Introduction
Dysregulated cellular metabolism is an emerging
hallmark of cancer as it supports cellular proliferation and
division via biomass generation (1,2). The
most extensively researched metabolic characteristic of cancer
cells is the Warburg effect, which favors glycolysis over oxidative
phosphorylation (OXPHOS) to generate energy, even in the presence
of sufficient oxygen (1). Genomic
analysis has revealed that >70% of all human cancer cases show
ubiquitous overexpression of glycolytic pathway genes (3,4).
Expression levels of genes involved in glucose metabolism,
including enolase 1, hexokinase 2 (HK2),
glyceraldehyde-3-phosphate dehydrogenase (GAPDH), glucose
transporter 1 (GLUT1), pyruvate kinase M2 (PKM2),
lactate dehydrogenase A (LDHA), and pyruvate dehydrogenase
kinase 1 (PDK1), are upregulated in most cancer tissues, and
hence, can be used as therapeutic targets (3,5,6). The
metabolic switch is directed by limiting the pyruvate utilization
in the mitochondrial tricarboxylic acid (TCA) cycle, which is
regulated by various cellular events, including altered growth
factor signaling, hypoxic or normoxic activation of
hypoxia-inducible factor 1α (HIF1α), oncogene activation and
loss-of-function of suppressor genes (5,7).
Andrographolide (AG), a diterpenoid lactone, was
first identified in Andrographis paniculata (Burm.f.) Nees,
an annual herbaceous plant belong to the Acanthaceae family,
commonly referred to as ‘green chiretta’ (8). AG is widely accepted as a natural
anticancer agent owing to its ability to inhibit cancer growth and
induce apoptotic cell death (9).
Several cellular pathways, including Wnt/β-catenin,
phosphatidylinositol-3-kinase (PI3K)-mammalian target of rapamycin,
vascular endothelial growth factor-mediated signaling, and tumor
necrosis factor-related apoptosis-inducing ligand-mediated
apoptosis pathways, have been identified as key pathways regulated
by AG (9,10). AG directly binds to Ras and nuclear
factor-κB and lowers their expression levels or activities
(11,12). AG targets HIF1α by downregulating
its expression or stability, thereby inhibiting its DNA-binding
activity (13–16). Recently, two separate groups have
shown that AG inhibits the glycolytic metabolic profile of cancer
cells by controlling the expression of enzymes involved in
glycolysis, including GLUT1, HK2, phosphofructokinase 1 (PFK1),
PKM2 and LDHA (17,18). The molecular mechanism by which AG
suppresses aerobic glycolysis has not yet been fully
characterized.
In the present study, it was found that AG
suppressed aerobic glycolysis and induced mitochondria-mediated
apoptosis in human lung cancer cells. In addition, AG decreased the
expression levels of several PDKs, including PDK1. Apoptotic cell
death was reduced in PDK1-overexpressing cells. Based on these
findings, it was hypothesized that the pro-apoptotic activity of AG
may be mediated by the negative regulation of PDK1 expression.
Materials and methods
Materials
AG was purchased from the Tokyo Chemical Industry. A
100 mM stock solution of AG was dissolved in dimethyl sulfoxide
(DMSO) and stored at −20°C. AG was diluted to the appropriate
concentration with a final DMSO concentration of less than 0.1%.
All other chemicals, including DMSO, sodium dodecyl sulfate (SDS),
Tween 20, and 3-(4,5-dimethyl-thiazole-2-yl)2,5-diphenyl
tetrazolium bromide (MTT), were purchased from MilliporeSigma. All
antibodies used in the present study for western blotting are
listed in Table I. Roswell Park
Memorial Institute (RPMI)-1640 medium, 1X phosphate-buffered saline
(PBS; pH 7.4), and 0.25% trypsin-ethylenediaminetetraacetic acid
(EDTA) were obtained from Welgene, Inc. Fetal bovine serum (FBS)
and penicillin/streptomycin were obtained from Gibco; Thermo Fisher
Scientific, Inc.
 | Table I.List of antibodies used in the
present study. |
Table I.
List of antibodies used in the
present study.
| Antibody | Company | Cat. no. | Dilution
factor |
|---|
| Phosphorylated | Abcam | ab177461 | 1:1,000 in 2% skim
milk |
| PDHA1 |
|
|
|
| PDHA1 | Santa Cruz
Biotechnology, Inc. | sc-377092 | 1:2,000 in 5% skim
milk |
| PDK1 | Enzo Life Sciences,
Inc. | ADI-KAP-PK112 | 1:2,000 in 5% skim
milk |
| PDK2 | Signalway antibody
LLC | 41330 | 1:1,000 in 5% skim
milk |
| PDK3 | Novus Biologicals,
LLC | NBP1-32581 | 1:1,000 in 5% skim
milk |
| PDK4 | Signalway antibody
LLC | 38562 | 1:1,000 in 5% skim
milk |
| GAPDH | Santa Cruz
Biotechnology, Inc. | sc-32233 | 1:2,000 in 5% skim
milk |
| PARP | Cell Signaling
Technology, Inc. | 9542 | 1:1,000 in 5% skim
milk |
| Cleaved
Caspase-3 | Cell Signaling
Technology, Inc. | 9661 | 1:1,000 in 2% skim
milk |
| Caspase-9 | Cell Signaling
Technology, Inc. | 9508 | 1:1,000 in 2% skim
milk |
| HSP90 | Santa Cruz
Biotechnology, Inc. | sc-13119 | 1:2,000 in 5% skim
milk |
| Anti-mouse IgG | Invitrogen; Thermo
Fisher Scientific, Inc. | RJ240410 | 1:2,000 in 5% skim
milk |
| Anti-rabbit
IgG | Invitrogen; Thermo
Fisher Scientific, Inc. | SA245916 | 1:2,000 in 5% skim
milk |
Cell lines and cultures
Human non-small cell lung cancer (NSCLC) cell lines,
namely A549 (adenocarcinoma), NCI-H292 (H292; mucoepidermoid
carcinoma) and NCI-H522 (H522; adenocarcinoma), were obtained from
the Korean Cell Line Bank (Seoul, Korea). Cells were cultured in
RPMI-1640 medium supplemented with 10% FBS and 1%
penicillin/streptomycin in a 5% CO2 incubator at 37°C
under 98% humidified conditions.
Cell viability assay
Cell viability was determined via the MTT (dissolved
in DMSO) assay. Cells (1×104 cells/well) were initially
seeded into a 96-well culture plate in triplicate, followed by
treatment with various concentrations of AG (10, 20, 50 and 100 µM)
the next day in a fresh medium. After 24 h of incubation, MTT
solution (5 mg/ml) was diluted in culture medium and added to each
well of the plate to a final concentration of 0.5 mg/ml, and the
culture plate was further incubated at 37°C for 4 h. Absorbance was
measured at 540 nm using the Spectramax M2 microplate reader
(Molecular Devices, LLC).
Lactate production assay
Lactate production in H292 cells was measured using
the lactate fluorometric assay kit (BioVision, Inc.). Briefly,
1×104 cells/well were seeded into a 96-well plate and
incubated overnight at 37°C. Prior to pretreatment with various
concentrations of AG (20, 50 and 75 µM), the medium was replaced
with phenol red and serum-free RPMI medium and incubated at 37°C
for 1 h. Finally, 1 µl of the medium from each well was received to
measure the absorbance at 570 nm using the Spectramax M2 microplate
reader (Molecular Devices, LLC).
Western blot analysis
After incubation, the cells were harvested and lysed
in ice-cold 1% NP-40 lysis buffer containing 150 mM NaCl, 10 mM
HEPES (pH 7.45), 5 mM sodium pyrophosphate, 5 mM sodium fluoride, 2
mM sodium orthovanadate, and a protease inhibitor cocktail (Roche
Applied Science). Whole cell extracts were obtained from the
supernatants after centrifugation of the cell lysate at 13,000 × g
for 15 min at 4°C. Protein concentration was quantitated using the
Bradford assay. Proteins (50 µg) were separated via
SDS-polyacrylamide gel electrophoresis (PAGE; 8–13%) and
transferred onto 0.45 µM nitrocellulose membranes (Amersham;
Cytiva). The membranes were blocked for 60 min at room temperature
using 5% (w/v) BD Difco skim milk powder (Thermo Fisher Scientific,
Inc.) and immunoblotted with primary antibodies at 4°C overnight at
optimal dilution (Table I). After
washing three with Tris-buffered saline with 0.1% Tween 20 buffer,
the membranes were incubated with horseradish peroxidase-conjugated
secondary antibodies at a dilution of 1:2,000 for 1 h at room
temperature. Immunoreactivity was detected using ECL Plus
(Amersham; Cytiva) and digitalized using Image Quant LAS 4000 (GE
Healthcare).
In vitro PDK activity assay
Kinase activity of PDK was slightly modified as
previously described (19,20). Briefly, 50 ng of recombinant PDK1
(Abcam) was incubated with 100 ng of recombinant pyruvate
dehydrogenase E1 subunit alpha 1 (PDHA1; Abcam) for 30 min at 37°C
in PDK1 buffer containing 20 mM Tris buffer (pH 7.5), 0.1 mM EDTA,
1 mM MgCl2, 2 mM dithiothreitol (DTT) and 250 µM ATP.
The samples were subjected to SDS-PAGE and immunoblotted using
antibodies against PDK1, p-PDHA1, and PDHA1 (Table I).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was isolated from H292 cells using the
TRIZOL® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) and quantified at 260 nm wavelength using the Nanodrop 2000
spectrophotometer (Thermo Fisher Scientific, Inc.), according to
the manufacturers' instructions (21). An equal amount of RNA (1 µg) from
each sample was used to synthesize cDNA, which was
reverse-transcribed using RevertAid reverse transcriptase (cat. no.
EP0441; Thermo Fisher Scientific, Inc.). RT was performed according
to the manufacturer's instructions. qPCR was performed using the
StepOnePlus real-time PCR system (Applied Biosystems; Thermo Fisher
Scientific, Inc.) with the RealHelix qPCR kit (cat. no. QP2-S500;
NanoHelix Co., Ltd.). PCR was performed with initial denaturation
at 95°C for 15 min, followed by 40 cycles of the reaction at 95°C
for 20 sec, 60°C for 30 sec, and 72°C for 30 sec. Relative mRNA
levels were normalized to actin levels. All the primers used in the
present study are listed in Table
II.
| Table II.List of primers used in the present
study. |
Table II.
List of primers used in the present
study.
| Gene name | Primer sequence
(5′→3′) |
|---|
| PDK1 | F:
AAGAATTCCATGAGGCTGGCGCGGCTGC |
| cloning | R:
ACCTCGAGCTAGGCACTGCGGAACGTCG |
| PDK1 | F:
CTATGAAAATGCTAGGCGTCT |
|
| R:
AACCACTTGTATTGGCTGTCC |
| PDK2 | F:
AGGACACCTACGGCGATGA |
|
| R:
TGCCGATGTGTTTGGGATGG |
| PDK3 | F:
GCCAAAGCGCCAGACAAAC |
|
| R:
CAACTGTCGCTCTCATTGAGT |
| β-actin | F:
CAAGAGATGGCCACGGCTGCT |
|
| R:
CACAGGACTCCATGCCCAGGA |
Flow cytometric analysis
Apoptotic cells were examined using the Annexin
V-fluorescein isothiocyanate (FITC) Apoptosis Detection Kit (Thermo
Fisher Scientific, Inc.). Cells (3×105 cells/well) were
seeded into a six-well plate and incubated overnight at 37°C. The
next day, cells were treated with various concentrations of AG (20,
50, and 75 µM) for 24 h and resuspended in 500 µl of binding buffer
(1X) to obtain a cell density of 2×105 cells/ml. The
cells were then incubated with 5 µl of Annexin V-FITC and 10 µl
propidium iodide (PI; 20 µg/ml) in cell suspension for 15 min at
room temperature. Fluorescence intensities of the samples were
determined using the FACS Canto II flow cytometer (BD Biosciences).
In the histogram of FACS analysis, apoptotic cells were represented
the combination of Q2 (PI+/Annexin V+, late
apoptotic cell) and Q3 (PI−/Annexin V+, early
apoptotic cell), and dead cells were represented the combination of
Q1 (PI+/Annexin V−, necrotic cell) and Q2
(PI+/Annexin V+, late apoptotic cell).
Determination of mitochondrial
reactive oxygen species (ROS) levels
The production of intracellular mitochondrial ROS
was determined using MitoSOX Red (Invitrogen; Thermo Fisher
Scientific, Inc.). Briefly, 1 µM MitoSOX Red was added to the
cultured cells in conditioned medium and incubated at 37°C for 10
min. Fluorescence intensity was determined using a fluorescence
microscope (AX10 Imager M1; Zeiss AG) and calculated using the
ImageJ software (version 1.53; National Institutes of Health).
Measurement of mitochondrial membrane
potential (MMP)
Cells (3×105 cells/well) were seeded into
a six-well plate and incubated overnight at 37°C. The next day, the
cells were treated with various concentrations of AG (20, 50 and 75
µM) for 12 h and incubated with 250 nM tetramethylrhodamine methyl
ester (TMRM; Thermo Fisher Scientific, Inc.) for 30 min. The cells
were washed thrice with PBS and observed under a fluorescence
microscope (AX10 Imager M1). Membrane depolarization was determined
by analyzing the captured images using the ImageJ software (version
1.53; National Institutes of Health).
Plasmid preparation and transfection
for PDK1 overexpression
PCR product of PDK1 was ligated into the
pMX–IRES-puromycin vector containing EcoRI and XhoI
restriction enzyme sites. The primers used to clone the PDK1
construct are listed in Table II.
Plat-A cells, a retroviral packaging cell line, were transfected
with 1 µg of pMX–IRES empty vector (EV) and pMX–IRES-PDK1 vector
(PDK1-OE) using Lipofectamine 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). The supernatant was collected after 24 h, and
the culture medium of H292 cells was replaced with filtered
retroviral supernatant containing 5 µg/ml polybren (Santa Cruz
Biotechnology, Inc.). The cells were selected using 2 µg/ml
puromycin after viral infection for 24 h and cultured for an
additional 2 weeks.
Bioinformatics analysis
The known target gene list of AG was downloaded from
PubChem (https://pubchem.ncbi.nlm.nih.gov/#query=andrographolide)
and analyzed using Cytoscape's JEPPETTO plugin (https://apps.cytoscape.org/apps/jepetto). Kyoto
Encyclopedia of Genes and Genomes analysis (https://www.genome.jp/kegg/) identified several
cancerous, metabolic and p53 signaling pathways as AG-related
pathways. Interaction between genes and chemicals by AG were
analyzed using BioCarta Pathways Dataset (https://maayanlab.cloud/Harmonizome/search?t=all&q=andrographolide).
The GSE74769 microarray dataset (obtained from the Gene Expression
Omnibus database; http://www.ncbi.nlm.nih.gov/geo/) was examined to
identify a potential link between glycolysis and AG. The
correlation between PDK mRNA expression and AG cytotoxicity in the
cells was performed using Spearman's correlation analysis.
Statistical analysis
All data are from at least three independent
experiments and expressed as the mean ± standard deviation.
Statistical differences were calculated via one-way analysis of
variance with Dunnett's post hoc test or unpaired Student's t-test.
GraphPad Prism software (version 5.0; GraphPad Software, Inc.) was
used for all statistical analyses, and P<0.05 was considered to
indicate a statistically significant difference.
Results
Cytotoxic effect of AG is related to
PDK1 expression
To validate the molecular pathways related to AG,
the known target gene list was downloaded from PubChem (https://pubchem.ncbi.nlm.nih.gov/#query=andrographolide)
and analyzed using Cytoscape's JEPPETTO plugin (https://apps.cytoscape.org/apps/jepetto). Kyoto
Encyclopedia of Genes and Genomes analysis (https://www.genome.jp/kegg/) identified several
cancerous, metabolic and p53 signaling pathways as AG-related
pathways. BioCarta analysis (https://maayanlab.cloud/Harmonizome/search?t=all&q=andrographolide)
revealed that the p53, DNA damage, cell cycle, apoptosis, and HIF
pathways are involved in AG treatment (Table SI, Table SII, Table SIII). Pathway analysis revealed no
direct evidence of a connection between glucose metabolism and AG.
Next, the GSE74769 microarray dataset (obtained from the Gene
Expression Omnibus database; http://www.ncbi.nlm.nih.gov/geo/) was examined to
identify a potential link between glycolysis and AG. The results
clearly revealed that AG treatment downregulated the expression
levels of numerous genes involved in pyruvate metabolism. Among
them, focus was addressed on PDK1, a key regulator of
pyruvate metabolism (22), as its
levels were reduced by >50%, similar to the previously reported
genes, including HK2, PFK1 and LDHA (Fig. S1) (17,18).
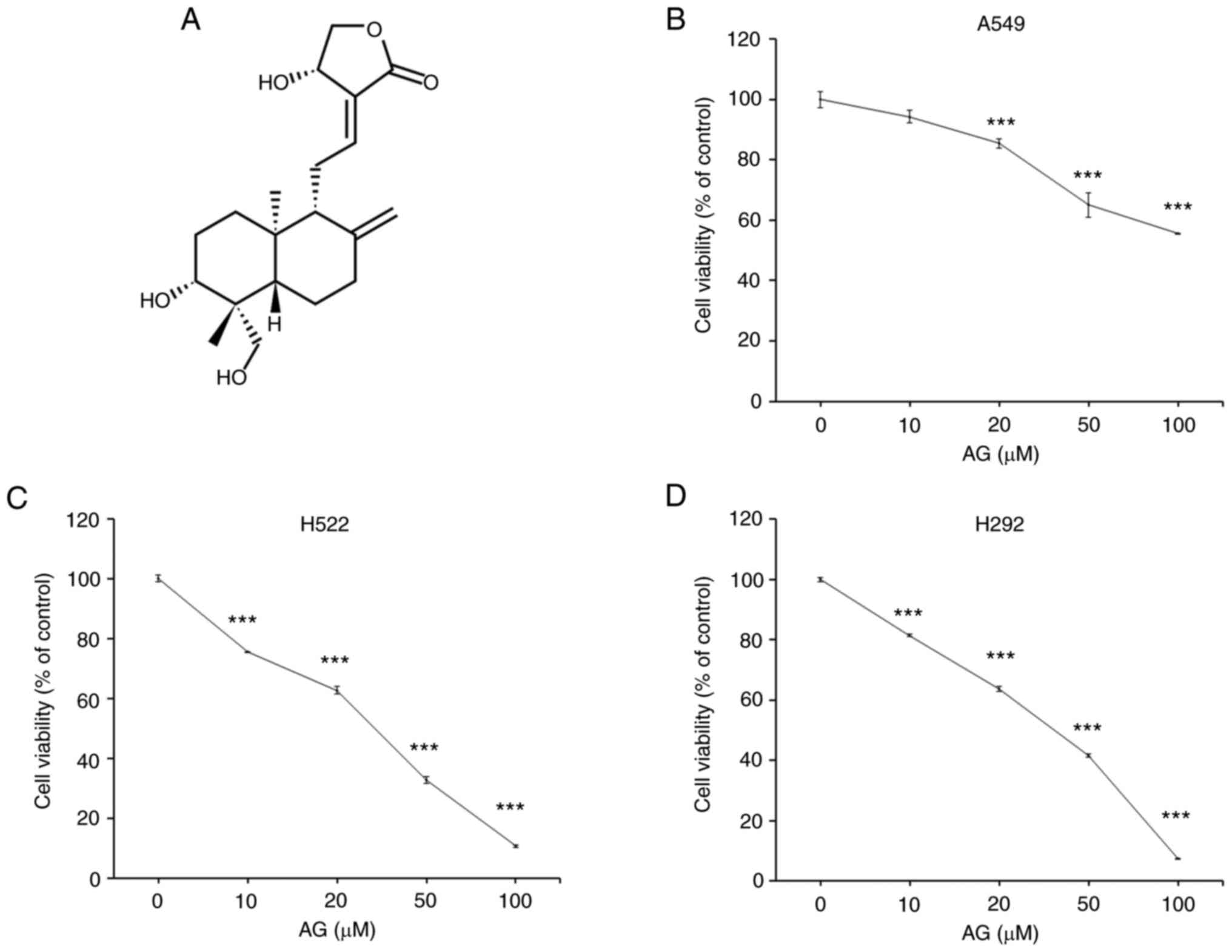
Next, the cytotoxic effects of AG were confirmed on
several NSCLC cell lines, namely the A549, H292 and H522 cell
lines. It was found that AG caused more severe cytotoxicity in H292
and H522 cells than that in A549 cells (Fig. 1). The GI50 (concentration
for 50% of maximal inhibition of cell proliferation) of each cell
line was 102.28 µM for A459, 46.17 µM for H292, and 43.59 µM for
H522. Examination of the Spearman's correlation between PDK mRNA
expression and AG cytotoxicity in these cell lines revealed that
PDK1 was most closely related to AG cytotoxicity (Fig. S2), whereas PDK4 expression was
lacking in these cell lines. H292 and H522 cells demonstrated
higher PDK1 expression than A549 cells at the protein level
(Fig. S3A) as well as the mRNA
level. Furthermore, AG exhibited strong cytotoxicity in HCT116 and
DLD-1 cells with high PDK1 expression (Fig. S3). These findings suggested that
PDK1 may be involved in AG-induced cytotoxicity in cancer
cells.
AG reduces the protein expression
levels of PDK1, 2 and 3
Pyruvate, the end product of glycolysis, can be
converted to lactate in the cytosol or to acetyl CoA to enter the
TCA cycle in mitochondria. Therefore, the effects of AG on
metabolic pathways following pyruvate synthesis were verified.
Similar to a recent study (17), AG
decreased lactate generation in a dose-dependent manner in H292
cells (Fig. 2A). The protein
expression of LDHA, an enzyme that catalyzes the conversion of
pyruvate to lactate, was unchanged (Fig. 2B). AG significantly decreased the
phosphorylation of PDHA1, an enzyme that converts pyruvate to
acetyl-CoA (Fig. 2B). Because the
phosphorylation of PDHA1 is regulated by PDKs, the protein
expression levels of PDKs were examined. As expected, AG induced a
decrease in the protein expression levels of PDK1, 2 and 3 in a
dose-dependent manner in H292 cells (Fig. 2C). In particular, the expression of
PDK1 was markedly reduced. To confirm that AG directly controlled
the activity of PDK1, an in vitro PDK activity assay was
performed using recombinant proteins for protein binding. However,
AG did not directly bind to PDK1 (Fig.
2D). Some binding inhibition was observed at a high
concentration of 10 µM or more, which was comparable to the
intracellular working concentration (20 µM). The positive control,
dichloroacetate, demonstrated a binding inhibitory effect, even at
a concentration of 1 mM, which was substantially lower than the
intracellular working concentration (10 mM). AG not only decreased
the protein expression levels of PDK1, 2 and 3 but also decreased
their mRNA levels in a dose-dependent manner in H292 cells
(Fig. 2E-G). These findings
suggested that AG was involved in the regulation of the
mitochondrial metabolic pathways by inhibiting the conversion of
pyruvate to acetyl-CoA via a decrease in PDK1 expression.
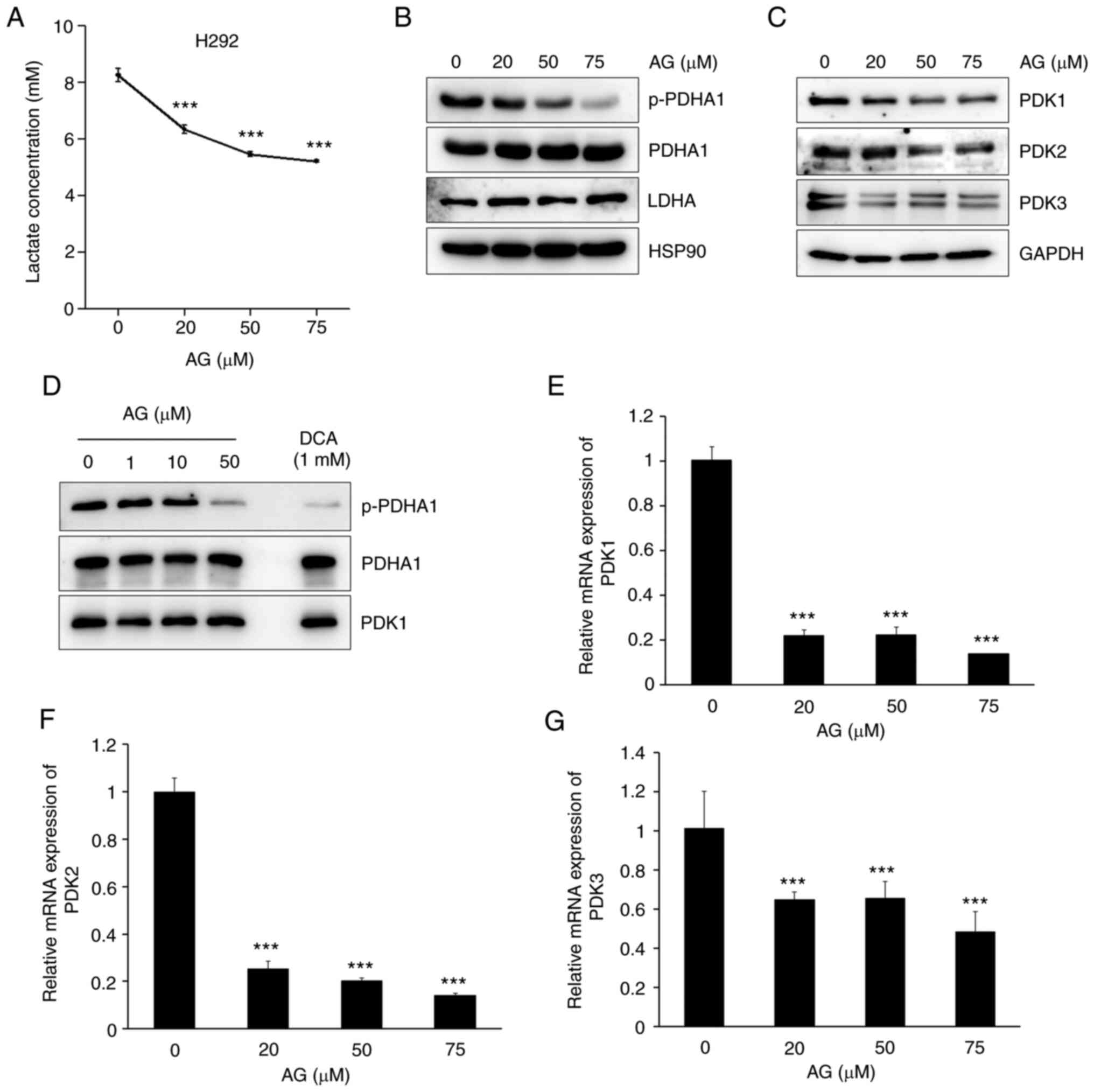 | Figure 2.AG inhibits the PDK protein and mRNA
expression. (A) H292 cells were treated with AG at the indicated
doses for 1 h. (A) AG reduced lactate generation in a
dose-dependent manner. (B) AG significantly inhibited the
expression of p-PDHA1, which is phosphorylated by PDKs. Total
amount of PDHA1 was not changed by AG treatment. Expression of LDHA
also was not affected by AG treatment. HSP90 represents the
normalized control of each well. (C) After H292 cells were treated
with various concentrations of AG (20, 50 and 75 µM) for 6 h, the
protein expression levels of mitochondrial PDK1, PDK2, and PDK3
were determined via western blotting. GAPDH represents the
normalized control of each well. Protein expression levels of PDKs
decreased in a dose-dependent manner after AG treatment. Notably,
PDK1 expression levels significantly decreased after AG treatment.
(D) In vitro PDK activity assay was performed to determine
whether PDK1 activity was regulated by AG. AG did not directly
regulate the activity of PDK1. (E-G) H292 cells were treated with
various concentrations of AG (20, 50 and 75 µM) for 6 h, and the
mRNA levels of PDK1, PDK2 and PDK3 were determined via reverse
transcription-quantitative polymerase chain reaction. AG
significantly decreased the mRNA levels of PDKs in H292 cells. Data
are represented as the mean ± standard deviation compared with the
control from three independent experiments. ***P<0.001 vs.
control. AG, andrographolide; PDK, pyruvate dehydrogenase kinase;
PDHA1, pyruvate dehydrogenase E1 subunit alpha 1; LDHA, lactate
dehydrogenase A; HSP90, heat shock protein 90; GAPDH,
glyceraldehyde 3-phosphate dehydrogenase; p-, phosphorylated. |
AG promotes apoptotic cell death via
mitochondrial ROS generation and loss of MMP
Metabolic reprogramming of cancer cells from aerobic
glycolysis to OXPHOS promotes mitochondrial ROS-dependent cell
death (23,24). Thus, it was investigated whether AG
induced apoptotic cell death and mitochondrial dysfunction in H292
and A549 cells, wherein different PDKs were expressed and different
cytotoxicities were observed (Fig.
1). When the number of AG-induced apoptotic cells after Annexin
V-FITC and PI staining was measured via FACS analysis, it was found
that the number of A549 cells was weakly increased, whereas the
number of H292 cells was significantly increased during apoptosis
(Fig. 3A and B). Similar results
were obtained in the histogram analyzed using the ImageJ software
(Fig. 3C). Levels of cleaved
caspase-3, cleaved caspase-9 and PARP, which are proteins involved
in the apoptotic signaling pathway, were also increased in
AG-treated H292 cells, but not in A549 cells, in a dose-dependent
manner (Fig. 3D).
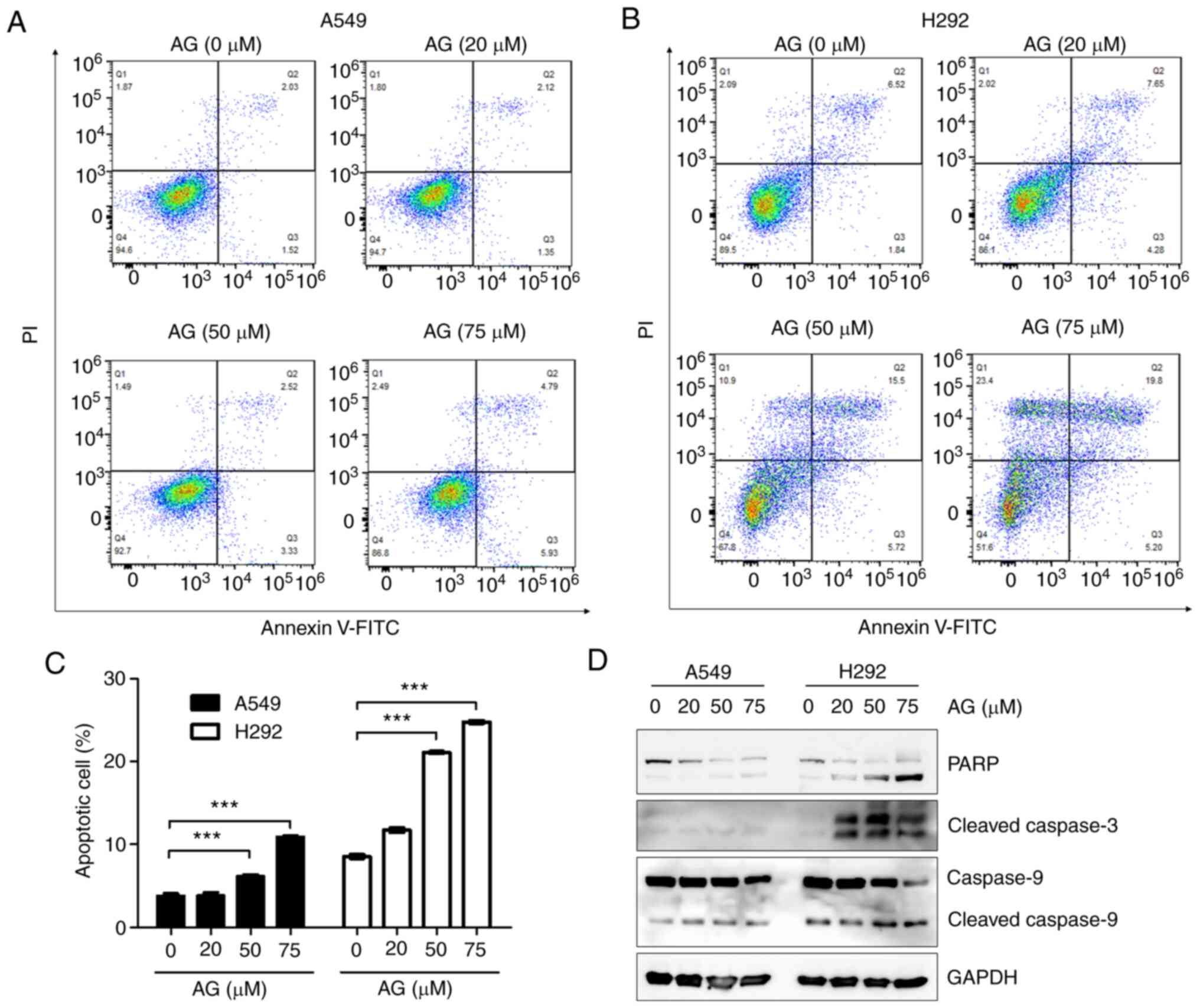 | Figure 3.AG induces more apoptotic cell death
in H292 cells than A549 cells. (A and B) A549 and H292 cells were
treated with various concentrations of AG (20, 50 and 75 µM) for 6
h, labeled with Annexin V-fluorescein isothiocyanate and propidium
iodide, and analyzed via flow cytometry. (C) Histogram showing the
percentage of apoptotic cells. Apoptotic cells were quantified by
combining Q2 (late-stage of apoptotic cells, PI+/Annexin
V+) and Q3 (early-stage of apoptotic cells,
PI−/Annexin V+) regions after FACS analysis.
AG significantly increased the apoptotic cell death in H292 cells,
but not in A549 cells, in a dose-dependent manner. (D) To determine
the protein levels of PARP, cleaved caspase-3, and cleaved
caspase-9 via western blot analysis, A549 and H292 cells were
treated with AG at the indicated dose for 24 h. Levels of all
apoptotic marker proteins were significantly increased by AG
treatment in H292 cells and only slightly increased in A549 cells.
Data are represented as the mean ± SD compared with the control
from three independent experiments. ***P<0.001 vs. control. AG,
andrographolide; PARP, cleaved poly (ADP ribose) polymerase. |
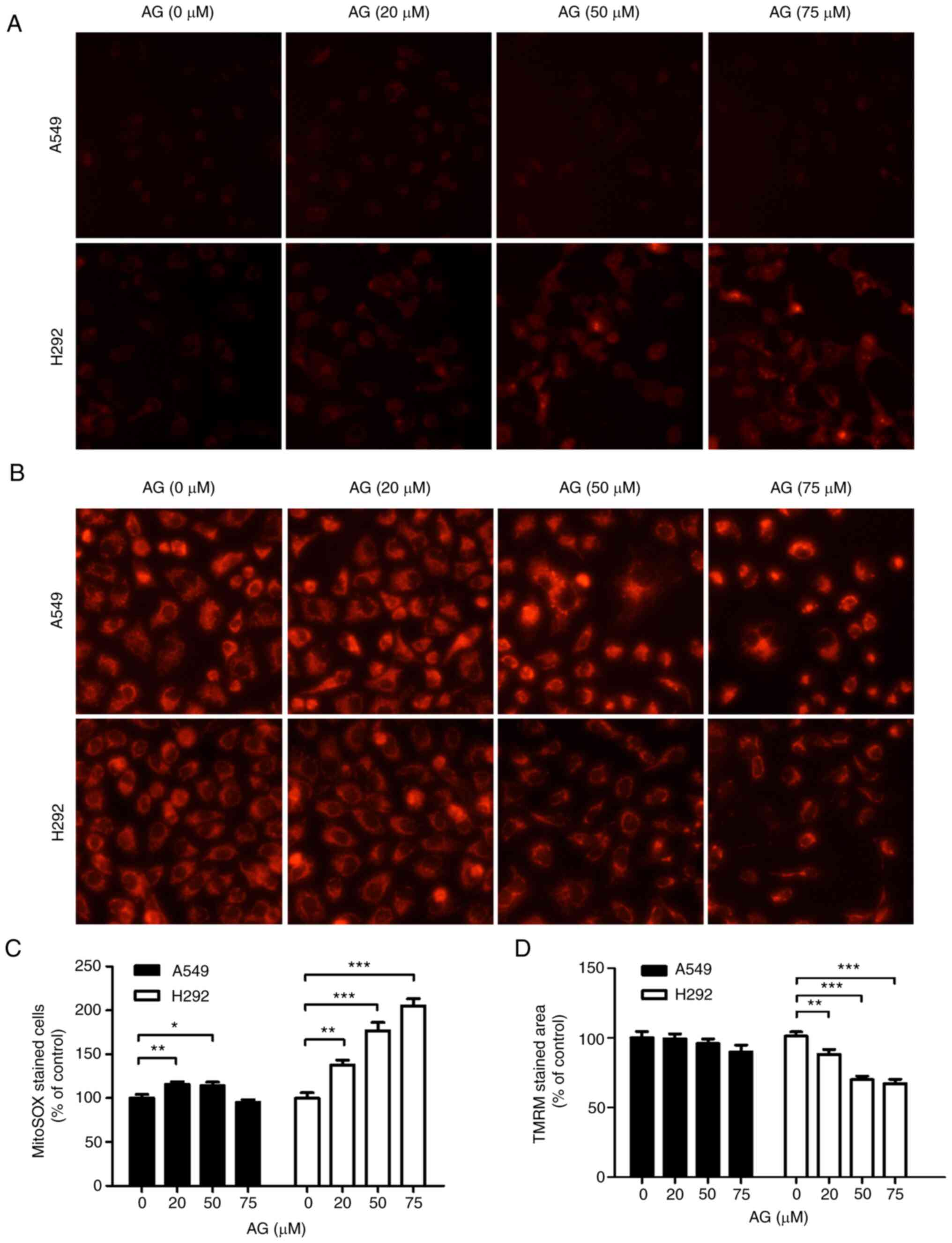
AG remarkably increased the generation of
mitochondrial ROS in H292 cells, as determined via MitoSOX staining
(Fig. 4A). As revealed in the
histogram of the relative amount of AG-induced mitochondrial ROS
production (Fig. 4C), a significant
increase in ROS levels was observed in H292 cells. TMRM staining
assay revealed that AG reduced the MMP in H292 cells in a
dose-dependent manner (Fig. 4B).
Histogram analysis revealed that AG decreased the MMP in H292 cells
but not in A549 cells (Fig. 4D).
These results suggested that AG-induced metabolic shift can lead to
apoptotic cell death in H292 cells by increasing mitochondrial ROS
levels and depleting the MMP.
Cytotoxic effect of AG is influenced
by PDK1 expression
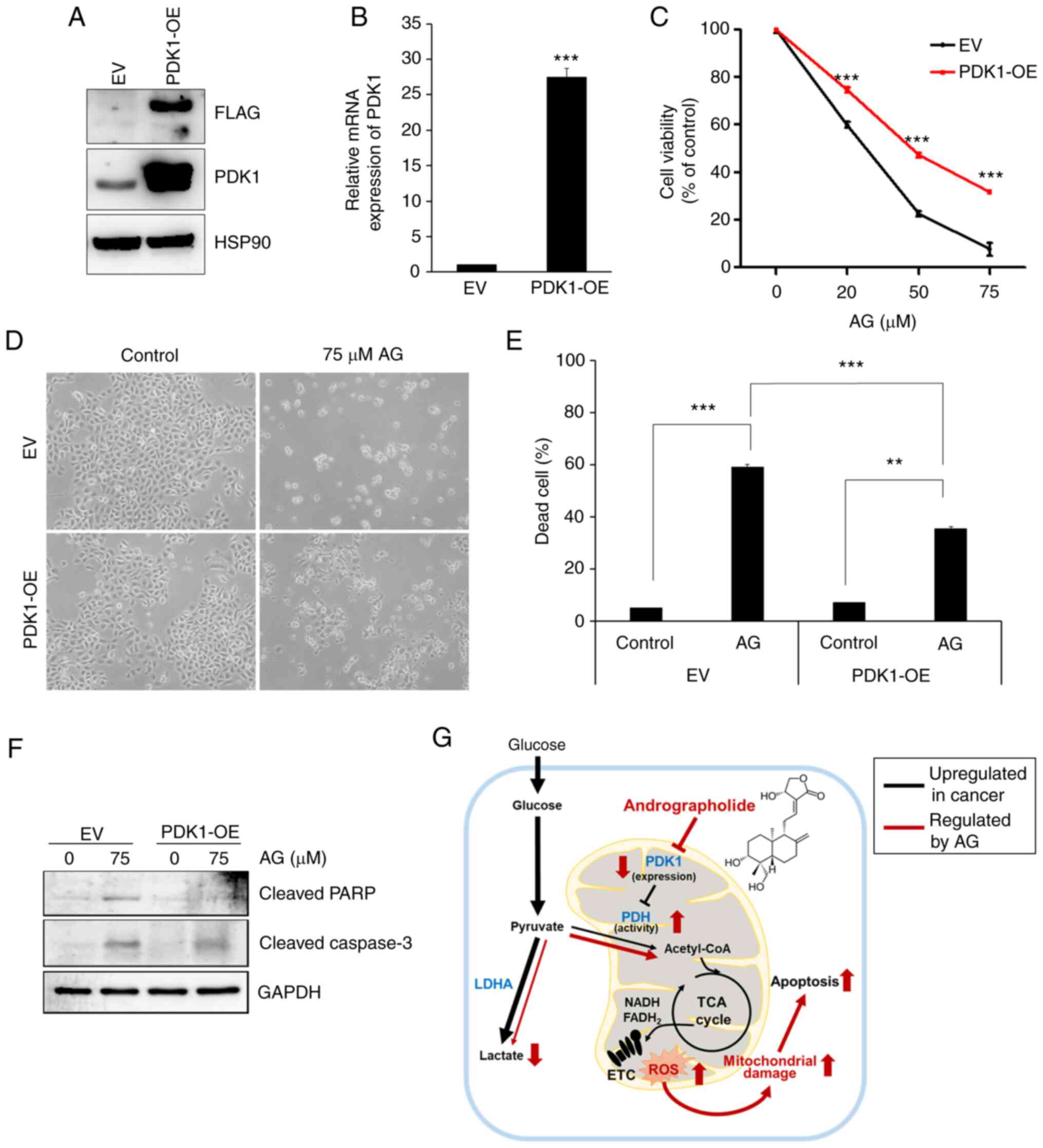
To determine whether the inhibition of PDK1
expression by AG is critical for cancer cell death, a
PDK1-overexpressing (PDK1-OE) cell line was constructed by
transfecting a virus with its expression vector. As revealed in
Fig. 5A, the PDK1 protein was
overexpressed in H292 cells, as determined by western blotting.
PDK1 mRNA was overexpressed in H292 cells, as determined by RT-qPCR
(Fig. 5B). In addition, AG-induced
cytotoxicity in the MTT assay was significantly suppressed in
PDK1-overexpressed H292 cells than that in the EV cells (Fig. 5C). At the highest concentration of
AG (75 µM), only 7.68% of empty vector cells survived, and 31.7% of
PDK1-overexpressed cells survived. The morphological changes in
PDK1-overexpressing H292 and EV cells after treatment with AG are
demonstrated in Fig. 5D. In
PDK1-overexpressing H292 cells, AG-induced cytotoxicity was
relatively weak compared with that in the control (EV) cells.
Histogram also revealed that the number of AG-induced dead cells
was slightly reduced in PDK-overexpressing H292 cells compared with
that in the control cells (Fig.
5E). Expression of apoptotic marker proteins, including
cleaved-PARP and cleaved-caspase 3, was suppressed in
PDK1-overexpressing H292 cells (Fig.
5F). These results indicated that PDK1 expression was partially
involved in the cytotoxic effects of AG. By suppressing PDK1
expression with AG, the metabolic shift from glycolysis to OXPHOS
caused a significant increase in mitochondrial ROS generation,
which is another key factor for AG-induced cancer cell death
(Fig. 5G).
Discussion
PDH complex connects glycolysis to mitochondrial
OXPHOS by converting pyruvate to acetyl-CoA, the main substrate of
TCA cycle (25). PDH activity is
inhibited by PDKs through phosphorylating serine residues in its E1
subunit (PDHA1) (22,26). Thus, PDKs are suggested to be master
regulators of aerobic glycolysis because they limit pyruvate
carboxylation to acetyl-CoA, thereby facilitating pyruvate
reduction to lactate (7,25). PDK1 is the most critical enzyme for
the development and metastasis of various types of malignant cancer
(25,27–30),
particularly NSCLC (31). Several
PDK1 inhibitors, such as dichloroacetate (DCA), AZD 7545, and
2,2-Dichloro-1-(4-isopropoxy-3-nitrophenyl)ethan-1-one (Cpd64),
have been studied to develop novel cancer therapies (32–34).
The use of several natural compounds as PDK1 inhibitors and
anticancer agents in various cancer types have been also reported,
including NSCLC (35–37). Initially, AG was expected to inhibit
PDK1 activity and expression. However, the in vitro PDK
activity assay revealed that AG had a very low inhibitory effect on
PDK1 activity. Recent metabolomic research has revealed an increase
in the levels of TCA cycle metabolites, including citrate,
cis-aconitic acid, and isocitrate, as well as glycolytic
metabolites in AG-treated red blood cells (RBCs) (38). Since mature RBCs do not synthesize
RNA or proteins, the metabolites can reflect the activity of
glycolytic and TCA cycle enzymes. The present findings are
consistent with the aforementioned study, as AG did not have any
effect on the enzyme activity.
PDK expression levels are tightly regulated by
several upstream regulators. The most well-known upstream regulator
of PDK expression is HIF1α, which also increases the expression
levels of other glycolytic enzymes, including GLUT1, HK2, PKM and
LDHA (26,39). HIF1α activates PDK1 and PDK3. It is
part of a series of reactions to achieve a low-oxygen state by
actively decreasing the mitochondrial oxygen consumption via the
induction of PDK expression and inhibition of PDH activity
(40). Other upstream regulators,
including transcription factors E2F1, Myc and Wnt, have been linked
to PDK expression, particularly PDK1 and PDK3 (41–43).
The expression levels of PDK2 and PDK4 are controlled in different
ways. Several metabolic regulators, including forkhead box O1
(FoxO1), liver X receptor (LXR), peroxisome proliferator-activated
receptor α (PPARα) and PPARγ coactivator α (PGC-1α), stimulate PDK2
and PDK4 expression levels (44,45).
Furthermore, p53 tumor suppressor, a master regulator of genome
stability and cell cycle, acts as a negative regulator of PDK2
expression (46).
In the present study, AG reduced PDK1-3 expression
at both the mRNA and protein levels in a dose-dependent manner. AG
inhibits the protein levels of HIF1α via the PI3K/Akt pathway and
ubiquitin-mediated proteolysis (13,14).
Thus, the downregulation of PDK1 and PDK3 expression levels can be
explained, at least in part, by an HIF1α-dependent pathway.
However, the expression of another target of HIF1α for glycolysis,
LDHA, was not altered by AG treatment. Extensive research is needed
to determine the reason for the differential regulation of these
genes. AG increases p53 expression levels in several cancer lines
(47–50) and decreases PPARγ expression levels
in several metabolic diseases (51–53).
Therefore, they may act as upstream regulators of PDK2
downregulation caused by AG treatment. Although the LXR, Myc and
Wnt signaling pathways are associated with AG, their expression
levels are increased by AG treatment (49,50,54).
Therefore, it was concluded that these pathways cannot be the
upstream regulators of PDK expression levels in AG-treated cancer
cells. PubMed database search revealed that E2F1 and FoxO1 pathways
were not previously identified as molecular targets for AG. The
expression of PDK4 was not examined in the present study since it
was not expressed in these cell lines and exerts a suppressive
effect on tumorigenesis in several cancer types, including NSCLC
(55,56).
Although AG treatment reduced the expression levels
of all three isoforms (PDK1-3) in a dose-dependent manner,
correlation analysis revealed that PDK1 may be the most potent
target of AG among them. This possibility was evaluated by
comparing the efficacy of AG in mitochondria-dependent apoptosis in
A549 and H292 cells, which have vastly different PDK1 expression
levels. PDK1-overexpressing cancers, particularly NSCLC (31,57),
are resistant to chemotherapy and radiotherapy (58–60).
In addition, PDK1 protects against apoptotic cell death by reducing
OXPHOS, ROS production and mitochondrial membrane stability
(57,60,61).
As expected, H292 cells expressing high levels of PDK1 experienced
more apoptotic cell death, mitochondrial ROS generation and MMP
depletion than A549 cells expressing low levels of PDK1.
PDK1-overexpressing H292 cells that exogenously expressed PDK1
regardless of AG treatment were generated to confirm the role of
PDK1 in AG-induced apoptotic cell death. Exogenously overexpressed
PDK1 is unaffected by AG regulation on PDK1 expression. Although
PDK1 overexpression did not completely abolish the AG-induced
apoptotic cell death, the portion of dead cell was significantly
recovered by exogenous PDK1 overexpression, as demonstrated in a
previous study by the authors (62). Based on these data, it was concluded
that AG may have another target to induce apoptotic cell death in
NSCLC, as reported in previous studies (9,10,63).
Nonetheless, the present findings suggested that PDK1
overexpression partially reverses AG-induced cytotoxicity and
apoptosis. To elucidate the precise mechanism of AG-stimulated
apoptotic cell death, further studies should examine the upstream
regulators that directly bind to AG and regulate multiple targets,
along with the correlations between PDK1 and other proapoptotic
factors.
In conclusion, it was revealed that AG induced
mitochondria-mediated apoptosis in a PDK1-dependent manner. AG
further inhibited PDK1 expression and increased PDH activity.
Aerobic glycolysis was changed to mitochondrial OXPHOS in cancer
cells. Moreover, PDK1-mediated metabolic shift resulted in
mitochondrial ROS generation, MMP loss and apoptotic cell death.
Therefore, PDK1-mediated metabolic change can explain AG-stimulated
apoptotic cell death as a basis for its anticancer effect; however,
this needs to be investigated further in future studies.
Supplementary Material
Supporting Data
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by Basic Science Research
Program through the National Research Foundation of Korea (NRF)
funded by the Ministry of Education (grant no.
NRF-2020R1I1A1A01068254) and by the National Research Foundation of
Korea (NRF) grants funded by the Korea government (MIST) (grant
nos. NRF-2022R1A2C2005130 and NRF-2021R1A4A1025662).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
ESY and YD performed most of the experiments and
data analysis and wrote the manuscript. TK and KTH conceived and
designed the study and involved in writing the manuscript. SC
performed the LDH assay. BK and JL constructed PDK1 overexpressing
cells. MKC performed FACS analysis. SJB constructed PDK1 expressing
vector. TK and SJB revised the manuscript. ESY, YD and KTH confirm
the authenticity of all raw data. All authors read and approved the
final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pavlova NN, Zhu J and Thompson CB: The
hallmarks of cancer metabolism: Still emerging. Cell Metab.
34:355–377. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Altenberg B and Greulich KO: Genes of
glycolysis are ubiquitously overexpressed in 24 cancer classes.
Genomics. 84:1014–1020. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Jiang B: Aerobic glycolysis and high level
of lactate in cancer metabolism and microenvironment. Genes Dis.
4:25–27. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
DeBerardinis RJ, Lum JJ, Hatzivassiliou G
and Thompson CB: The biology of cancer: Metabolic reprogramming
fuels cell growth and proliferation. Cell Metab. 7:11–20. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lin X, Xiao Z, Chen T, Liang SH and Guo H:
Glucose metabolism on tumor plasticity, diagnosis, and treatment.
Front Oncol. 10:3172020. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Woolbright BL, Rajendran G, Harris RA and
Taylor JA III: Metabolic flexibility in cancer: Targeting the
pyruvate dehydrogenase kinase: Pyruvate dehydrogenase axis. Mol
Cancer Ther. 18:1673–1681. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rajani M, Shrivastava N and Ravishankara
MN: A rapid method for isolation of andrographolide from
Andrographis paniculata nees (kalmegh). Pharm Biol.
38:204–209. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Farooqi AA, Attar R, Sabitaliyevich UY,
Alaaeddine N, de Sousa DP, Xu B and Cho WC: The prowess of
andrographolide as a natural weapon in the war against cancer.
Cancers (Basel). 12:21592020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Islam MT, Ali ES, Uddin SJ, Islam MA, Shaw
S, Khan IN, Saravi SSS, Ahmad S, Rehman S, Gupta VK, et al:
Andrographolide, a diterpene lactone from Andrographis
paniculata and its therapeutic promises in cancer. Cancer Lett.
420:129–145. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hocker HJ, Cho KJ, Chen CY, Rambahal N,
Sagineedu SR, Shaari K, Stanslas J, Hancock JF and Gorfe AA:
Andrographolide derivatives inhibit guanine nucleotide exchange and
abrogate oncogenic Ras function. Proc Natl Acad Sci USA.
110:10201–10206. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nguyen VS, Loh XY, Wijaya H, Wang J, Lin
Q, Lam Y, Wong WS and Mok YK: Specificity and inhibitory mechanism
of andrographolide and its analogues as antiasthma agents on NF-κB
p50. J Nat Prod. 78:208–217. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lin HH, Tsai CW, Chou FP, Wang CJ, Hsuan
SW, Wang CK and Chen JH: Andrographolide down-regulates
hypoxia-inducible factor-1α in human non-small cell lung cancer
A549 cells. Toxicol Appl Pharmacol. 250:336–345. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li J, Zhang C, Jiang H and Cheng J:
Andrographolide inhibits hypoxia-inducible factor-1 through
phosphatidylinositol 3-kinase/AKT pathway and suppresses breast
cancer growth. Onco Targets Ther. 8:427–435. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Li GF, Qin YH and Du PQ: Andrographolide
inhibits the migration, invasion and matrix metalloproteinase
expression of rheumatoid arthritis fibroblast-like synoviocytes via
inhibition of HIF-1α signaling. Life Sci. 136:67–72. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lin HC, Su SL, Lu CY, Lin AH, Lin WC, Liu
CS, Yang YC, Wang HM, Lii CK and Chen HW: Andrographolide inhibits
hypoxia-induced HIF-1α-driven endothelin 1 secretion by activating
Nrf2/HO-1 and promoting the expression of prolyl hydroxylases 2/3
in human endothelial cells. Environ Toxicol. 32:918–930. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen Z, Tang WJ, Zhou YH, Chen ZM and Liu
K: Andrographolide inhibits non-small cell lung cancer cell
proliferation through the activation of the mitochondrial apoptosis
pathway and by reprogramming host glucose metabolism. Ann Transl
Med. 9:17012021. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li X, Tian R, Liu L, Wang L, He D, Cao K,
Ma JK and Huang C: Andrographolide enhanced radiosensitivity by
downregulating glycolysis via the inhibition of the PI3K-Akt-mTOR
signaling pathway in HCT116 colorectal cancer cells. J Int Med Res.
48:3000605209461692020.PubMed/NCBI
|
|
19
|
Devkota AK, Kaoud TS, Warthaka M and Dalby
KN: Fluorescent peptide assays for protein kinases. Curr Protoc Mol
Biol. Chapter 18: Unit 18.17. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mullinax TR, Stepp LR, Brown JR and Reed
LJ: Synthetic peptide substrates for mammalian pyruvate
dehydrogenase kinase and pyruvate dehydrogenase phosphatase. Arch
Biochem Biophys. 243:655–659. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Anwar S, Shamsi A, Mohammad T, Islam A and
Hassan MI: Targeting pyruvate dehydrogenase kinase signaling in the
development of effective cancer therapy. Biochim Biophys Acta Rev
Cancer. 1876:1885682021. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yadav N, Kumar S, Marlowe T, Chaudhary AK,
Kumar R, Wang J, O'Malley J, Boland PM, Jayanthi S, Kumar TK, et
al: Oxidative phosphorylation-dependent regulation of cancer cell
apoptosis in response to anticancer agents. Cell Death Dis.
6:e19692015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Michelakis ED, Webster L and Mackey JR:
Dichloroacetate (DCA) as a potential metabolic-targeting therapy
for cancer. Br J Cancer. 99:989–994. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
McFate T, Mohyeldin A, Lu H, Thakar J,
Henriques J, Halim ND, Wu H, Schell MJ, Tsang TM, Teahan O, et al:
Pyruvate dehydrogenase complex activity controls metabolic and
malignant phenotype in cancer cells. J Biol Chem. 283:22700–22708.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang X, Shen X, Yan Y and Li H: Pyruvate
dehydrogenase kinases (PDKs): An overview toward clinical
applications. Biosci Rep. 41:BSR202044022021. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Hur H, Xuan Y, Kim YB, Lee G, Shim W, Yun
J, Ham IH and Han SU: Expression of pyruvate dehydrogenase kinase-1
in gastric cancer as a potential therapeutic target. Int J Oncol.
42:44–54. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fujiwara S, Kawano Y, Yuki H, Okuno Y,
Nosaka K, Mitsuya H and Hata H: PDK1 inhibition is a novel
therapeutic target in multiple myeloma. Br J Cancer. 108:170–178.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tan J, Lee PL, Li Z, Jiang X, Lim YC, Hooi
SC and Yu Q: B55β-associated PP2A complex controls PDK1-directed
myc signaling and modulates rapamycin sensitivity in colorectal
cancer. Cancer Cell. 18:459–471. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Qu C, Yan C, Cao W, Li F, Qu Y, Guan K, Si
C, Yu Z and Qu Z: miR-128-3p contributes to mitochondrial
dysfunction and induces apoptosis in glioma cells via targeting
pyruvate dehydrogenase kinase 1. IUBMB Life. 72:465–475. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu T and Yin H: PDK1 promotes tumor cell
proliferation and migration by enhancing the Warburg effect in
non-small cell lung cancer. Oncol Rep. 37:193–200. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang W, Hu X, Chakravarty H, Yang Z and
Tam KY: Identification of novel pyruvate dehydrogenase kinase 1
(PDK1) inhibitors by kinase activity-based high-throughput
screening for anticancer therapeutics. ACS Comb Sci. 20:660–671.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Garon EB, Christofk HR, Hosmer W, Britten
CD, Bahng A, Crabtree MJ, Hong CS, Kamranpour N, Pitts S,
Kabbinavar F, et al: Dichloroacetate should be considered with
platinum-based chemotherapy in hypoxic tumors rather than as a
single agent in advanced non-small cell lung cancer. J Cancer Res
Clin Oncol. 140:443–452. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yang Z, Zhang SL, Hu X and Tam KY:
Inhibition of pyruvate dehydrogenase kinase 1 enhances the
anti-cancer effect of EGFR tyrosine kinase inhibitors in non-small
cell lung cancer. Eur J Pharmacol. 838:41–52. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jin L, Kim EY, Chung TW, Han CW, Park SY,
Han JH, Bae SJ, Lee JR, Kim YW, Jang SB and Ha KT: Hemistepsin A
suppresses colorectal cancer growth through inhibiting pyruvate
dehydrogenase kinase activity. Sci Rep. 10:219402020. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kwak CH, Jin L, Han JH, Han CW, Kim E, Cho
M, Chung TW, Bae SJ, Jang SB and Ha KT: Ilimaquinone induces the
apoptotic cell death of cancer cells by reducing pyruvate
dehydrogenase kinase 1 activity. Int J Mol Sci. 21:60212020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kwak CH, Lee JH, Kim EY, Han CW, Kim KJ,
Lee H, Cho M, Jang SB, Kim CH, Chung TW and Ha KT: Huzhangoside A
suppresses tumor growth through inhibition of pyruvate
dehydrogenase kinase activity. Cancers (Basel). 11:7122019.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Alapid AAI, Abd Majid R, Ibraheem ZO,
Mediani A, Ismail IS, Unyah NZ, Alhassan Abdullahi S, Nordin N,
Nasiru Wana M and Basir R: Investigation of andrographolide effect
on non-infected red blood cells using the 1H-NMR-based metabolomics
approach. Metabolites. 11:4862021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim JW, Tchernyshyov I, Semenza GL and
Dang CV: HIF-1-mediated expression of pyruvate dehydrogenase
kinase: A metabolic switch required for cellular adaptation to
hypoxia. Cell Metab. 3:177–185. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Papandreou I, Cairns RA, Fontana L, Lim AL
and Denko NC: HIF-1 mediates adaptation to hypoxia by actively
downregulating mitochondrial oxygen consumption. Cell Metab.
3:187–197. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang LY, Hung CL, Chen YR, Yang JC, Wang
J, Campbell M, Izumiya Y, Chen HW, Wang WC, Ann DK and Kung HJ:
KDM4A coactivates E2F1 to regulate the PDK-dependent metabolic
switch between mitochondrial oxidation and glycolysis. Cell Rep.
16:3016–3027. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kim JW, Gao P, Liu YC, Semenza GL and Dang
CV: Hypoxia-inducible factor 1 and dysregulated c-Myc cooperatively
induce vascular endothelial growth factor and metabolic switches
hexokinase 2 and pyruvate dehydrogenase kinase 1. Mol Cell Biol.
27:7381–7393. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Pate KT, Stringari C, Sprowl-Tanio S, Wang
K, TeSlaa T, Hoverter NP, McQuade MM, Garner C, Digman MA, Teitell
MA, et al: Wnt signaling directs a metabolic program of glycolysis
and angiogenesis in colon cancer. EMBO J. 33:1454–1473. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Piao L, Sidhu VK, Fang YH, Ryan JJ, Parikh
KS, Hong Z, Toth PT, Morrow E, Kutty S, Lopaschuk GD and Archer SL:
FOXO1-mediated upregulation of pyruvate dehydrogenase kinase-4
(PDK4) decreases glucose oxidation and impairs right ventricular
function in pulmonary hypertension: Therapeutic benefits of
dichloroacetate. J Mol Med (Berl). 91:333–346. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Sugden MC and Holness MJ: Mechanisms
underlying regulation of the expression and activities of the
mammalian pyruvate dehydrogenase kinases. Arch Physiol Biochem.
112:139–149. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Contractor T and Harris CR: p53 negatively
regulates transcription of the pyruvate dehydrogenase kinase Pdk2.
Cancer Res. 72:560–567. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gao H, Li H, Liu W, Mishra SK and Li C:
Andrographolide induces apoptosis in gastric cancer cells through
reactivation of p53 and inhibition of Mdm-2. Dokl Biochem Biophys.
500:393–401. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Shi MD, Lin HH, Lee YC, Chao JK, Lin RA
and Chen JH: Inhibition of cell-cycle progression in human
colorectal carcinoma Lovo cells by andrographolide. Chem Biol
Interact. 174:201–210. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Othman NS and Mohd Azman DK:
Andrographolide induces G2/M cell cycle arrest and apoptosis in
human glioblastoma DBTRG-05MG cell line via ERK1/2/c-Myc/p53
signaling pathway. Molecules. 27:66862022. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zhang J, Li C, Zhang L, Heng Y, Wang S,
Pan Y, Cai L, Zhang Y, Xu T, Chen X, et al: Andrographolide, a
diterpene lactone from the traditional Chinese medicine
andrographis paniculate, induces senescence in human lung
adenocarcinoma via p53/p21 and Skp2/p27. Phytomedicine.
98:1539332022. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jin L, Fang W, Li B, Shi G, Li X, Yang Y,
Yang J, Zhang Z and Ning G: Inhibitory effect of andrographolide in
3T3-L1 adipocytes differentiation through the PPARγ pathway. Mol
Cell Endocrinol. 358:81–87. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lin HC, Lii CK, Chen HC, Lin AH, Yang YC
and Chen HW: Andrographolide inhibits oxidized LDL-induced
cholesterol accumulation and foam cell formation in macrophages. Am
J Chin Med. 46:87–106. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Shu J, Huang R, Tian Y, Liu Y, Zhu R and
Shi G: Andrographolide protects against endothelial dysfunction and
inflammatory response in rats with coronary heart disease by
regulating PPAR and NF-κB signaling pathways. Ann Palliat Med.
9:1965–1975. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tapia-Rojas C, Schuller A, Lindsay CB,
Ureta RC, Mejías-Reyes C, Hancke J, Melo F and Inestrosa NC:
Andrographolide activates the canonical Wnt signalling pathway by a
mechanism that implicates the non-ATP competitive inhibition of
GSK-3β: Autoregulation of GSK-3β in vivo. Biochem J. 466:415–430.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li G, Li M, Hu J, Lei R, Xiong H, Ji H,
Yin H, Wei Q and Hu G: The microRNA-182-PDK4 axis regulates lung
tumorigenesis by modulating pyruvate dehydrogenase and lipogenesis.
Oncogene. 36:989–998. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Atas E, Oberhuber M and Kenner L: The
implications of PDK1-4 on tumor energy metabolism, aggressiveness
and therapy resistance. Front Oncol. 10:5832172020. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
De Rosa V, Iommelli F, Terlizzi C,
Leggiero E, Camerlingo R, Altobelli GG, Fonti R, Pastore L and Del
Vecchio S: Non-canonical role of PDK1 as a negative regulator of
apoptosis through macromolecular complexes assembly at the
ER-mitochondria interface in oncogene-driven NSCLC. Cancers
(Basel). 13:41332021. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Yao S, Shang W, Huang L, Xu R, Wu M and
Wang F: The oncogenic and prognostic role of PDK1 in the
progression and metastasis of ovarian cancer. J Cancer. 12:630–643.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Peng F, Wang JH, Fan WJ, Meng YT, Li MM,
Li TT, Cui B, Wang HF, Zhao Y, An F, et al: Glycolysis gatekeeper
PDK1 reprograms breast cancer stem cells under hypoxia. Oncogene.
37:11192018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Lu H, Lu Y, Xie Y, Qiu S, Li X and Fan Z:
Rational combination with PDK1 inhibition overcomes cetuximab
resistance in head and neck squamous cell carcinoma. JCI Insight.
4:e1311062019. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Jin L, Cho M, Kim BS, Han JH, Park S, Lee
IK, Ryu D, Kim JH, Bae SJ and Ha KT: Drug evaluation based on
phosphomimetic PDHA1 reveals the complexity of activity-related
cell death in A549 non-small cell lung cancer cells. BMB Rep.
54:563–568. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Kim BS, Chung TW, Choi HJ, Bae SJ, Cho HR,
Lee SO, Choi JH, Joo JK and Ha KT: Caesalpinia sappan induces
apoptotic cell death in ectopic endometrial 12Z cells through
suppressing pyruvate dehydrogenase kinase 1 expression. Exp Ther
Med. 21:3572021. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Zeng B, Wei A, Zhou Q, Yuan M, Lei K, Liu
Y, Song J, Guo L and Ye Q: Andrographolide: A review of its
pharmacology, pharmacokinetics, toxicity and clinical trials and
pharmaceutical researches. Phytother Res. 36:336–364. 2022.
View Article : Google Scholar : PubMed/NCBI
|