Introduction
Lung cancer is the most common type of malignant
tumor, and is the primary cause of cancer-related death worldwide
(1). As the primary cause of
mortality in men and women, the number of deaths from lung cancer
was 135,720, which accounted for 22.5% of all cancer-related deaths
in the United States in 2020 (2).
Small cell lung cancer (SCLC) and non-SCLC (NSCLC), the two primary
types of lung cancer, account for >85% of all lung cancer cases
(3). Despite the progress in clinical
management that has been made in the last few years, the 5-year
overall survival (OS) rate of NSCLC is 15–21% (1,2). The
primary cause of the low 5-year survival rate is that the majority
of patients are diagnosed with advanced stage cancer with distant
metastases at the first presentation (3). Additionally, cancer recurrence and drug
resistance contribute to the high mortality rates of NSCLC
(4). The poor prognosis of NSCLC can
be attributed to the complicated and unclear molecular mechanisms
underlying its development and progression (5,6).
Therefore, it is vital to identify novel biomarkers or therapeutic
targets to improve the prognosis of NSCLC.
Multiple factors may participate in the development
and progression of NSCLC, including proliferation, autophagy,
apoptosis, invasion, metastasis and drug resistance (7,8). One of
the most fundamental characteristics of cancer cells is its ability
to sustain chronic proliferation. However, proliferation and
division of malignant cells is uncontrolled (9). Autophagy serves an important and
complicated role in tumor development. Upregulation of autophagy in
cancer therapy can promote the survival or death of tumor cells
(10). Abnormal regulation of cell
death, whether too little or too much, may contribute to several
diseases. Aberrant initiation of apoptosis may lead to malignant
transformation of NSCLC cells (11,12).
Metastasis of NSCLC is a significant obstacle reducing the OS of
NSCLC patients, and is considered a core step in the malignant
progress of NSCLC (13). Although
chemotherapy prolongs the OS of patients with NSCLC, tumor cells
may acquire resistance, resulting in poor therapeutic effects,
tumor metastasis and recurrence (7).
Therefore, it is necessary to identify and determine the
relationship between novel biomarkers and malignant behavior in
NSCLC.
CircRNAs are a novel group of non-coding RNAs that
do not possess 3′ and 5′ends, but instead form a closed-loop
dissimilar to linear RNAs (14).
CircRNAs were first detected in a virus by Sanger in 1976, and were
initially deemed as irrelevant byproducts without any significant
biological functions for a period of time (15). In the last decade, owing to the rapid
advance of RNA-sequencing technologies, researchers have
re-evaluated the crucial functions of circRNAs in the regulation of
gene expression and in multiple diseases, such as carcinomas
(16). Additionally, previous studies
have indicated that circRNAs are conserved, stable and abundantly
expressed in tissues and exosomes (17,18).
According to reverse transcription quantitative (RT-qPCR) and
reverse transcription-droplet digital (RT-ddPCR) qualification, 343
differentially-expressed circRNAs were identified between the
plasma of patients with gastric cancer and healthy controls
(19). CircRNAs are closely
associated with tumorigenesis, development, proliferation,
apoptosis, invasion and migration of various physiological and
pathological processes in tumors (20). CircRNAs are extensively and stably
expressed in the plasma and exosomes, indicating that they may
serve as promising biomarkers in the prognosis and therapeutics of
malignancies (21). The present
review summarizes the relationship between circRNAs and the
biological behaviors of NSCLC.
Classification and functions of
circRNAs
Classification of circRNAs
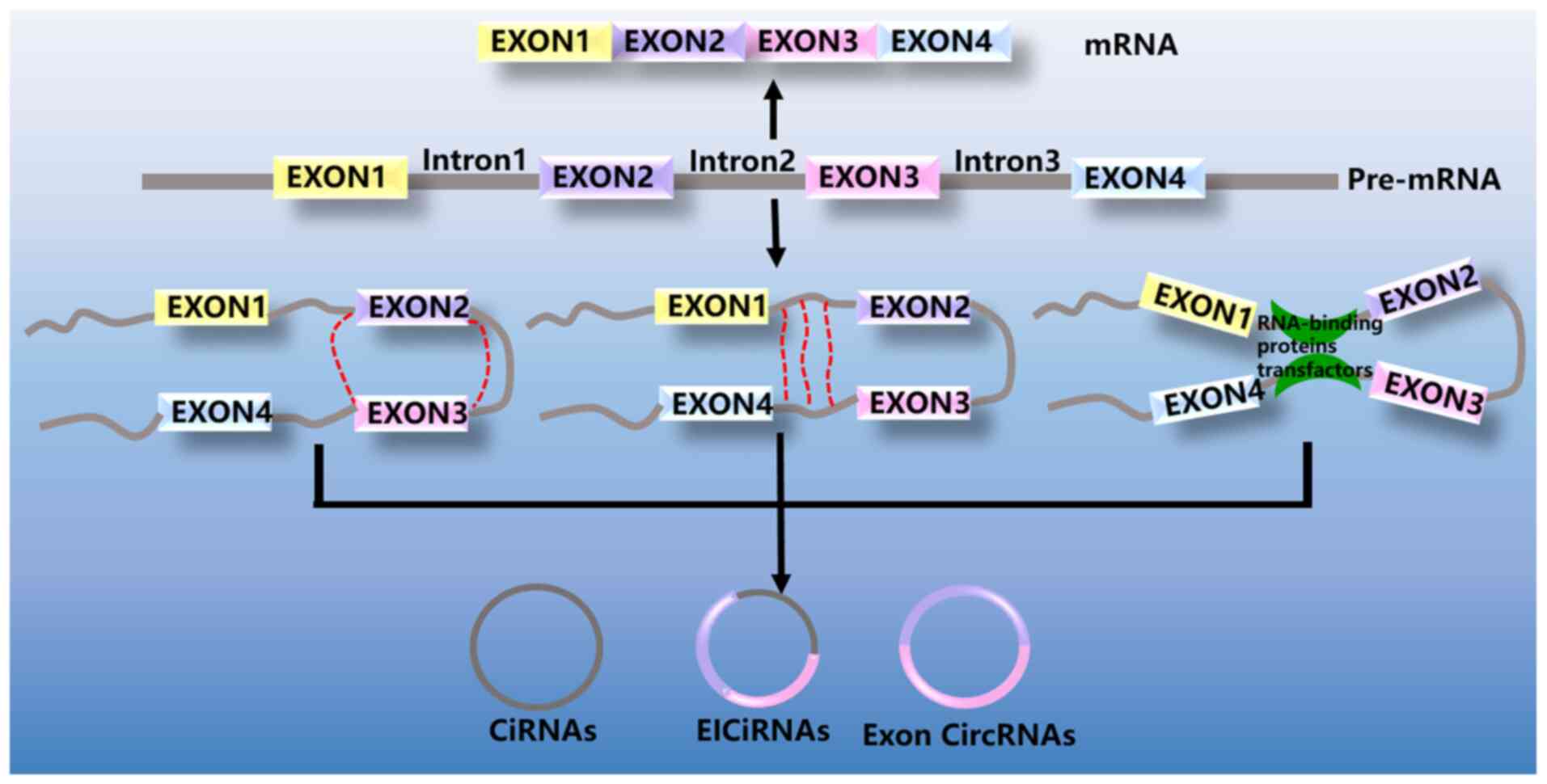
CircRNAs can be divided into four groups, exonic
circRNAs (ecircRNAs), intronic circRNAs (ciRNAs), exon-intron
circRNA (EIciRNAs) (Fig. 1) and tRNA
intronic circRNAs (tricRNAs). EcircRNAs are generated from single
or several exons. The majority of circRNAs are ecircRNAs,
accounting for >80% of currently identified circRNAs. CircRNAs
are primarily localized in the cytoplasm and may act as miRNA
sponges, indirectly participating in the regulation of gene
expression. CiRNAs are intron-derived circRNAs. CiRNAs are
abundantly present in the nucleus and may modulate the expression
of their parental genes. EIciRNAs contain both introns and exons
that can regulate their parental genes in a cis manner.
TricRNAs are derived from tRNA introns and can form stable circRNAs
via pre-tRNA splicing (15,22).
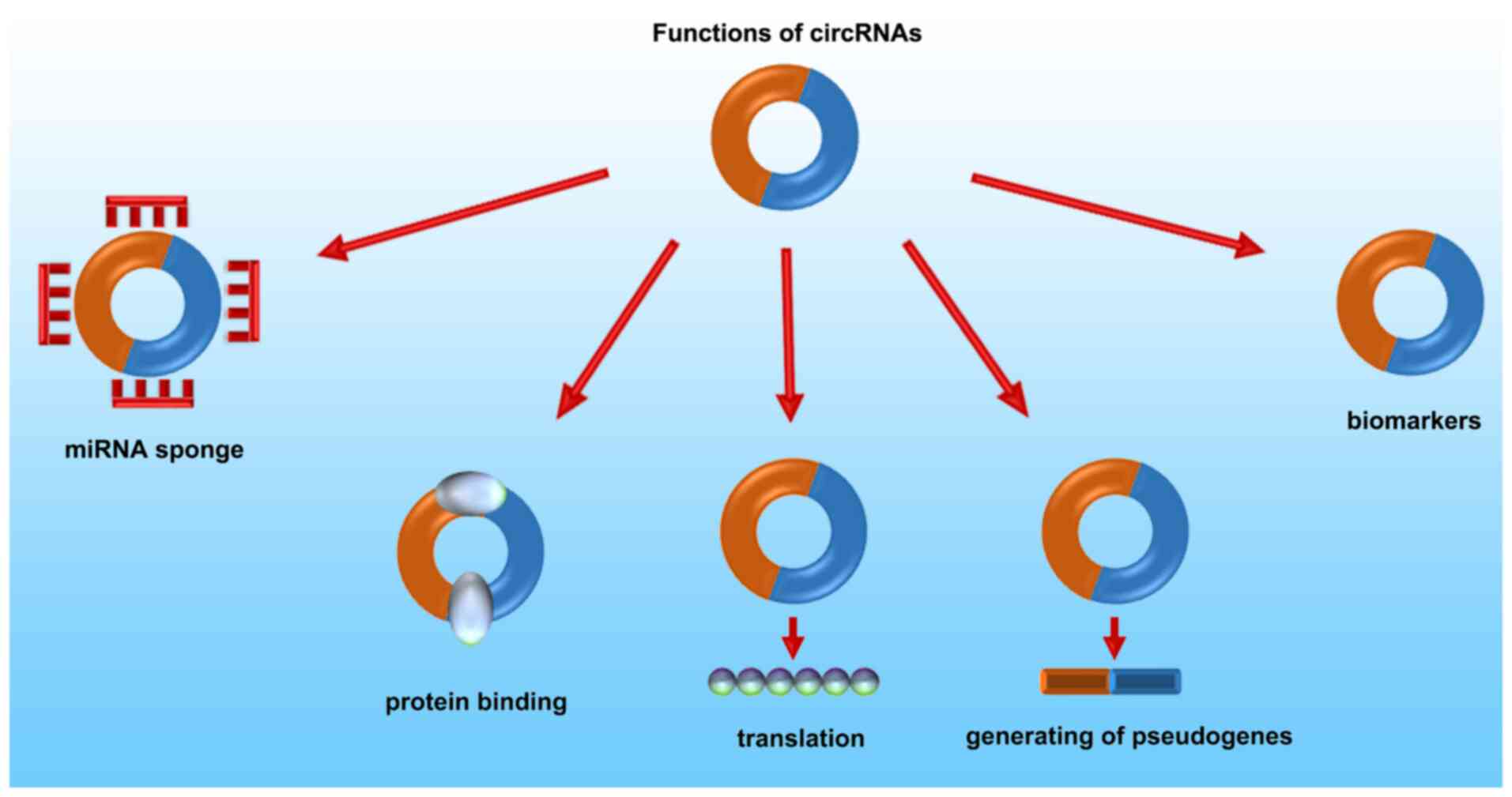
Functions of circRNAs
With the number of studies on circRNAs increasing,
our understanding of the biological functions of circRNAs is ever
growing. As shown in Fig. 2, an
increasing number of studies have shown that circRNAs exhibit
multiple functions, such as functioning as miRNA sponges,
interacting with proteins, translation into proteins and regulation
of transcription (23).
CircRNAs can sponge miRNAs
Recently, several studies have found that circRNA
are primarily located in the cytoplasm. CircRNAs compete with
miRNAs to regulate gene expression via miRNA response elements
(MREs) (24). CircRNAs can increase
the levels of the target genes of miRNAs, and circRNAs with this
competitive function are termed competing endogenous (ce)RNAs. As a
well-studied function of circRNAs, ceRNAs are widely involved in
various circRNA-related diseases, particularly in cancer. For
example, the expression of miR-7 is affected by ciRS-7 which
possesses >70 selectively conserved binding sites for miR-7.
When ciRS-7 efficiently binds to miR-7, the expression of miR-7 is
attenuated and the activity of miR-7-target genes is increased. The
ciRS-7/miR-7 axis participates in numerous diseases, such as breast
cancer (25), cervical cancer
(26), gastric carcinoma (27) and hepatocellular carcinoma (28,29).
CircRNA zinc finger protein 609 (Circ-ZNF609) improves vascular
endothelial dysfunction through upregulating the expression of
myocyte enhancer factor 2A by serving as a ceRNA of miR-615-5p
(30).
CircRNAs can bind with RNA-binding
proteins (RBPs)
CircRNAs can also bind to proteins directly. Thus,
they can act as protein sponges, similar in principal to their
function as miRNA sponges. CircRNAs contain a high density of
binding sites for RBPs, and they may affect the activity of related
proteins through binding with them directly (31). A circular transcript from forkhead box
O3 (Circ-Foxo3) is related with cell cycle progression. Circ-Foxo3
affects cell cycle progression via regulation of a G1/S transition
through binding with cyclin-dependent kinase 2 (CDK2) and p21
(32). Circ-Foxo3 facilitates Foxo3
expression via interacting with the MDM proto-oncogene (MDM2) and
p53, which leads to MDM2-induced p53 ubiquitination and subsequent
degradation (33).
CircRNAs can be translated into
proteins
Although most circRNAs serve as miRNA sponges and
indirectly regulate the expression of mRNAs, emerging evidence has
shown that certain circRNAs are translatable (34). CircRNAs may contain an open-reading
frame (ORF), N6-methyladenosine modifications and/or internal
ribosome entry site (IRES) elements. Hence, circRNAs can be
translated into proteins accordingly (35,36).
Moreover, circRNAs can translate into proteins via a rolling circle
amplification mechanism in eukaryotic cells (37). Circ-ZNF609 contains an ORF and it can
be translated into a protein via a splicing event (38). CircRNA F-box and WD repeat domain
containing 7, expression of which is high in the brain, encodes
F-box and WD repeat domains containing 7–185aa, and inhibits
proliferation and cell cycle progression in cancer cells (39). A circular form of SNF2 histone linker
PHD RING helicase, which contains an ORF driven by the IRES, is
translated into SNF2 histone linker PHD RING helicase-146aa, which
is a cancer suppressor in human glioblastoma (40).
CircRNAs can regulate
transcription
CiRNAs are primarily located in the nucleus, and
possess very little MRE activity for sponging miRNAs (41). Knockdown of ciRNAs decreases the
expression of their parental genes. CiRNAs can regulate PolII
transcription in a cis manner via an RNA-RNA interaction, and can
mediate the expression of their parental genes (42,43).
EIciRNAs interact with UI small nuclear ribonucleoprotein (snRNP)
and PolII to regulate gene expression via RNA-RNA interactions. UI
snRNP is indispensable for EIciRNA-mediated regulation of
expression of parental genes (44).
Ci-ankrd52, which accumulates at transcription sites, is an
intron-derived circRNA that is produced from ANKRD52, and
Ci-ankrd52 can interact with PolII to regulate transcription of the
parent genes (45).
CircRNAs enable derivation of
pseudogenes
Pseudogenes are non-functional residues that were
formed during the evolution of a gene family, and they serve as
essential markers in the field of evolutionary and comparative
genomics (46). Pseudogenes may
participate in cellular differentiation and cancer progression
(47). There is an exon-exon junction
in a reversed order in circRNA-derived pseudogenes. In both mice
and humans, numerous circRNA-derived pseudogenes have been detected
by a computational pipeline (CIRCpseudo) (48). In all mouse strains and in the rat
reference genome, there are dozens of low-confidence circular SATB
homeobox 1-derived pseudogenes. In the gorilla and chimp genomes,
researchers identified the homologous sequences of human circular
protein kinase, DNA-activated, catalytic submit-derived and
circular calmodulin regulated spectrin associated protein 1-derived
pseudogenes (49). Though reverse
transcription may be involved in the non-colinear exon-exon
junctions of pseudogenes, the mechanism of reverse transcription
and translocation of circRNAs are still not distinct (23).
CircRNAs may serve as promising
biomarkers
CircRNAs are stably expressed both intracellularly
and in the plasma, due to their unique annular structure (14). RNA-seq analyses indicated that
>1,000 circRNAs have been identified in human exosomes and may
transfer biological activity to other cells (50). CircRNAs generated from cancer cells
can enter into the blood circulation and can be detected easily,
and may thus be used to distinguish between cancerous and healthy
individuals (14). Exosomal circRNAs
are thus potential biological markers of various types of cancer
including NSCLC (17).
Roles of circRNAs in cancer
CircRNAs are extensively implicated in the
pathological progression of multiple types of cancer, including
gastric cancer, hepatocellular carcinoma, lung cancer, colorectal
cancer and bladder cancer, amongst others (51). Moreover, these circRNAs exhibit dual
roles; serving as oncogenes and tumor suppressors dependent on the
type and potentially stage of cancer (52,53). In
the following section, the roles of circRNAs in NSCLC are discussed
in additional detail.
CircRNAs and NSCLC
Expression of circRNAs in NSCLC
With the development of next-generation sequencing
technologies and advances in bioinformatics analysis, a large
number of studies have shown that circRNAs are ectopically
expressed in several types of tumors, including NSCLC. A total of
957 abnormally expressed circRNAs were identified by human circRNA
microarray analysis in NSCLC tissues when compared with the
adjacent normal tissue (54). In
another study, 356 circRNAs were dysregulated in lung
adenocarcinoma, including 204 upregulated circRNAs and 152
downregulated circRNAs (55). By
utilizing circRNA chips, Mu et al (56) identified and annotated a total of
10,566 circRNAs in the peripheral whole blood of patients with lung
adenocarcinoma. Amongst these, 78.14% of the circRNAs were exonic,
and 3,009 circRNAs were upregulated, whereas 1,381 circRNAs were
downregulated.
CircRNAs can be used as diagnostic
biomarkers in NSCLC
CircRNAs produced by cancer cells can enter into the
blood circulation and can be detected easily (57). Thus, they can be used to distinguish
between patients with cancer from healthy individuals. Exosomal
circRNAs are potential biological markers in a range of cancer
types (14). With the development of
circRNA research, a plethora of circRNAs may eventually be used as
clinically diagnostic markers for the diagnosis of early-stage
NSCLC (Table I).
 | Table I.CircRNAs act as prognostic or
diagnostic biomarkers of non-small cell lung cancer. |
Table I.
CircRNAs act as prognostic or
diagnostic biomarkers of non-small cell lung cancer.
|
|
|
|
|
|
| ROC curve |
|
|---|
|
|
|
|
|
|
|
|
|
|---|
| First author,
year | CircRNA name | circBase ID | Dysregulation | Correlation with
clinical characteristics | Kaplan-Meier OS
curves | AUC | Sensitivity | Specificity | (Refs.) |
|---|
| Zhang et al,
2018 | Circ_0014130 |
| Up | TNM stage,
lymphatic metastasis | – | 0.878 | 0.87 | 0.848 | (58) |
| Chen et al,
2019 | Circ_100146 |
| Up | Pathological
classification, differentiation grade | – | 0.643 | 0.725 | 0.575 | (59) |
| Qiu et al,
2018 | CircPRKCI |
| Up | T stage, TNM
stage | Poor
prognosis/independent factor (Multivariate analyses: HR=2.664, 95%
CI: 1.327–5.347, P=0.006) | – | – | – | (60) |
| Chen et al,
2018 | Circ_100395 |
| Down | Cancer metastasis,
TNM stage | Poor prognosis | – | – | – | (63) |
| Han et al,
2018 | CircBANP |
| Up | Cancer metastasis,
TNM stage | Poor prognosis | – | – | – | (64) |
| Zhang et al,
2018 and Su et al, 2018 | Circular
RNAciRS-7/CDR1as |
hsa_circ_0001946 | Up | TNM stage,
lymphatic Metastasis, pathological classification | Shorter OS |
|
|
| (68,71) |
| Zhu et al,
2017 | Circ_0013958 |
hsa_circ_0013958 | Up | TNM stage,
lymphatic metastasis | – | 0.815 | 0.755 | 0.796 | (72) |
| Wang et al,
2019 | CircVANGL1 |
| Up | T stage, TNM
stage | Poor prognosis | – | – | – | (85) |
| Li et al,
2018 | CircPVT1 |
| Up | Distant
metastasis | – | 0.803 | 0.825 | 0.675 | (86) |
| Wang et al,
2019 | CircCRIM1 |
| Down | TNM stage,
lymphatic metastasis | Poor prognosis | – | – | – | (94) |
| SHI et al,
2020 | CircLARP4 |
| Down | – | Poor prognosis | – | – | – | (95) |
The expression of hsa_circ_0014130 is associated
with Tumor-Node-Metastasis (TNM) stage and lymphatic metastasis of
NSCLC. Receiver operating characteristic (ROC) curves were used to
determine the diagnostic potential of hsa_circ_0014130. The area
under the ROC curve (AUC) was 0.878, the optimum critical value of
hsa_circ_0014130 was 0.573, the sensitivity was 87% and specificity
was 84.8%. Thus, hsa_circ_0014130 may serve as a biomarker for
distinguishing NSCLC from normal tissues (58). CircRNA 100146 was shown to be
augmented in 26 cases of NSCLC, and was associated with
pathological stage and differentiation of lung cancer. ROC curve
analysis indicated that the AUC was 0.643 (95% confidence interval:
0.521–0.764), the sensitivity was 72.5% and the specificity was
57.5%. Thus, circRNA 100146 may also be used as a diagnostic marker
in NSCLC (59).
CircRNAs may serve as therapeutic
targets in NSCLC
CircRNAs are stably expressed both intracellularly
and in the plasma due to their annular structure. RNA-seq analyses
indicated that >1,000 circRNAs are present in human exosomes and
may transfer biological activity to other cells (50). Numerous circRNAs have been reported to
be involved in the tumorigenesis and progression of NSCLC, and are
being extensively assessed as potential therapeutic targets for the
treatment of NSCLC (Table I).
Circular protein kinase C iota (CircPRKCI) is
generated from exons 15 and 16 of the PRKCI gene
(chr3:170013698-170015181) and is located at the 3q26.2 amplicon.
CircPRKCI acts as a tumor promoting factor in lung adenocarcinoma
(LAD), and circPRKCI is positively correlated with T stage and TNM
stage in patients with LAD (60).
Knockdown of circPRKCI led to a decrease in tumor size and tumor
weight in nude mice. Patient-derived tumor xenografts (PDTXs) can
be used as a translational model. Intratumoral injection of
cholesterol-conjugated si-circPRKCI was used to clarify the
therapeutic potential of circPRKCI. The findings showed that the
growth of PDTX was decreased in the si-circPRKCI group. These
results highlight the therapeutic potential of circPRKCI (60). At present, EGFR tyrosine kinase
inhibitors (EGFR-TKIs) are widely used to treat NSCLC patients with
EGFR-sensitive mutations. The combination of EGFR-TKIs (gefitinib)
and knockdown of circPRKCI resulted in a more notable inhibitory
effect than gefitinib or knockdown of circPRKCI alone. This
suggests that a combination of EGFR-TKIs and attenuation of
circPRKCI may exert a synergistic effect on reducing cancer
progression (60).
Circular coiled-coil domain containing 66
(circCCDC66) is primarily located in the endoplasmic reticulum.
CircCCDC66 is involved in several types of cancer and serves as a
diagnostic and therapeutic biomarker (61). CircCCDC66 is highly expressed in LAD
and in EGFR-resistant H1975 cells. EGFR is the primary target of
TKIs for tyrosine kinase mutations, such as gefitinib and erlotinib
in LAD chemotherapy. Knockdown of focal adhesion kinase (FAK) and
hepatocyte growth factor reduces circRNA CCDC66 expression,
separately. FAK was associated with metastasis and EMT. Meanwhile,
administration of a FAK inhibitor, Y15, reduced metastasis.
Conversely, nicotinic acetylcholine receptor α7 (nAchRα7)
negatively regulates the expression of CCDC66β and circRNA CCDC66.
The regulatory effect of nAchRα7 on circRNA CCDC66 is greater than
that of FAK. Furthermore, knockdown of circRNA CCDC66 suppresses
EMT and invasion, and augments cisplatin resistance in H23 cells.
CircCCDC66 may thus serve as a novel therapeutic target for
regulating EGFR-mediated tumorigenesis in NSCLC (62).
Prognostic potential of circRNAs in
NSCLC
It is very important to evaluate the prognosis of
patients with cancer. Aberrant expression of circRNAs has been
reported to show extensive associations with clinical features of
patients with NSCLC. Meanwhile, circRNAs are considered to possess
valuable prognostic value as biomarkers in NSCLC.
According to a study with 69 cases of NSCLC, which
used RT-qPCR qualification, hsa_circ_100395 expression was found to
be lower in patients with advanced TNM stage. Additionally,
Kaplan-Meier survival curve analysis showed that the survival rate
of patients with lower expression of hsa_circ_100395 was lower
(63). Circular BTG3 associated
nuclear protein (Circ-BANP) was shown to be upregulated in lung
cancer tissues and cell lines, and was higher in patients with
stage III–IV cancer or in the metastatic tissue. Higher expression
of circ-BANP was associated with reduced OS based on Kaplan-Meier
curve analysis. Thus, circ-BANP may serve as an independent
prognostic biomarker (64).
Based on the above studies, circRNAs may be used for
the diagnosis, treatment and evaluation of prognosis of patients
with NSCLC. In subsequent studies, increased attention should be
paid to the molecular mechanisms by which circRNAs regulate cancer
development/progression and in the clinical application of
targeting circRNAs.
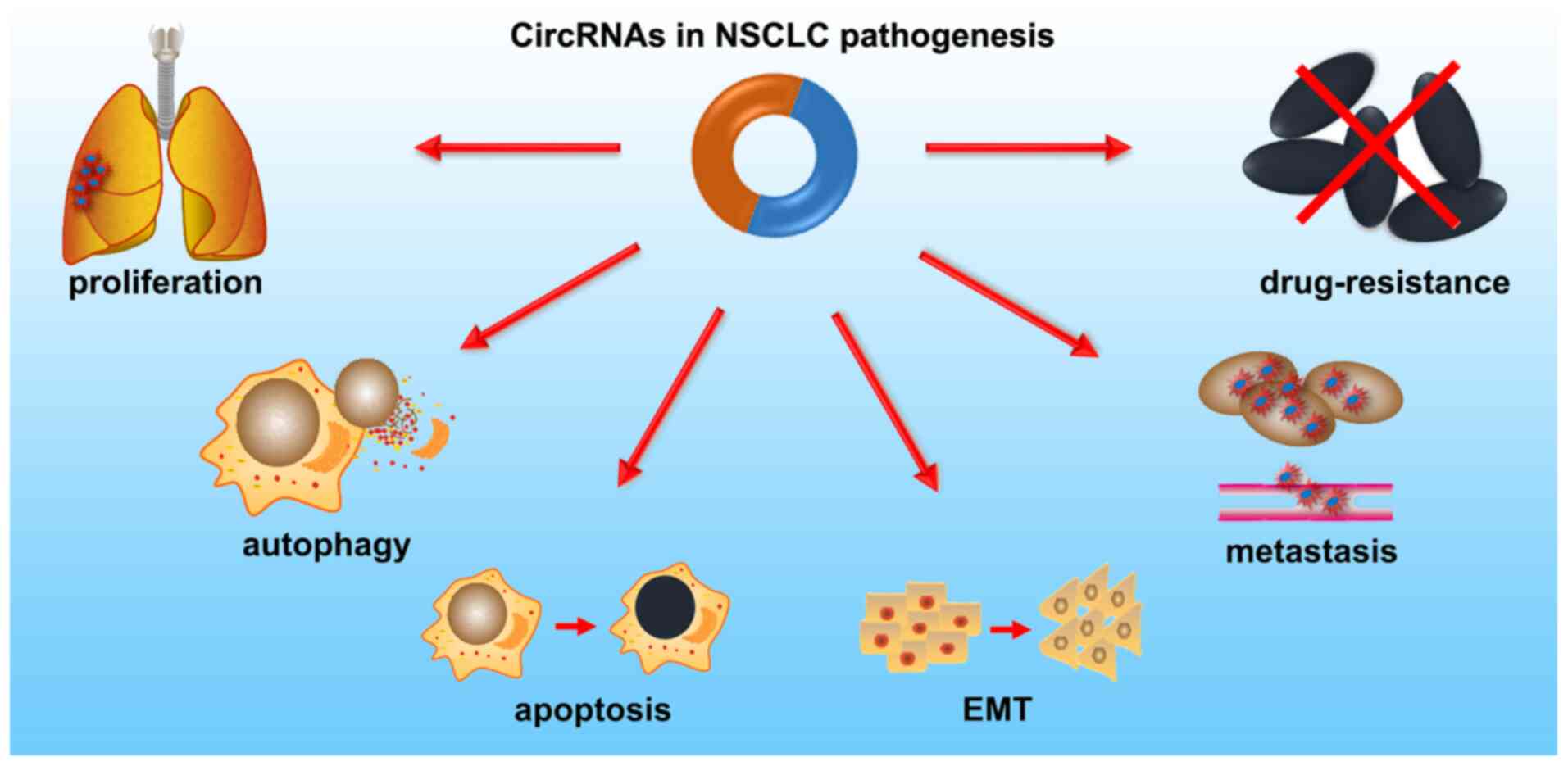
Functions of circRNAs in NSCLC
NSCLC pathogenesis is modulated by oncogenic or
tumor suppressive circRNAs, via regulation of cell proliferation,
autophagy, apoptosis, invasion, migration and EMT (Fig. 3). Additionally, circRNAs can act as
independent prognostic biomarkers, and serve an important role in
multidrug resistance (MDR) in NSCLC. The functions of circRNAs in
NSCLC are discussed in the upcoming sections and are summarized in
Table II.
| Table II.Clinical value of circRNAs involved
in the pathogenesis and progression of NSCLC. |
Table II.
Clinical value of circRNAs involved
in the pathogenesis and progression of NSCLC.
| First author,
year | CircRNA name | circBase ID | Dysregulation | Function | Target
gene/pathway | Effect in
NSCLC | (Refs.) |
|---|
| Chen et al,
2019 | Circ_100146 | – | Up | miRNA sponge | miR-615-5p↓,
miR-361-3p↓, SF3B3↑ | Proliferation↑,
invasion↑, apoptosis↓ | (59) |
| Qiu et al,
2018 | CircPRKCI | – | Up | miRNA sponge | miR-545↓, miR-589↓,
E2F7↑ | Proliferation↑,
migration↑, | (60) |
| Joseph et
al, 2018 | CircCCDC66 | – | Up | – | – | Invasion↑, EMT↑,
drug resistance (cisplatin) ↑ | (62) |
| Chen et al,
2018 | Circ_100395 | – | Down | miRNA sponge | miR-1228↓,
TCF21↑ | Proliferation↑,
migration↑, invasion↑ | (63) |
| Han et al,
2018 | CircBANP | – | Up | miRNA sponge | miR-503↓,
LARP1↑ | Proliferation↑,
migration↑, invasion↑ | (64) |
| Zhang et al,
2018 and Su et al, 2018 | Circular
RNAciRS-7/CDR1as |
hsa_circ_0001946 | Up | miRNA sponge | miR-7↓, EGFR↑,
CCNE1↑, PIK3CD↑, NF-kB↑ | Proliferation↑,
migration↑. invasion↑, apoptosis↓ | (68,71) |
| Zhu et al,
2017 | Circ_0013958 |
hsa_circ_0013958 | Up | miRNA sponge | miR-134↓,
CCND1↑ | Proliferation↑,
invasion↑, apoptosis↓ | (72) |
| Chi et al,
2019 | CircPIP5K1A |
hsa_circ_0014130 | Up | miRNA sponge | miR-600↓,
HIF-1α↑ | Apoptosis↓,
proliferation↑ | (74) |
| Nan et al,
2018 | CircNOL10 |
hsa_circ_0000977 | Down | Protein binding,
regulation of transcriptional | SCML1↑, HN
polypeptide family | Apoptosis↑,
proliferation↓, invasion↓ | (75) |
| Chen et al,
2020 | CircHIPK3 |
hsa_circ_0000284 | Up | miRNA sponge | miR-124-3p↓,
STAT3/PRKAA/AMPKa↑ | Proliferation↑,
invasion↑, migration↑, autophagy↑ | (77) |
| Wang et al,
2019 | CircVANGL1 | – | Up | miRNA sponge | miR-195↓, Bax↑,
Bcl-2↑ | Migration↑,
invasion↑, apoptosis↓ | (85) |
| Li et al,
2018 | CircPVT1 | – | Up | miRNA sponge | miR-125b↓,
E2F2↑ | Proliferation↑,
migration↑, invasion↑, apoptosis↓ | (86) |
| Gao and Ye,
2020 | CircSOX4 | – | Up | miRNA sponge | miR-1270↓,
PLAGL2↑ | Proliferation↑,
invasion↑, migration↑ | (91) |
| Wang et al,
2018 | CircPTK2 | – | Down | miRNA sponge | miR-429↓,
miR-200b-3p↓, TIF1γ↑ | Invasion↓,
TGF-β-induced EMT↓ | (92) |
| Wang et al,
2019 | CircCRIM1 |
hsa_circ_0002346 | Down | miRNA sponge | miR-93↓, miR-182↓,
LIFR↑, MMP13↑, PI3K/AKT/ JAK1↑ | Invasion↓,
metastasis↓ | (94) |
| Shi et al,
2020 | CircLARP4 | – | Down | Protein
binding | SMAD7↑ | Invasion↓,
migration↓ | (95) |
| Wan et al,
2016 | CircITCH | – | Down | miRNA sponge,
regulation of parental genes | miR-7↓, miR-214↓,
Wnt/β-catenin↓, ITCH↑ | Proliferation↓ | (96) |
| Huang et al,
2019 | Circ_0001946 | – | Down | miRNA sponge | miR-7-5p↓,
miR-671-5p↓, miR-3156-5p↓, miR-1270↓, NER↑ | Proliferation↓,
invasion↓, migration↓, drug resistance (cisplatin) ↓,
apoptosis↑ | (100) |
| Li et al,
2019 | Circ_0002483 | – | Down | miRNA sponge | miR-182-5p↓,
miR-520q-3p↓, miR-582-3p↓, miR-587↓, GRB2↑, FOXO1↑, FOXO3↑ | Proliferation↓,
invasion↓, drug resistance (Taxol)↓ | (101) |
| Xiao et al,
2020 | Circ_103762 | – | Up | Protein
binding | CHOP↓ | Proliferation↑,
migration↑, invasion↑, drug resistance (MDR) ↑ | (102) |
CircRNAs in NSCLC proliferation and
cell cycle progression
Cell cycle progression is an important factor in
maintaining cell proliferation. The phenomenon of normal cells
inhibiting division due to contact inhibition, is termed
density-dependent inhibition of growth. When cells reach a finite
density, they halt proliferation and the cell cycle arrests at the
G0 phase of the cell cycle (65).
Unrestricted cell proliferation and reduced apoptosis results in
unlimited growth and distant metastasis of tumors. The
proliferation of cancer cells represents a typical prognostic
marker in the diagnosis of cancer (66). Abnormal expression of circRNAs leads
to growth of NSCLC cells (67),
suggesting that circRNAs serve a potential role in NSCLC treatments
targeting unlimited proliferation.
CircRNAs modulate proliferation and
cell cycle progression via sponging of miRNAs in NSCLC
CircRNAs serve their biological function in numerous
ways. CircRNA-mediated sponging of miRNAs is the most-extensively
studied circRNA mechanism.
CiRS-7, also known as cerebellar
degeneration-related protein 1 antisense RNA (CDR1as), can absorb
miRNAs, such as miR-7 and miR-671, and thus reduces the levels of
CDR1 transcripts. CiRS-7 possesses over 70 binding sites with
miR-7. Several reports have demonstrated that the CiRS-7/miR-7 axis
contributes to several pathological processes, including NSCLC.
CiRS-7 may increase cell viability and induce cell growth in NSCLC.
CiRS-7 also significantly increases the expression of
growth-related genes, including EGFR, cyclin E1 (CCNE1) and
phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit δ
(PIK3CD) (68). Lower expression
levels of EGFR maintains cell cycle arrest, facilitating mitosis
and preventing cell apoptosis (69).
CCNE1 induces rapid progression of cells through the G1/S phase via
activation of CDK2 (70). The effect
of ciRS-7 on monitoring cell cycle progression is reversed by
overexpression of miR-7. CiRS-7 also regulates cell proliferation,
invasion, migration and apoptosis via targeting miR-7 to modulate
nuclear factor-κB (NF-kB) (71).
Cyclin D1 (CCND1) is an important target of
abnormally expressed circRNAs in NSCLC, such as circ_0013958
(72). Abnormal expression of
circRNAs can regulate the cell cycle process and proliferation in
NSCLC. The primary function of CCND1 is to promote cell
proliferation. CCND1 can bind to and activate cyclin-dependent
kinase CDK4, which is unique to the G1 phase (73). Circ_0013958 was identified as a sponge
of miR-134, and circ_0013958 promotes the development of NSCLC via
upregulation of oncogenic CCND1 (72).
A high-throughput microarray assay revealed that
circular phosphatidylinositol-4-phophate 5-kinase type 1 alpha
(circPIP5K1A) was significantly upregulated in NSCLC (58). CircPIP5K1A regulates the progression
of NSCLC via activation of several signaling pathways. For example,
circPIP5K1A promotes proliferation via a miR-600/hypoxia inducible
factor-1α axis in NSCLC (74).
CircRNAs modulate proliferation and
cell cycle progression via binding with RBPs and regulating
transcription in NSCLC
Circular nucleolar protein 10 (circNOL10) is
primarily expressed in the nucleus, and is generated from exons
6–12 of pre-NOL10 mRNA. The expression of circNOL10 is
cooperatively regulated by pre-NOL10 methylation and by epithelial
splicing regulatory protein 1, a splicing factor. CircNOL10
expression is low in lung cancer. CircNOL10 directly promotes the
expression of sex comb on midleg-like 1 (SCML1) by suppression of
ubiquitination, and also promotes the transcriptional regulatory
effect of SCML1 on the humanin polypeptide family, ultimately
inhibiting the progression of lung cancer (75).
CircRNAs participate in RNA splicing
in NSCLC
The notable associations between the expression of
circ-UBR5 and differentiation degree of NSCLC has been established.
The differentiation of NSCLC is decreased following knockdown of
circ-UBR5. Circ-UBR5 may thus be used to evaluate tumor
differentiation, and as an indicator for the pathological grading
of NSCLC. Circ-UBR5 binds to splicing regulatory factors, including
KH domain containing RNA binding (QKI), NOVA alternative splicing
regulator 1 and U1 snRNA. Circ-UBR5 additionally participates in
differentiation via modulation of RNA splicing (76).
CircRNAs and NSCLC autophagy
Autophagy is a process of transporting damaged,
denatured or aging proteins and organelles to lysosomes for
digestion and degradation in cells. Autophagy serves an important
and complicated role in tumor development. Upregulation of
autophagy in cancer therapy can promote the survival or death of
tumor cells (10). Abnormal activity
of the mTOR signaling pathway, pathophysiological p53 expression
and endoplasmic reticulum stress serve key roles in autophagy of
NSCLC (12). Further studies have
shown that autophagy is one of the most important pathogenic events
in NSCLC development, leading to drug resistance, metastasis and
poor prognosis (10).
CircRNAs modulate autophagy via
sponging miRNAs in NSCLC
Circular homeodomain-interacting protein kinase 3
(circHIPK3) is derived from exon 2 of the HIPK3 gene, and circHIPK3
is primarily localized in the cytoplasm. Through an RFP-GFP-LC3B
assay, Chen et al (77)
reported that knockdown of circHIPK3 elevated autophagic flux in
autophagy-induced cell lines (A549 and H838). CircHIPK3 may sponge
miR-124-3p, a well-known tumor suppressor and autophagy regulator,
and therefore indirectly regulate IL-6 receptor and STAT3. As a
downstream factor of IL6R, STAT3 suppresses autophagy (78). Downregulation of circHIPK3 induces
autophagy by modulation of miR-124-3p/STAT3/PRKAA/AMPKα signaling
in NSCLC (77).
CircRNAs and NSCLC apoptosis
The unique morphology of cell death was first termed
apoptosis by Kerr in 1972. Apoptosis is the process of programmed
cell death which serves a crucial role in cell biology and life.
The regulation of apoptosis must be strictly controlled (79). The imbalance in the expression ratio
of pro-apoptotic proteins and anti-apoptotic proteins, such as the
Bcl-2 protein family, p53 or inhibitor of apoptosis proteins are
crucial for regulating cell death (80). The Caspases are generally divided into
two groups. Caspase-1, 4, 5, 13 and 14, which primarily participate
in the inflammatory process, and Caspase-2, 3, 6, 7, 8, 9 and 10,
that either participate in initiation or execution of cell death
(81). An increasing number of
studies have illustrated that circRNAs regulate apoptosis in NSCLC,
such as circ_0003645 (82) and
circ_0074027 (83).
CircRNAs modulate apoptosis by
sponging miRNAs in NSCLC
CircRNAs can affect the expression of
apoptosis-related proteins by sponging miRNAs. The activation,
expression and regulation of a series of proteins including the
Caspase family of proteins, Bax and Bcl-2 family of proteins are
involved in apoptosis. Circular VANGL planar cell polarity protein
1 (CircVANGL1) is generated from exons 3–4 of the VANGL1 gene.
CircVANGL1 was reported as an oncogene in bladder cancer (84). Additionally, circVANGL1 was shown to
reduce cell apoptotic rates in NSCLC. Silencing of circVANGL1
increased Bax expression and decreased Bcl-2 expression, and this
effect was achieved by sponging of miR-195 in NSCLC (85).
Circular PVT1 (circPVT1) is generated from exon 3 of
its host gene PVT1, and is flanked by two long introns (35,269 and
41,466 bp) on each side. In a total of 68 cases of NSCLC, the
expression of circPVT1 was >2× higher than that in normal or
paired paratumoral tissues (41/68 cases). Additionally, circPVT1
expression was significantly increased in 7 NSCLC cell lines
compared with a human bronchial epithelial cell line (HBE cells). A
luciferase assay showed that luciferase activity was promoted by
c-Fos interacting with the circPVT1 promoter region. This results
in upregulation of circPVT1 in NSCLC. C-Fos-induced circPVT1
modulates cell proliferation, invasion and migration, and induces
cell apoptosis in NSCLC. CircPVT1 regulates carcinogenesis by
downregulating miR-125b and upregulating E2F transcription factor 2
(E2F2) (86).
CircRNAs and NSCLC EMT, invasion and
metastasis
Tumor metastasis refers to the process in which
malignant tumor cells infiltrate into the surrounding tissues from
their origin. The progression of tumor cell metastasis is divided
into three stages: Adhesion, degradation and migration. Malignant
tumor cells break through the basement membrane, move from its
primary site (primary tumor) into lymphatic vessels, blood vessels
or body cavities to ‘target’ tissues or organs, and form a distant
secondary tumor with the same/similar histological type to that of
the primary tumor (87). The EMT
program is considered a key step and is closely involved in
pathological states of tumor progression (88). EMT is considered as the driving factor
of invasion and metastasis (89).
Metastasis of NSCLC cells is a significant obstacle reducing the OS
of NSCLC patients, and is considered a core step in the malignant
progression of NSCLC (87). Thus, it
is crucial that we improve our understanding of the mechanisms
underlying metastasis. Moreover, it is widely accepted that
circRNAs are related to the invasion and metastasis of NSCLC.
CircRNAs modulate EMT, invasion and
metastasis through sponging miRNAs in NSCLC
Circular SRY-box transcription factor 4 (circ-SOX4)
possesses a covalently closed cyclic structure, and has been shown
to be upregulated in NSCLC. Reduced expression of Circ-SOX4
decreases the number of invasive and metastatic cells, and
decreases the expression of the EMT related proteins, including
N-cadherin, Vimentin, ZEB1, Slug, Twist, Snail, matrix
metalloproteinase (MMP)2, MMP7 and MMP9. Overexpression of
circ-SOX4 increases β-catenin expression in the cell nucleus and
reduces its expression in the cell cytoplasm (increases
translocation). Thus, circ-SOX4 results in activation of the Wnt
pathway. c-MYC is upregulated upon Wnt pathway stimulation.
Furthermore, c-MYC harbors two binding sites with circ-SOX4, and
can increase its expression, highlighting the presence of a
positive feedback loop between circ-SOX4 and c-MYC. Invasion,
metastasis and EMT of NSCLC is promoted by circ-SOX4 via increased
activity of the Wnt/β-catenin pathway, through increasing the
expression of c-MYC (90). Gao and Ye
(91) showed that circ-SOX4 was
upregulated in LUAD Western blotting showed that expression of Wnt
pathway related proteins and EMT representative proteins was
increased when circ-SOX4 was overexpressed. These effects were
altered via upregulated expression of miR-1270 and decreased
expression of PLAG1 like zinc finger 2.
Three circRNAs, hsa_circ_0005273, hsa_circ_0008305,
and hsa_circ_0003221, which are spliced from different exons of the
pre-mRNA of PTK2, are all termed circular protein tyrosine kinase 2
(CircPTK2). In NSCLC, hsa_circ_0008305 expression is low in
patients with distant metastasis. Hsa_circ_0008305 interacts with
miR-429 and miR-200b-3p in NSCLC. The expression of tripartite
motif containing 33 (TIF1γ) is reduced following overexpression of
miR-429 and miR-200b-3p, as they can bind to the 3′-untralsted
region (UTR) of TIF1γ. TIF1γ may mediate EMT and the TGF-β/Smad
pathway by ubiquitinating Smad4. CircPTK2 participates in
TGF-β-induced EMT and invasion. Mechanically, circPTK2 represses
miR-429/miR-200b-3p expression and increases TIF1γ expression
(92).
The process of tumor cell metastasis requires
destruction of any physical barriers, such as the basement membrane
and the extracellular matrix (ECM). MMPs are important proteases
that degrade the ECM, and they serve an important role in tumor
invasion and metastasis (93).
Circular cysteine rich transmembrane BMP regulator 1 (circCRIM1),
also known as hsa_circ_0002346, is an exon-related circRNA. LUAC
patients with TNM stage II and III and lymph node metastasis
exhibited lower circCRIM1 expression levels than patients with TNM
stage I LUAD. Functional experiments revealed that circCRIM1
represses invasion and metastasis in LADC. CircCRIM1 serves as a
miR-182/93 sponge. Resulting in upregulation of leukemia inhibitory
factor receptor and increases MMP13 expression via activation of
the PI3K/AKT/JAK1 signaling pathway (94).
CircRNAs modulate EMT, invasion and
metastasis through binding with RBPs in NSCLC
Circular La ribonucleoprotein 4 (circLARP4), derived
from the LARP4 gene, acts as a La-related RBP. In NSCLC,
downregulated expression of circLARP4 is associated with a worse
prognosis. Overexpression of circLARP4 suppresses the metastatic
ability of SPCA1 cells. Additionally, the protein levels of the
SMAD family member 7 (SMAD7) is upregulated following circLARP4
overexpression. Thus, circLARP4 negatively regulates invasion and
metastasis of NSCLC by upregulating of SMAD7 (95).
CircRNAs modulate EMT, invasion and
metastasis through sponging miRNAs and regulating expression of
their parental genes
Dysregulation of the Wnt/β-catenin signaling pathway
modulates EMT progression in a range of cancer types. CircRNAs
modulate the Wnt/β-catenin pathway through different mechanisms.
Circ-ITCH is located on chromosome 20q11.22. Circ-ITCH expression
is significantly decreased in lung cancer tissues, and its
expression is positively correlated with its parental gene, ITCH.
Circ-ITCH inhibits the activity of the Wnt/β-catenin pathway.
Western blot analysis showed that overexpression of circ-ITCH
suppressed the protein expression levels of β-catenin.
Subsequently, mRNA expression of c-Myc and CCND1, the two
downstream binding partners of β-catenin, was reduced following
circ-ITCH upregulation/overexpression. Thus, circ-ITCH interacts
with miR-7 and miR-214, and participates in the progression of lung
cancer (96).
CircRNAs and NSCLC drug
resistance
Chemotherapy is a common method for treating cancer,
including NSCLC. It has been found that cells that are resistant to
certain chemotherapeutic drugs may also possess resistance to other
structurally unrelated drugs via different mechanisms. This
phenomenon of broad drug resistance is termed MDR (97). Whilst certain factors in tumor cells
underlying the development of resistance to chemoradiation and
targeted therapy have been characterized, the process and the
underlying molecular mechanisms are still not completely
understood. Recent studies have described the roles of circRNAs in
drug resistant NSCLC (98,99).
CircRNAs modulate drug resistance
through sponging miRNAs in NSCLC
One circRNA can interact with multiple miRNAs to
moderate the nucleotide excision repair (NER) signaling pathway.
Hsa_circ_0001946 is an exon-derived circRNA that is produced from
CDR1 with a length of 1,485 nt. Hsa_circ_0001946 is located in
chrX: 139865339-139866824. FISH analysis indicated that
hsa_circ_0001946 is primarily present in the cytoplasm.
Hsa_circ_0001946 functions as a tumor suppressor in NSCLC.
Upregulation of hsa_circ_0001946 enhances the cisplatin sensitivity
of A549 cells. Moreover, silencing of hsa_circ_0001946 activates
the NER signaling pathway, which decreased cisplatin sensitivity of
lung cancer. Hsa_circ_0001946 is implicated in regulation of the
sensitivity of NSCLC cells to cisplatin via modulation of the NER
signaling pathway. Hsa_circ_0001946 sponges four miRNAs
(hsa-miR-7-5p, hsa-miR-671-5p hsa-miR-1270 and hsa-miR-3156-5p) to
moderate NER signaling (100).
Circ_0002483 is located at chr8:141862969-141921766.
Circ_0002483 is significantly downregulated in NSCLC tissue samples
and in Taxol-resistant NSCLC cell lines. Lower levels of
circ_0002483 is correlated with a poorer prognosis in patients with
NSCLC. A Cell Counting Kit-8 assay showed that overexpression of
circ_0002483 notably increased the sensitivity of NSCLC cells to
Taxol. Using dual-luciferase reporter assays and an RNA
immunoprecipitation assay, circ_0002483 was confirmed to
competitively bind to miR-182-5p. Knockdown of miR-182-5p increases
sensitivity to Taxol in A549 and H1299 cells. A luciferase assay
indicated that miR-182-5p could bind to the 3′UTR of growth factor
receptor bound protein 2 (GRB2), forkhead box O1 (FOXO1) and
forkhead box O3 (FOXO3). Co-transfection of miR-182-5p and
circ_0002483 restored GRB2, FOXO1 and FOXO3 expression and induced
resistance to Taxol in NSCLC cells. These findings suggest that
circ_0002483, serves as a miR-182-5p sponge, promotes GRB2, FOXO
and FOXO3 expression and enhances the sensitivity of A549 and H1299
cells to the chemotherapeutic drug Taxol (101).
CircRNAs modulate drug resistance
through binding with RBPs in NSCLC
CircRNA_103762 is significantly highly expressed in
NSCLC tissues and cell lines, and its upregulated expression is
closely correlated with shorter survival rates in patients with
NSCLC. CircRNA_103762 is also upregulated in cisplatin-resistant
H358/CDDP lung cancer cells. CircRNA_103762 represses the
expression of DNA damage inducible transcript 3 and facilitates MDR
in NSCLC (102).
CircRNAs modulate drug resistance
through regulation of translation of their parental genes
Hsa_circ_0004350 and hsa_circ_0092857 are
transcribed from eukaryotic translation initiation factor 3 subunit
A (EIF3a). Hsa_circ_0004350 is located on chromosome
10:120.832.401–120.833.449 and hsa_circ_0092857 on chromosome
10:120.809.312-120.810.833, including three exons and two introns,
and are differentially expressed in A549 and A549/DDP cells.
Hsa_circ_0004350 and hsa_circ_0092857 are prominently associated
with translation regulation based on analysis of data obtained from
Metascape. Gene Ontology analysis showed that the overlapping RBPs
of the two circEIF3as were regulators of translation, and they may
exhibit functional synergy with their parental gene, EIF3a.
Abnormal expression of hsa_circ_0004350 and hsa_circ_0092857 may
impact the cisplatin resistance of lung cancer cells (103).
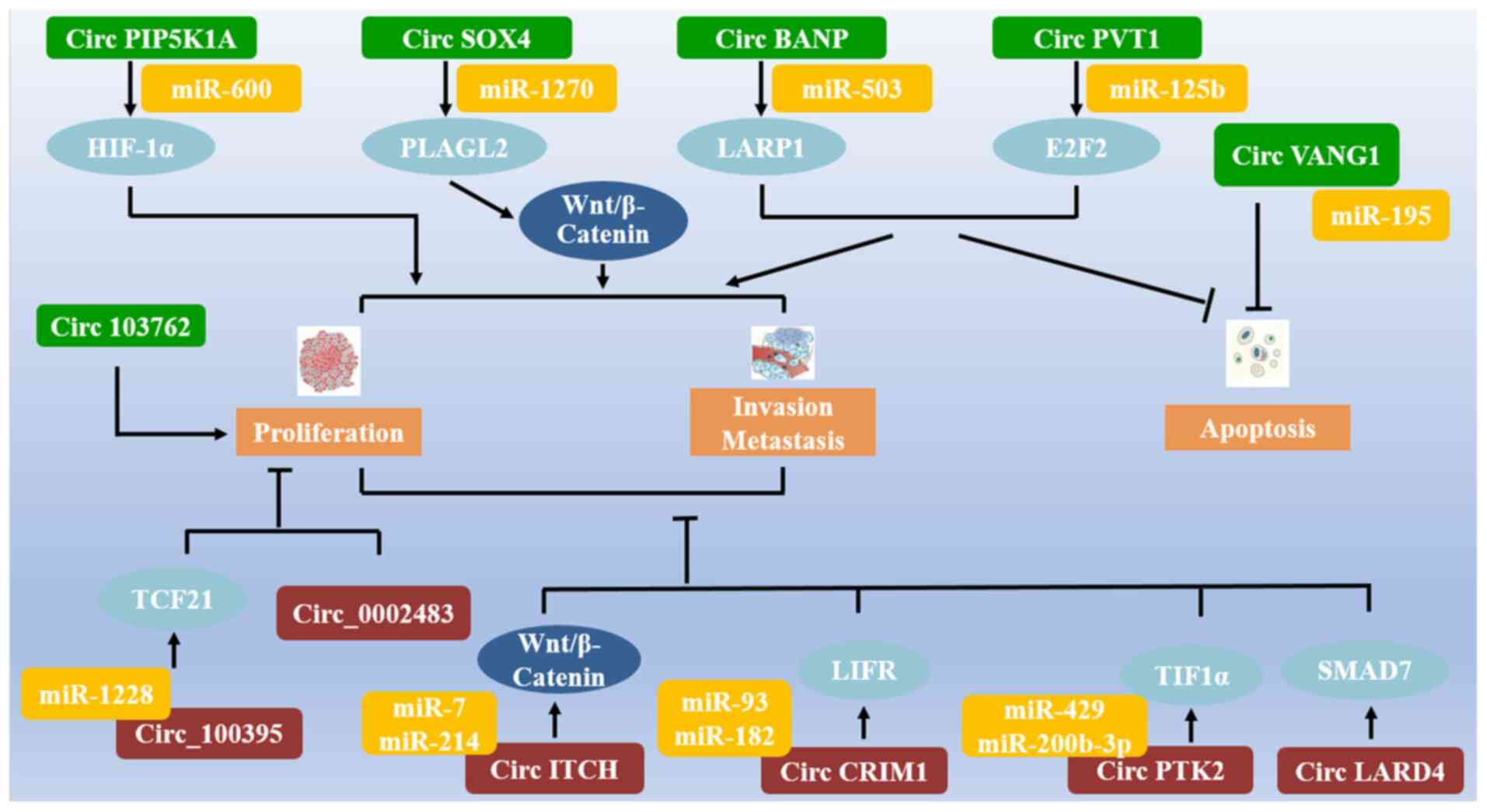
Conclusion
This review summarizes the findings of recent
studies on circRNAs that may function as carcinogenic or tumor
suppressor genes in NSCLC (Fig. 4).
Several circRNAs participate in regulating the pathological
progression of NSCLC. Compared with coding RNAs, miRNAs and
lncRNAs, circRNA research is in its initial stages, and several
problems still need to be addressed. To date several functions of
circRNAs and their participation in the regulation of the
progression of cancer have been identified, although considerably
more remain undetermined. For NSCLC, in order to improve the
prognosis and OS of patients, novel targeted therapeutic approaches
are required. Further development of targeted circRNAs may become
potential pivotal elements to improve our understanding of NSCLC.
These mentioned circRNAs may serve as biomarkers of diagnosis and
prediction in NSCLC. They may also serve as a means of non-invasive
treatments. In future studies, additional attention should be paid
to the role of circRNAs in the clinical diagnosis and treatment of
NSCLC.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 81972522),
Youth Talent Support Program of Liaoning Province (grant no.
XLYC1907011), Key R&D Program of Liaoning Province (grant no.
2018225014) and Technological innovation fund of Shenyang
Technology Division (grant nos. RC190008 and 19-112-4-023).
Availability of data and materials
Not applicable.
Authors' contributions
SL, YL, GQ, YL, XL, FM, NL and TX wrote the original
manuscript. YW, BQ and SX reviewed and edited the manuscript. All
authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
NSCLC
|
non-small cell lung cancer
|
|
circRNAs
|
circular RNAs
|
|
miRNA
|
microRNA
|
|
TNM
|
tumor node metastasis
|
|
EMT
|
epithelial-mesenchymal transition
|
|
SCLC
|
small cell lung cancer
|
|
ceRNA
|
competing endogenous RNA
|
|
ecircRNAs
|
exonic circRNAs
|
|
ciRNAs
|
intronic circRNAs
|
|
EIciRNAs
|
exon-intron circRNA
|
|
tricRNAs
|
tRNA intronic circRNAs
|
|
MRE
|
microRNA response element
|
References
|
1
|
Hirsch FR, Scagliotti GV, Mulshine JL,
Kwon R, Curran WJ Jr, Wu YL and Paz-Ares L: Lung cancer: Current
therapies and new targeted treatments. Lancet. 389:299–311. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tsim S, O'Dowd CA, Milroy R and Davidson
S: Staging of non-small cell lung cancer (NSCLC): A review. Respir
Med. 104:1767–1674. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Giovannetti E, Toffalorio F, De Pas T and
Peters GJ: Pharmacogenetics of conventional chemotherapy in
non-small-cell lung cancer: A changing landscape? Pharmacogenomics.
13:1073–1086. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yu Y and He J: Molecular classification of
non-small-cell lung cancer: Diagnosis, individualized treatment,
and prognosis. Front Med. 7:157–171. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Minguet J, Smith KH and Bramlage P:
Targeted therapies for treatment of non-small cell lung
cancer-recent advances and future perspectives. Int J Cancer.
138:2549–2561. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rotow J and Bivona TG: Understanding and
targeting resistance mechanisms in NSCLC. Nat Rev Cancer.
17:637–658. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Herbst RS, Morgensztern D and Boshoff C:
The biology and management of non-small cell lung cancer. Nature.
553:446–454. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hallstrom TC and Nevins JR: Balancing the
decision of cell proliferation and cell fate. Cell Cycle.
8:532–535. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Choi AM, Ryter SW and Levine B: Autophagy
in human health and disease. N Engl J Med. 368:651–662. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Elmore S: Apoptosis: A review of
programmed cell death. Toxicol Pathol. 35:495–516. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu G, Pei F, Yang F, Li L, Amin AD, Liu
S, Buchan JR and Cho WC: Role of autophagy and apoptosis in
non-small-cell lung cancer. Int J Mol Sci. 18:3672017. View Article : Google Scholar
|
|
13
|
Perlikos F, Harrington KJ and Syrigos KN:
Key molecular mechanisms in lung cancer invasion and metastasis: A
comprehensive review. Crit Rev Oncol Hematol. 87:1–11. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Meng S, Zhou H, Feng Z, Xu Z, Tang Y, Li P
and Wu M: CircRNA: Functions and properties of a novel potential
biomarker for cancer. Mol Cancer. 16:942017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Petkovic S and Müller S: RNA
circularization strategies in vivo and in vitro. Nucleic Acids Res.
43:2454–2465. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Dong Y, He D, Peng Z, Peng W, Shi W, Wang
J, Li B, Zhang C and Duan C: Circular RNAs in cancer: An emerging
key player. J Hematol Oncol. 10:22017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Y, Liu J, Ma J, Sun T, Zhou Q, Wang
W, Wang G, Wu P, Wang H, Jiang L, et al: Exosomal circRNAs:
Biogenesis, effect and application in human diseases. Mol Cancer.
18:1162019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li T, Shao Y, Fu L, Xie Y, Zhu L, Sun W,
Yu R, Xiao B and Guo J: Plasma circular RNA profiling of patients
with gastric cancer and their droplet digital RT-PCR detection. J
Mol Med (Berl). 96:85–96. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Geng Y, Jiang J and Wu C: Function and
clinical significance of circRNAs in solid tumors. J Hematol Oncol.
11:982018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhang M and Xin Y: Circular RNAs: A new
frontier for cancer diagnosis and therapy. J Hematol Oncol.
11:212018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Z, Yang T and Xiao J: Circular RNAs:
Promising biomarkers for human diseases. EBioMedicine. 34:267–274.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li X, Yang L and Chen LL: The biogenesis,
functions, and challenges of circular RNAs. Mol Cell. 71:428–442.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bezzi M, Guarnerio J and Pandolfi PP: A
circular twist on microRNA regulation. Cell Res. 27:1401–1402.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang H, Cai K, Wang J, Wang X, Cheng K,
Shi F, Jiang L, Zhang Y and Dou J: MiR-7, inhibited indirectly by
lincRNA HOTAIR, directly inhibits SETDB1 and reverses the EMT of
breast cancer stem cells by downregulating the STAT3 pathway. Stem
Cells. 32:2858–2868. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu S, Zhang P, Chen Z, Liu M, Li X and
Tang H: MicroRNA-7 downregulates XIAP expression to suppress cell
growth and promote apoptosis in cervical cancer cells. FEBS Lett.
587:2247–2253. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kong D, Piao YS, Yamashita S, Oshima H,
Oguma K, Fushida S, Fujimura T, Minamoto T, Seno H, Yamada Y, et
al: Inflammation-induced repression of tumor suppressor miR-7 in
gastric tumor cells. Oncogene. 31:3949–3960. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fang Y, Xue JL, Shen Q, Chen J and Tian L:
MicroRNA-7 inhibits tumor growth and metastasis by targeting the
phosphoinositide 3-kinase/Akt pathway in hepatocellular carcinoma.
Hepatology. 55:1852–1862. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Sang M, Meng L, Sang Y, Liu S, Ding P, Ju
Y, Liu F, Gu L, Lian Y, Li J, et al: Circular RNA ciRS-7
accelerates ESCC progression through acting as a miR-876-5p sponge
to enhance MAGE-A family expression. Cancer Lett. 426:37–46. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu C, Yao MD, Li CP, Shan K, Yang H, Wang
JJ, Liu B, Li XM, Yao J, Jiang Q and Yan B: Silencing of circular
RNA-ZNF609 ameliorates vascular endothelial dysfunction.
Theranostics. 7:2863–2877. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Conn SJ, Pillman KA, Toubia J, Conn VM,
Salmanidis M, Phillips CA, Roslan S, Schreiber AW, Gregory PA and
Goodall GJ: The RNA binding protein quaking regulates formation of
circRNAs. Cell. 160:1125–1134. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Du WW, Yang W, Liu E, Yang Z, Dhaliwal P
and Yang BB: Foxo3 circular RNA retards cell cycle progression via
forming ternary complexes with p21 and CDK2. Nucleic Acids Res.
44:2846–2858. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Du WW, Fang L, Yang W, Wu N, Awan FM, Yang
Z and Yang BB: Induction of tumor apoptosis through a circular RNA
enhancing Foxo3 activity. Cell Death Differ. 24:357–370. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang Y and Wang Z: Efficient backsplicing
produces translatable circular mRNAs. RNA. 21:172–179. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen CY and Sarnow P: Initiation of
protein synthesis by the eukaryotic translational apparatus on
circular RNAs. Science. 268:415–417. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Perriman R and Ares M Jr: Circular mRNA
can direct translation of extremely long repeating-sequence
proteins in vivo. RNA. 4:1047–1054. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Abe N, Matsumoto K, Nishihara M, Nakano Y,
Shibata A, Maruyama H, Shuto S, Matsuda A, Yoshida M, Ito Y and Abe
H: Rolling circle translation of circular RNA in living human
cells. Sci Rep. 5:164352015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Legnini I, Di Timoteo G, Rossi F, Morlando
M, Briganti F, Sthandier O, Fatica A, Santini T, Andronache A, Wade
M, et al: Circ-ZNF609 is a circular RNA that can be translated and
functions in myogenesis. Mol Cell. 66:22–37.e9. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang Y, Gao X, Zhang M, Yan S, Sun C, Xiao
F, Huang N, Yang X, Zhao K, Zhou H, et al: Novel role of FBXW7
circular RNA in repressing glioma tumorigenesis. J Natl Cancer
Inst. 110:304–315. 2018. View Article : Google Scholar
|
|
40
|
Zhang M, Huang N, Yang X, Luo J, Yan S,
Xiao F, Chen W, Gao X, Zhao K, Zhou H, et al: A novel protein
encoded by the circular form of the SHPRH gene suppresses glioma
tumorigenesis. Oncogene. 37:1805–1814. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Westholm JO, Miura P, Olson S, Shenker S,
Joseph B, Sanfilippo P, Celniker SE, Graveley BR and Lai EC:
Genome-wide analysis of drosophila circular RNAs reveals their
structural and sequence properties and age-dependent neural
accumulation. Cell Rep. 9:1966–1980. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Holdt LM, Stahringer A, Sass K, Pichler G,
Kulak NA, Wilfert W, Kohlmaier A, Herbst A, Northoff BH, Nicolaou
A, et al: Circular non-coding RNA ANRIL modulates ribosomal RNA
maturation and atherosclerosis in humans. Nat Commun. 7:124292016.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Li Z, Huang C, Bao C, Chen L, Lin M, Wang
X, Zhong G, Yu B, Hu W, Dai L, et al: Exon-intron circular RNAs
regulate transcription in the nucleus. Nat Struct Mol Biol.
22:256–264. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Salzman J, Gawad C, Wang PL, Lacayo N and
Brown PO: Circular RNAs are the predominant transcript isoform from
hundreds of human genes in diverse cell types. PLoS One.
7:e307332012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin
QF, Xing YH, Zhu S, Yang L and Chen LL: Circular intronic long
noncoding RNAs. Mol Cell. 51:792–806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Harrison PM, Zheng D, Zhang Z, Carriero N
and Gerstein M: Transcribed processed pseudogenes in the human
genome: An intermediate form of expressed retrosequence lacking
protein-coding ability. Nucleic Acids Res. 33:2374–2383. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kalyana-Sundaram S, Kumar-Sinha C, Shankar
S, Robinson DR, Wu YM, Cao X, Asangani IA, Kothari V, Prensner JR,
Lonigro RJ, et al: Expressed pseudogenes in the transcriptional
landscape of human cancers. Cell. 149:1622–1634. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhang Z, Harrison PM, Liu Y and Gerstein
M: Millions of years of evolution preserved: A comprehensive
catalog of the processed pseudogenes in the human genome. Genome
Res. 13:2541–2558. 2015. View Article : Google Scholar
|
|
49
|
Dong R, Zhang XO, Zhang Y, Ma XK, Chen LL
and Yang L: CircRNA-derived pseudogenes. Cell Res. 26:747–750.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J,
Chen D, Gu J, He X and Huang S: Circular RNA is enriched and stable
in exosomes: A promising biomarker for cancer diagnosis. Cell Res.
25:981–984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Lei B, Tian Z, Fan W and Ni B: Circular
RNA: A novel biomarker and therapeutic target for human cancers.
Int J Med Sci. 16:292–301. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li Z, Ruan Y, Zhang H, Shen Y, Li T and
Xiao B: Tumor-suppressive circular RNAs: Mechanisms underlying
their suppression of tumor occurrence and use as therapeutic
targets. Cancer Sci. 110:3630–3638. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Li Y, Ge YZ, Xu L and Jia R: Circular RNA
ITCH: A novel tumor suppressor in multiple cancers. Life Sci.
254:1171762020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Jiang MM, Mai ZT, Wan SZ, Chi YM, Zhang X,
Sun BH and Di QG: Microarray profiles reveal that circular RNA
hsa_circ_0007385 functions as an oncogene in non-small cell lung
cancer tumorigenesis. J Cancer Res Clin Oncol. 144:667–674. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li C, Zhang L, Meng G, Wang Q, Lv X, Zhang
J and Li J: Circular RNAs: Pivotal molecular regulators and novel
diagnostic and prognostic biomarkers in non-small cell lung cancer.
J Cancer Res Clin Oncol. 145:2875–2889. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Mu Y, Xie F, Huang Y, Yang D, Xu G, Wang C
and Wu Q: Circular RNA expression profile in peripheral whole blood
of lung adenocarcinoma by high: Throughput sequencing. Medicine
(Baltimore). 98:e176012019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen LL and Yang L: Regulation of circRNA
biogenesis. RNA Biol. 12:381–388. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Zhang S, Zeng X, Ding T, Guo L, Li Y, Ou S
and Yuan H: Microarray profile of circular RNAs identifies
hsa_circ_0014130 as a new circular RNA biomarker in non-small cell
lung cancer. Sci Rep. 8:28782018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chen L, Nan A, Zhang N, Jia Y, Li X, Ling
Y, Dai J, Zhang S, Yang Q, Yi Y and Jiang Y: Circular RNA 100146
functions as an oncogene through direct binding to miR-361-3p and
miR-615-5p in non-small cell lung cancer. Mol Cancer. 18:132019.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Qiu M, Xia W, Chen R, Wang S, Xu Y, Ma Z,
Xu W, Zhang E, Wang J, Fang T, et al: The circular RNA circPRKCI
promotes tumor growth in lung adenocarcinoma. Cancer Res.
78:2839–2851. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Hsiao KY, Lin YC, Gupta SK, Chang N, Yen
L, Sun HS and Tsai SJ: Noncoding effects of circular RNA CCDC66
promote colon cancer growth and metastasis. Cancer Res.
77:2339–2350. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Joseph NA, Chiou SH, Lung Z, Yang CL, Lin
TY, Chang HW, Sun HS, Gupta SK, Yen L, Wang SD, et al: The role of
HGF-MET pathway and CCDC66 cirRNA expression in EGFR resistance and
epithelial-to-mesenchymal transition of lung adenocarcinoma cells.
J Hematol Oncol. 11:742018. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Chen D, Ma W, Ke Z and Xie F: CircRNA
hsa_circ_100395 regulates miR-1228/TCF21 pathway to inhibit lung
cancer progression. Cell Cycle. 17:2080–2090. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Han J, Zhao G, Ma X, Dong Q, Zhang H, Wang
Y and Cui J: CircRNA circ-BANP-mediated miR-503/LARP1 signaling
contributes to lung cancer progression. Biochem Biophys Res Commun.
503:2429–2435. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Pack LR, Daigh LH and Meyer T: Putting the
brakes on the cell cycle: Mechanisms of cellular growth arrest.
Curr Opin Cell Biol. 60:106–113. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Juríková M, Danihel Ľ, Polák Š and Varga
I: Ki67, PCNA, and MCM proteins: Markers of proliferation in the
diagnosis of breast cancer. Acta Histochem. 118:544–552. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhao F, Han Y, Liu Z, Zhao Z, Li Z and Jia
K: circFADS2 regulates lung cancer cells proliferation and invasion
via acting as a sponge of miR-498. Biosci Rep. 38:BSR201805702018.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Zhang X, Yang D and Wei Y: Overexpressed
CDR1as functions as an oncogene to promote the tumor progression
via miR-7 in non-small-cell lung cancer. Onco Targets Ther.
11:3979–3987. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Zhu XF, Liu ZC, Xie BF, Li ZM, Feng GK,
Yang D and Zeng YX: EGFR tyrosine kinase inhibitor AG1478 inhibits
cell proliferation and arrests cell cycle in nasopharyngeal
carcinoma cells. Cancer Lett. 169:27–32. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Nakayama K, Rahman MT, Rahman M, Nakamura
K, Ishikawa M, Katagiri H, Sato E, Ishibashi T, Iida K, Ishikawa N
and Kyo S: CCNE1 amplification is associated with aggressive
potential in endometrioid endometrial carcinomas. Int J Oncol.
48:506–516. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Su C, Han Y, Zhang H, Li Y, Yi L, Wang X,
Zhou S, Yu D, Song X, Xiao N, et al: CiRS-7 targeting miR-7
modulates the progression of non-small cell lung cancer in a manner
dependent on NF-κB signalling. J Cell Mol Med. 22:3097–3107. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhu X, Wang X, Wei S, Chen Y, Chen Y, Fan
X, Han S and Wu G: hsa_circ_0013958: A circular RNA and potential
novel biomarker for lung adenocarcinoma. FEBS J. 284:2170–2182.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Donnellan R and Chetty R: Cyclin D1 and
human neoplasia. Mol Pathol. 51:1–7. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Chi Y, Luo Q, Song Y, Yang F, Wang Y, Jin
M and Zhang D: Circular RNA circPIP5K1A promotes non-small cell
lung cancer proliferation and metastasis through miR-600/HIF-1α
regulation. J Cell Biochem. 120:19019–19030. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Nan A, Chen L, Zhang N, Jia Y, Li X, Zhou
H, Ling Y, Wang Z, Yang C, Liu S and Jiang Y: Circular RNA
circNOL10 inhibits lung cancer development by promoting
SCLM1-mediated transcriptional regulation of the humanin
polypeptide family. Adv Sci (Weinh). 6:18006542018. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Qin M, Wei G and Sun X: Circ-UBR5: An
exonic circular RNA and novel small nuclear RNA involved in RNA
splicing. Biochem Biophys Res Commun. 503:1027–1034. 2019.
View Article : Google Scholar
|
|
77
|
Chen X, Mao R, Su W, Yang X, Geng Q, Guo
C, Wang Z, Wang J, Kresty LA, Beer DG, et al: Circular RNA
circHIPK3 modulates autophagy via MIR124-3p-STAT3-PRKAA/AMPKα
signaling in STK11 mutant lung cancer. Autophagy. 16:659–671. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
You L, Wang Z, Li H, Shou J, Jing Z, Xie
J, Sui X, Pan H and Han W: The role of STAT3 in autophagy.
Autophagy. 11:729–739. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Espinosa-Oliva AM, García-Revilla J,
Alonso-Bellido IM and Burguillos MA: Brainiac caspases: Beyond the
wall of apoptosis. Front Cell Neurosci. 13:5002019. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Pistritto G, Trisciuoglio D, Ceci C,
Garufi A and D'Orazi G: Apoptosis as anticancer mechanism: Function
and dysfunction of its modulators and targeted therapeutic
strategies. Aging (Albany NY). 8:603–619. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Wong RS: Apoptosis in cancer: From
pathogenesis to treatment. J Exp Clin Cancer Res. 30:872011.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
An J, Shi H, Zhang N and Song S: Elevation
of circular RNA circ_0003645 forecasts unfavorable prognosis and
facilitates cell progression via miR-1179/TMEM14A pathway in
non-small cell lung cancer. Biochem Biophys Res Commun.
511:921–925. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Gao P, Wang Z, Hu Z, Jiao X and Yao Y:
Circular RNA circ_0074027 indicates a poor prognosis for NSCLC
patients and modulates cell proliferation, apoptosis, and invasion
via miR-185-3p mediated BRD4/MADD activation. J Cell Biochem.
121:2632–2642. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Zeng Z, Zhou W, Duan L, Zhang J, Lu X, Jin
L and Yu Y: Circular RNA circ-VANGL1 as a competing endogenous RNA
contributes to bladder cancer progression by regulating
miR-605-3p/VANGL1 pathway. J Cell Physiol. 234:3887–3896. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wang L, Ma H, Kong W, Liu B and Zhang X:
Up-regulated circular RNA VANGL1 contributes to progression of
non-small cell lung cancer through inhibition of miR-195 and
activation of Bcl-2. Biosci Rep. 39:BSR201824332019. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Li X, Zhang Z, Jiang H, Li Q, Wang R, Pan
H, Niu Y, Liu F, Gu H, Fan X and Gao J: Circular RNA circPVT1
promotes proliferation and invasion through sponging miR-125b and
activating E2F2 signaling in non-small cell lung cancer. Cell
Physiol Biochem. 51:2324–2340. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Kramer N, Walzl A, Unger C, Rosner M,
Krupitza G, Hengstschläger M and Dolznig H: In vitro cell migration
and invasion assays. Mutat Res. 752:10–24. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Goossens S, Vandamme N, Van Vlierberghe P
and Berx G: EMT transcription factors in cancer development
re-evaluated: Beyond EMT and MET. Biochim Biophys Acta Rev Cancer.
1868:584–591. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Diepenbruck M and Christofori G:
Epithelial-mesenchymal transition (EMT) and metastasis: Yes, no,
maybe? Curr Opin Cell Biol. 43:7–13. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Wang L, Zheng C, Wu X, Zhang Y, Yan S,
Ruan L and Dai H: Circ-SOX4 promotes non-small cell lung cancer
progression by activating the Wnt/beta-catenin pathway. Mol Oncol.
2020.
|
|
91
|
Gao N and Ye B: Circ-SOX4 drives the
tumorigenesis and development of lung adenocarcinoma via sponging
miR-1270 and modulating PLAGL2 to activate WNT signaling pathway.
Cancer Cell Int. 20:22020. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Wang L, Tong X, Zhou Z, Wang S, Lei Z,
Zhang T, Liu Z, Zeng Y, Li C, Zhao J, et al: Circular RNA
hsa_circ_0008305 (circPTK2) inhibits TGF-β-induced
epithelial-mesenchymal transition and metastasis by controlling
TIF1γ in non-small cell lung cancer. Mol Cancer. 17:1402018.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Winer A, Adams S and Mignatti P: Matrix
metalloproteinase inhibitors in cancer therapy: Turning past
failures into future successes. Mol Cancer Ther. 17:1147–1155.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Wang L, Liang Y, Mao Q, Xia W, Chen B,
Shen H, Xu L, Jiang F and Dong G: Circular RNA circCRIM1 inhibits
invasion and metastasis in lung adenocarcinoma through the microRNA
(miR)-182/miR-93-leukemia inhibitory factor receptor pathway.
Cancer Sci. 110:2960–2972. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Shi JQ, Wang B, Cao XQ, Wang YX, Cheng X,
Jia CL, Wen T, Luo BJ and Liu ZD: Circular RNA_LARP4 inhibits the
progression of non-small-cell lung cancer by regulating the
expression of SMAD7. Eur Rev Med Pharmacol Sci. 24:1863–1869.
2020.PubMed/NCBI
|
|
96
|
Wan L, Zhang L, Fan K, Cheng ZX, Sun QC
and Wang JJ: Circular RNA-ITCH suppresses lung cancer proliferation
via inhibiting the Wnt/β-catenin pathway. Biomed Res Int.
2016:15794902016. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Zhitomirsky B and Assaraf YG: Lysosomes as
mediators of drug resistance in cancer. Drug Resist Updat.
24:23–33. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Fraipont F, Gazzeri S, Cho WC and Eymin B:
Circular RNAs and RNA splice variants as biomarkers for prognosis
and therapeutic response in the liquid biopsies of lung cancer
patients. Front Genet. 10:3902019. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Dong Y, Xu T, Zhong S, Wang B, Zhang H,
Wang X, Wang P, Li G and Yang S: Circ_0076305 regulates cisplatin
resistance of non-small cell lung cancer via positively modulating
STAT3 by sponging miR-296-5p. Life Sci. 239:1169842019. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Huang MS, Liu JY, Xia XB, Liu YZ, Li X,
Yin JY, Peng JB, Wu L, Zhang W, Zhou HH and Liu ZQ:
Hsa_circ_0001946 inhibits lung cancer progression and mediates
cisplatin sensitivity in non-small cell lung cancer via the
nucleotide excision repair signaling pathway. Front Oncol.
9:5082019. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Li X, Yang B, Ren H, Xiao T, Zhang L, Li
L, Li M, Wang X, Zhou H and Zhang W: Hsa_circ_0002483 inhibited the
progression and enhanced the Taxol sensitivity of non-small cell
lung cancer by targeting miR-182-5p. Cell Death Dis. 10:9532019.
View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Xiao G, Huang W, Zhan Y, Li J and Tong W:
CircRNA_103762 promotes multidrug resistance in NSCLC by targeting
DNA damage inducible transcript 3 (CHOP). J Clin Lab Anal.
34:e232522020. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Huang MS, Yuan FQ, Gao Y, Liu JY, Chen YX,
Wang CJ, He BM, Zhou HH and Liu ZQ: Circular RNA screening from
EIF3a in lung cancer. Cancer Med. 8:4159–4168. 2019. View Article : Google Scholar : PubMed/NCBI
|