Introduction
Thyroid carcinoma (TC) is an endocrine carcinoma
with increasing incidence worldwide in 2018 (1). Papillary TC (PTC) is a differentiated
type of TC that accounts for ~80% of all cases of TC (2). The majority of patients with PTC have a
favorable prognosis with a ~90% overall survival rate due to the
improvements in therapeutic strategies, such as surgical excision,
radiotherapy and hormone suppression (3). Targeted treatments provide novel
insights into the mechanism underlying PTC progression, such as the
use of tyrosine kinase inhibitors (4). Small-molecule MAPK inhibitors increase
the susceptibility of PTC cells to radioiodine (5). Nevertheless, certain patients still
exhibit a poor prognosis due to the presence of distant metastasis.
Thus, developing novel therapeutic targets is essential for
limiting PTC progression.
Circular RNAs (circRNAs) are the most recently
discovered type of non-coding RNA generated by the back-splicing of
precursor mRNAs with covalently closed loop structures (6). These are extensively expressed in
eukaryotes and have been shown to participate in multiple
biological processes, including sponging microRNAs (miRNA/miRs),
interacting with RNA-binding proteins and regulating gene
expression levels (7–9). Previous studies have reported that
circRNAs exhibit tumor promoting or suppressive effects in several
types of cancer, such as hepatocellular carcinoma, glioma and lung
adenocarcinoma (10–12). Additionally, in PTC, the dysregulated
expression of circRNAs is responsible for several adverse
clinicopathological features, such as capsular infiltration,
advanced pathological T stage and lymph node metastasis (13). Liu et al (14) demonstrated that circRap guanine
nucleotide exchange factor 5 (RAPGEF5) serves a tumorigenic role in
PTC via the miR-198/FGFR1 axis. Microarray analysis has shown that
a novel circRNA hsa_circ_0001666 (hsa_circ_000742), derived from
the linear gene family with sequence similarity (FAM)120B, is
upregulated in PTC (15). To the best
of our knowledge, however, the biological functions of
hsa_circ_0001666 in PTC have not been studied.
miRNAs are evolutionarily conserved small non-coding
RNAs that regulate multiple biological processes. Several miRNAs,
such as miR-221, miR-146b-5p and miR-1178, have been reported to
modulate PTC development and progression (16–18). For
example, miR-206 suppresses drug resistance to Euthyrox by
targeting MAP4K3 (19). A recent
study showed that miR-330-5p mediates the positive regulation of
long intergenic non-protein-coding RNA 003311 on cancer stem-like
features in PTC (20). miR-326
suppresses PTC cell proliferation and metastasis by inhibiting
MAPK1 and erb-b2 receptor tyrosine kinase (ERBB) 4 (21).
ETS variant transcription factor 4 (ETV4) serves a
tumor promoting role in non-small-cell lung, breast and pancreatic
cancer (22–24). The present study aimed to discover
potential common binding miRNAs with hsa_circ_0001666 and ETV4.
Emerging evidence indicates that circRNAs may sponge miRNAs to
regulate gene expression levels (14,25).
However, whether hsa_circ_0001666 serves as a miRNA sponge in PTC
remains unknown.
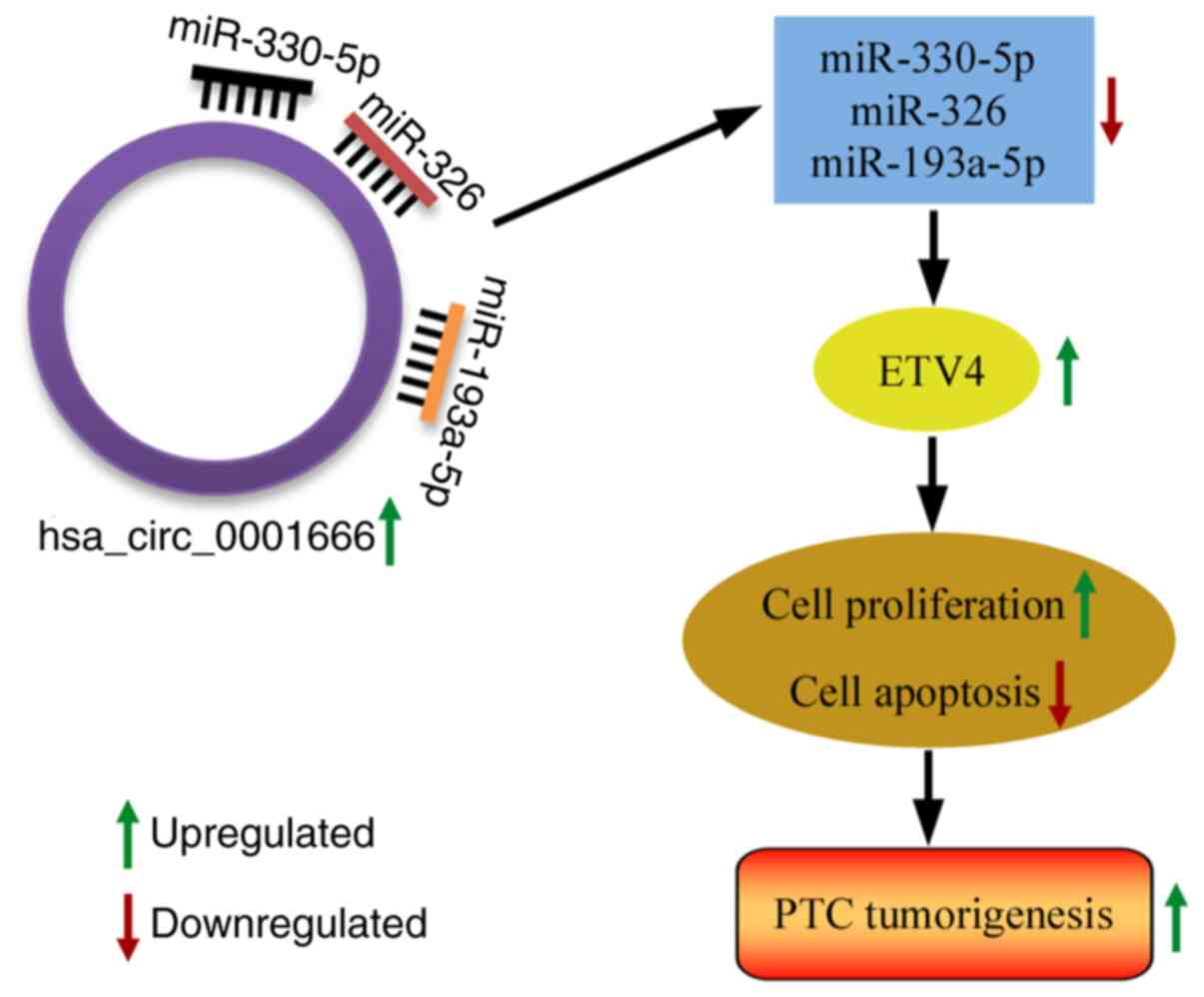
The expression of hsa_circ_0001666 in PTC and its
biological functions were here investigated. A
miR-330-5p/miR-193a-5p/miR-326/ETV4 axis was used to elucidate the
regulatory mechanism of hsa_circ_0001666.
Materials and methods
Clinical samples
Patients with PTC (n=60, female and male; age, 28–71
years) were enrolled at Shengjing Hospital of China Medical
University (Shenyang, China) between April 2019 and June 2020. The
distance between tumor and matching normal adjacent tissue was ~2
cm. Patients who were diagnosed with PTC for the first time and
with no prior treatment history associated with cancer were
included. Patients who received prolonged hormonotherapy or
immunosuppressive therapy were excluded. In addition, patients with
complicated chronic diseases, pregnant or lactating women, and
children were also excluded. All tissues samples were collected and
immediately frozen in liquid nitrogen for further examination. The
present study was approved by the Institutional Review Board of
Shengjing Hospital of China Medical University and performed in
accordance with the Declaration of Helsinki. All patients provided
written informed consent. The collected tumor tissues and adjacent
normal tissues were used to measure hsa_circ_0001666 expression
levels and analyze the associations between hsa_circ_0001666 and
clinicopathological features.
Cell culture
Three human PTC cell lines, TPC-1 (Procell Life
Science & Technology Co., Ltd.), IHH-4 (CoBioer Biosciences
Co., Ltd.) and GLAG-66 (Guangzhou Cellcook Biotech Co., Ltd.), and
human thyroid follicular epithelial cell line Nthy-ori 3-1
(Shanghai Zhongqiaoxinzhou Biotech) were obtained. All cell lines
were STR profiled every year if no other problems were reported.
Mycoplasma testing was also performed every 3 months. TPC-1 and
Nthy-ori 3-1 cells were cultured in RPMI-1640 medium
(Sigma-Aldrich; Merck KGaA). GLAG-66 cells were cultured in DMEM
(Sigma-Aldrich; Merck KGaA). IHH-4 cells were incubated in
DMEM/RPMI-1640 (Sigma-Aldrich; Merck KGaA) mixed medium. All media
were supplemented with 10% FBS (Sigma-Aldrich; Merck KGaA) and the
cells were cultured at 37°C in a humidified incubator with 5%
CO2.
Fluorescence in situ hybridization
(FISH)
FISH staining was performed to determine the
cellular distribution of hsa_circ_0001666 using a FISH kit
(Guangzhou RiboBio Co., Ltd.). TPC-1 cells were seeded in a 24-well
plate (3×104 cells/well) and fixed in 4%
paraformaldehyde at room temperature for 10 min. Then, specific
FISH probes against hsa_circ_0001666 were used to hybridize its
junction site. The nuclei were counterstained with DAPI (0.005
mg/ml) at room temperature for 10 min. 18S probes were used as the
positive control. Images of FISH staining were captured using a
light Olympus microscope (Olympus Corporation) at ×400
magnification.
Reverse transcription (RT)PCR
TPC-1 cells (2×105/well) were plated in
6-well plates. For RNase R treatment, total RNAs were extracted
from TPC-1 cells and incubated in the presence or absence of RNase
R (Guangzhou Jisai Biological Technology Co., Ltd.) at 37°C for 30
min and subsequently reverse-transcribed using Super M-MLV Reverse
Transcriptase kit (BioTeke Corporation) according to the
manufacturer's protocol at 42°C for 50 min. Genomic (g)DNA from
cells was isolated using a genome DNA isolation kit (BioTeke
Corporation) according to the manufacturer's protocol. The
complementary (c)DNA or gDNA was analyzed by RT-PCR using specific
primers (Table SII); PCR products
were separated on a 1.5% agarose gel and analyzed using Sanger
sequencing (Sangon Biotech Co., Ltd.).
Cell transfection
For hsa_circ_0001666 knockdown, five pairs of small
interfering (si)RNAs against hsa_circ_0001666 and corresponding
negative control (NC) siRNA (JTS Scientific Ltd.) were constructed
and transfected into TPC-1 or IHH-4 cells at a final concentration
of 23 nM for 48 h.
For miRNA overexpression, miR-330-5p, miR-193a-5p,
miR-326 or corresponding NC mimics (JTS Scientific Ltd.) were
transfected into PTC cells for 48 h at a concentration of 23
nM.
In addition, miR-330-5p, miR-193a-5p, miR-326 or
corresponding NC inhibitors were acquired from JTS Scientific Ltd.
The ETV4 overexpression plasmid or empty vector was constructed by
GenScript. For the rescue experiments, TPC-1 cells were
co-transfected with hsa_circ_0001666 siRNAs (23 nM) and miRNA
inhibitor (23 nM) or NC inhibitor for 48 h. ETV4 overexpressing
pcDNA3.1 vector or empty pcDNA3.1 vector at a final concentration
of 455 ng/ml together with hsa_circ_0001666 siRNAs (23 nM) were
transfected into TPC-1 cells for 48 h.
All oligonucleotide sequences (Table SI) were transfected into PTC cells
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) following the manufacturer's instruction at room
temperature. All subsequent experiments were performed at 48 h post
transfection. The transfection efficiency for inhibiting gene
expression was ~70% and for overexpressing gene expression was
~900%.
Cell Counting Kit (CCK)8 assay
CCK8 assays were performed to investigate the
viability of cells (Beyotime Institute of Biotechnology). In brief,
TPC-1 or IHH-4 cells were seeded in a 96-well plate
(4×103 cells/well) and incubated with CCK8 solution (10
µl) for 1 h. A microplate reader was used to measure the absorbance
at 450 nm.
5-ethynyl-2′-deoxyuridine (Edu)
staining assay
An Edu assay kit (Nanjing KeyGen Biotech Co., Ltd.)
was utilized to assess cell proliferation. Briefly, transfected
TPC-1 and IHH-4 cells (3×104/well) were collected and
incubated with Edu reagent according to the manufacturer's
protocol. Subsequently, cells were counterstained with Hoechst
33342 (1:2,000) for 15 min at room temperature, harvested and
observed under a light microscope at ×400 magnification.
Flow cytometry
Cells were seeded onto 6-well plates
(2×105 cells/well). After transfection for 48 h, cells
were collected to determine cell cycle distribution or cell
apoptosis. For cell cycle analysis, cells were pre-treated with 70%
cold ethanol at 4°C overnight and incubated with PI solution for 30
min at 4°C in the dark according to the manufacturer's protocol
(Nanjing KeyGen Biotech Co., Ltd.). Cell apoptosis was measured
using Annexin V-FITC/PI apoptosis detection kit (Nanjing KeyGen
Biotech Co., Ltd.). Flow cytometry instrument NovoCyte (ACEA
Bioscience, Inc.) was used to detect the cells and NovoExpress
1.3.4 software (ACEA Bioscience, Inc.) was used to analyze
data.
Luciferase reporter assay
Bioinformatics analysis predicted that
hsa_circ_0001666 was complementary to three miRNAs (miR-330-5p,
miR-193a-5p and miR-326) based on StarBase v3.0 (starbase.sysu.edu.cn/index.php) and
Circular RNA Interactome software (circinteractome.nia.nih.gov/index.html). In addition,
these three miRNAs were predicted to bind with ETV4 3′-UTR based on
StarBase. In order to confirm this association, wild-type and
mutant sequences of the targeting region of hsa_circ_0001666 or
ETV4 were constructed and cloned into a pmirGLO vector (Promega
Corporation). Subsequently, 293T cells were co-transfected with the
wild-type or mutant luciferase reporter plasmids targeting
hsa_circ_0001666 or ETV4, together with miR-330-5p, miR-193a-5p or
miR-326 mimics using Lipofectamine® 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) for 48 h. Binding activity was
measured using a Dual luciferase reporter assay kit (Promega
Corporation), relative to firefly/Renilla luciferase activity.
RNA binding protein
immunoprecipitation (RIP) assay
RIP experiments were performed using a Magna RIP
RNA-Binding Protein Immunoprecipitation kit (EMD Millipore)
according to the manufacturer's protocol. TPC-1 and IHH-4 cell
lysates were incubated with argonaute RISC catalytic component 2
(AGO2) antibody stock solution (cat. no. 10686-1-AP; Wuhan Sanying
Biotechnology) for 30 min at room temperature, and relative
hsa_circ_0001666 expression levels were determined by
RT-quantitative (q)PCR.
Animal experiments
Animal care was performed in line with the Guide for
the Care and Use of Laboratory Animals (26) and all experiments were approved by
Shengjing Hospital of China Medical University. A total of 24 male
BALB/c nude mice (age, 4 weeks; weight, 14–16 g) were purchased
from Beijing HFK Bioscience Co., Ltd. and housed in a 12-h
light/dark cycle with a humidity of 45–55% at 22±1°C, and had free
access to food and water. Briefly, TPC-1 or IHH-4 cells with
hsa_circ_0001666 expression stably knocked down or stably
expressing sh-NC cells at density of 1×107 in 100 µl
RPMI-1640 or DMEM/RPMI-1640 (both Sigma-Aldrich; Merck KGaA) per
mouse were subcutaneously injected into the right side of the
axilla of nude mice. Tumor size was measured every 4 days and
calculated as length × width2 × 0.5. The maximum tumor
size was <0.5 cm3. All mice survived during
experiments. Mice were sacrificed at 28 days by injecting sodium
pentobarbital (200 mg/kg) intraperitoneally and tumor tissue was
harvested.
Histological analysis
The xenograft tumors were fixed in 4%
paraformaldehyde for 48 h at room temperature. After being embedded
in paraffin, the xenograft tumors were sectioned into 5-µm slices
for histological examination. For cell apoptosis analysis in
vivo, tissue sections were blocked in 3%
H2O2 for 10 min at room temperature. TUNEL
staining was performed using an in situ Cell Death Detection
kit (Roche Diagnostics) according to the manufacturer's protocols.
For cell proliferation analysis in vivo, the sections were
blocked in goat serum stock solution (Beijing Solarbio Science
& Technology Co., Ltd.) for 15 min at room temperature and
incubated with primary antibody against Ki67 (1:100; cat. no.
A2094; Abclonal Biotech Co., Ltd.) in PBS overnight at 4°C, and
then labeled horseradish peroxidase (HRP)-conjugated goat
anti-rabbit secondary antibody (1:500; cat. no. 31460; Thermo
Fisher Scientific, Inc.) in PBS was applied for 1 h at 37°C. After
counterstaining with hematoxylin (Beijing Solarbio Science &
Technology Co., Ltd.) for 3 min at room temperature. the slices
were visualized using a light microscope to observe three fields of
view at ×400 magnification.
RT-qPCR
Total RNA from all PTC cell lines, clinical tumor,
adjacent normal or xenograft tumor tissues were extracted using
TRIpure (BioTeke Corporation) and reverse-transcribed into cDNA
using a Super M-MLV Reverse Transcriptase kit (BioTeke Corporation)
according to the manufacturer's procedure at 42°C for 50 min.
RT-qPCR was performed using SYBR Green (Sigma-Aldrich; Merck KGaA).
The thermocycling conditions for hsa_circ_0001666 were as follows:
94°C for 5 min, 40 cycles of 94°C for 15 sec, 60°C for 25 sec and
72°C for 30 sec. The thermocycling conditions for miRNAs were as
follows: 94°C for 4 min, 40 cycles of 94°C for 15 sec, 60°C for 20
sec and 72°C for 15 sec. The thermocycling conditions for FAM120B
and ETV4 were as follows: 94°C for 5 min, 40 cycles of 94°C for 15
sec, 60°C for 25 sec and 72°C for 30 sec. The relative expression
levels of hsa_circ_0001666, miR-330-5p, miR-193a-5p, miR-326,
FAM120B and ETV4 were determined using the 2−ΔΔCt method
(27). β-actin was used as the
internal control for circRNA and mRNA and U6 was used as the
internal control for miRNA. The relative gene expression levels in
cell lines and xenograft tumors were normalized to the control. All
primer sequences are listed in Table
SII.
Western blotting
Total proteins from TPC-1 and IHH-4 cells or tissues
were isolated using cell lysis buffer and quantified using a BCA
assay kit (both Beyotime Institute of Biotechnology). Equal
quantities of proteins (20 µl) were loaded on 12 or 15% SDS-PAGE
gel and transferred to a PVDF membrane (EMD Millipore). After
blocking in 5% non-fat milk for 1 h at room temperature, membranes
were incubated with primary antibodies against cyclin D1 (1:1,000;
cat. no. A19038; Abclonal Biotech Co., Ltd.), cyclin E (1:1,000;
cat. no. AF0144; Affinity Biosciences), Caspase-3 (1:1,000; cat.
no. A19654), Caspase-9 (1:1,000; cat. no. A2636), ETV4 (1:1,000;
cat. no. A5797) or β-actin (1:10,000; cat. no. AC004; all Abclonal
Biotech Co., Ltd.) in 5% non-fat milk dilution overnight at 4°C.
Membranes were incubated with HRP-conjugated goat anti-rabbit
(1:5,000; cat. no. A0208) or goat anti-mouse secondary antibodies
(1:5,000; cat. no. A0216; Beyotime Institute of Biotechnology) in
5% non-fat milk for 45 min at 37°C. Protein signals were visualized
with an ECL kit (Beyotime Institute of Biotechnology), captured and
analyzed using Gel-Pro-Analyzer 4.0 software (Media Cybernetics,
Inc.).
Statistical analysis
Data were presented as mean ± SD. Differences
between groups were compared using unpaired Student's t-test or
one-way ANOVA followed by post hoc Bonferroni's correction using
GraphPad Prism 8.0 software (GraphPad Software, Inc.). Comparisons
of hsa_circ_0001666 expression levels between clinical tumors and
adjacent normal tissues were analyzed by paired Student's t-test.
Associations between hsa_circ_0001666 expression level and
clinicopathological features were analyzed using Chi-square or
Fisher's exact test. The numbers of independent repeats for each
experiment were as follows: In vitro, three; in vivo,
six; CCK8, five; RIP and RT-qPCR, three. P<0.05 was considered
to indicate a statistically significant difference.
Results
Identification and expression of
hsa_circ_0001666 in PTC and clinical significance
Dysregulated expression of circRNAs has been
reported to be involved in tumor progression (10). Through microarray analysis, a novel
circRNA, hsa_circ_0001666, has previously been found to be
significantly upregulated in human PTC tumors (15). In order to validate whether exons 2, 3
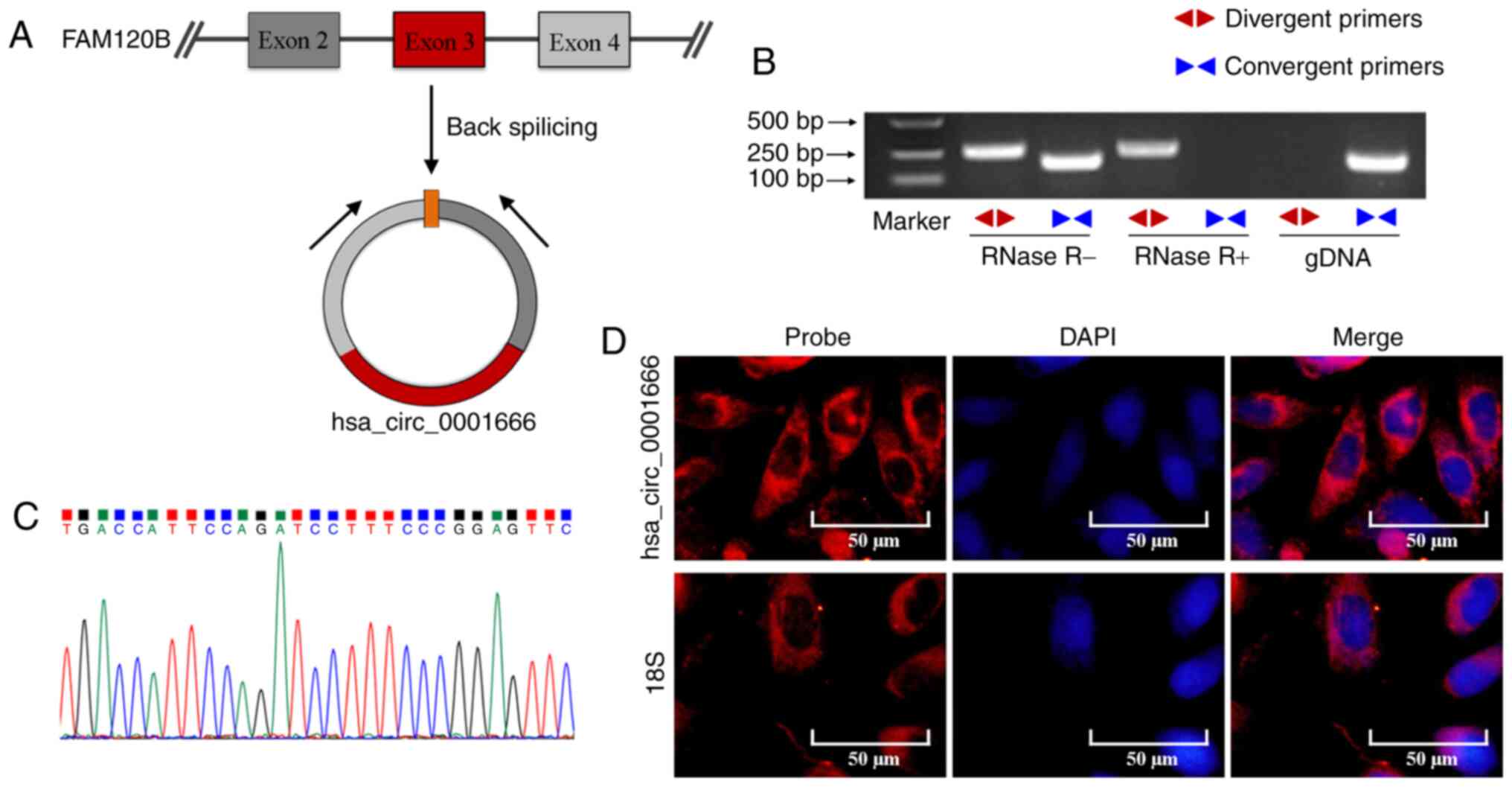
and 4 of the FAM120B gene formed hsa_circ_0001666 (Fig. 1A), divergent and convergent primers
were designed to amplify hsa_circ_0001666 and FAM120B.
hsa_circ_0001666 was detected by the divergent primers in cDNAs in
the presence or absence of RNase R treatment, but not in gDNA from
PTC cells; by contrast, the linear FAM120B mRNA amplified by
convergent primers was absent in cDNA following RNase R treatment
(Fig. 1B). Sanger sequencing of the
PCR products further confirmed the back-splicing junction of
hsa_circ_0001666 (Fig. 1C), which
supported the closed circular structure of hsa_circ_0001666.
Furthermore, FISH staining showed that that hsa_circ_0001666 was
primarily localized in the cytoplasm of PTC cells (Fig. 1D). Next, the expression of
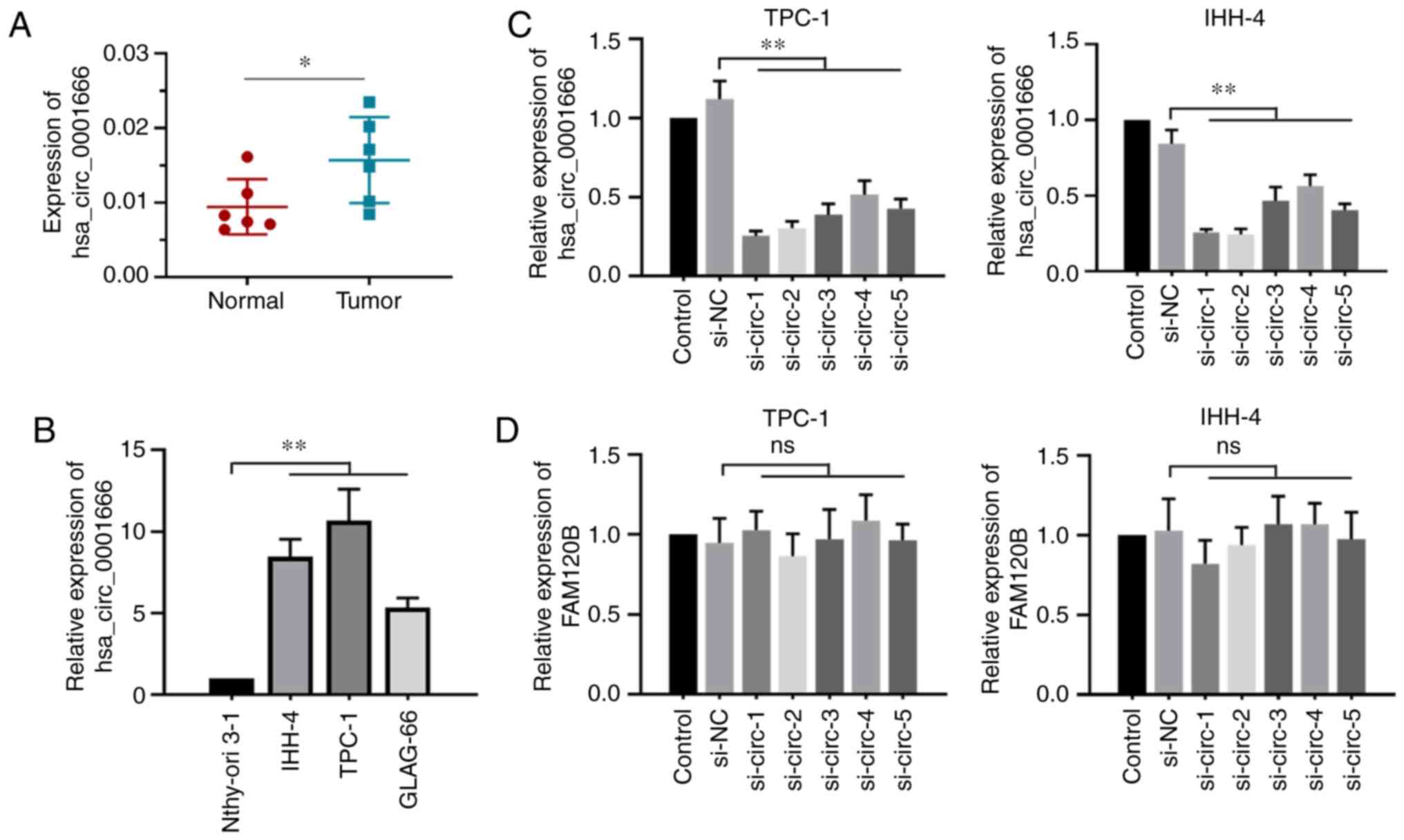
hsa_circ_0001666 in PTC was confirmed by RT-qPCR analysis. Results
showed that hsa_circ_0001666 was significantly upregulated in PTC
tissues compared with the adjacent normal specimens (Fig. 2A), consistent with results in the PTC
cell lines (Fig. 2B). The association
between hsa_circ_0001666 expression levels and clinicopathological
features of patients with PTC were analyzed. The analysis showed
that hsa_circ_0001666 expression level was associated with lymph
node metastasis, but not with sex, age and tumor stage (Table I). These results highlighted the
potential oncogenic role of hsa_circ_0001666 in PTC.
 | Table I.Association between hsa_circ_0001666
expression levels and clinicopathological features of patients with
papillary thyroid carcinoma. |
Table I.
Association between hsa_circ_0001666
expression levels and clinicopathological features of patients with
papillary thyroid carcinoma.
|
|
| hsa_circ_0001666
expression levels |
|
|---|
|
|
|
|
|
|---|
| Clinical
parameter | Cases (n=60) | Low | High | P-value |
|---|
| Sex |
|
|
|
|
|
Male | 11 | 7 | 4 |
|
|
Female | 49 | 23 | 26 | 0.5062 |
| Age, years |
|
|
|
|
|
≤60 | 48 | 27 | 21 |
|
|
>60 | 12 | 3 | 9 | 0.1042 |
| Tumor stage |
|
|
|
|
| T1 | 55 | 30 | 25 |
|
|
T2-T4 | 5 | 0 | 5 | 0.0522 |
| Lymph node
metastasis |
|
|
|
|
| N0 | 32 | 20 | 12 |
|
| N1 | 28 | 10 | 18 | 0.0384a |
Silencing hsa_circ_0001666 inhibits
cell proliferation and promotes apoptosis in vitro
In order to investigate the function of
hsa_circ_0001666 in PTC cells, five specific siRNAs targeting
hsa_circ_0001666 were prepared. Results showed that these siRNAs
significantly decreased hsa_circ_0001666 expression levels but did
not affect FAM120B mRNA expression levels (Fig. 2C and D). The two siRNAs (si-circ-1 and
si-circ-2) with the most efficient interference ability were used
for subsequent experiments.
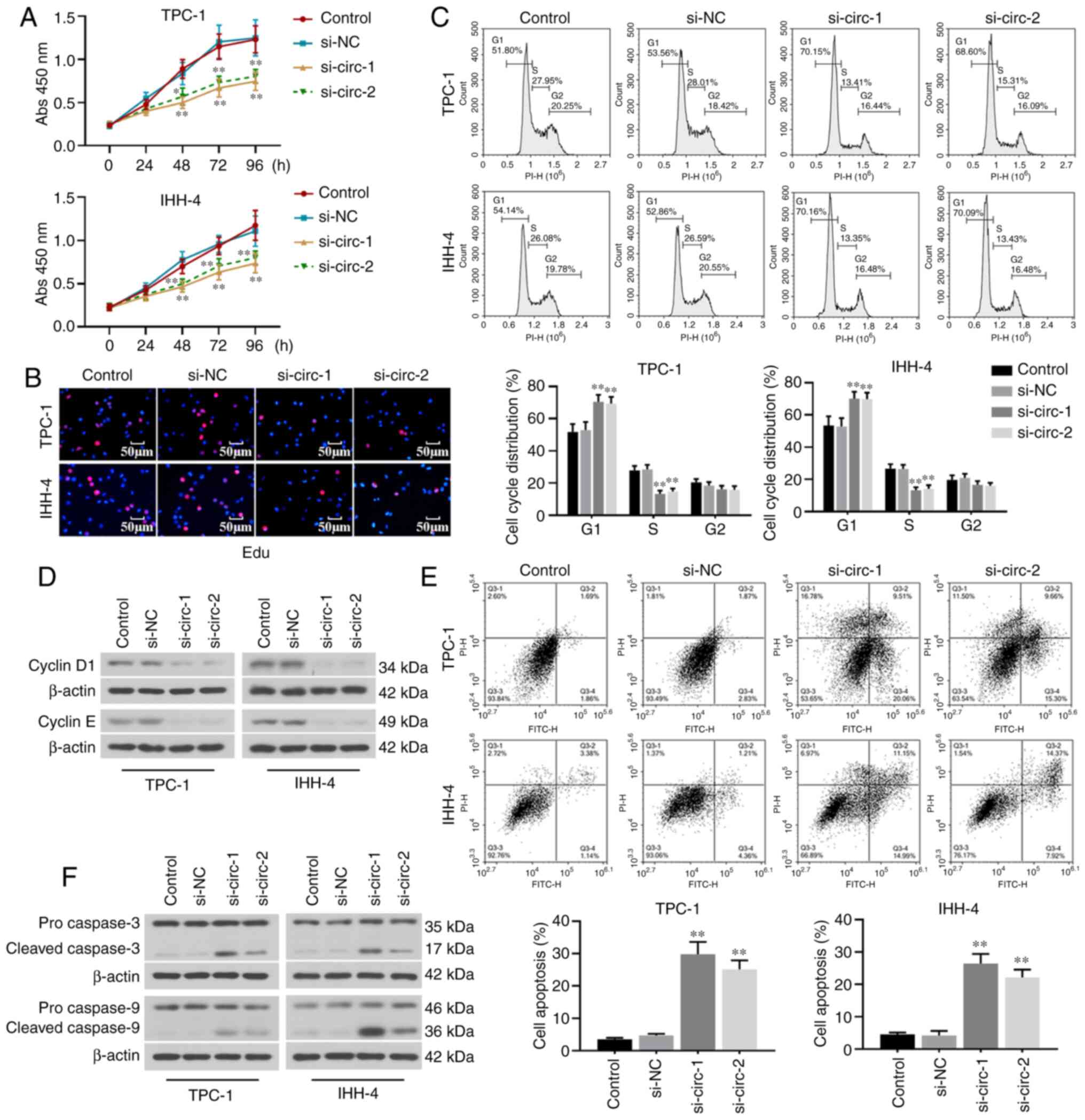
The effect of hsa_circ_0001666 on cell proliferation
was evaluated. Results from the CCK8 and Edu staining assays
demonstrated that hsa_circ_0001666 knockdown significantly
inhibited cell viability and proliferation (Fig. 3A and B). The cell cycle was arrested
at the G1 phase in TPC-1 and IHH-4 cells transfected
with hsa_circ_0001666 siRNAs (Fig.
3C). Moreover, the expression levels of cell cycle-associated
proteins (cyclins D1 and E) were significantly decreased in the
transfected cells (Fig. 3D).
The role of hsa_circ_0001666 in the apoptosis of PTC
cells was next assessed. Hsa_circ_0001666 knockdown promoted the
apoptosis of PTC cells, as shown by flow cytometry analysis
(Fig. 3E). Furthermore, the
expression levels of pro-apoptotic proteins, including cleaved
caspase 3 and caspase 9, were increased following knockdown of
hsa_circ_0001666 in PTC cells (Fig.
3F). These results showed that hsa_circ_0001666 knockdown
suppressed proliferation and promoted apoptosis of PTC cells in
vitro.
Hsa_circ_0001666 directly regulates
the miR-330-5p/miR-193a-5p/miR-326/ETV4 axis
Considering the cytoplasmic distribution of
hsa_circ_0001666 in PTC cells, it was speculated that it may exert
its functions by sponging miRNAs. In total, three potential miRNAs
(miR-330-5p, miR-193a-5p and miR-326) were predicted to be
complementary to hsa_circ_0001666 (Fig.
4A). The transfection efficiency for miR-330-5p, miR-193a-5p or
miR-326 mimics is shown in Fig.
S1A-C. Luciferase reporter assays demonstrated that miR-330-5p,
miR-193a-5p or miR-326 mimics inhibited the luciferase activity of
cells co-transfected with hsa_circ_0001666 wild-type plasmid.
However, there were no statistically significant changes in the
luciferase activity of cells co-transfected with the mutant
hsa_circ_0001666 (Fig. 4B). RIP
assays were performed to demonstrate a significant enrichment of
AGO2 binding to hsa_circ_0001666 compared with IgG in TPC-1 and
IHH-4 cells (Fig. 4C). Based on the
predicted results, these three miRNAs were shown to have binding
sites with ETV4 mRNA (Fig. 4D).
Results suggested that the binding activity of the wild-type ETV4
was decreased by these miRNAs compared with the mutant ETV4
(Fig. 4E). It was also demonstrated
that both the mRNA and protein expression levels of ETV4 were
significantly downregulated by these three miRNAs (Fig. 4F and G). Therefore, the data
illustrated that hsa_circ_0001666 shared a common binding motif on
miR-330-5p, miR-193a-5p and miR-326 with ETV4 and directly
regulated the miR-330-5p/miR-193a-5p/miR-326 axis to upregulate
ETV4 expression.
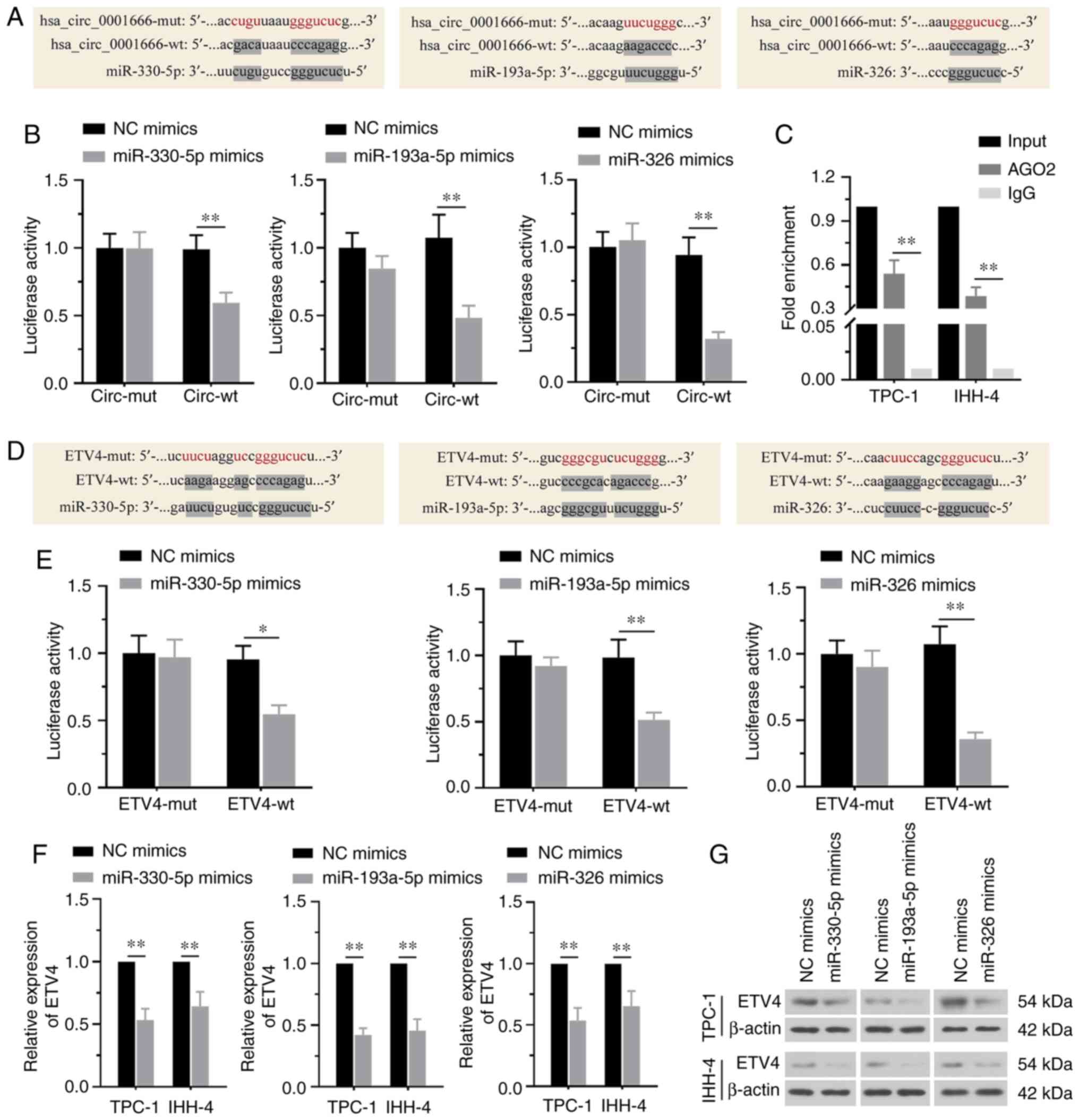 | Figure 4.Hsa_circ_0001666 directly targets the
miR-330-5p/miR-193a-5p/miR-326/ETV4 axis. (A) Sequence alignment of
the binding sites between hsa_circ_0001666 and miR-330-5p,
miR-193a-5p or miR-326. (B) Luciferase reporter assays showed that
hsa_circ_0001666 directly bound to these miRs. (C) RNA binding
protein immunoprecipitation analysis demonstrated that
hsa_circ_0001666 bound to miRs. (D) Sequence alignment of the
binding sites between ETV4 and miR-330-5p, miR-193a-5p or miR-326.
(E) Luciferase reporter assays confirmed that miR-330-5p,
miR-193a-5p, miR-326 targeted ETV4. (F) mRNA and (G) protein
expression levels of ETV4 were decreased by the three miR mimics.
*P<0.05, **P<0.01. miR, microRNA; ETV4, ETS variant
transcription factor 4; mut, mutant; wt, wild-type; NC, negative
control; circ, circular; AGO2, argonaute RISC catalytic component
2. |
Hsa_circ_0001666 facilitates PTC cell
growth by regulating the miR-330-5p/miR-193a-5p/miR-326/ETV4
axis
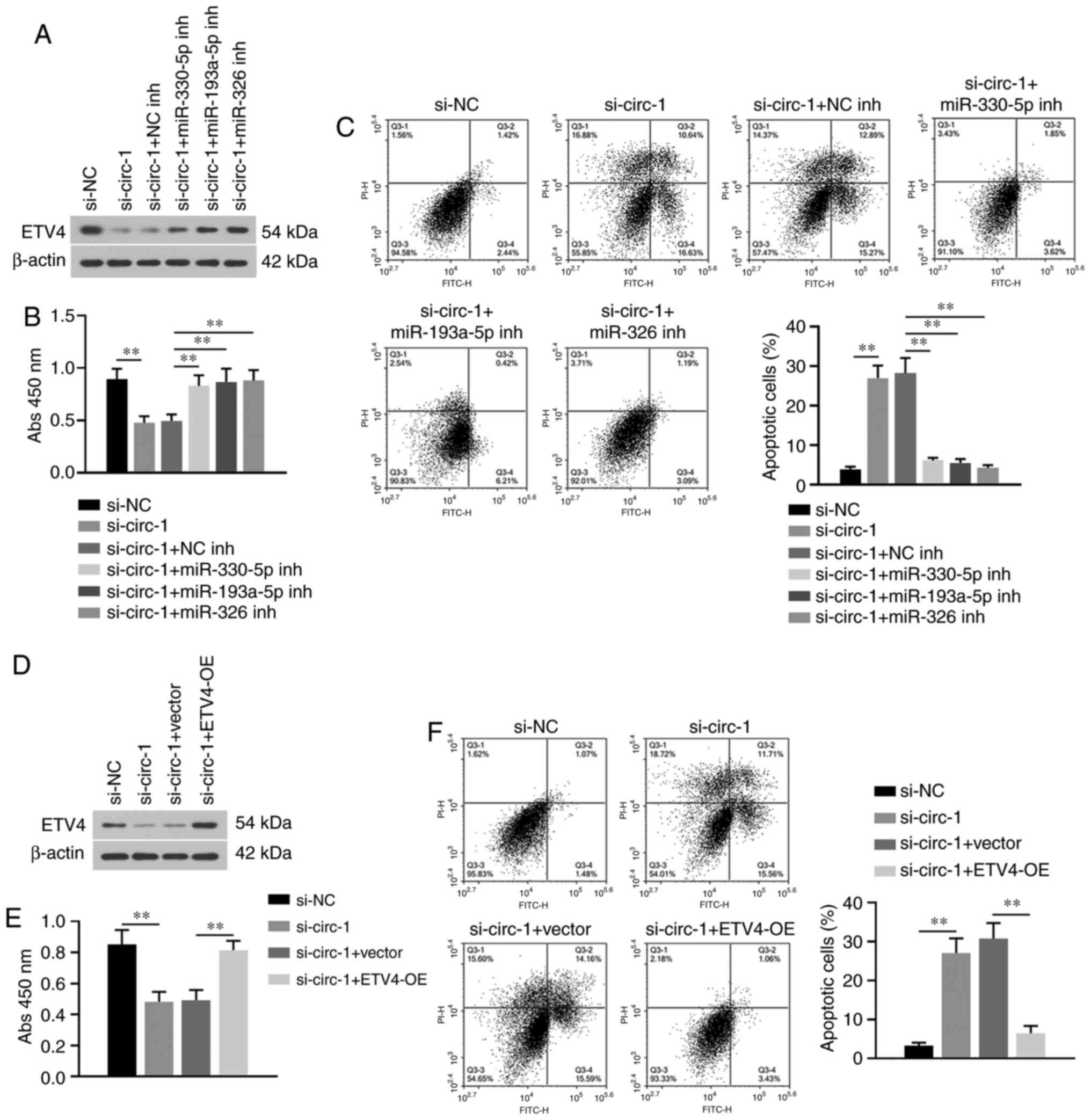
In order to determine whether the
miR-330-5p/miR-193a-5p/miR-326/ETV4 axis mediated the effect of
hsa_circ_0001666 on PTC cell proliferation and apoptosis, rescue
experiments were performed. The transfection efficiency for
miR-330-5p, miR-193a-5p or miR-326 inhibitor is presented in
Fig. S1D-F. Specific inhibitors
targeting miR-330-5p, miR-193a-5p or miR-326 resulted in
upregulation of ETV4 following knockdown of hsa_circ_0001666
(Fig. 5A). The decreased cell
viability and increased apoptosis in PTC cells with
hsa_circ_0001666 knock down were reversed by these miRNA inhibitors
(Fig. 5B and C). In addition, the
role of ETV4 in regulation of hsa_circ_0001666 on PTC progression
was further confirmed. Transfection of the ETV4 overexpression
plasmid resulted in increased expression levels of ETV4 and also
reversed the inhibitory effect of hsa_circ_0001666 knockdown on PTC
cell proliferation (Fig. 5D and E).
Moreover, the increased apoptotic rates were also suppressed by
overexpression of ETV4 (Fig. 5F).
Altogether, the data revealed that hsa_circ_0001666 targeted the
miR-330-5p/miR-193a-5p/miR-326/ETV4 axis to promote PTC cell
growth.
Silencing hsa_circ_0001666 inhibits
tumor growth in vivo
TPC-1 or IHH-4 cells with hsa_circ_0001666 stably
knocked down were injected into nude mice to determine the effect
of hsa_circ_0001666 in vivo. Tumor sizes were significantly
smaller in mice injected with by hsa_circ_0001666 knockdown cells
(Fig. 6A). Knockdown of
hsa_circ_0001666 induced a decrease in hsa_circ_0001666 expression
levels and increased expression of miR-330-5p, miR-193a-5p and
miR-326 in the tumors (Fig. 6B-E).
Furthermore, a significant decrease in both ETV4 mRNA and protein
expression levels was observed in the tumor tissue of mice injected
with hsa_circ_0001666 knockdown cells (Fig. 6F and G). Histological analysis showed
that hsa_circ_0001666 knockdown decreased Ki67 expression levels
and increased the proportion of apoptotic cells in the xenograft
tumors (Fig. 6H and I). The in
vivo results supported the hypothesis that hsa_circ_0001666
exhibited an oncogenic effect on PTC tumor growth. The overall
findings of the present study are summarized in Fig. 7.
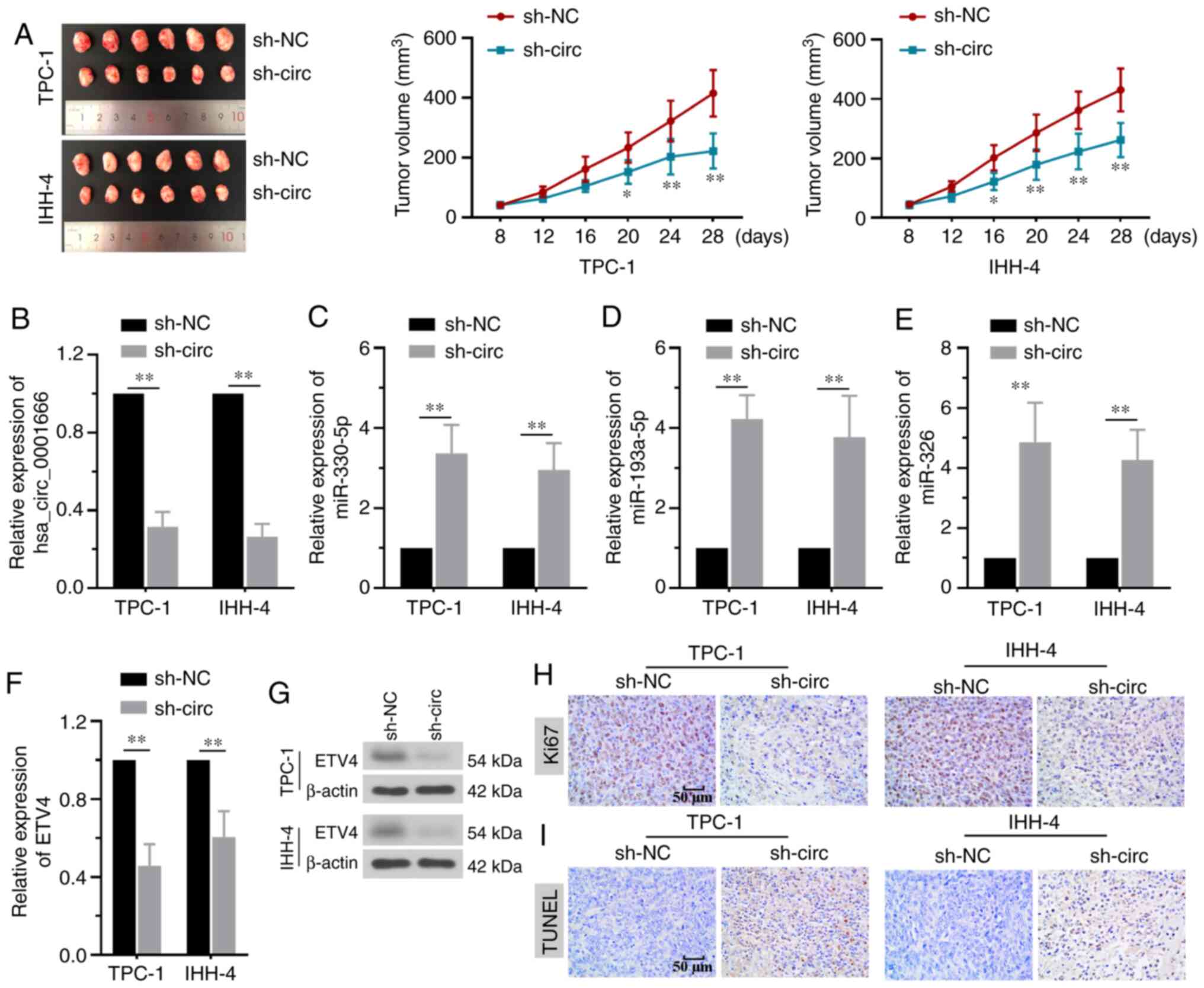 | Figure 6.Silencing hsa_circ_0001666 inhibits
tumor growth in vivo. (A) TPC-1 or IHH-4 cells transfected
with sh-NC or sh-hsa_circ_0001666 were subcutaneously injected into
nude mice, which were sacrificed 28 days later. Knockdown of
hsa_circ_0001666 decreased tumor size. (B) Inhibition of
hsa_circ_0001666 expression decreased its expression and increased
(C) miR-330-5p, (D) miR-193a-5p and (E) miR-326 expression levels.
ETV4 (F) mRNA and (G) protein expression levels were downregulated
by hsa_circ_0001666 shRNA. (H) Ki67 expression was decreased by
knocking down hsa_circ_0001666, as shown by immunohistochemistry.
(I) Apoptosis was increased by hsa_circ_0001666 silencing, as shown
by TUNEL staining. *P<0.05, **P<0.01 vs. sh-NC. circ,
circular; sh, short hairpin; NC, negative control; miR, microRNA;
ETV4, ETS variant transcription factor 4. |
Discussion
Abnormal expression of circRNAs in several types of
cancer has been shown to serve an important role in aggressive
tumor behavior, such as that of glioma, urothelial carcinoma and
cervical cancer (25,28,29). The
present study demonstrated that hsa_circ_0001666 expression was
significantly upregulated in PTC tissue compared with matched
adjacent normal tissue. In order to uncover the biological role of
hsa_circ_0001666 and the underlying mechanism in PTC progression,
loss-of-function experiments were performed using PTC cells and
xenograft animals. Hsa_circ_0001666 knockdown suppressed
proliferation and promoted apoptosis of PTC cells both in
vitro and in vivo. The inhibitory effect of
hsa_circ_0001666 silencing on PTC was reversed by miRNA inhibitors
or ETV4 overexpression. The results suggested that hsa_circ_0001666
functioned as a sponge to mediate the
miR-330-5p/miR-193a-5p/miR-326/ETV4 axis and promote PTC tumor
progression.
Emerging evidence has shown that circRNAs are novel
molecules associated with tumorigenesis, metastasis or drug
resistance in PTC (30–32). It has previously been shown that
circFOXM1 acts as a tumor promoter in PTC by regulating
miR-1179/HMGB1 (33). Another
circRNA, circRAPGEF5, was found to upregulate FGFR1 expression and
facilitate PTC proliferation and metastasis via competitively
binding with miR-198 (14).
Microarray analysis by Peng et al (15) demonstrated that hsa_circ_0001666
expression is increased in PTC; this circRNA is located at chr6:
70726457-170739638. Bioinformatics analysis suggested that
hsa_circ_0001666 functions as a sponge due to the presence of
multiple miRNA binding sites in breast cancer (34). To the best of our knowledge, however,
studies regarding the biological features of hsa_circ_0001666 have
not been performed. The present study demonstrated that
hsa_circ_0001666 was upregulated in PTC tissue and cell lines.
Silencing hsa_circ_0001666 suppressed proliferation and
G1/S phase transition and promoted cell apoptosis in
vitro. Similarly, downregulated expression levels of cell cycle
proteins and upregulated expression levels of pro-apoptotic
proteins were observed. Xenograft experiments confirmed the
oncogenic effect of hsa_circ_0001666 on tumor growth in
vivo.
circRNA-miRNA-mRNA interactions mediate
tumorigenesis (35,36). Hsa_circ_0001666 was here shown to
possess binding sites with three potential miRNAs (miR-330-5p,
miR-193a-5p and miR-326). RIP assay confirmed the interaction
between hsa_circ_0001666 and miRNAs. Luciferase reporter assays
confirmed these three miRNAs directly bound with hsa_circ_0001666.
These three miRNAs have been investigated in various types of
cancer. For example, miR-330-5p was shown to serve as a tumor
suppressor in esophageal adenocarcinoma via regulating MMP1
(37). miR-326 inhibits the growth of
breast cancer cells via regulating the ERBB/PI3K signaling pathway
(38). miR-193a-5p directly targets
SPOCK1 in hepatocellular carcinoma to repress its malignancy
(39). Additionally, miR-330-5p and
miR-326 have both been demonstrated to play tumor-suppressive roles
in PTC progression (20,21). However, research on miR-193a-5p in PTC
is limited. The present study showed that silencing miR-330-5p,
miR-193a-5p or miR-326 using specific inhibitors negatively
regulated the effect of hsa_circ_0001666 siRNAs on PTC cell
proliferation and apoptosis.
It has been shown that miRNAs exert an important
role in biological processes via regulating gene expression, such
as miR-221 and miR-330-5p (16,37). In
the present study, three miRNAs directly targeted ETV4, a
transcriptional factor of E26 transformation-specific family, which
is overexpressed in breast, lung and pancreatic cancer (22–24). Xu
et al (40) showed that ETV4
bound to FOSL1 promoter to increase cell migration in a
PI3K/AKT-dependent manner in clear cell renal cell carcinoma. In
hepatocellular carcinoma, ETV4 overexpression increases resistance
to sorafenib and cisplatin to protect against apoptosis (41); it was also reported to be associated
with the clinical response to MEK inhibitors in patients with
melanoma (42), supporting its use as
a potential biomarker in multiple types of cancer. The present
study demonstrated that hsa_circ_0001666 silencing-induced effects
on cell proliferation and apoptosis were reversed by ETV4
overexpression. Taken together, these data suggested that
hsa_circ_0001666 serves as a sponge to promote tumor malignancy via
the miR-330-5p/miR-193a-5p/miR-326/ETV4 pathway. In addition to
ETV4, target genes, such as tripartite motif containing 25,
transcription factor 3 and dual specificity phosphatase 28, were
also identified to bind with the miRNAs in the present study. In
future studies, the effect of these potential targets on the
regulation of hsa_circ_0001666 in PTC progression should be
assessed.
The present study has several limitations. For
example, as a novel circRNA, data regarding hsa_circ_0001666 in
cancer is limited. The present study showed that hsa_circ_0001666
functioned as an oncogene, promoting PTC progression both in
vitro and in vivo. However, whether hsa_circ_0001666
improves diagnostic sensitivity and specificity in combination with
traditional biomarkers in PTC tumors remains to be determined. In
addition, considering the structural specificity of circRNAs, the
circRNA overexpressing plasmid is challenging to construct because
of the instability of its loop-forming efficiency and accuracy.
Thus, the biological functions of circRNAs using both
overexpressing and knockdown vectors should be assessed when
technology for modulation of circRNA expression has matured.
Furthermore, due to the small size of PTC tumors and the emergence
of COVID-19, clinical tumors and adjacent normal tissue were
difficult to access. Only a limited quantity was obtained to assess
the potential association with clinicopathological features. Thus,
additional samples should be gathered for further analysis.
In summary, the upregulated expression of
hsa_circ_0001666 exhibited an oncogenic role in the malignant
behavior of PTC tumors. Hsa_circ_0001666 functioned as a miRNA
sponge to inhibit proliferation and enhance apoptosis of PTC cells
by positively regulating ETV4. The findings of the present study
highlight the potential role of circRNA hsa_circ_0001666 as a
molecular target for PTC diagnosis and treatment.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
Science and Technology Planning Project of Shenyang (grant no.
19-112-4-013) and Medical Education Science Research of China
Medical University (grant no. YDJK2020042).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
YQ and JH conceptualized and designed the research.
YQ, JH, YZ and LW performed the experiments. YY and BY contributed
reagents and collected clinical samples. YQ and ZT analyzed the
data. YQ and JH wrote and revised the manuscript. YQ, JH, YY and ZT
confirm the authenticity of all the raw data. All authors read and
approved the final manuscript and agree to be accountable for all
aspects of the research in ensuring that the accuracy or integrity
of any part of the work are appropriately investigated and
resolved.
Ethics approval and consent to
participate
The present study was approved by Shengjing Hospital
of China Medical University (approval no. 2019PS322K). All patients
provided written informed consent. Animal experiments were approved
by Shengjing Hospital of China Medical University (approval no.
2019PS296K).
Patient consent for publication
Not applicable.
Competing interests
The authors declare they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ancker OV, Krüger M, Wehland M, Infanger M
and Grimm D: Multikinase inhibitor treatment in thyroid cancer. Int
J Mol Sci. 21:102019. View Article : Google Scholar
|
|
3
|
Jiang C, Cheng T, Zheng X, Hong S, Liu S,
Liu J, Wang J and Wang S: Clinical behaviors of rare variants of
papillary thyroid carcinoma are associated with survival: A
population-level analysis. Cancer Manag Res. 10:465–472. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fallahi P, Ferrari SM, Galdiero MR,
Varricchi G, Elia G, Ragusa F, Paparo SR, Benvenga S and Antonelli
A: Molecular targets of tyrosine kinase inhibitors in thyroid
cancer. Semin Cancer Biol. 26:S1044–S1579. 2020.
|
|
5
|
Chakravarty D, Santos E, Ryder M, Knauf
JA, Liao XH, West BL, Bollag G, Kolesnick R, Thin TH, Rosen N, et
al: Small-Molecule MAPK inhibitors restore radioiodine
incorporation in mouse thyroid cancers with conditional BRAF
activation. J Clin Invest. 121:4700–4711. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jeck WR and Sharpless NE: Detecting and
characterizing circular RNAs. Nat Biotechnol. 32:453–461. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Thomson DW and Dinger ME: Endogenous
microRNA sponges: Evidence and controversy. Nat Rev Genet.
17:272–283. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang F, Nazarali AJ and Ji S: Circular
RNAs as potential biomarkers for cancer diagnosis and therapy. Am J
Cancer Res. 6:1167–1176. 2016.PubMed/NCBI
|
|
9
|
Ebbesen KK, Kjems J and Hansen TB:
Circular RNAs: Identification, biogenesis and function. Biochim
Biophys Acta. 1859:163–168. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yu J, Xu QG, Wang ZG, Yang Y, Zhang L, Ma
JZ, Sun SH, Yang F and Zhou WP: Circular RNA cSMARCA5 inhibits
growth and metastasis in hepatocellular carcinoma. J Hepatol.
68:1214–1227. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chen L, Nan A, Zhang N, Jia Y, Li X, Ling
Y, Dai J, Zhang S, Yang Q, Yi Y, et al: Circular RNA 100146
functions as an oncogene through direct binding to miR-361-3p and
miR-615-5p in non-small cell lung cancer. Mol Cancer. 18:132019.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang Q, Guo H, Wang S, Ma Y, Chen H, Li
H, Li J, Li X, Yang F, Qiu M, et al: A novel circular RNA,
circXPO1, promotes lung adenocarcinoma progression by interacting
with IGF2BP1. Cell Death Dis. 11:10312020. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Guo D, Li F, Zhao X, Long B, Zhang S, Wang
A, Cao D, Sun J and Li B: Circular RNA expression and association
with the clinicopathological characteristics in papillary thyroid
carcinoma. Oncol Rep. 44:519–532. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu W, Zhao J, Jin M and Zhou M:
circRAPGEF5 contributes to papillary thyroid proliferation and
metastatis by regulation miR-198/FGFR1. Mol Ther Nucleic Acids.
14:609–616. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Peng N, Shi L, Zhang Q, Hu Y, Wang N and
Ye H: Microarray profiling of circular RNAs in human papillary
thyroid carcinoma. PLoS One. 12:e01702872017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kim HJ, Kim YH, Lee DS, Chung JK and Kim
S: In vivo imaging of functional targeting of miR-221 in papillary
thyroid carcinoma. J Nucl Med. 49:1686–1693. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jia M, Shi Y, Li Z, Lu X and Wang J:
MicroRNA-146b-5p as an oncomiR promotes papillary thyroid carcinoma
development by targeting CCDC6. Cancer Lett. 443:145–156. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu G, Zhou W, Lin X, Sun Y, Li J, Xu H,
Shi P, Gao L and Tian X: circRASSF2 acts as ceRNA and promotes
papillary thyroid carcinoma progression through miR-1178/TLR4
signaling pathway. Mol Ther Nucleic Acids. 19:1153–1163. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu F, Yin R, Chen X, Chen W, Qian Y, Zhao
Y, Jiang Y, Ma D, Hu T, Yu T, et al: Over-Expression of miR-206
decreases the euthyrox-resistance by targeting MAP4K3 in papillary
thyroid carcinoma. Biomed Pharmacother. 114:1086052019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao Y, Wang F, Zhang L, Kang M, Zhu L, Xu
L, Liang W and Zhang W: LINC00311 promotes cancer stem-like
properties by targeting miR-330-5p/TLR4 pathway in human papillary
thyroid cancer. Cancer Med. 9:1515–1528. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Nie FR, Li QX, Wei HF and Ma Y: MiR-326
inhibits the progression of papillary thyroid carcinoma by
targeting MAPK1 and ERBB4. Neoplasma. 67:604–613. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang Y, Ding X, Liu B, Li M, Chang Y, Shen
H, Xie SM, Xing L and Li Y: ETV4 overexpression promotes
progression of non-small cell lung cancer by upregulating PXN and
MMP1 transcriptionally. Mol Carcinog. 59:73–86. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen Y, Sumardika IW, Tomonobu N,
Kinoshita R, Inoue Y, Iioka H, Mitsui Y, Saito K, Ruma IM, Sato H,
et al: Critical role of the MCAM-ETV4 axis triggered by
extracellular S100A8/A9 in breast cancer aggressiveness. Neoplasia.
21:627–640. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tyagi N, Deshmukh SK, Srivastava SK, Azim
S, Ahmad A, Al-Ghadhban A, Singh AP, Carter JE, Wang B and Singh S:
ETV4 facilitates cell-cycle progression in pancreatic cells through
transcriptional regulation of cyclin D1. Mol Cancer Res.
16:187–196. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang Q, Chen Z, Zhao L and Xu H: Circular
RNA hsa_circ_0000515 acts as a miR-326 sponge to promote cervical
cancer progression through up-regulation of ELK1. Aging (Albany
NY). 11:9982–9999. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
National Research Council, . Guide for the
Care and Use of Laboratory Animals. 8th edition. National Academies
Press; Washington, DC: 2011
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen J, Chen T, Zhu Y, Li Y, Zhang Y, Wang
Y, Li X, Xie X, Wang J, Huang M, et al: circPTN sponges
miR-145-5p/miR-330-5p to promote proliferation and stemness in
glioma. J Exp Clin Cancer Res. 38:3982019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang C, Tao W, Ni S and Chen Q: Circular
RNA circ-foxo3 induced cell apoptosis in urothelial carcinoma via
interaction with miR-191-5p. Onco Targets Ther. 12:8085–8094. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Gui X, Li Y, Zhang X, Su K and Cao W:
Circ_LDLR promoted the development of papillary thyroid carcinoma
via regulating miR-195-5p/LIPH axis. Cancer Cell Int. 20:2412020.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guan H, Guo Y, Liu L, Ye R, Liang W, Li H,
Xiao H and Li Y: INAVA promotes aggressiveness of papillary thyroid
cancer by upregulating MMP9 expression. Cell Biosci. 8:262018.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Liu F, Zhang J, Qin L, Yang Z, Xiong J,
Zhang Y, Li R, Li S, Wang H and Yu B: Circular RNA EIF6
(Hsa_circ_0060060) sponges miR-144-3p to promote the
cisplatin-resistance of human thyroid carcinoma cells by autophagy
regulation. Aging (Albany NY). 10:3806–3820. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ye M, Hou H, Shen M, Dong S and Zhang T:
Circular RNA circFOXM1 plays a role in papillary thyroid carcinoma
by sponging miR-1179 and regulating HMGB1 expression. Mol Ther
Nucleic Acids. 19:741–750. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Afzali F and Salimi M: Unearthing
regulatory axes of breast cancer circRNAs networks to find novel
targets and fathom pivotal mechanisms. Interdiscip Sci. 11:711–722.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen
B, Luo Y, Lyu D, Li Y, Shi G, et al: Circular RNA profiling reveals
an abundant circHIPK3 that regulates cell growth by sponging
multiple miRNAs. Nat Commun. 7:112152016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bibby BAS, Miranda CS, Reynolds JV,
Cawthorne CJ and Maher SG: Silencing microRNA-330-5p increases MMP1
expression and promotes an invasive phenotype in oesophageal
adenocarcinoma. BMC Cancer. 19:7842019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ghaemi Z, Soltani BM and Mowla SJ:
MicroRNA-326 functions as a tumor suppressor in breast cancer by
targeting ErbB/PI3K signaling pathway. Front Oncol. 9:6532019.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li P, Xiao Z, Luo J, Zhang Y and Lin L:
MiR-139-5p, miR-940 and miR-193a-5p inhibit the growth of
hepatocellular carcinoma by targeting SPOCK1. J Cell Mol Med.
23:2475–2488. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Xu L, Hu H, Zheng LS, Wang MY, Mei Y, Peng
LX, Qiang YY, Li CZ, Meng DF, Wang MD, et al: ETV4 is a theranostic
target in clear cell renal cell carcinoma that promotes metastasis
by activating the pro-metastatic gene FOSL1 in a PI3K-AKT dependent
manner. Cancer Lett. 482:74–89. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xiaohui C, Xin LI and Dehua WU: E26
transformation-specific variant 4 promotes sorafenib and cisplatin
resistance in hepatocellular carcinoma cells in vitro. Nan Fang Yi
Ke Da Xue Xue Bao. 39:875–882. 2019.(In Chinese). PubMed/NCBI
|
|
42
|
Gupta A, Towers C, Willenbrock F, Brant R,
Hodgson DR, Sharpe A, Smith P, Cutts A, Schuh A, Asher R, et al:
Dual-Specificity protein phosphatase DUSP4 regulates response to
MEK inhibition in BRAF wild-type melanoma. Br J Cancer.
122:506–516. 2020. View Article : Google Scholar : PubMed/NCBI
|