Introduction
Pancreatic cancer is one of deadliest cancers
(1). According to GLOBOCAN 2020
estimates, there were 495,773 new cases of pancreatic cancer and
466,003 deaths in 2020 (2). Due to
insidious onset, special anatomical position, low resection rate
and high recurrence rate, the 5-year survival rate of patients is
<8% (3–5). Therefore, it is imperative to clarify
the mechanism of pancreatic cancer progression in order to identify
potential therapeutic targets and thus improve the treatment
efficacy and survival rate of the patients.
It is reported that, 80% of DNA sequences could be
transcribed into RNA, yet merely <2% of RNAs are translated into
proteins; RNAs which have no protein-coding function are defined as
non-coding RNAs (ncRNAs) (6). Long
non-coding RNAs (lncRNAs) belong to ncRNAs, with a transcript
longer than 200 nucleotides (7).
Previously, lncRNAs were considered as ‘junk’. However, in previous
years, growing studies have indicated that lncRNAs have crucial
functions in biological processes (8,9).
Since there are numerous lncRNAs specifically expressed or
dysregulated in multiple cancers, lncRNAs are regarded as promising
diagnostic biomarkers and treatment targets for cancer. LncRNA FGD5
antisense RNA 1 (FGD5-AS1) has been reported to be involved in the
pathogenesis of acute myocardial infarction and periodontitis
(10,11). Additionally, FGD5-AS1 is involved
in the progression of clear renal cell carcinoma, colorectal
cancer, and non-small cell lung carcinoma (12–15).
Nonetheless, the role of FGD5-AS1 in pancreatic cancer has not been
clarified.
Recognized as a highly conserved signaling pathway,
Wnt/β-catenin is crucial in the development, organogenesis, tissue
regeneration and other physiological or pathological processes
(16). In cancer biology, the
activation of the Wnt/β-catenin signaling is pivotal in regulating
the sustaining proliferation, metastasis and chemoresistance of
cancer cells (17). In the present
study, the expression characteristics, biological functions and
mechanism of FGD5-AS1 in pancreatic cancer were explored. It was
revealed that FGD5-AS1 was highly expressed in pancreatic cancer
tissues, and its overexpression facilitated the activation of the
Wnt/β-catenin pathway and cancer progression via suppression of
miR-577. The present study identified FGD5-AS1 as a novel
carcinogenic lncRNA in pancreatic cancer, and partly explained the
mechanism of the Wnt/β-catenin pathway dysregulation in pancreatic
cancer.
Materials and methods
Clinical samples and ethical
statement
The collection and use of human samples was carried
out strictly following The Declaration of Helsinki (7th revision,
2008). The present study obtained approval from the Ethics
Committee of the Sixth Hospital of Shanxi Medical University
(approval no. 201310012; Taiyuan, China). The pancreatic cancer
tissue and adjacent non-tumorous tissue samples were collected from
37 patients (21–78 years old; 21 males and 16 females) who had
undergone surgery at the Sixth Hospital of Shanxi Medical
University from January 2014 to June 2018. Patients who had no
other digestive system tumors or tumor history, had never received
any chemotherapy or radiotherapy, and were confirmed as pancreatic
cancer by pathology and genetics were included. Patients with
serious injury to the heart, liver, kidney and other important
organs, history of autoimmune diseases, and chronic or acute
infectious diseases were excluded. All of the patients had provided
a written informed consent prior to inclusion in the study.
Bioinformatics analysis
The expression pattern of FGD5-AS1 in pancreatic
cancer tissues and normal tissues was predicted using the Gene
Expression Profiling Interactive Analysis (GEPIA) database
(http://gepia.cancer-pku.cn/). The
potential downstream target miRNAs of FGD5-AS1 were predicted by
the starBase database (http://starbase.sysu.edu.cn/panCancer.php) and LncBase
Predicted v.2 database (http://carolina.imis.athena-innovation.gr) (18).
Cell culture
Normal pancreatic ductal epithelial cells (HPDE6-C7
cells) and human pancreatic cancer cell lines (PANC-1, BXPC-3,
CAPAN-1 and SW1990) were obtained from Shanghai Institute of
Biochemistry and Cell Biology (Shanghai, China). Cells were
cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen;
Thermo Fisher Scientific, Inc.) containing 0.1 mg/ml streptomycin,
100 U/ml penicillin and 10% fetal bovine serum (FBS; all from
Hyclone; Cytiva) in an incubator at 37°C in 5% CO2.
Cell transfection
Control siRNA (si-NC), FGD5-AS1 siRNA (si-FGD5-AS1),
miR-577 mimics (miR-577), miR-577 inhibitors (miR-577 in), microRNA
mimics control (miR-con), and microRNA inhibitor control (miR-con
in) were obtained from Shanghai GenePharma Co., Ltd. Cells were
cultured in 6-well plates, and when cell confluence reached 80–90%,
the pancreatic cancer cells were cultured in the fresh serum-free
and antibiotic-free medium for 8 h. Subsequently, 100 nM siRNAs or
50 nM miRNAs were transfected with pancreatic cancer cells using
Lipofectamine® 2000 (Invitrogen, Thermo Fisher
Scientific, Inc.) at room temperature according to the supplier's
protocol. A total of 12 h following transfection, the medium was
replaced by complete medium, and the cell culture was continued for
24 h. Subsequently, the transfection efficiency was evaluated by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR). The sequences of the oligonucleotides were as follows:
FGD5-AS1 siRNA1: 5′-UUGGUCGUUGUCAACUUCCCA-3′; FGD5-AS1 siRNA2:
5′-UAUUGUAUGAAUACACUGCUA-3′; si-NC: 5′-UUCUCCGAACGUGUCACGUTT-3′;
miR-577 mimics: 3′-UAGAUAAAAUAUUGGUACCUG-5′; microRNA mimics
control: 5′- UAAGUAGUCUGAAAUAGUUAC-3′; miR-577 inhibitors:
3′-CAGGUACCAAUAUUUUAUCUA-5′. microRNA inhibitor control:
5′-AUCAGUUCAAUCAUGUAUCAU-3′.
RT-qPCR
Total RNA was extracted using TRIzol reagent
(Invitrogen, Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol. To remove the DNA, 2 µg of RNA was treated
with DNase and then reverse-transcribed into cDNA using SuperScript
First Strand cDNA System (Invitrogen; Thermo Fisher Scientific,
Inc.) according to the manufacturer's protocol. Subsequently, with
cDNA as the template, RT-qPCR was performed using an ABI 7500 Fast
Real-Time PCR System with SYBR® PremixExTaq™ kit (Takara
Biotechnology Co., Ltd.). The qPCR thermocycling conditions were as
follows: pre-denaturation at 95°C for 10 min; followed by 40 cycles
of denaturation at 95°C for 15 sec, and annealing and extension at
60°C for 60 sec. The primers were designed and synthesized by BGI
(Shenzhen, China). The expression levels of FGD5-AS1, miR-577,
low-density lipoprotein receptor-related protein 6 (LRP6) and
β-catenin were determined with 2−∆∆Cq method (19). The levels of U6 were used to
normalize the miR-577 expression, and the levels of
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were used to
normalize the expression levels of lncRNA and mRNA. The sequences
of the primers are listed in Table
I.
 | Table I.Primer sequences for RT-qPCR. |
Table I.
Primer sequences for RT-qPCR.
| Genes | Primer sequences
(5′-3′) |
|---|
| FGD5-AS1 | F:
AGAAGCGGAGGGGTGAAAAT |
|
| R:
CCGCCTTATAGTTGGCCCTC |
| LRP6 | F:
AGGCACTTACTTCCCTGCAA |
|
| R:
GGGCACAGGTTCTGAATCAT |
| β-catenin | F:
GACATCAACGTGGTGACCTG |
|
| R:
GCTGGCTCTGTGATTTCCTC |
| GAPDH | F:
GGAGCGAGATCCCTCCAAAAT |
|
| R:
GGCTGTTGTCATACTTCTCATGG |
| miR-577 | F:
TGCGGTAGATAAAATATTGG |
|
| R:
CCAGTGCAGGGTCCGAGGT |
| U6 | F:
GCTCGCTTCGGCAGCACA |
|
| R:
GAGGTATTCGCACCAGAGGA |
MTT assay
An MTT assay was utilized to determine cell
proliferation rates. Cells in each group were transferred into
96-well plates (5×104 cells/well), and routinely
cultured. Following 12, 24, 48 and 72 h of culture, the cells were
incubated with MTT reagent (Beyotime Institute of Biotechnology)
for 4 h at 37°C. Subsequently, the formazan crystals were dissolved
in dimethyl sulfoxide. Finally, a spectrophotometer was utilized to
measure the absorbance of the cells at 570 nm.
Transwell assay
In the migration assay, SW1990 cells were harvested
and resuspended with serum-free medium, and then 200 µl of cell
suspension (containing ~5×104 cells) was added into the
upper compartment of each Transwell chamber (8-µm pore size; BD
Biosciences) in 24-well plates, and complete DMEM (containing
glucose, L-glutamine, and sodium bicarbonate) with 10% FBS (600 µl
per well) were added in the lower compartment. The cells were
cultured for 24 h at 37°C, and then the cells on the upper surface
of the membrane were gently wiped off with cotton swabs. The cells
on the lower surface were fixed in 10% formalin for 20 min at room
temperature and then stained with 0.1% crystal violet solution for
20 min at room temperature. Subsequently, the cells were counted
under a light microscope. In the invasion assay, the procedures
were the same as the migration assay using Transwell chambers
covered with a layer of Matrigel (BD Biosciences) at 4°C
overnight.
Dual-luciferase reporter gene
assay
To determine the binding association between
FGD5-AS1 and miR-577, the wild-type (WT) or mutant (MUT) FGD5-AS1
sequence was subcloned into the psi-CHECK2 reporter vector (Promega
Corporation). The cells were subsequently transferred to 24-well
plates. A total of 24 h later, miR-577 mimics or miR-NC were
co-transfected into SW1990 cells with WT or MUT FGD5-AS1 reporter
vectors, respectively, using Lipofectamine 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.). Luciferase activity was determined 48 h
following transfection. Luciferase activity was determined by a
Dual-Luciferase® Reporter Assay system (Promega
Corporation). Renilla luciferase activity was used as a
control for firefly luciferase activity. Similarly, the targeting
relationship between miR-577 and LRP6 (or β-catenin) was
determined. To determine the activity of Wnt/β-catenin pathway, the
TOPflash and FOPflash system (Upstate Biotechnology, Inc.) was
used. This system was used to evaluate β-catenin-dependent
signaling which drives the expression of T-cell factor (TCF).
TOPflash is a TCF reporter plasmid containing WT TCF binding sites
driven by the thymidine kinase minimal prompter and an upstream
luciferase reporter gene. FOPflash contains mutated TCF binding
sites driven by the same thymidine kinase promoter and upstream
luciferase reporter gene. FOPflash was used as a control for
TOPflash activity.
RNA immunoprecipitation (RIP)
assay
The interaction between FGD5-AS1 and miR-577 was
analyzed using the EZ-Magna RNA binding protein immunoprecipitation
kit (cat. no. 17–700; EMD Millipore). Briefly, following
transfection, 1×106 SW1990 cells were lysed in the RIP
lysis buffer (included in the kit), and then 100 µl of cell lysate
was mixed with 50 µl of magnetic beads coupled with 5 µg of
anti-Ago2 antibody (cat. no. MABE253; EMD Millipore) or anti-IgG
antibody (cat. no. AP101; EMD Millipore) in RIP buffer. Following
incubation at 4°C for 8 h, the sample was centrifuged at 12,000 × g
for 30 sec at 4°C, and the supernatant was discarded, and then the
immunoprecipitate was obtained and incubated at 55°C for 30 min
with proteinase K (EMD Millipore) to remove the protein. The RNA
was isolated using TRIzol reagent according to the manufacturer's
protocol. Subsequently, the RNA was reversely transcribed into cDNA
(the procedure was the same as aforementioned in RT-qPCR section).
Finally, the level of FGD5-AS1 in the immunoprecipitate was
analyzed by RT-qPCR.
RNA pull-down assay
Biotin-labeled miR-577 (Bio-miR-577) and the control
miRNAs (Bio-miR-NC) were incubated with 500 µl cell lysates at 4°C
overnight. Then, streptavidin-coated 400 µl of magnetic beads
(Thermo Fisher Scientific, Inc.) were added in the cell lysates,
and incubated at 4°C overnight, to enrich the complex containing
the biotin. Next, the complex was eluted, and then the RNA in the
complex was extracted using TRIzol reagent as aforementioned.
Subsequently, the FGD5-AS1 enrichment in the complex was evaluated
via qPCR.
Western blot assay
Cells were lysed in RIPA buffer (Pierce; Thermo
Fisher Scientific, Inc.) containing protease inhibitors. Then the
lysates were centrifuged at 12,000 × g for 15 min at 4°C and the
supernatant was collected. The supernatant was added with loading
buffer and boiled, and the protein samples were prepared. Next, 10%
sodium dodecyl sulfate-polyacrylamide gel electrophoresis was
employed to separate the protein samples (20 µg per lane), and then
the proteins were electrotransferred onto polyvinylidene fluoride
(PVDF) membranes (EMD Millipore). Subsequently, the proteins were
blocked with 5% skimmed milk for 2 h at room temperature, and then
incubated overnight with primary antibodies anti-Axin2 (product
code ab109307; 1:1,000), anti-c-Myc (product code ab32072;
1:1,000), anti-cyclin D1 (product code ab16663; 1:1,000), β-actin
(product code ab8227; 1:1,000; all from Abcam) at 4°C. After being
rinsed three times with Tris-buffered saline and 0.05 % Tween-20
(TBST), the proteins were incubated with horseradish
peroxidase-coupled secondary antibodies goat anti-rabbit IgG
H&L (HRP) (ab205718; 1:5,000; Abcam) at room temperature for 30
min. Then, TBST was employed again, to wash the membranes 3 times.
Ultimately, a hypersensitive ECL kit (Beyotime Institute of
Biotechnology) was utilized to develop the protein bands on an
X-ray film.
Lung metastasis assay in vivo
A total of 20 nude mice (male; 4 weeks old; weighing
~16 g) were obtained commercially from the National Laboratory
Animal Center (Beijing, China). Mice were housed and maintained at
~20°C in a relative humidity of 40–70% with a 12-h light/dark cycle
and received food and water ad libitum. All the mice were
randomly divided into two groups (si-NC group vs. si-FGD5-AS1
group, 10 mice per group). SW1990 cells were transfected with si-NC
or FGD5-AS1 siRNA1. Then the cells were suspended in sterile
phosphate-buffered saline and injected into the tail vein of each
4-week-old mouse (1×107 cells/mouse) (20). The animal health and behaviour were
monitored weekly. A total of 3 weeks later, all 20 mice were
anesthetized by intraperitoneal injection with 10% chloral hydrate
(300 mg/kg), and then sacrificed by decapitation. The lung tissues
were fixed in 4% paraformaldehyde at 4°C for 24 h and embedded in
paraffin; after which, 4-µm sections were cut and stained with
hematoxylin for 5 min and eosin for 2 min at room temperature.
Metastatic nodules in the lungs were evaluated by pathological
examination under a light microscope (magnification, ×200). No
mouse exhibited signs of peritonitis, pain or discomfort following
the administration of 10% chloral hydrate. The animal experiments
were approved (approval no. 8217113963) by the Institutional Animal
Care and Use Committee of Chinese PLA General Hospital (Beijing,
China).
Statistical analysis
Data analysis was carried out using the SPSS 20.0
software (IBM Corp.). The data was presented as the mean ± standard
deviation (SD). Pearson's correlation analysis was used to
determine the correlation between the expression levels of FGD5-AS1
and miR-577 in pancreatic cancer tissues. The Kolmogorov-Smirnov
test was used for normality and equal variance of the data.
Unpaired Student's t-tests were utilized to perform comparisons.
For data that had skewed distribution, comparisons between two
groups were performed by Wilcoxon signed-rank test. P<0.05 was
considered to indicate a statistically significant difference.
Results
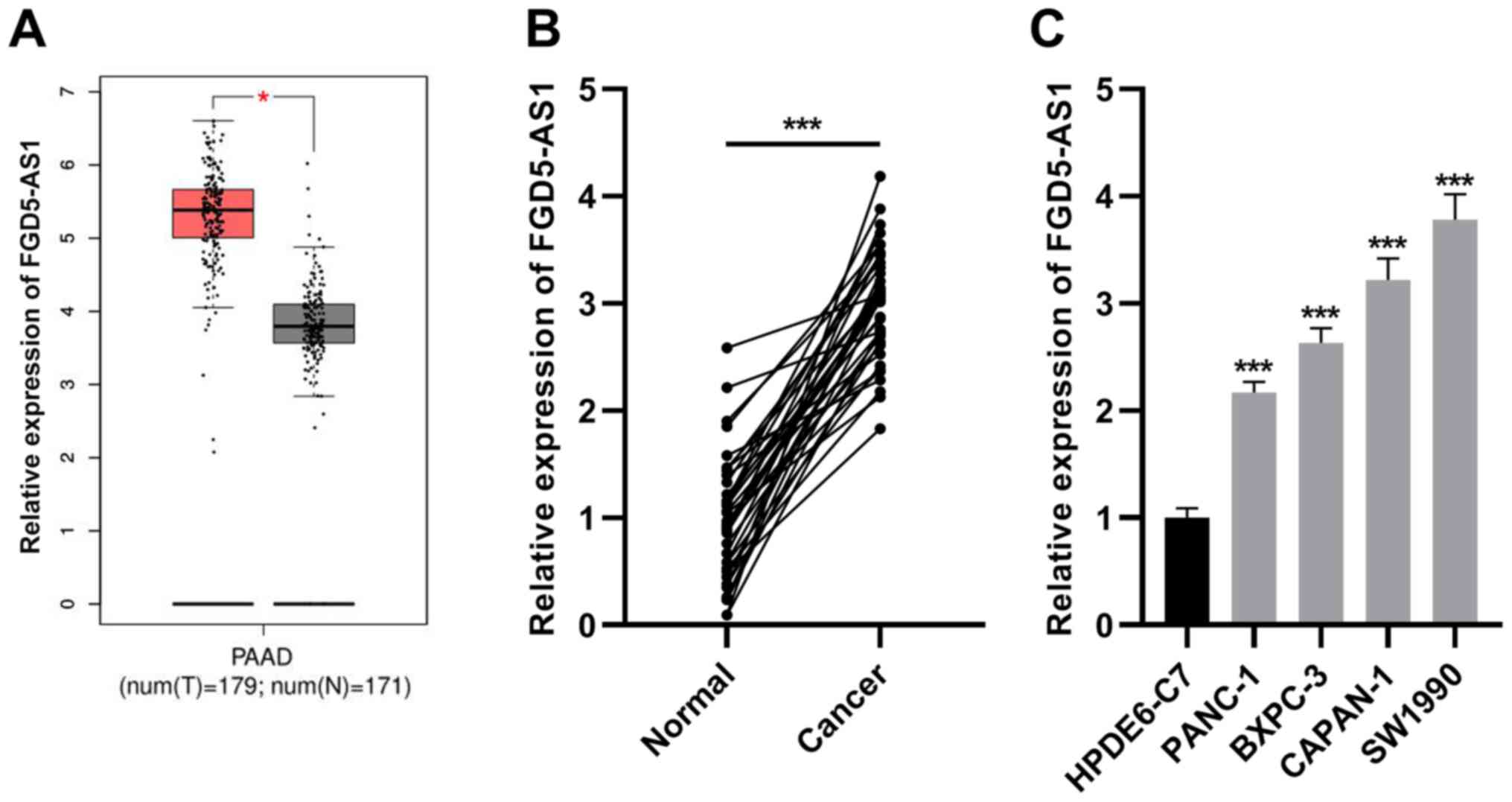
FGD5-AS1 expression is increased in
pancreatic cancer
GEPIA database (http://gepia.cancer-pku.cn/) revealed that FGD5-AS1
was differentially expressed in pancreatic cancer tissues and
normal tissues, and FGD5-AS1 expression in cancerous tissues was
higher than that in normal pancreatic tissues (Fig. 1A). Consistently, RT-qPCR revealed
that FGD5-AS1 expression was significantly elevated in pancreatic
cancer tissues in comparison to the adjacent non-tumorous tissues
(Fig. 1B). Subsequently, RT-qPCR
was utilized for detecting FGD5-AS1 expression in the pancreatic
cancer cell lines (SW1990, PANC-1, BXPC-3 and CAPAN-1) and
pancreatic ductal epithelial cell line (HPDE6-C7), and it was
revealed that compared with HPDE6-C7 cells, FGD5-AS1 expression was
increased in the pancreatic cancer cell lines (Fig. 1C).
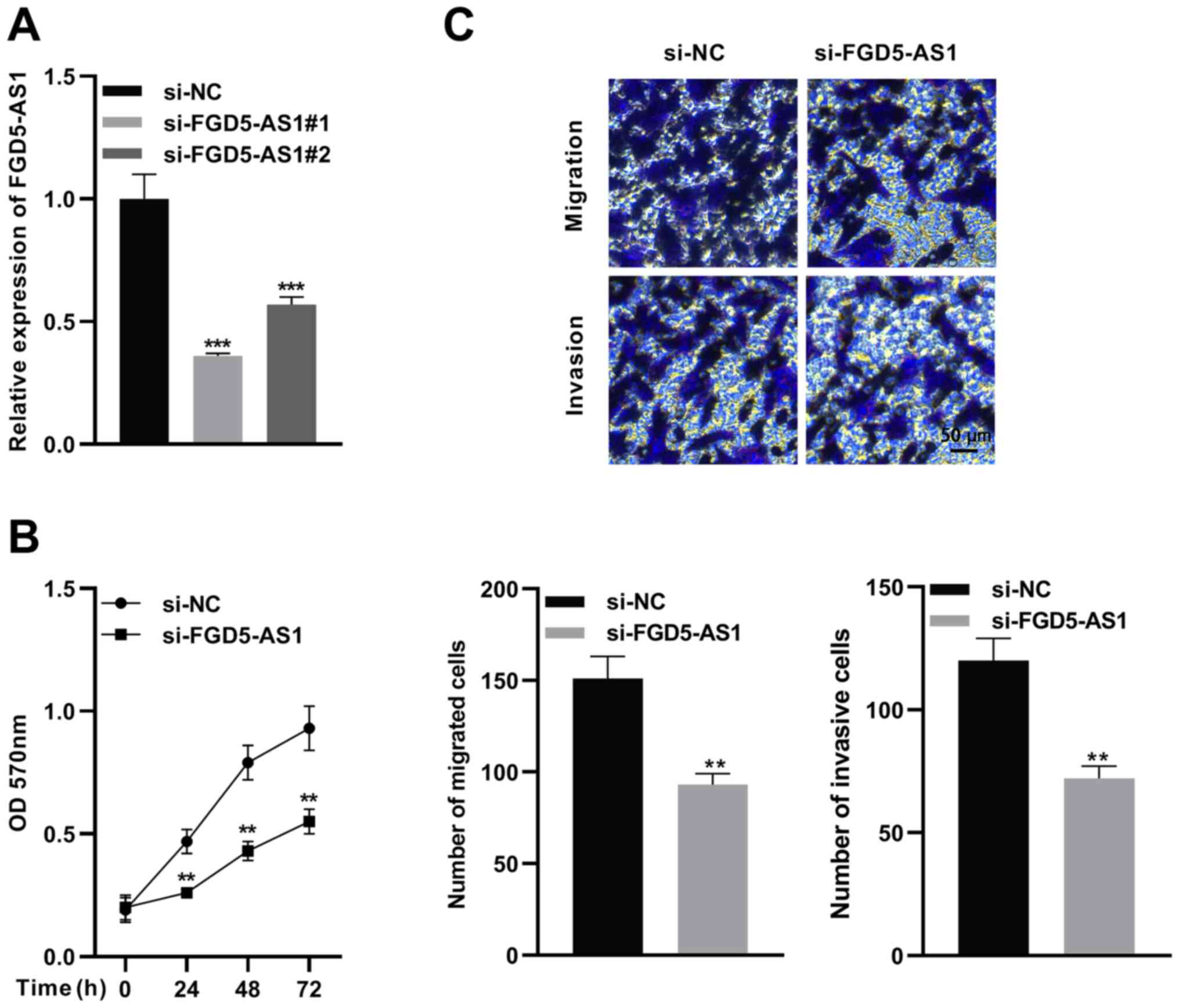
FGD5-AS1 knockdown suppresses SW1990
cell proliferation, migration and invasion
To determine the functions of FGD5-AS1 in pancreatic
cancer progression, two siRNAs for FGD5-AS1 were designed. Then,
FGD5-AS1 siRNA or control siRNA was transfected into SW1990 cells,
and the knockdown efficiency was verified by RT-qPCR (Fig. 2A). Next, the more effective siRNA
(si-FGD5-AS1 1) was selected for subsequent functional experiments.
MTT and Transwell assays were utilized to detect proliferation,
migration and invasion of SW1990 cells. It was revealed that
FGD5-AS1 knockdown significantly suppressed viability, migration
and invasion of SW1990 cells in contrast to the si-NC group
(Fig. 2B and C).
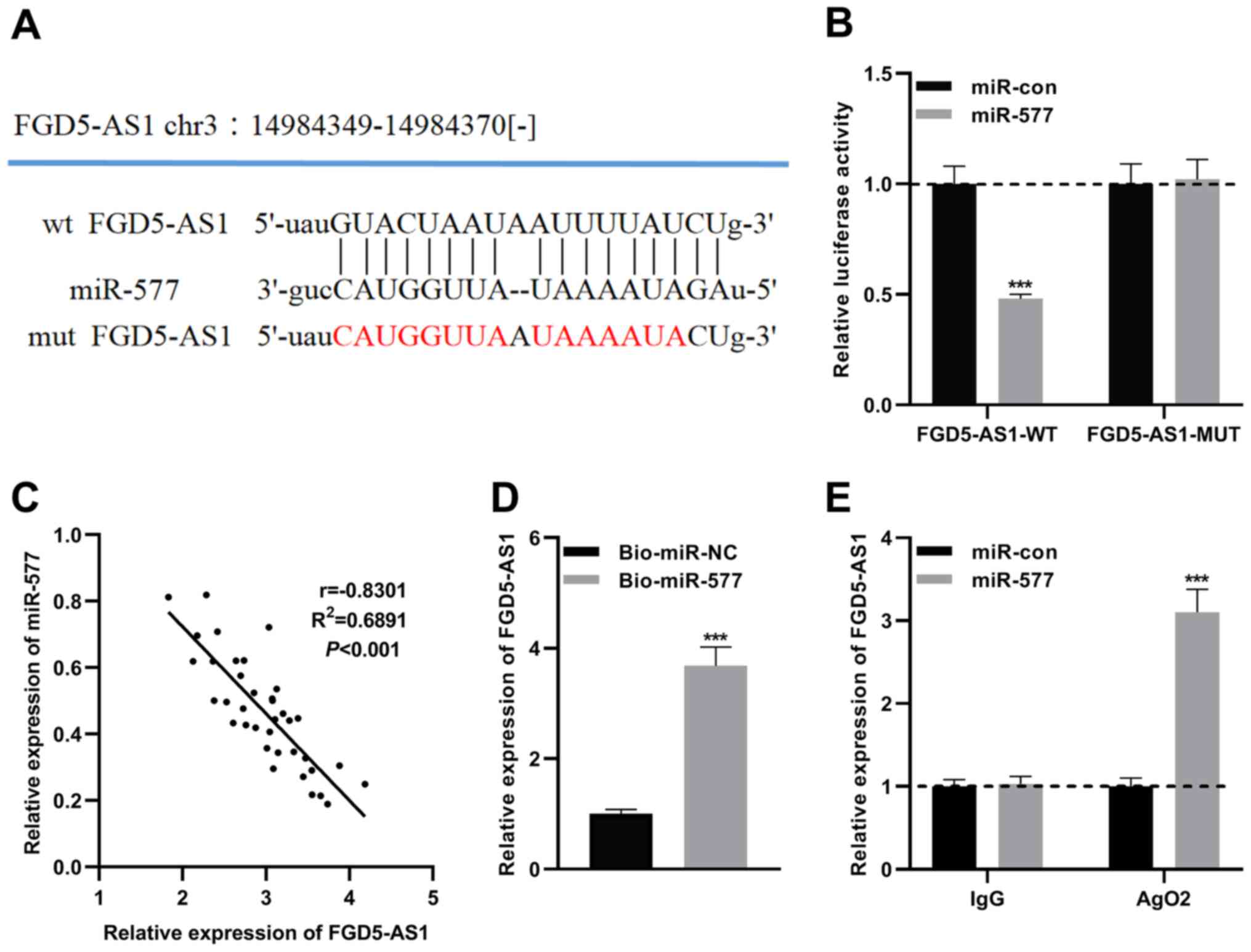
miR-577 is a downstream target of
FGD5-AS1
To clarify the mechanism of FGD5-AS1 in pancreatic
cancer progression, the potential downstream target miRNAs of
FGD5-AS1 were predicted by the starBase v2.0 database (http://starbase.sysu.edu.cn/panCancer.php) and LncBase
Predicted v.2 database (http://carolina.imis.athena-innovation.gr) (Tables SI and SII). Notably, a potential binding site
between miR-577 and FGD5-AS1 was predicted (Figs. 3A and S1). Recent research has revealed that
miR-577 is downregulated in pancreatic ductal adenocarcinoma, and
the low expression of miR-577 is significantly associated with poor
prognosis (21). Therefore, the
role of miR-577 in pancreatic cancer was investigated. To verify
the targeting relationship between miR-577 and FGD5-AS1, MUT
FGD5-AS1 luciferase reporter vector (FGD5-AS1-MUT) or WT FGD5-AS1
luciferase reporter vector (FGD5-AS1-WT) was co-transfected into
SW1990 cells with miR-577 mimics or control miRNA, and it was
revealed that miR-577 could only suppress the luciferase activity
of FGD5-AS-WT (Figs. 3B and
S2). Subsequently, Pearson's
correlation analysis revealed that FGD5-AS1 and miR-577 expression
levels were negatively correlated in pancreatic cancer tissues
(Fig. 3C). Additionally, RNA
pull-down and RIP assays were performed, and it was revealed that
ectogenic miR-577 enriched FGD5-AS1, and miR-577 and FGD5-AS1 were
enriched in Ago2-containing microribonucleoproteins (Fig. 3D and E). All of the aforementioned
findings indicated that miR-577 is a direct downstream target of
FGD5-AS1.
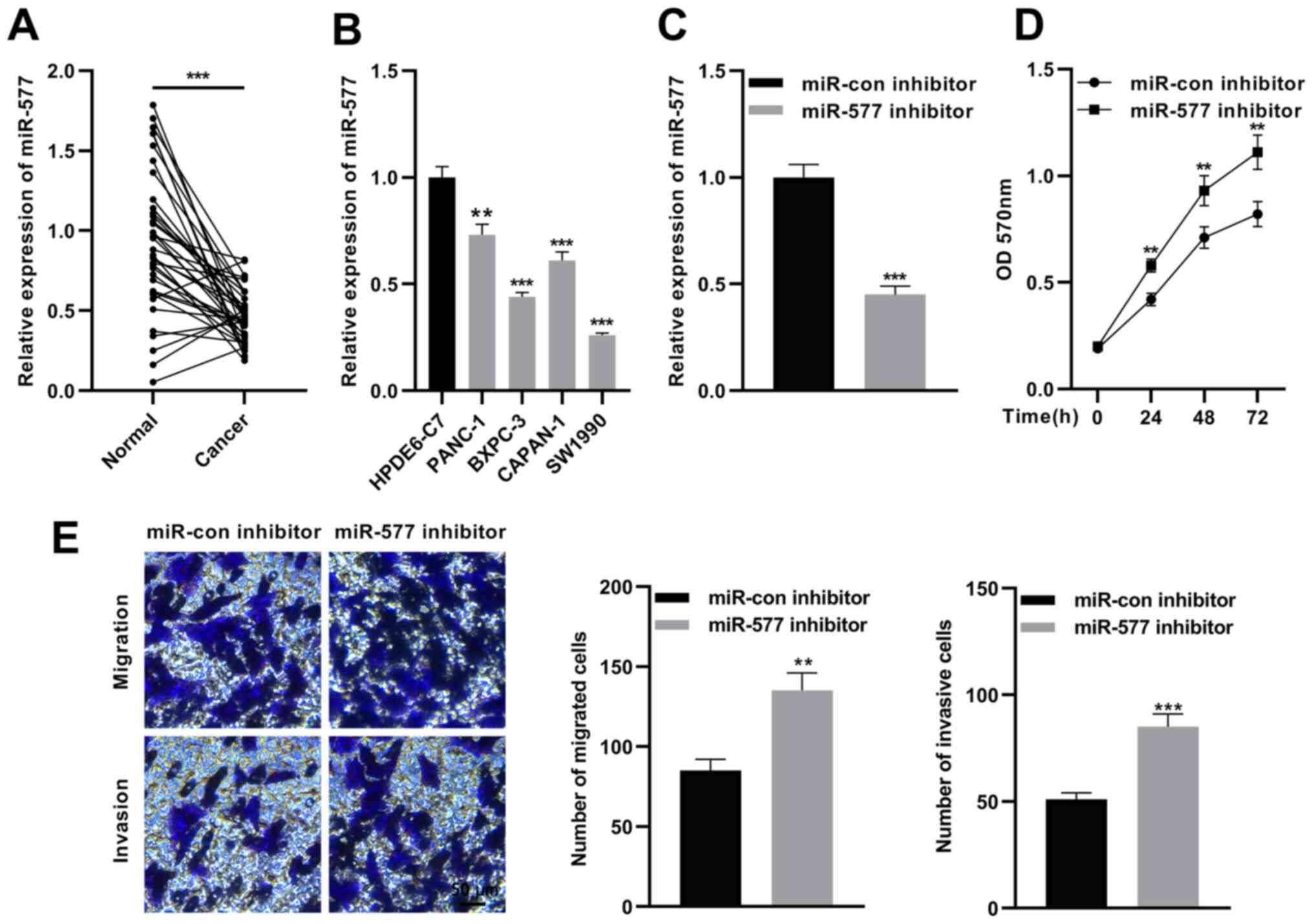
Inhibition of FGD5-AS1 promotes SW1990
cell proliferation, invasion and migration
The expression of FGD5-AS1 in pancreatic cancer was
then investigated. RT-qPCR revealed that miR-577 expression was
significantly reduced in pancreatic cancer tissues in comparison
with para-tumorous tissues (Fig.
4A), and compared with HPDE6-C7 cells, miR-577 expression was
significantly decreased in pancreatic cancer cell lines (Fig. 4B). Next, miR-577 inhibitors or
miR-con inhibitors were transfected into SW1990 cells (Fig. 4C). Subsequently, MTT and Transwell
assays were used to detect proliferation, migration and invasion of
SW1990 cells. As revealed in Fig. 4D
and E, miR-577 inhibition significantly increased
proliferation, migration and invasion of SW1990 cells. These
results suggested that miR-577 is a tumor suppressor in pancreatic
cancer.
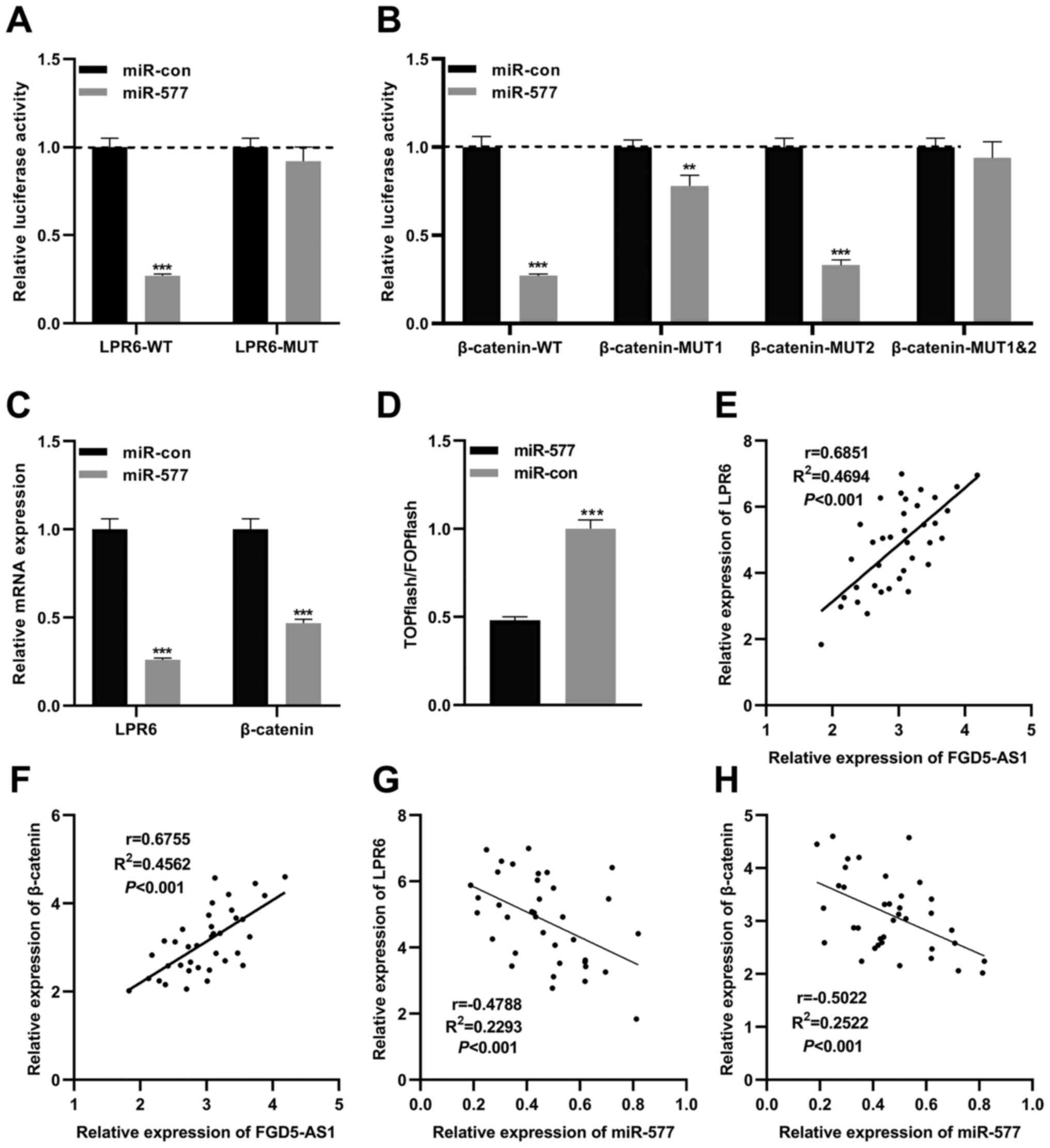
miR-577 regulates the Wnt/β-catenin
signaling in pancreatic cancer via targeting LRP6 and
β-catenin
A previous study reported that miR-577 inhibits the
growth of glioblastoma multiforme via modulating the Wnt/β-catenin
pathway; both β-catenin and LRP6, two crucial components of the
Wnt/β-catenin signaling, are directly targeted by miR-577 (22). Therefore, in the present study, it
was hypothesized that miR-577 was also associated with the
Wnt/β-catenin signaling in pancreatic cancer. Based on the previous
study (22), WT luciferase
reporter vectors (LRP6-WT and β-catenin-WT) and MUT luciferase
reporter vectors (LRP6-MUT, β-catenin-MUT1, β-catenin-MUT2 and
β-catenin-MUT1&2) were constructed. Subsequently, reporter
vectors and miR-577 mimics were co-transfected into SW1990 cells,
and the dual-luciferase reporter assays were performed. As revealed
in Fig. 5A, miR-577 mimics
significantly reduced the luciferase activity of LRP6-WT, yet
exerted no significant effect on that of LRP6-MUT. miR-577
significantly suppressed the luciferase activity of β-catenin-WT,
β-catenin-MUT1 and β-catenin-MUT2, yet did not suppress that of
β-catenin-MUT1&2 (Fig. 5B).
The aforementioned data suggested that miR-577 could bind to LRP6
3′ untranslated region (UTR) and β-catenin 3′UTR in pancreatic
cancer cells. As expected, following transfection of miR-577 mimics
into SW1990 cells, RT-qPCR revealed that the expression levels of
β-catenin mRNA and LRP6 mRNA were significantly reduced (Fig. 5C). Subsequently, miR-577 mimics (or
control miRNA) and the Wnt/β-catenin signaling reporter vector
TOPflash were co-transfected into SW1990 cells, and the luciferase
activity was detected 2 days following transfection. It was
revealed that miR-577 mimics notably decreased the luciferase
activity of pancreatic cancer cells, while control miRNA exerted no
significant effect, which suggested that miR-577 indeed suppressed
the activity of the Wnt/β-catenin signaling (Fig. 5D). Furthermore, Pearson's
correlation analysis revealed that FGD5-AS1 expression was
positively correlated with LRP6 expression and β-catenin expression
in pancreatic cancer tissues (Fig. 5E
and F), whereas miR-577 expression was negatively correlated
with them (Fig. 5G and H). These
results indicated that LRP6 and β-catenin were directly negatively
regulated by miR-577, and FGD5-AS1 could probably indirectly
modulate the activity of the Wnt/β-catenin pathway.
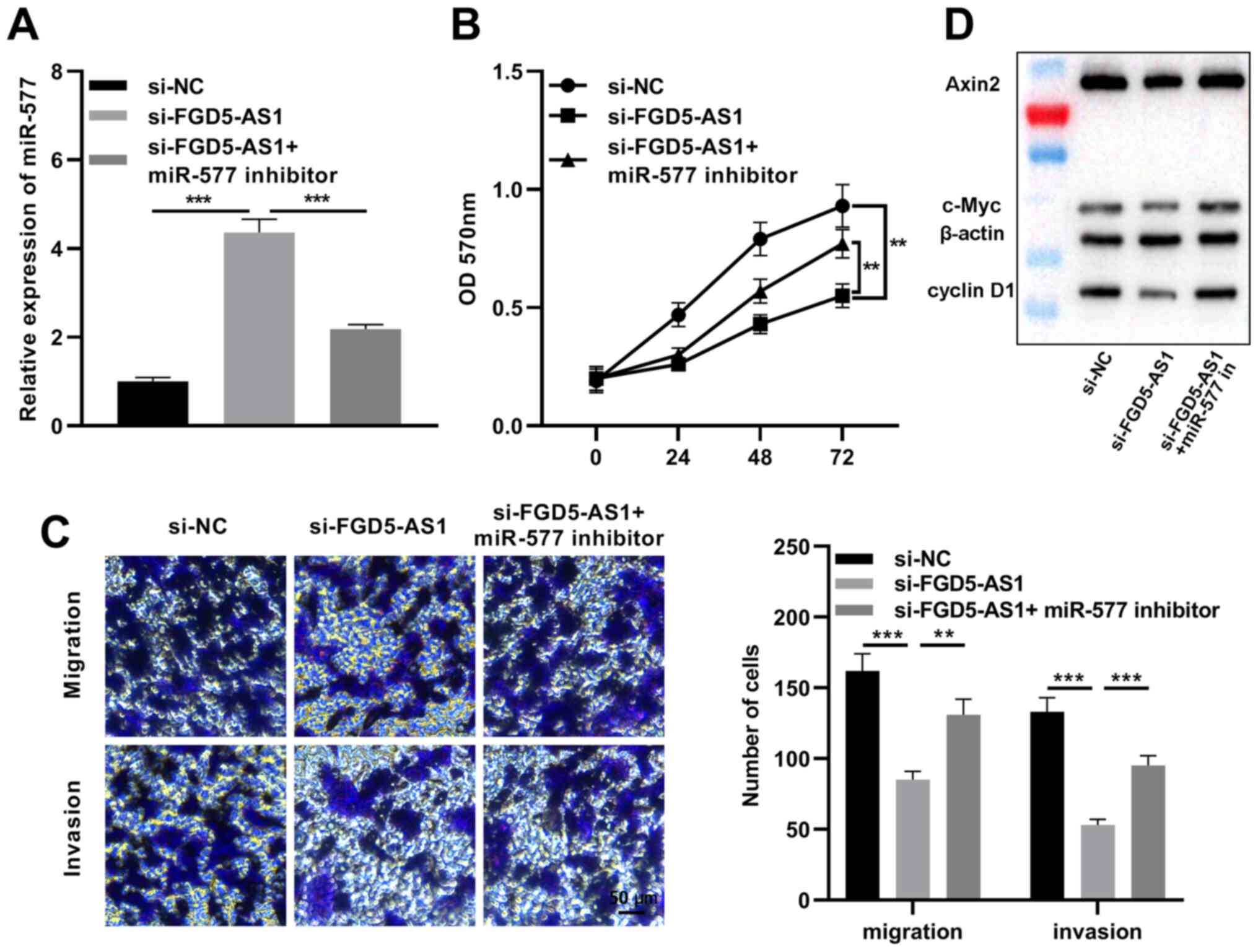
miR-577 reverses the effects of
FGD5-AS1 on pancreatic cancer cells
Subsequently, miR-577 inhibitors were transfected
into SW1990 cells with FGD5-AS1 knockdown (Fig. 6A). The MTT assay revealed that
miR-577 inhibition partially reversed the inhibitory effect of
FGD5-AS1 knockdown on SW1990 cell proliferation (Fig. 6B). The Transwell assay indicated
that miR-577 inhibition partially reversed the reduction caused by
FGD5-AS1 knockdown in the migration and invasion of SW1990 cells
(Fig. 6C). Additionally, to delve
deeper into the association between the Wnt/β-catenin signaling
pathway and the FGD5-AS1/miR-577 axis, western blotting was
conducted to detect the expression levels of Axin2, cyclin D1 and
c-Myc, which are considered as Wnt/β-catenin pathway-related
proteins. It was revealed that FGD5-AS1 knockdown significantly
decreased Axin2, cyclin D1 and c-Myc expression levels in SW1990
cells, and miR-577 inhibition partially counteracted these effects
of FGD5-AS1 knockdown (Fig. 6D).
The aforementioned findings suggested that FGD5-AS1 could regulate
the activity of the Wnt/β-catenin signaling via modulating
miR-577.
FGD5-AS1 enhances the lung metastasis
of pancreatic cancer cells in vivo
To confirm that FGD5-AS1 was indeed implicated in
the metastasis of pancreatic cancer, a lung metastasis model was
constructed with nude mice. After SW1990 cells were injected into
the mice via the caudal vein, the metastatic nodules in the lung
tissues were detected. It was revealed that SW1990 cells with
FGD5-AS1 knockdown reduced metastatic potential (Fig. S3 and Table SIII), forming less metastatic
nodules. This result further supported that FGD5-AS1 participates
in the progression of pancreatic cancer.
Discussion
It is estimated that there were approximately 44,330
mortalities caused by pancreatic cancer in the USA in 2018, and by
2030 it will be the second leading cause of cancer-related
mortality in the USA (23,24). The anatomical position of pancreas
is special, and the majority of patients with pancreatic cancer
have already reached the advanced stage when they are diagnosed. In
addition, the surgical resection rate is low and pancreatic cancer
cells are not sensitive to chemotherapy (25). Therefore, the prognosis of
pancreatic cancer is extremely poor. In recent years, molecular
targeted therapy has been gradually applied to cancer treatment and
has improved the prognosis of cancer patients (26). Therefore, it is of significant
clinical value to explore the mechanism of pancreatic cancer as
well as to explore novel therapeutic targets.
Multiple lncRNAs are aberrantly expressed in
cancerous tissues and mediate cancer progression by regulating
epigenetic modification, alternative splicing, transcription and
protein translation and lncRNAs have drawn widespread attention as
new therapeutic targets and biomarkers (27,28).
In pancreatic cancer, lncRNAs play crucial roles in regulating
various malignant biological behaviors of cancer cells, including
proliferation, migration, invasion, epithelial-mesenchymal
transition (EMT), chemoresistance and radioresistance (29). For example, lncRNA BX111 is
overexpressed in pancreatic cancer tissues, and highly expressed
BX111 is associated with advanced TNM stage, distant metastasis,
lymphatic vessel invasion and short overall survival rate of
patients; BX111 knockdown has been revealed to suppress pancreatic
cancer cell growth, invasion and the EMT process (30). In pancreatic cancer, lncRNA HOTTIP
is highly expressed, and its high expression is associated with
shorter overall survival and disease-free survival (31). The role of FGD5-AS1 in cancer
biology has gradually been revealed in recent years (12–15).
The present study, for the first time, to the best of our
knowledge, demonstrated that the expression level of FGD5-AS1 was
enhanced in pancreatic cancer tissues and cells. In addition, it
was revealed that FGD5-AS1 knockdown significantly suppressed the
malignant phenotype of pancreatic cancer cells. Our demonstrations
suggested that FGD5-AS1 is a potential biomarker and treatment
target for pancreatic cancer.
miRNAs are involved in the tumorigenesis and
progression of multiple malignancies including pancreatic cancer.
miRNAs are non-coding RNAs with 21–25 nucleotides in length, which
target the complementary sequences of mRNAs in the 3′-UTR to cause
translation inhibition or mRNA degradation, thereby regulating
pathological and physiological processes, such as cell
proliferation, migration, differentiation, apoptosis and
angiogenesis (32). Recent studies
have reported that miR-577 is downregulated in multiple cancers,
and its aberrant expression is related to tumorigenesis. For
instance, miR-577 expression was revealed to be decreased in
colorectal cancer samples and cell lines, and miR-577
overexpression inhibited tumor cell proliferation and metastasis
(33,34). Furthermore, miR-577 inhibited the
proliferation and EMT process via suppressing Rab25 expression in
breast cancer (35). miR-577 may
also suppress the progression of hepatocellular carcinoma,
osteosarcoma and glioblastoma via targeting the Wnt/β-catenin
signaling pathway (22,36,37).
The present study revealed that the expression level of miR-577 was
decreased in pancreatic cancer tissues and cells; additionally,
miR-577 inhibition significantly enhanced proliferation, migration
and invasion capabilities of pancreatic cancer cells, and miR-577
suppressed the activity of the Wnt/β-catenin signaling. Our data
indicated that miR-577 is a tumor suppressor in pancreatic
cancer.
The Wnt/β-catenin signaling pathway contributes to
the progression of pancreatic cancer. It was reported that the Wnt
signal inhibits the apoptosis of pancreatic cancer cells by
increasing the expression of survivin, a member of apoptosis
suppressors (38). Reportedly,
SMARCAD1 promotes the growth and metastasis of pancreatic cancer by
activating the Wnt/β-catenin pathway (39). In addition, the activation of the
Wnt/β-catenin pathway has also been revealed to induce the
resistance of pancreatic cancer cells to gemcitabine (40–42).
In the present study, it was confirmed that miR-577 targets LPR6
and β-catenin in pancreatic cancer cells, and LPR6 and β-catenin
expression levels were positively regulated by FGD5-AS1, which
helps to clarify the mechanism of the activation of the
Wnt/β-catenin pathway in pancreatic cancer.
lncRNAs may sponge miRNAs via miRNA response
elements, and function as competing endogenous RNAs (ceRNAs),
thereby indirectly modulating the translation of mRNAs (9). There are several studies suggesting
that FGD5-AS1 functions as a ceRNA to regulate the biological
behaviors of cancer cells. In particular, FGD5-AS1 facilitated
non-small cell lung carcinoma cell proliferation via suppressing
miR-107 to upregulate FGFRL1 (43); FGD5-AS1 facilitated oral squamous
cell carcinoma progression by sponging miR-520b and inducing the
expression of USP21 (44). The
present study revealed that FGD5-AS1 was a ceRNA for miR-577, and
it functioned as a molecular sponge to decoy miR-577; additionally,
FGD5-AS1 knockdown in SW1990 cells suppressed the expression levels
of the Wnt/β-catenin pathway-related proteins Axin2, c-Myc and
cyclin D1. These results further indicated that the interaction
between FGD5-AS1 and miR-577 was involved in the aberrant
activation of the Wnt/β-catenin signaling in pancreatic cancer.
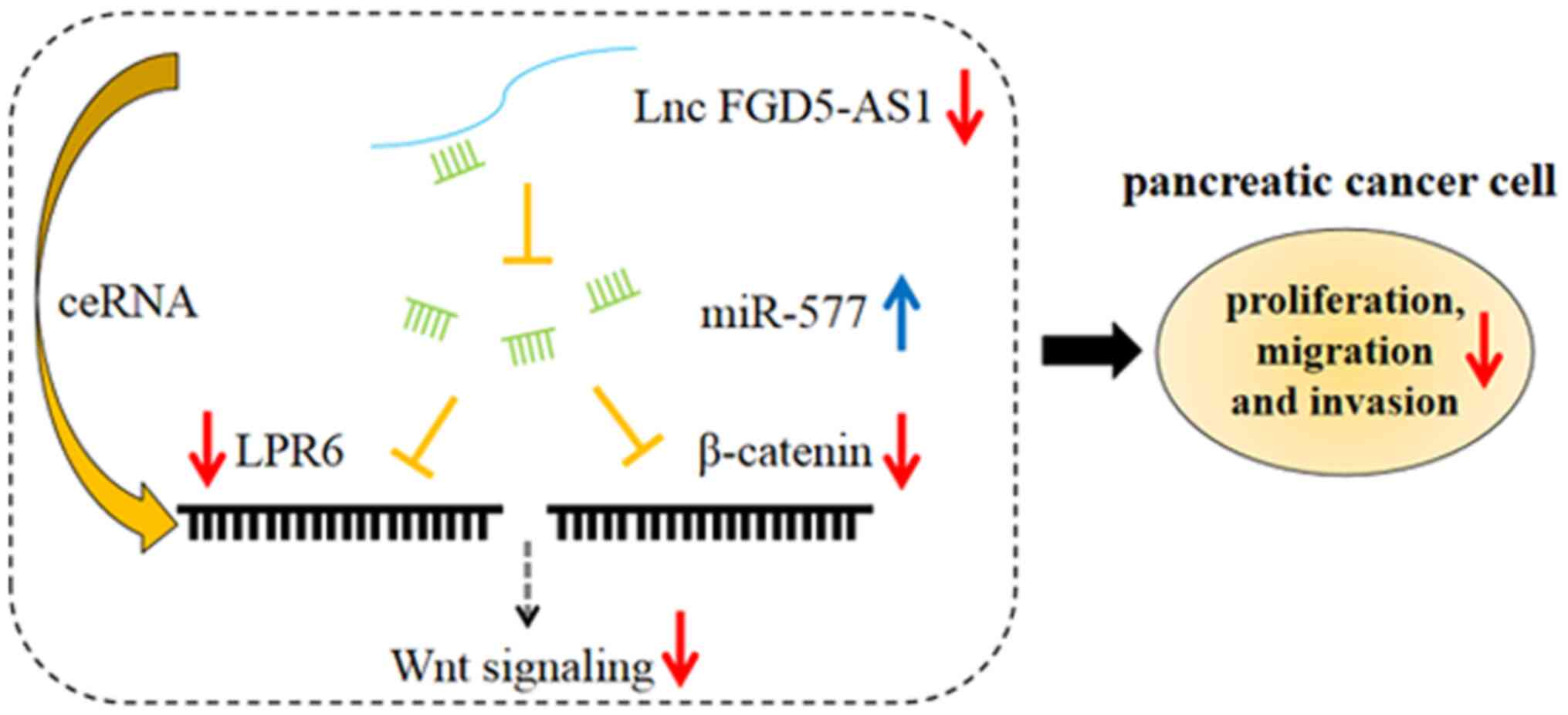
To sum up, in the present study, for the first time
to the best of our knowledge, it was demonstrated that FGD5-AS1 was
highly expressed in pancreatic cancer, and FGD5-AS1 promoted
pancreatic cancer cell proliferation, migration and invasion. By
contrast, miR-577 functioned as a tumor suppressor in pancreatic
cancer, inhibiting cancer cell proliferation, migration and
invasion. Additionally, FGD5-AS1 activated the Wnt/β-catenin
signaling via suppressing miR-577 (Fig. 7). Our study helped to clarify the
mechanism of pancreatic cancer progression and provided useful
insights into the treatment of this disease. There are some
limitations in the present study. The number of subjects was
limited, and thus it was impossible to analyze the pathological
characteristics and survival time more accurately. Therefore,
whether FGD5-AS1 can be extended to a wider range of applications
has yet to be determined. Notably, excluding functioning as a
ceRNA, lncRNA FGD5-AS1 may exert its biological effect via
functioning as a transcription regulator or protein-binding RNA,
and whether FGD5-AS1 could regulate the progression of pancreatic
cancer via these mechanisms requires further investigation in
subsequent studies.
Supplementary Material
Supporting Data
Supporting Data
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Science
Foundation of China (NSFC; grant no. 81802390), the Natural Science
Foundation of Beijing (BJNSF; grant no. 7202187) and the Military
Medicine Youth Special Project of PLA General Hospital (grant no.
QNF19037).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WTZ and WJY made substantial contributions to the
conception and design, and also critically reviewed the study. WTZ,
WJY, JJZ, QS and YKW made substantial contributions to the study
execution and acquisition of data. WTZ, WJY, JPJ, BQ and XWT made
substantial contributions to the data analysis and interpretation.
WTZ and WJY confirmed the authenticity of all the raw data and
participated in revising the present study. All authors read and
approved the final version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Ethics
Committee of the Sixth Hospital of Shanxi Medical University
(Taiyuan, China) and written informed consents were acquired from
all enrolled patients. The animal experiments were approved
(approval no. 8217113963) by the Institutional Animal Care and Use
Committee of Chinese PLA General Hospital (Beijing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Klaiber U, Hackert T and Neoptolemos JP:
Adjuvant treatment for pancreatic cancer. Transl Gastroenterol
Hepatol. 4:272019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global cancer statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kong K, Guo M, Liu Y and Zheng J: Progress
in animal models of pancreatic ductal adenocarcinoma. J Cancer.
11:1555–1567. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ma YY, Shi JJ, Chen JB, Xu KC and Niu LZ:
Irreversible electroporation for liver metastasis from pancreatic
cancer: A case report. World J Clin Cases. 8:390–397. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Perinel J and Adham M: Palliative therapy
in pancreatic cancer-palliative surgery. Transl Gastroenterol
Hepatol. 4:282019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Liu G, Jiang Z, Qiao M and Wang F:
Lnc-GIHCG promotes cell proliferation and migration in gastric
cancer through miR-1281 adsorption. Mol Genet Genomic Med.
7:e7112019. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fu R, Wang X, Hu Y, Du H, Dong B, Ao S,
Zhang L, Sun Z, Zhang L, Lv G, et al: Solamargine inhibits gastric
cancer progression by regulating the expression of lncNEAT1_2 via
the MAPK signaling pathway. Int J Oncol. 54:1545–1554.
2019.PubMed/NCBI
|
|
8
|
Evans JR, Feng FY and Chinnaiyan AM: The
bright side of dark matter: lncRNAs in cancer. J Clin Invest.
126:2775–2782. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu XH, Sun M, Nie FQ, Ge YB, Zhang EB,
Yin DD, Kong R, Xia R, Lu KH, Li JH, et al: Lnc RNA HOTAIR
functions as a competing endogenous RNA to regulate HER2 expression
by sponging miR-331-3p in gastric cancer. Mol Cancer. 13:922014.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shen LS, Hu XF, Chen T, Shen GL and Cheng
D: Integrated network analysis to explore the key mRNAs and lncRNAs
in acute myocardial infarction. Math Biosci Eng. 16:6426–6437.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li S, Liu X, Li H, Pan H, Acharya A, Deng
Y, Yu Y, Haak R, Schmidt J, Schmalz G, et al: Integrated analysis
of long noncoding RNA-associated competing endogenous RNA network
in periodontitis. J Periodontal Res. 53:495–505. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lei Y, Shi Y, Duan J, Liu Y, Lv G, Shi R,
Zhang F, Yang Q and Zhao W: Identification of alternative splicing
and lncRNA genes in pathogenesis of small cell lung cancer based on
their RNA sequencing. Adv Clin Exp Med. 28:1043–1050. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu H, Lu J, Zhao H, Chen Z, Cui Q, Lin Z,
Wang X, Wang J, Dong H, Wang S, et al: Functional long noncoding
RNAs (lncRNAs) in clear cell kidney carcinoma revealed by
reconstruction and comprehensive analysis of the lncRNA-miRNA-mRNA
regulatory network. Med Sci Monit. 24:8250–8263. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hamilton MJ, Girke T and Martinez E:
Global isoform-specific transcript alterations and deregulated
networks in clear cell renal cell carcinoma. Oncotarget.
9:23670–23680. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li D, Jiang X, Zhang X, Cao G, Wang D and
Chen Z: Long noncoding RNA FGD5-AS1 promotes colorectal cancer cell
proliferation, migration, and invasion through upregulating CDCA7
via sponging miR-302e. In Vitro Cell Dev Biol Anim. 55:577–585.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Fan J, Wei Q, Liao J, Zou Y, Song D, Xiong
D, Ma C, Hu X, Qu X, Chen L, et al: Noncanonical Wnt signaling
plays an important role in modulating canonical Wnt-regulated
stemness, proliferation and terminal differentiation of hepatic
progenitors. Oncotarget. 8:27105–27119. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang L, Cheng H, Yue Y, Li S, Zhang D and
He R: H19 knockdown suppresses proliferation and induces apoptosis
by regulating miR-148b/WNT/β-catenin in ox-LDL-stimulated vascular
smooth muscle cells. J Biomed Sci. 25:112018. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zimmerman M, Hu X and Liu K: Experimental
metastasis and CTL adoptive transfer immunotherapy mouse model. J
Vis Exp. 45:20772010.
|
|
21
|
Yang J, Cong X, Ren M, Sun H, Liu T, Chen
G, Wang Q, Li Z, Yu S and Yang Q: Circular RNA hsa_circRNA_0007334
is predicted to promote MMP7 and COL1A1 expression by functioning
as a miRNA sponge in pancreatic ductal adenocarcinoma. J Oncol.
2019:76308942019. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang W, Shen C, Li C, Yang G, Liu H, Chen
X, Zhu D, Zou H, Zhen Y, Zhang D, et al: miR-577 inhibits
glioblastoma tumor growth via the Wnt signaling pathway. Mol
Carcinog. 55:575–585. 2016. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ilic M and Ilic I: Epidemiology of
pancreatic cancer. World J Gastroenterol. 22:9694–9705. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Grant TJ, Hua K and Singh A: Molecular
pathogenesis of pancreatic cancer. Prog Mol Biol Transl Sci.
144:241–275. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Masiak-Segit W, Rawicz-Pruszyński K,
Skórzewska M and Polkowski WP: Surgical treatment of pancreatic
cancer. Pol Przegl Chir. 90:45–53. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lee YT, Tan YJ and Oon CE: Molecular
targeted therapy: Treating cancer with specificity. Eur J
Pharmacol. 834:188–196. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Duguang L, Jin H, Xiaowei Q, Peng X,
Xiaodong W, Zhennan L, Jianjun Q and Jie Y: The involvement of
lncRNAs in the development and progression of pancreatic cancer.
Cancer Biol Ther. 18:927–936. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chandra Gupta S and Nandan Tripathi Y:
Potential of long non-coding RNAs in cancer patients: From
biomarkers to therapeutic targets. Int J Cancer. 140:1955–1967.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Y, Yang X, Kang X and Liu S: The
regulatory roles of long noncoding RNAs in the biological behavior
of pancreatic cancer. Saudi J Gastroenterol. 25:145–151. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Deng SJ, Chen HY, Ye Z, Deng SC, Zhu S,
Zeng Z, He C, Liu ML, Huang K, Zhong JX, et al: Hypoxia-induced
LncRNA-BX111 promotes metastasis and progression of pancreatic
cancer through regulating ZEB1 transcription. Oncogene.
37:5811–5828. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fu Z, Chen C, Zhou Q, Wang Y, Zhao Y, Zhao
X, Li W, Zheng S, Ye H, Wang L, et al: LncRNA HOTTIP modulates
cancer stem cell properties in human pancreatic cancer by
regulating HOXA9. Cancer Lett. 410:68–81. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou L, Liang X, Zhang L, Yang L, Nagao N,
Wu H, Liu C, Lin S, Cai G and Liu J: MiR-27a-3p functions as an
oncogene in gastric cancer by targeting BTG2. Oncotarget.
7:51943–51954. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang Y, Lu Z, Wang N, Feng J, Zhang J,
Luan L, Zhao W and Zeng X: Long noncoding RNA DANCR promotes
colorectal cancer proliferation and metastasis via miR-577
sponging. Exp Mol Med. 50:1–17. 2018. View Article : Google Scholar
|
|
34
|
Du C, Wang HX, Chen P and Chen CH:
STAT3-induced upregulation of lncRNA DUXAP8 functions as ceRNA for
miR-577 to promote the migration and invasion in colorectal cancer
through the regulation of RAB14. Eur Rev Med Pharmacol Sci.
23:6105–6118. 2019.PubMed/NCBI
|
|
35
|
Yin C, Mou Q, Pan X, Zhang G, Li H and Sun
Y: MiR-577 suppresses epithelial-mesenchymal transition and
metastasis of breast cancer by targeting Rab25. Thorac Cancer.
9:472–479. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang LY, Li B, Jiang HH, Zhuang LW and Liu
Y: Inhibition effect of miR-577 on hepatocellular carcinoma cell
growth via targeting β-catenin. Asian Pac J Trop Med. 8:923–929.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Jiang Z, Jiang C and Fang J: Up-regulated
lnc-SNHG1 contributes to osteosarcoma progression through
sequestration of miR-577 and activation of WNT2B/Wnt/β-catenin
pathway. Biochem Biophys Res Commun. 495:238–245. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Modi S, Kir D, Banerjee S and Saluja A:
Control of apoptosis in treatment and biology of pancreatic cancer.
J Cell Biochem. 117:279–288. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu F, Xia Z, Zhang M, Ding J, Feng Y, Wu
J, Dong Y, Gao W, Han Z, Liu Y, et al: SMARCAD1 promotes pancreatic
cancer cell growth and metastasis through Wnt/β-catenin-mediated
EMT. Int J Biol Sci. 15:636–646. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nagano H, Tomimaru Y, Eguchi H, Hama N,
Wada H, Kawamoto K, Kobayashi S, Mori M and Doki Y: MicroRNA-29a
induces resistance to gemcitabine through the Wnt/β-catenin
signaling pathway in pancreatic cancer cells. Int J Oncol.
43:1066–1072. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Zhan T, Chen X, Tian X, Han Z, Liu M, Zou
Y, Huang S, Chen A, Cheng X, Deng J, et al: MiR-331-3p links to
drug resistance of pancreatic cancer cells by activating
WNT/β-catenin signal via ST7L. Technol Cancer Res Treat.
19:15330338209458012020. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhou C, Yi C, Yi Y, Qin W, Yan Y, Dong X,
Zhang X, Huang Y, Zhang R, Wei J, et al: LncRNA PVT1 promotes
gemcitabine resistance of pancreatic cancer via activating
Wnt/β-catenin and autophagy pathway through modulating the
miR-619-5p/Pygo2 and miR-619-5p/ATG14 axes. Mol Cancer. 19:1182020.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Fan Y, Li H, Yu Z, Dong W, Cui X, Ma J and
Li S: Long non-coding RNA FGD5-AS1 promotes non-small cell lung
cancer cell proliferation through sponging hsa-miR-107 to
up-regulate FGFRL1. Biosci Rep. 40:BSR201933092020. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Liu L, Zhan Y, Huang Y and Huang L: LncRNA
FGD5-AS1 can be predicted as therapeutic target in oral cancer. J
Oral Pathol Med. 49:243–252. 2020. View Article : Google Scholar : PubMed/NCBI
|