Introduction
Prostate cancer (PCa) is the second most common
cancer type worldwide and was ranked fifth with regards to
cancer-related mortality rates in men in 2018 globally (1). Localized PCa can be treated using
radical prostatectomy or radiation therapy (2). However, the disease control of
metastatic PCa (mPCa) remains unsatisfactory (3). Although hormonal therapy has been widely
used for mPCa, recurrence nearly always occurs after the initial
period of treatment response and the cancer inevitably progresses
to metastasis castrate-resistant PCa, which is extremely difficult
to treat (4,5). Therefore, it is crucial to identify the
exact molecular mechanism of the progression of mPCa, which may
provide novel diagnostic and therapeutic targets.
Over the last decades, the technology of gene
microarray and bioinformatic analysis has been applied for the
examination of genetic alterations, which has enabled the
identification of differentially expressed genes (DEGs) between
mPCa and normal prostate tissues (6).
In the present study, different microarray datasets were downloaded
from the Gene Expression Omnibus (GEO) and multivariate statistical
techniques were used for analysis. Analysis of the protein-protein
interaction (PPI) network, enrichment analysis of Gene Ontology
(GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway
were performed to predict the hub genes and the molecular mechanism
of mPCa among the DEGs, which could guide future experiments in
vitro and in vivo.
CD44 is a cell-surface receptor that is expressed in
the majority of normal and cancer tissues (7). CD44 is a cell-surface marker that is
associated with the stemness, initiation and invasiveness of tumor
cells (8). It has been reported that
cell adhesion is primarily mediated by the CD44 signaling pathway,
which is initiated by cleavages of CD44 ectodomain. Cleavages of
CD44 ectodomain induced CD44 intracellular domain cleavage, and the
subsequently generated intracellular domain fragment, can regulate
signaling transcription (9). It has
been revealed that MMPs are involved in the cleavage of CD44
ectodomain (10). Previous findings
have observed that PC-3 cells expressed CD44, while LNCaP cells did
not (11). It was also reported that
CD44 could regulate cell proliferation, invasion and migration via
pyruvate dehydrogenase kinase 1 (PDK1) and
6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 4 (PFKB4) in
PCa cells (12). Additionally, MMP
inhibitor (SB-3CT) could decrease glycolytic activity via the
inhibition of CD44 in PCa cells, and combination therapy with
SB-3CT and docetaxel was more effective in inhibiting PCa compared
with monotherapy (12).
Based on the results of previous studies in
vitro, the present study performed additional experiments in
vivo to further determine the role of CD44 in the progression
of PCa and the role of SB-3CT in PCa.
Materials and methods
Information of microarray data
GEO (http://www.ncbi.nlm.nih.gov/geo) is a public
repository of high-throughput functional genomics data (13,14). In
total, three datasets (GSE3325, GSE6919 and GSE38241) were
downloaded from GEO to compare gene expression between metastasis
prostate cancer tissues and normal prostate tissues. The organism
of the selected datasets was homo sapiens, and the experiment type
was expression profiling array. The probes of each dataset were
annotated by the gene symbol, according to the information of the
platform. The dataset of GSE3325 contained six mPCa tissue samples
and six normal prostate tissue samples (15), while GSE6919 contained 25 mPCa tissue
samples and 17 normal prostate tissue samples (16,17), and
GSE38241 contained 18 mPCa tissue samples and 21 normal prostate
tissue samples (18).
Identification of differentially
expressed genes
The DEGs between mPCa and normal prostate samples
were screened using statistical software R (https://www.r-project.org; version 3.6.0). Data
standardization and quality detection were performed prior to
analysis. DEGs were identified using the Empirical Bayes method
according to the ‘limma’ package of Bioconductor (https://bioconductor.org) (19). In addition, |log2FC|>1 was set as
the cut-off value and adjusted P<0.05 was considered
significant. A volcano plot of the DEGs was conducted based on the
‘ggplot2’ package of R (https://CRAN.R-project.org/package=ggplot2; version
3.2.1). A heatmap of the DEGs was constructed using the ‘pheatmap’
package of R (https://CRAN.R-project.org/package=pheatmap; version
1.0.12).
Enrichment analysis of GO term and
KEGG pathway
GO term enrichment analysis of DEGs including
biological process (BP), cellular component (CC) and molecular
function (MF) was conducted using the ‘clusterProfiler’ package of
Bioconductor (20). The KEGG pathway
enrichment analysis of DEGs was performed using the DAVID database
(https://david.ncifcrf.gov; version 6.8),
which provides functional annotation tools online for understanding
biological processes. P<0.05 was considered to indicate a
statistically significant difference (21,22).
Analysis of PPI network and hub gene
identification
PPI network analysis was performed to identify hub
genes and to evaluate the interactions among DEGs using the online
database of Search Tool for the Retrieval of Interacting Genes
(STRING; https://string-db.org; version 11.0) and
Cytoscape software (www.cytoscape.org; version 3.7.1) (23,24).
Firstly, the network of DEGs was mapped using STRING database with
an interaction score >0.4. Then, the network was visualized
using Cytoscape software. The top 10 hub genes among the DEGs were
identified using the cytoHubba plugin of Cytoscape (25).
PCa cell line and cell culture
LNCap and PC-3 cells (obtained from the Cell Bank of
Type Culture Collection of Chinese Academy of Sciences) were
cultured in RPMI-1640 medium (Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Thermo Fisher Scientific, Inc.). The
cells were cultured in an incubator at 37°C and 5% CO2.
Negative control groups were designed and performed in the
experiments, and experiments were repeated at least three
times.
Cell transfection
The sequences of short hairpin (sh)RNA targeting
PDK1 or PFKFB4 and the negative control were inserted
into the lentiviral vector. A non-targeting sequence (forward
sequence
5′-CCGGCAACAAGATGAAGAGCACCAACTCGAGTTGGTGCTCTTCATCTTGTTGTTTTT-3′)
was used as the negative control. Lentiviruses with the packaging
plasmid (PG-P1-VSVG, PG-P2-REV, PG-P3-RRE and pGLV3/H1/GFP) and
shRNA plasmid were produced via the transfection of 293T cells.
Supernatants with lentiviral were collected after transfection and
filtered using a 0.45-µm strainer. The lentiviral-expressing CD44
was harvested by inserting the sequences of CD44 into a pLVX-EF1α
vector. An empty vector was used as the negative control. The
lentiviral vector was used to infect PC-3 cells for 24 h at 37°C.
Using the same method, LNCaP cells were transfected with CD44
overexpression lentiviral vector. PC-3 and LNCaP cells infected by
lentiviral vector were cultured for 72 h before subsequent
experiments. Plasmid was purchased from BioVector NTCC Inc.
Western blot analysis
Cell extraction was performed using lysis buffer
(Thermo Fisher Scientific, Inc.) and protein was separated via 10%
SDS-PAGE. The concentration of protein was quantified by the
bicinchoninic acid method. The protein extracted from the NC,
SB-3CT, SB-3CT + Docetaxel (5 mg/kg), and SB-3CT + Docetaxel (10
mg/kg) groups were loaded in different western blot lanes,
respectively. Then, the separated protein was transferred onto a
polyvinylidene fluoride membrane. After blocking with 5% skimmed
milk, the membrane was incubated with primary antibodies specific
for PDK1 (cat. no. ab110025; 1:500 dilution, Abcam), PFKFB4 (cat.
no. ab137785; 1:500 dilution, Abcam) or GAPDH (cat. no. ab181602;
1:1,000 dilution, Abcam), followed by incubation with secondary
antibody IgG (cat. no. R4880; 1:1,000 dilution, Sigma-Aldrich;
Merck KGaA). The protein was visualized using enhanced
chemiluminescence and analysed using software by Labworks Analysis
Software.
Immunohistochemical staining
Immunohistochemical staining was performed on
paraffin-embedded tumor tissue sections using anti-PDK1 (cat. no.
ab110025; 1:1,000 dilution, Abcam) or anti-PFKFB4 (cat. no.
ab137785; 1:1,000 dilution, Abcam) antibodies according to the
manufacturer's protocol. The tissue sample was fixed with 4%
paraformaldehyde at 4°C for 12 h. After staining, the sections (5
µm) were observed at ×100 and ×400 magnification using a light
microscope. Positive cells were distinguished by strong staining of
the membrane.
Tumor xenograft model in vivo
Animal experiments were permitted by the Ethics
Committee of the People's Hospital of Guangxi Zhuang Autonomous
Region (approval no. 2014-010). A total of 40 BALB/c nude mice
(age, 4–6 weeks; male; weight, 20–25 g) were obtained from
Guangdong Medical Laboratory Animal Center and maintained in a
specific pathogen-free environment which consisted of individually
ventilated cages and isolator modules. The mice were injected with
treated PCa cells in the armpit. PC-3 cells, PC-3 cells infected
with shRNA-PDK1 or shRNA-PFKFB4, and LNCaP cells infected with
vector-expressing CD44 or negative control were subcutaneously
injected into BALB/c nude mice. The BALB/c nude mice injected with
PC-3 cells were considered as the negative control. The tumor
volume was observed and measured up to 33 days. Tumor weight was
measured after mice were euthanized.
For the evaluation of CD44 in the treatment of PCa
in vivo, PC-3 cells were subcutaneously injected into BALB/c
nude mice and SB-3CT or SB-3CT combined with docetaxel (5 or 10
mg/kg) was injected into the mice via the tail vein. The tumor
volume and weight were measured, and the tumor tissues were
dissected for western blotting and immunohistochemical staining.
BALB/c nude mice injected with PC-3 cells were considered the
negative control.
During the experiment, 40 BALB/c nude mice were
used. Mice were kept on a 12 h light-dark cycle at 50-60% humidity
and 23–25°C and fed chow and water ad libitum. The health
and behavior of the mice were monitored twice daily, which included
weight, water and food intake, and animal posture. Sign of weight
loss, rapid breathing, bloating, reduced food intake, and visible
tumor under the skin was regarded as illness, which led to the
euthanasia of mice. All 40 mice were euthanized as tumors were
observed under the skin, using pentobarbital sodium (100 mg/kg) in
the study. The pentobarbital sodium was injected via the tail vein.
The maximum tumor size in the mice allowed to grow was 2,000
mm3 (not exceed 20 mm in any direction) before
euthanasia. In the research, ulceration of tumors was not observed
in any of the mice, and metastatic tumors to the lung were evident
in 8 of the 40 mice.
Statistical analysis
The statistical analysis was performed using SPSS
software (version 19.0; IBM Corp.) and the graphs were created
using GraphPad Prism software (version 6.0; GraphPad Software,
Inc.). Data were analysed using the Student's t-test (independent
t-test) or a one-way ANOVA with Tukey's post hoc test. P<0.05
was considered to indicate a statistically significant
difference.
Results
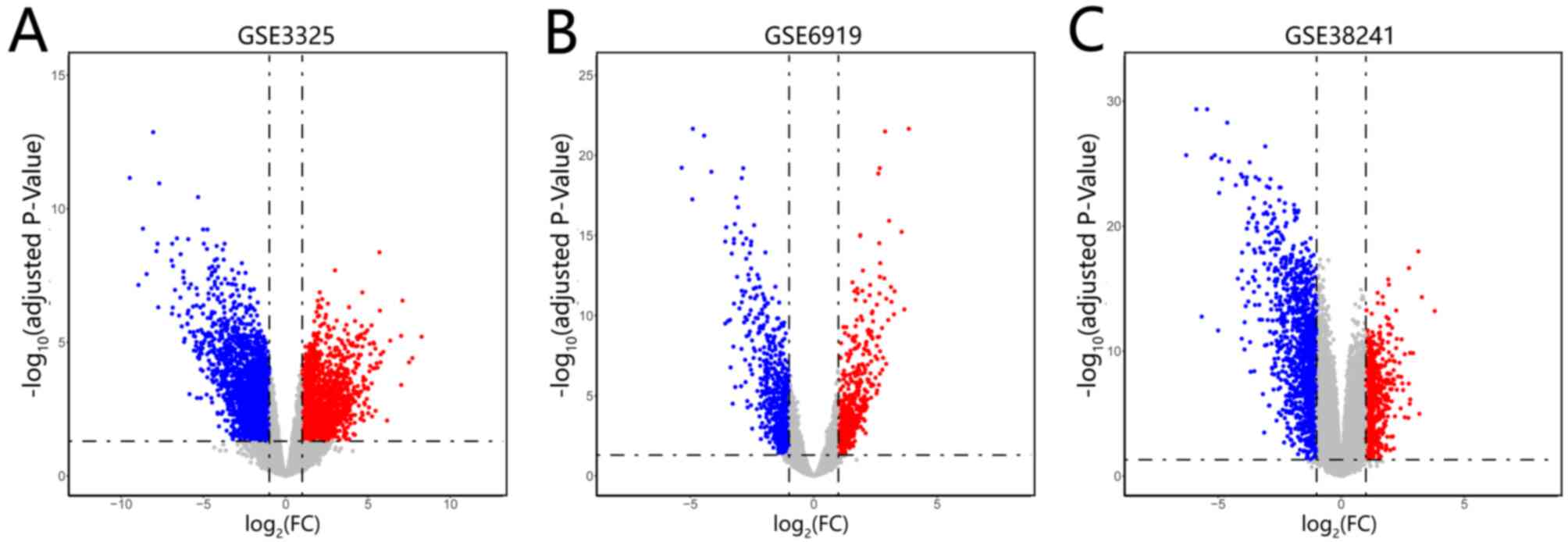
Identification of DEGs in mPCa
The datasets of GSE3325, GSE6919 and GSE38241 were
downloaded from the GEO platform. A total of 4,790, 1,144 and 1,920
DEGs were screened from GSE3325, GSE6919 and GSE38241, respectively
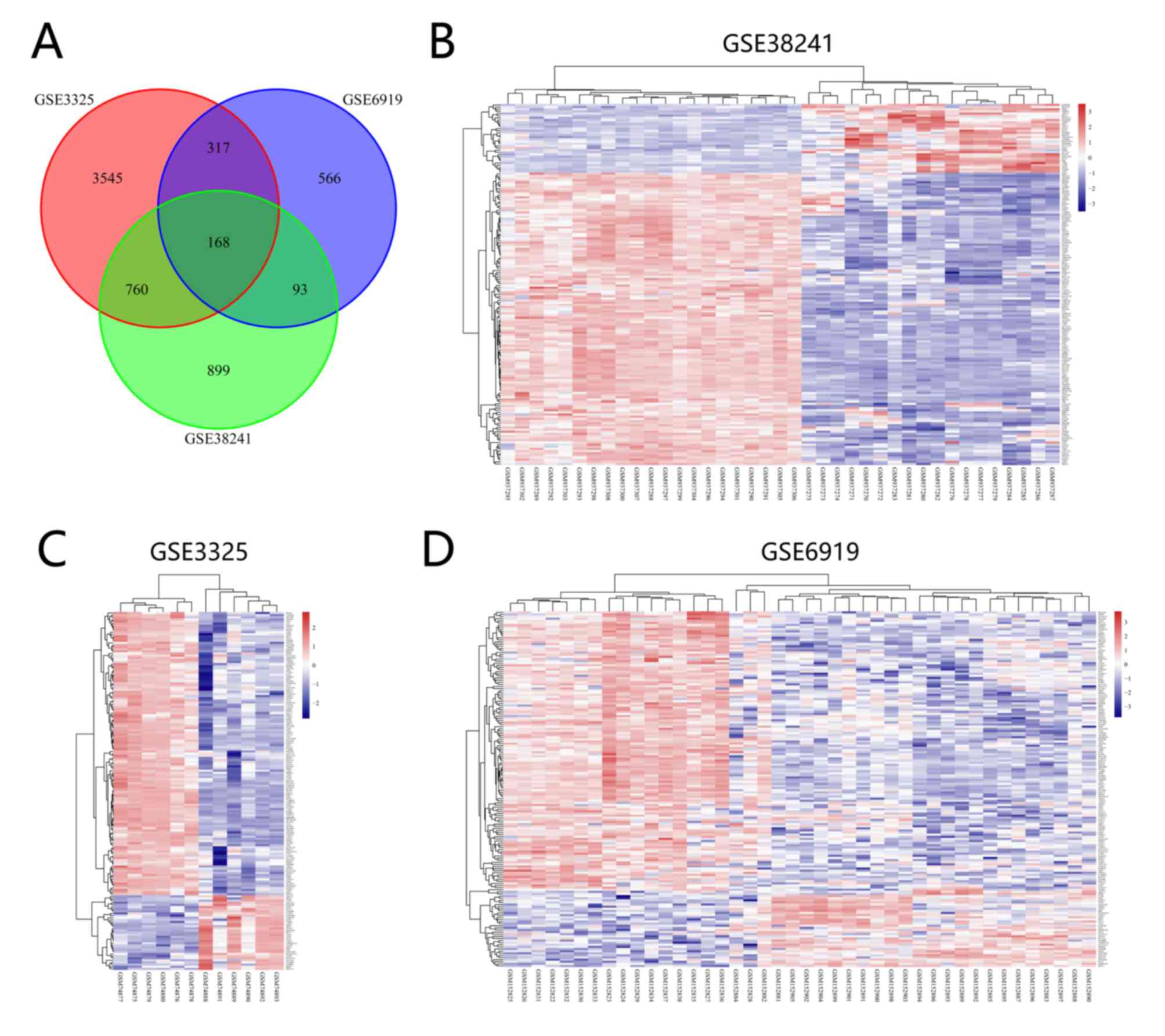
(Fig. 1A-C). In addition, 168 common
DEGs were identified among the three datasets (Fig. 2A) and the expression levels of the
common DEGs in the three datasets are presented (Fig. 2B-D).
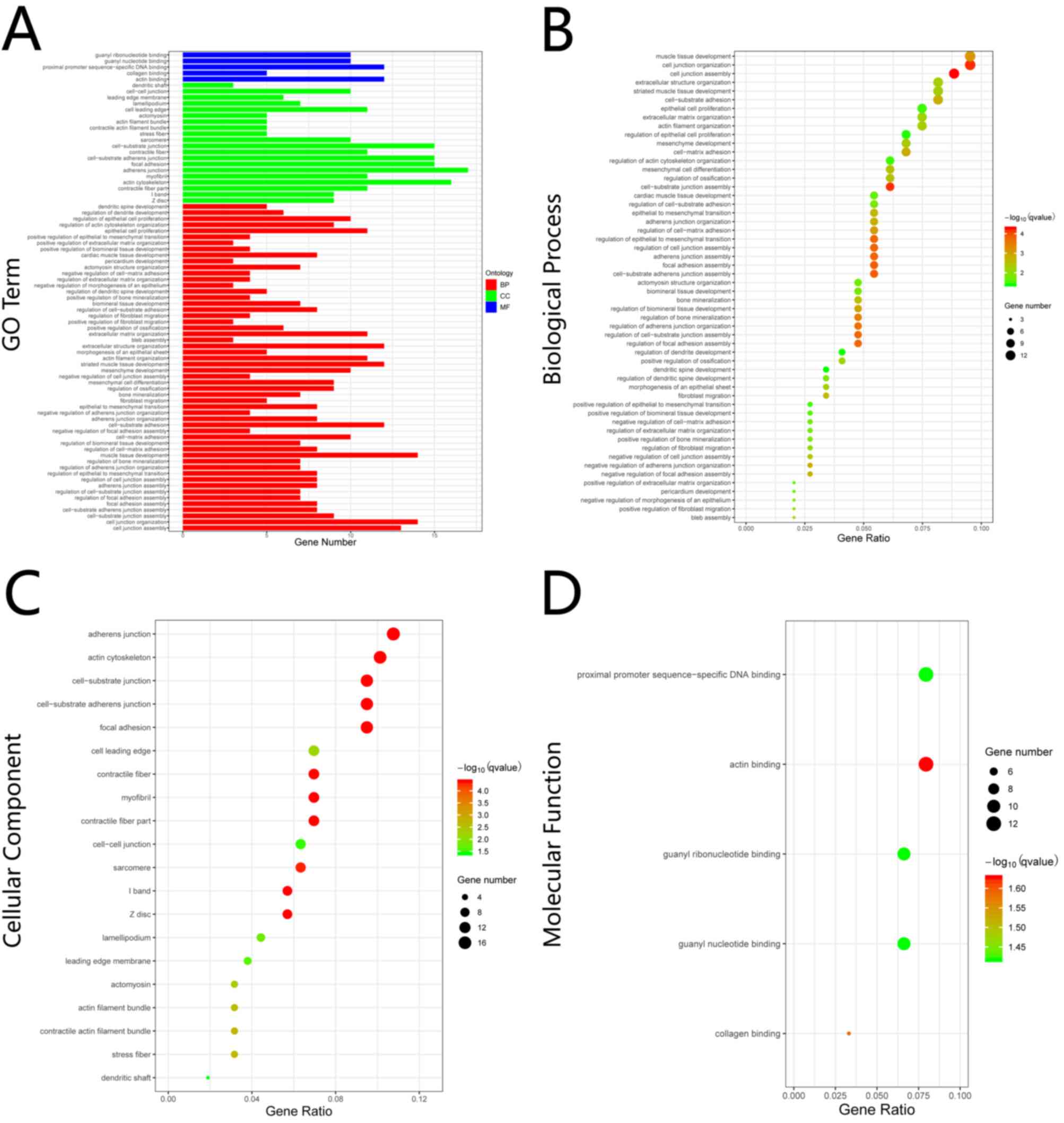
Enrichment analysis of GO term
The results of GO enrichment analysis varied with
regards to the GO term and the different expression of common DEGs
(Fig. 3A). The result of GO
enrichment analysis in BP showed that the common DEGs were
significantly enriched in ‘cell junction’, ‘cell adhesion’,
‘epithelial to mesenchymal transition’ and ‘epithelial cell
proliferation’, among others (Fig.
3B). With regards to CC, the common DEGs were significantly
enriched in ‘adherens junction’, ‘cell junction’, ‘focal adhesion’
and ‘myofibril’, among others (Fig.
3C). For MF, the common DEGs were significantly enriched in
‘actin binding’, ‘collagen binding’, ‘proximal promoter
sequence-specific DNA binding’, ‘guanyl nucleotide binding’ and
‘guanyl ribonucleotide binding’ (Fig.
3D).
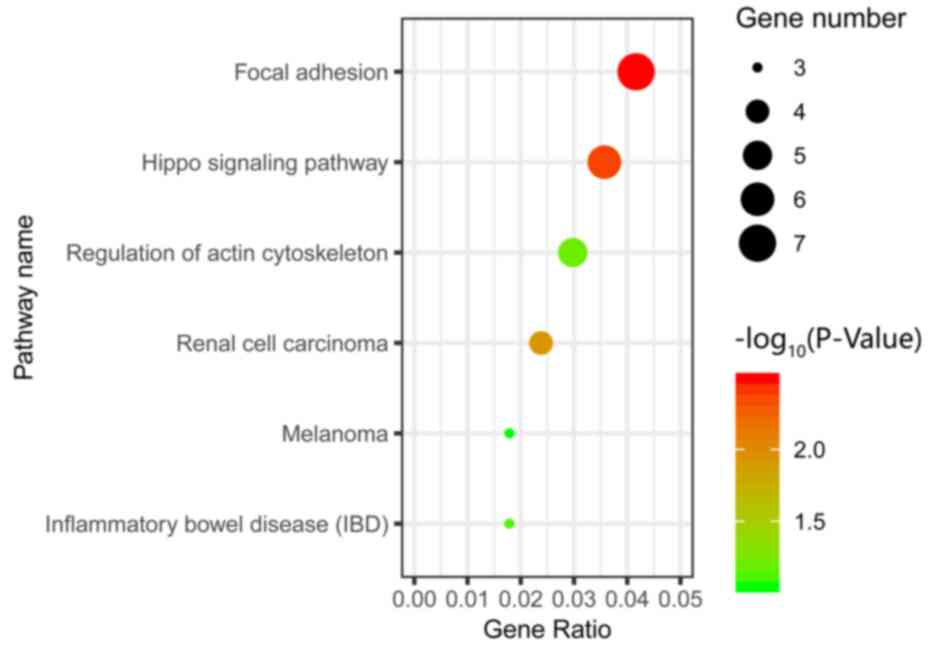
Enrichment analysis of KEGG
pathway
The results of KEGG pathway enrichment analysis
demonstrated that the common DEGs were significantly enriched in
the ‘Focal adhesion’, ‘Hippo’ and ‘Renal cell carcinoma’ signaling
pathways (Fig. 4).
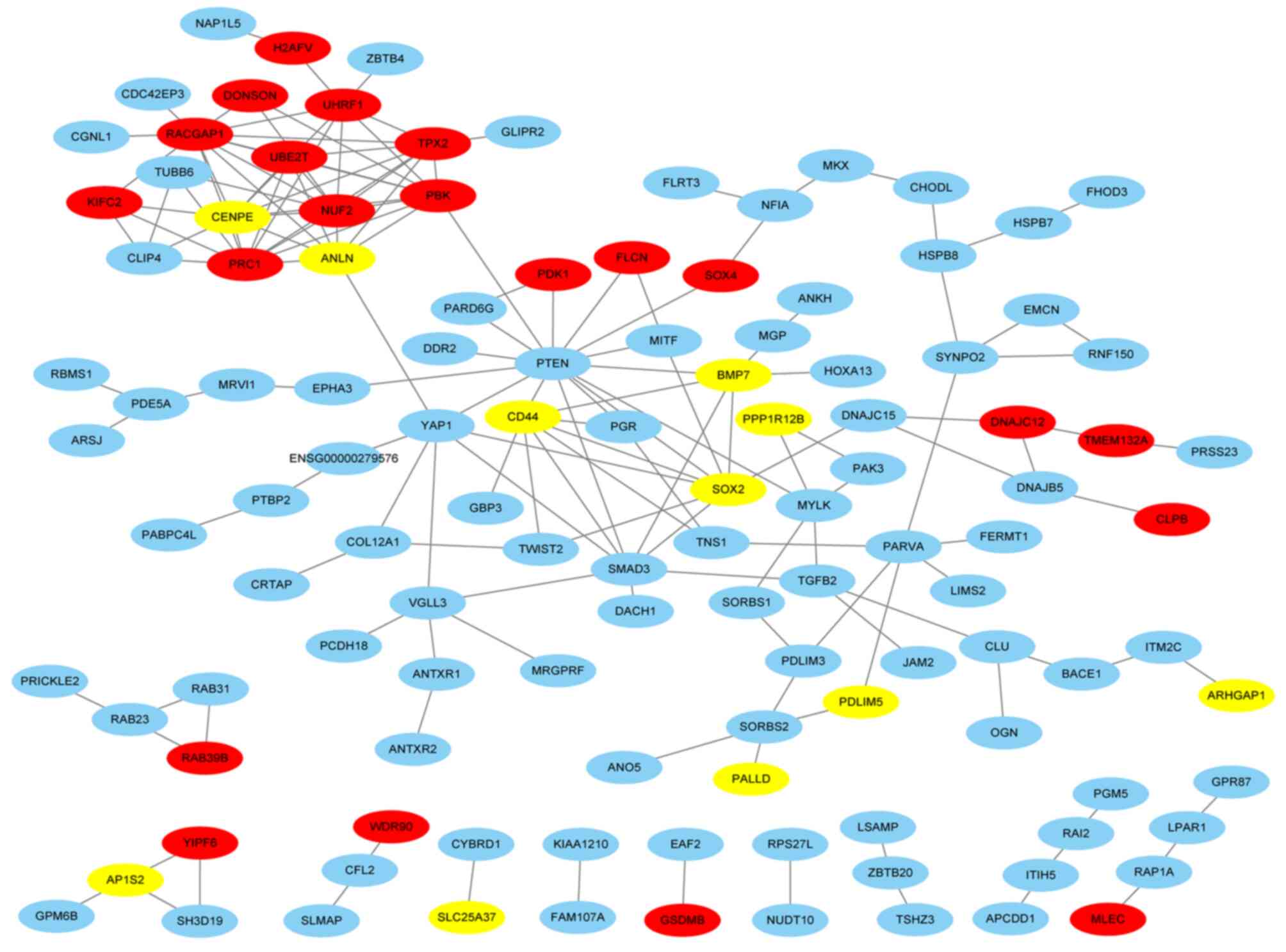
Analysis of PPI network and hub
gene
The PPI network consisted of 167 nodes and 168 edges
based on STRING database, and the network was visualized using
Cytoscape software (Fig. 5). The top
10 common DEGs with the highest degree were screened as the hub
genes of mPCa and their names and functions are presented in
Table I.
 | Table I.Functional roles of top 10 hub genes
with a degree ≥8. |
Table I.
Functional roles of top 10 hub genes
with a degree ≥8.
| No. | Gene symbol | Full name | Function
(Ref.) |
|---|
| 1 | PTEN | Phosphatase and
tensin homolog | PTEN acts as a
tumor suppressor through regulating AKT/PKB signaling pathway
negatively (41) |
| 2 | RACGAP1 | Rac
GTPase-activating protein 1 | RACGAP1 can
regulate the progression of cytokinesis, cell growth and
differentiation (42) |
| 3 | PRC1 | Protein regulator
of cytokinesis 1 | PRC1 is associated
with cytokinesis (43) |
| 4 | PBK | PDZ binding
kinase | High expression of
PBK is associated with tumorigenesis (44) |
| 5 | CENPE |
Centromere-associated protein E | CENPE is necessary
for stable spindle microtubule capture (45) |
| 6 | NUF2 | NUF2 component of
NDC80 kinetochore complex | NUF2 is associated
with centromeres of mitotic (46) |
| 7 | TPX2 | TPX2 microtubule
nucleation factor | TPX2 is implicated
as a regulator of cell apoptosis (47) |
| 8 | SOX2 | SRY-box
transcription factor 2 | Transcription
factors encoded by SOX2 play a regulatory role in embryonic
development (48) |
| 9 | CD44 | CD44 molecule | CD44 is associated
with cell-cell interaction, cell migration and adhesion (10) |
| 10 | UHRF1 | Ubiquitin-like with
PHD and ring finger domains 1 | UHRF1 is
overexpressed in various types of cancer (49) |
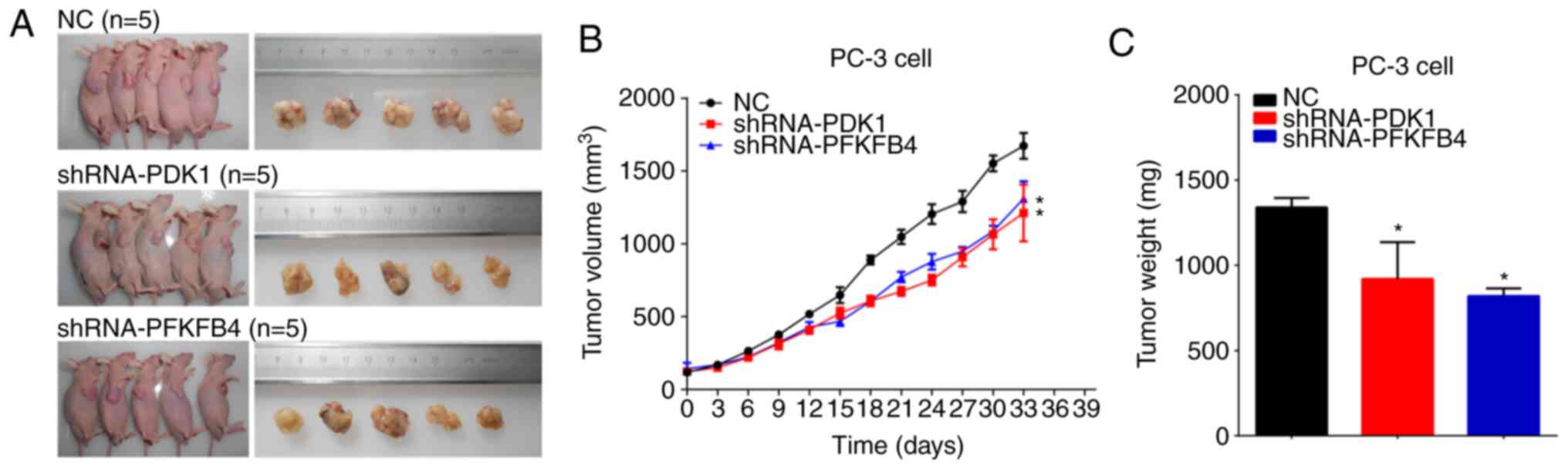
Knockdown of PDK1 or PFKFB4 inhibits
tumorigenicity of PCa cells in vivo
The BALB/c nude mice were injected subcutaneously
with PC-3 cells infected with shRNA-PDK1 or shRNA-PFKFB4. Tumors
dissected from mice were imaged and measured (Fig. 6A). The maximum tumor size was 1,775.74
mm3. It was found that knockdown of PDK1 or PFKFB4
inhibited the tumor growth of PCa cells in vivo (Fig. 6B). Similar results were observed for
tumor weight (Fig. 6C).
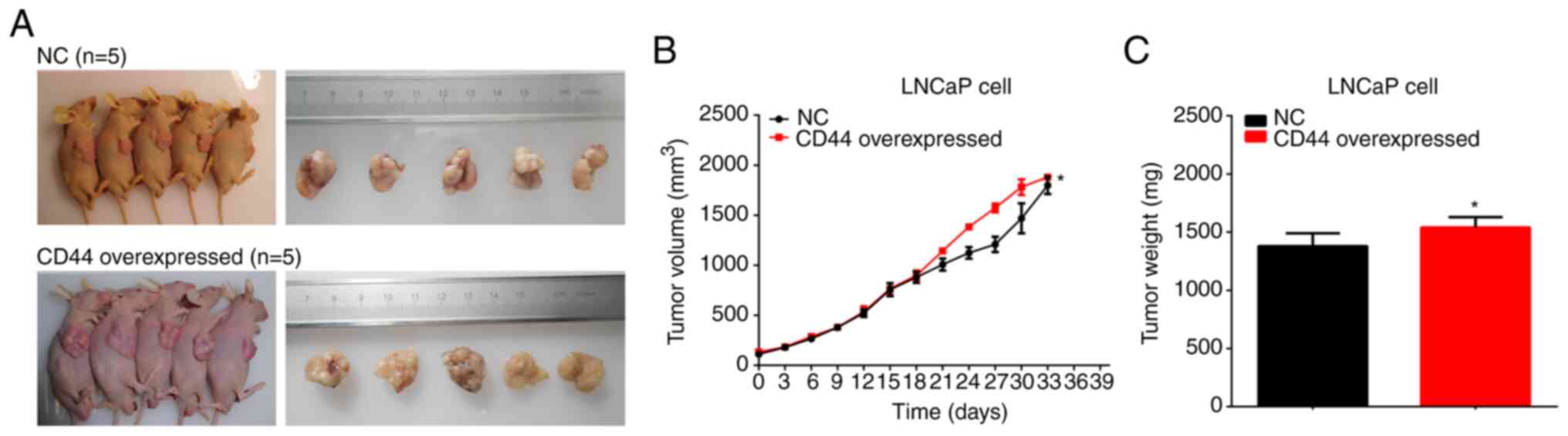
Overexpression of CD44 promotes
tumorigenicity of PCa cells in vivo
The BALB/c nude mice were subcutaneously injected
with LNCaP cells transfected with CD44 overexpression vector or NC.
Tumors dissected from mice were imaged and measured (Fig. 7A). The maximum tumor size was 1,932.95
mm3. The results indicated that overexpression of CD44
promoted the tumor growth and tumor weight of PCa xenografts in
vivo (Fig. 7B and C).
Inhibition of CD44 suppresses
tumorigenicity of PCa cells in vivo and the CD44 inhibitor (SB-3CT)
combined with docetaxel inhibits the tumorigenicity of PCa
The BALB/c nude mice were subcutaneously injected
with PC-3 cells. After inoculation, SB-3CT or SB-3CT combined with
docetaxel (5 or 10 mg/kg) was injected into mice via the tail vein.
Tumors dissected from mice were imaged and measured (Fig. 8A). The maximum tumor size was 1,775.74
mm3. It was identified that combined therapy with CD44
inhibitor (SB-3CT) and docetaxel could significantly inhibit tumor
growth compared with treatment with the CD44 inhibitor (SB-3CT)
alone. Moreover, it was found that a high concentration of
docetaxel (10 mg/kg) could achieve higher inhibitory effects
compared with the low concentration (5 mg/kg) (Fig. 8B and C). The expression levels of PDK1
and PFKFB4 in tumor tissue were examined, and were found to be
significantly downregulated both in the monotherapy and combined
therapy groups. Similar results were also observed in the results
of immunohistochemical staining (Fig.
8D-G). Based on these aforementioned results, it was suggested
that SB-3CT combined with a high dose of docetaxel could inhibit
tumor growth more effectively than SB-3CT alone.
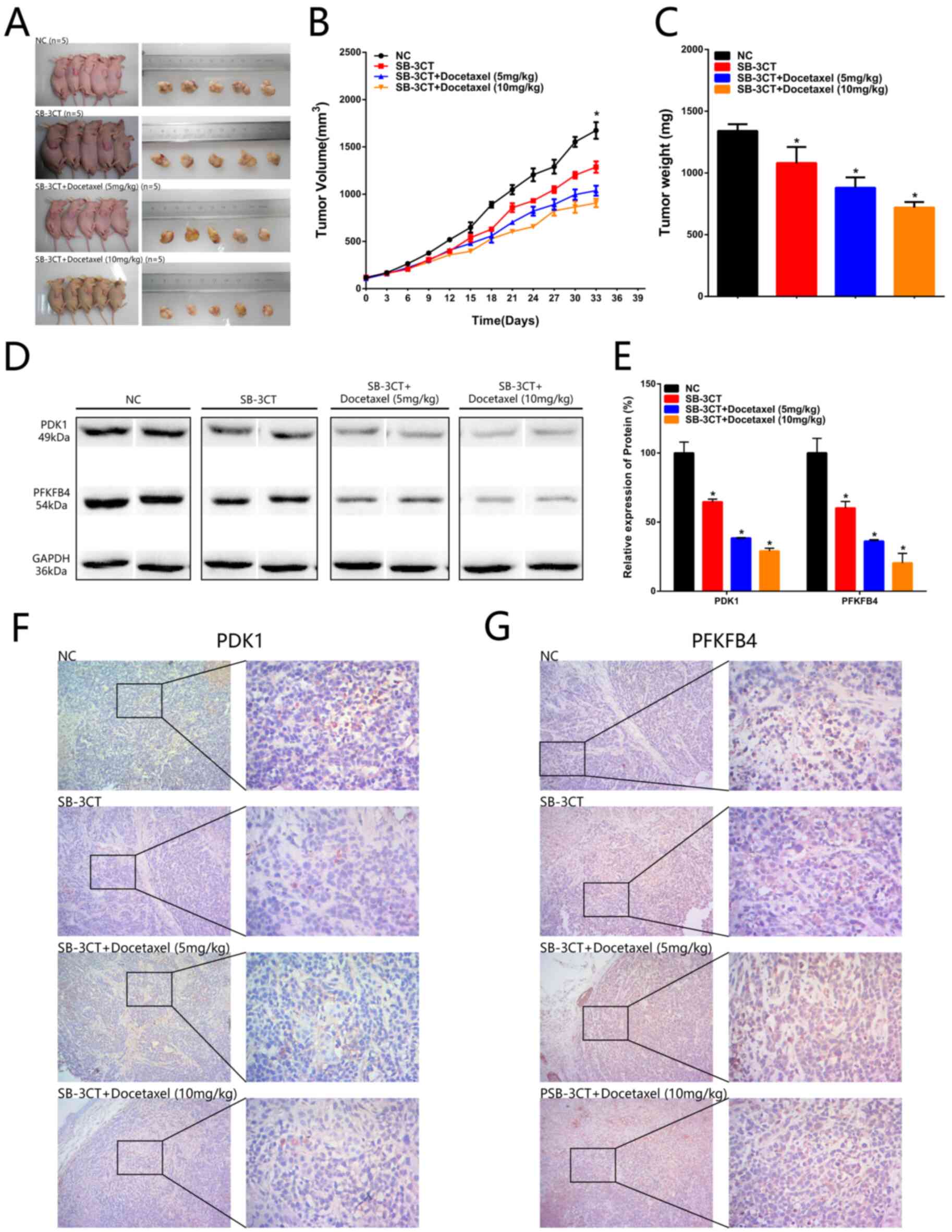 | Figure 8.Inhibition of CD44 suppresses
tumorigenicity of prostate cancer cells in vivo and the CD44
inhibitor (SB-3CT) combined with docetaxel inhibits tumorigenicity
of prostate cancer. (A) Tumors dissected from BALB/c nude mice are
presented (n=5 for each group). (B) Tumor volume curve of NC,
SB-3CT, SB-3CT + Docetaxel (5 mg/kg) and SB-3CT + Docetaxel (10
mg/kg) treatment groups. (C) Tumor weight of NC, SB-3CT, SB-3CT +
Docetaxel (5 mg/kg) and SB-3CT + Docetaxel (10 mg/kg) treatment
groups. (D and E) PDK1 and PFKFB4 expression levels were
downregulated in the PC-3 + SB-3T group, PC-3 + SB-3T + Docetaxel
(5 mg/kg) group and PC-3 + SB-3T + Docetaxel (10 mg/kg) group
compared with the PC-3 + NC group. (F) PDK1 expression levels were
downregulated in the PC-3 + SB-3T group, PC-3 + SB-3T + Docetaxel
(5 mg/kg) group and PC-3 + SB-3T + Docetaxel (10 mg/kg) group
compared with the PC-3 + NC group. (G) PFKFB4 expression levels
were downregulated in the PC-3 + SB-3T group, PC-3 + SB-3T +
Docetaxel (5 mg/kg) group and PC-3 + SB-3T + Docetaxel (10 mg/kg)
group compared with the PC-3 + NC group. Data are analysed using an
ANOVA with Tukey's post hoc test. Data are presented as the mean ±
SD. *P<0.05. NC, negative control; PDK1, pyruvate dehydrogenase
kinase 1; PFKB4,
6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 4. |
Discussion
Although significant progress has been achieved in
the management of mPCa, the pathogenesis of mPCa has not been fully
elucidated due to the potentially complex biological traits of
cancer. Microarray technology enables researchers to screen hub
genes and primary pathways that are associated with mPCa, and has
proven to be a helpful technology (26). Therefore, microarray technology has
been used to identify genetic alterations involved in the
pathogenesis and progression of diseases.
In the present study, three microarray databases
were accessed to identify DEGs between mPCa tissues and normal
prostate tissues. A total of 168 common DEGs were obtained for
further analysis. To reveal interactions among the common DEGs, GO
and KEGG pathways, enrichment analysis was performed. GO enrichment
analysis indicated that the DEGs were mostly enriched in ‘cell
junction’ and ‘cell adhesion’. The results of GO analysis are
consistent with previous studies, which reported that ‘cell
junction’ is associated with paracellular diffusion regulation and
that ‘cell adhesion’ serves a crucial role in the transformation
and progression of cancer (27–29). The
KEGG pathway enrichment analysis indicated that the DEGs were
mostly enriched in ‘Hippo signaling pathway’, ‘focal adhesion’ and
‘renal cell carcinoma’. Previous studies have reported that the
hippo signaling pathway is involved in the regulation of cell
proliferation and cell apoptosis, and is upregulated in tumors
(30,31). In the present study, the top 10 common
DEGs with highest degree were screened as the hub genes, including
PTEN, Rac GTPase-activating protein 1, Protein regulator of
cytokinesis 1, PDZ binding kinase, Centromere-associated protein E,
NUF2 component of NDC80 kinetochore complex, TPX2 microtubule
nucleation factor, SOX2, CD44 and ubiquitin-like with PHD and ring
finger domains 1. Previous findings have revealed that CD44 is an
adhesion molecule and is involved in the processes of invasion and
metastasis in tumor cells (10). The
present bioinformatics analysis results were consistent with these
aforementioned findings. Based on the current results of the
bioinformatics analysis, it was suggested that CD44 could regulate
cell proliferation, migration and invasion in PCa.
The results of our previous study revealed that
inhibition of CD44 using SB-3CT could suppress proliferation,
invasion and migration in PCa cells by regulating PDK1 and PFKFB4
expression levels (12). Based on the
present bioinformatics analysis results and our previous study,
additional experiments in vivo were performed, including
tumor formation assay and tumor metastasis experiments. In the
present study, the results of tumor xenograft implantation
demonstrated that knockdown of PDK1 or PFKFB4 in PC-3 cells
inhibited the tumorigenicity of PCa in vivo. Further
experiments indicated that inhibition of CD44 using an MMP
inhibitor (SB-3CT) in PC-3 cells suppressed the tumorigenicity of
PCa and inhibited the expression levels of PDK1 and PFKFB4 in PC-3
cells. SB-3CT, a selective MMP inhibitor, has been reported to
block CD44 cleavage and inhibit downstream signaling pathway
(32). Taken together, the present
results indicated that CD44 suppressed the tumorigenicity of PCa
via PDK1 and PFKFB4 in vivo, which was consistent with the
results of previous study in vitro (12). As aforementioned, previous studies
have reported that PC-3 cells expressed CD44, while LNCaP cells did
not (11). In the present study, CD44
was overexpressed in LNCaP cells and this overexpression promoted
the tumorigenicity of PCa. Tumor metastasis experiments were also
performed. In the present study, metastatic tumors were found in
the lung of the mice. However, the difference between two groups in
the number of metastatic tumors in the lung was not
significant.
Currently, hormonal therapy and chemotherapy are the
first line choice for the treatment of mPCa, and adverse events
were frequent in the two protocols (33). The combination of hormonal therapy and
docetaxel became a novel therapy for mPCa. The results of the
present study demonstrated that CD44 regulated the tumorigenicity
of PCa in vivo, which suggested that inhibition of CD44
using SB-3CT is a novel potential treatment of PCa. According to
current combined therapy of mPCa, it was suggested that the
combination of CD44 inhibitor and docetaxel may be a beneficial
strategy. Our previous study revealed that the combination of
docetaxel and SB-3CT could significantly decrease the viability of
PC-3 cells compared with single treatment of docetaxel at the
concentration of 5 or 10 mg/kg (12).
The present study evaluated the effect of combined therapy with
CD44 inhibitor (SB-3CT) and docetaxel. The results indicated that
treatment with CD44 inhibitor and docetaxel inhibited tumor growth
and decreased expression levels of PDK1 and PFKFB4. Moreover, it
was identified that treatment with high concentration of docetaxel
induced a more positive response compared with the low
concentration. Although the combined therapy was effective,
sequential therapy is another potential therapy, but requires
further investigation (34).
CD44 is a cell-surface receptor for hyaluronic acid
and extracellular matrix components, and it serves a critical role
in connecting the microenvironments in cancer. CD44 enables cancer
cells to perceive the changes of microenvironments and can mediate
the transduction of growth factor and cytokine signaling which can
promote cell invasion and metastasis. Growth factors from
microenvironments mediated by CD44, including EDF, FGF, HGF, VEGF,
TGF-β, can also regulate tumorigenicity (35). It has been reported that CD44 could
regulate the activation of macrophages in tumors, which was
associated with tumorigenicity (36,37). As
aforementioned, the regulation of microenvironments or macrophages
by CD44 could affect tumorigenicity, but this required further
examination. CD44 can also regulate EMT and reactive oxygen species
(ROS) metabolism. Moreover, CD44 may regulate glucose metabolism in
PCa (38). With increased glycolytic
activity, reduced mitochondrial respiration leads to decreased ROS
levels (39). Previous in
vitro studies have reported that inhibition of CD44 expression
could decrease glucose consumption and increase ROS level. Based on
these results, it was suggested that CD44 could regulate the
tumorigenicity of PCa cells via the regulation of ROS via PDK1 or
PFKFB4.
The 70-kDa ribosomal protein S6 kinase, known as
p70S6K, is a dual pathway kinase which acts downstream of PI3K
pathway and mTOR pathway in response to growth factors and
cytokines to regulate cell growth and inhibit cell apoptosis.
Previous findings showed that p70S6K was regulated by PDK1 in PI3K
pathway and could suppress BAD-induced cell apoptosis by the
phosphorylation of Ser-136 on BAD (40). In our study, it was suggested that
CD44 could suppress the tumorigenicity of prostate cancer by
decreasing PDK1 and PFKFB4. Consequently, we hypothesized that CD44
could regulate the apoptosis of prostate cancer cells through
p70S6K. In future, the effect of CD44 on the apoptosis of prostate
cancer and its mechanism may be the focus of future research.
However, the present study has some limitations.
CD44 has two variable regions and various CD44 isoforms produced by
alternative splicing, which may have diverse effects on cancer
progression. Thus, the CD44 isoforms should be analyzed and
examined further in PCa tissues. Our previous findings demonstrated
the difference between combined treatment and docetaxel alone in
vitro. In addition, docetaxel was the first-line treatment for
castration-resistant prostate cancer, which had been confirmed by
lots of experiments in vivo and clinical practices. The main
purpose of the present study was confirming the benefits of
combined treatment with docetaxel and SB-3CT, a novel compound for
prostate cancer treatment. Based on the aforementioned findings,
SB-3CT alone was designed as the control group; however, it would
be more reasonable to add docetaxel as monotherapy as a further
control group.
In conclusion, a total of 168 common DEGs were
identified and 10 hub genes were considered as biomarkers for mPCa.
Further experimental results indicated that CD44 regulated the
tumorigenicity of PCa via PDK1 and PFKFB4 in vivo. The
present results demonstrated that the combination of SB-3CT and
docetaxel was more effective in the inhibition of tumor growth,
which suggested that combination therapy is a potential therapeutic
strategy for mPCa.
Acknowledgements
Not applicable.
Funding
This work was funded by the National Natural Science
Foundation of China (grant no. 81460387).
Availability of data and materials
The data are available on reasonable request from
the corresponding author.
Authors' contributions
WL contributed to the experiment conception and
design, data analysis, and manuscript draft. JL, ZC, and XL
conducted the experiments. WL, JL, DN, and GH contributed to
manuscript draft and data analysis. NH, ZL, and JL contributed to
interpretation of data, manuscript draft and manuscript revision.
NH, ZL, and JL are responsible for confirming the authenticity of
all the raw data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The Ethics Committee of the People's Hospital of
Guangxi Zhuang Autonomous Region approved the study (approval no.
2014-010). Informed consent was provided by all the
participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cocci A, Cito G, Romano A, Larganà G,
Vignolini G, Minervini A, Di Maida F, Campi R, Carini M, Mondaini N
and Russo GI: Radical prostatectomy and simultaneous penile
prosthesis implantation: A narrative review. Int J Impot Res.
32:274–280. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weiner AB, Nettey OS and Morgans AK:
Management of metastatic hormone-sensitive prostate cancer (mHSPC):
An evolving treatment paradigm. Curr Treat Options Oncol. 20:62019.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Heidenreich A, Bastian PJ, Bellmunt J,
Bolla M, Joniau S, van der Kwast T, Mason M, Matveev V, et al: EAU
guidelines on prostate cancer. Part II: Treatment of advanced,
relapsing, and castration-resistant prostate cancer. Eur Urol.
65:467–479. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wülfing C, Bögemann M, Goebell PJ,
Hammerer P, Machtens S, Pfister D, Schwentner C, Steuber T, von
Amsberg G and Schostak M: Treatment situation in metastastic
Castration Naive Prostate Cancer (mCRPC) and the implications on
clinical routine. Urologe A. 58:1066–1072. 2019.(In German).
View Article : Google Scholar
|
|
6
|
Dailey AL: Metabolomic bioinformatic
analysis. Methods Mol Biol. 1606:341–352. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Spring FA, Dalchau R, Daniels GL,
Mallinson G, Judson PA, Parsons SF, Fabre JW and Anstee DJ: The Ina
and Inb blood group antigens are located on a glycoprotein of
80,000 MW (the CDw44 glycoprotein) whose expression is influenced
by the In(Lu) gene. Immunology. 64:37–43. 1988.PubMed/NCBI
|
|
8
|
Prochazka L, Tesarik R and Turanek J:
Regulation of alternative splicing of CD44 in cancer. Cell Signal.
26:2234–2239. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Miletti-González KE, Murphy K, Kumaran MN,
Ravindranath AK, Wernyj RP, Kaur S, Miles GD, Lim E, Chan R,
Chekmareva M, et al: Identification of function for CD44
intracytoplasmic domain (CD44-ICD). J Biol Chem. 287:18995–19007.
2012. View Article : Google Scholar
|
|
10
|
Nagano O and Saya H: Mechanism and
biological significance of CD44 cleavage. Cancer Sci. 95:930–935.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tai S, Sun Y, Squires JM, Zhang H, Oh WK,
Liang CZ and Huang J: PC3 is a cell line characteristic of
prostatic small cell carcinoma. Prostate. 71:1668–1679. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Li W, Qian L, Lin J, Huang G, Hao N, Wei
X, Wang W and Liang J: CD44 regulates prostate cancer
proliferation, invasion and migration via PDK1 and PFKFB4.
Oncotarget. 8:65143–65151. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res. 41((Database Issue)):
D991–D995. 2013.PubMed/NCBI
|
|
14
|
Edgar R, Domrachev M and Lash AE: Gene
Expression Omnibus: NCBI gene expression and hybridization array
data repository. Nucleic Acids Res. 30:207–210. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Varambally S, Yu J, Laxman B, Rhodes DR,
Mehra R, Tomlins SA, Shah RB, Chandran U, Monzon FA, Becich MJ, et
al: Integrative genomic and proteomic analysis of prostate cancer
reveals signatures of metastatic progression. Cancer Cell.
8:393–406. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chandran UR, Ma C, Dhir R, Bisceglia M,
Lyons-Weiler M, Liang W, Michalopoulos G, Becich M and Monzon FA:
Gene expression profiles of prostate cancer reveal involvement of
multiple molecular pathways in the metastatic process. BMC Cancer.
7:642007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu YP, Landsittel D, Jing L, Nelson J, Ren
B, Liu L, McDonald C, Thomas R, Dhir R and Finkelstein S: Gene
expression alterations in prostate cancer predicting tumor
aggression and preceding development of malignancy. J Clin Oncol.
22:2790–2799. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Aryee MJ, Liu W, Engelmann JC, Nuhn P,
Gurel M, Haffner MC, Esopi D, Irizarry RA, Getzenberg RH, Nelson
WG, et al: DNA methylation alterations exhibit intraindividual
stability and interindividual heterogeneity in prostate cancer
metastases. Sci Transl Med. 5:169ra102013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Smyth GK: Linear models and empirical
bayes methods for assessing differential expression in microarray
experiments. Stat Appl Genet Mol Biol. 3:Article32004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yu G, Wang LG, Han Y and He QY:
clusterProfiler: An R package for comparing biological themes among
gene clusters. OMICS. 16:284–287. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar
|
|
22
|
Huang da W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: Paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009. View Article : Google Scholar
|
|
23
|
Shannon P, Markiel A, Ozier O, Baliga NS,
Wang JT, Ramage D, Amin N, Schwikowski B and Ideker T: Cytoscape: A
software environment for integrated models of biomolecular
interaction networks. Genome Res. 13:2498–2504. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Szklarczyk D, Gable AL, Lyon D, Junge A,
Wyder S, Huerta-Cepas J, Simonovic M, Doncheva NT, Morris JH, Bork
P, et al: STRING v11: Protein-protein association networks with
increased coverage, supporting functional discovery in genome-wide
experimental datasets. Nucleic Acids Res. 47:D607–D613. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chin CH, Chen SH, Wu HH, Ho CW, Ko MT and
Lin CY: cytoHubba: Identifying hub objects and sub-networks from
complex interactome. BMC Syst Biol. 8 (Suppl 4):S112014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Albertson DG and Pinkel D: Genomic
microarrays in human genetic disease and cancer. Hum Mol Genet 12
Spec No. 2:R145–R152. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hejmej A and Bilinska B: A role of
junction-mediated interactions in cells of the male reproductive
tract: Impact of prenatal, neonatal, and prepubertal exposure to
anti-androgens on adult reproduction. Histol Histopathol.
29:815–830. 2014.PubMed/NCBI
|
|
28
|
Hejmej A and Bilinska B: The effects of
flutamide on cell-cell junctions in the testis, epididymis, and
prostate. Reprod Toxicol. 81:1–16. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yu CC, Chen LC, Lin VC, Huang CY, Cheng
WC, Hsieh AR, Chang TY, Lu TL, Lee CH, Huang SP and Bao BY: Effect
of genetic variants in cell adhesion pathways on the biochemical
recurrence in prostate cancer patients with radical prostatectomy.
Cancer Med. 8:2777–2783. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Pan D: The hippo signaling pathway in
development and cancer. Dev Cell. 19:491–505. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Saucedo LJ and Edgar BA: Filling out the
Hippo pathway. Nat Rev Mol Cell Biol. 8:613–621. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Thorne RF, Legg JW and Isacke CM: The role
of the CD44 transmembrane and cytoplasmic domains in co-ordinating
adhesive and signalling events. J Cell Sci. 117:373–380. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tannock IF, de Wit R, Berry WR, Horti J,
Pluzanska A, Chi KN, Oudard S, Théodore C, James ND, Turesson I, et
al: Docetaxel plus prednisone or mitoxantrone plus prednisone for
advanced prostate cancer. N Engl J Med. 351:1502–1512. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Takeuchi H, Mmeje CO, Jinesh GG, Taoka R
and Kamat AM: Sequential gemcitabine and tamoxifen treatment
enhances apoptosis and blocks transformation in bladder cancer
cells. Oncol Rep. 34:2738–2744. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yan Y, Zuo X and Wei D: Concise review:
Emerging role of CD44 in cancer stem cells: A promising biomarker
and therapeutic target. Stem Cells Transl Med. 4:1033–1043. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rao G, Wang H, Li B, Huang L, Xue D, Wang
X, Jin H, Wang J, Zhu Y, Lu Y, et al: Reciprocal interactions
between tumor-associated macrophages and CD44-positive cancer cells
via Osteopontin/CD44 promote tumorigenicity in colorectal cancer.
Clin Cancer Res. 19:785–797. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Takeuchi H, Tanaka M, Tanaka A, Tsunemi A
and Yamamoto H: Predominance of M2-polarized macrophages in bladder
cancer affects angiogenesis, tumor grade and invasiveness. Oncol
Lett. 11:3403–3408. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Li W, Cohen A, Sun Y, Squires J, Braas D,
Graeber TG, Du L, Li G, Li Z, Xu X, et al: The role of CD44 in
glucose metabolism in prostatic small cell neuroendocrine
carcinoma. Mol Cancer Res. 14:344–353. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Takeuchi H, Taoka R, Mmeje CO, Jinesh GG,
Safe S and Kamat AM: CDODA-Me decreases specificity protein
transcription factors and induces apoptosis in bladder cancer cells
through induction of reactive oxygen species. Urol Oncol.
34:337.e11–e18. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Harada H, Andersen JS, Mann M, Terada N
and Korsmeyer SJ: p70S6 kinase signals cell survival as well as
growth, inactivating the pro-apoptotic molecule BAD. Proc Natl Acad
Sci USA. 98:9666–9670. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chu EC and Tarnawski AS: PTEN regulatory
functions in tumor suppression and cell biology. Med Sci Monit.
10:RA235–RA241. 2004.PubMed/NCBI
|
|
42
|
Glotzer M: Cytokinesis: Centralspindlin
moonlights as a membrane anchor. Curr Biol. 23:R145–R147. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Jiang W, Jimenez G, Wells NJ, Hope TJ,
Wahl GM, Hunter T and Fukunaga R: PRC1: A human mitotic
spindle-associated CDK substrate protein required for cytokinesis.
Mol Cell. 2:877–885. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gaudet S, Branton D and Lue RA:
Characterization of PDZ-binding kinase, a mitotic kinase. Proc Natl
Acad Sci USA. 97:5167–5172. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Testa JR, Zhou JY, Bell DW and Yen TJ:
Chromosomal localization of the genes encoding the kinetochore
proteins CENPE and CENPF to human chromosomes 4q24->q25 and
1q32->q41, respectively, by fluorescence in situ hybridization.
Genomics. 23:691–693. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Nabetani A, Koujin T, Tsutsumi C,
Haraguchi T and Hiraoka Y: A conserved protein, Nuf2, is implicated
in connecting the centromere to the spindle during chromosome
segregation: A link between the kinetochore function and the
spindle checkpoint. Chromosoma. 110:322–334. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Holland AJ and Cleveland DW: Losing
balance: The origin and impact of aneuploidy in cancer. EMBO Rep.
13:501–514. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Johansson H and Simonsson S: Core
transcription factors, Oct4, Sox2 and Nanog, individually form
complexes with nucleophosmin (Npm1) to control embryonic stem (ES)
cell fate determination. Aging (Albany NY). 2:815–822. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mudbhary R, Hoshida Y, Chernyavskaya Y,
Jacob V, Villanueva A, Fiel MI, Chen X, Kojima K, Thung S, Bronson
RT, et al: UHRF1 overexpression drives DNA hypomethylation and
hepatocellular carcinoma. Cancer Cell. 25:196–209. 2014. View Article : Google Scholar : PubMed/NCBI
|