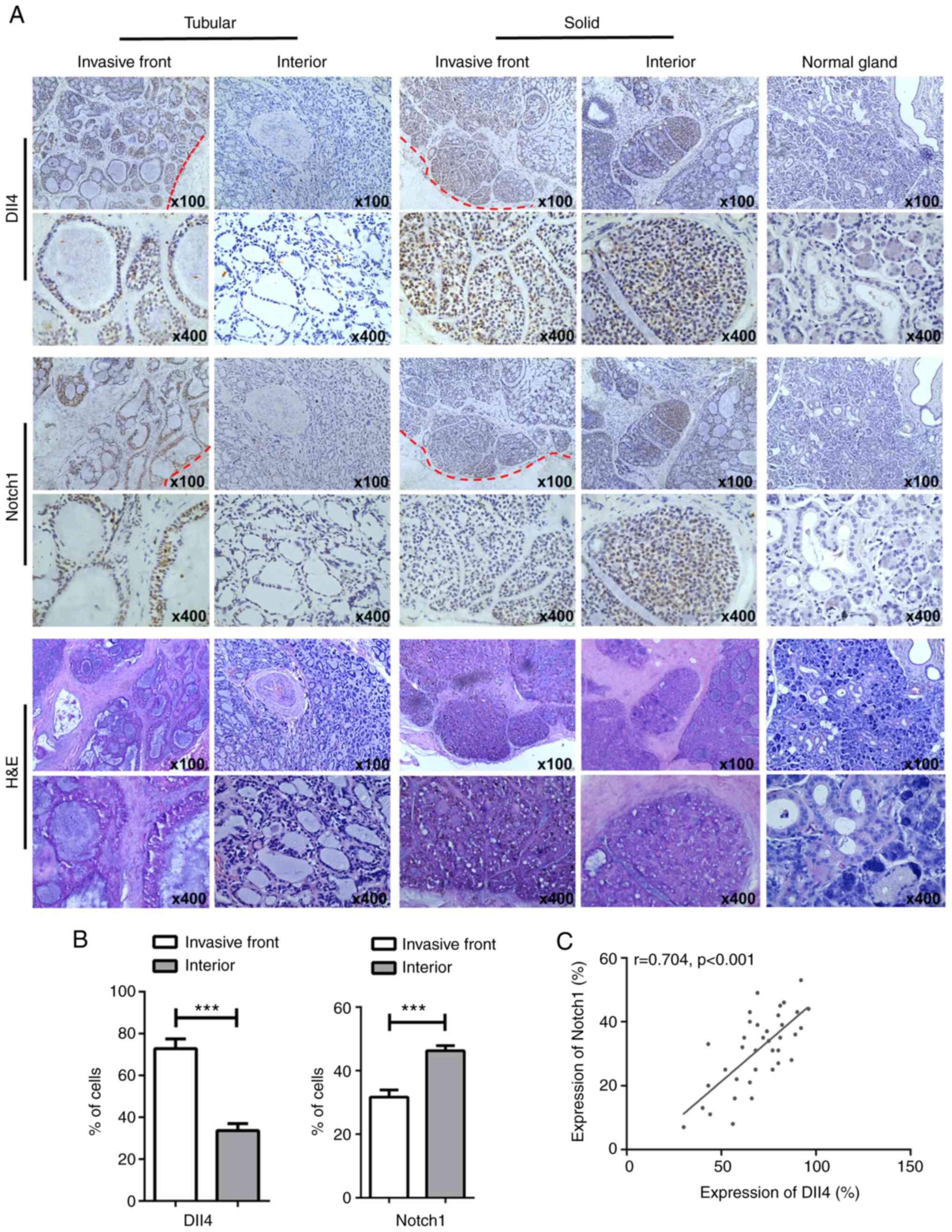
human SACC tissues was evaluated using IHC. The results revealed
that Dll4 staining was predominantly observed in the cytoplasm and
membrane and Notch1 staining was observed in the cytoplasm,
membrane and nucleus (Fig. 1A).
Furthermore, Dll4-positive staining was observed in 46 of the 84
patients with SACC (54.76%) and Notch1-positive staining was
observed in 37 patients (44.04%). Of note, Dll4 expression was much
higher in tumour cells at the invasive front (vs. at the centre of
the SACC mass; P=0.001; Fig. 1B)
and there was a close correlation between Dll4 and Notch1
expression at the invasive front (r=0.74, P<0.001; Fig. 1C).
Tables I and
II show the relationships between
Dll4/Notch1 expression and the patients' clinicopathological
characteristics. High expression of Dll4 and Notch1 at the invasive
front was significantly related to solid tumour type, high TNM
grade, and high rates of metastasis and recurrence (all P<0.05),
but not with age (P=0.794 for Dll4 and P=0.922 for Notch1), sex
(P=0.752 for Dll4 and P=0.703 for Notch1) or tumour site (P=0.875
for Dll4 and P=0.603 for Notch1). Thus, Dll4 and Notch1 expression
appears to play important roles at the invasive front of SACC.
 | Table I.Association between Dll4 expression
and clinicopathological characteristics of patients with SACC. |
Table I.
Association between Dll4 expression
and clinicopathological characteristics of patients with SACC.
|
|
| Dll4 expression, n
(%) |
|
|---|
|
|
|
|
|
|---|
| Parameter | N (%) | Low | High | P-value |
|---|
| Sex |
|
|
| 0.752 |
|
Male | 36 (42.86) | 17 (44.70) | 19 (41.30) |
|
|
Female | 48 (57.14) | 21 (55.30) | 27 (58.70) |
|
| Age, years |
|
|
| 0.794 |
|
≥60 | 54 (65.79) | 25 (65.79) | 29 (63.04) |
|
|
<60 | 30 (34.21) | 13 (34.21) | 17 (36.96) |
|
| Tumour
location |
|
|
| 0.875 |
| Major
salivary | 39 (46.43) | 18 (47.37) | 21 (45.65) |
|
| Minor
salivary | 45 (53.57) | 20 (52.63) | 25 (54.35) |
|
| Pathological
type |
|
|
| 0.030a |
|
Cribriform/tubular | 61 (72.62) | 32 (84.21) | 29 (63.04) |
|
|
Solid | 23 (27.38) | 6 (15.79) | 17 (36.96) |
|
| TNM stage |
|
|
| 0.007a |
|
I–II | 27 (32.14) | 18 (47.37) | 9 (19.57) |
|
|
III–IV | 57 (67.86) | 20 (52.63) | 37 (80.43) |
|
| Distant
metastasis |
|
|
| 0.027a |
|
Yes | 18 (21.43) | 4 (10.53) | 14 (30.43) |
|
| No | 66 (78.57) | 34 (89.47) | 32 (69.57) |
|
| Recurrence |
|
|
| 0.030a |
|
Yes | 15 (17.86) | 3 (7.89) | 12 (26.09) |
|
| No | 69 (82.14) | 35 (92.11) | 34 (73.91) |
|
| Table II.Association between Notch1 expression
and clinicopathological characteristics of patients with SACC. |
Table II.
Association between Notch1 expression
and clinicopathological characteristics of patients with SACC.
|
|
| Notch1 expression,
n (%) |
|
|---|
|
|
|
|
|
|---|
| Parameter | N (%) | Low | High | P-value |
|---|
| Sex |
|
|
| 0.703 |
|
Male | 36 (42.86) | 21 (44.68) | 15 (40.54) |
|
|
Female | 48 (57.14) | 26 (55.32) | 22 (59.46) |
|
| Age, years |
|
|
| 0.922 |
|
≥60 | 54 (65.79) | 30 (63.83) | 24 (64.86) |
|
|
<60 | 30 (34.21) | 17 (36.17) | 13 (35.14) |
|
| Tumour
location |
|
|
| 0.603 |
| Major
salivary | 39 (46.43) | 23 (48.93) | 16 (43.24) |
|
| Minor
salivary | 45 (53.57) | 24 (51.07) | 21 (54.35) |
|
| Pathological
type |
|
|
| 0.016a |
|
Cribriform/tubular | 61 (72.62) | 39 (82.98) | 22 (59.46) |
|
|
Solid | 23 (27.38) | 8 (17.02) | 15 (40.54) |
|
| TNM stage |
|
|
| 0.021a |
|
I–II | 27 (32.14) | 20 (42.55) | 7 (18.92) |
|
|
III–IV | 57 (67.86) | 27 (57.45) | 30 (81.08) |
|
| Distant
metastasis |
|
|
| 0.029a |
|
Yes | 18 (21.43) | 6 (12.77) | 12 (32.43) |
|
| No | 66 (78.57) | 41 (87.23) | 25 (68.57) |
|
| Recurrence |
|
|
| 0.012a |
|
Yes | 15 (17.86) | 4 (8.51) | 11 (29.73) |
|
| No | 69 (82.14) | 43 (91.49) | 26 (70.27) |
|
Dll4 is primarily expressed in leader
cells and Notch1 is mainly expressed in follower cells
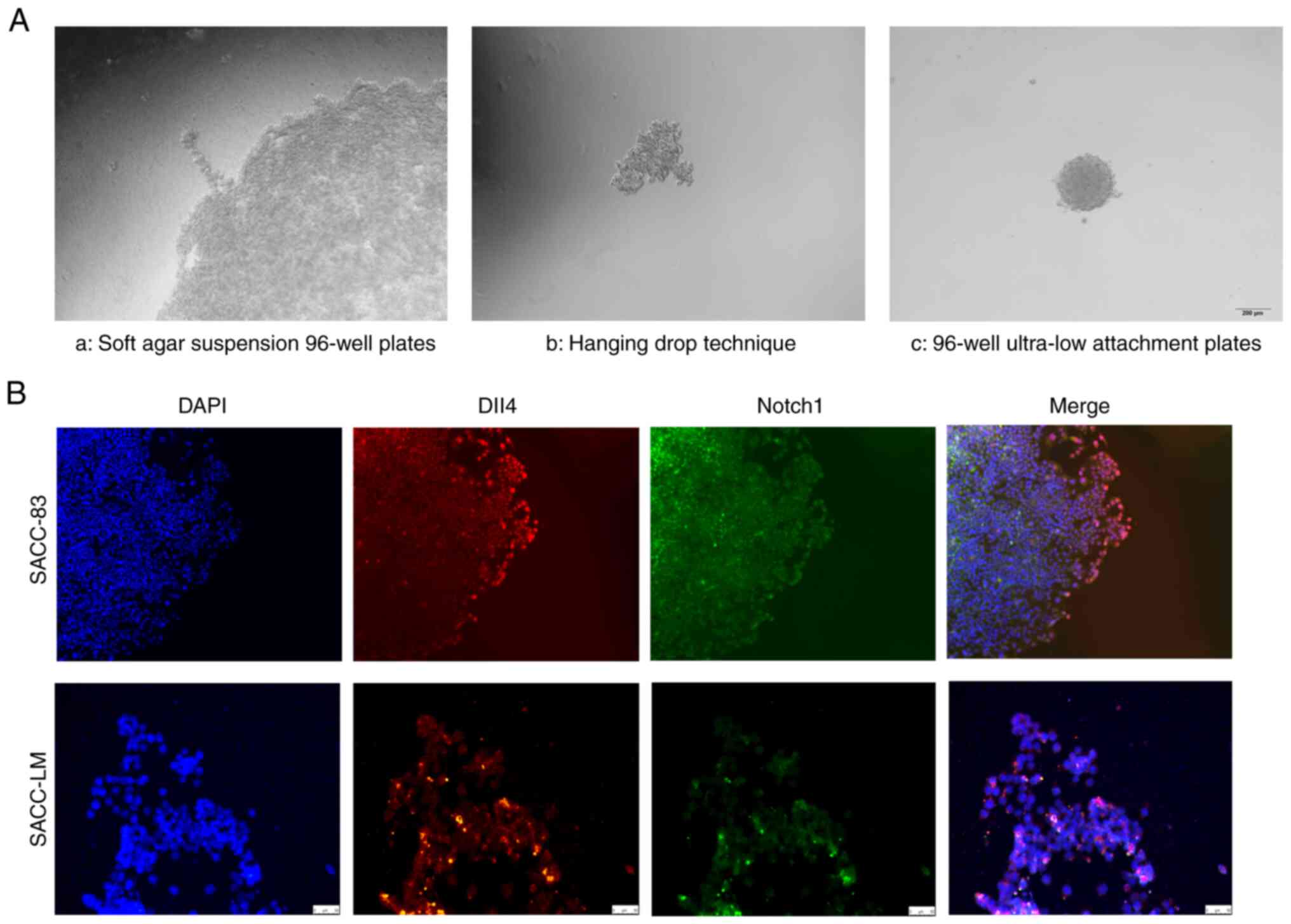
Multicellular spheroids were generated using the
hanging drop technique, agar-coated suspension technique, and
ultra-low 96-well attachment plates, and then embedded in
extracellular matrix or collagen I. After 24–72 h, several invasive
multicellular strands had extended from the spheroid margin, which
simulated in vitro collective invasion (Fig. 2A), similar to that also observed in
our previous study (12). IFC
staining revealed that leader cells at the invasive front expressed
Dll4, while follower cells within the multicellular spheroids
expressed Notch1 (Fig. 2B). Thus,
Dll4 may be a marker for leader cells and Notch1 may be a marker
for follower cells in the collective invasion of SACC.
Increased extracellular matrix
stiffness may promote SACC collective invasion and enhance
Dll4/Notch1 expression
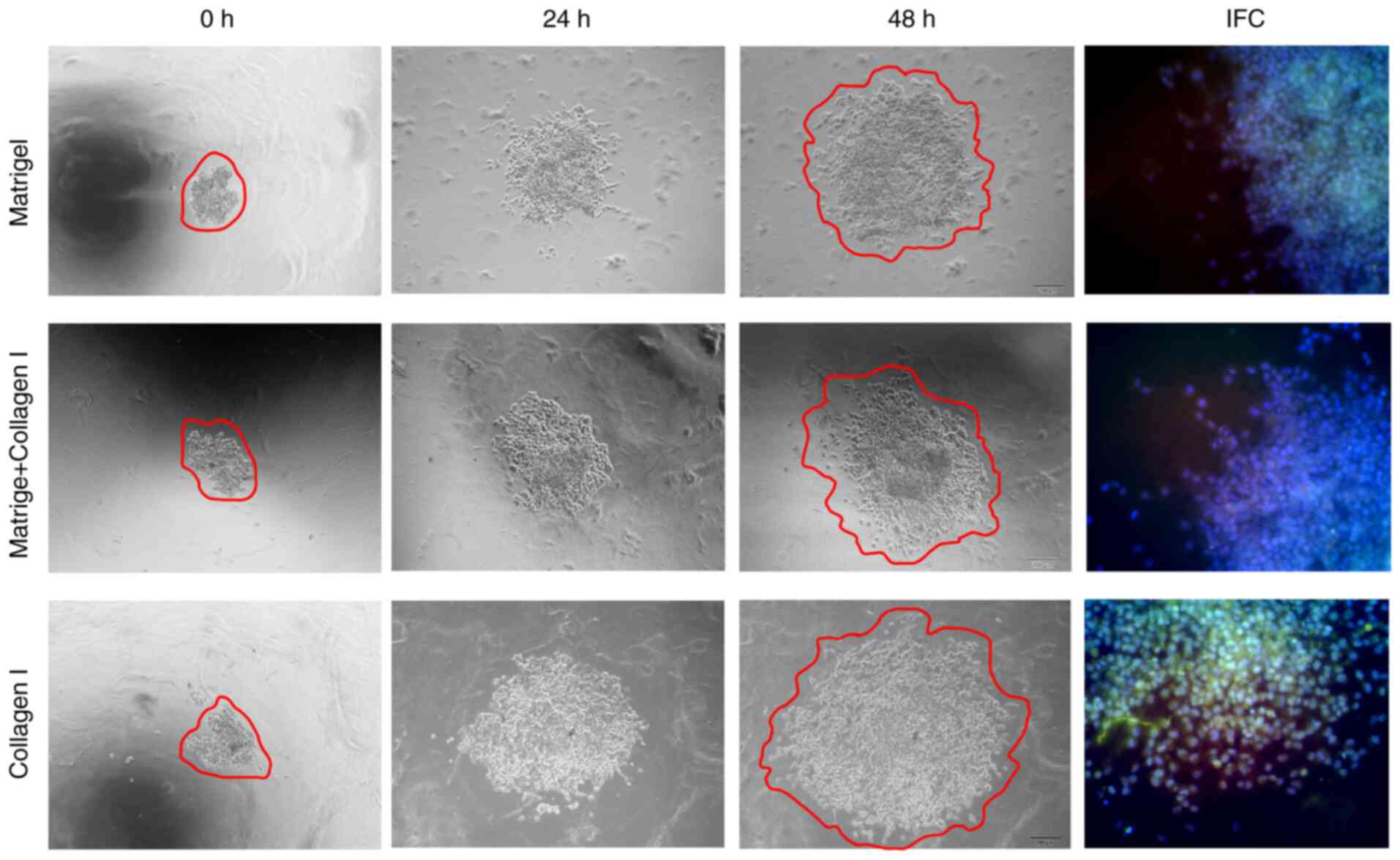
The stiffness of the 3D culture model was gradually
increased by decreasing the proportion of Matrigel and increasing
the proportion of collagen I. Greater stiffness was associated with
an increased number and range of invasive branches, as well as
increased expression of Dll4 and Notch1 based on IFC staining
(Fig. 3). These findings indicated
that increased extracellular matrix stiffness may promote
collective invasion and increase Dll4/Notch1 expression in SACC
cells.
Knockdown of Dll4 inhibits the
migratory, invasive and pro-angiogenic abilities of SACC cells
The three siRNA duplexes against Dll4 were
characterized and the siRNA with the highest efficiency was used
for subsequent experiments (Fig.
4A). Wound healing and Transwell assays revealed that
transfection with Dll4 siRNA-3 reduced the migratory and invasive
abilities of the SACC cells, relative to cells that were
transfected with control siRNA (Fig. 4B
and C). Additionally, the tube formation assay indicated that
Dll4 siRNA-3-transfected SACC cells induced less microvessel
formation of HUVECs (Fig. 4D).
Moreover, Dll4 siRNA-3-transfected SACC cells had decreased mRNA
expression levels of Notch1, Hey1, Hes1, Twist1, MMP9 and CXCR4
(Fig. 4E). Therefore, Dll4
knockdown appeared to inhibit the migratory, invasive and
pro-angiogenic capabilities of SACC cells.
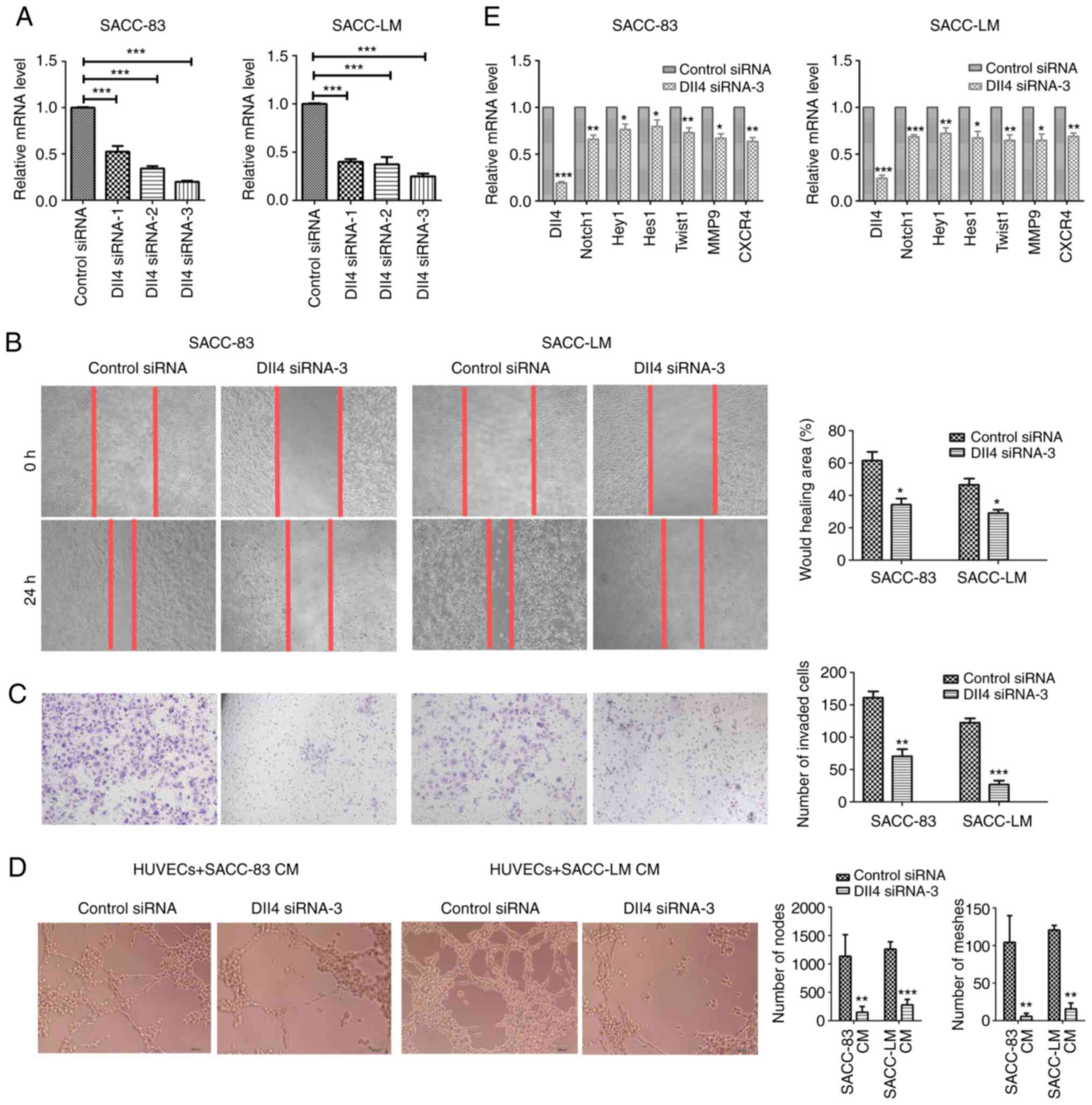 | Figure 4.Dll4 knockdown inhibits the migration
and invasion of SACC cells. (A) The relative Dll4 mRNA expression
in transfected cells was evaluated using RT-qPCR. (B) The migration
of SACC cells transfected with Dll4 siRNA-3 or control siRNA was
determined using a wound healing assay (magnification, ×100). (C)
The invasion of SACC cells transfected with Dll4 siRNA-3 or control
siRNA was measured using a Transwell assay (magnification, ×100).
(D) The tube formation ability of HUVECs treated with CM of SACC
cells was quantified by measuring the number of nodes and the
number of meshes (magnification, ×100). (E) In SACC cells, RT-qPCR
revealed that Dll4 knockdown was associated with the downregulation
of Dll4, Notch1, Hey1, Hes1, Twist1, MMP9 and CXCR4 mRNA
expression. The data are presented as the mean ± standard deviation
(n=3). *P<0.05, **P<0.01, ***P<0.001 vs. control siRNA or
as indicated. SACC, salivary adenoid cystic carcinoma; Dll4, δ-like
ligand 4; CM, conditioned medium; RT-qPCR, reverse
transcription-quantitative polymerase chain reaction; siRNA, small
interfering RNA; HUVECs, human umbilical vein endothelial cells;
Hey1, hairy/enhancer-of-split related with YRPW motif protein 1;
Hes1, transcription factor HES-1; Twist1, Twist-related protein 1;
CXCR4, C-X-C chemokine receptor type 4. |
Hypoxia promotes SACC collective
invasion by modulating the Dll4/Notch1 signalling pathway
A hypoxic environment is a common characteristic of
solid tumours and contributes to the invasion and metastasis of
tumour cells (22). Thus, whether
hypoxia could regulate SACC collective invasion by modulating the
Dll4/Notch1 signalling pathway was analysed. Under hypoxic
conditions, it was observed that the spheroids developed
multicellular strands from their margins, which exhibited
mesenchymal features. IFC staining suggested that Dll4 and Notch1
were highly expressed in the invading cells, especially in the
leader cells. However, siRNA silencing of Dll4 reduced the
crab-like invasive processes of the leader cells and decreased the
expression of Dll4 and Notch1 (Fig.
5A). The hypoxic conditions appeared to enhance the migratory
and invasive abilities of SACC cells, which could be partially
inhibited by silencing Dll4 (Fig. 5B
and C). Furthermore, under hypoxic conditions, Dll4/Notch1 and
their downstream molecules (Hey1, Hes1, HIF-1α, MMP9 and Twist1)
expression levels were increased, but lower expression levels were
observed in SACC cells transfected with Dll4 siRNA (Fig. 5D). These results implied that the
Dll4/Notch1 signalling pathway was modulated by hypoxia to
contribute to the collective invasion of SACC.
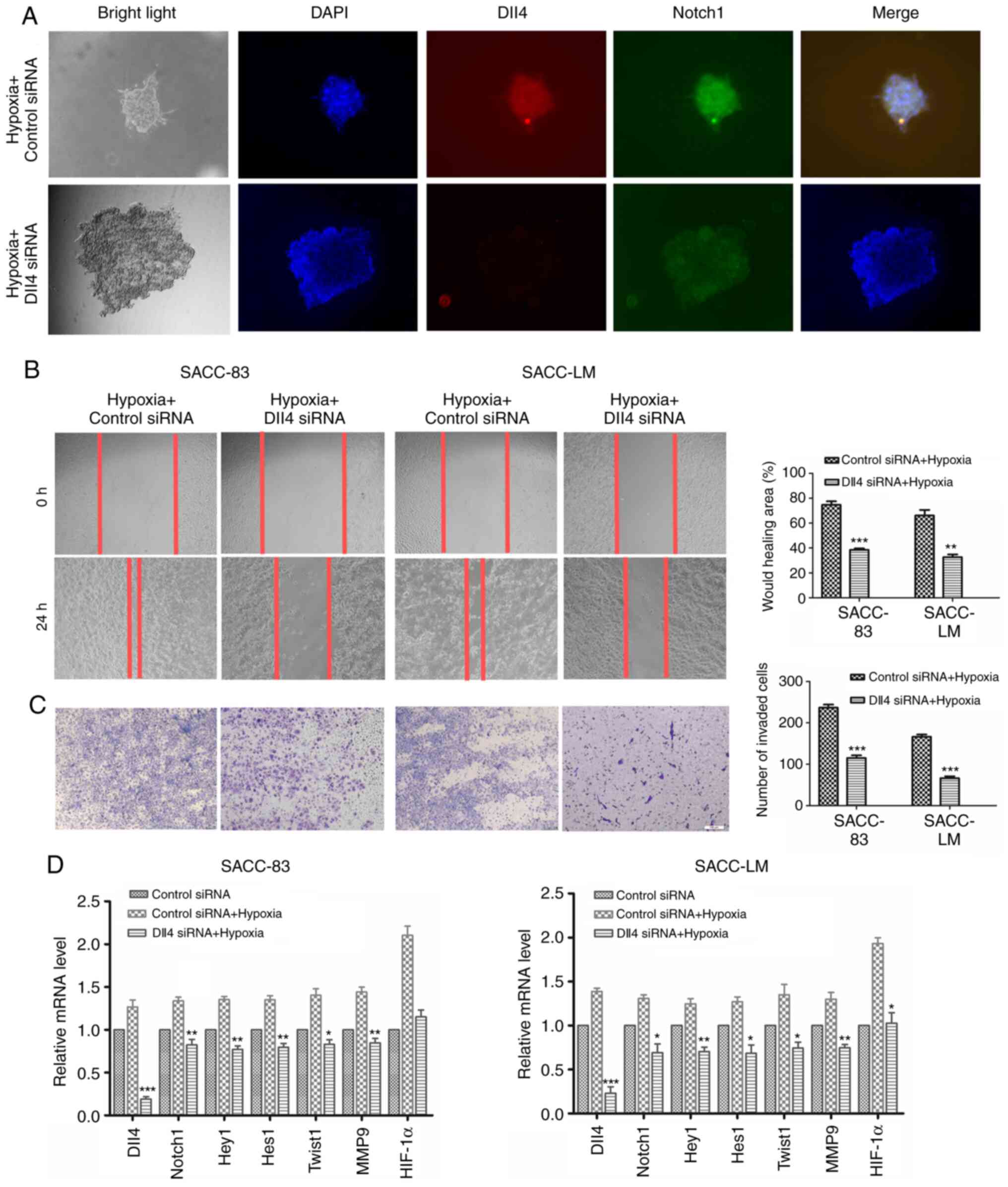 | Figure 5.Hypoxia regulates SACC collective
invasion via the Dll4/Notch1 signalling pathway. (A)
Immunofluorescence results for Dll4 and Notch1 expression in SACC
cells with or without Dll4 knockdown under hypoxia (magnification,
×40). (B) The migration of SACC cells transfected with Dll4 siRNA-3
or control siRNA under hypoxia (magnification, ×100). (C) The
invasion of SACC cells transfected with Dll4 siRNA-3 or control
siRNA under hypoxia (magnification, ×100). (D) The relative mRNA
expression levels of Dll4, Notch1, Hey1, Hes1, Twist1, MMP9 and
HIF1-α in SACC cells transfected with Dll4 siRNA-3 or control siRNA
under hypoxia. The data are presented as the mean ± standard
deviation (n=3). *P<0.05, **P<0.01, ***P<0.001 vs. control
siRNA + Hypoxia. SACC, salivary adenoid cystic carcinoma; Dll4,
δ-like ligand 4; siRNA, small interfering RNA; Hey1,
hairy/enhancer-of-split related with YRPW motif protein 1; Hes1,
transcription factor HES-1; Twist1, Twist-related protein 1;
HIF1-α, hypoxia-inducible factor 1-α. |
Co-culture with HUVECs promotes
collective invasion in SACC by regulating the Dll4/Notch1
signalling pathway
Previous research has suggested that Dll4 could
mediate the HUVEC phenotype and promote mature vessel formation
(23). Thus, whether HUVECs could
also modulate Dll4/Notch1 expression and SACC collective invasion
was investigated. Co-culture with HUVECs upregulated the expression
of Dll4 and Notch1 in SACC cells, and led to enhanced migratory and
invasive abilities. Furthermore, increased expression levels of
Dll4/Notch1 and their downstream molecules (Hey1, Hes1, VEGFA, MMP9
and Twist1) were also observed in SACC cells co-cultured with
HUVECs (Fig. 6B-D). Silencing of
Dll4 partially inhibited the crab-like invasive processes of the
leader cells during co-culture with HUVECs, reduced the migratory
and invasive abilities of cells, and downregulated the expression
of Hey1, Hes1, VEGFA, MMP9 and Twist1 (Fig. 6A-D). These results implied that the
Dll4/Notch1 signalling pathway was important in HUVEC-related
induction of SACC collective invasion.
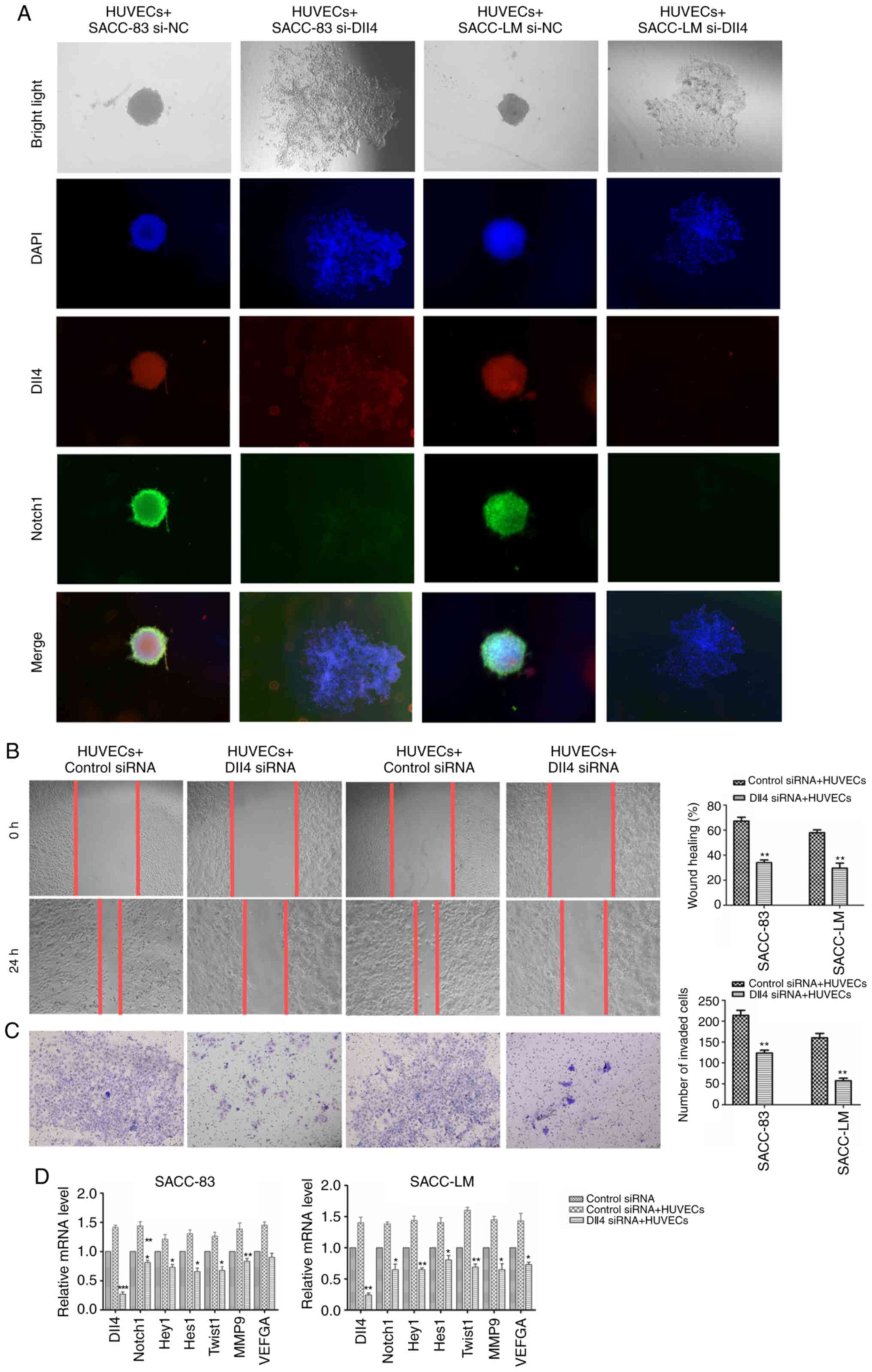 | Figure 6.HUVECs regulates SACC collective
invasion via the Dll4/Notch1 signalling pathway. (A)
Immunofluorescence results for Dll4 expression (red) and Notch1
expression (green) in SACC cells transfected with Dll4 siRNA-3 or
control siRNA during co-culture with HUVECs (magnification, ×40).
(B) The migration of SACC cells transfected with Dll4 siRNA-3 or
control siRNA during co-culture with HUVECs (magnification, ×100).
(C) The invasion of SACC cells transfected with Dll4 siRNA-3 or
control siRNA during co-culture with HUVECs (magnification, ×100).
(D) The relative mRNA expression levels of Dll4, Notch1, Hey1,
Hes1, Twist1, MMP9 and VEGFA in SACC cells transfected with Dll4
siRNA-3 or control siRNA during co-culture with HUVECs. The data
are presented as the mean ± standard deviation (n=3). *P<0.05,
**P<0.01, ***P<0.001 vs. control siRNA + HUVECs. SACC,
salivary adenoid cystic carcinoma; Dll4, δ-like ligand 4; siRNA,
small interfering RNA; HUVECs, human umbilical vein endothelial
cells; Hey1, hairy/enhancer-of-split related with YRPW motif
protein 1; Hes1, transcription factor HES-1; Twist1, Twist-related
protein 1. |
Discussion
Unlike the classic single-cell invasion that
accompanies the EMT, collective invasion plays an important role in
the invasion and metastasis of various epithelial tumours (24,25).
Early research regarding this topic used H&E staining of SACC
specimens and IHC staining for EMT-related molecules, which
revealed that collective invasion was an important mechanism for
SACC invasion (12,26). The present study revealed that Dll4
and Notch1 were highly expressed at the invasive front of SACC,
while knockdown of Dll4 inhibited SACC collective invasion and
reduced the in vitro migratory and invasive abilities of
SACC cells. Furthermore, greater extracellular matrix stiffness,
hypoxia and co-culture with HUVECs appeared to promote SACC
collective invasion by upregulating Dll4. Therefore, the present
results suggested that Dll4 and Notch1 facilitated the collective
invasion of SACC cells.
Initially, the expression of Dll4 and Notch1 in SACC
tissues was evaluated, which revealed that Dll4 and Notch1 were
highly expressed at the invasive front of SACC specimens. In
addition, higher expression of Dll4 and Notch1 at the invasive
front was associated with higher TNM staging and higher rates of
metastasis and recurrence. Similarly, Torab et al (27) reported that Dll4 was upregulated in
leader cells using a 3D microtumour invasion model and specimens
from patients with bladder cancer. Another study revealed that
leader cells could be identified based on Dll4 expression using a
computational model, and that creating a cell-free region reduced
mechanical stress, induced Dll4 expression, and induced leader cell
formation during collective cell migration (15). These results suggested that the
Dll4/Notch1 signalling pathway may play an important role in the
collective invasion of SACC, and that Dll4/Notch1 expression might
be useful for predicting the prognosis of patients with SACC.
3D models of SACC collective invasion were created
using agarose suspension culture, hanging drop culture, and
ultra-low adhesion 96-well plate suspension culture. The IFC
results revealed high expression of Dll4 in the leader cells and
high expression of Notch1 in the follower cells. Furthermore,
silencing of Dll4 using siRNA reduced the migratory, invasive and
pro-angiogenic abilities of SACC-83 and SACC-LM cells. Another
study suggested that downregulating Dll4 notably decreased the
invasion, migration and metastatic properties of oesophageal cancer
cells via the PI3K/Akt/E-cadherin pathway (28). Downregulation of Dll4/Notch4 has
also been reported to reduce endocrine therapy resistance and
metastasis in breast cancer (29).
Moreover, it was observed in the present study that knockdown of
Dll4 downregulated the expression of Hey1, Hes1, MMP9, CXCR4 and
Twist1, which are downstream factors in the Dll4/Notch1 signalling
pathway. These results also indicated that Dll4 contributed to the
collective invasion of SACC cells.
A hypoxic environment induces tumour invasion and
metastasis in several cancer types, including SACC (30,31).
Thus, it was evaluated whether the Dll4/Notch1 signalling pathway
promoted SACC collective invasion under hypoxic conditions. The
results indicated that hypoxic conditions increased and elongated
the long fusiform and crab claw-like invasive processes of leader
cells, which suggested that hypoxic conditions promoted the
collective invasion of SACC cells. Furthermore, the migratory and
invasive abilities of SACC-LM were enhanced under hypoxic
conditions, while silencing of Dll4 using siRNA partially mitigated
the pro-invasive effects of the hypoxic conditions. Co-culture with
HUVECs also upregulated Dll4/Notch1 expression in the leader and
follower cells, while knockdown of Dll4 decreased the crab-like
invasive processes of leader cells in the HUVEC co-culture. Using a
two-dimensional cell culture system, SACC cell migration and
invasion was markedly enhanced during HUVEC co-culture, although
Dll4 silencing partially inhibited the SACC cells' enhanced
migration, invasion and expression of VEGFA. Mendonça et al
(32) demonstrated that mice with
endothelial-specific Dll4 loss-of-function mutations had increased
tumour hypoxia, but also a significantly decreased the number and
burden of macro-metastases, while Dll4/Notch signalling appeared to
mediate the tumour hypoxia-driven increase in EMT. When considered
with the present results, it appears that hypoxia and HUVECs might
promote SACC collective invasion via the Dll4/Notch1 signalling
pathway.
In conclusion, Dll4 and Notch1 were highly expressed
in leader and follower cells, and appeared to play important roles
in SACC collective invasion. Furthermore, increased extracellular
matrix stiffness, hypoxia and co-culture with HUVECs appeared to
promote SACC collective invasion by upregulating Dll4. These
results suggested that Dll4 expression may define the leader cells
in SACC and may be required for collective cell invasion, which
also involves Notch1. Therefore, Dll4 might be a novel target for
SACC treatment. Further studies should be performed using more
surgical specimens from a range of clinic centres to improve the
validity of the present results.
Acknowledgements
Not applicable.
Funding
The present study was supported by National Natural
Science Foundation of China grants (grant nos. 82073000, 81902779
and 81972542), National Science Foundation of Sichuan Province
(grant nos. 2020JDRC0018 and 2020YFS0171), Clinical Project of West
China College of Stomatology, Sichuan University (grant no.
LCYJ2019-8) and Exploration and research projects of West China
College of Stomatology Sichuan University (grant no.
LCYJ2020-YJ-1).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
KW performed most of the experiments and wrote the
manuscript. HYF, XP and MZ assisted in experiments. XHY, JSW, BJC
and JJ analyzed the data, assisted in manuscript writing and
confirmed the authenticity of all the raw data. XHL and YLT
conceived the study and performed the final corrections. All the
authors read and approved the final manuscript.
Ethics approval and consent to
participate
The samples and clinical data were obtained with
informed consent from the patients, and the protocols were approved
by the Institutional Ethics Committee of the West China Medical
Center, Sichuan University (Chengdu, China; approval no.
WCHSIRB-D-2017-208).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Nascimento AG, Amaral AL, Prado LA,
Kligerman J and Silveira TR: Adenoid cystic carcinoma of salivary
glands. A study of 61 cases with clinicopathologic correlation.
Cancer. 57:312–319. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fordice J, Kershaw C, El-Naggar A and
Goepfert H: Adenoid cystic carcinoma of the head and neck:
Predictors of morbidity and mortality. Arch Otolaryngol Head Neck
Surg. 125:149–152. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Friedl P and Gilmour D: Collective cell
migration in morphogenesis, regeneration and cancer. Nat Rev Mol
Cell Biol. 10:445–457. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Brabletz T: To differentiate or not-routes
towards metastasis. Nat Rev Cancer. 12:425–436. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Savagner P: Epithelial-mesenchymal
transitions: From cell plasticity to concept elasticity. Curr Top
Dev Biol. 112:273–300. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Christiansen JJ and Rajasekaran AK:
Reassessing epithelial to mesenchymal transition as a prerequisite
for carcinoma invasion and metastasis. Cancer Res. 66:8319–8326.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Clark AG and Vignjevic DM: Modes of cancer
cell invasion and the role of the microenvironment. Curr Opin Cell
Biol. 36:13–22. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gialeli C, Theocharis AD and Karamanos NK:
Roles of matrix metalloproteinases in cancer progression and their
pharmacological targeting. FEBS J. 278:16–27. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cheung KJ, Gabrielson E, Werb Z and Ewald
AJ: Collective invasion in breast cancer requires a conserved basal
epithelial program. Cell. 155:1639–1651. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Han P, Liu J, Lei Y, Lin Z, Tian D and Yan
W: Netrin-1 promotes the collective cell migration of liver cancer
cells in a 3D cell culture model. J Physiol Biochem. 75:489–498.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu JS, Li ZF, Wang HF, Yu XH, Pang X, Wu
JB, Wang SS, Zhang M, Yang X, Cao MX, et al: Cathepsin B defines
leader cells during the collective invasion of salivary adenoid
cystic carcinoma. Int J Oncol. 54:1233–1244. 2019.PubMed/NCBI
|
|
13
|
Ding XY, Ding J, Wu K, Liu C, Yan HX, Chen
C, Wang S, Tang H, Gao CK, Guo LN, et al: Cross-talk between
endothelial cells and tumor via delta-like ligand 4/Notch/PTEN
signaling inhibits lung cancer growth. Oncogene. 31:2899–2906.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang YZ, Qin F, Han ZG, Liu Q, Zhou L and
Wang YW: Prognostic significance of DLL4 expression in papillary
thyroid cancer. Eur Rev Med Pharmacol Sci. 19:2901–2905.
2015.PubMed/NCBI
|
|
15
|
Riahi R, Sun J, Wang S, Long M, Zhang DD
and Wong PK: Notch1-Dll4 signalling and mechanical force regulate
leader cell formation during collective cell migration. Nat Commun.
6:65562015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tsukamoto H, Kato T, Enomoto A, Nakamura
N, Shimono Y, Jijiwa M, Asai N, Murakumo Y, Shibata K, Kikkawa F
and Takahashi M: Expression of Ret finger protein correlates with
outcomes in endometrial cancer. Cancer Sci. 100:1895–1901. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Iwakoshi A, Murakumo Y, Kato T, Kitamura
A, Mii S, Saito S, Yatabe Y and Takahashi M: RET finger protein
expression is associated with prognosis in lung cancer with
epidermal growth factor receptor mutations. Pathol Int. 62:324–330.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Korff T and Augustin HG: Integration of
endothelial cells in multicellular spheroids prevents apoptosis and
induces differentiation. J Cell Biol. 143:1341–1352. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xu Z, Wang Z, Jia X, Wang L, Chen Z, Wang
S, Wang M, Zhang J and Wu M: MMGZ01, an anti-DLL4 monoclonal
antibody, promotes nonfunctional vessels and inhibits breast tumor
growth. Cancer Lett. 372:118–127. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ishigami S, Arigami T, Uenosono Y, Okumura
H, Kurahara H, Uchikado Y, Setoyama T, Kita Y, Kijima Y, Nishizono
Y, et al: Clinical implications of DLL4 expression in gastric
cancer. J Exp Clin Cancer Res. 32:462013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lu X and Kang Y: Hypoxia and
hypoxia-inducible factors: Master regulators of metastasis. Clin
Cancer Res. 16:5928–5935. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Harrington LS, Sainson RC, Williams CK,
Taylor JM, Shi W, Li JL and Harris AL: Regulation of multiple
angiogenic pathways by Dll4 and Notch in human umbilical vein
endothelial cells. Microvasc Res. 75:144–154. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Khalil AA, Ilina O, Gritsenko PG, Bult P,
Span PN and Friedl P: Collective invasion in ductal and lobular
breast cancer associates with distant metastasis. Clin Exp
Metastasis. 34:421–429. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Westcott JM, Prechtl AM, Maine EA, Dang
TT, Esparza MA, Sun H, Zhou Y, Xie Y and Pearson GW: An
epigenetically distinct breast cancer cell subpopulation promotes
collective invasion. J Clin Invest. 125:1927–1943. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gao XL, Wu JS, Cao MX, Gao SY, Cen X,
Jiang YP, Wang SS, Tang YJ, Chen QM, Liang XH and Tang Y:
Cytokeratin-14 contributes to collective invasion of salivary
adenoid cystic carcinoma. PLoS One. 12:e01713412017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Torab P, Yan Y, Yamashita H, Warrick JI,
Raman JD, DeGraff DJ and Wong PK: Three-dimensional microtumors for
probing heterogeneity of invasive bladder cancer. Anal Chem.
92:8768–8775. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Guo X, Duan Y, Ye X, Hu L, Xu T, Tong L
and Yu M: Stable silencing of dll4 gene suppresses the growth and
metastasis of esophagus cancer cells by attenuating Akt
phosphorylation. Oncol Rep. 40:495–503. 2018.PubMed/NCBI
|
|
29
|
McClements L, Annett S, Yakkundi A,
O'Rourke M, Valentine A, Moustafa N, Alqudah A, Simões BM, Furlong
F, Short A, et al: FKBPL and its peptide derivatives inhibit
endocrine therapy resistant cancer stem cells and breast cancer
metastasis by downregulating DLL4 and Notch4. BMC Cancer.
19:3512019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
He C, Wang L, Zhang J and Xu H:
Hypoxia-inducible microRNA-224 promotes the cell growth, migration
and invasion by directly targeting RASSF8 in gastric cancer. Mol
Cancer. 16:352017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Wang HF, Wang SS, Zheng M, Dai LL, Wang K,
Gao XL, Cao MX, Yu XH, Pang X, Zhang M, et al: Hypoxia promotes
vasculogenic mimicry formation by vascular endothelial growth
factor A mediating epithelial-mesenchymal transition in salivary
adenoid cystic carcinoma. Cell Prolif. 52:e126002019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mendonça L, Trindade A, Carvalho C,
Correia J, Badenes M, Gigante J and Duarte A: Metastasis is
impaired by endothelial-specific Dll4 loss-of-function through
inhibition of epithelial-to-mesenchymal transition and reduction of
cancer stem cells and circulating tumor cells. Clin Exp Metastasis.
36:365–380. 2019. View Article : Google Scholar : PubMed/NCBI
|