Introduction
Epithelial ovarian cancer is the second most
prevalent gynecological malignancy with a considerably high
mortality rate (1). Unfortunately,
most patients are usually diagnosed at an advanced stage due to a
lack of effective screening strategies (2). Currently, surgical resection and
platinum-based combination chemotherapy are the principal
treatments for this disease. Despite great progress in ovarian
cancer treatment, the majority of patients eventually experience
chemoresistance, which could lead to tumor recurrence and
metastasis (3,4). Therefore, the search for effective
therapies remains to be resolved.
Triptolide (TPL), a diterpene triepoxide extracted
from the Chinese herb Tripterygium wilfordii, has been
reported to present marked effects in various cancers, such as
hepatocellular (5) and pancreatic
carcinoma (6), bladder (7), breast (8)
and ovarian cancers (9–13). The anticancer properties of TPL have
been revealed in numerous contexts, including the induction of cell
cycle arrest and apoptosis and the inhibition of cell
proliferation, metastasis and angiogenesis (5–13). Our
previous study demonstrated that TPL synergistically inhibited the
growth of cisplatin (DDP)-resistant ovarian cancer cells by
reactive oxygen species (ROS)-mediated apoptosis induction
(12). Recently, TPL has been
considered to be a potential autophagy modulator (14). However, the autophagic effect of TPL
on resistant human epithelial ovarian cancer cells is not
clear.
Autophagy is a highly conserved cellular catabolic
pathway that plays a homeostatic role in normal cells. There is a
growing interest in utilizing autophagic mechanisms to suppress
tumor growth (15). Intriguingly,
autophagy has multifaceted and context-specific roles in cancer.
Altered autophagic activity has been revealed to favor cancer cell
survival in response to stress stimuli, such as starvation, ROS,
and chemotherapy, and mediate resistance to these treatments
(16). On the other hand, excessive
autophagy is known as type II programmed cell death and has been
exploited as a potential strategy for cancer therapy and tumor
chemosensitization, especially in apoptosis-defective tumor cells
(17). The regulation of autophagy is
highly complex. Notably, signal transducer and activator of
transcription-3 (STAT3) is a redox-sensitive transcription factor
that participates in the regulation of autophagic processes upon
activation by Janus family kinases (JAKs) (18,19).
Activated STAT3 dimerizes, transfers into the nucleus and regulates
the transcription of numerous target genes, including certain
autophagy-related genes (20). To
date, accumulating evidence and studies have established the role
of the aberrantly active Janus kinase 2 (JAK2)/STAT3 pathway in
ovarian cancer progression and chemoresistance (21,22).
Inhibition of the JAK2/STAT3 pathway may shed new light on cancer
treatment and provide new targets for ovarian cancer therapy
(21).
In the present study, the autophagic role of TPL in
DDP-resistant ovarian cancer was evaluated using SKOV3/DDP cells
and the effects of TPL-induced autophagy on chemosensitization
in vivo were further studied. In addition, systematic
research on the underlying mechanisms involved in TPL-induced
autophagy against SKOV3/DDP cells was performed in vitro, by
various molecular biological techniques, such as flow cytometry,
gene silencing, western blotting, immunofluorescence staining and
co-immunoprecipitation.
Materials and methods
Antibodies and reagents
Antibodies to phosphorylated (p)-JAK2 (product no.
3776; 1:1,000), JAK2 (product no. 3230; 1:1,000), p-STAT3 (Y705)
(product no. 9145; used at 1:1,000 for western blotting and 1:100
for immunofluorescence staining), STAT3 (product no. 12640;
1:1,000), myeloid cell leukemia-1 (Mcl-1; product no. 94296;
1:1,000), LC3 (product no. 4599; used at 1:1,000 for western
blotting and 1:6,400 for immunohistochemistry), p62 (product no.
88588; used at 1:1,000 for western blotting and 1:1,600 for
immunohistochemistry), Beclin1 (product no. 3495; 1:1,000),
caspase-3 (product no. 9662; used at 1:1,000 for western blotting
and immunohistochemistry), β-actin (product no. 3700; 1:1,000) and
Ki67 (product no. 12202; 1:800) were purchased from Cell Signaling
Technology, Inc. HRP-labeled anti-rabbit (cat. no. SA00001-2;
1:5,000) and anti-mouse IgG (cat. no. SA00001-1; 1:5,000) were
obtained from ProteinTech, Group, Inc. Chemicals, including
triptolide (TPL), chloroquine (CQ), 3-methyladenine (3-MA),
N-acetyl-L-cysteine (NAC), cisplatin (DDP), interleukin (IL)-6 and
AG490 were purchased from Sigma-Aldrich; Merck KGaA. Cell Counting
Kit-8 (CCK-8) assay and Reactive Oxygen Species (ROS) Assay Kit
were obtained from Beyotime Institute of Biotechnology.
Cell culture
Cisplatin-resistant SKOV3/DDP and parental SKOV3
cells were purchased from China Center for Type Culture Collection
and preserved at the Key Laboratory of Molecular Center of Jiangxi
Province. The cells were routinely cultured in RPMI-1640 medium
containing fetal bovine serum (FBS; 10%), penicillin/streptomycin
(100 U/ml) in a 5% humidified CO2 atmosphere at 37°C.
Cisplatin (0.3 µg/ml) was added to the SKOV3/DDP culture media to
maintain its acquired resistance.
Autophagy flux analysis
To track autophagic flux, SKOV3/DDP cells were
transfected with mRFP-GFP-LC3 adenovirus (Hanbio Biotechnology Co.,
Ltd.) following the manufacturer's recommendations and performed as
previously described (23). Briefly,
SKOV3/DDP cells (1×106 cells/ml) were treated with TPL
(0, 25, 50 and 100 nM) for 12 h, or treated with AG490 (50 µM) for
24 h at 37°C, respectively. After drug treatment, the formation of
autophagosomes (yellow puncta) and autolysosomes (red puncta) were
detected using a confocal laser scanning microscope under an ×400
magnification.
Cell viability assay
Cell viability was measured using Cell Counting
Kit-8 (CCK-8) assay. Briefly, SKOV3/DDP cells (1×104
cells/well) were treated with TPL (100 nM), CQ (10 µM), 3-MA (10
mM) or a combination of these compounds for 24 h at 37°C. Following
treatment, CCK-8 solution was added and incubated at 37°C for 4 h.
The optical density (OD) value of each well was measured at 450 nm
using a Microplate Reader (Molecular Devices, LLC), and the cell
viability was then calculated.
Western blot analysis
Immunoblotting analyses were performed as previously
described (12). The whole cell
protein extracts were prepared using the RIPA lysis buffer
(Solarbio Life Science), and the protein concentrations were
quantified with BCA Protein Assay Kit. Equal amounts of protein (30
µg/lane) were separated on 10–15% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to a polyvinylidene difluoride (PVDF) membrane (Thermo
Fisher Scientific, Inc.). Following blocking for 2 h using 5%
non-fat milk at room temperature, the membrane was incubated
overnight with the specific primary antibodies against p-JAK2,
JAK2, p-STAT3 (Y705), STAT3, Mcl-1, LC3, p62, Beclin1, caspase-3,
and the internal control β-actin at 4°C. After washing with PBS,
the membranes were incubated with the corresponding horseradish
peroxidase (HRP)-conjugated goat anti-rabbit IgG or anti-mouse IgG
at room temperature for 1 h. The blot bands were detected using an
enhanced chemiluminescence (ECL) kit (Thermo Fisher Scientific,
Inc.), and imaged and quantified using the ChemiDoc XRS System
(Bio-Rad Laboratories, Inc.).
Measurement of intracellular ROS
Reactive Oxygen Species Assay Kit was applied to
measure the level of intracellular ROS in SKOV3/DDP cells. Briefly,
SKOV3/DDP cells (1×106 cells/ml) were treated with TPL
(100 nM) alone or in presence of NAC (5 mM) at 37°C for 12 h, then
washed three times in PBS and incubated with DCFH-DA at 37°C for 20
min in the dark. The DCF fluorescence intensity was examined and
analyzed by cell flow cytometry (FACSCalibur; BD Biosciences).
Immunofluorescence staining
Immunofluorescence staining was carried out to
analyze the expression of p-STAT3 (Y705) in SKOV3/DDP cells.
Briefly, SKOV3/DDP cells (1×106 cells/ml) were treated
with or without 100 nM TPL at 37°C for 24 h, and then fixed with 4%
paraformaldehyde for 20 min on ice and permeablized with 0.5%
Triton X-100 for 10 min at 37°C. Then, the cells were incubated
with anti-p-STAT3 (Y705) antibody at 4°C overnight and subsequently
with CoraLite 488-conjugated Affinipure goat anti-rabbit IgG
secondary antibody (cat. no. SA00013-2; 1:500; ProteinTech Group,
Inc.) for 1 h at room temperature in the dark. Finally, the cells
were incubated with DAPI at room temperature for 10 min to stain
nuclei, and the and the images were obtained using a confocal
microscope (Olympus FV1000; Olympus Corporation) at a magnification
of ×400.
Co-immunoprecipitation (Co-IP)
assay
Co-IP assay was performed as previously reported
(24). Briefly, 1,000 µg of whole
cell lysates were prepared with 1 ml of IP lysis buffer (cat. no.
87787; Pierce IP Lysis Buffer; Thermo Fisher Scientific, Inc.)
after drug treatment. Then the lysate was incubated with the
antibody against Beclin1 and 100 µl of Protein A/G magnetic beads
(cat. no. 88803; Pierce; Thermo Fisher Scientific, Inc.) at 4°C
overnight on a rocking platform. The immunoprecipitated pellets
were collected by centrifugation at 12,000 × g for 5 min at 4°C.
Then the pellets were washed three times with the cold lysis
buffer, boiled in 2X SDS loading buffer and analyzed by western
blot analysis with anti-Mcl-1.
Beclin1 siRNA transfection
After SKOV3/DDP cells were grown into 6-well plates
and reached 50% confluence, cells were transiently transfected with
the Beclin1 small interfering (si)RNA or negative control siRNA
(100 pmol) using Lipofectamine 2000 ((Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's instructions.
Briefly, 5 µl (100 pmol) Beclin1 siRNA and 5 µl Lipofectamine 2000
were diluted in 50 µl RPMI-1640 medium, respectively, and were
incubated for 5 min at room temperature. Then, the diluted Beclin1
siRNA and Lipofectamine 2000 were mixed and incubated for 15 min at
room temperature. Next, the complexes were added to cells and
incubated for 6 h at 37°C in a 5% CO2 incubator.
Thereafter, the cells were washed with PBS and suspended in the
RPMI-1640 medium plus 10% FBS. After 48 h of transfection, cells
were treated or untreated with 100 nM TPL for an additional 24 h
and collected for western blot analysis. The Beclin1 siRNA
(5′-GGATGACAGTGAACAGTTA-3′) and negative control siRNA
(5′-UUCUCCGAACGUGUCACGUTT-3′) were designed and synthesized by
Guangzhou Ribobio Co., Ltd.
Ectopic expression of Mcl-1 by
transient transfection
Proliferating SKOV3/DDP cells in a 6-well plate were
transiently transfected with the pcDNA3.1-3×FLAG vector (GV141;
Shanghai Genechem, Co., Ltd.) encoding Mcl-1 or the empty vector (1
µg/µl) at 50% confluence using Lipofectamine 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. Briefly, 5 µl (1 µg/µl) Mcl-1-vector and 10 µl
Lipofectamine 3000 were diluted in 125 µl RPMI-1640 medium,
respectively, and were incubated for 5 min at room temperature.
Then, the diluted Mcl-1-vector and Lipofectamine 3000 mixture were
maintained at room temperature for 15 min. Next, the complexes were
added to each well and incubated for 6 h at 37°C. Subsequently, the
cells were washed with PBS and replaced with RPMI-1640 medium
containing 10% FBS. After 48 h of transfection, cells were treated
or untreated with 100 nM TPL for an additional 24 h and harvested
for western blot analysis.
In vivo xenograft study
The female athymic BALB/CA-u nude mice (4-6 weeks
old; 18±2 g; n=25) were purchased from Hunan SJA Laboratory Animal
Co., Ltd. and housed at the SPF-level laboratory animal center in
Nanchang University (26-28°C; 55% humidity; 10-h light/14-h dark
cycle). All mice had free access to sterilized food and water. All
animal experiments were approved by the Ethics Committee of
Nanchang University and performed in compliance with the approved
guidelines. A tumor xenograft was established in a murine model as
described in our previous study (11,13).
Briefly, single-tumor cells of SKOV3/DDP suspensions
(5×106) were subcutaneously injected into the right
armpit of each BALB/CA-u nude mice. Seven days after cell
implantation, the mice bearing tumors were randomized into five
groups (n=5/group) and initially treated as follows: i) Vehicle
group (50 ml/kg/day PBS every day, i.p.); ii) TPL group (0.15
mg/kg/day TPL every day, i.p.); iii) DDP group (4 mg/kg/day DDP on
the 1st and 8th days, i.p.); iv) TPL + DDP group (0.15 mg/kg/day
TPL every day, 4 mg/kg/day DDP on the 1st and 8th days, i.p.); and
v) TPL + DDP + CQ group (0.15 mg/kg/day TPL every day, 4 mg/kg/day
DDP on the 1st and 8th days, 25 mg/kg/day CQ every day, i.p.). All
drugs were intraperitoneally (i.p.) injected into the mice as
aforementioned (day 0 was considered as the beginning day of drug
treatment). Tumor volumes and body weight were verified every other
day. The tumor volume was calculated using the standard formula:
Volume (mm3) = width2 (mm2) ×
length (mm) ×0.5. After treatment for 10 days, all the mice were
euthanized. The tumors were harvested, weighed and subjected to
further immunohistochemical analysis as previously described
(11,13).
Statistical analysis
All experiments were performed at least three times,
and the results are presented as the mean ± standard deviation
(SD). Statistical analyses were performed using GraphPad Prism
version 6.0 software (GraphPad Software, Inc.).
Unpaired Student's t-test was employed for
comparisons of means between two groups, and one-way ANOVA with
Tukey's post hoc test were employed for the comparison of multiple
groups. The criterion for statistical significance was indicated as
P<0.05.
Results
TPL treatment triggers autophagic cell
death in SKOV3/DDP cells
TPL has been revealed to stimulate autophagy in some
cancer cells (25–28), while the effect of TPL on autophagy
regulation in drug-resistant ovarian cancer cells has not been
reported. Therefore, mRFP-GFP tandem fluorescent-tagged LC3 was
first used to track autophagic flux in SKOV3/DDP cells. As revealed
in Fig. 1A, a significant increase in
the number of autophagosomes (yellow puncta) and autolysosomes (red
puncta) were observed following TPL treatment under a confocal
microscope. Next, the expression of two specific autophagy markers,
the proteins LC3 and p62 were assessed. Western blot analysis
revealed that TPL treatment markedly increased the ratio of
LC3-II/LC3-I but reduced p62 expression in a dose-dependent manner
in SKOV3/DDP cells (Fig. 1B). These
results clearly revealed that TPL treatment enhanced autophagic
flux in ovarian cancer cells, which was consistent with previous
studies (25–28).
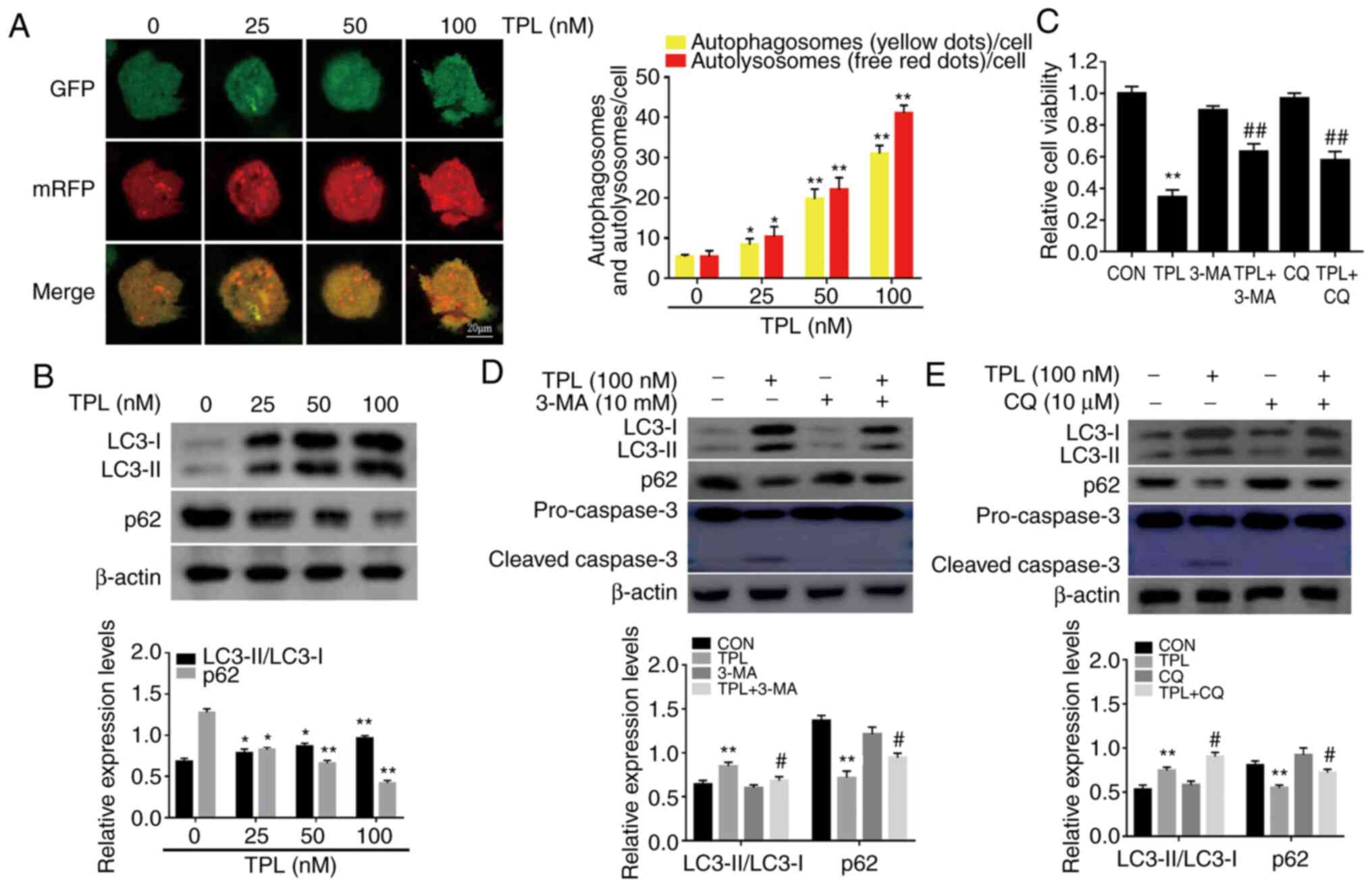 | Figure 1.TPL treatment induces autophagic cell
death in DDP-resistant ovarian cancer SKOV3/DDP cells. (A)
SKOV3/DDP cells transfected with mRFP-GFP-LC3 adenovirus were
treated with the indicated doses of TPL for 12 h, and then analyzed
using a confocal microscope (magnification, ×400). Representative
images respectively revealed the autophagosomes (yellow puncta) and
the autolysosomes (red puncta). Scale bar, 20 µm. *P<0.05 and
**P<0.01 vs. the control (untreated) group. (B) SKOV3/DDP cells
treated with indicated concentrations of TPL for 24 h were
subjected to immunoblotting for the levels of LC3 and p62. (C-E)
SKOV3/DDP cells were pre-incubated with or without autophagy
inhibitor 3-MA (10 mM) or CQ (10 µM) for 1 h, followed by 24-h TPL
(100 nM) treatment. (C) TPL cytotoxicity was detected by CCK-8
assay. (D and E) LC3, p62 and cleaved caspase-3 levels were
examined by western blot analysis. *P<0.05 and **P<0.01 vs.
the Con group. #P<0.05 and ##P<0.01 vs.
the TPL group. TPL, triptolide; DDP, cisplatin; 3-MA,
3-methyladenine; CQ, chloroquine; Con, control. |
Our group previously revealed that TPL induced
apoptosis-mediated cell death in DDP-resistant ovarian cancer cells
(9,10,12,13). Given
the dual role of autophagy in cancer cell death, widely used
autophagy inhibitors (3-MA and CQ) were applied to further confirm
whether this autophagic response mediates the cytotoxic effects of
TPL. As revealed in Fig. 1C, cell
viability was significantly restored following treatment with TPL
in the presence of 3-MA or CQ. Moreover, the expression of cleaved
caspase-3 was also significantly suppressed when TPL-induced
autophagy was inhibited by 3-MA or CQ (Fig. 1D and E). These data indicated that
autophagy contributed to TPL-induced SKOV3/DDP cell death. The
autophagic effect of TPL on parental SKOV3 cells was then
evaluated. Consistent with the effect observed in SKOV3/DDP cells,
TPL also prompted autophagy of SKOV3 cells (Fig. S1A). In addition, it was also revealed
that the cytotoxic effect of TPL on SKOV3 cells was significantly
reduced by pretreatment with 3-MA (Fig.
S1B). Thus, the data aforementioned suggested that the
autophagy induced by TPL was cytotoxic to ovarian cancer cells,
rather than cytoprotective.
TPL potentiates the antitumor effect
of DDP in vivo partially via autophagy induction
Our group previously demonstrated that TPL could
enhance the suppressive effects of DDP on tumor growth in a
SKOV3/DDP xenograft model (11,13). To
further evaluate the effects of TPL-induced autophagy on
chemosensitization in vivo, the impact of the combination of
TPL + DDP + CQ on the growth of SKOV3/DDP xenografts was
determined. Mice-bearing SKOV3/DDP cells were randomly separated
into five groups: Vehicle, TPL, DDP, TPL + DDP, and TPL + DDP + CQ.
All animals tolerated TPL treatment well. The mice did not exhibit
any obvious change in body weight (Fig.
2A). The tumor volume and tumor weight in the TPL + DDP group
were significantly reduced compared to those in the TPL + DDP + CQ
group or any other single treatment group (Fig. 2B and C). In agreement with this
observation, proliferation-related Ki-67 (the brown color area) was
greatly downregulated in the TPL + DDP group (Fig. 2D). Additionally, the
immunohistochemical staining results also revealed that autophagy
inhibition significantly reduced the chemosensitization effect of
TPL on SKOV3/DDP cells in vivo (Fig. 2D). It was concluded from these results
that the chemosensitization effect of TPL was dependent on
autophagy.
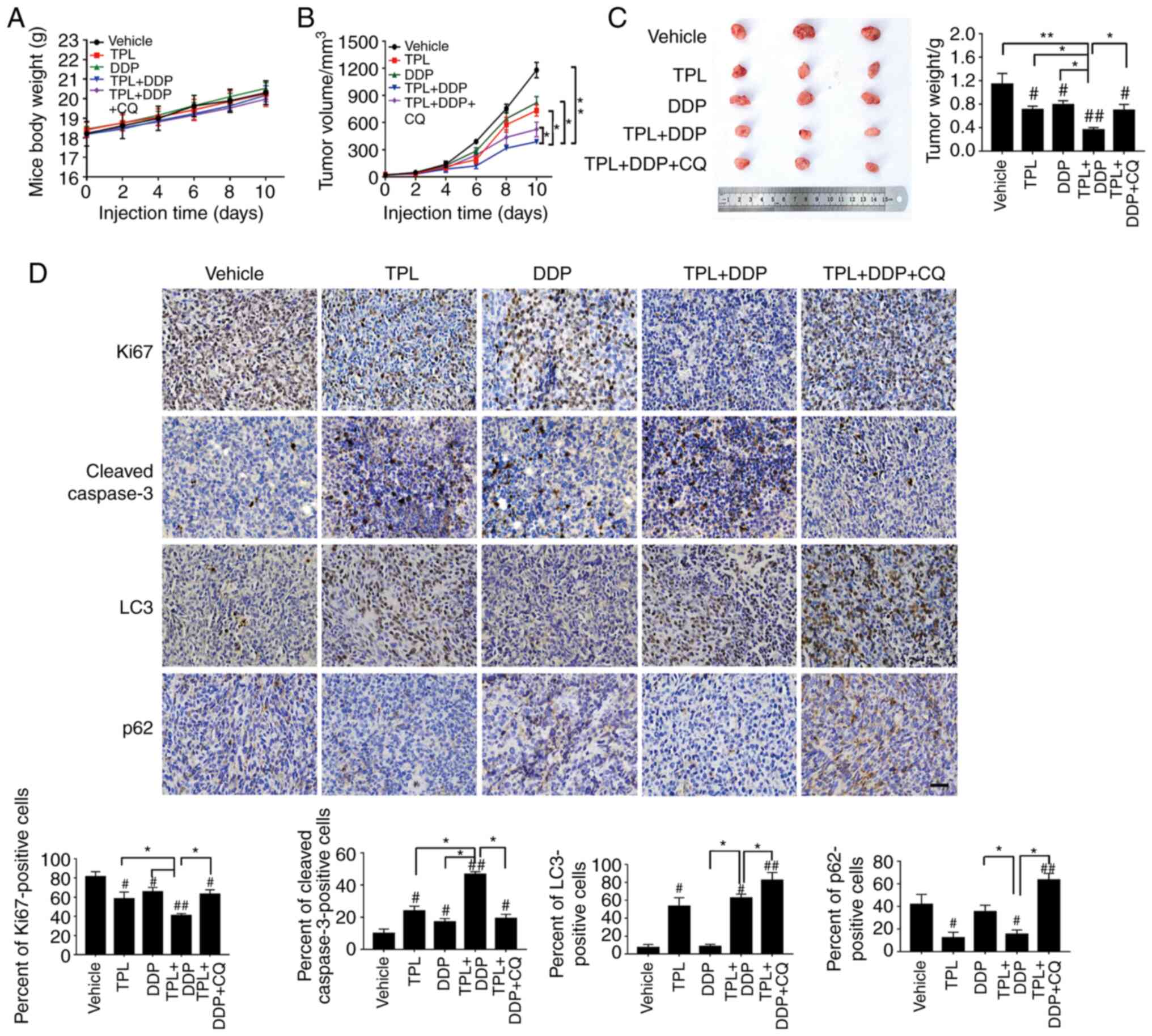 | Figure 2.TPL potentiates the antitumor effect
of DDP through autophagy induction in vivo. Mice bearing
SKOV3/DDP tumor cells were intraperitoneally injected with PBS,
TPL, DDP, TPL + DDP, and TPL + DDP + CQ. (A) The body weight of
mice and (B) tumor volume were measured every two days during the
administration period. At the end of experiment, tumors were
removed, photographed and (C) weighed. (D) Immunohistochemical
analysis (Ki67, cleaved caspase-3, LC3 and p62 staining) of tumor
tissue sections isolated from the indicated groups of mice
(magnification, ×400). Scale bar, 100 µm. *P<0.05 and
**P<0.01 vs. the TPL + DDP group. #P<0.05 and
##P<0.01 vs. the Vehicle group. TPL, triptolide; DDP,
cisplatin; CQ, chloroquine; Con, control. |
TPL-induced SKOV3/DDP cell autophagy
is closely related to intracellular ROS generation
Then, systematic research on the underlying
mechanisms involved in TPL-induced autophagy against SKOV3/DDP
cells was performed in vitro. ROS have been documented as
common mediators of cell apoptosis and autophagy (29). In a previous study by our group, it
was found that TPL induced ROS-related apoptosis in DDP-resistant
ovarian cancer cells (12).
Therefore, DCFH-DA was first used as a probe to examine the
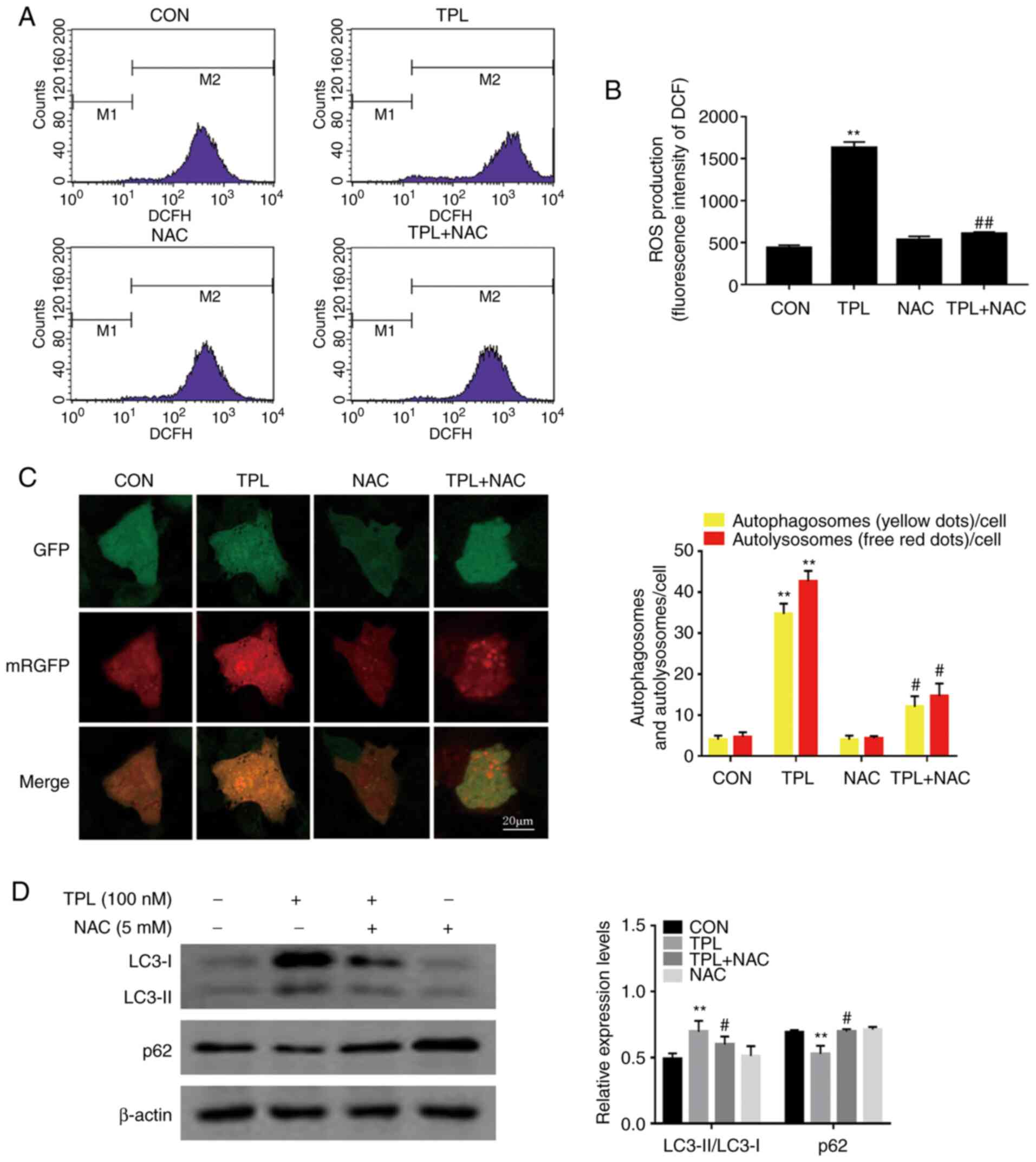
intracellular ROS levels. As revealed in Fig. 3A and B, the mean DCF fluorescence
intensity of SKOV3/DDP cells was significantly increased in
response to 100 nM of TPL treatment, as compared with the control
group. Additionally, this increase was significantly attenuated
when the cells were pretreated with the well-known antioxidant NAC
(Fig. 3A and B). These data indicated
that TPL promoted the generation of cellular ROS in SKOV3/DDP
cells. To evaluate the role of ROS in TPL-induced autophagy,
SKOV3/DDP cells were incubated with TPL in the presence or absence
of NAC. Notably, pretreatment with NAC strongly reduced the number
of autophagosomes and autolysosomes compared with TPL treatment
alone (Fig. 3C). Consistently, NAC
strongly blocked the TPL-induced upregulation of LC3-II/LC3-I
expression levels while enhancing p62 expression (Fig. 3D). Overall, these findings suggested
that the autophagic inducing effect of TPL on SKOV3/DDP cells was
associated with ROS generation.
ROS generation is upstream of
JAK2/STAT3 pathway inactivation in TPL-induced autophagy
The JAK2/STAT3 pathway is constitutively activated
in ovarian cancer (21,22), has a well-established role in
autophagy regulation and can be influenced by ROS (30). First, to evaluate the association
between activation of the JAK2/STAT3 pathway and autophagy in
SKOV3/DDP cells, AG490 (a JAK2/STAT3 signaling inhibitor) was used.
As revealed in Fig. 4A and B, the
level of autophagy significantly increased after treatment with
AG490, suggesting that inhibition of the JAK2/STAT3 pathway may
play an important role in autophagy induction.
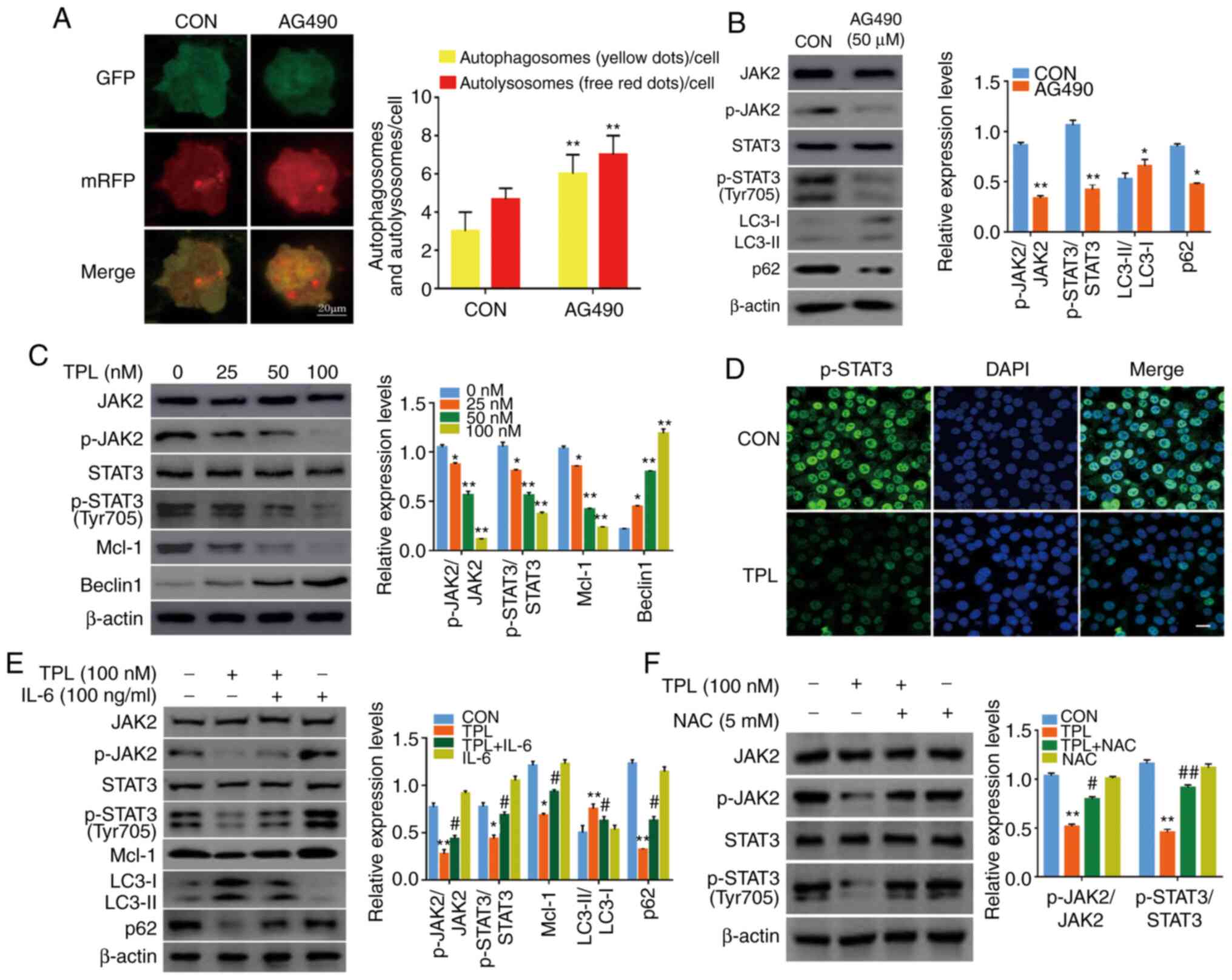 | Figure 4.ROS generation is upstream of
JAK2/STAT3 inactivation in TPL-induced autophagy. (A and B)
SKOV3/DDP cells were treated with or without 50 µM AG490 for 24 h,
and (A) the autophagosomes (yellow puncta) and the autolysosomes
(red puncta) were detected using a fluorescence microscope. Scale
bar, 20 µm. (B) The expression levels of p-JAK2, p-STAT3 (Y705),
LC3 and p62 were compared by western blot analysis. (C) Western
blotting revealed the expression of JAK2, p-JAK2, STAT3, p-STAT3
(Y705), Mcl-1 and Beclin1 in SKOV3/DDP cells after treatment with
variable doses of TPL for 24 h. (D) SKOV3/DDP cells were treated
with or without 100 nM TPL for 24 h, then the expression of p-STAT3
(Y705) was examined using immunofluorescence staining
(magnification, ×400). The cell nuclei were stained with DAPI
(blue). Scale bar, 20 µm. (E) SKOV3/DDP cells were incubated
without or with TPL (100 nM) in the presence or absence of IL-6
(100 ng/ml) for 24 h. Mcl-1, LC3 and p62 were detected by western
blot analysis. (F) SKOV3/DDP cells were treated with NAC (5 mM) for
1 h prior to exposure to TPL (100 nM) for 24 h, and then the p-JAK2
and p-STAT3 (Y705) expression were evaluated with western blotting.
*P<0.05 and **P<0.01 vs. the Con group. #P<0.05
and ##P<0.01 vs. the TPL group. ROS, reactive oxygen
species; JAK2, Janus kinase 2; STAT3, signal transducer and
activator of transcription-3; TPL, triptolide; DDP, cisplatin;
p-phosphorylated; Mcl-1, myeloid cell leukemia-1; IL, interleukin;
NAC, N-acetyl-l-cysteine; Con, Control. |
The effect of TPL treatment on the JAK2/STAT3
signaling pathway was next examined. Western blot analysis revealed
that TPL treatment led to noticeable inhibition of p-JAK2 and
p-STAT3 (Y705) (Fig. 4C).
Additionally, immunofluorescence analysis revealed that TPL
incubation significantly decreased the nuclear level of p-STAT3
(Y705) (Fig. 4D). These results
indicated that TPL treatment inhibited the JAK2/STAT3 pathway in
SKOV3/DDP cells.
To further reveal the importance of the JAK2/STAT3
pathway in modulating TPL-induced autophagy, SKOV3/DDP cells were
pretreated with IL-6 (a JAK2/STAT3 signaling activator). It was
revealed that IL-6 strongly attenuated the LC3 conversion and p62
degradation accompanied by the upregulation of p-JAK2 and p-STAT3
(Y705) compared to those of the TPL group (Fig. 4E). These data indicated that TPL
triggered autophagy in SKOV3/DDP cells at least partially by
modulating JAK2/STAT3 signaling. Notably, the TPL-induced
reductions in p-JAK2 and p-STAT3 (Y705) could be restored in the
presence of NAC (Fig. 4F). Thus,
ROS-mediated inhibition of the JAK2/STAT3 signaling pathway was
involved in TPL-induced autophagy.
JAK2/STAT3 pathway inactivation
contributes to TPL-induced autophagy by regulating the
Beclin1/Mcl-1 interaction
Next, the downstream signaling molecules of STAT3
involved in the regulation of TPL-induced autophagy were
investigated. It has been reported that Mcl-1, a well-known
antiapoptotic protein that is transcriptionally regulated by STAT3,
negatively regulates autophagy by binding to Beclin1 (24,31).
Therefore, the protein levels of Mcl-1 and Beclin1 were analyzed
and it was revealed that TPL efficiently decreased Mcl-1 levels and
increased Beclin1 protein levels in a dose-dependent manner
(Fig. 4C). Next, an
immunoprecipitation assay was conducted to monitor the interaction
between Beclin1 and Mcl-1 in SKOV3/DDP cells. It was revealed that
Mcl-1 and Beclin1 immunoprecipitated with each other in SKOV3/DDP
cells under basal conditions (Fig.
5A), whereas the interaction markedly decreased in the presence
of TPL (Fig. 5B).
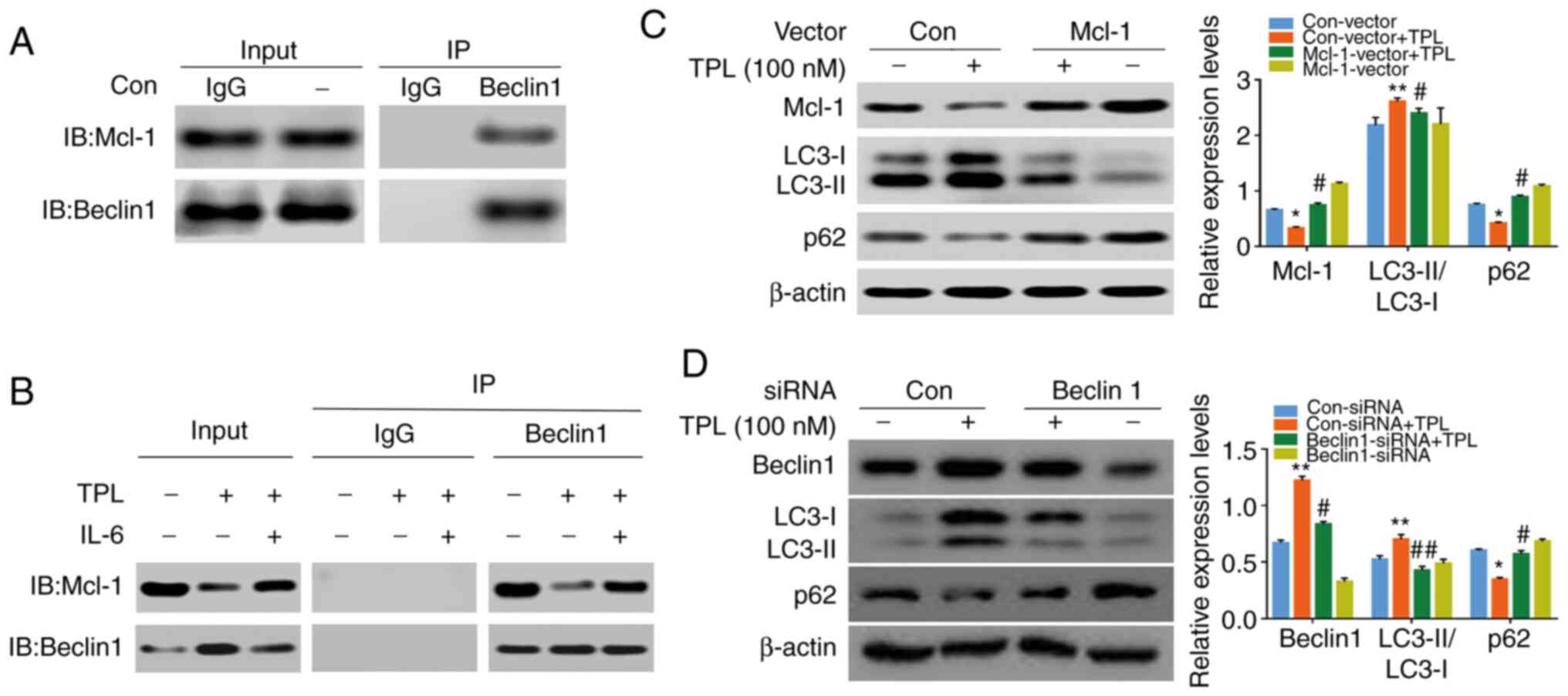 | Figure 5.JAK2/STAT3 inactivation contributes
to TPL-induced autophagy via regulation of Beclin1/Mcl-1
interaction. (A) Coimmunoprecipitation analysis of Beclin1 and
Mcl-1 in SKOV3/DDP cells. (B) SKOV3/DDP cells were treated with TPL
(100 nM) in the absence or presence of IL-6 (100 ng/ml) for 24 h.
Cell lysates were subjected to immunoprecipitation with
anti-Beclin1 antibody and interaction with Mcl-1 was determined by
western blotting. (C) SKOV3/DDP cells transfected with Mcl-1
overexpression plasmid were treated with or without TPL (100 nM)
for 24 h, followed by western blot analysis to detect the levels of
Mcl-1, LC3 and p62. (D) SKOV3/DDP cells transfected with the
Beclin1 siRNA or negative control siRNA were left cultured with or
without TPL (100 nM) for 24 h, and then the amount of Beclin1, LC3
and p62 were analyzed by western blotting. *P<0.05 and
**P<0.01 vs. the Con-vector group or Con-siRNA group.
#P<0.05 and ##P<0.01 vs. the Con-vector
+ TPL group or Con-siRNA + TPL group. JAK2, Janus kinase 2; STAT3,
signal transducer and activator of transcription-3; TPL,
triptolide; Mcl-1, myeloid cell leukemia-1; DDP, cisplatin; IL,
interleukin; Con, Control. |
Notably, IL-6 significantly prevented TPL-mediated
Mcl-1 downregulation (Fig. 4E) and
the Beclin1/Mcl-1 interaction (Fig.
5B). Furthermore, transient overexpression of Mcl-1 (Fig. 5C) and siRNA-mediated Beclin1
downregulation (Fig. 5D)
significantly attenuated LC3-II accumulation and p62 degradation
induced by TPL. Collectively, these results indicated that
inhibition of the JAK2/STAT3/Mcl-1 pathway and the subsequent
disruption of the Beclin1/Mcl-1 interaction are vital to
TPL-induced autophagy.
Discussion
Recent studies have revealed that numerous
anticancer drugs, including TPL, can concurrently activate
autophagy, as they induce apoptotic cell death in tumor cells
(32–34). Over the last several years, our prior
studies have demonstrated the antitumor and sensitization
properties of TPL against epithelial ovarian cancer in vitro
and in vivo, and apoptosis has been revealed to mediate this
process (9–13). However, the exact role of TPL-induced
autophagy in DDP-resistant human epithelial ovarian cancer cells
remains unclear. Thus, DDP-resistant human ovarian cancer SKOV3/DDP
cells were used as an experimental model to address this issue. The
present study demonstrated for the first time, to the best of our
knowledge, that TPL induced lethal autophagy in SKOV3/DDP cells via
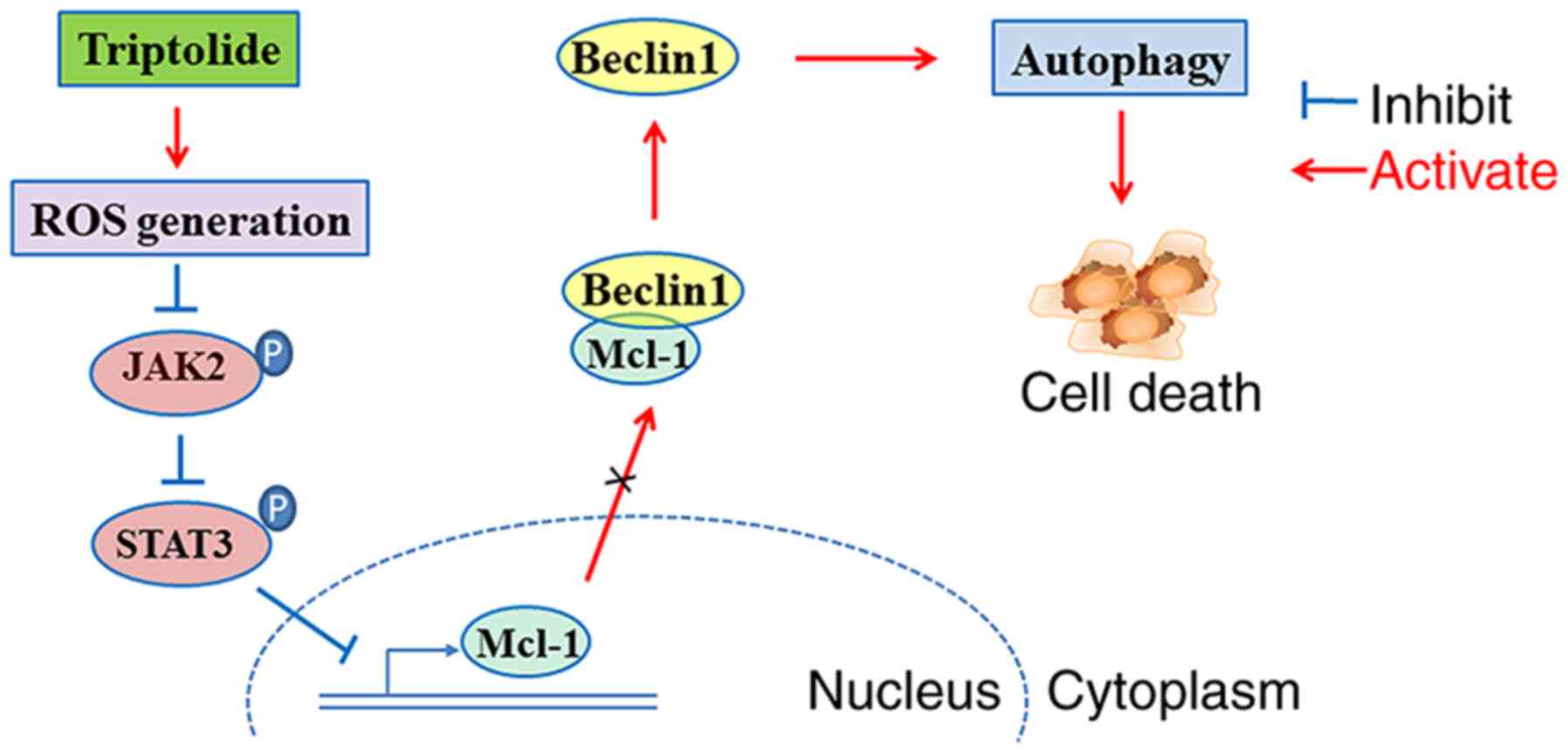
the ROS/JAK2/STAT3 pathway (Fig.
6).
Autophagy and apoptosis are distinct cellular
processes that are closely interrelated (35). In the present study, the data clearly
revealed that TPL strongly elicited typical autophagic features in
cells, as evidenced by the altered expression of autophagic
proteins, such as LC3-II, p62 and Beclin1, and the increased
numbers of autophagosomes and autolysosomes. Usually, normal cells
have a low basal level of autophagy to maintain homeostasis and
suppress tumor formation (36). On
the other hand, a certain basal autophagic flux is necessary for
highly-proliferating tumor cells to ensure metabolic requirements
(37). Following chemotherapy, the
basally-enhanced autophagic flux is generally regarded as an
adaptive survival response that mediates tumor chemoresistance
(37). However, extended autophagic
flux above the optimal survival limit may have cytotoxic effects on
cancer cells and ultimately facilitate cell death (38,39).
Clearly, inducing autophagic cell death could potentially be used
as a novel strategy to manage resistant tumors (40). Further in vitro experiments
revealed that pretreatment with the autophagy inhibitors 3-MA or CQ
significantly abolished the inhibitory effect of TPL on cell
proliferation. Consistently, it was observed that TPL treatment
effectively enhanced the growth-inhibiting effect of DDP by
enhancing autophagic activity in the SKOV3/DDP xenograft model.
These data strongly indicated that enhanced autophagy may operate
as a death mechanism in TPL-treated DDP-resistant SKOV3/DDP cells.
It is highly likely that the extent of autophagic flux induced by
TPL in SKOV3/DDP cells exceeds the lethal autophagic flux
threshold. These observations were consistent with those of
previous studies showing that TPL induced autophagy to induce
apoptosis and inhibit angiogenesis in human osteosarcoma MG63 cells
(26), and neuroblastoma SH-SY5Y
cells (25). Some studies have
reported that protective autophagy is enhanced by TPL in malignant
glioma U251 cells (27) and cervical
cancer SiHa cells (28). Accumulating
evidence suggests that the level of basal autophagic flux and
lethal autophagic flux threshold, which play a crucial role in
governing cell fate, may vary according to the different tumor
tissue cell types (37). These
disparate effects may be, in part, attributed to the differences in
tumor types and lack of a standard test method for autophagic flux
threshold (37,38). Future studies will be required to
delineate these questions.
Although TPL has been demonstrated to exert potent
autophagic-regulating effects via different mechanisms (14), the exact molecular mechanism remains
poorly understood. Previously, our group demonstrated that TPL
triggered the ROS-dependent activation of apoptotic cell death in
DDP-resistant ovarian cancer cells (12). ROS are highly reactive oxygen-derived
molecules that influence cell survival, autophagy and apoptosis
through redox signaling (29).
Usually, cancer cells exhibit increased ROS generation that may
promote cell survival and drug resistance (41). However, an excessively high level of
ROS may cause excessive oxidative stress and lead to cell death
through apoptosis, autophagy and necrosis (42). Novel anticancer agents have been
developed to generate ROS and trigger oxidative stress-induced
tumor cell death (42,43). This finding suggests that ROS may play
an important role in TPL-mediated autophagy induction in SKOV3/DDP
cells. Consistent with our hypothesis, pretreatment with the
antioxidant NAC substantially inhibited ROS generation and rescued
TPL-induced autophagy, as supported by our results revealing that
NAC markedly blocked TPL-induced autophagosome/autolysosome
formation, LC3 conversion and p62 degradation in SKOV3/DDP cells.
These data were consistent with those of previous studies which
revealed that costunolide and cinobufagin induced autophagy in
cancer cells through ROS production (33,44).
The JAK2/STAT3 pathway is one of the well-known
pathways associated with ovarian tumor growth and chemoresistance
(21,22). Accumulating evidence indicates that
STAT3 is an important regulator of autophagy (20). Upon stimulation, STAT3 is activated
and phosphorylated, and then STAT3 transfers into the nucleus to
regulate the transcription of various genes, including certain
autophagy-associated genes (20).
Consistently, the present results revealed that p-JAK2 and p-STAT3
(Y705) were overexpressed in SKOV3/DDP cells. Notably, treatment
with AG490 strongly enhanced the level of autophagy in SKOV3/DDP
cells, suggesting that inhibition of the JAK2/STAT3 pathway is
involved in the regulation of autophagy. It is intriguing that
activated nuclear STAT3 can upregulate Mcl-1 expression, which
leads to autophagy inhibition (45).
A previous study by Tai et al (24) revealed that sorafenib downregulated
STAT3 activity and Mcl-1 expression and induced autophagy in
multiple HCC cell lines. In contrast, Mcl-1 overexpression
abrogated the effect of sorafenib on autophagy. Furthermore, Mcl-1
has been reported to negatively regulate the autophagy induction
through binding to Beclin1. In accordance with these findings, the
present results revealed that TPL suppressed the JAK2/STAT3
signaling pathway and further downregulated Mcl-1, along with the
subsequent reduction in the Beclin1/Mcl-1 interaction. These
inhibitory effects of TPL could be strongly reversed by the
JAK2/STAT3 signaling activator IL-6. Moreover, ectopic expression
of Mcl-1 or genetic knockdown of Beclin1 impaired the
autophagy-inducing effect of TPL. Thus, it is reasonable to
conclude that TPL triggered Beclin1-dependent autophagy in
SKOV3/DDP cells through inhibition of the JAK2/STAT3/Mcl-1
signaling cascade and disruption of the Beclin1/Mcl-1 interaction.
Notably, it was further revealed that the generation of ROS by TPL
precedes the inhibition of the JAK2/STAT3 pathway. It is well
documented that ROS influence the enzymatic activity of JAK2 by
oxidative modification of specific cysteine residues on JAK2
(30,46). The mechanism by which ROS are involved
in the regulation of the JAK2/STAT3 pathway requires further
study.
Collectively, it was proposed that TPL induced ROS
generation to suppress the JAK2/STAT3 signaling cascade, causing a
decline in Mcl-1 expression and alleviating Mcl-1-mediated
inhibition of Beclin1, consequently triggering autophagic cell
death in SKOV3/DDP cells (Fig. 6). In
addition, TPL enhanced the sensitivity of SKOV3/DDP cells to DDP by
increasing the autophagic response in vivo. The present
results not only deepen the understanding of the molecular
mechanism of TPL-mediated antitumor effects but also support the
use of TPL as a therapeutic option for patients with resistant
ovarian tumors in the future.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81760729), the
Project of the Jiangxi Provincial Natural Science Foundation in
China (grant no. 20202BABL206102), the Project of the Jiangxi
Provincial Health Commission (grant no. 20204271), the Youth
Science Foundation of Science and Technology Program of the Second
Affiliated Hospital of Nanchang University (grant no.
2019YNQN12001).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Author's contributions
BT and YZ conceived and designed the present study.
FL, YZ, JC, CL, XZ, XW, FX, QZ performed the experiments. YZ, CL,
FL, XW, JC, XZ, FX, QZ and BT performed the data acquisition and
analysis. YZ wrote the manuscript. YZ, CL, XW, FL and BT revised
the work critically for important intellectual content. All the
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were approved by the Ethics
Committee of Nanchang University (Nanchang, China) and performed in
compliance with the approved guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
TPL
|
triptolide
|
|
DDP
|
cisplatin
|
|
3-MA
|
3-methyladenine
|
|
CQ
|
chloroquine
|
|
NAC
|
N-acetyl-l-cysteine
|
|
ROS
|
reactive oxygen species
|
|
JAKs
|
Janus family kinases
|
|
STAT3
|
signal transducer and activator of
transcription-3
|
|
Mcl-1
|
myeloid cell leukemia-1
|
|
FBS
|
fetal bovine serum
|
|
CCK-8
|
Cell Count Kit-8
|
|
OD
|
optical density
|
|
SDS-PAGE
|
sodium dodecyl sulfate-polyacrylamide
gel electrophoresis
|
|
PVDF
|
polyvinylidene difluoride
|
|
IP
|
immunoprecipitation
|
|
SPF
|
Specific-Pathogen-Free
|
|
SD
|
standard deviation
|
References
|
1
|
Lheureux S, Gourley C, Vergote I and Oza
AM: Epithelial ovarian cancer. Lancet. 393:1240–1253. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Orr B and Edwards RP: Diagnosis and
treatment of ovarian cancer. Hematol Oncol Clin North Am.
32:943–964. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Armbruster S, Coleman RL and Rauh-Hain JA:
Management and treatment of recurrent epithelial ovarian cancer.
Hematol Oncol Clin North Am. 32:965–982. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pignata S, Pisano C, Di Napoli M, Cecere
SC, Tambaro R and Attademo L: Treatment of recurrent epithelial
ovarian cancer. Cancer. 125 (Suppl 24):S4609–S4615. 2019.
View Article : Google Scholar
|
|
5
|
Wang H, Ma D, Wang C, Zhao S and Liu C:
Triptolide inhibits invasion and tumorigenesis of hepatocellular
carcinoma MHCC-97H cells through NF-κB Signaling. Med Sci Monit.
22:1827–1836. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kim ST, Kim SY, Lee J, Kim K, Park SH,
Park YS, Lim HY, Kang WK and Park JO: Triptolide as a novel agent
in pancreatic cancer: The validation using patient derived
pancreatic tumor cell line. BMC Cancer. 18:11032018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang T, Ding Y, Yang Y, Wang Z, Gao W, Li
D, Wei J and Sun Y: Synergistic antitumour effects of triptolide
plus 10-hydroxycamptothecin onbladder cancer. Biomed Pharmacother.
115:1088992019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Liu H, Tang L, Li X and Li H: Triptolide
inhibits vascular endothelial growth factor-mediated angiogenesis
in human breast cancer cells. Exp Ther Med. 16:830–836.
2018.PubMed/NCBI
|
|
9
|
Hu H, Luo L, Liu F, Zou D, Zhu S, Tan B
and Chen T: Anti-cancer and sensibilisation effect of triptolide on
human epithelial ovarian cancer. J Cancer. 7:2093–2099. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hu H, Huang G, Wang H, Li X, Wang X, Feng
Y, Tan B and Chen T: Inhibition effect of triptolide on human
epithelial ovarian cancer via adjusting cellular immunity and
angiogenesis. Oncol Rep. 39:1191–1196. 2018.PubMed/NCBI
|
|
11
|
Huang G, Hu H, Zhang Y, Zhu Y, Liu J, Tan
B and Chen T: Triptolide sensitizes cisplatin-resistant human
epithelial ovarian cancer by inhibiting the phosphorylation of AKT.
J Cancer. 10:3012–3020. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhong YY, Chen HP, Tan BZ, Yu HH and Huang
XS: Triptolide avoids cisplatin resistance and induces apoptosis
via the reactive oxygen species/nuclear factor-κB pathway in
SKOV3PT platinum-resistant human ovarian cancer cells.
Oncol Lett. 6:1084–1092. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hu H, Zhu S, Tong Y, Huang G, Tan B and
Yang L: Antitumor activity of triptolide in SKOV3 cells and
SKOV3/DDP in vivo and in vitro. Anticancer Drugs. 31:483–491. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wei YM, Wang YH, Xue HQ, Luan ZH, Liu BW
and Ren JH: Triptolide, A potential autophagy modulator. Chin J
Integr Med. 25:233–240. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kocaturk NM, Akkoc Y, Kig C, Bayraktar O,
Gozuacik D and Kutlu O: Autophagy as a molecular target for cancer
treatment. Eur J Pharm Sci. 134:116–137. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Poillet-Perez L and White E: Role of tumor
and host autophagy in cancer metabolism. Genes Dev. 33:610–619.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Denton D and Kumar S: Autophagy-dependent
cell death. Cell Death Differ. 26:605–616. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pietrocola F, Izzo V, Niso-Santano M,
Vacchelli E, Galluzzi L, Maiuri MC and Kroemer G: Regulation of
autophagy by stress-responsive transcription factors. Semin Cancer
Biol. 23:310–322. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hao D, Wen X, Liu L, Wang L, Zhou X, Li Y,
Zeng X, He G and Jiang X: Sanshool improves UVB-induced skin
photodamage by targeting JAK2/STAT3-dependent autophagy. Cell Death
Dis. 10:192019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
You L, Wang Z, Li H, Shou J, Jing Z, Xie
J, Sui X, Pan H and Han W: The role of STAT3 in autophagy.
Autophagy. 11:729–739. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yoshikawa T, Miyamoto M, Aoyama T, Soyama
H, Goto T, Hirata J, Suzuki A, Nagaoka I, Tsuda H, Furuya K and
Takano M: JAK2/STAT3 pathway as a therapeutic target in ovarian
cancers. Oncol Lett. 15:5772–5780. 2018.PubMed/NCBI
|
|
22
|
Gritsina G, Xiao F, O'Brien SW, Gabbasov
R, Maglaty MA, Xu RH, Thapa RJ, Zhou Y, Nicolas E, Litwin S, et al:
Targeted blockade of JAK/STAT3 signaling inhibits ovarian carcinoma
growth. Mol Cancer Ther. 14:1035–1047. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zheng X, Chen W, Hou H, Li J, Li H, Sun X,
Zhao L and Li X: Ginsenoside 20(S)-Rg3 induced autophagy to inhibit
migration and invasion of ovarian cancer. Biomed Pharmacother.
85:620–626. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tai WT, Shiau CW, Chen HL, Liu CY, Lin CS,
Cheng AL, Chen PJ and Chen KF: Mcl-1-dependent activation of Beclin
1 mediates autophagic cell death induced by sorafenib and SC-59 in
hepatocellular carcinoma cells. Cell Death Dis. 4:e4852013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Krosch TC, Sangwan V, Banerjee S, Mujumdar
N, Dudeja V, Saluja AK and Vickers SM: Triptolide-mediated cell
death in neuroblastoma occurs by both apoptosis and autophagy
pathways and results in inhibition of nuclear factor-kappa B
activity. Am J Surg. 205:387–396. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Li X, Lu Q, Xie W, Wang Y and Wang G:
Anti-tumor effects of triptolide on angiogenesis and cell apoptosis
in osteosarcoma cells by inducing autophagy via repressing
Wnt/β-Catenin signaling. Biochem Biophys Res Commun. 496:443–449.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu X, Zhao P, Wang X, Wang L, Zhu Y and
Gao W: Triptolide induces glioma cell autophagy and apoptosis via
upregulating the ROS/JNK and downregulating the Akt/mTOR signaling
pathways. Front Oncol. 9:3872019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Qin G, Li P and Xue Z: Triptolide induces
protective autophagy and apoptosis in human cervical cancer cells
by downregulating Akt/mTOR activation. Oncol Lett. 16:3929–3934.
2018.PubMed/NCBI
|
|
29
|
Kaminskyy VO and Zhivotovsky B: Free
radicals in cross talk between autophagy and apoptosis. Antioxid
Redox Signal. 21:86–102. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cao Y, Wang J, Tian H and Fu GH:
Mitochondrial ROS accumulation inhibiting JAK2/STAT3 pathway is a
critical modulator of CYT997-induced autophagy and apoptosis in
gastric cancer. J Exp Clin Cancer Res. 39:1192020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Germain M, Nguyen AP, Le Grand JN, Arbour
N, Vanderluit JL, Park DS, Opferman JT and Slack RS: MCL-1 is a
stress sensor that regulates autophagy in a developmentally
regulated manner. EMBO J. 30:395–407. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gali-Muhtasib H, Hmadi R, Kareh M, Tohme R
and Darwiche N: Cell death mechanisms of plant-derived anticancer
drugs: Beyond apoptosis. Apoptosis. 20:1531–1562. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ma K, Zhang C, Huang MY, Li WY and Hu GQ:
Cinobufagin induces autophagy-mediated cell death in human
osteosarcoma U2OS cells through the ROS/JNK/p38 signaling pathway.
Oncol Rep. 36:90–98. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chang CH, Lee CY, Lu CC, Tsai FJ, Hsu YM,
Tsao JW, Juan YN, Chiu HY, Yang JS and Wang CC: Resveratrol-induced
autophagy and apoptosis in cisplatin-resistant human oral cancer
CAR cells: A key role of AMPK and Akt/mTOR signaling. Int J Oncol.
50:873–882. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cooper KF: Till death do us part: The
marriage of autophagy and apoptosis. Oxid Med Cell Longev.
2018:47012752018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fitzwalter BE and Thorburn A: Recent
insights into cell death and autophagy. FEBS J. 282:4279–4288.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bhat P, Kriel J, Shubha Priya B, Basappa,
Shivananju NS and Loos B: Modulating autophagy in cancer therapy:
Advancements and challenges for cancer cell death sensitization.
Biochem Pharmacol. 147:170–182. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kriel J and Loos B: The good, the bad and
the autophagosome: Exploring unanswered questions of
autophagy-dependent cell death. Cell Death Differ. 26:640–652.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fulda S and Kögel D: Cell death by
autophagy: Emerging molecular mechanisms and implications for
cancer therapy. Oncogene. 34:5105–5113. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cirone M, Gilardini Montani MS, Granato M,
Garufi A, Faggioni A and D'Orazi G: Autophagy manipulation as a
strategy for efficient anticancer therapies: Possible consequences.
J Exp Clin Cancer Res. 38:2622019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Galadari S, Rahman A, Pallichankandy S and
Thayyullathil F: Reactive oxygen species and cancer paradox: To
promote or to suppress? Free Radic Biol Med. 104:144–164. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zou Z, Chang H, Li H and Wang S: Induction
of reactive oxygen species: An emerging approach for cancer
therapy. Apoptosis. 22:1321–1335. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Gao L, Loveless J, Shay C and Teng Y:
Targeting ROS-Mediated crosstalk between autophagy and apoptosis in
cancer. Adv Exp Med Biol. 1260:1–12. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fang Y, Li J, Wu Y, Gui J and Shen Y:
Costunolide inhibits the growth of OAW42-A multidrug-resistant
human ovarian cancer cells by activating apoptotic and autophagic
pathways, production of reactive oxygen species (ROS), cleaved
caspase-3 and cleaved caspase-9. Med Sci Monit. 25:3231–3237. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ma Z, Bao X and Gu J: Furowanin A-induced
autophagy alleviates apoptosis and promotes cell cycle arrest via
inactivation STAT3/Mcl-1 axis in colorectal cancer. Life Sci.
218:47–57. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sun Q, Lu NN and Feng L: Apigetrin
inhibits gastric cancer progression through inducing apoptosis and
regulating ROS-modulated STAT3/JAK2 pathway. Biochem Biophys Res
Commun. 498:164–170. 2018. View Article : Google Scholar : PubMed/NCBI
|