Introduction
Lung cancer is one of the most common malignant
cancer types and is the leading cause of cancer-associated
mortality worldwide (1,2). In China, it is estimated that the number
of lung cancer cases reached 2.2 million and 1.8 million
individuals succumbed to lung cancer in 2020 (3). Moreover, lung cancer is typically
classified as small-cell lung cancer (SCLC) and non-SCLC (NSCLC)
(2), with NSCLC being the major type
of lung cancer and accounting for 85% of all cases (2). Despite significant improvements in NSCLC
treatment, including surgery (4),
chemotherapy (5,6), radiotherapy (7), targeted therapy (8–10) and
immunotherapy (11), the 5-year
survival rate of patients with NSCLC is only 15% (2,12). This is
due to the absence of early sensitivity and specificity diagnostic
biomarkers (13), drug resistance
(14) and distant metastasis
(15). Therefore, it is essential to
investigate the molecular mechanism underlying NSCLC
progression.
The transmembrane protein (TMEM) family includes
types of proteins that span the biological membranes, most of which
extend through the lipid bilayer of the plasma membrane, while
others are located in the cell membranes, such as mitochondrial,
endoplasmic reticulum, lysosomal, and Golgi membranes (16,17).
Although the functions of most TMEM proteins remain unknown, the
associations between TMEM proteins and tumor progression have been
a hotspot of research (17,18). Recently, several TMEM proteins have
been revealed to act as oncogenes or tumor suppressors in NSCLC,
and play a crucial role in the tumorigenesis and progression of
NSCLC (17,18). For example, TMEM209, localized to the
nuclear envelope and in the cytoplasm of lung cancer cells, is
upregulated in lung cancer and promotes cell proliferation and
tumor progression by interacting with nucleoporin 205 kDa (19). TMEM88 is a Wnt regulatory protein, and
its cytosolic form and the methylation of TMEM88 are highly
expressed in lung cancer, which accelerates the tumorigenesis and
progression of lung cancer (20,21).
Moreover, Liu and Zhu (22) reported
that TMEM106A was lowly expressed in NSCLC and suppressed cell
proliferation, migration and invasion, as well as induced cell
apoptosis via the PI3K/AKT/NF-κB signaling pathway. TMEM17 has also
been demonstrated to be aberrantly expressed in various solid tumor
types, such as lung (23) and breast
cancer (24). Furthermore, TMEM17 has
been revealed to be downregulated in NSCLC and to suppress cell
invasion and metastasis by inactivating the ERK/ribosomal protein
S6 kinase A1/snail family transcriptional repressor 1 (Snail1)
pathway (23). It has also been
reported that the TMEM39AS41 peptide contributed to cell death and
suppressed tumor growth via the suppression of
inflammation/autophagy pathways in lung cancer (25). In addition, our previous results
revealed that a TMEM229A Q200del mutation was associated with
histopathologic type and lymphatic metastasis in lung
adenocarcinoma (Zhang et al, unpublished data). However, to
the best of our knowledge, the expression level and biological role
of TMEM229A in NSCLC has not been previously studied.
The present study aimed to investigate the
expression level of TMEM229A in NSCLC tissues and cell lines, and
examine the relationship between TMEM229A expression and
clinicopathological features. To this end, the biological roles and
mechanisms of TMEM229A in the tumorigenesis of NSCLC were
determined, providing a potential therapeutic target for the
treatment of NSCLC.
Materials and methods
Ethical approval for the study
protocol
The present study was approved by the Ethics
Committee of The First People's Hospital of Huzhou, Huzhou, China
(approval no. 2019012), and informed consent was obtained from all
patients.
Patient samples
A total of 240 NSCLC and matched para-carcinoma
tissues (>5 cm away from the tumor) were obtained from patients
who underwent surgical resections at the First People's Hospital of
Huzhou (Huzhou, China) between January 2015 and December 2018. The
NSCLC patients consisted of 113 females and 127 males (age range,
27–82 years; mean age, 63). Among them, an NSCLC tissue microarray
(TMA), containing 189 pairs of tissues that were formalin-fixed
paraffin-embedded (FFPE), was used for immunohistochemical (IHC)
analysis, and 51 pairs of fresh tissues were used for reverse
transcription-quantitative (RT-q)PCR and western blot analysis.
These patients had not received chemotherapy or radiation therapy
prior to surgical resection. According to the 8th edition of Union
for International Cancer Control (UICC) (26,27), the
pathological diagnosis was confirmed and classified by two
certified pathologists (Professors Qilin Shi and Hui Xia,
Department of Pathology, The First People's Hospital of Huzhou).
All patients and clinical information were collected
retrospectively and analyzed in this study, including sex, age,
smoking history, tumor size, histological type, tumor
differentiation, lymph node metastasis, cancer thrombus and TNM
stage (Table I). Informed consent was
obtained from all patients.
 | Table I.Association between TMEM229A
expression and clinicopathological characteristics of patients with
non-small cell lung cancer. |
Table I.
Association between TMEM229A
expression and clinicopathological characteristics of patients with
non-small cell lung cancer.
|
|
| TMEM229A
expression |
|
|
|---|
|
|
|
|
|
|
|---|
| Variables | N=189 | Low | High | χ2 | P-value |
|---|
| Sex |
|
|
| 0.058 | 0.810 |
|
Female | 90 | 53 | 37 |
|
|
|
Male | 99 | 60 | 39 |
|
|
| Age |
|
|
| 1.883 | 0.170 |
|
>65 | 88 | 48 | 40 |
|
|
|
≤65 | 101 | 65 | 36 |
|
|
| Smoking
history |
|
|
| 0.690 | 0.406 |
|
Ever | 95 | 54 | 41 |
|
|
|
Never | 94 | 59 | 35 |
|
|
| Tumor size
(cm) |
|
|
| 0.009 | 0.925 |
|
>3.0 | 54 | 32 | 22 |
|
|
|
≤3.0 | 135 | 81 | 54 |
|
|
| Histological
type |
|
|
| 0.707 | 0.863 |
|
Squamous cell carcinoma | 47 | 28 | 19 |
|
|
|
Adenocarcinoma | 141 | 94 | 57 |
|
|
| Large
cell carcinoma | 1 | 1 | 0 |
|
|
| Tumor
differentiation |
|
|
| 3.627 | 0.043 |
|
Well/Moderate | 146 | 82 | 64 |
|
|
|
Poor | 43 | 31 | 12 |
|
|
| Lymphatic
invasion |
|
|
| 7.062 | 0.008 |
|
Present | 66 | 48 | 18 |
|
|
|
Absent | 123 | 65 | 58 |
|
|
| Cancer
thrombus |
|
|
| 6.354 | 0.012 |
|
Present | 78 | 55 | 23 |
|
|
|
Absent | 111 | 58 | 53 |
|
|
| Stage |
|
|
| 4.217 | 0.040 |
|
I+II | 115 | 62 | 53 |
|
|
|
III+IV | 74 | 51 | 23 |
|
|
Kaplan-Meier Plotter
Kaplan-Meier Plotter (http://kmplot.com/analysis/) is a public web server
used for the validation of the gene prognostic power using
expression data from independent datasets (28). All the related data were downloaded
from the Gene Expression Omnibus (GEO) database and The Cancer
Genome Atlas (TCGA) (28,29). A total of 1,233 samples of nine
independent datasets were integrated, including 1,100 samples in
GEO datasets [GSE4573 (30), GSE14814
(31), GSE8894 (32), GSE19188 (33), GSE3141 (34), GSE31210 (35), GSE29013 (36) and GSE37745 (37)] and 133 samples in TCGA datasets
(38). TMEM229A expression levels in
patients with lung adenocarcinoma (n=601) and patients with
squamous cell lung carcinoma (n=632) were collected using the
Kaplan-Meier Plotter online bioinformatics datasets. Additionally,
patients with lung adenocarcinoma and squamous cell carcinoma were
divided into low or high TMEM229A expression groups, according to
TMEM229A mRNA expression based on the median value. The overall
survival was analyzed using the Kaplan-Meier method.
Cell culture and treatment
The normal human lung epithelial cell (BEAS-2B) and
seven lung cancer cell lines (A549, H23, H226, 95D, H1975, PC-9 and
H1299) were purchased from the American Type Culture Collection.
Cells were cultured in DMEM (cat. no. SH30243.01; HyClone; Cytiva)
or RPMI-1640 medium (cat. no. SH30809.01; HyClone; Cytiva)
supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc.),
100 IU/ml penicillin and 100 µg/ml streptomycin (Sigma-Aldrich;
Merck KGaA) in a humidified incubator under 5% CO2 at
37°C. To inhibit the ERK signaling pathway, cells were treated with
100% dimethylsulfoxide (DMSO) as a negative control or PD98059 (ERK
inhibitor; 20 µM; Beyotime Institute of Biotechnology) for 2 h
prior to plasmid transfection in a humidified incubator under 5%
CO2 at 37°C.
Plasmid construction and cell
transfection
To construct the full-length human TMEM229A
proteins, the corresponding TMEM229A cDNA was cloned into a
pcDNA3.1/myc-his vector (cat. no. V800-20; Invitrogen;
Thermo Fisher Scientific, Inc.). In total, two specific small
interfering (si)RNAs of TMEM229A (20 µM) and scrambled siRNA
(referred to as si-Ctrl; 20 µM) were designed and synthesized by
Guangzhou RiboBio Co., Ltd.. Once cells reached 60–70% confluence,
cell transfections of different plasmids [pcDNA3.1/myc-his
(referred to as vector), pcDNA3.1-TMEM229A/myc-his (referred
to as overexpressed (OE)-TMEM229A), si-Ctrl, si-TMEM229A] were
carried out using Lipofectamine® 2000 reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's instructions. The medium was replaced with complete
medium and transfection was performed in a humidified incubator
under 5% CO2 at 37°C for 6 h. In a 6-well plate, 2 µg
vector or OE-TMEM229A and 50 nM si-Ctrl or si-TMEM229A plasmids
were added to each well. At 24 h after transfection, cells were
harvested for the subsequent experiments. The siRNA sequences used
in the experiment were as follows: si-TMEM229A#1 forward,
5′-GGAUGCGCCUCUACUUCUAdTdT-3′ and reverse,
5′-UAGAAGUAGAGGCGCAUCCdTdT-3′; si-TMEM229A#2 forward,
5′-CCUUCGUCUUCAAUUUCCUdTdT-3′ and reverse,
5′-AGGAAAUUGAAGACGAAGGdTdT-3′; and si-Ctrl forward,
5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse,
5′-ACGUGACACGUUCGGAGAATT-3′.
Real-time cellular analysis
(RTCA)
The RTCA xCELLLigence system (ACEA Biosciences,
Inc.; Agilent Technologies, Inc.) was used to uninterruptedly
monitor cell morphology, proliferation and migration in
vitro in a non-invasive manner, as previously described
(39,40). A cell index was used to indicate the
cell number and cell adhesion. As cells adhered to the surface of
an E-plate and influenced the electrical impedance across the
array, the xCELLLigence system provided an electronic record and
converted it into the cell index. Cell proliferation and migration
assays were performed according to the manufacturer's instructions
(ACEA Biosciences, Inc.; Agilent Technologies, Inc.).
Briefly, for cell proliferation assays, 50 µl
culture medium was added to measure the background, and then
6×103 A549 cells, 1×104 H23 cells or
1×104 H1299 cells transfected with different plasmids
were mixed with 100 µl culture medium and seeded into the E-plate.
For cell migration and invasion assays, 165 µl culture medium was
added to the lower chamber and 30 µl serum-free culture medium was
added to the upper chamber, and then the E-plate was left to stand
for 1 h in a humidified incubator under 5% CO2 at 37°C.
The cells were mixed with 100 µl serum-free culture medium and
seeded into the E-plate. The data were recorded using xCELLLigence
software 2.0 (ACEA Biosciences, Inc.; Agilent Technologies, Inc.)
and analyzed using GraphPad Prism 5.0 (GraphPad Software,
Inc.).
Cell proliferation assay
Cell proliferation was also assessed using a Cell
Counting Kit-8 (CCK-8; Beyotime Institute of Biotechnology) assay,
according to the manufacturer's instructions. Briefly,
6×103 A549, 1×104 H23 or 1×104
H1299 cells transfected with different plasmids were seeded into
96-well plates and cultured for 24, 48 and 72 h at 37°C, followed
by incubation with 10 µl of CCK-8 reagent for 1 h. The absorbance
values were measured using a Spectra Max 190 reader (Molecular
Devices, LLC) at 450 nm.
Clonogenic assay
The transfected A549, H23 or H1299 cells were seeded
into 6-well plates at a density of 1×103 cells/well and
cultured with culture medium containing 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.) for 10 days. Cells were fixed in 4%
paraformaldehyde for 30 min at room temperature and followed by
staining with 1% crystal violet stain solution (Beyotime Institute
of Biotechnology) overnight at room temperature. Cell colonies were
imaged under an inverted fluorescence microscope with a
magnification of ×20 magnification (ZEISS Axio Vert.A1; Carl Zeiss
AG) and counted if there were >50 cells in the assay.
Soft agar assay
The cell proliferative ability was also examined
using a soft agar assay, as previously described (41,42). At 24
h-post-transfection, A549, H23 or H1299 cells were plated in
24-well plates, with the bottom layer containing 0.7% low-melting
point agarose (cat. no. 16520100; Invitrogen; Thermo Fisher
Scientific, Inc.). Cells (1,000-2,000 per well) were seeded into
the medium with 0.35% agarose. After 14 days of incubation in a
humidified incubator under 5% CO2 at 37°C, cell colonies
were imaged and counted if there were >50 cells in the assay
under an inverted fluorescence microscope with a magnification of
×20 (ZEISS Axio Vert.A1; Carl Zeiss AG).
Transwell assay
Cell migratory and invasive abilities were
determined using a Transwell plate chamber (8 µm; Corning; Corning,
Inc.), according to the manufacturer's protocols. For the migration
assay, the transfected cells (5×104) were mixed in 100
µl serum-free culture medium and seeded into the upper chamber, and
600 µl complete culture medium was added to the lower chamber. For
the invasion assay, the upper chamber was firstly coated with
Matrigel® Matrix Basement Membrane (cat. no. 356234; BD
Biosciences) for 4 h in a humidified incubator under 5%
CO2 at 37°C. Then, cells were seeded into the upper
chamber, and complete medium was added to the lower chamber.
Following 48 h incubation in a humidified incubator under 5%
CO2 at 37°C, the cells in the lower chamber were fixed
in 4% paraformaldehyde for 30 min at room temperature and stained
with 1% crystal violet stain solution (Beyotime Institute of
Biotechnology) overnight at room temperature. The images were
captured and the cells were decolorized with 30% acetic acid. The
absorbance values were measured using a Spectra Max 190 reader
(Molecular Devices, LLC) at 570 nm.
RNA extraction and RT-qPCR
Total RNA from tissues and cells was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.), as previously described (43).
Briefly, samples were treated with 20% chloroform, followed by
incubation at room temperature for 5 min. Then, samples were
centrifuged and the aqueous phase was transferred to a new tube,
followed by the addition of an equal amount of isopropanol and
incubation at room temperature for 10 min. The pellets were
dissolved in nuclease-free water. cDNA was reverse transcribed
using the PrimeScript RT reagent kit (Takara Biotechnology Co.,
Ltd.), according to the manufacturer's protocol. RT-qPCR was
performed using UltraSYBR Green PCR Master mix (cat. no. CW0957H;
CWBio) on an ABI 7500 system (Applied Biosystems; Thermo Fisher
Scientific, Inc.). The thermocycling conditions used for the qPCR
were as follows: Initial denaturation at 95°C for 10 min, followed
by 40 cycles at 95°C for 15 sec and 60°C for 1 min. Gene expression
was normalized to 18S ribosomal RNA (18sRNA), and the relative
expression level was calculated using the 2−ΔΔCq method
(44). The primer sequences used in
the experiment were as follows: TMEM229A forward,
5′-CGACCTACCCCGCTTTCTTTT-3′ and reverse,
5′-GCTCCCACCGTAGATGAAGAT-3′; N-cadherin forward,
5′-AGCCAACCTTAACTGAGGAGT-3′ and reverse,
5′-GGCAAGTTGATTGGAGGGATG-3′; E-cadherin forward,
5′-ATTTTTCCCTCGACACCCGAT-3′ and reverse, 5′-TCCCAGGCGTAGACCAAGA-3′;
MMP2 forward, 5′-GATACCCCTTTGACGGTAAGGA-3′ and reverse,
5′-CCTTCTCCCAAGGTCCATAGC-3′; Snail1 forward,
5′-ACTGCAACAAGGAATACCTCAG-3′ and reverse,
5′-GCACTGGTACTTCTTGACATCTG-3′; and 18sRNA forward,
5′-GTAACCCGTTGAACCCCATT-3′ and reverse,
5′-CCATCCAATCGGTAGTAGCG-3′.
Western blotting
Tissues or cells were lysed in RIPA buffer (cat. no.
P0013B; Beyotime Institute of Biotechnology) containing protease
and phosphatase inhibitors (Beyotime Institute of Biotechnology).
The total protein concentration was quantified with a BCA assay kit
(Beyotime Institute of Biotechnology). An equal amount of total
protein (30 µg) was loaded and separated using 8–12% SDS-PAGE. The
proteins were transferred onto 0.45-µm PVDF membranes (EMD
Millipore). The membranes were blocked with 5% non-fat milk at room
temperature for 1 h, followed by incubation with primary antibodies
overnight at 4°C. The membranes were subsequently washed with PBS
(cat. no. SH30256.01; HyClone; Cytiva) containing 0.1% Tween-20,
and incubated with the corresponding HRP-conjugated secondary
antibodies [(goat-anti mouse IgG (H+L) antibody; 1:5,000; cat. no.
A0216) or (goat-anti rabbit IgG (H+L) antibody; 1:5,000; cat. no.
A0208) both from Beyotime Institute of Biotechnology] at room
temperature for 1 h. The images were visualized using an ECL
reagent (cat. no. P0018S; Beyotime Institute of Biotechnology) and
measured using a Tanon 5200 system (Tanon Science & Technology
Co., Ltd.). The relative expression level of proteins was
normalized to β-actin using ImageJ vl.6.0 software (National
Institutes of Health).
The primary antibodies used in the experiment were
as follows: Anti-TMEM229A (1:500; cat. no. ab107780; Abcam);
anti-E-cadherin (1:1,000; cat. no. 20874-1-AP; ProteinTech Group,
Inc.); anti-N-cadherin (1:1,000; cat. no. 22018-1-AP; ProteinTech
Group, Inc.); anti-MMP2 (1:1,000; cat. no. 10373-2-AP; ProteinTech
Group, Inc.); anti-Snail1 (1:1,000; cat. no. 13099-1-AP;
ProteinTech Group, Inc.); anti-ERK1/2 (1:2,000; cat. no. 4695; Cell
Signaling Technology, Inc.); anti-phosphorylated (p)-ERK1/2
(1:2,000; cat. no. 4370; Cell Signaling Technology, Inc.); anti-AKT
(1:2,000; cat. no. 4691; Cell Signaling Technology, Inc.);
anti-p-AKT (1:2,000; Ser473; cat. no. 4060; Cell Signaling
Technology, Inc.); anti-p-p38 (1:2,000; Thr180/Tyr182; cat. no.
4631; Cell Signaling Technology, Inc.); anti-p-JNK (1:2,000;
Thr183/Tyr185; cat. no. 4668; Cell Signaling Technology, Inc.);
anti-p-p65 (1:1,000; Ser536; cat. no. 3033; Cell Signaling
Technology, Inc.), and anti-β-actin (1:5,000; cat. no. M1210-2;
Hangzhou HuaAn Biotechnology Co., Ltd.).
IHC staining
The tissues were fixed with 10% formalin overnight
at room temperature, and the FFPE tissues were sliced into 5-µm
thickness sections, followed by dewaxing, hydrating and
inactivating of endogenous peroxidase. Sections were then incubated
with a polyclonal rabbit anti-TMEM229A antibody (1:500; cat. no.
PA5-63294; Invitrogen; Thermo Fisher Scientific, Inc.) overnight at
4°C in a humidified chamber. The sections were subsequently washed
with PBS (cat. no. SH30256.01; HyClone; Cytiva), and incubated with
the SP Rabbit & Mouse HRP-conjugated secondary antibody kit
(cat. no. CW2069S; CWBio) at room temperature for 1 h. Next, the
sections were incubated with DAB (included in the aforementioned
kit) at room temperature for 20 min. After washing with water, the
sections were counterstained with hematoxylin staining solution
(cat. no. C0107; Beyotime Institute of Biotechnology) at room
temperature for 5 min. The TMA sections were further imaged and
digitally scanned at a magnification of ×400 using a KF-PRO-005
platform (Ningbo Jiangfeng Bio-information Technology Co., Ltd.)
into whole slide digital images. The scoring of the slides was
conducted using HALO Multiplex IHC analysis software version
v3.1.1076.308 (Indica Labs). The intensity of TMEM229A was scored
as 1 (absent or weak), 2 (moderate) or 3 (high). The percentage of
positive cells was assigned as 1 (<25%), 2 (25–50%) and 3
(>50%). The score of each slice was multiplied to acquire a
final score of 1–9. A composite score of <3 was defined as
TMEM229A low expression, and a score of ≥3 was defined as TMEM229A
high expression.
Statistical analysis
All data are presented as the mean ± SEM of three
independent experiments. The associations between TMEM229A
expression and clinicopathological characteristics of patients were
analyzed using the χ2 test, Fisher's exact test and
Pearson rank correlation analysis. Kaplan-Meier survival analysis
was performed using the log-rank test. Statistical significance
between the groups was determined using SPSS 21.0 software (IBM
Corp.) with an unpaired Student's t-test or one-way ANOVA followed
by a Tukey's post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
TMEM229A is lowly expressed in
NSCLC
To determine the expression pattern of TMEM229A in
NSCLC, the mRNA expression level of TMEM229A was measured in 51
paired NSCLC and corresponding healthy tissues via RT-qPCR.
Downregulation of TMEM229A was observed in 40 pairs of tissues,
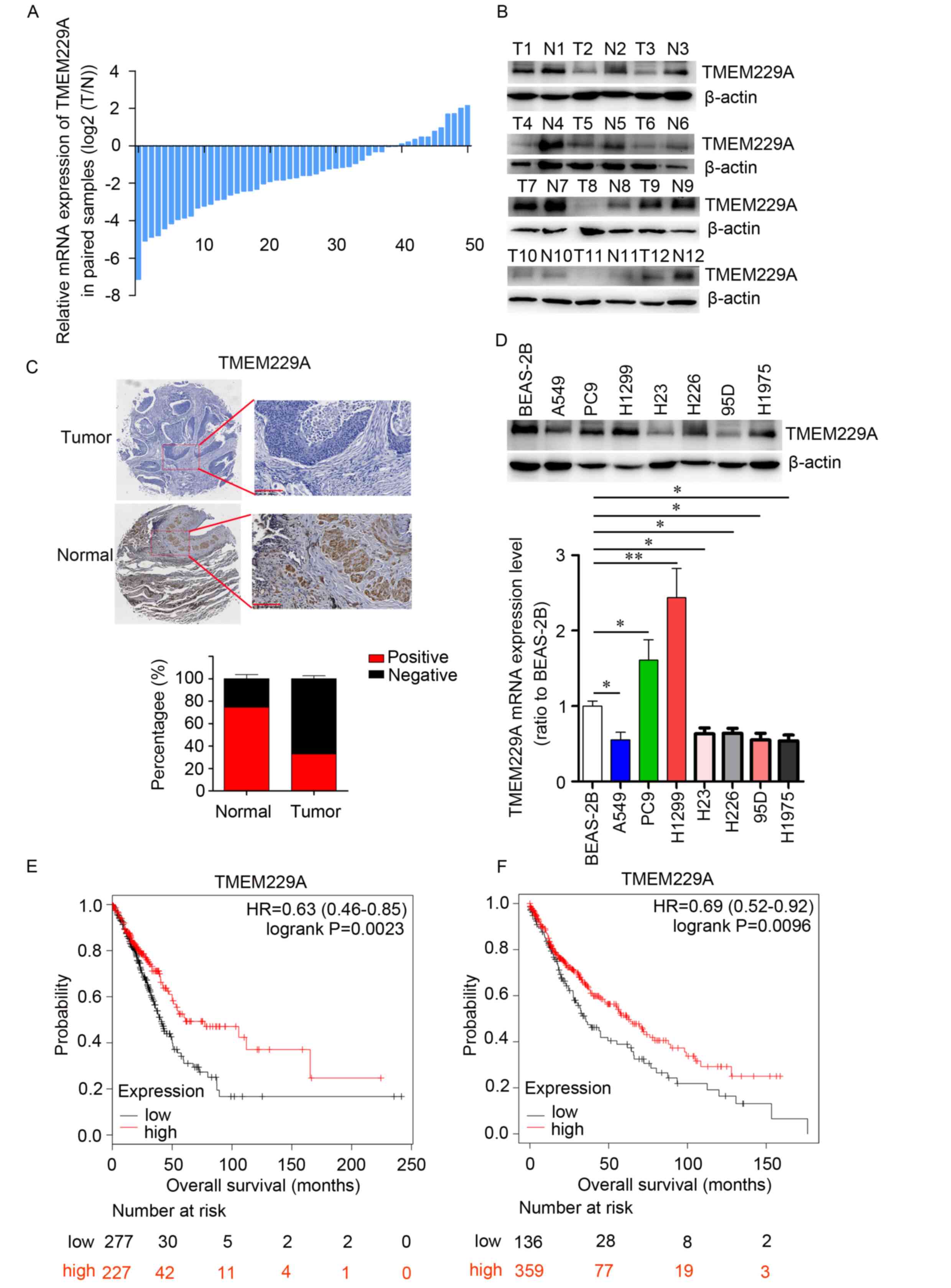
accounting for 78% of the total specimens examined (Fig. 1A). Consistent with the mRNA expression
pattern, a reduced protein expression level of TMEM229A was
identified in NSCLC tissues compared with the matched adjacent
healthy tissues in 10/12 paired samples, as determined using
western blotting (Fig. 1B). The low
expression of TMEM229A protein in NSCLC tissues was further
confirmed via IHC staining analysis (Fig.
1C). Moreover, TMEM229A expression was measured in BEAS-2B
cells and seven NSCLC cell lines, including A549, H23, H1299, H226,
95D, PC-9 and H1975. The mRNA and protein expression levels of
TMEM229A were decreased in several NSCLC cell lines (A549, H23,
95D, H226 and H1975) compared with BEAS-2B cells (Fig. 1D). Owing to the higher invasiveness of
95D, A549 and H23 were selected for the follow-up overexpressing
experiments. Collectively, these data indicated that TMEM229A
expression was downregulated in NSCLC.
Correlation between TMEM229A
expression and the clinicopathological characteristics of
NSCLC
To evaluate the clinical significance of TMEM229A in
NSCLC, a NSCLC TMA containing 189 specimens was stained with
TMEM229A antibody and scored in a standard manner, as previously
described (45). The results
indicated that 113 patients were classified into the low TMEM229A
expression group, while 76 patients were classified into the high
TMEM229A expression group (Fig. 1C
and Table I). Further analysis of
TMEM229A expression and the clinicopathologic characteristics
revealed that low TMEM229A expression was associated with tumor
differentiation (P=0.043), lymph node metastasis (P=0.008), cancer
thrombus (P=0.012) and TNM stage (P=0.04). However, there were no
significant associations with sex, age, smoking history, tumor size
and histological type (Table I).
Subsequently, Kaplan-Meier Plotter was used to study
the association between TMEM229A expression level and overall
survival in patients with NSCLC. All the related data were
downloaded from the GEO database, the EGA and TCGA. Patients with
lung adenocarcinoma and those with squamous cell carcinoma were
divided into low- or high-TMEM229A expression groups, according to
TMEM229A mRNA expression. The results demonstrated that lung
adenocarcinoma cases and squamous cell lung carcinoma cases with
low TMEM229A expression had a poor prognosis with a shorter overall
survival (P=0.0023 and P=0.0096, respectively; Fig. 1E and F). Thus, these data indicated
that low TMEM229A expression was associated with poor prognosis in
patients with lung adenocarcinoma and in patients with squamous
cell lung carcinoma.
TMEM229A overexpression suppresses
NSCLC cell proliferation, migration and invasion in vitro
To investigate the effect of TMEM229A on cell
proliferation, TMEM229A was overexpressed in A549 and H23 cells
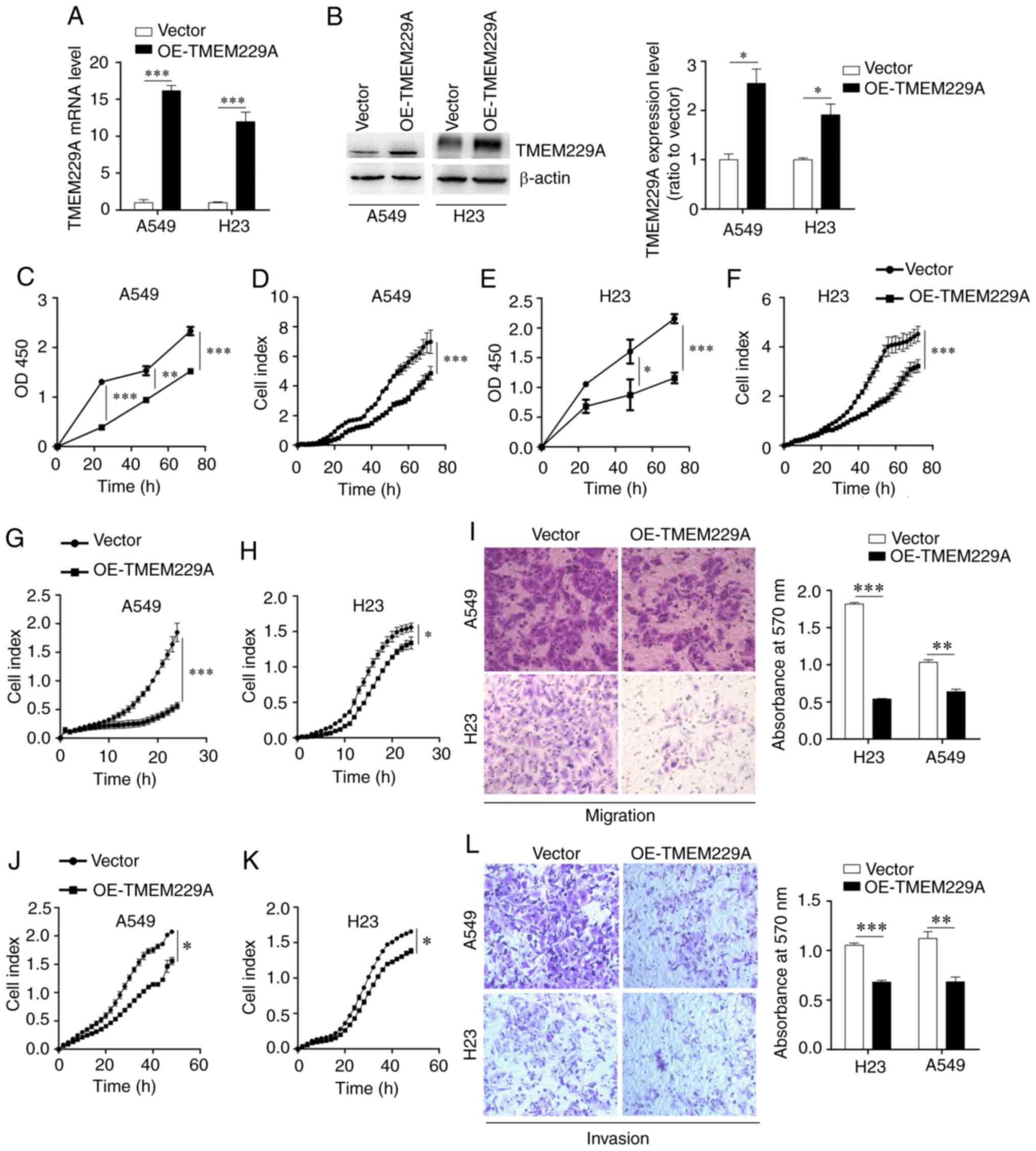
(Fig. 2A and B), which had relatively
low expression levels of TMEM229A. Subsequently, the proliferative
capacity of TMEM229A-overexpressing cells was measured using CCK-8
and RTCA assays. The results demonstrated that overexpression of
TMEM229A significantly inhibited A549 and H23 cell proliferation in
a time-dependent manner (Fig.
2C-F).
In addition, two TMEM229A siRNAs and si-Ctrl were
transfected into H1299 cells, which demonstrated relatively high
expression levels of TMEM229A. The RT-qPCR and western blot results
revealed that TMEM229A expression was markedly decreased in the
si-TMEM229A-transfected H1299 cells compared with
si-Ctrl-transfected cells (Fig. S1A and
B), and TMEM229A knockdown significantly promoted H1299 cell
proliferation (Fig. S1C and D).
Furthermore, the effects of TMEM229A overexpression or knockdown on
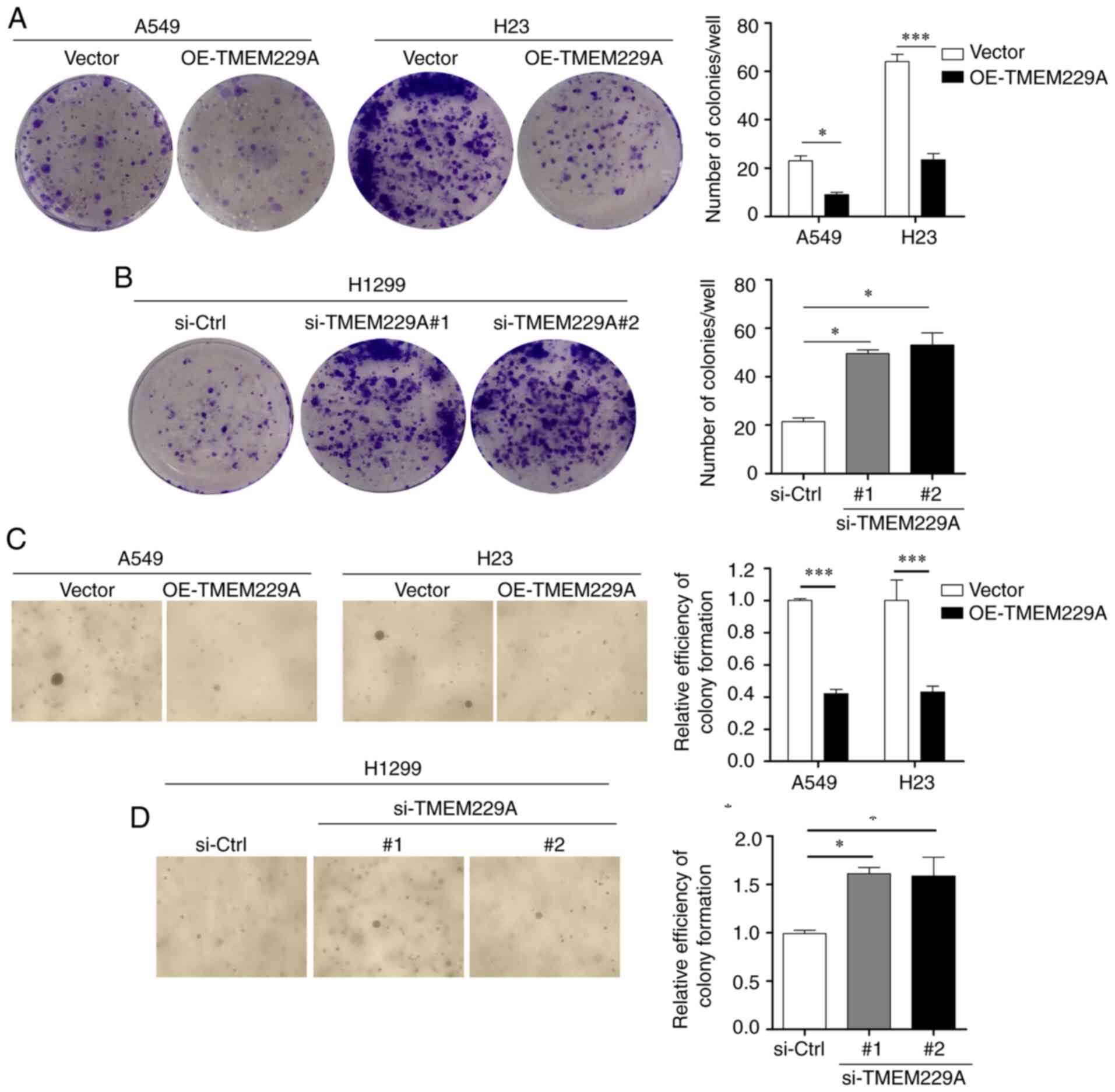
cell proliferation were verified using cell colony and soft agar
assays. The data indicated that overexpression of TMEM229A
suppressed the number of cell colonies, while TMEM229A knockdown
promoted these (Fig. 3A-D). Overall,
these findings indicated that TMEM229A plays a crucial role in the
proliferation of NSCLC cells.
The potential influence of TMEM229A overexpression
or knockdown on cell migration and invasion was then examined. It
was revealed that overexpression of TMEM229A significantly
suppressed the migration and invasion of NSCLC cells, as determined
using Transwell and RTCA assays (Fig.
2G-L), whereas knockdown of TMEM229A promoted the migratory and
invasive abilities of H1299 cells compared with those in the
control group (Fig. S1E-G).
TMEM229A regulates the
epithelial-mesenchymal transition (EMT) phenotype
Considering the critical role of EMT in the
metastasis of NSCLC (46), the
present study examined the role of TMEM229A on EMT, and detected
the expression levels of E-cadherin, N-cadherin, MMP2 and Snail1.
As demonstrated in Fig. 4A and B,
TMEM229A overexpression significantly increased E-cadherin
expression and reduced N-cadherin, Snail1 and MMP2 expression,
while TMEM229A knockdown exerted the opposite effects. These data
indicated that TMEM229A influenced cell migration and invasion by
modulating the EMT phenotype.
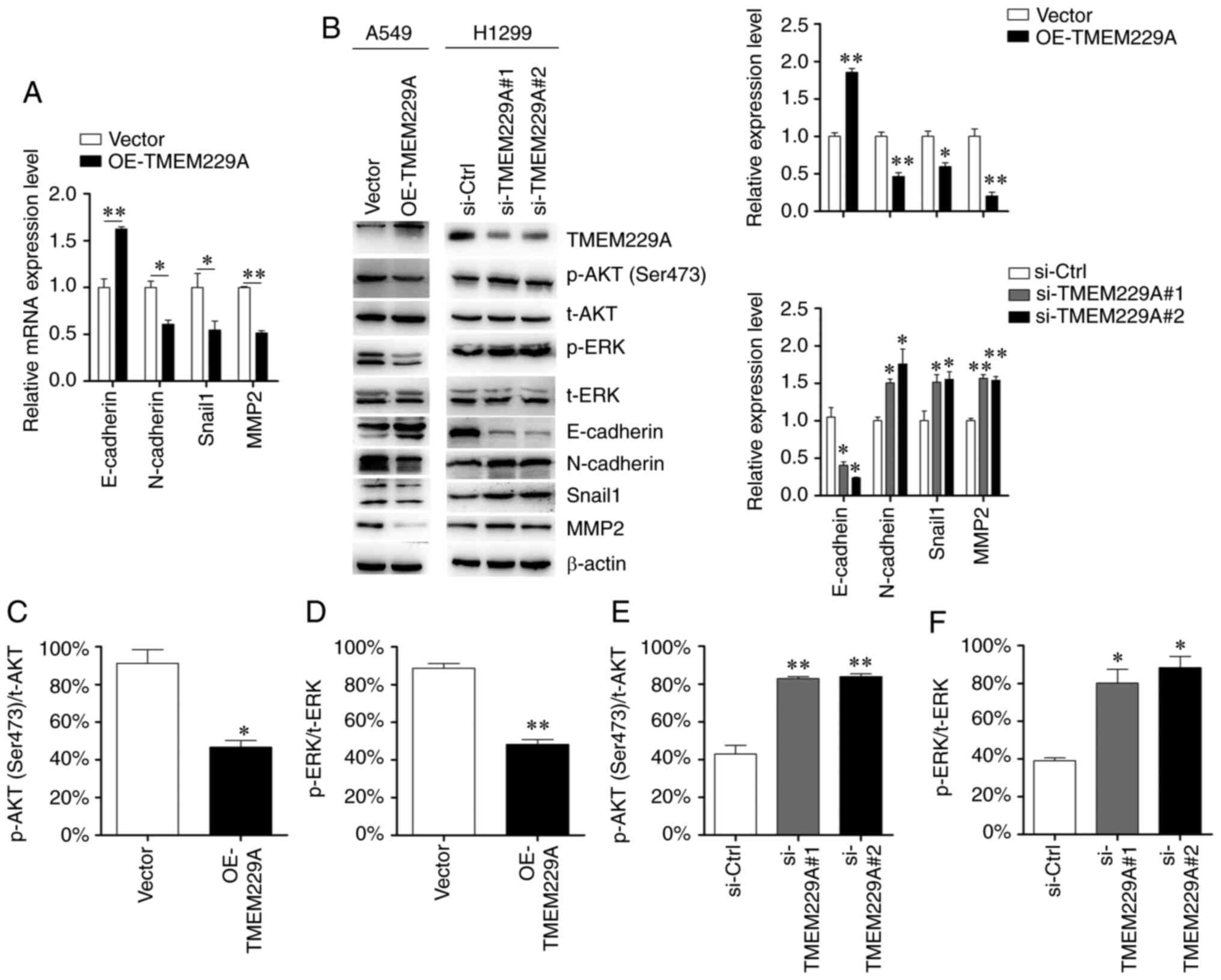 | Figure 4.TMEM229A regulates
epithelial-mesenchymal transition via p-ERK and p-AKT. (A) mRNA
expression levels of E-cadherin, N-cadherin, MMP2 and Snail1 were
detected using reverse transcription-quantitative PCR. (B-F)
Protein expression levels of E-cadherin, N-cadherin, MMP2, Snail1,
p-ERK, p-AKT (Ser 473), t-ERK and t-AKT were detected using western
blot analysis. The ratios of p-AKT (Ser473)/t-AKT and p-ERK/t-ERK
were analyzed. *P<0.05 and **P<0.01. TMEM229A, transmembrane
protein 229A; OE, overexpressed; Snail1, snail family
transcriptional repressor 1; si-Ctrl, scrambled siRNA; si-, small
interfering; p-, phosphorylated; t-, total. |
TMEM229A knockdown promotes ERK and
AKT phosphorylation
To investigate the mechanisms via which TMEM229A
promotes NSCLC progression, key factors of common signaling
pathways were screened in NSCLC cells, including p38/MAPK, ERK,
AKT, JNK and NF-κB. The expression levels of p-p38, p-ERK, p-AKT
(Ser473), p-JNK and p-p65 were primarily analyzed in the present
study. It was revealed that knockdown of TMEM229A upregulated p-ERK
and p-AKT (Ser473) expression in NSCLC cells, while overexpression
of TMEM229A exerted the opposite effect (Fig. 4B-F). Moreover, other signaling
pathways showed no obvious changes (Fig.
S2).
Subsequently, si-TMEM229A-transfected cells were
treated with the ERK inhibitor PD98059, and the results
demonstrated that PD98059 suppressed the TMEM229A-induced ERK
phosphorylation, and partly reversed the expression levels of
E-cadherin, N-cadherin, MMP2 and Snail1 (Fig. 5A). Moreover, the Transwell assay
results indicated that the promoting effect on cell invasiveness
caused by TMEM229A knockdown was also partially reversed by PD98059
(Fig. 5B).
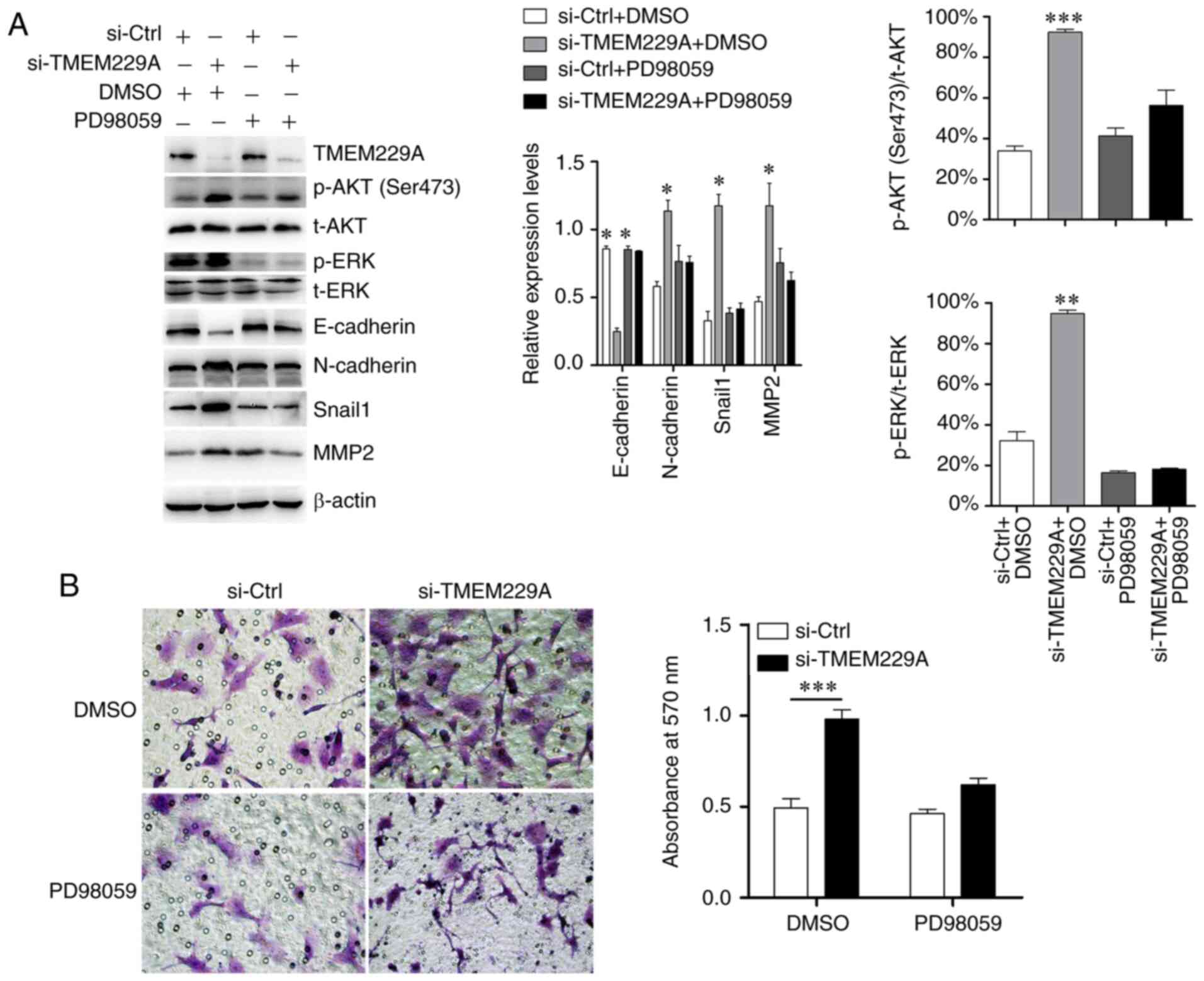 | Figure 5.TMEM229A inhibits NSCLC progression
partly via the ERK signaling pathway. (A) Western blotting
examination of p-ERK, E-cadherin, N-cadherin, Snail1 and MMP2
expression after TMEM229A knockdown with or without PD98059
treatment (ERK inhibitor; 20 µM). The ratios of p-AKT
(Ser473)/t-AKT and p-ERK/t-ERK were analyzed. (B) Cell invasion was
measured using a Transwell assay in H1299 cells after knockdown of
TMEM229A with or without PD98059 treatment (ERK inhibitor; 20 µM).
*P<0.05, **P<0.01 and ***P<0.001. TMEM229A, transmembrane
protein 229A; NSCLC, non-small cell lung cancer; si-Ctrl, scrambled
siRNA; p-, phosphorylated; t-, total; Snail1, snail family
transcriptional repressor 1; si-, small interfering. |
Discussion
TMEM229A is a seven-transmembrane protein with
poorly defined biology. At present, research has revealed that
TMEM229A serves a critical role in the development and
differentiation of teeth (47).
However, the current understanding of the expression and possible
mechanism of TMEM229A is lacking, particularly with regard to that
in cancer. Therefore, the present study aimed to investigate the
expression level and biological role of TMEM229A in NSCLC. It was
identified that TMEM229A was lowly expressed in NSCLC tissues and
several cell lines, and low TMEM229A expression was positively
associated with poor tumor differentiation, positive lymph node
metastasis, positive cancer thrombus and advanced stage of patients
with NSCLC, suggesting that TMEM229A may exert a suppressive role
in NSCLC development.
Metastasis is the major cause of the poor prognosis
of NSCLC (48). Accumulating evidence
has revealed that multiple TMEM proteins, such as TMEM209A
(19), TMEM88 (20,21),
TMEM106A (22), TMEM17 (23), TMEM98 (49) and TMEM48 (50), can serve as prognostic markers for
NSCLC. In the present study, survival analysis using the
Kaplan-Meier Plotter online bioinformatics datasets revealed that
patients with low TMEM229A expression had a poorer prognosis
compared with those with high TMEM229A expression, suggesting that
TMEM229A could be considered as a prognostic marker for patients
with NSCLC.
EMT also plays a critical role in the NSCLC
metastasis, and its associated proteins and MMPs are considered as
key regulators in NSCLC invasion and metastasis (51–54). The
present results demonstrated that overexpression of TMEM229A
suppressed cell migration and invasion, and the EMT phenotype, as
well as upregulated E-cadherin expression and downregulated
N-cadherin, Snail1 and MMP2 expression in vitro.
Collectively, these results indicated that TMEM229A promoted the
progression of NSCLC by regulating the EMT phenotype.
The ERK signaling pathway exerts an oncogenic role
in regulating the proliferation and migration of NSCLC cells
(55). Moreover, a previous study
reported that TMEM17 suppressed NSCLC progression via the ERK
signaling pathway (23). The present
study primarily screened for key factors of common signaling
pathways (56–58), and revealed that overexpression of
TMEM229A reduced the expression levels of p-ERK and p-AKT.
Furthermore, other signaling pathways demonstrated no obvious
changes. It was also identified that the promoting effect on cell
invasiveness caused by TMEM229A knockdown was partly reversed by
PD98059, but PD98059 alone did not affect cell invasive capacity.
These results were similar to those of a previous study (23), indicating that TMEM229A suppresses
NSCLC progression partly by inactivating the ERK pathway.
However, the present study had certain limitations.
For example, the mechanism of TMEM229A and the ERK signaling
pathway was not fully determined. Thus, future studies will further
investigate the underlying mechanisms of TMEM229A on NSCLC
progression.
In summary, the present study demonstrated the
suppressive role of TMEM229A in NSCLC development and progression.
Mechanistically, TMEM229A inhibited NSCLC tumorigenesis, at least
partly, via the inhibition of the ERK signaling pathway. These
results suggest that TMEM229A may be a potential therapeutic target
for NSCLC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was kindly supported by Zhejiang
Provincial Natural Science Foundation of China under grant no.
LGF20H160016, the Scientific Technology Projects of Health and
Medicine of Zhejiang Province under grant nos. WKJ-ZJ-1830 and
2019KY207, and Huzhou Science and Technology Fund under grant no.
2020GYB01.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
XW, HY and XZ designed and conceived the study. XZ,
YH and YJ performed the experiments. XZ, YB, DX and QC analyzed the
data. XZ wrote and DX revised the manuscript. All authors have read
the final manuscript.
Ethics approval and consent to
participate
The study was approved (approval no. 2019012) by the
Ethics Committee of The First People's Hospital of Huzhou, Huzhou,
China. Written informed consent was provided by all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2020. CA Cancer J Clin. 70:7–30. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cao W, Chen HD, Yu YW, Li N and Chen WQ:
Changing profiles of cancer burden worldwide and in China: A
secondary analysis of the global cancer statistics 2020. Chin Med J
(Engl). 134:783–791. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Whitson BA, Groth SS, Duval SJ, Swanson SJ
and Maddaus MA: Surgery for early-stage non-small cell lung cancer:
A systematic review of the video-assisted thoracoscopic surgery
versus thoracotomy approaches to lobectomy. Ann Thorac Surg.
86:2008–2018. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Horinouchi H, Atagi S, Oizumi S, Ohashi K,
Kato T, Kozuki T, Seike M, Sone T, Sobue T, Tokito T, et al:
Real-world outcomes of chemoradiotherapy for unresectable stage III
non-small cell lung cancer: The SOLUTION study. Cancer Med.
9:6597–6608. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tachihara M, Dokuni R, Okuno K, Tokunaga
S, Nakata K, Katsurada N, Yamamoto M, Nagano T, Kobayashi K, Tanaka
Y, et al: Phase II study of adjuvant chemotherapy with pemetrexed
and cisplatin with a short hydration method for completely resected
nonsquamous non-small cell lung cancer. Thorac Cancer.
11:2536–2541. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Delaney GP and Barton MB: Evidence-based
estimates of the demand for radiotherapy. Clin Oncol (R Coll
Radiol). 27:70–76. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Garg A, Batra U, Choudhary P, Jain D,
Khurana S, Malik PS, Muthu V, Prasad KT, Singh N, Suri T and Mohan
A: Clinical predictors of response to EGFR-tyrosine kinase
inhibitors in EGFR-mutated non-small cell lung cancer: A real-world
multicentric cohort analysis from India. Curr Probl Cancer.
44:1005702020. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu C, Yu H, Chang J, Chen H, Li Y, Zhao
W, Zhao K, Zhu Z, Sun S, Fan M and Wang J: Crizotinib in Chinese
patients with ROS1-rearranged advanced non-small-cell lung cancer
in routine clinical practice. Target Oncol. 14:315–323. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Y, Zeng L, Zhou C, Li Y, Wu L, Xia
C, Jiang W, Hu Y, Liao D, Xiao L, et al: Detection of
nonreciprocal/reciprocal ALK translocation as poor predictive
marker in patients with first-line crizotinib-treated
ALK-rearranged NSCLC. J Thorac Oncol. 15:1027–1036. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bylicki O, Paleiron N, Margery J, Guisier
F, Vergnenegre A, Robinet G, Auliac JB, Gervais R and Chouaid C:
Targeting the PD-1/PD-L1 immune checkpoint in EGFR-mutated or
ALK-translocated non-small-cell lung cancer. Target Oncol.
12:563–569. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Balata H, Fong KM, Hendriks LE, Lam S,
Ostroff JS, Peled N, Wu N and Aggarwal C: Prevention and early
detection for NSCLC: Advances in thoracic oncology 2018. J Thorac
Oncol. 14:1513–1527. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yang SR, Schultheis AM, Yu H, Mandelker D,
Ladanyi M and Büttner R: Precision medicine in non-small cell lung
cancer: Current applications and future directions. Semin Cancer
Biol. Jul 27–2020.(Epub ahead of print). doi:
10.1016/j.semcancer.2020.07.009. View Article : Google Scholar
|
|
14
|
Imakita T, Matsumoto H, Hirano K, Morisawa
T, Sakurai A and Kataoka Y: Impact on prognosis of rebiopsy in
advanced non-small cell lung cancer patients after epidermal growth
factor receptor-tyrosine kinase inhibitor treatment: A systematic
review. BMC Cancer. 19:1052019. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hsu PC, Tian B, Yang YL, Wang YC, Liu S,
Urisman A, Yang CT, Xu Z, Jablons DM and You L: Cucurbitacin E
inhibits the Yes-associated protein signaling pathway and
suppresses brain metastasis of human non-small cell lung cancer in
a murine model. Oncol Rep. 42:697–707. 2019.PubMed/NCBI
|
|
16
|
Rao J, Wu X, Zhou X, Deng R and Ma Y:
TMEM205 is an independent prognostic factor and is associated with
immune cell infiltrates in hepatocellular carcinoma. Front Genet.
11:5757762020. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schmit K and Michiels C: TMEM proteins in
cancer: A review. Front Pharmacol. 9:13452018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Marx S, Dal Maso T, Chen JW, Bury M,
Wouters J, Michiels C and Le Calvé B: Transmembrane (TMEM) protein
family members: Poorly characterized even if essential for the
metastatic process. Semin Cancer Biol. 60:96–106. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Fujitomo T, Daigo Y, Matsuda K, Ueda K and
Nakamura Y: Critical function for nuclear envelope protein TMEM209
in human pulmonary carcinogenesis. Cancer Res. 72:4110–4118. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang X, Yu X, Jiang G, Miao Y, Wang L,
Zhang Y, Liu Y, Fan C, Lin X, Dong Q, et al: Cytosolic TMEM88
promotes invasion and metastasis in lung cancer cells by binding
DVLS. Cancer Res. 75:4527–4537. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ma R, Feng N, Yu X, Lin H, Zhang X, Shi O,
Zhang H, Zhang S, Li L, Zheng M, et al: Promoter methylation of
Wnt/β-Catenin signal inhibitor TMEM88 is associated with
unfavorable prognosis of non-small cell lung cancer. Cancer Biol
Med. 14:377–386. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu J and Zhu H: TMEM106A inhibits cell
proliferation, migration, and induces apoptosis of lung cancer
cells. J Cell Biochem. Nov 19–2018.(Epub ahead of print). doi:
10.1002/jcb.28057.
|
|
23
|
Zhang X, Zhang Y, Miao Y, Zhou H, Jiang G
and Wang E: TMEM17 depresses invasion and metastasis in lung cancer
cells via ERK signaling pathway. Oncotarget. 8:70685–70694. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhao Y, Song K, Zhang Y, Xu H, Zhang X,
Wang L, Fan C, Jiang G and Wang E: TMEM17 promotes malignant
progression of breast cancer via AKT/GSK3β signaling. Cancer Manag
Res. 10:2419–2428. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Park S, Kim M, Hong Y, Lee H, Tran Q, Kim
C, Kwon SH, Park J, Park J and Kim SH: Myristoylated TMEM39AS41, a
cell-permeable peptide, causes lung cancer cell death. Toxicol Res.
36:123–130. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Asamura H, Chansky K, Crowley J, Goldstraw
P, Rusch VW, Vansteenkiste JF, Watanabe H, Wu YL, Zielinski M, Ball
D, et al: The international association for the study of lung
cancer lung cancer staging project: Proposals for the revision of
the N descriptors in the forthcoming 8th edition of the TNM
classification for lung cancer. J Thorac Oncol. 10:1675–1684. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Eberhardt WE, Mitchell A, Crowley J, Kondo
H, Kim YT, Turrisi A III, Goldstraw P, Rami-Porta R; International
Association for Study of Lung Cancer Staging, ; Prognostic Factors
Committee, Advisory Board Members and Participating Institutions:
The IASLC lung cancer staging project: Proposals for the revision
of the M descriptors in the forthcoming eighth edition of the TNM
classification of lung cancer. J Thorac Oncol. 10:1515–1522. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nagy Á, Munkácsy G and Győrffy B:
Pancancer survival analysis of cancer hallmark genes. Sci Rep.
11:60472021. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Győrffy B, Surowiak P, Budczies J and
Lánczky A: Online survival analysis software to assess the
prognostic value of biomarkers using transcriptomic data in
non-small-cell lung cancer. PLoS One. 8:e822412013. View Article : Google Scholar
|
|
30
|
Raponi M, Zhang Y, Yu J, Chen G, Lee G,
Taylor JM, Macdonald J, Thomas D, Moskaluk C, Wang Y and Beer DG:
Gene expression signatures for predicting prognosis of squamous
cell and adenocarcinomas of the lung. Cancer Res. 66:7466–7472.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhu CQ, Ding K, Strumpf D, Weir BA,
Meyerson M, Pennell N, Thomas RK, Naoki K, Ladd-Acosta C, Liu N, et
al: Prognostic and predictive gene signature for adjuvant
chemotherapy in resected non-small-cell lung cancer. J Clin Oncol.
28:4417–4424. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee ES, Son DS, Kim SH, Lee J, Jo J, Han
J, Kim H, Lee HJ, Choi HY, Jung Y, et al: Prediction of
recurrence-free survival in postoperative non-small cell lung
cancer patients by using an integrated model of clinical
information and gene expression. Clin Cancer Res. 14:7397–7404.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hou J, Aerts J, den Hamer B, van Ijcken W,
den Bakker M, Riegman P, van der Leest C, van der Spek P, Foekens
JA, Hoogsteden HC, et al: Gene expression-based classification of
non-small cell lung carcinomas and survival prediction. PLoS One.
5:e103122010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bild AH, Yao G, Chang JT, Wang Q, Potti A,
Chasse D, Joshi MB, Harpole D, Lancaster JM, Berchuck A, et al:
Oncogenic pathway signatures in human cancers as a guide to
targeted therapies. Nature. 439:353–357. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yamauchi M, Yamaguchi R, Nakata A, Kohno
T, Nagasaki M, Shimamura T, Imoto S, Saito A, Ueno K, Hatanaka Y,
et al: Epidermal growth factor receptor tyrosine kinase defines
critical prognostic genes of stage I lung adenocarcinoma. PLoS One.
7:e439232012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Xie Y, Xiao G, Coombes KR, Behrens C,
Solis LM, Raso G, Girard L, Erickson HS, Roth J, Heymach JV, et al:
Robust gene expression signature from formalin-fixed
paraffin-embedded samples predicts prognosis of non-small-cell lung
cancer patients. Clin Cancer Res. 17:5705–5714. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Botling J, Edlund K, Lohr M, Hellwig B,
Holmberg L, Lambe M, Berglund A, Ekman S, Bergqvist M, Pontén F, et
al: Biomarker discovery in non-small cell lung cancer: Integrating
gene expression profiling, meta-analysis, and tissue microarray
validation. Clin Cancer Res. 19:194–204. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cancer Genome Atlas Research Network, .
Comprehensive genomic characterization of squamous cell lung
cancers. Nature. 489:519–525. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yan G, Du Q, Wei X, Miozzi J, Kang C, Wang
J, Han X, Pan J, Xie H, Chen J and Zhang W: Application of
real-time cell electronic analysis system in modern pharmaceutical
evaluation and analysis. Molecules. 23:32802018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang X, Guo H, Bao Y, Yu H, Xie D and
Wang X: Exosomal long non-coding RNA DLX6-AS1 as a potential
diagnostic biomarker for non-small cell lung cancer. Oncol Lett.
18:5197–5204. 2019.PubMed/NCBI
|
|
41
|
Guo Z, Zhang X, Zhu H, Zhong N, Luo X,
Zhang Y, Tu F, Zhong J, Wang X, He J and Huang L: TELO2 induced
progression of colorectal cancer by binding with RICTOR through
mTORC2. Oncol Rep. 45:523–534. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Liu J, Li J, Wang K, Liu H, Sun J, Zhao X,
Yu Y, Qiao Y, Wu Y, Zhang X, et al: Aberrantly high activation of a
FoxM1-STMN1 axis contributes to progression and tumorigenesis in
FoxM1-driven cancers. Signal Transduct Target Ther. 6:422021.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhang X, Hu J, Zhong L, Wang N, Yang L,
Liu CC, Li H, Wang X, Zhou Y, Zhang Y, et al: Quercetin stabilizes
apolipoprotein E and reduces brain Aβ levels in amyloid model mice.
Neuropharmacology. 108:179–192. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Stack EC, Wang C, Roman KA and Hoyt CC:
Multiplexed immunohistochemistry, imaging, and quantitation: A
review, with an assessment of Tyramide signal amplification,
multispectral imaging and multiplex analysis. Methods. 70:46–58.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Liao H, Liang Y, Kang L, Xiao Y, Yu T and
Wan R: miR4543p inhibits nonsmall cell lung cancer cell
proliferation and metastasis by targeting TGFB2. Oncol Rep.
45:672021. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Kang J, Bai R, Liu K, Ji XP, Li Y and Han
JY: Identification of significantly different modules between
permanent and deciduous teeth by network and pathway analyses.
Genet Mol Res. 15:2016. View Article : Google Scholar
|
|
48
|
Perlikos F, Harrington KJ and Syrigos KN:
Key molecular mechanisms in lung cancer invasion and metastasis: A
comprehensive review. Crit Rev Oncol Hematol. 87:1–11. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mao M, Chen J, Li X and Wu Z: siRNA-TMEM98
inhibits the invasion and migration of lung cancer cells. Int J
Clin Exp Pathol. 8:15661–15669. 2015.PubMed/NCBI
|
|
50
|
Akkafa F, Koyuncu I, Temiz E, Dagli H,
Dilmec F and Akbas H: miRNA-mediated apoptosis activation through
TMEM 48 inhibition in A549 cell line. Biochem Biophys Res Commun.
503:323–329. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ombrato L and Malanchi I: The EMT
universe: Space between cancer cell dissemination and metastasis
initiation. Crit Rev Oncog. 19:349–361. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Son H and Moon A: Epithelial-mesenchymal
transition and cell invasion. Toxicol Res. 26:245–252. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Hida Y and Hamada J: Differential
expressions of matrix metalloproteinases, a disintegrin and
metalloproteinases, and a disintegrin and metalloproteinases with
thrombospondin motifs and their endogenous inhibitors among
histologic subtypes of lung cancers. Anticancer Agents Med Chem.
12:744–752. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Engelman JA, Chen L, Tan X, Crosby K,
Guimaraes AR, Upadhyay R, Maira M, McNamara K, Perera SA, Song Y,
et al: Effective use of PI3K and MEK inhibitors to treat mutant
Kras G12D and PIK3CA H1047R murine lung cancers. Nat Med.
14:1351–1356. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Goodwin JM, Svensson RU, Lou HJ, Winslow
MM, Turk BE and Shaw RJ: An AMPK-independent signaling pathway
downstream of the LKB1 tumor suppressor controls Snail1 and
metastatic potential. Mol Cell. 55:436–450. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Qiao M, Sheng S and Pardee AB: Metastasis
and AKT activation. Cell Cycle. 7:2991–2996. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Kim SO and Kim MR: [6]-Gingerol prevents
disassembly of cell junctions and activities of MMPs in invasive
human pancreas cancer cells through ERK/NF-κ B/Snail signal
transduction pathway. Evid Based Complement Alternat Med.
2013:7618522013. View Article : Google Scholar : PubMed/NCBI
|