Introduction
Breast cancer (BC) affects 2.3 million women per
year and is responsible for the highest number of cancer-related
deaths among women globally (1) BC
contributes to 30% of female cancers in the USA and it is estimated
that 276,480 new cases of BC and 42,170 deaths will be attributed
to BC for the year 2020 (2).
Since BC is a multifactorial disease, incidence,
survival and mortality rates differ across countries. The latter is
due to the involvement of a multitude of modifiable and
non-modifiable risk factors such as ethnicity, mutations in the
BRCA1/2 genes, diet, alcohol, sedentary lifestyle, radiation
and lifetime exposure to estrogens (3–5). For
example, the 5-year survival rate for BC is 90% in the USA
(2) whereas in certain underdeveloped
countries the 5-year survival rate can be as low as 57% (6). Incidence and survival rates not only
differ between different countries, but also vary among the
different types of BC. The best survival rates are seen in women
with the subtype estrogen receptor (ER)+ and/or
progesterone (PR)+ and HER-2, while the worst survival
rates are observed in women with the triple-negative subtype
(ER− PR−, HER-2−) (7).
The microbiota found in our bodies is estimated to
outnumber our cells by a factor of 10 and it is considered crucial
for the correct functioning of our physiology, mostly due to the
production of metabolites by the microbes (8). Humans have therefore evolved to be
dependent on these bacteria and have cultivated a fruitful
symbiotic relationship between them. Resulting from this, the Human
Microbiome Project was created to further document the effect that
the microbial genomes have on our physiology (8). Initially, this project aimed to
investigate the microbiota of only the skin, mouth, nose, colon and
vagina and to examine how the bacteria can manipulate the function
of the human body to shift from a healthy state to a diseased one.
Interestingly, body sites that have been previously considered as
sterile such as the pancreas, the prostate, the lungs and the
breast have been found to harvest unique microbial environments
(9). Given the fact that the breast
has a naturally occurring nutrient rich and fatty environment and
knowing that bacteria found in the skin have direct access to the
mammary ducts through the nipple, it is not surprising that the
breast contains a vast array of bacteria in its tissues (10).
When in a healthy state, the microbiota and our body
exist in perfect balance i.e. symbiosis. However, when this
delicate balance is disrupted, a microbial imbalance develops known
as dysbiosis, potentially leading to numerous malignancies such as
colorectal, skin, liver, stomach, and lung cancers (4,11). In
fact, studies suggest that the microbial flora is responsible for
at least 18% of all malignancies worldwide (4). In the past few years, an increasingly
well-established relationship has been recognised between gut
microbiota and colorectal cancer (11). The mechanisms implicated in the
carcinogenesis depends vastly on the different types of microbiota
involved. Some bacteria alter immunological functions while others
synthesize genotoxins or change the regulation of circulating
steroid hormones (10).
Since BC is linked to many modifiable risk factors
and affects many women worldwide, it has become a growing priority
to identify new biomarkers to facilitate the screening and
management of BC. Recently, emerging evidence in the literature
supports that alterations in the microbiota of the mammary and gut
tissue are associated with the development of BC (10). It is therefore important for more
research to be carried out to identify the bacteria and molecular
mechanisms involved in breast carcinogenesis. The aim of the
current review is to provide recent evidence regarding the role of
the mammary and gut microbiota in the development, prevention and
management of BC.
Microbiota in normal vs. malignant breast
tissues
The highly diverse microbiota in
normal breast tissue
Research in the past few years has provided evidence
that the breast tissue harbors a diverse and unique community of
microbes (4). There are many theories
regarding the origins of the breast microbiota, ranging from
translocation of microbiota from the skin through the nipple, to
the translocation of microbiota during lactation (4). Whatever the origin, the breast tissue
contains a high diversity of bacteria. Using Shannon's diversity
index (SDI), an index used to characterize species diversity,
studies have concluded that the mammary microbiota has an average
SDI of 3.6 (12). This is a
relatively high diversity since the gut and oral cavity, which are
considered to have a variety of bacterial communities, have an SDI
of 3.9 and 6.5 respectively, while the vagina which is known to
have a low diversity of bacteria has an SDI between 0.46 and 2.9
(10).
The knowledge that breast tissue has its own unique
microbiome has sparked interest as to its involvement in the
development of BC (4). Even though
this area of research has yet to be fully explored, there have been
various studies that have proved to be fruitful.
A study conducted by Urbaniak et al (10) was conducted to investigate if healthy
mammary tissues from women in Canada and Ireland contained their
own microbiome. In this study, breast tissue samples were collected
from women undergoing breast surgery to either remove a cancerous
or benign tumor or to have breast reductions. Interestingly, the
results showed that the breast tissues have their own unique
microbiome. By using 16S rRNA sequencing and culture, the
researchers identified various bacteria and found the most abundant
phyla in normal breast tissues to be Proteobacteria,
Firmicutes and Actinobacteria (10) (Table I).
Interestingly, while Proteobacteria were seen in abundance
in breast tissue, they are normally known to be a minority in other
body sites, such as the vagina, bladder and GI tract. This could be
because Proteobacteria have adapted to the fatty acid
environment of the breast tissue (10). More specifically, the Canadian samples
showed that the most abundant taxa were from the genera
Bacillus, Acinetobacter Enterobacteriaceae, Pseudomonas,
Staphylococcus, Propionibacterium, Comamonadaceae,
Gammaproteobacteria and Prevotella (Table I). On the other hand, the Irish
samples demonstrated that the most abundant taxa were
Enterobacteriaceae, Staphylococcus Listeria welshimeri,
Propionibacterium and Pseudomonas (10) (Table
I).
 | Table I.Summary of the main bacteria found in
healthy breast tissue, non-cancerous adjacent tissue and breast
cancer (BC) tissue. |
Table I.
Summary of the main bacteria found in
healthy breast tissue, non-cancerous adjacent tissue and breast
cancer (BC) tissue.
| Main bacteria in
normal breast tissue | Study
participants | Authors
(Refs.) |
|---|
| Most abundant
Phyla: | Total number of
participants: 81 women | Urbaniak et
al (10) |
|
Proteobacteria | Canadian
participants: 43 women |
|
|
Firmicutes | • 38 underwent
resection for benign |
|
|
Actinobacteria | (n=11)
or cancerous tumors (n=27) |
|
| Most abundant Taxa
in Canadian samples: | • 5 women had no
history of BC and |
|
|
Bacillus (Phylum:
Firmicutes) (11.4%) |
underwent breast reduction
surgery |
|
|
Acinectobacter (Phylum:
Proteobacteria) (10%) | Irish participants:
38 women |
|
|
Enterobacteriaceae
(Phylum: Proteobacteria) (8.3%) | • 33 underwent
lumpectomies or |
|
|
Pseudomonas (Phylum:
Proteobacteria) (6.5%) |
mastectomies for cancerous
tumors |
|
|
Staphylococcus (Phylum:
Firmicutes) (6.5%) | • 5 had no history
of BC and |
|
|
Propionibacterium
(Phylum: Actinobacteria) (5.8%) |
underwent breast reduction
surgery |
|
|
Comamonadaceae (Phylum:
Proteobacteria) (5.7%) |
|
|
|
Gammaproteobacteria
(Phylum: Proteobacteria) (5.0%) |
|
|
|
Prevotella (Phylum:
Bacteroidetes) (5.0%) |
|
|
| Most abundant Taxa
in Irish samples: |
|
|
|
Enterobacteriaceae
(Phylum: Proteobacteria) (30.8%) |
|
|
|
Staphylococcus (Phylum:
Firmicutes) (12.7%) |
|
|
|
Listeria welshimeri
(Phylum: Firmicutes) (12.1%) |
|
|
|
Propionibacterium
(Phylum: Actinobacteria) (10.1%) |
|
|
|
Pseudomonas (Phylum:
Proteobacteria) (5.3%) |
|
|
|
| Bacteria in
healthy breast tissue or non-cancerous adjacent tissue or tumor
tissue of BC patients | Study
participants | Authors
(Refs.) |
|
| Increased levels of
bacteria in breast tissue of healthy | Total number of
participants: 81 women | Urbaniak et
al (14) |
| women: | Tissue samples were
obtained from: |
|
|
Lactococcus (Phylum:
Firmicutes) | • 58 women who had
lumpectomies or |
|
|
Streptococcus (Phylum:
Firmicutes) |
mastectomies for benign (n=13)
or |
|
|
Prevotella (Phylum:
Bacteroidetes) |
cancerous (n=45) tumors |
|
| Increased levels of
bacteria in non-cancerous | • 23 women who
underwent breast |
|
| adjacent tissue and
tumor tissue in BC patients: |
reductions or breast
enhancement |
|
|
Bacillus (Phylum:
Firmicutes) |
surgeries (healthy
control) |
|
|
Enterobacteriaceae
(Phylum: Proteobacteria) |
|
|
| (e.g.,
Escherichia coli) |
|
|
|
Staphylococcus (Phylum:
Firmicutes) |
|
|
| Increased levels in
nipple aspirate fluid (NAF) of | Total number of
participants: 48 women | Chan et al
(15) |
| healthy women: | undergoing skin
sampling |
|
|
Sphingomonadaceae
(Phylum: Proteobacteria) | Tissue samples were
obtained from: |
|
| Increased levels of
nipple aspirate fluid (NAF) of | • 25 women with a
history of ductal breast |
|
| BC survivors: |
carcinoma with at least one
intact nipple |
|
|
Alistipes (Phylum:
Bacteroidetes) |
(metastatic breast cancer
patients were excluded) |
|
|
| • 23 healthy
control |
|
| Bacteria found at
increased levels in malignant vs. | Total number of
participants: 33 women | Hieken et al
(17) |
| benign
disease: | undergoing
non-mastectomy breast surgery |
|
|
Fusobacterium (Phylum:
Fusobacteria) | for benign disease
(n=16) or cancer (n=17) |
|
|
Atopobium (Phylum:
Actinobacteria) | Tissue samples were
obtained from |
|
|
Gluconacetobacter
(Phylum: Proteobacteria) | 33 women with
benign disease (n=16) |
|
|
Hydrogenophaga (Phylum:
Proteobacteria) | and cancer (n=17)
and were compared to |
|
|
Lactobacillus (Phylum:
Firmicutes) | 28 adjacent normal
tissues from the patients with benign disease without atypia (n=13)
and from patients with invasive breast cancer (n=15) |
|
| Increased levels of
bacteria in normal breast tissue: | Total number of
participants: Tissue | Thompson et
al (18) |
|
Actinobacteria | samples were
obtained from 668 women |
|
| Increased levels of
bacteria in BC tissue samples: | and included in the
TCGA data portal |
|
|
Proteobacteria | • 668 tumor tissues
were derived from the |
|
|
| TCGA
data portal |
|
|
| • 72 non-cancerous
adjacent tissues were |
|
|
| derived
from the TCGA data portal |
|
| No significant
difference either in the overall diversity | Total number of
participants: 78 women | Wang et al
(20) |
| or in the number of
microbes between cancerous and | Tissue samples were
obtained from: |
|
| healthy
simples | • 57 women who
underwent mastectomy |
|
| Increased levels of
Methylobacterium (Phylum: | due to
invasive breast carcinoma (samples |
|
|
Proteobacteria) in tissues of
healthy individuals | of 15
women could not be obtained) |
|
| compared to BC
patients | • 21 women
underwent cosmetic procedures |
|
|
| such as
bilateral reduction mammoplasty |
|
|
| and
mastopexy (sample of 1 woman could not be obtained) |
|
| Increased levels of
bacteria in tumor tissues of BC | Total number of
participants: 8 women | Urbaniak et
al (14) |
| women compared to
tissues of healthy women: | undergoing breast
surgery |
|
| Escherichia
coli (Phylum: Proteobacteria) | Tissue samples were
collected from: |
|
| Bacillus
cereus (Phylum: Firmicutes) | • 58 women who had
lumpectomies or |
|
|
|
mastectomies for benign (n=13)
or |
|
|
|
cancerous (n=45) tumors |
|
|
| • 23 were healthy
controls who underwent |
|
|
| breast
reductions or enhancements |
|
Bacteria in non-cancerous adjacent
tissue and cancer tissue of BC women compared to breast tissue of
healthy women
Xuan et al (13) reported that in addition to the
presence of different species between tumor and normal breast
tissues, tumor tissues seem to have a dramatic reduction in
bacterial load compared to normal tissues. In addition, the inverse
correlation between bacterial load and tumor stage implies that
bacterial load could be used in conjunction with current methods to
monitor the progression of BC and to facilitate staging of the
disease. However, further research is warranted to evaluate and
determine a possible role of the bacterial load in the diagnosis or
staging of BC. Furthermore, in the study by Xuan et al
(13) it was observed that one third
of antibacterial genes were downregulated in tumor tissues compared
to healthy tissues. These results suggest that bacteria may play a
role in regulating immune responses within healthy breast tissue
and these responses may be disrupted during tumorigenesis (13).
Following on from previous research, Urbaniak et
al (14) used 16S rRNA amplicon
sequencing and showed that the bacterial profiles differ between
tissues from healthy controls (women undergoing breast reduction or
breast enhancement surgeries), normal adjacent tissue from women
with BC and tumor tissue from women with BC. The researchers found
that women with BC had higher relative abundances of Bacillus,
Enterobacteriaceae and Staphylococcus, among others. The
researchers also reported that Escherichia coli (a member of
the Enterobacteriaceae family) and Staphylococcus
epidermidis, isolated from BC patients, were shown to induce
DNA double-stranded breaks in HeLa cells using the histone-2AX
(H2AX) phosphorylation assay. They also found that microbial
profiles are similar between normal adjacent tissue and tissue
sampled directly from the tumor in BC women. Interestingly they
also found that Lactococccus, Streptococcus and
Prevotella were higher in the tissues of healthy women
compared to BC patients (Table I).
Interestingly, all of the aforementioned bacteria have shown to
confer some protective properties against BC. Lactococcus
spp. is known to have anti-carcinogenic properties through
potentially activating natural killer (NK) cells and enhancing
cellular immunity, thus being potentially protective against BC.
Streptococcus, especially S. thermophilus has also
been shown to have protective properties against cancer by
producing anti-oxidant metabolites that neutralize reactive oxygen
species and thus protect the host from DNA damage. In addition,
Prevotella is able to produce the short-chain fatty acid
(SCFA) propionate, which has shown to have anti-inflammatory
properties (14).
In another study, Chan et al (15) investigated the composition of nipple
aspirate fluid (NAF) on BC survivors and reported a higher level of
β-glucuronidase levels as well as a higher abundance of
Alistipes in the NAF of BC survivors compared to healthy
controls. On the other hand, they found an unclassified genus of
the Sphingomonadaceae family in the NAF of healthy
individuals. Additionally, they also found Sphingobium
yanoikuyae to be more abundant in normal breast tissue when
compared to ER+ breast tumor tissue (Table I). While the benefits of
Sphingomonadaceae are not well known, this family has also
been known to break down aromatic hydrocarbons as well as
polycyclic aromatic hydrocarbons, both of which are known to be
associated with BC (15,16). Due to their ability to metabolize
aromatic hydrocarbons, they may confer protective properties
against BC, as they were only observed in NAF of healthy breast
tissue. However, more research is warranted to confirm this
association (15).
Hieken et al (17) explored the different microbiota
composition between benign vs. malignant breast tumor tissue. The
study reported significant differences in the breast tissue
microbiome between women with benign vs. women with malignant
disease. Specific genus-level taxa that were significantly enriched
in breast tissue from women with malignant disease included
Fusobacterium, Atopobium, Gluconacetobacter, Hydrogenophaga
and Lactobacillus (17)
(Table I).
In a study by Thompson et al (18), the researchers aimed to compare the
microbiota in BC tissues compared to non-cancerous adjacent
tissues. They found Proteobacteria to be enriched in breast
tumor tissue samples while Actinobacteria were found in
abundance in non-cancerous adjacent tissues. In addition, Thompson
et al (18) found E.
coli to be prevalent in cancerous and adjacent non-cancerous
tissues but with a higher abundance in non-cancerous adjacent
tissue of BC women (Table I). This
was inconsistent with the observations by Urbaniak et al
(14) who reported higher levels of
E. coli in BC compared to healthy controls. Thompson et
al (18) also observed an
increase in Streptococcus pyogenes, in tumor breast tissue
samples among others. Other recent studies have also established a
relationship between Streptococcus spp. and
β-glucuronidase/β-glucosidase enzymes that are responsible for
promoting the re-circulation of estrogen by breaking down the
estrogen-glucoronate conjugate (18,19).
Therefore, S. pyogenes may increase systemic estrogen levels
which have been continuously proven to increase the risk for BC
(18) (Table I).
In another study, Wang et al (20) observed the microbiome of breast
tissue, urine and oral rinse from BC patients and compared them
with those of healthy women (women who underwent cosmetic
procedures such as bilateral reduction mammoplasty and mastopexy).
Unlike previous studies, they found no significant differences
either in the overall diversity or in the number of microbes
between cancerous and healthy samples. Additionally, tissues from
healthy participants were shown to have an increased abundance of
Methylobacterium while in BC patients the same bacterium was
observed to be significantly decreased (20) (Table
I).
In addition, Parida and Sharma (16) reported that Enterobacteriaceae
(specifically E. coli), and Bacillus cereus were more
abundant in tumor tissues than in normal breast tissues. E.
coli, (B2 phylotype), has cancer-promoting properties as it has
the ability to produce colibacin, a known genotoxin. Colibacin is
known to induce double-stranded DNA breaks and has not only been
implicated in colorectal cancer but has now also been linked to BC
(16). DNA damage induced by microbes
may not be enough for tumorigenesis to occur, but if coupled with
other molecular errors the result could be detrimental to host
health (14). On the other hand, even
though B. cereus does not necessarily induce DNA damage, it
still possesses pro-carcinogenic properties. For example, B.
cereus is able to metabolize progesterone into
5-alpha-3,20-dione (5aP), which has been seen to be much higher in
BC and has been observed in vitro to induce BC cell
proliferation (14,16).
Some of the limitations of the studies investigating
the microbiota in normal and malignant breast tissue are the low
number of participants as well as the fact that the microbiota from
BC tissues was commonly compared to that of women with benign
disease or women that underwent breast reduction or breast
augmentation surgeries (Table I). It
is questionable whether the tissues assessed by women undergoing
breast reduction should be considered as ‘healthy.’ The potential
absence of epithelium or stroma in these fatty tissues could render
these not directly comparable to a breast cancer which is comprised
predominantly of epithelium and stroma, or healthy breast tissue
that has a varying abundance of these cell types.
Bacteria found in different BC subtypes
The microbiota of ER+
malignant tumors compared to healthy tissues
Xuan et al (13) investigated the potential role of
breast microbiota on ER+ BC by comparing the DNA of the
microbiota found in the breast tumor tissue compared to the normal
adjacent tissue from the same patients and healthy women that
underwent reduction mammoplasty. Sphingomonas yanoikuyae was
the most abundant bacterium found in normal adjacent breast tissue,
while the bacterium Methylobacterium radiotolerans was
abundant in ER+ breast tumor tissues (13). Interestingly, while S.
yanoikuyae was detected in almost half of normal adjacent
tissue samples, it was not detected at all in the paired
corresponding tumor tissues, whereas M. radiotolerans was
detected in both the normal adjacent and the tumor tissue samples.
This observation suggests that S. yanoikuyae may have a
protective function in the breast, potentially due to its ability
to express certain ligands that activate important cancer
immuno-surveillance mediators (13)
(Table II).
| Table II.Summary of some of the most important
bacteria found in different breast cancer (BC) subtypes. |
Table II.
Summary of some of the most important
bacteria found in different breast cancer (BC) subtypes.
| Bacteria found in
normal adjacent tissues compared to ER+ breast tumors of
BC patients | Study
participants | Authors
(Refs.) |
|---|
| Most abundant
bacterium in normal adjacent tissues | Total number of
participants: 91 women | Xuan et al
(13) |
| of BC
patients: | Tissues samples
were collected from: |
|
|
Sphingomonas
yanoikuyae | • 65 women with
ER+ BC |
|
| Most abundant
bacterium in ER+ breast tumor tissues: | • 26 healthy women
that underwent |
|
|
Methylbacterium
radiotolerans |
reduction mammoplasty with no
evidence of BC |
|
|
| Microbiota of
ER+ and PR+ malignant tumors vs.
ER− PR− tumors | Study
participants | Authors
(Refs.) |
|
| Higher microbial
diversity and abundance in | Total number of
participants: 78 women | Wang et al
(20) |
| hormone-positive BC
vs. hormone-negative BC | Tissues samples
were collected from: |
|
| Increased levels of
bacterium in hormone-negative | • 57 women who
underwent mastectomy |
|
| BC compared to
hormone positive BC: | due to
invasive breast carcinoma (samples |
|
|
Methylbacterium | of 15
women could not be obtained) |
|
|
| • 21 women who
underwent cosmetic procedures such as bilateral reduction
mammoplasty and mastopexy (sample of 1 woman could not be
obtained) |
|
|
| Microbiota of
HER-2-positive (HER-2+), hormone receptor-positive
(PR+/ER+), triple-negative (PR−,
ER−, HER-2−) and triple-positive
(PR+, ER+, HER-2+) BC | Study
participants | Authors
(Refs.) |
|
| Microbiota found in
all four BC subtypes: | Total number of
participants: 168 women | Banerjee et
al (21) |
|
Sphyngomonas | Tissue samples were
collected from: |
|
|
Mycobacterium | • 50 women with
hormone receptor-positive |
|
| Higher levels in
PR+/ER+ and triple-positive compared | breast
cancer |
|
| to
PR−/ER− and triple-negative subtypes: | • 34 women with
HER-2+ breast cancer |
|
|
Brevundiomanas |
samples |
|
| Increased levels of
bacterium in PR− and ER− BC | • 24 women with
triple-positive breast |
|
| compared to
ER+PR+ tissues: | cancer
samples |
|
|
Mycobacterium | • 40 women
triple-negative breast cancer |
|
|
|
samples |
|
|
| • 20 women who
underwent breast |
|
|
|
reduction surgery |
|
|
| Due to HIPAA
regulations that the study was subjected to, there was no
information on the type of treatment that the individuals with BC
received. |
|
The microbiota in ER+ and
PR+ malignant tumors compared to ER−
PR− tumors
Wang et al (20) observed that the microbial diversity
and abundance was higher in hormone-positive BC than in
hormone-negative BC. This could be due to the ability of some
microbes to metabolize and regulate the bioavailability of steroid
hormones in the former (16,20). Additionally, they found
Methylbacterium to be significantly decreased in
hormone-positive BC when compared to hormone-negative BC (20) (Table
II).
The microbiota of HER-2-positive
(HER-2+), hormone receptor-positive
(PR+/ER+), triple-negative (PR−,
ER−, HER-2−) and triple-positive
(PR+, ER+, HER-2+) BC
A study by Banerjee et al (21) was carried out to investigate the
specific microbial signatures in 4 different BC subtypes i.e.,
HER-2-positive (HER-2+), hormone receptor-positive
(PR+/ER+), triple-negative (PR−,
ER−, HER-2−) and triple-positive
(PR+, ER+, HER-2+) BC. In
addition, women who had undergone breast reduction surgery were
used as a control. The researchers observed that, among others,
Sphyngomonas and Mycobacterium, were present in all
four BC subtypes. Additionally, they found that each subtype had a
unique microbial signature; Triple-negative had the least complex
microbial signature, while PR+/ER+ was
associated with the most complex microbial signature. Additionally,
higher levels of Brevundimonas were noted in
PR+/ER+ and in triple-positive compared to
PR−/ER− and triple-negative subtypes. In
hormone-negative BC (PR− and ER−) an
increased abundance of Mycobacterium was observed (21) (Table
II). Identifying BC subtype signatures through microbiota
composition could pave way for potential BC screening techniques.
Nevertheless, it is evident that investigations regarding
microbiota residing in normal breast tissue and its role in BC
development is still in its infancy, but can potentially have a
huge role in developing screening and therapy techniques for
BC.
Overall, the limitations of the studies
investigating the microbiota composition in different BC subtypes
were similar to the studies comparing the microbiota in BC vs.
normal tissue; i.e. the number of participants in these studies was
low and they used women that underwent cosmetic surgeries as
healthy controls (Table II).
Microbiota in the gut and its association
with BC
Even though microbiota can reside in multiple body
habitats, most of the microbial biomass is located in the GI tract
and as such it plays a huge role in the maintenance of the host's
health as well as in the development of various diseases (22). Therefore, the role that the gut
microbiota plays in BC development should also be explored.
Differences in gut microbiota
composition between healthy individuals and women with BC
The dysregulation of the gut microbiota has been
associated with BC since 1990, where a study found that women with
BC had an abundance of multiple different microbes in the GI tract
(Table III) (23). Since then, more recent studies have
continuously showed that the gut microbiome composition does indeed
change in women with BC compared to those without (22).
| Table III.Summary of the intestinal microbiota
found in women with breast cancer (BC), post-menopausal women with
BC, and healthy post-menopausal women. |
Table III.
Summary of the intestinal microbiota
found in women with breast cancer (BC), post-menopausal women with
BC, and healthy post-menopausal women.
| Bacteria found in
the gut of women with BC | Study
participants | Authors
(Refs.) |
|---|
| Escherichia
coli | Total number of
participants: 66 women | Minelli et
al (23) |
|
Clostridium | Faeces samples were
collected from: |
|
|
Enterobacterium | • 18 women with
BC |
|
|
Lactobacillus | • 18 women with
uterine myoma |
|
|
Bacteroides | • 30 healthy
women |
|
|
| Bacteria found
in the gut of healthy post-menopausal women vs. post-menopausal
women with BC | Study
participants | Authors
(Refs.) |
|
| Bacteria found at
increased levels in the gut of | Total number of
participants: 133 women | Zhu et al
(24) |
| post-menopausal
women with BC compared to healthy | Faeces samples were
collected from: |
|
| post-menopausal
women: | • 18 pre-menopausal
women with breast |
|
|
Escherichia coli |
cancer |
|
|
Klebsiella
sp_1_1_55 | • 25 pre-menopausal
healthy controls |
|
|
Prevotella amnii | 44
post-menopausal women with breast |
|
|
Enterococcus
gallinarum |
cancer |
|
|
Actinomyces sp.
HPA0247 | • 46
post-menopausal healthy controls |
|
|
Shewanella
putrefaciens | None of the BC
patients received any form |
|
|
Erwinia amylovora | of chemotherapy,
radiation or surgery |
|
| Bacteria found at
decreased levels in the gut of post-menopausal women with BC
compared to healthy post-menopausal women: | before fecal sample
collection |
|
|
Eubacterium
eligens |
|
|
|
Lactobacillus
vaginalis |
|
|
| Bacteria found at
increased levels in the gut of healthy post-menopausal women
compared to post-menopausal women with BC: |
|
|
|
Eubacterium
eligens |
|
|
|
Roseburia
inulinivorans |
|
|
A more recent study by Zhu et al (24) showed that the composition and
functions of the gut microbial community differ between
post-menopausal BC patients and post-menopausal healthy controls.
In this study, the researchers performed a comprehensive shotgun
metagenomic analysis of 18 pre-menopausal BC patients, 25
pre-menopausal healthy controls, 44 post-menopausal BC patients and
46 post-menopausal healthy controls. The results of the study
showed that the microbial diversity was higher in BC patients than
in controls. Relative species abundance in gut microbiota did not
differ significantly between pre-menopausal BC patients and
pre-menopausal controls. However, the relative abundance of 45
species differed significantly between post-menopausal BC patients
and post-menopausal controls: 38 species were enriched in
post-menopausal BC patients including Escherichia coli,
Klebsiella sp_1_1_55, Prevotella amnii, Enterococcus gallinarum,
Actinomyces sp. HPA0247, Shewanella putrefaciens, and
Erwinia amylovora. On the other hand, 7 species were less
abundant in post-menopausal BC patients compared to healthy
controls including Eubacterium eligens and Lactobacillus
vaginalis (24).
Interestingly Zhu et al (24) reported that Erwina amylovora
and Shewanella putrefaciens were shown to have a positive
correlation, albeit a weak one, with estradiol, suggesting
potential involvement of both microbes in the metabolism of
estrogen. Due to their potential involvement in estrogen
metabolism, such gut microbiota could be used in the future as
novel biomarkers for BC (24)
(Table III).
In the same study, Zhu et al (24) also observed that Eubacterium
eligens and Roseburia inulinivorans were more abundant
in post-menopausal healthy controls than in post-menopausal women
with BC. Interestingly, R. inulinivorans is a bacterium with
known anticarcinogenic properties; i.e. it produces butyrate which
has anti-inflammatory properties mainly by inhibiting the
activation of nuclear factor (NF)-κB in intestinal epithelial cells
through tumor necrosis factor (TNF)-α (24,25).
Therefore, a reduction in butyrate in the colon caused by decreased
levels of R. inulinivorans in post-menopausal women can
increase inflammation and therefore increase their risk of BC
(24). It was also observed that in
both pre-menopausal women and post-menopausal women, genes of the
iron transport system were increased, however, in post-menopausal
women genes involved in lipopolysaccharide (LPS) biosynthesis were
also seen to be increased. Both the iron transport system and LPS
biosynthesis increase systemic inflammation and therefore increase
the risk for BC (24) (Table III).
Microbial metabolites with anticancer
potency in the gut microbiota
It has recently been proposed that the microbiota in
the GI tract may act as an endocrine organ, due to the gut's
ability to produce various metabolites such as SCFAs and to
regulate hormone metabolism, all of which affect distal organs
through the blood stream (26). Some
of these microbial metabolites have been shown to influence the
progression of BC (27). A
well-documented microbe metabolite linked to BC is cadaverine, a
biogenic amine whose biosynthesis is decreased in early stage BC
(27,28). Cadaverine is considered an anticancer
bacterial metabolite because of its ability to suppress the
aggressiveness, metastasis and progression of tumor cells.
Cadaverine has also been shown to induce the metabolism of tumor
cells that have begun glycolysis by decreasing cellular oxygen
consumption (27,28). Additionally, other bacterial
metabolites such as lithocholic acid (LCA), a secondary bile acid,
have been observed to inhibit BC progression by creating oxidative
and nitrosative stress (27). More
specifically, LCA inhibits epithelial-mesenchymal transition,
metastasis and BC progression by activating nuclear factor
erythroid 2-related factor 2 (NRF2) and has pro-apoptotic effects
on BC tumor cells (27,28).
SCFAs are products of fibre being fermented in the
colon by certain bacteria (27). The
most predominant SCFAs are butyrate, propionate, and acetate, which
are well known for their anticancer effects (27). These metabolites have shown to be
protective against cancer as they are involved in mediating cell
cycle arrest, and inducing apoptosis (28). Therefore, changing the composition of
the gut microbiome will inevitably also change the
anti-carcinogenic metabolites being produced, thereby preventing or
promoting BC progression (27).
The role of the gut microbiome in
estrogen metabolism
As previously discussed, the gut microbiome may be
considered an endocrine organ, due partly to its role in estrogen
metabolism. The microbiota in the GI tract is known to be one of
the most important regulators in estrogen circulation (27). This is especially relevant in
hormone-positive BC, as there is an undisputable relationship
between endogenous estrogen burden and the development of BC in
post-menopausal women (there is a similar relationship with
pre-menopausal women but the correlation has not proven to be as
strong) (28,29).
Estrogen metabolism in the GI tract is attributed to
what is currently being referred to as the estrobalome. The
estrobalome is defined as the aggregate of bacterial genes that
result in enzymes that can metabolize estrogens (28,30).
Free-circulating estrogens are subjected to hepatic first-pass
metabolism where they are conjugated and then eliminated via urine
or feces (30). However, a
significant portion of the conjugated estrogen is reabsorbed back
into circulation before it gets excreted. This suggests that
bacteria in the gut possess β-glucuronidase/β-glucosidase activity
allowing them to deconjugate estrogens, permitting them to become
biologically active again and re-enter the circulation (30). The re-circulated estrogens then
interact with breast tissue facilitating cellular growth and thus
contributing to the initiation and progression of BC, as well as
increasing the risk of ER-positive BC in post-menopausal women
(27,28). This is especially relevant as certain
bacteria that have been previously shown to be abundant in BC are
coincidentally also related to an increase in activity of
β-glucuronidase/β-glucosidase activity. Such bacteria include S.
pyogenes, Clostridia spp., Baccilus spp. and E. coli
(18,31). It was also further seen that the
relationship between microbiota and estrogen metabolism can be used
to predict the risk of BC in post-menopausal women due to the
diversity and composition of gut microbiota being associated with
patterns of estrogen metabolism (32). Therefore, a change in microbiota
composition can alter normal host function, favoring BC
development.
The role of the gut microbiome in
breast cancer metastasis
A study by Rosean et al (33) used a mouse model for hormone
receptor-positive (HR+) mammary cancer in order to
investigate whether commensal dysbiosis, more specifically,
pre-existing dysbiosis in the microbiome can influence the
progression and outcome in HR+ BC. The researchers
demonstrated that a pre-established disruption of commensal
homeostasis results in enhanced circulating tumor cells and
subsequent dissemination to the tumor-draining lymph nodes and in
distant sites such as the lungs; all of which promote the poor
outcomes seen in HR+ BC. Due to the results being so
promising, Buchta Rosean et al (33) concluded that commensal dysbiosis may
have therapeutic implications and could potentially be used as a
biomarker for HR+ BC (33). The study by Buchta Rosean et al
(33) currently presents one of the
strongest evidence to date that gut dysbiosis may promote breast
cancer metastasis, and further studies should be conducted to
investigate this association.
Diet and microbiota in BC
Diet is a well-established modifiable risk factor
for BC, as changes in diet have been linked with increasing overall
health and survival rates in patients that already have BC
(34). A diet rich in fat has been
shown to affect β-glucuronidase activity by increasing bile acid
secretion, thus promoting the growth of E. coli and
Enterobacter, both of which are potent β-glucuronidases and
thus contribute to the estrogen burden (30,35).
However, a fibre-rich diet can potentially be onco-protective;
soluble fibre is fermented by SCFAs and creates a favorable
environment for beneficial bacteria such as Bifidobacterium
(30).
Probiotic therapies for BC
Due to the newly established role that the
microbiota potentially plays in the development of BC, it is
important to explore how interventions that affect the composition
of the microbiome (e.g., via the use of probiotics) may affect the
process of breast carcinogenesis. The most common microorganisms
used in probiotics are lactic acid bacteria (LAB) which have been
shown to have multiple health benefits for the host (36). Various other studies have been
conducted both in vitro and in vivo demonstrating the
effects of probiotics on BC (Table
IV).
| Table IV.Summary of in vitro and animal
studies investigating the role of probiotics in breast cancer
(BC). |
Table IV.
Summary of in vitro and animal
studies investigating the role of probiotics in breast cancer
(BC).
| In vitro
studies |
|---|
|
|---|
| Probiotic
strain | Cell lines | Effect on BC | Authors
(Refs.) |
|---|
| Enterococcus
feacalis | In vitro
(MCF-7 cell line) | ↑Apoptosis | Hassan et al
(37) |
| Staphylococcus
hominis |
| ↓Cell
proliferation |
|
| Lactobacillus
crispatus | In vitro
(MDA-MB-231 cell line) | ↑Cytotoxic effects
on triple-negative | Esfandiary et
al (38) |
|
SJ-3C-US |
| BC cells |
|
| Lactobacillus
rhamnosus |
| L. rhamnosus
showed a greater |
|
| GG |
| downregulation of
hypoxia-associated |
|
|
|
| genes (HF1α,
HSP90, SLCA2A1) |
|
| Lactobacillus
plantarum | In vitro
(MCF-7 cell line) | Inhibition of
growth of cell lines | Bharti et al
(39) |
| Lactobacillus
acidophilus |
| L. plantarum
showed the highest |
|
|
|
| cytotoxic effect on
the MCF-7 cell line |
|
| Lactobacillus
plantarum | In vitro
(MDA-MB-231 cell line) | Downregulation of
the NF-κB pathway leading to apoptosis of ER-negative BC cell
lines | Kadirareddy et
al (40) |
|
| Animal
studies |
|
| Probiotic
strain | Animal
model | Effect on
BC | Authors
(Refs.) |
|
| Lactobacillus
helveticus | In vivo
BALB/c mouse | ↑IL-10 | de Moreno de |
| R389 | model | ↓IL-6 | LeBlanc et
al (41) |
|
|
| Induction of
cellular apoptosis |
|
|
|
| Delayed BC
development |
|
| Lactobacillus
reuteri | In vivo mice
models: | Delay in onset of
pre-neoplastic features | Lakritz et
al (42) |
|
ATCC-PTA-6475 | Swiss models
fed | Activates
CD4+ and CD25+ cells |
|
|
| ‘Westernized chow’
and |
|
|
|
| erbB2 (HER2)
mice |
|
|
|
| supplemented
with |
|
|
|
| L.
reuteri |
|
|
| Lactobacillus
casei | In vivo
orally | ↓Growth rate of
tumors | Soltan Dallal et
al (45) |
|
| administered,
BALB/c | ↑Survival compared
with the control |
|
|
| mouse model | ↑IFN-γ and
IL-12 |
|
|
|
| Affects the
stimulation of Th1 cytokine |
|
|
|
| production |
|
|
|
| ↑NK cell
cytotoxicity in spleen cells of |
|
|
|
| mice with invasive
ductal carcinoma. |
|
| Lactobacillus
brevis | In vivo
orally | Improved disease
prognosis in mice | Yazdi et al
(46) |
| (enriched with
selenium | administered,
BALB/c | with highly
metastatic breast tumors |
|
| nanoparticles) | mouse model | ↑NK
cytotoxicity |
|
|
|
| ↑IFN-γ and
IL-17 |
|
|
|
| ↓Metastasis to the
liver |
|
| Lactobacillus
acidophilus | In vivo
orally | ↑Increased survival
time of BALB/c | Imani Fooladi et
al (36) |
|
| administered,
BALB/c | mice |
|
|
| mouse model | ↑IFN-γ |
|
|
|
| ↓IL-4 |
|
| Lactobacillus
acidophilus | In vivo
BALB/c mouse | ↓Cell mitosis | Zamberi et
al (47) |
| Lactobacillus
casei | model with
triple-negative | ↑Cancer cell
apoptosis |
|
| Lactococcus
lactis | 4T1 BC cells fed
Kefir | Inhibition of
inflammation in tumor |
|
|
| water | environment |
|
|
|
| Modulation of
immune system |
|
|
|
| Regulation of
angiogenesis proteins and genes |
|
Studies in cell lines
Hassan et al (37) investigated the role of Enterococcus
feacalis and Staphylococcus hominis in vitro. They
isolated these bacteria from the breast milk of healthy women and
conducted the rest of the investigation in various antibiotic-free
media. They used three different cell forms of the bacteria (live,
heat-killed and cytoplasmic fractions) and found that all three
forms of the bacteria caused a significant decrease in cell
proliferation via induction of both cell cycle arrest and apoptosis
in BC cells such as in MCF-7 cells (37).
Another study by Esfandiary et al (38) investigated the effect of the probiotic
strains Lactobacillus crispatus SJ-3C-US and
Lactobacillus rhamnosus GG on MDA-MB-231 cell lines. They
found that both strains induced cytotoxic effects on
triple-negative BC cells and that L. rhamnosus especially,
downregulated genes associated with hypoxia, such as SLCA2A1
(38), which is known to code for the
production of GLUT 1. By downregulating the production of GLUT 1,
metastasis can be prevented through the MMP-2 and JNK pathways
(9).
Lactobacillus plantarum has also shown
promise in various studies, such as in the study conducted by
Bharti et al (2015), where the highest cytotoxic effect on
MCF-7 cancer cell lines was seen with the administration of L.
plantarum (39). Kadirareddy
et al (40) demonstrated that
L. plantarum induced apoptosis in MDA-MB-231 cells by the
production of conjugated linoleic acid which downregulated the
NF-κB pathway causing proteasome degradation of IκBα, inhibition of
p65 nuclear translocation, release of cytochrome c from the
mitochondria and finally overexpression of Bax protein (40). These are but a few examples of in
vitro studies in which probiotics strains have been used to
investigate their potential role in breast cancer prevention.
Studies in animal models
The efficacy of orally administered probiotics in
tumor cell inhibition was also confirmed by animal studies. For
example, in an in vivo mouse model, the intake of milk
fermented with Lactobacillus helveticus R389 played a role
in delaying breast tumor cell development. The latter was
associated with increased regulatory cytokines such as interleukin
(IL)-10 and decreased IL-6 levels in serum and mammary mouse cells,
as well as by induction of cellular apoptosis. IL-10 is known to
induce the expression of TNF-α related apoptosis-inducing ligand
(TRAIL) and death receptor 4 (DR4), as well as recruiting
lymphocytes, all of which contribute to the apoptosis of mammary
cells (41).
Additionally, Lakritz et al (42) observed the effect that
Lactobacillus reuteri has on the development of mammary
cancer in two different mouse models. In the first model, Swiss
mice were fed Westernized chow (mimicking a westernized diet)
thereby putting the mice at increased risk for the development of
mammary tumors. The second animal model used was FVB strain erbB2
(HER2) mutant mice which are genetically susceptible to mammary
tumors. Both of these mouse models were exposed to L.
reuteri through their water supplies and compared to controls.
Interestingly, exposure to L. reuteri in the experimental
groups was found to be sufficient in delaying the onset of
pre-neoplastic features in both models. This hypothesis is
consistent with other research studies supporting that lactic
acid-producing bacteria decrease the risk of BC in women (42–44). In
addition to inhibiting early stage tumorigenesis, L. reuteri
has also been reported to increase the sensitivity of BC cells to
apoptosis (27,42). This study demonstrates that by
modulating the tumor microenvironment through the use of
probiotics, one can potentially regulate systemic immune cell
responses impacting cancer progression. Such findings may have
future implications in the public health sector since such
administrations could be used to decrease BC risk (42).
Additionally, a study conducted by Soltan Dallal
et al (45) investigated the
effect that orally administrated Lactobacillus casei has on
NK cytotoxicity and production of cytokines in the spleen cells of
BALB/C mice that have invasive ductal carcinoma. The results showed
that L. casei was able to decrease the growth rate of
tumors, prolong survival, stimulate Th1 cytokine production and
increase NK cell cytotoxicity in the spleen cells of mice with
invasive ductal carcinoma demonstrating its potential in BC therapy
(45).
Yazdi et al (46) conducted a unique study in which BALB/c
mice were administered with Lactobacillus brevis that was
enriched with selenium nanoparticles. The results showed that this
not only improved the prognosis of mice with highly metastatic BC,
but it also increased NK cytotoxicity, IFN-γ and IL-17 as well as
decreased tumor metastasis to the liver (46). Subsequently, another study confirmed
that the oral administration of Lactobacillus acidophilus
significantly increased the survival time of BALB/c mice with
mammary tumors. L. acidophilus promoted the proliferation of
immune cells and increased the production of IFN-γ and decreased
the production of IL-4. IFN-γ is crucial in the activation of NK
cells, which are known to be the first line defence against tumor
cells (36). High levels of IFN-γ not
only increase NK cytotoxicity but may also increase tumor cell
visibility [through its role in major histocompatability complex
(MHC) expression], as well as inhibiting intra-tumoral angiogenesis
(36).
In another study by Zamberi et al (47), BALB/c mice were injected with
triple-negative 4T1 BC cells, and were treated with Kefir water
that is naturally enriched with L. acidophilus, L. casei,
and Lactobacillus lactis. The results showed a decrease in
cell mitosis and an increase in apoptosis in mice that drank the
water enriched with the specific probiotic strains. In addition,
they found that the various strains also inhibited the inflammation
in the tumor environment, modulated the immune system and regulated
genes and proteins of angiogenesis (47). Moreover, Zamberi et al
(47) and the Soltan Dallal et
al (45) demonstrate how
especially L. casei, not only decreased the growth of
tumors, but also increased apoptosis and inhibited inflammation,
further illustrating the emerging importance of this microbe in BC
prevention and possibly treatment.
Limitations and future directions of
studies with probiotics
While the aforementioned studies presented
promising results, it should be noted that some of the studies had
limitations. For example, most of the studies were either conducted
in vitro or using mouse models and therefore could not
investigate the different factors that affect the diversity of the
microbiome in humans, such as diet, age or genetic background
(48).
In their very comprehensive perspective review Suez
et al (48), reports that even
though probiotics are commonly used by the general public there are
still conflicting clinical results for many probiotic strains and
formulations. Furthermore, the researchers stress the importance of
how large scale randomized blind clinical trials should be designed
to assess a number of questions such as gut colonization by
probiotics, the importance of specific bacterial strains and their
interactions with the indigenous microbiome and the safety and
impact on the host.
Evidence from the literature discusses how the
safety and adverse effects of probiotics have neither been
extensively researched nor correctly documented in most of the
clinical trials. In fact, it has been observed that the use of
probiotics in critically ill adults in intensive care units, young
infants, or neonates with a very low birth weight is associated
with an increased risk of infection. In addition, patients that
underwent antibiotic treatments showed increased colonization by
probiotic strains, which was linked to persistent dysbiosis induced
by long term probiotics (48). A
systematic review by Befeta et al (49), reported that most clinical trials did
not report the adverse effects accurately and support how the
increasing use of probiotics both in clinical practice and
over-the-counter increases the urgency to design and conduct
clinical trials that adequately report and document any adverse
effects caused by probiotics (49).
Therefore, it is important to note that research on
the effects of probiotics on BC is still at its infancy and larger
and more elaborate studies should be conducted to conclude if they
have a protective long-term anticancer effects and to find the
optimum dosage, strain and regimen for a treatment with probiotics
to be effective.
Conclusion
In recent years, the microbiota has proven to play
an important role in diseases and has sparked interest regarding
its involvement in the development, management and screening of BC.
Preliminary studies have provided evidence showing diverse
microbiota belonging to the phyla Proteobacteria, Firmicutes
and Actinobacteria being present in normal breast tissue
(10,14,15). Some
bacteria were found to be more abundant in malignant breast tissue
compared to benign breast tissue and these include, among others,
Fusobaterium, Atopodium, Gluconacetobacter, Hydrogenophaga,
Lactobacillus (17), E.
coli and B. cereus (16)
(Table I). On the other hand,
Lactococcus, Streptococcus, Prevotella and S. yanoikuyae
were more abundant in healthy breast tissue compared to malignant
tissue, suggesting that they may confer various protective effects
against BC (13,14) (Tables I
and II).
In addition, differences in the microbiota were
reported between different types of BC. Sphyngomona and
Mycobacterium were reported in all four BC subtypes
(HER-2-positive (HER-2+), hormone receptor-positive
(PR+/ER+), triple-negative (PR−,
ER−, HER-2−) and triple-positive
(PR+, ER+, HER-2+) BC),
Brevudioomonas was present at higher levels in
PR+/ER+ and triple-positive BC compared to
PR−/ER− and triple-negative subtypes, whereas
Mycobacterium was found at increased levels in
PR− and ER− BC compared to
ER+PR+ tissues (21). In addition, a study has reported that
Methylobabacterium radiotolerans was the most abundant
bacterium in ER+ tissues (13), whereas another study supported that
Methylbacterium was found at increased levels in
hormone-negative BC compared to hormone-positive BC (20) (Table
II).
Overall, evidence in the literature has suggested
an association between bacterial species and breast cancer
pathogenesis. However, it should be noted that these studies were
association studies and in addition suffered from a few limitations
such as the low participant number and the use of tissues from
women that underwent breast reduction or augmentation surgeries as
normal tissue controls in some of the studies (Tables I and II). Therefore, care should be taken in
interpreting the findings from these studies.
Other studies have also provided evidence for an
association between gut microbiota and BC. Certain bacteria such as
Escherichia coli, Klebsiella sp_1_1_55, Prevotella amnii,
Enterococcus gallinarum, Actinomyces sp. HPA0247, Shewanella
putrefaciens, Erwinia amylovora were seen to be present at
higher levels in post-menopausal women with BC compared to healthy
post-menopausal women (24). On the
other hand, bacteria such as Eubacterium eligens and
Lactobacillus vaginalis were found to be at decreased levels
in the gut of post-menopausal women with BC compared to healthy
menopausal women (24) (Table III). Similarly, to the studies
investigating the microbiota in normal and BC tissue, only a few
studies investigated the association between gut microbiota and BC
and these studies used a small number of participants (Table III) again supporting the notion that
caution should be taken in interpreting the results of these
studies.
Evidence from the literature also suggests an
association between gut microbiota and BC progression possibly
through metabolites that certain bacteria produce, as well as their
role in dysregulating estrogen metabolism. Currently, we have
enough knowledge to confirm that bacterial metabolites are crucial
to the molecular pathways of carcinogenesis. These metabolites can
either promote carcinogenesis through genotoxins and other
metabolites or can be protective, through the production of
SCFAs.
At the present time, the situation is still complex
and there are discrepancies between studies with regards to the
direct association of specific bacteria to BC (13,20),
supporting the need for larger randomised controlled studies to be
carried out to derive more conclusive results. Furthermore,
specific bacteria used in probiotics mainly Lactobacillus
spp but also Enterococcus feacalis and Staphylococcus
hominis may confer protective advantages against BC as shown in
preliminary studies in cell lines and animal models and larger
randomised controlled trials will have to be conducted to derive
conclusive results (Table IV).
By understanding the identity and the molecular
mechanisms regulated by pro-carcinogenic and anti-carcinogenic
bacteria, as well as the association between the microbiota in the
gut and the breast it will easier to explore new management routes
that may be more effective and produce less toxic side effects than
current treatments. Additionally, by knowing how bacterial load
changes in BC and which specific microbial signatures are present
in the different BC subtypes, new methods will be developed to
improve screening (i.e. by the use of biomarkers) and consequently
prevention of BC (e.g. by the use of probiotics consisting of
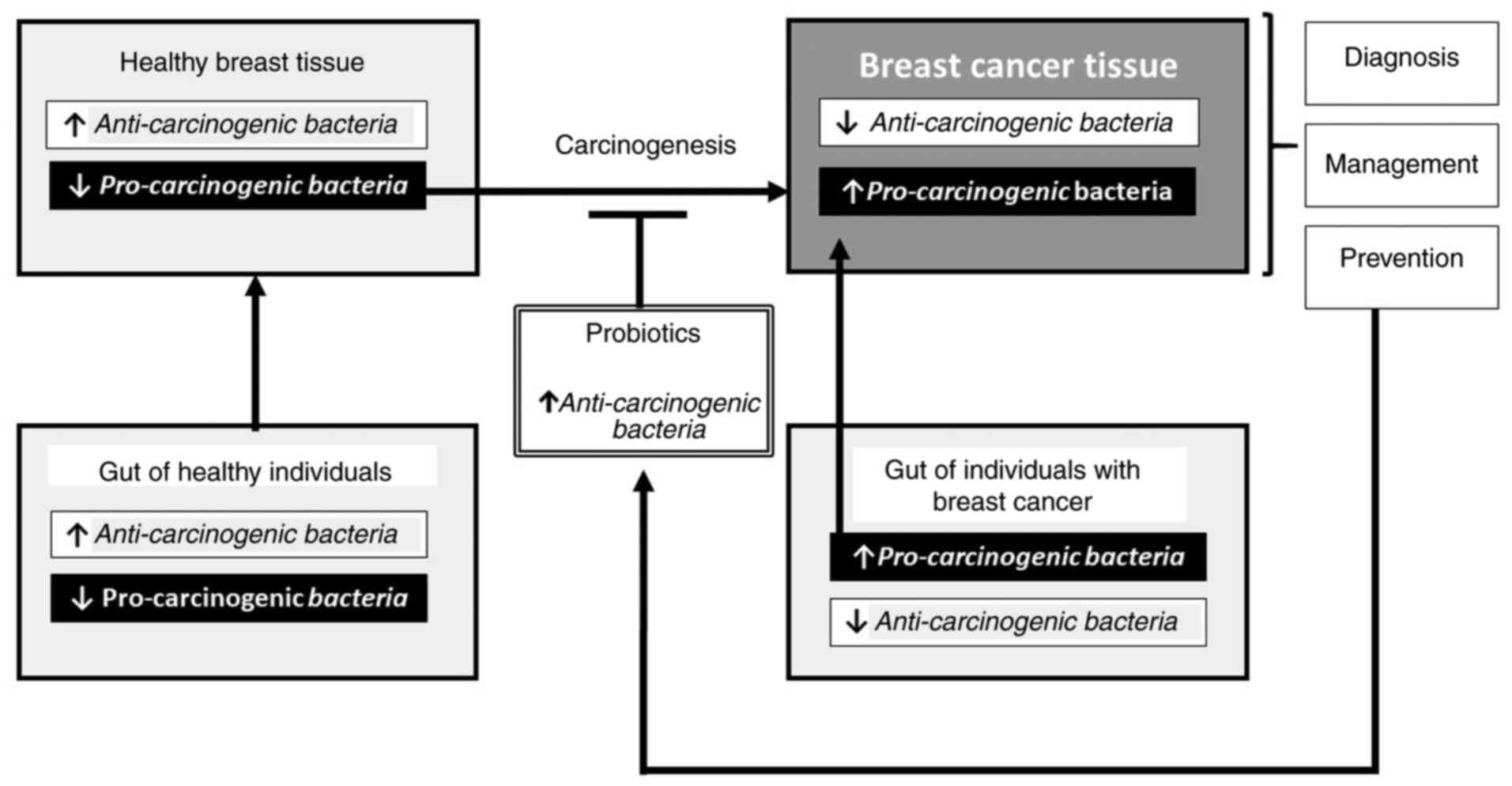
anti-carcinogenic bacteria) (Fig. 1).
This will therefore lead to the development of less invasive and
more effective ways to detect BC and as a result, a visible
decrease in morbidity and mortality associated with BC should be
expected to be seen.
Currently there are various clinical trials ongoing
investigating various intervention strategies. For example, an
interventional study was created to observe the effect that
probiotics have on the immune system of patients suffering from
breast adenocarcinoma stage I–III. This study used a probiotic
formula that included 13 species of bacteria. Some of these
beneficial bacteria include Lactobacillus plantarum
Lactobacillus rhamnosus and Lactobacillus casei
(50). Additionally, an observational
case-control, cross sectional study has been developed to observe
the relationship between BC risk in women with stage I and II and
the gut/mammary microbiota, as well as investigating the possible
contribution of environmental contaminants in the alteration of the
microbiota (51). These are only a
few examples of the many clinical trials that are currently in
progress. The remaining (52–55) are documented in Table V to investigate the link between BC
and the microbiota.
| Table V.Summary of ongoing clinical trials
regarding breast cancer (BC) and microbiota. |
Table V.
Summary of ongoing clinical trials
regarding breast cancer (BC) and microbiota.
| Study ID | Type of study | Year study
started | Expected year of
completion | Status | Aim | Probiotic/Drug
used | (Refs.) |
|---|
| NCT03358511 | Interventional
study | 2017 | 2020 | Completed | To investigate if
probiotics are able to affect the body's immune system on BC. All
subjects that had operable stage I–III breast adenocarcinoma were
given Primal Defense Ultra® Probiotic formula* for 2–4
weeks before surgery | Saccharomyces
boulardii, Lactobacillus plantarum, Bacillus subtilis,
Bifidobacterium lactis, Bifidobacterium bifidum, Lactobacillus
rhamnosus, Bifidobacterium breve, lactobacillus casei,
Lactobacillus salivarius, Lactobacillus acidophilus, Lactobacillus
brevis, Bifidobacterium longum, and Lactobacillus
paracasei | Clinicaltrials.org (50) |
| NCT03586297 | Observational
prospective cohort study | 2017 | 2022 | Recruiting | To observe the gut
and intratumoral micro biome composition as well as antitumor
immune responses in women with triple-negative BC receiving
standard of care neoadjuvant therapy. | No probiotics
used | Clinicaltrials.org (52) |
| NCT03885648 | Observational
case-control, cross sectional study with 200 participants | 2018 | 2022 | Recruiting | To investigate if
the risk of breast cancer in women with stage I and II is
associated with the composition of the gut/mammary microbiota and
if exposure to environ mental contaminants could contribute to the
alteration of the microbiota. | No probiotic was
used in this study | Clinicaltrials.org (51) |
| NCT04138979 | Observational
cross-sectional case control study | 2019 | 2020 | Recruiting | To explore the link
between gut microbiota and chemotherapy used in BC |
Cyclophosphamide | Clinicaltials.org (53) |
| NCT03290651 | Interventional,
double blind randomized, placebo controlled pilot trial | 2019 | 2021 | Recruiting | To test the
hypothesis that by taking an oral probiotic, it can lead to the
displacement of harmful bacteria in the breast and to reduce
inflammation linked to BC | Probiotic contains
Lactobacillus rhamnosus GR-1 and Lactobacillus reuteri
RC-14 | Clinicaltrials.org (54) |
| NCT04139993 | Interventional
non-randomized study | 2020 | 2025 | Not yet
recruiting | To evaluate the
safety and tolerability of a novel oral Microbiome Restoration
Therapy™, RBX7455 and to evaluate intratumoral immunomodulatory
effects in patients with stage I–III BC before undergoing
definitive surgery | Novel oral
Microbiome Restoration Therapy™: RBX7455 | Clinicaltrials.org (55) |
Overall, more research in the field of the
association between the microbiota and cancer development will
significantly improve the prevention of BC since particular
metabolites of the microbiome could be used as biomarkers for
screening. In addition, probiotics (especially those of the
Lactobacillus spp.) could be used in the management of women
diagnosed with BC in order to prevent BC metastasis or relapse
(13). Even though such research is
still at its infancy, the microbiota both in the mammary tissue and
in the gut seem to have a promising future in the management and
prevention of BC.
Acknowledgements
Not applicable.
Funding
No funding was used for the development of the
current manuscript.
Availability of data and materials
All information included in this review has been
documented by relevant references.
Authors' contributions
DT, SED and CC were all involved in the
conceptualization of the current manuscript. DT and SED were
involved in the literature search and review of the resources that
were used in the current manuscript. All authors were involved in
the writing and revision of the manuscript and they all approved
the final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Authors' information
Dr Constantina Constantinou: Orcid ID:
0000-0001-6167-4023.
References
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. Feb 4–2021.(Epub ahead
of print). doi: 10.3322/caac.21660. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brown SB and Hankinson SE: Endogenous
estrogens and the risk of breast, endometrial, and ovarian cancers.
Steroids. 99:8–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fernández MF, Reina-Pérez I, Astorga JM,
Rodríguez-Carrillo A, Plaza-Díaz J and Fontana L: Breast cancer and
its relationship with the microbiota. Int J Environ Res Public
Health. 15:17472018. View Article : Google Scholar
|
|
5
|
Momenimovahed Z and Salehiniya H:
Epidemiological characteristics of and risk factors for breast
cancer in the world. Breast Cancer (Dove Med Press). 11:151–164.
2019.PubMed/NCBI
|
|
6
|
da Costa Vieira RA, Biller G, Uermura G,
Ruiz CA and Curado MP: Breast cancer screening in developing
countries. Clinics (Sao Paulo). 72:244–253. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Howlader N, Cronin KA, Kurian AW and
Andridge R: Differences in breast cancer survival by molecular
subtypes in the United States. Cancer Epidemiol Biomarkers Prev.
27:619–626. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Turnbaugh PJ, Ley RE, Hamady M,
Fraser-Liggett CM, Knight R and Gordon JI: The human microbiome
project. Nature. 449:804–810. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Parida S and Sharma D: Microbial
alterations and risk factors of breast cancer: Connections and
mechanistic insights. Cells. 9:10912020. View Article : Google Scholar
|
|
10
|
Urbaniak C, Cummins J, Brackstone M,
Macklaim JM, Gloor GB, Baban CK, Scott L, O'Hanlon DM, Burton JP,
Francis KP, et al: Microbiota of human breast tissue. Appl Environ
Microbiol. 80:3007–3014. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Toumazi D and Constantinou C: A fragile
balance: The important role of the intestinal microbiota in the
prevention and management of colorectal cancer. Oncology.
98:593–602. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chung YR, Kim HJ, Kim YA, Chang MS, Hwang
KT and Park SY: Diversity index as a novel prognostic factor in
breast cancer. Oncotarget. 8:97114–97126. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xuan C, Shamonki JM, Chung A, Dinome ML,
Chung M, Sieling PA and Lee DJ: Microbial dysbiosis is associated
with human breast cancer. PLoS One. 9:e837442014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Urbaniak C, Gloor GB, Brackstone M, Scott
L, Tangney M and Reid G: The microbiota of breast tissue and its
association with breast cancer. Appl Environ Microbiol.
82:5039–5048. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chan AA, Bashir M, Rivas MN, Duvall K,
Sieling PA, Pieber TR, Vaishampayan PA, Love SM and Lee DJ:
Characterization of the microbiome of nipple aspirate fluid of
breast cancer survivors. Sci Rep. 6:280612016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Parida S and Sharma D: The power of small
changes: Comprehensive analyses of microbial dysbiosis in breast
cancer. Biochim Biophys Acta Rev Cancer. 1871:392–405. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hieken TJ, Chen J, Hoskin TL,
Walther-Antonio M, Johnson S, Ramaker S, Xiao J, Radisky DC,
Knutson KL, Kalari KR, et al: The microbiome of aseptically
collected human breast tissue in benign and malignant disease. Sci
Rep. 6:307512016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Thompson KJ, Ingle JN, Tang X, Chia N,
Jeraldo PR, Walther-Antonio MR, Kandimalla KK, Johnson S, Yao JZ,
Harrington SC, et al: A comprehensive analysis of breast cancer
microbiota and host gene expression. PLoS One. 12:e01888732017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Flores R..Shi J, Fuhrman B, Xu X, Veenstra
TD, Gail MH, Gajer P, Ravel J and Goedert JJ: Fecal microbial
determinants of fecal and systemic estrogens and estrogen
metabolites: A cross-sectional study. J Transl Med. 10:2532012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang H, Altemus J, Niazi F, Green H,
Calhoun BC, Sturgis C, Grobmyer SR and Eng C: Breast tissue, oral
and urinary microbiomes in breast cancer. Oncotarget.
8:88122–88138. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Banerjee S, Tian T, Wei Z, Shih N, Feldman
MD, Peck KN, DeMichele AM, Alwine JC and Robertson ES: Distinct
microbial signatures associated with different breast cancer types.
Front Microbiol. 9:9512018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Parida S and Sharma D: The
microbiome-estrogen connection and breast cancer risk. Cells.
8:16422019. View Article : Google Scholar
|
|
23
|
Minelli EB, Beghini AM, Vesentini S,
Marchiori L, Nardo G, Cerutti R and Mortani E: Intestinal
microflora as an alternative metabolic source of estrogens in women
with uterine leiomyoma and breast cancer. Ann N Y Acad Sci.
595:473–479. 1990. View Article : Google Scholar
|
|
24
|
Zhu J, Liao M, Yao Z, Liang W, Li Q, Liu
J, Yang H, Ji Y, Wei W, Tan A, et al: Breast cancer in
postmenopausal women is associated with an altered gut metagenome.
Microbiome. 6:1362018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Inan MS, Rasoulpour RJ, Yin L, Hubbard AK,
Rosenberg DW and Giardina C: The luminal short-chain fatty acid
butyrate modulates NF-kappaB activity in a human colonic epithelial
cell line. Gastroenterology. 118:724–734. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Heinken A and Thiele I: Systematic
prediction of health-relevant human-microbial co-metabolism through
a computational framework. Gut Microbes. 6:120–130. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Eslami-S Z, Majidzadeh-A K, Halvaei S,
Babapirali F and Esmaeili R: Microbiome and breast cancer: New role
for an ancient population. Front Oncol. 10:1202020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang Z, Tang H, Chen P, Xie H and Tao Y:
Demystifying the manipulation of host immunity, metabolism, and
extraintestinal tumors by the gut microbiome. Signal Transduct
Target Ther. 4:412019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mani S: Microbiota and breast cancer. Prog
Mol Biol Transl Sci. 151:217–229. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Komorowski AS and Pezo RC: Untapped
‘-omics’: The microbial metagenome, estrobolome, and their
influence on the development of breast cancer and response to
treatment. Breast Cancer Res Treat. 179:287–300. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Flores R, Shi J, Gail MH, Gajer P, Ravel J
and Goedert JJ: Association of fecal microbial diversity and
taxonomy with selected enzymatic functions. PLoS One. 7:e397452012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Fuhrman BJ, Feigelson HS, Flores R, Gail
MH, Xu X, Ravel J and Goedert JJ: Associations of the fecal
microbiome with urinary estrogens and estrogen metabolites in
postmenopausal women. J Clin Endocrinol Metab. 99:4632–4640. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Buchta Rosean C, Bostic RR, Ferey JCM,
Feng TY, Azar FN, Tung KS, Dozmorov MG, Sirmnova E, Bos PD and
Rutlowski MR: Preexisting commensal dysbiosis is a host-intrinsic
regulator of tissue inflammation and tumour cell dissemination in
hormone receptor-positive breast cancer. Cancer Res. 79:3662–3675.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Malik SS, Saeed A, Baig M, Asif N, Masood
N and Yasmin A: Anticarcinogenecity of microbiota and probiotics in
breast cancer. Int J Food Prop. 21:655–666. 2018. View Article : Google Scholar
|
|
35
|
Yang J, Tan Q, Fu Q, Zhou Y, Hu Y, Tang S,
Zhou Y, Zhang J, Qiu J and Lv Q: Gastrointestinal microbiome and
breast cancer: Correlations, mechanisms and potential clinical
implications. Breast Cancer. 24:220–228. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Imani Fooladi AA, Yazdi MH, Pourmand MR,
Mirshafiey A, Hassan ZM, Azizi T, Mahdavi M and Soltan Dallal MM:
Th1 cytokine production induced by Lactobacillus acidophilus
in BALB/c Mice bearing transplanted breast tumor. Jundishapur J
Microbiol. 8:e173542015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hassan Z, Mustafa S, Rahim RA and Isa NM:
Anti-breast cancer effects of live, heat-killed and cytoplasmic
fractions of Enterococcus faecalis and Staphylococcus
hominis isolated from human breast milk. In Vitro Cell Dev Biol
Anim. 52:337–348. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Esfandiary A, Taherian-Esfahani Z,
Abedin-Do A, Mirfakhraie R, Shirzad M, Ghafouri-Fard S and
Motevaseli E: Lactobacilli modulate hypoxia-inducible factor
(HIF)-1 regulatory pathway in triple negative breast cancer cell
line. Cell J. 18:237–244. 2016.PubMed/NCBI
|
|
39
|
Bharti V, Singh S, Ahirwal L, Mehta A and
Jain N: Cytotoxicity of live whole cell, heat killed cell and cell
free extract of lactobacillus strain in u-87 human glioblastoma
cell line and mcf-7 breast cancer cell line. Int J Probiotics
Prebiotics. 10:153–158. 2015.
|
|
40
|
Kadirareddy RH, Vemuri SG and Palempalli
UM: Probiotic conjugated linoleic acid mediated apoptosis in breast
cancer cells by downregulation of NFκB. Asian Pac J Cancer Prev.
17:3395–3403. 2016.PubMed/NCBI
|
|
41
|
De Moreno de LeBlanc A, Matar C, Thériault
C and Perdigón G: Effects of milk fermented by Lactobacillus
helveticus R389 on immune cells associated to mammary glands in
normal and a breast cancer model. Immunobiology. 210:349–358. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Lakritz JR, Poutahidis T, Levkovich T,
Varian BJ, Ibrahim YM, Chatzigiagkos A, Mirabal S, Alm EJ and
Erdman SE: Beneficial bacteria stimulate host immune cells to
counteract dietary and genetic predisposition to mammary cancer in
mice. Int J Cancer. 135:529–540. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Van't Veer P, Dekker JM, Lamers JW, Kok
FJ, Schouten EG, Brants HA, Sturmans F and Hermus RJ: Consumption
of fermented milk products and breast cancer: A case-control study
in The Netherlands. Cancer Res. 49:4020–4023. 1989.PubMed/NCBI
|
|
44
|
Chagas CE, Rogero MM and Martini LA:
Evaluating the links between intake of milk/dairy products and
cancer. Nutr Rev. 70:294–300. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Soltan Dallal MM, Yazdi MH, Holakuyee M,
Hassan ZM, Abolhassani M and Mahdavi M: Lactobacillus casei
ssp.casei induced Th1 cytokine profile and natural killer cells
activity in invasive ductal carcinoma bearing mice. Iran J Allergy
Asthma Immunol. 11:183–189. 2012.PubMed/NCBI
|
|
46
|
Yazdi MH, Mahdavi M, Setayesh N, Esfandyar
M and Shahverdi AR: Selenium nanoparticle-enriched Lactobacillus
brevis causes more efficient immune responses in vivo and
reduces the liver metastasis in metastatic form of mouse breast
cancer. DARU. 21:332013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zamberi NR, Abu N, Mohamed NE, Nordin N,
Keong YS, Beh BK, Zakaria ZA, Nik Abdul Rahman NM and Alitheen NB:
The antimetastatic and antiangiogenesis effects of kefir water on
murine breast cancer cells. Integr Cancer Ther. 15:NP53–NP66. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Suez J, Zmora N, Segal E and Elinav E: The
pros, cons and many knowns of probiotics. Nat Med. 25:716–729.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Bafeta A, Koh M, Riveros C and Ravaud P:
Harms reporting in randomized controlled trials of interventions
aimed at modifying microbiota: A systematic review. Ann Intern Med.
169:240–247. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Mayo Clinic, . Engineering Gut Microbiome
to Target Breast Cancer. https://clinicaltrials.gov/ct2/show/NCT03358511NLM
identifier: NCT03358511. October 28–2020
|
|
51
|
Universidad de Granada, . Breast and
Cancer Its Relationship With the Microbiota (MICROMA). https://clinicaltrials.gov/ct2/show/NCT03885648NLM
identifier: NCT03885648. October 28–2020
|
|
52
|
Hackensack Meridian Health, . Gut and
Intratumoral Microbiome Effect on the Neoadjuvant
Chemotherapy-induced Immunosurveillance in Triple Negative Breast
Cancer. https://clinicaltrials.gov/ct2/show/NCT03586297NLM
identifier: NCT03586297. October 28–2020
|
|
53
|
First Affiliated Hospital of Harbin
Medical University, . Intestinal Microbiota of Breast Cancer
Patients Undergoing Chemotherapy. https://clinicaltrials.gov/ct2/show/NCT04138979NLM
identifier: ID: NCT04138979. October 28–2020
|
|
54
|
Lawson Health Research Institute, .
Probiotics and Breast Health. https://clinicaltrials.gov/ct2/show/NCT03290651NLM
identifier: NCT03290651. October 28–2020
|
|
55
|
Mayo Clinic, . A Pilot Trial of
Preoperative Oral Microbiota-based Investigational New Drug
(RBX7455). https://clinicaltrials.gov/ct2/show/NCT04139993NLM
identifier: NCT04139993. October 28–2020
|