Lung cancer is the leading cause of cancer-related
death worldwide; more than 1.7 million people succumbed to lung
cancer in 2018 (1). Based on origin,
lung cancer can be divided into small cell lung cancer (SCLC) and
non-SCLC (NSCLC), of which NSCLC accounts for 80–85% of the cases
(1). Early lung cancer often lacks
symptoms, which may lead to delayed diagnosis and treatment. In the
late stage, both SCLC and NSCLC can metastasize to other organs;
SCLC can metastasize considerably more rapidly, and patients
develop metastatic symptoms (bone pain, nervous system changes such
as dizziness and seizures, jaundice, enlarged lymph nodes, and/or
other conditions such as syndrome of inappropriate antidiuretic
hormone and Horner syndrome) faster than those with NSCLC. Due to
the large proportion of patients diagnosed with locally advanced or
widely metastatic cancer at the time of diagnosis, the 5-year
relative survival rate for NSCLC is poor, from 68% in patients with
stage IB disease to 0–10% in patients with stage IVA-IVB disease
(2). Although SCLC is characterized
by rapid responses to chemotherapy and sensitivity to radiotherapy,
given the early treatment resistance, the 5-year overall survival
(OS) is <10% (3). Traditional
treatments for lung cancer include chemotherapy, radiotherapy and
surgery. Fortunately, advances in the knowledge of lung cancer and
technologies for its detection have promoted marked progress of
theories and molecular methods in diagnosing lung cancer and have
revolutionized the relevant therapeutics. Researchers have already
extensively described the characteristics of the lung cancer
genome, and several major pathways sensitive to targeted therapy
have been identified (4). Drugs that
target these pathways have improved response and survival in
patients with metastatic disease (5),
some of which have replaced chemotherapy as first-line treatment
drugs. Unfortunately, the efficacy in most patients is limited by
the emergence of resistance mechanisms, while these interventions
are effective initially (6,7). Therefore, investigation of effective
strategies to eliminate these resistant tumor cells is urgently
needed.
In recent decades, a series of studies have reported
the importance of the immune system in malignant disease control,
and immunotherapy has gradually attracted the attention of
researchers (8,9). Inducing passive or active antitumor
responses by the immune system against malignant tumors is an
attractive therapeutic strategy. As a critical part of immune
surveillance, natural killer (NK) cells exhibit cytotoxic activity
against diverse tumor cell types; furthermore, NK cells bridge the
innate and adaptive immune responses (10). With the development of methods to
regulate NK cell function and enhance tumor sensitivity to NK cell
cytotoxicity and the ability to expand NK cells in vitro and
manipulate their homing, numerous NK cell-based immunotherapy
methods and strategies have been developed (9). In physiological conditions, lung tissue
has a considerable amount of NK cells, which may be important
antitumor effector cells of lung tissue. Therefore, immunotherapy
strategies based on NK cells may confer great clinical benefit to
lung cancer treatment. In the present review, the distribution and
function of NK cells, the control effect of NK cells on lung
cancer, and the effect of the lung cancer tumor microenvironment
(TME) on NK cells were briefly introduced and some NK cell-based
immunotherapy strategies were described. Given the advances
summarized in the present review, an exciting future for NK
cell-based cancer immunotherapy is foreseen and the challenges that
remain to be tackled are presented. Although enormous steps have
been taken in understanding NK cell biology, more work is required
to fully explore the anticancer potential of these cells.
A search for scientific papers published between
1975 and 2020 focusing on NK cells, lung cancer and NK cell-based
immunotherapy was performed in PubMed. The search terms used were
‘NK cell’, ‘lung’, ‘cancer’, ‘immunotherapy’, ‘tumor
microenvironment’, ‘cytokine’, ‘monoclonal antibodies’, ‘adoptive
transfer’, ‘CAR’, alone and in combination. A total of 176
scientific papers were selected, 117 of which were original
studies.
NK cells are innate lymphocytes that can directly
eliminate target cells without prior exposure (11,12) and
play a key role in antiviral and antitumor immunity. NK cells,
mainly present in the peripheral blood, comprise approximately 15%
of all circulating lymphocytes (13),
while they are also distributed in multiple tissues including the
liver, lung, skin, kidney and bone marrow. Moreover, based on the
expression of CD49a (i.e., integrin α1), CD69 and CD103 (i.e.,
integrin aE) (14–17), NK cells can be subdivided into
circulating and tissue-resident NK cells. Tissue-resident NK cells
usually display high expression of CD49a, CD103, and CD69 (18). More commonly, researchers subdivide
human NK cells into two major subsets with distinct maturation and
functional properties according to the expression of CD56 and the
antibody binding-Fc receptor CD16 (13). CD56brightCD16−
NK cells (approximately 10% of NK cells in the peripheral blood)
are specialized in secreting cytokines and are abundantly located
in secondary lymphoid organs (lymph nodes, tonsils, and spleen)
(19), most of which exhibit
characteristics of tissue-resident lymphocytes and tissue-specific
adaptations. Furthermore, they can also reveal cytotoxicity under
prolonged stimulation with cytokines such as interleukin (IL)-15,
IL-12, and IL-18 (13,20–24).
CD56dimCD16+ NK cells (approximately 90–95%
of NK cells in the peripheral blood) (12) are potent cytolytic effector cells,
which can rapidly secrete pro-inflammatory cytokines such as
interferon (IFN)-γ and cytotoxic mediators such as granzyme once
activated. Most of them exhibit characteristics of circulating
cells, but they can also show a resident phenotype while located in
the lymph nodes, mucosa, and other parts.
NK cells exhibit marked mobility, thereby
circulating between organs to promote immune surveillance (38). NK cells can respond to multiple
chemokines and be recruited to different tissues or inflammation
sites owing to the expression of several chemokine receptors
(39). The lung is a critical organ
of body-environment interaction and is rich in NK cells (40,41). It is
generally considered that NK cells originate and develop in the
bone marrow and then migrate to the lung (42). The proportion of NK cells in the lung
is similar to, and even slightly higher, than that in the
peripheral blood, accounting for 10–20% of the lymphocytes in the
lung (43). IL-15 secreted by
bronchial epithelial cells and alveolar macrophages may be
responsible for the high proportion of NK cells (44,45)
because it is the main cytokine supporting NK cell cytotoxicity,
homeostasis, and development (46,47).
Unlike the liver and secondary lymphoid organs rich in
CD56brightCD16− NK cell subpopulation, most
NK cells in the lung exhibit the CD56dimCD16+
phenotype (48), indicating that most
are circulating subsets and highly differentiated (49,50).
Despite their high differentiation, human NK cells in the lung
exhibit a weaker response to target cell stimulation than
peripheral blood NK cells (51),
which may be attributable to the inhibition of alveolar macrophages
(52) and soluble factors of the
lower respiratory tract (51).
Perhaps because the pulmonary mucosa is continuously exposed to the
environment and autoantigens, NK cells with restricted function in
physiological conditions may be more conducive to the maintenance
of pulmonary homeostasis (49).
Although circulating CD56dimCD16+ NK cells
are the major subpopulation in the lung (51), CD49a+ tissue-resident NK
cells (mainly CD56brightCD16− NK cells) also
account for approximately 15% of human NK cells in this organ
(53). Studies have revealed that
CD56brightCD49a+ NK cells in the lung
strongly co-express CD103 and CD69, significantly different from
CD56brightCD16−NK cells in the peripheral
blood (18,53). In in vitro experiments,
CD49a+ tissue-resident NK cells exhibited a higher
ability to degranulate and produce IFN-γ when in contact with
virus-infected autologous macrophages than NK cells in the
peripheral blood (50). Collectively,
these results indicated that circulating NK cells in the lung have
a larger number and highly differentiated phenotype but exhibit
depressed function, while tissue-resident NK cells have stronger
function. It is necessary to further study the characteristics of
circulating and tissue-resident NK cells in the lung to understand
their roles in the physiological condition and in the occurrence
and progression of lung tumors, which may provide novel directions
for the development of therapeutic strategies.
In the 1980s, some studies revealed that cancer
incidence was higher among individuals with NK cell dysfunction
(54,55). Since then, studies have increasingly
confirmed that the antitumor effect of NK cells can act against
multiple tumor types (56,57), including head and neck (58), pharyngeal (59), colorectal (60), and lung (38,61)
cancers. The direct evidence that NK cells act against lung cancer
is supported by Kras-driven spontaneous lung cancer and cancer cell
implantation experiments in mice (61,62), both
of which revealed that mice lacking NK cells have a greater lung
tumor burden. However, the antitumor effect is limited to the early
stage of Kras-driven lung cancer in mice due to NK cell dysfunction
in the advanced stage (61).
In the past few years, some studies have revealed
that NK cells can infiltrate lung cancer and that the number of
tumor-infiltrating NK (TINK) cells is significantly associated to
postoperative patient survival, indicating that the infiltration of
NK cells into tumors may benefit patient prognosis (63–65).
Similar phenomena have been observed in patients with breast cancer
(66) and renal clear cell carcinoma
(67). Interestingly, most TINK cells
are of the CD56brightCD16− NK type (68), and they only exist in the intratumoral
fibrous septum and the interface between stromal and surrounding
tumor cells, which appear to indirectly be in contact with cancer
cells (68,69). Conversely, in renal clear cell
carcinoma, NK cells infiltrate the entire tumor tissue (70). Although the mechanism of TINK
enrichment remains unclear, homing restriction and an
immunosuppressive microenvironment may play an important role
(71–73). A previous study revealed that the
proportion of CD56brightCD16− NK cells in
tumoral and non-tumoral lung tissues is similar (74), indicating that the enrichment of
CD56brightCD16− NK cells in tumors may be
driven by the rejection of CD56dimCD16+ NK
cells by the tumor. Conversely, it may be related to the chemokine
spectrum of CD56dimCD16+ NK cell subsets. For
example, it has already been confirmed that the adhesion signal of
heterodimerization of chemokine receptor CCR5 (i.e., MIP-1β
receptor), which is only expressed by CD56bright
CD16− NK cells, could force leukocytes to stay in the
tissue (75,76). In contrast, the viability of
CD56dimCD16+ NK cells may be impaired in the
TME. Interestingly, it is generally considered that tumor rejection
is mainly due to direct killing by lymphocytes. However, some
studies have revealed that IFN-γ and other lymphocyte-derived
cytokines such as TNF-α can promote tumor rejection to control
tumor progression (77,78), which indicates that tumor rejection is
a more complicated event than previously considered and that the
cytokine secretion function of tumor-infiltrating
CD56brightCD16− NK cells cannot be ignored in
tumor control.
In summary, NK cells can infiltrate and eliminate
tumor cells; therefore, targeting NK cells through immunotherapy is
an attractive anticancer strategy. Based on the available
literature, it can be theorized that the localization of NK cells
in tumors (NK cells are most successful in the treatment of
hematopoietic malignancies such as leukemia because NK cells are
abundant in the peripheral blood) may be an essential factor of NK
cell-based immunotherapy. Exploring the role of NK cells in
survival and the lung tumor environment may enable the development
of methods to improve the ability of NK cells to migrate and
infiltrate into tumor tissues, thereby effectively improving the
antitumor immunity of the body.
Although it has been determined that NK cells have
antitumor effects, malignant tumors continue to develop in the
presence of NK cells, which does not mean that NK cells do not
contribute to tumor control, but that their antitumor activity may
be impaired to some extent (79,80). In
this regard, the TME, which is composed of cell components, growth
factors, proteases, extracellular matrix, and lymphatic and
vascular systems, plays an important role (81). The TME allows tumor cells to obtain
cancer markers, establishing a chronic inflammatory environment
that maintains tumor growth and induces dysfunction of NK cells in
various ways (79,82). An increasing number of studies have
revealed that the phenotype and function of NK cells are altered in
the tumor microenvironment (68–69,79). A
comprehensive understanding of the factors and mechanisms that
cause NK cell changes in the TME may help reveal means to restoring
their antitumor potential.
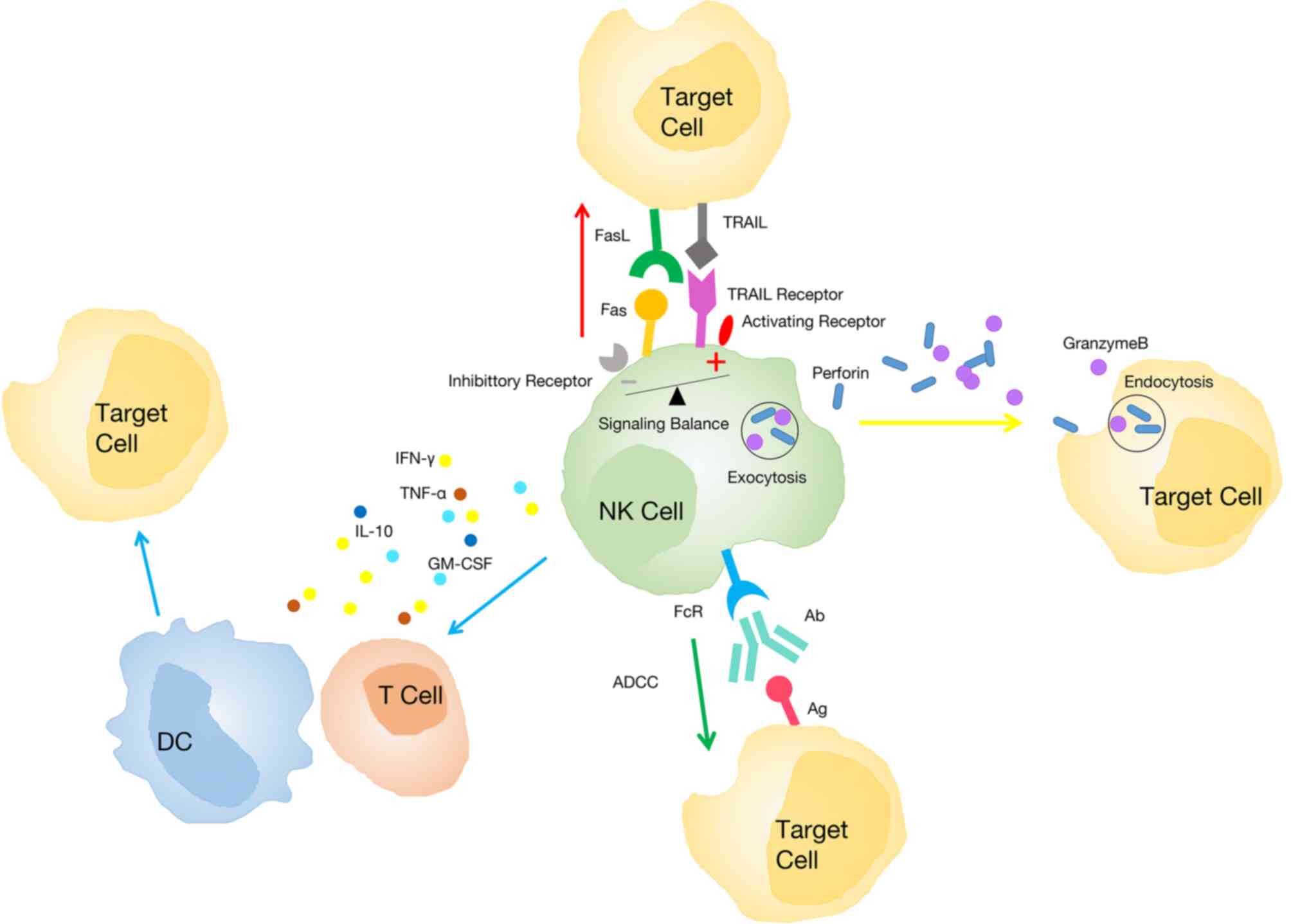
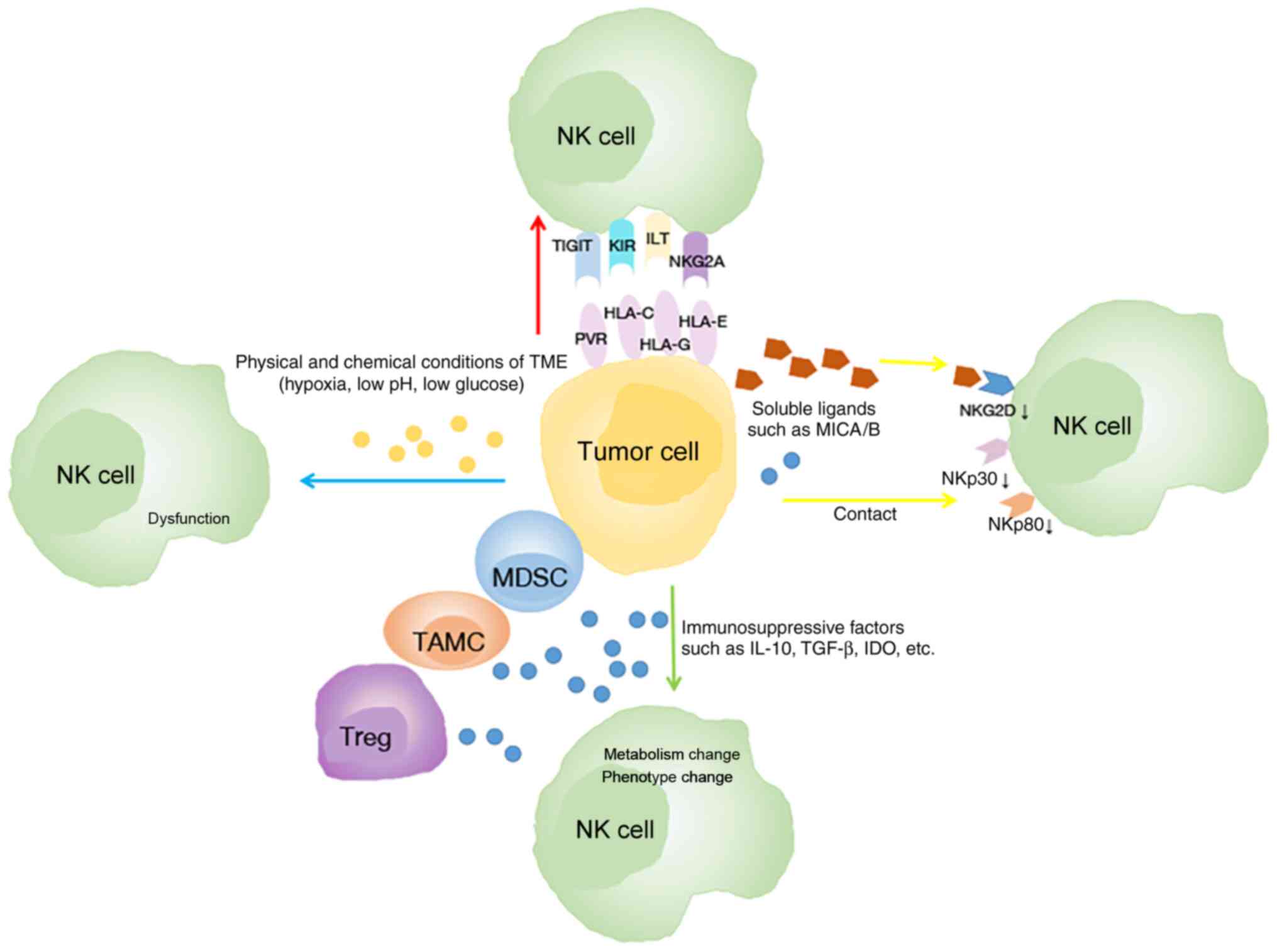
Several mechanisms have been revealed to be related
to the phenotype and function alterations of NK cells (Fig. 2). Firstly, tumor cells can affect the
phenotype of NK cells depending on cell-to-cell contact (83). In lung cancer, an in vitro
Transwell experiment revealed that the communication between NK and
tumor cells is associated with the downregulation of active
receptors including NKp30, NKp80, DNAM-1, and NKG2D on the surface
of TINKs (69). Another study
revealed that the expression of CD155 on tumor cells is related to
the downregulation of DNAM-1 on NK cells in NSCLC (84). Conversely, the inhibitory receptors of
NK cells are upregulated in cancer. In humans, the expression of
T-cell immunoglobulin and ITIM domain (TIGIT) on NK cells was
further upregulated in tumor regions compared with peritumoral
regions in colorectal tumors (85).
Secondly, the modification of the NK cell phenotype
can be altered due to the high expression of immunosuppressive
factors such as transforming growth factor (TGF)-β, indoleamine
2,3-dioxygenase (IDO), IL-4, and prostaglandin E2 (PGE2) (86–89). TGF-β
has been revealed to be overexpressed in lung cancer cells
(90,91), and its expression level can be a
prognostic marker in lung cancer (92). Some mechanisms by which TGF-β inhibits
the function of NK cells have been identified in lung cancer: i)
TGF-β changes the receptor spectrum of NK cells in patients with
lung cancer (68,69,93). TGF-β
downregulates the expression of NK activating receptors NKp80,
NKp30, and NKG2D (68,69,94),
thereby inhibiting the cytolytic activity of NK cells (86,94).
Neutralizing TGF-β inhibits the downregulation of NKG2D expression
and restores the antitumor response of NK cells (93,95);
conversely, TGF-β upregulates inhibitory receptors including NKG2A
(96) and programmed cell death-1
(PD-1) (97) in NK cells in tumors
(96,98). ii) TGF-β affects the metabolism of NK
cells in lung cancer. In Kras-driven lung cancer, high levels of
TGF-β in the TME cause aberrant expression of the fructose
1,6-biophosphatase (FBP1) protein in NK cells, thereby inhibiting
NK cell glycolysis and reducing cellular activity, eventually
leading to NK cell dysfunction (61);
iii) TGF-β mediates NK cell polarization toward angiogenesis
(99). Furthermore, another study
revealed that the expression of PGE2, which inhibited the antitumor
activity of NK cells in NSCLC tumor tissue, was significantly
increased (100). Notably, immune
cells which are a major component of the TME, inhibit NK cell
function mainly by secreting immunosuppressive molecules (101–103).
Studies have revealed that the number of myeloid-derived suppressor
cells (MDSCs) (104) and regulatory
T cells (Tregs) (105) in the lung
cancer TME is higher than that in normal tissues and peripheral
blood adjacent to cancer, both of which can secrete TGF-β (86,93,106,107).
In addition, the physical and chemical conditions of
the TME, including hypoxia, low pH and low glucose concentration,
can also impair NK cell function (108). Previous studies have confirmed that
hypoxia downregulates the expression of NCR and NKG2D on NK cells
(109) and damages their
cytotoxicity (110). In NSCLC, high
HIF-1α levels of tumor negatively impact (111,112)
the OS of patients; the associated mechanisms may include adenosine
generation and accumulation, lactate accumulation and extracellular
acidosis. Both adenosine accumulation and extracellular acidosis
can block NK cell activation, proliferation and cytotoxicity
(113,114), while lactate accumulation mainly
inhibits the cytotoxic activity of NK cells and increases the
number of MDSCs that inhibit NK cytotoxicity (115).
Clearly, various components of the TME affect the
antitumor functional activity of NK cells in different ways during
the progression of lung cancer. Among them, TGF-β is the main
inhibitor of NK cell function. Notably, intratumoral NK cells may
have a negative effect on other immune cells located in the TME
after their own antitumor function decreases. For example, DC
maturation was impaired due to the lack of IFN-γ secretion by NK
cells and Tregs were profusely recruited through CCL22 secretion
induced by NK cells (116,117). An important question is whether the
modifications in the intratumoral NK phenotype and function are
reversible. If they are, enhancing NK cell function with
immuno-stimulatory cytokines such as IL-15 or by neutralization of
immunosuppressive factors produced in the environment may improve
the efficacy of NK cell-based immunotherapy and further ameliorate
the clinical outcome of lung cancer.
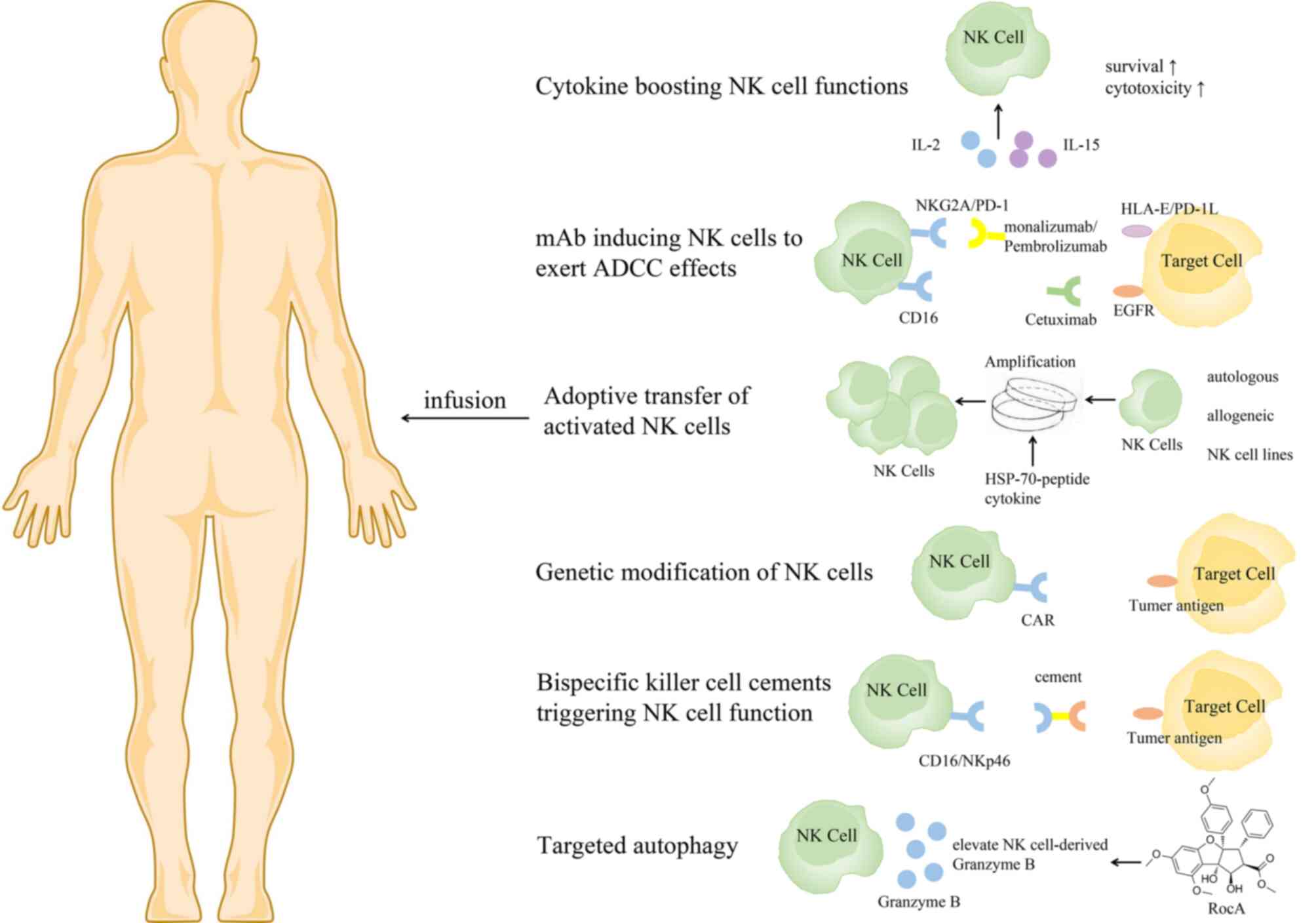
Based on the important role of NK cells in tumor
control, NK cell immunotherapy has developed rapidly. Several
approaches have recently been proposed to boost NK cell antitumor
function, to support in vivo persistence and homeostatic
proliferation, and to promote homing to the tumor microenvironment
(33) (Fig.
3).
It has been reported that IL-2, IL-15 and IL-18 can
enhance the proliferation ability of NK cells and improve their
antitumor function (48,118). Meta-analysis results have revealed
that IL-2 treatment can significantly improve OS in patients with
NSCLC (119). However, IL-2 is not
the best choice because Tregs can be preferentially activated by
IL-2, thereby inhibiting NK cell proliferation and cytotoxicity
(120,121). IL-15 is an alternative form of IL-2,
which preferentially stimulates NK cells without activating Tregs
(122). Treatment of drug-resistant
solid tumors (including NSCLC) with subcutaneous injection of
recombinant human (rh) IL-15 could significantly promote the
proliferation of peripheral blood NK cells, especially the
proliferation of CD56brightNK cells, in a phase I
non-randomized trial (123). As a
super-agonist of IL-15, ALT-803 could encourage the growth of NK
cells, induce the expression of NKG2D and the production and
release of IFN-γ, and enhance the role of ADCC (124). In a phase 1b clinical trial,
patients with metastatic NSCLC treated with ALT-803 and nivolumab
exhibited a high tumor response rate and the treatment was well
tolerated (125).
mAbs that induce NK cells to exert ADCC effects
include mAbs targeting tumor-associated antigens such as rituximab
or cetuximab that recognize CD20 and epidermal growth factor
receptor (EGFR) and antibodies against inhibitory molecules such as
monalizumab that recognizes NKG2A (126–129).
Nowadays, immunotherapy targeting the PD-1/PD-L1 inhibitory axis is
considered a treatment pillar in NSCLC (130–135). A
recent study revealed that TINK cells from patients with NSCLC
expressed increasing immune checkpoint receptor PD-1 on their
surface, which correlated with their dysfunction (132). Notably, treatment with PD-1 blocking
antibodies could reverse PD-L1-mediated inhibition of NK cells
(132), highlighting the critical
role of PD-1+ NK cells in immune checkpoint blockade for
NSCLC. In addition, TIGIT immune checkpoint inhibitors have been
revealed to prevent NK cell depletion and elicit effective
tumor-specific T-cell immunity in an NK cell-dependent manner
(85).
Adoptive transfer of NK cells with high yield and
high quality is the most direct means to restoring and improving
the function of the immune system. The outcome of adoptive transfer
of NK cells can vary due to differences in the strategies used for
the separation, expansion and activation of NK cells (136,137).
NK cells can be derived from either autologous or allogeneic
sources, either peripheral blood mononuclear cell (PBMC), stem cell
(including umbilical cord blood, embryonic stem cells and induced
pluripotent stem cells) or NK cell lines. It has been proven that
transferring autologous NK cells into patients is safe (138). An experiment using adoptive NK cells
to treat melanoma revealed that the adoptively transferred NK cells
persisted in the peripheral circulation of patients for at least 1
week post-transfer and exhibited high levels of lytic activity
in vitro but had no effect on tumor regression (139). The limited effect may be
attributable to the KIR-ligands of tumors always matching the
autologous NK cell KIR repertoire and the suppression by self MHC
class I that enables malignant cells to evade NK-mediated
elimination. Thus, strategies have been developed to overcome this
limitation, such as use of an anti-KIR antibody (140).
The source of cells is another issue to consider.
Given the small percentage of NK cells of PBMCs, purified NK cells
must be expanded ex vivo to attain the requirements for
clinical use, but developing strategies to yield an adequate cell
number remains a major challenge. Thus, stem cell-derived NK cells
are gradually becoming a focus of research. Differentiation of
mature, functional NK cells can be achieved through the co-culture
of bone-marrow- or umbilical cord blood-derived CD34+
hematopoietic stem cells with IL-2/IL-15 and various growth factors
(145). Compared with PBMC-derived
and stem cell-derived NK cells, NK cell lines are easier to expand.
Several cytotoxic cell lines including KHYG-1, NK-92, and NKL are
gradually becoming a powerful tool for NK cell-based immunotherapy
(136,137). However, the lack of in vivo
persistence and CD16 expression in most cell lines limits their
clinical use. Fortunately, transgene expression can promote the
expression of CD16 (145). Moreover,
an in vitro experiment revealed that NK92-CD16 cells have
greater cytotoxic potential against tyrosine kinase
inhibitor-resistant NSCLC cells than their parental NSCLC cells
(146).
Furthermore, to obtain more robust cytotoxic
activity of NK cells, immunostimulatory molecules such as cytokines
are usually used in combination with NK cells in clinical research.
Studies have confirmed that adoptive transfer of NK cells
stimulated by IL-15 is effective in the treatment of patients with
advanced NSCLC (130,147). Notably, ex vivo heat shock
protein (HSP)-70-peptide stimulates NK cells and improves their
function. HSPs are usually synthesized when cells react to various
stress-inducing or toxic factors (148). Most HSPs are molecular chaperones,
and promoting the synthesis of HSPs can improve the function of the
chaperone machinery and lead to reduction of cell sensitivity to
repeated action of the same or other stressful agents (149). HSPs are overexpressed in various
cancers, and their increased expression is generally associated
with tumor cell survival, invasion, metastasis and chemoresistance
(150–152). A substantial number of studies have
reported a relatively higher risk of lung cancer with increased
expression of HSP-70, and their levels correlated with the grade
and stage of lung tumors (153,154).
Another study revealed that membrane-bound HSP-70 acts as a
tumor-specific marker enhancing NK cell activity (155). Subsequently, several clinical trials
have revealed that ex vivo HSP-70-peptide-activated,
autologous NK cells are well tolerated and deliver positive
clinical responses in patients with advanced NSCLC (156–158).
This may be a promising treatment for lung cancer.
Genetic modification can induce profound and
sustained genetic changes in NK cells (159). Among them, chimeric antigen receptor
(CAR) NK cells have attracted increasing attention (160,161).
The advantages of CAR NK cells over CAR T cells are MHC
independence, lack of graft-versus-host response and a relatively
limited lifespan (avoiding the need to insert suicide genes into
CAR NK cell constructs). Similar to adoptive transfer, the sources
of CAR NK cells are diverse and include peripheral blood NK cells,
primary cord blood-derived NK cells and the NK cell line NK-92;
recent studies have tested their effectiveness (162,163). A
previous study has revealed that cord blood NK cells engineered to
express IL-15 and a CD19-targeted CAR have long-term persistence
and potent antitumor activity and are easy to produce (164). In a phase 2 trial, among 11 patients
with relapsed or refractory CD19-positive lymphoid tumors, eight
patients exhibited a response to treatment with cord blood-derived
CAR-NK cells without the development of major toxic effects
(165).
Transformed cell line NK-92, originating from
undifferentiated NK-cell precursors, is also commonly used
(166). To date, NK-92 has been
intensively studied; both preclinical mouse studies and phase I
clinical testing have confirmed its safety in patients and
cytotoxicity against several tumor types, particularly against lung
tumors (167–169). In an in vitro study, a novel
chimeric costimulatory converting receptor-modified NK92, which
comprised the extracellular domain of PD-1, transmembrane and
cytoplasmic domains of NKG2D, and the cytoplasmic domain of 4-1BB,
exhibited enhanced antitumor activity against human lung cancer
H1299 cells (170). Given that CAR
NK cells have favorable application prospects, some measures need
to be implemented to develop a more intelligent next generation.
First, non-viral vector methods should be developed to avoid the
insertion mutations induced by retroviral transfection. Second,
establishing improved clinical-grade protocols for purifying NK
cells to avoid T-cell contamination which may lead to
graft-versus-host disease or lymphoproliferative disorders
(171,172). Third, establishing CAR ligand
bi-specific CAR molecules or silencing NK inhibitory receptors
during the design of CAR-NK cells may further improve the efficacy
of CAR-NK cell therapy.
Recently, bispecific killer-cell cements have been
designed to promote lysis of tumor cells by NK cells (173,174).
These NK cell adaptors enable the killing effect of NK cells on
tumor cells by targeting activation receptors NKp46 and CD16 via
tumor antigens (such as CD19, CD20, or EGFR) and Fc fragments,
respectively. Two CD16-based bispecific antibodies with EGFR
variants and wild-type EGFR (AFM22 and AFM24) are in preclinical
development. In vivo, these antibodies effectively control
tumor growth in mouse models of solid and invasive tumors (81).
Targeting autophagy is a new strategy in cancer
immunotherapy. A recent study revealed that rocaglamide inhibits
autophagy and restores the level of NK cell-derived granzyme B in
NSCLC, enhances NK cell-mediated lysis of lung cancer cells, and
causes tumor regression in vivo (175).
Various applications of NK cells to lung cancer
treatment are increasingly being attempted, denoting that there is
major progress in NK cell research (Table
I). Additional studies in patients with lung cancer are still
required to realize the antitumoral potential of NK cells and
establish its clinical applications.
In recent years, molecular targeted therapy and
immune checkpoint inhibitor therapy have led to marked progress in
the treatment of lung cancer. However, a considerable number of
patients remain unresponsive to treatment, and the need for new
treatment strategies is still urgent. Numerous studies have
confirmed the critical role of NK cells in lung cancer control.
Immunotherapy targeting NK cells may be an effective strategy for
lung cancer treatment. The growing insight into the NK cell
potential for lung cancer treatment provides a platform for the
development of NK-based immunotherapy. However, numerous obstacles
remain to be overcome to derive the full benefit of the NK cell
antitumor potential. First, the poor ability of NK cells to reach
tumor tissues limits their application in solid tumor therapy,
which is a common problem with cellular immunotherapy strategies.
As aforementioned, when NK cells are present in tumor tissues, they
are preferentially localized in the matrix without coming into
contact with the tumor cells. Second, changes in NK cell-activated
receptors and ligands in tumors may result in decreased antitumor
activity. Finally, the TME remains the main obstacle to the
effectiveness of the adoptive transfer of NK cells. Despite these
challenges, as more data are gathered on the lung cancer TME,
immune regulatory cell populations, cancer-related changes in NK
cell biology, function, and transport, NK cell immunotherapy will
become increasingly effective. Key components to the success of
future trials include the incorporation of modalities that harness
NK cell cytotoxicity while promoting in vivo survival,
homeostatic proliferation, and trafficking to the tumor and the
development of drugs that trigger NK cell tumor killing via ADCC or
sensitization of the target and drugs that promote NK cell tumor
homing, such as the development of monoclonal antibody-chemokine
fusion proteins to promote the infiltration of cytolytic NK cells
into tumor tissues. Efforts should be made to solve the problems of
clonal expansion and genetic modification of NK cells. Currently,
several phase I and II clinical trials for the treatment of
targeted NK cells for lung cancer are underway, including
chemotherapy combined with NK cell adoptive transfer therapy
(NCT03366064, NCT03410368), immune checkpoint inhibitors combined
with NK cell adoptive transfer (NCT03958097), surgery combined with
NK cell adoptive transfer therapy (NCT02843815), and CAR NK cell
therapy (NCT03656705). In the future, combined standard
radiotherapy, chemotherapy or radiochemotherapy, targeted therapy,
ex vivo stimulation or CAR-NK cells and other targeted NK
cell methods may eventually change the treatment mode of lung
cancer, providing hope to patients with limited treatment
options.
Not applicable.
The present work was supported by the National
Natural Science Foundation of China (grant no. 81902895), the
National Key Sci-Tech Special Project of China (grant no.
2018ZX10302207), the Beijing Natural Science Foundation (grant no.
M21007) and the Beijing Hospital Authority (grant no.
DFL20191801).
Not applicable.
YZ conceptualized and wrote the manuscript. XL and
JD contributed to the literature review and organization of the
manuscript. All authors confirm the authenticity of all the raw
data. All authors read and approved the final manuscript.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Yuan M, Huang LL, Chen JH, Wu J and Xu Q:
The emerging treatment landscape of targeted therapy in
non-small-cell lung cancer. Signal Transduct Target Ther. 4:612019.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Goldstraw P, Chansky K, Crowley J,
Rami-Porta R, Asamura H, Eberhardt WE, Nicholson AG, Groome P,
Mitchell A, Bolejack V, et al: The IASLC lung cancer staging
project: Proposals for revision of the TNM stage groupings in the
forthcoming (eighth) edition of the TNM classification for lung
cancer. J Thorac Oncol. 11:39–51. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Planchard D, Popat S, Kerr K, Novello S,
Smit EF, Faivre-Finn C, Mok TS, Reck M, Van Schil PE, Hellmann MD,
et al: Metastatic non-small cell lung cancer: ESMO clinical
practice guidelines for diagnosis, treatment and follow-up. Ann
Oncol. 29 (Suppl 4):iv192–iv237. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen Z, Fillmore CM, Hammerman PS, Kim CF
and Wong KK: Non-small-cell lung cancers: A heterogeneous set of
diseases. Nat Rev Cancer. 14:535–546. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Mayekar MK and Bivona TG: Current
landscape of targeted therapy in lung cancer. Clin Pharmacol Ther.
102:757–764. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xie Q, Yu Z, Lu Y, Fan J, Ni Y and Ma L:
MicroRNA-148a-3p inhibited the proliferation and
epithelial-mesenchymal transition progression of non-small-cell
lung cancer via modulating Ras/MAPK/Erk signaling. J Cell Physiol.
234:12786–12799. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fan Q, Hu X, Zhang H, Wang S, Zhang H, You
C, Zhang CY, Liang H, Chen X and Ba Y: miR-193a-3p is an important
tumour suppressor in lung cancer and directly targets KRAS. Cell
Physiol Biochem. 44:1311–1324. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Steven A, Fisher SA and Robinson BW:
Immunotherapy for lung cancer. Respirology. 21:821–833. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shimasaki N, Jain A and Campana D: NK
cells for cancer immunotherapy. Nat Rev Drug Discov. 19:200–218.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Vivier E, Raulet DH, Moretta A, Caligiuri
MA, Zitvogel L, Lanier LL, Yokoyama WM and Ugolini S: Innate or
adaptive immunity? The example of natural killer cells. Science.
331:44–49. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Herberman RB, Nunn ME, Holden HT and
Lavrin DH: Natural cytotoxic reactivity of mouse lymphoid cells
against syngeneic and allogeneic tumors. II. Characterization of
effector cells. Int J Cancer. 16:230–239. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kiessling R, Klein E, Pross H and Wigzell
H: ‘Natural’ killer cells in the mouse. II. Cytotoxic cells with
specificity for mouse Moloney leukemia cells. Characteristics of
the killer cell. Eur J Immunol. 5:117–121. 1975. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cooper MA, Fehniger TA and Caligiuri MA:
The biology of human natural killer-cell subsets. Trends Immunol.
22:633–640. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sojka DK, Plougastel-Douglas B, Yang L,
Pak-Wittel MA, Artyomov MN, Ivanova Y, Zhong C, Chase JM, Rothman
PB, Yu J, et al: Tissue-resident natural killer (NK) cells are cell
lineages distinct from thymic and conventional splenic NK cells.
Elife. 3:e016592014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tang L, Peng H, Zhou J, Chen Y, Wei H, Sun
R, Yokoyama WM and Tian Z: Differential phenotypic and functional
properties of liver-resident NK cells and mucosal ILC1s. J
Autoimmun. 67:29–35. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Björkström NK, Ljunggren HG and
Michaëlsson J: Emerging insights into natural killer cells in human
peripheral tissues. Nat Rev Immunol. 16:310–320. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lugthart G, Melsen JE, Vervat C, van
Ostaijen-Ten Dam MM, Corver WE, Roelen DL, van Bergen J, van Tol
MJ, Lankester AC and Schilham MW: Human lymphoid tissues harbor a
distinct CD69+CXCR6+ NK cell population. J Immunol. 197:78–84.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Marquardt N, Kekäläinen E, Chen P, Lourda
M, Wilson JN, Scharenberg M, Bergman P, Al-Ameri M, Hård J, Mold
JE, et al: Unique transcriptional and protein-expression signature
in human lung tissue-resident NK cells. Nat Commun. 10:38412019.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cooper MA, Fehniger TA, Turner SC, Chen
KS, Ghaheri BA, Ghayur T, Carson WE and Caligiuri MA: Human natural
killer cells: A unique innate immunoregulatory role for the
CD56(bright) subset. Blood. 97:3146–3151. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fehniger TA and Caligiuri MA: Interleukin
15: Biology and relevance to human disease. Blood. 97:14–32. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wagner JA, Rosario M, Romee R,
Berrien-Elliott MM, Schneider SE, Leong JW, Sullivan RP, Jewell BA,
Becker-Hapak M, Schappe T, et al: CD56bright NK cells exhibit
potent antitumor responses following IL-15 priming. J Clin Invest.
127:4042–4058. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fauriat C, Long EO, Ljunggren HG and
Bryceson YT: Regulation of human NK-cell cytokine and chemokine
production by target cell recognition. Blood. 115:2167–2176. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Caligiuri MA, Zmuidzinas A, Manley TJ,
Levine H, Smith KA and Ritz J: Functional consequences of
interleukin 2 receptor expression on resting human lymphocytes.
Identification of a novel natural killer cell subset with high
affinity receptors. J Exp Med. 171:1509–1526. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nagler A, Lanier LL and Phillips JH:
Constitutive expression of high affinity interleukin 2 receptors on
human CD16-natural killer cells in vivo. J Exp Med. 171:1527–1533.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Leibson PJ: Signal transduction during
natural killer cell activation: Inside the mind of a killer.
Immunity. 6:655–661. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lanier LL: On guard-activating NK cell
receptors. Nat Immunol. 2:23–27. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yoon SR, Kim TD and Choi I: Understanding
of molecular mechanisms in natural killer cell therapy. Exp Mol
Med. 47:e1412015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Moretta L, Bottino C, Pende D, Mingari MC,
Biassoni R and Moretta A: Human natural killer cells: Their origin,
receptors and function. Eur J Immunol. 32:1205–1211. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ljunggren HG and Kärre K: In search of the
‘missing self’: MHC molecules and NK cell recognition. Immunol
Today. 11:237–244. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Osińska I, Popko K and Demkow U: Perforin:
An important player in immune response. Cent Eur J Immunol.
39:109–115. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Smyth MJ and Johnstone RW: Role of TNF in
lymphocyte-mediated cytotoxicity. Microsc Res Tech. 50:196–208.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Screpanti V, Wallin RP, Ljunggren HG and
Grandien A: A central role for death receptor-mediated apoptosis in
the rejection of tumors by NK cells. J Immunol. 167:2068–2073.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Crinier A, Narni-Mancinelli E, Ugolini S
and Vivier E: SnapShot: Natural killer cells. Cell.
180:1280–1280.e1. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee H, Schlereth SL, Park EY, Emami-Naeini
P, Chauhan SK and Dana R: A novel pro-angiogenic function for
interferon-γ-secreting natural killer cells. Invest Ophthalmol Vis
Sci. 55:2885–2892. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Martín-Fontecha A, Thomsen LL, Brett S,
Gerard C, Lipp M, Lanzavecchia A and Sallusto F: Induced
recruitment of NK cells to lymph nodes provides IFN-gamma for T(H)1
priming. Nat Immunol. 5:1260–1265. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shankaran V, Ikeda H, Bruce AT, White JM,
Swanson PE, Old LJ and Schreiber RD: IFNgamma and lymphocytes
prevent primary tumour development and shape tumour immunogenicity.
Nature. 410:1107–1111. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zhang M, Wen B, Anton OM, Yao Z, Dubois S,
Ju W, Sato N, DiLillo DJ, Bamford RN, Ravetch JV and Waldmann TA:
IL-15 enhanced antibody-dependent cellular cytotoxicity mediated by
NK cells and macrophages. Proc Natl Acad Sci USA.
115:E10915–E10924. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Malmberg KJ, Carlsten M, Björklund A,
Sohlberg E, Bryceson YT and Ljunggren HG: Natural killer
cell-mediated immunosurveillance of human cancer. Semin Immunol.
31:20–29. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Fu X, Yang B, Lao S, Fan Y and Wu C: Human
memory-like NK cells migrating to tuberculous pleural fluid via
IP-10/CXCR3 and SDF-1/CXCR4 axis produce IFN-γ in response to
Bacille Calmette Guerin. Clin Immunol. 148:113–123. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Culley FJ: Natural killer cells in
infection and inflammation of the lung. Immunology. 128:151–163.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Grégoire C, Chasson L, Luci C, Tomasello
E, Geissmann F, Vivier E and Walzer T: The trafficking of natural
killer cells. Immunol Rev. 220:169–182. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Trinchieri G: Biology of natural killer
cells. Adv Immunol. 47:187–376. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hervier B, Russick J, Cremer I and
Vieillard V: NK cells in the human lungs. Front Immunol.
10:12632019. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ge N, Nishioka Y, Nakamura Y, Okano Y,
Yoneda K, Ogawa H, Sugita A, Yanagawa H and Sone S: Synthesis and
secretion of interleukin-15 by freshly isolated human bronchial
epithelial cells. Int Arch Allergy Immunol. 135:235–242. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shi FD, Ljunggren HG, La Cava A and Van
Kaer L: Organ-specific features of natural killer cells. Nat Rev
Immunol. 11:658–671. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ma A, Koka R and Burkett P: Diverse
functions of IL-2, IL-15, and IL-7 in lymphoid homeostasis. Annu
Rev Immunol. 24:657–679. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Floros T and Tarhini AA: Anticancer
cytokines: Biology and clinical effects of interferon-α2,
interleukin (IL)-2, IL-15, IL-21, and IL-12. Semin Oncol.
42:539–548. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Tomasello E, Yessaad N, Gregoire E,
Hudspeth K, Luci C, Mavilio D, Hardwigsen J and Vivier E: Mapping
of NKp46(+) cells in healthy human lymphoid and non-lymphoid
tissues. Front Immunol. 3:3442012. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang J, Li F, Zheng M, Sun R, Wei H and
Tian Z: Lung natural killer cells in mice: Phenotype and response
to respiratory infection. Immunology. 137:37–47. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Marquardt N, Kekäläinen E, Chen P,
Kvedaraite E, Wilson JN, Ivarsson MA, Mjösberg J, Berglin L,
Säfholm J, Manson ML, et al: Human lung natural killer cells are
predominantly comprised of highly differentiated hypofunctional
CD69-CD56dim cells. J Allergy Clin Immunol.
139:1321–1330.e4. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Robinson BW, Pinkston P and Crystal RG:
Natural killer cells are present in the normal human lung but are
functionally impotent. J Clin Invest. 74:942–950. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Weissman DN, deShazo RD and Banks DE:
Modulation of natural killer cell function by human alveolar
macrophages. J Allergy Clin Immunol. 78((4 Pt 1)): 571–577. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Cooper GE, Ostridge K, Khakoo SI,
Wilkinson TMA and Staples KJ: Human CD49a(+) lung natural killer
cell cytotoxicity in response to influenza a virus. Front Immunol.
9:16712018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Roder JC, Haliotis T, Klein M, Korec S,
Jett JR, Ortaldo J, Heberman RB, Katz P and Fauci AS: A new
immunodeficiency disorder in humans involving NK cells. Nature.
284:553–555. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Sullivan JL, Byron KS, Brewster FE and
Purtilo DT: Deficient natural killer cell activity in x-linked
lymphoproliferative syndrome. Science. 210:543–545. 1980.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Pross HF and Lotzová E: Role of natural
killer cells in cancer. Nat Immun. 12:279–292. 1993.PubMed/NCBI
|
|
57
|
Imai K, Matsuyama S, Miyake S, Suga K and
Nakachi K: Natural cytotoxic activity of peripheral-blood
lymphocytes and cancer incidence: An 11-year follow-up study of a
general population. Lancet. 356:1795–1799. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Schantz SP and Ordonez NG: Quantitation of
natural killer cell function and risk of metastatic poorly
differentiated head and neck cancer. Nat Immun Cell Growth Regul.
10:278–288. 1991.PubMed/NCBI
|
|
59
|
Schantz SP, Savage HE, Racz T, Taylor DL
and Sacks PG: Natural killer cells and metastases from pharyngeal
carcinoma. Am J Surg. 158:361–366. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Tartter PI, Steinberg B, Barron DM and
Martinelli G: The prognostic significance of natural killer
cytotoxicity in patients with colorectal cancer. Arch Surg.
122:1264–1268. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Cong J, Wang X, Zheng X, Wang D, Fu B, Sun
R, Tian Z and Wei H: Dysfunction of natural killer cells by
FBP1-induced inhibition of glycolysis during lung cancer
progression. Cell Metab. 28:243–255.e5. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Takeda K, Nakayama M, Sakaki M, Hayakawa
Y, Imawari M, Ogasawara K, Okumura K and Smyth MJ: IFN-γ production
by lung NK cells is critical for the natural resistance to
pulmonary metastasis of B16 melanoma in mice. J Leukoc Biol.
90:777–785. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Villegas FR, Coca S, Villarrubia VG,
Jiménez R, Chillón MJ, Jareño J, Zuil M and Callol L: Prognostic
significance of tumor infiltrating natural killer cells subset CD57
in patients with squamous cell lung cancer. Lung Cancer. 35:23–28.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Takanami I, Takeuchi K and Giga M: The
prognostic value of natural killer cell infiltration in resected
pulmonary adenocarcinoma. J Thorac Cardiovasc Surg. 121:1058–1063.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Jin S, Deng Y, Hao JW, Li Y, Liu B, Yu Y,
Shi FD and Zhou QH: NK cell phenotypic modulation in lung cancer
environment. PLoS One. 9:e1099762014. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Muntasell A, Rojo F, Servitja S,
Rubio-Perez C, Cabo M, Tamborero D, Costa-García M, Martínez-Garcia
M, Menéndez S, Vazquez I, et al: NK cell infiltrates and HLA class
I expression in primary HER2+ breast cancer predict and
uncouple pathological response and disease-free survival. Clin
Cancer Res. 25:1535–1545. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Eckl J, Buchner A, Prinz PU, Riesenberg R,
Siegert SI, Kammerer R, Nelson PJ and Noessner E: Transcript
signature predicts tissue NK cell content and defines renal cell
carcinoma subgroups independent of TNM staging. J Mol Med (Berl).
90:55–66. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Carrega P, Morandi B, Costa R, Frumento G,
Forte G, Altavilla G, Ratto GB, Mingari MC, Moretta L and Ferlazzo
G: Natural killer cells infiltrating human nonsmall-cell lung
cancer are enriched in CD56 bright CD16(−) cells and display an
impaired capability to kill tumor cells. Cancer. 112:863–875. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Platonova S, Cherfils-Vicini J, Damotte D,
Crozet L, Vieillard V, Validire P, André P, Dieu-Nosjean MC,
Alifano M, Régnard JF, et al: Profound coordinated alterations of
intratumoral NK cell phenotype and function in lung carcinoma.
Cancer Res. 71:5412–5422. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Schleypen JS, Baur N, Kammerer R, Nelson
PJ, Rohrmann K, Gröne EF, Hohenfellner M, Haferkamp A, Pohla H,
Schendel DJ, et al: Cytotoxic markers and frequency predict
functional capacity of natural killer cells infiltrating renal cell
carcinoma. Clin Cancer Res. 12((3 Pt 1)): 718–725. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Melero I, Rouzaut A, Motz GT and Coukos G:
T-cell and NK-cell infiltration into solid tumors: A key limiting
factor for efficacious cancer immunotherapy. Cancer Discov.
4:522–526. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Cantoni C, Huergo-Zapico L, Parodi M,
Pedrazzi M, Mingari MC, Moretta A, Sparatore B, Gonzalez S, Olive
D, Bottino C, et al: NK cells, tumor cell transition, and tumor
progression in solid malignancies: New Hints for NK-based
immunotherapy? J Immunol Res. 2016:46842682016. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Gonzalez-Gugel E, Saxena M and Bhardwaj N:
Modulation of innate immunity in the tumor microenvironment. Cancer
Immunol Immunother. 65:1261–1268. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Lavin Y, Kobayashi S, Leader A, Amir ED,
Elefant N, Bigenwald C, Remark R, Sweeney R, Becker CD, Levine JH,
et al: Innate immune landscape in early lung adenocarcinoma by
paired single-cell analyses. Cell. 169:750–765.e17. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Buckley CD and Simmons DL: Sticky moments
with sticky molecules. Immunol Today. 21:601–603. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Campbell JJ, Qin S, Unutmaz D, Soler D,
Murphy KE, Hodge MR, Wu L and Butcher EC: Unique subpopulations of
CD56+ NK and NK-T peripheral blood lymphocytes identified by
chemokine receptor expression repertoire. J Immunol. 166:6477–6482.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Böhm W, Thoma S, Leithäuser F, Möller P,
Schirmbeck R and Reimann J: T cell-mediated, IFN-gamma-facilitated
rejection of murine B16 melanomas. J Immunol. 161:897–908.
1998.PubMed/NCBI
|
|
78
|
Spear P, Barber A, Rynda-Apple A and
Sentman CL: Chimeric antigen receptor T cells shape myeloid cell
function within the tumor microenvironment through IFN-γ and
GM-CSF. J Immunol. 188:6389–6398. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Cremer I, Fridman WH and Sautès-Fridman C:
Tumor microenvironment in NSCLC suppresses NK cells function.
Oncoimmunology. 1:244–246. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Guillerey C and Smyth MJ: NK cells and
cancer immunoediting. Curr Top Microbiol Immunol. 395:115–145.
2016.PubMed/NCBI
|
|
81
|
Belli C, Trapani D, Viale G, D'Amico P,
Duso BA, Della Vigna P, Orsi F and Curigliano G: Targeting the
microenvironment in solid tumors. Cancer Treat Rev. 65:22–32. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Vitale M, Cantoni C, Pietra G, Mingari MC
and Moretta L: Effect of tumor cells and tumor microenvironment on
NK-cell function. Eur J Immunol. 44:1582–1592. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Fauriat C, Just-Landi S, Mallet F,
Arnoulet C, Sainty D, Olive D and Costello RT: Deficient expression
of NCR in NK cells from acute myeloid leukemia: Evolution during
leukemia treatment and impact of leukemia cells in NCRdull
phenotype induction. Blood. 109:323–330. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Carlsten M, Norell H, Bryceson YT, Poschke
I, Schedvins K, Ljunggren HG, Kiessling R and Malmberg KJ: Primary
human tumor cells expressing CD155 impair tumor targeting by
down-regulating DNAM-1 on NK cells. J Immunol. 183:4921–4930. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Zhang Q, Bi J, Zheng X, Chen Y, Wang H, Wu
W, Wang Z, Wu Q, Peng H, Wei H, et al: Blockade of the checkpoint
receptor TIGIT prevents NK cell exhaustion and elicits potent
anti-tumor immunity. Nat Immunol. 19:723–732. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Castriconi R, Cantoni C, Della Chiesa M,
Vitale M, Marcenaro E, Conte R, Biassoni R, Bottino C, Moretta L
and Moretta A: Transforming growth factor beta 1 inhibits
expression of NKp30 and NKG2D receptors: Consequences for the
NK-mediated killing of dendritic cells. Proc Natl Acad Sci USA.
100:4120–4125. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Marcenaro E, Della Chiesa M, Bellora F,
Parolini S, Millo R, Moretta L and Moretta A: IL-12 or IL-4 prime
human NK cells to mediate functionally divergent interactions with
dendritic cells or tumors. J Immunol. 174:3992–3998. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Pietra G, Manzini C, Rivara S, Vitale M,
Cantoni C, Petretto A, Balsamo M, Conte R, Benelli R, Minghelli S,
et al: Melanoma cells inhibit natural killer cell function by
modulating the expression of activating receptors and cytolytic
activity. Cancer Res. 72:1407–1415. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Massagué J: TGFbeta in cancer. Cell.
134:215–230. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Bruno A, Focaccetti C, Pagani A,
Imperatori AS, Spagnoletti M, Rotolo N, Cantelmo AR, Franzi F,
Capella C, Ferlazzo G, et al: The proangiogenic phenotype of
natural killer cells in patients with non-small cell lung cancer.
Neoplasia. 15:133–142. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Kim TK, Mo EK, Yoo CG, Lee CT, Han SK,
Shim YS and Kim YW: Alteration of cell growth and morphology by
overexpression of transforming growth factor beta type II receptor
in human lung adenocarcinoma cells. Lung Cancer. 31:181–191. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Li J, Shen C, Wang X, Lai Y, Zhou K, Li P,
Liu L and Che G: Prognostic value of TGF-beta in lung cancer:
Systematic review and meta-analysis. BMC Cancer. 19:6912019.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Laouar Y, Sutterwala FS, Gorelik L and
Flavell RA: Transforming growth factor-beta controls T helper type
1 cell development through regulation of natural killer cell
interferon-gamma. Nat Immunol. 6:600–607. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Lee JC, Lee KM, Kim DW and Heo DS:
Elevated TGF-beta1 secretion and down-modulation of NKG2D underlies
impaired NK cytotoxicity in cancer patients. J Immunol.
172:7335–7340. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Kopp HG, Placke T and Salih HR:
Platelet-derived transforming growth factor-beta down-regulates
NKG2D thereby inhibiting natural killer cell antitumor reactivity.
Cancer Res. 69:7775–7783. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Sun C, Xu J, Huang Q, Huang M, Wen H,
Zhang C, Wang J, Song J, Zheng M, Sun H, et al: High NKG2A
expression contributes to NK cell exhaustion and predicts a poor
prognosis of patients with liver cancer. Oncoimmunology.
6:e12645622016. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Beldi-Ferchiou A, Lambert M, Dogniaux S,
Vély F, Vivier E, Olive D, Dupuy S, Levasseur F, Zucman D, Lebbé C,
et al: PD-1 mediates functional exhaustion of activated NK cells in
patients with Kaposi sarcoma. Oncotarget. 7:72961–72977. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Li F, Wei H, Wei H, Gao Y, Xu L, Yin W,
Sun R and Tian Z: Blocking the natural killer cell inhibitory
receptor NKG2A increases activity of human natural killer cells and
clears hepatitis B virus infection in mice. Gastroenterology.
144:392–401. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Bassani B, Baci D, Gallazzi M, Poggi A,
Bruno A and Mortara L: Natural killer cells as key players of tumor
progression and angiogenesis: Old and novel tools to divert their
pro-tumor activities into potent anti-tumor effects. Cancers
(Basel). 11:4612019. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Kim JI, Lakshmikanthan V, Frilot N and
Daaka Y: Prostaglandin E2 promotes lung cancer cell migration via
EP4-betaArrestin1-c-Src signalsome. Mol Cancer Res. 8:569–577.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Balsamo M, Vermi W, Parodi M, Pietra G,
Manzini C, Queirolo P, Lonardi S, Augugliaro R, Moretta A,
Facchetti F, et al: Melanoma cells become resistant to
NK-cell-mediated killing when exposed to NK-cell numbers compatible
with NK-cell infiltration in the tumor. Eur J Immunol.
42:1833–1842. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Li T, Yang Y, Hua X, Wang G, Liu W, Jia C,
Tai Y, Zhang Q and Chen G: Hepatocellular carcinoma-associated
fibroblasts trigger NK cell dysfunction via PGE2 and IDO. Cancer
Lett. 318:154–161. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Li T, Yi S, Liu W, Jia C, Wang G, Hua X,
Tai Y, Zhang Q and Chen G: Colorectal carcinoma-derived fibroblasts
modulate natural killer cell phenotype and antitumor cytotoxicity.
Med Oncol. 30:6632013. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Brandau S, Trellakis S, Bruderek K,
Schmaltz D, Steller G, Elian M, Suttmann H, Schenck M, Welling J,
Zabel P and Lang S: Myeloid-derived suppressor cells in the
peripheral blood of cancer patients contain a subset of immature
neutrophils with impaired migratory properties. J Leukoc Biol.
89:311–317. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Granville CA, Memmott RM, Balogh A,
Mariotti J, Kawabata S, Han W, Lopiccolo J, Foley J, Liewehr DJ,
Steinberg SM, et al: A central role for Foxp3+ regulatory T cells
in K-Ras-driven lung tumorigenesis. PLoS One. 4:e50612009.
View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Smyth MJ, Teng MW, Swann J, Kyparissoudis
K, Godfrey DI and Hayakawa Y: CD4+CD25+ T regulatory cells suppress
NK cell-mediated immunotherapy of cancer. J Immunol. 176:1582–1587.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Ghiringhelli F, Ménard C, Terme M, Flament
C, Taieb J, Chaput N, Puig PE, Novault S, Escudier B, Vivier E, et
al: CD4+CD25+ regulatory T cells inhibit natural killer cell
functions in a transforming growth factor-beta-dependent manner. J
Exp Med. 202:1075–1085. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Lardner A: The effects of extracellular pH
on immune function. J Leukoc Biol. 69:522–530. 2001.PubMed/NCBI
|
|
109
|
Balsamo M, Manzini C, Pietra G, Raggi F,
Blengio F, Mingari MC, Varesio L, Moretta L, Bosco MC and Vitale M:
Hypoxia downregulates the expression of activating receptors
involved in NK-cell-mediated target cell killing without affecting
ADCC. Eur J Immunol. 43:2756–2764. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Sarkar S, Germeraad WT, Rouschop KM,
Steeghs EM, van Gelder M, Bos GM and Wieten L: Hypoxia induced
impairment of NK cell cytotoxicity against multiple myeloma can be
overcome by IL-2 activation of the NK cells. PLoS One.
8:e648352013. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Li C, Lu HJ, Na FF, Deng L, Xue JX, Wang
JW, Wang YQ, Li QL and Lu Y: Prognostic role of hypoxic inducible
factor expression in non-small cell lung cancer: A meta-analysis.
Asian Pac J Cancer Prev. 14:3607–3612. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Yang SL, Ren QG, Wen L and Hu JL:
Clinicopathological and prognostic significance of
hypoxia-inducible factor-1 alpha in lung cancer: A systematic
review with meta-analysis. J Huazhong Univ Sci Technolog Med Sci.
36:321–327. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Vaupel P and Multhoff G: Adenosine can
thwart antitumor immune responses elicited by radiotherapy:
Therapeutic strategies alleviating protumor ADO activities.
Strahlenther Onkol. 192:279–287. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Pötzl J, Roser D, Bankel L, Hömberg N,
Geishauser A, Brenner CD, Weigand M, Röcken M and Mocikat R:
Reversal of tumor acidosis by systemic buffering reactivates NK
cells to express IFN-γ and induces NK cell-dependent lymphoma
control without other immunotherapies. Int J Cancer. 140:2125–2133.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Husain Z, Huang Y, Seth P and Sukhatme VP:
Tumor-derived lactate modifies antitumor immune response: Effect on
myeloid-derived suppressor cells and NK cells. J Immunol.
191:1486–1495. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Sautès-Fridman C, Cherfils-Vicini J,
Damotte D, Fisson S, Fridman WH, Cremer I and Dieu-Nosjean MC:
Tumor microenvironment is multifaceted. Cancer Metastasis Rev.
30:13–25. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Mailloux AW and Young MR: NK-dependent
increases in CCL22 secretion selectively recruits regulatory T
cells to the tumor microenvironment. J Immunol. 182:2753–2765.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Srivastava S, Pelloso D, Feng H, Voiles L,
Lewis D, Haskova Z, Whitacre M, Trulli S, Chen YJ, Toso J, et al:
Effects of interleukin-18 on natural killer cells: Costimulation of
activation through Fc receptors for immunoglobulin. Cancer Immunol
Immunother. 62:1073–1082. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Mi D, Ren W and Yang K: Adoptive
immunotherapy with interleukin-2 & induced killer cells in
non-small cell lung cancer: A systematic review &
meta-analysis. Indian J Med Res. 143:S1–S10. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Davis ZB, Felices M, Verneris MR and
Miller JS: Natural killer cell adoptive transfer therapy:
Exploiting the first line of defense against cancer. Cancer J.
21:486–491. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Ito S, Bollard CM, Carlsten M, Melenhorst
JJ, Biancotto A, Wang E, Chen J, Kotliarov Y, Cheung F, Xie Z, et
al: Ultra-low dose interleukin-2 promotes immune-modulating
function of regulatory T cells and natural killer cells in healthy
volunteers. Mol Ther. 22:1388–1395. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Waldmann TA: The biology of interleukin-2
and interleukin-15: Implications for cancer therapy and vaccine
design. Nat Rev Immunol. 6:595–601. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Miller JS, Morishima C, McNeel DG, Patel
MR, Kohrt HEK, Thompson JA, Sondel PM, Wakelee HA, Disis ML, Kaiser
JC, et al: A first-in-human phase I study of subcutaneous
outpatient recombinant human IL15 (rhIL15) in adults with advanced
solid tumors. Clin Cancer Res. 24:1525–1535. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Wong HC, Jeng EK and Rhode PR: The
IL-15-based superagonist ALT-803 promotes the antigen-independent
conversion of memory CD8+ T cells into innate-like
effector cells with antitumor activity. Oncoimmunology.
2:e264422013. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Wrangle JM, Velcheti V, Patel MR,
Garrett-Mayer E, Hill EG, Ravenel JG, Miller JS, Farhad M, Anderton
K, Lindsey K, et al: ALT-803, an IL-15 superagonist, in combination
with nivolumab in patients with metastatic non-small cell lung
cancer: A non-randomised, open-label, phase 1b trial. Lancet Oncol.
19:694–704. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Bowles JA, Wang SY, Link BK, Allan B,
Beuerlein G, Campbell MA, Marquis D, Ondek B, Wooldridge JE, Smith
BJ, et al: Anti-CD20 monoclonal antibody with enhanced affinity for
CD16 activates NK cells at lower concentrations and more
effectively than rituximab. Blood. 108:2648–2654. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Decaup E, Rossi C, Gravelle P, Laurent C,
Bordenave J, Tosolini M, Tourette A, Perrial E, Dumontet C, Poupot
M, et al: A tridimensional model for NK cell-mediated ADCC of
follicular lymphoma. Front Immunol. 10:19432019. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Mallmann-Gottschalk N, Sax Y, Kimmig R,
Lang S and Brandau S: EGFR-specific tyrosine kinase inhibitor
modifies NK cell-mediated antitumoral activity against ovarian
cancer cells. Int J Mol Sci. 20:46932019. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
André P, Denis C, Soulas C,
Bourbon-Caillet C, Lopez J, Arnoux T, Bléry M, Bonnafous C,
Gauthier L, Morel A, et al: Anti-NKG2A mAb Is a checkpoint
inhibitor that promotes anti-tumor immunity by unleashing both T
and NK cells. Cell. 175:1731–1743.e13. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Lin M, Luo H, Liang S, Chen J, Liu A, Niu
L and Jiang Y: Pembrolizumab plus allogeneic NK cells in advanced
non-small cell lung cancer patients. J Clin Invest. 130:2560–2569.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Youn JI, Park SM, Park S, Kim G, Lee HJ,
Son J, Hong MH, Ghaderpour A, Baik B, Islam J, et al: Peripheral
natural killer cells and myeloid-derived suppressor cells correlate
with anti-PD-1 responses in non-small cell lung cancer. Sci Rep.
10:90502020. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Trefny MP, Kaiser M, Stanczak MA, Herzig
P, Savic S, Wiese M, Lardinois D, Läubli H, Uhlenbrock F and
Zippelius A: PD-1+ natural killer cells in human
non-small cell lung cancer can be activated by PD-1/PD-L1 blockade.
Cancer Immunol Immunother. 69:1505–1517. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Paz-Ares L, Luft A, Vicente D, Tafreshi A,
Gümüş M, Mazières J, Hermes B, Çay Şenler F, Csőszi T, Fülöp A, et
al: Pembrolizumab plus chemotherapy for squamous non-small-cell
lung cancer. N Engl J Med. 379:2040–2051. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Mok TSK, Wu YL, Kudaba I, Kowalski DM, Cho
BC, Turna HZ, Castro G Jr, Srimuninnimit V, Laktionov KK,
Bondarenko I, et al: Pembrolizumab versus chemotherapy for
previously untreated, PD-L1-expressing, locally advanced or
metastatic non-small-cell lung cancer (KEYNOTE-042): A randomised,
open-label, controlled, phase 3 trial. Lancet. 393:1819–1830. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Lisberg A, Cummings A, Goldman JW,
Bornazyan K, Reese N, Wang T, Coluzzi P, Ledezma B, Mendenhall M,
Hunt J, et al: A phase II study of pembrolizumab in EGFR-Mutant,
PD-L1+, tyrosine kinase inhibitor naïve patients with advanced
NSCLC. J Thorac Oncol. 13:1138–1145. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Chabannon C, Mfarrej B, Guia S, Ugolini S,
Devillier R, Blaise D, Vivier E and Calmels B: Manufacturing
natural killer cells as medicinal products. Front Immunol.
7:5042016. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Carotta S: Targeting NK cells for
anticancer immunotherapy: Clinical and preclinical approaches.
Front Immunol. 7:1522016. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Sakamoto N, Ishikawa T, Kokura S, Okayama
T, Oka K, Ideno M, Sakai F, Kato A, Tanabe M, Enoki T, et al: Phase
I clinical trial of autologous NK cell therapy using novel
expansion method in patients with advanced digestive cancer. J
Transl Med. 13:2772015. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Parkhurst MR, Riley JP, Dudley ME and
Rosenberg SA: Adoptive transfer of autologous natural killer cells
leads to high levels of circulating natural killer cells but does
not mediate tumor regression. Clin Cancer Res. 17:6287–6297. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Benson DM Jr, Bakan CE, Zhang S, Collins
SM, Liang J, Srivastava S, Hofmeister CC, Efebera Y, Andre P,
Romagne F, et al: IPH2101, a novel anti-inhibitory KIR antibody,
and lenalidomide combine to enhance the natural killer cell versus
multiple myeloma effect. Blood. 118:6387–6391. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Ruggeri L, Capanni M, Urbani E, Perruccio
K, Shlomchik WD, Tosti A, Posati S, Rogaia D, Frassoni F, Aversa F,
et al: Effectiveness of donor natural killer cell alloreactivity in
mismatched hematopoietic transplants. Science. 295:2097–2100. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Calvo T, Reina-Ortiz C, Giraldos D, Gascón
M, Woods D, Asenjo J, Marco-Brualla J, Azaceta G, Izquierdo I,
Palomera L, et al: Expanded and activated allogeneic NK cells are
cytotoxic against B-chronic lymphocytic leukemia (B-CLL) cells with
sporadic cases of resistance. Sci Rep. 10:193982020. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Miller JS, Soignier Y,
Panoskaltsis-Mortari A, McNearney SA, Yun GH, Fautsch SK, McKenna
D, Le C, Defor TE, Burns LJ, et al: Successful adoptive transfer
and in vivo expansion of human haploidentical NK cells in patients
with cancer. Blood. 105:3051–3057. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Cheng M, Chen Y, Xiao W, Sun R and Tian Z:
NK cell-based immunotherapy for malignant diseases. Cell Mol
Immunol. 10:230–252. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Jochems C, Hodge JW, Fantini M, Fujii R,
Morillon YM II, Greiner JW, Padget MR, Tritsch SR, Tsang KY,
Campbell KS, et al: An NK cell line (haNK) expressing high levels
of granzyme and engineered to express the high affinity CD16
allele. Oncotarget. 7:86359–86373. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Park HR, Ahn YO, Kim TM, Kim S, Kim S, Lee
YS, Kim M, Keam B, Kim DW and Heo DS: NK92-CD16 cells are cytotoxic
to non-small cell lung cancer cell lines that have acquired
resistance to tyrosine kinase inhibitors. Cytotherapy. 21:603–611.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Iliopoulou EG, Kountourakis P, Karamouzis
MV, Doufexis D, Ardavanis A, Baxevanis CN, Rigatos G, Papamichail M
and Perez SA: A phase I trial of adoptive transfer of allogeneic
natural killer cells in patients with advanced non-small cell lung
cancer. Cancer Immunol Immunother. 59:1781–1789. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Wu C: Heat shock transcription factors:
Structure and regulation. Annu Rev Cell Dev Biol. 11:441–469. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Kregel KC: Heat shock proteins: Modifying
factors in physiological stress responses and acquired
thermotolerance. J Appl Physiol (1985). 92:2177–2186. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Guzhova IV and Margulis BA: HSP70-based
anti-cancer immunotherapy. Hum Vaccin Immunother. 12:2529–2535.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Lazarev VF, Nikotina AD, Mikhaylova ER,
Nudler E, Polonik SG, Guzhova IV and Margulis BA: Hsp70 chaperone
rescues C6 rat glioblastoma cells from oxidative stress by
sequestration of aggregating GAPDH. Biochem Biophys Res Commun.
470:766–771. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Calderwood SK and Gong J: Heat shock
proteins promote cancer: It's a protection racket. Trends Biochem
Sci. 41:311–323. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Mittal S and Rajala MS: Heat shock
proteins as biomarkers of lung cancer. Cancer Biol Ther.
21:477–485. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Jin HO, Hong SE, Kim JY, Kim MR, Chang YH,
Hong YJ, Lee JK and Park IC: Induction of HSP27 and HSP70 by
constitutive overexpression of Redd1 confers resistance of lung
cancer cells to ionizing radiation. Oncol Rep. 41:3119–3126.
2019.PubMed/NCBI
|
|
155
|
Hromadnikova I, Pirkova P and Sedlackova
L: Influence of in vitro IL-2 or IL-15 alone or in combination with
Hsp-70-derived 14-mer peptide (TKD) on the expression of NK cell
activatory and inhibitory receptors. Mediators Inflamm.
2013:4052952013. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Multhoff G, Seier S, Stangl S, Sievert W,
Shevtsov M, Werner C, Pockley AG, Blankenstein C, Hildebrandt M,
Offner R, et al: Targeted natural killer cell-based adoptive
immunotherapy for the treatment of patients with NSCLC after
radiochemotherapy: A randomized phase II clinical trial. Clin
Cancer Res. 26:5368–5379. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Specht HM, Ahrens N, Blankenstein C, Duell
T, Fietkau R, Gaipl US, Günther C, Gunther S, Habl G, Hautmann H,
et al: Heat shock protein 70 (Hsp70) peptide activated natural
killer (NK) cells for the treatment of patients with non-small cell
lung cancer (NSCLC) after radiochemotherapy (RCTx)-from preclinical
studies to a clinical phase II trial. Front Immunol. 6:1622015.
View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Shevtsov M, Pitkin E, Ischenko A, Stangl
S, Khachatryan W, Galibin O, Edmond S, Lobinger D and Multhoff G:
Ex vivo Hsp70-activated nk cells in combination with PD-1
inhibition significantly increase overall survival in preclinical
models of glioblastoma and lung cancer. Front Immunol. 10:4542019.
View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Daher M and Rezvani K: Next generation
natural killer cells for cancer immunotherapy: The promise of
genetic engineering. Curr Opin Immunol. 51:146–153. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Xie G, Dong H, Liang Y, Ham JD, Rizwan R
and Chen J: CAR-NK cells: A promising cellular immunotherapy for
cancer. EBioMedicine. 59:1029752020. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Wang W, Jiang J and Wu C: CAR-NK for tumor
immunotherapy: Clinical transformation and future prospects. Cancer
Lett. 472:175–180. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Hu Y, Tian ZG and Zhang C: Chimeric
antigen receptor (CAR)-transduced natural killer cells in tumor
immunotherapy. Acta Pharmacol Sin. 39:167–176. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Oei VYS, Siernicka M, Graczyk-Jarzynka A,
Hoel HJ, Yang W, Palacios D, Almåsbak H, Bajor M, Clement D, Brandt
L, et al: Intrinsic functional potential of NK-cell subsets
constrains retargeting driven by chimeric antigen receptors. Cancer
Immunol Res. 6:467–480. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Liu E, Tong Y, Dotti G, Shaim H, Savoldo
B, Mukherjee M, Orange J, Wan X, Lu X, Reynolds A, et al: Cord
blood NK cells engineered to express IL-15 and a CD19-targeted CAR
show long-term persistence and potent antitumor activity. Leukemia.
32:520–531. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Liu E, Marin D, Banerjee P, Macapinlac HA,
Thompson P, Basar R, Nassif Kerbauy L, Overman B, Thall P, Kaplan
M, et al: Use of CAR-transduced natural killer cells in
CD19-positive lymphoid tumors. N Engl J Med. 382:545–553. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Gunesch JT, Angelo LS, Mahapatra S,
Deering RP, Kowalko JE, Sleiman P, Tobias JW, Monaco-Shawver L,
Orange JS and Mace EM: Genome-wide analyses and functional
profiling of human NK cell lines. Mol Immunol. 115:64–75. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Liu Q, Xu Y, Mou J, Tang K, Fu X, Li Y,
Xing Y, Rao Q, Xing H, Tian Z, et al: Irradiated chimeric antigen
receptor engineered NK-92MI cells show effective cytotoxicity
against CD19+ malignancy in a mouse model. Cytotherapy.
22:552–562. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Yang S, Cao B, Zhou G, Zhu L, Wang L,
Zhang L, Kwok HF, Zhang Z and Zhao Q: Targeting B7-H3 immune
checkpoint with chimeric antigen receptor-engineered natural killer
cells exhibits potent cytotoxicity against non-small cell lung
cancer. Front Pharmacol. 11:10892020. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Tonn T, Schwabe D, Klingemann HG, Becker
S, Esser R, Koehl U, Suttorp M, Seifried E, Ottmann OG and Bug G:
Treatment of patients with advanced cancer with the natural killer
cell line NK-92. Cytotherapy. 15:1563–1570. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Lu C, Guo C, Chen H, Zhang H, Zhi L, Lv T,
Li M, Niu Z, Lu P and Zhu W: A novel chimeric PD1-NKG2D-41BB
receptor enhances antitumor activity of NK92 cells against human
lung cancer H1299 cells by triggering pyroptosis. Mol Immunol.
122:200–206. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Shah NN, Baird K, Delbrook CP, Fleisher
TA, Kohler ME, Rampertaap S, Lemberg K, Hurley CK, Kleiner DE,
Merchant MS, et al: Acute GVHD in patients receiving IL-15/4-1BBL
activated NK cells following T-cell-depleted stem cell
transplantation. Blood. 125:784–792. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Klingemann H: Challenges of cancer therapy
with natural killer cells. Cytotherapy. 17:245–249. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Gauthier L, Morel A, Anceriz N, Rossi B,
Blanchard-Alvarez A, Grondin G, Trichard S, Cesari C, Sapet M,
Bosco F, et al: Multifunctional natural killer cell engagers
targeting NKp46 trigger protective tumor immunity. Cell.
177:1701–1713.e16. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Hodgins JJ, Khan ST, Park MM, Auer RC and
Ardolino M: Killers 2.0: NK cell therapies at the forefront of
cancer control. J Clin Invest. 129:3499–3510. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Yao C, Ni Z, Gong C, Zhu X, Wang L, Xu Z,
Zhou C, Li S, Zhou W, Zou C and Zhu S: Rocaglamide enhances NK
cell-mediated killing of non-small cell lung cancer cells by
inhibiting autophagy. Autophagy. 14:1831–1844. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Lin M, Liang SZ, Wang XH, Liang YQ, Zhang
MJ, Niu LZ, Chen JB, Li HB and Xu KC: Clinical efficacy of
percutaneous cryoablation combined with allogenic NK cell
immunotherapy for advanced non-small cell lung cancer. Immunol Res.
65:880–887. 2017. View Article : Google Scholar : PubMed/NCBI
|